
Submitted:
24 July 2024
Posted:
24 July 2024
You are already at the latest version
Abstract
Some of neurodegenerative diseases may be characterized by continuing behavioral and cognitive dysfunction that contains memory loss and/or apathy. Alzheimer's disease is the most typical type of such neurodegenerative disease characterized by deficit of cognition and alteration of behavior. Despite the huge efforts against Alzheimer's disease, there has been yet no successful treatment for this disease. Interestingly, several possible risk genes to cognitive dysfunction are frequently expressed within brain cells, which may also be linked to cholesterol metabolism, lipid transport, exosomes and/or caveolae formation, suggesting that caveolae may be a therapeutic target to consider cognitive dysfunctions. Interestingly, modulation of autophagy/mitophagy with the alteration of glucagon-like peptide-1 (GLP-1) and N-methyl-d-aspartate (NMDA) receptors signaling may offer a novel approach to prevent and alleviate the cognitive dysfunction. A paradigm that both GLP-1 and NMDA receptors at caveolae sites is promising and crucial for the treatment of cognitive dysfunctions has been presented here, which might also be able to modify the progression of Alzheimer's disease. This research direction may open the potential to move a clinical care toward disease-modifying treatment strategies with maximal benefits for patients without detrimental adverse events for neurodegenerative diseases.
Keywords:
caveolae
; caveolin
; glucagon-like peptide-1
; NMDA
; autophagy
; mitophagy
; cognitive dysfunction
; Alzheimer's disease
1. Introduction
Given the risk of worsening and changing to dementia, concern to cognitive dysfunctions has recently expanded. Cognitive dysfunctions are characterized by a reduction in the weight/volume of the brain due to brain cortical atrophy with widening of the brain grooves. This brain atrophy predominantly encompasses the hippocampus. At this time, there is almost no treatment for the cognitive dysfunctions [1,2]. Alzheimer's disease is the most general type of cognitive dysfunctions, disturbing almost a quarter of the senior over 80 years old [1,2]. Alzheimer's disease is also a chronic neurodegenerative disease, which is considered as one of the most difficult medical problems with substantial financial and/or social costs. The discovery programs for the treatment of cognitive dysfunctions under clinical trials targeting inflammatory mediators, N-methyl-d-aspartate (NMDA) receptors, nicotinic acetylcholine receptors, secretase modulators, cholinergic system, tau, and glucagon-like peptide-1 (GLP-1) have been explored. Among them, the current drug for the treatment of Alzheimer's disease, memantine, a NMDA receptor antagonist, and acetylcholinesterase inhibitors may momentarily amend the cognitive decline, even without the termination of the dementia progression. Therefore, novel techniques are immediately required to considerably improve the cognitive impairment with safety [3]. Attention in the preclinical stages of dementia also remains meaningfully to deepen within the research and/or medical areas. The identification for the modifiable risk factors as well as disease-modifying treatments for Alzheimer's disease has been emergently required, which could target the early or mild stages of cognitive dysfunctions [4,5].
Cholesterol is one of the most essential biomolecules in cellular physiology due to its involvement in several key biological processes. For example, cholesterol forms caveolae and/or lipid rafts that expedite the localization of several key molecules including receptors and downstream enzymes in cells of central nervous system (CNS). Cholesterol also plays an important function in the plasticity of synapse [6]. Remarkably, the CNS is different from the other peripheral organs in terms of cholesterol metabolism and/or requirements [7]. The depletion of cholesterol from the plasma membrane of some neurons might lead to the complete loss of caveolae structure, which may drastically impair their chemo-migratory responses [8]. The majority of risk genes to Alzheimer's disease are commonly expressed within brain microglia, which may be linked to cholesterol metabolism, lipid transport, endocytosis, exocytosis and/or caveolae formation [9]. In addition, through caveolae and cholesterol-rich lipid raft endocytic pathways within CNS, it has been shown that some extracellular vesicles enter oligodendrocyte progenitor cells exerting a pro-maturation effect onto oligodendrocytes [10]. Accordingly, learning the roles of cellular membrane biophysics with neuronal/microglial function should improve our understanding of the pathology for the cognitive dysfunctions.
2. Caveolae
Caveolae are approximately 100-nm flask-shaped invaginations of the plasma membrane with the presence of caveolin proteins [11]. Caveolae may also hold several signaling proteins including various transmembrane receptors, basically through their interaction with some of caveolins, which have been theorized to act as crucial hubs for facilitating intracellular signal transduction [12]. (Figure 1) Therefore, caveolae might be involved in the synapse formation and its plasticity. For example, neuron-targeted caveolin-1 may improve the NMDA- and/or BDNF-mediated signaling, which could increase the growth of dendritic cells [13]. Consequently, by which the mechanical changes in hippocampal neurons may be associated with enhancements in hippocampal-function of learning and/or memory [14]. In addition, a different study has revealed that caveolin-1 can interact with the function of several metabotropic glutamate receptors in the hippocampus assisting their expression [15]. Caveolin-1 might also regulate the transport of metabotropic glutamate receptors to the membrane surface, which may be responsible for the synaptic facilitation in the formation of long-term potentiation (LTP) [16]. Caveolin-1 may also be involved in the endocytosis of AMPA receptor in the hippocampus, which might reverse the metabotropic glutamate receptors-related long-term depression (LTD) [17]. Conversely, overexpression of caveolin-1 could promote the endocytosis of AMPA receptor from caveolae-like structures wihin neurons of hippocampus, which may assist the contribution of caveolin-1 in the receptor trafficking [18]. Interestingly, it has been revealed that the AMPA receptor trafficking possibly mediated by caveolin-1 could improve the primary behavioral disorders [17,19]. Therefore, caveolin-1 might be important for the augmentation of metabotropic glutamate receptors-related LTD, and for further bringing AMPA receptor trafficking.
In general, the key protein of caveolae is the caveolin-1 that is a 23 kDa of membrane protein with a hairpin-like structure ubiquitously expressed in various tissue cells, while caveolin-3 is extremely expressed in striated muscle cells [20]. The amino- and carboxy-terminal domains are preoccupied with the cytoplasm, whereas the hairpin loop is in transmembrane zone. In addition to create caveolae structure through the hetero-oligomeric complex with caveolin-2, the juxtamembrane region in the amino-terminus of the caveolin-1 may specially work as a scaffold protein, associating with a variety of signaling molecules including G-proteins [21,22]. Moreover, caveolin-1 can significantly cooperate with insulin receptors maintaining a role in energy/metabolic regulation [23]. Interestingly, it has been revealed that caveolin-1 could indorse structural plasticity of neurons as well as neurogenesis in brain, suggesting that caveolae may be a fruitful therapeutic target to consider cognitive dysfunctions [24]. In addition, caveolin-1 has been a possible therapeutic target for a diabetes-induced cognitive dysfunction [25]. Remarkably, the alteration of caveolin-1 could induce memory deficits that take after the neurological phenotype of Alzheimer's disease [26]. Caveolin-1 can physically associate with the mature amyloid precursor protein [27]. Roles of caveolins at caveolae might affect the neurological condition conceivably through an anti-oxidative stress-dependent mechanism in the process of neurodegeneration [26,28,29]. It has been shown that caveolin-1 may be a novel candidate with neuroprotective and/or anti-oxidative effects through modulating neuronal ferroptosis-mediated mitochondrial homeostasis [30]. Further studies are indispensable to evaluate the precise effect of caveolae dysfunction for the cascade of cognitive dysfunctions.
3. Two Relevant Key Receptors to Cognition in Caveolae
Activation of GLP-1 receptors and their receptor internalization may result in the intracellular signaling that is regulated by availability of downstream molecules. In addition, it has been shown that GLP-1 receptor co-localizes with the caveolin-1 at caveolae, which may be involved in controlling the receptor activity by assembling signaling complexes with the receptor trafficking [31]. In fact, the sequence of GLP-1 receptor contains a classical caveolin-1 binding motif within the second intracellular loop [32], by which GLP-1 receptor can directly associate with the caveolin-1 [33]. This interaction may be required for the internalization of receptors in caveolae and also for the removal of GLP-1 receptor [34,35]. On the contrary, GLP-1 receptor is not internalized even after the agonist stimulation in cells expressing the P132L-altered caveolin-1, a mutated form of the caveolin-1, which may result in misfolded oligomers accumulated within the Golgi complex [36]. The activated standard GLP-1 receptor generally undergoes agonist-mediated endocytosis, which may lead either to recycle the receptor back to the plasma membrane or to degradative autophagosome pathway [37]. Since caveolin-1 may be required for the internalization of GLP-1 receptor after an agonist stimulation, it is plausible that caveolin-1 might also affect the fate of GLP-1 receptor determining either degradation or recycling of GLP-1 receptor [38]. Therefore, the association between GLP-1 receptor and caveolin-1 is crucial not only for receptor trafficking, but also for the instigation of various intracellular signaling pathways. Remarkably, the GLP-1 receptors in the hippocampus of brain are also involved in the modulation of NMDA receptor activity through the modulation of cAMP-response element binding protein (CREB) [39]. In addition, GLP-1 analogues may also be useful for the treatment of Alzheimer's disease, because they have been linked to neuroprotective and/or anti-inflammatory properties [40]. Modulation of GLP-1 activity could affect the aggregation of amyloid-beta in the brain of Alzheimer's disease [40]. The GLP-1 receptor agonists have been shown potentially to have favorable actions on the brain with several neurological deficits via the phosphoinositide-3 kinase (PI3K)/AKT/mTOR signaling pathway [41]. (Figure 2) It has also been shown that downstream ionotropic glutamate receptors such as NMDA receptor signaling in hippocampus may be required for the GLP-1 receptor mediated behaviors, suggesting that a signaling from GLP-1 receptors to a NMDA-receptor pathway may be at least modulating a behavior in the realm of neuronal control. [42]. NMDA receptors are kinds of ionic glutamine receptors involved in brain development and/or functions such as learning and memory formation. Synaptic NMDA receptors are indeed important for synaptic plasticity and neuronal survival, which might also play an indispensable role in memory and learning [43]. This NMDA receptor may be responsible for mediating LTP and maintaining brain health. Physiological precise functions of NMDA receptors may be shaped by their subunit composition within CNS [44,45]. Noteworthy, recent studies in mice have detected a reduction in the expression of the NMDA receptor to the dendritic spine, leading to the formation of extrasynapse, which is associated with advanced neuronal cell death and accelerated age-related cognitive decline [46]. In addition, targeting the NMDA receptor has been proposed to be therapeutically advantageous in addressing various neurodegenerative disorders. Up to now, a great deal of the related targets have been explored [47]. Illuminating a novel functional harvest of GLP-1 receptor and NMDA receptor in neural connectivity of brain hippocampus, this may enlighten novel neurological targets for the treatment of cognitive dysfunctions. However, translating studies for agonists/antagonists of these receptors into fruitful clinical medications has been a hard mission. This is principally owing to the complexity of receptors-signaling, as their functions might profoundly be associated with various fundamental functions of brain. Abnormality either of hypofunction or hyper-function of those signaling may be harmful to whole brain health [48]. For example, the use of NMDA receptor agonists/antagonists in various neurodegenerative disorders has not yet to produce the satisfactory consequence, suggesting an inadequate understanding of the receptor’s function in brain [49]. In addition, some of these agonists/antagonists have been obvious by detrimental adverse effects such as schizophrenia-like psychological effects [50]. The significance of glutamate-induced excito-toxic neuronal cells death in the pathogenesis of neurodegenerative disorders has been well recognized. Although memantine and amantadine are clinically accessible, they are not without adverse effects at all. Similarly, Ifenprodil, a selective allosteric modulator targeting a NMDA receptor, has exhibited positive results with good profiles of small adverse effects, only suggesting symptomatic improvement. A better understanding of the mechanism of agonists/antagonists to GLP-1 and NMDA receptors is anticipated.
4. Autophagy/Mitophagy
It is of great importance to identify effective interventions that could delay or prevent cognitive dysfunctions. In view of the fact that the hyperactivation of mTOR in patients with Alzheimer’s disease may impair the autophagy contributing to the accumulation of plaques and tangles, the dysregulation of autophagy is a key pathological feature of Alzheimer’s disease [51]. In this regard, it has been shown that the p62 molecule, an autophagy receptor, is a multifunctional protein that has been associated in the pathology of Alzheimer’s disease due to its ability to attach with neurofibrillary tangles for degradation [52]. Additionally, the mitochondrial dysfunction with increase of damaged mitochondria, an impairment of mitophagy/autophagy, has been well-known to contribute to the cognitive dysfunctions [53]. Therefore, improved autophagy and/or mitophagy could reduce the pathology of Alzheimer’s disease [54]. Interestingly, a promising applicant material for this is the disaccharide trehalose, which has shown the effectiveness in inhibiting experimental neurodegeneration in several models of Alzheimer’s disease [55]. Some mechanisms of therapeutic action have been identified, which suggests that trehalose can regulate autophagy by encouraging rapid lysosomal enlargement and membrane permeabilization correlating with the calcium-dependent calcineurin activation [56]. By stimulating autophagy/mitophagy, this substance could consequently enhance the removal of cytotoxic proteins including amyloid-β aggregates as well as damaged mitochondria, suggesting that autophagy/mitophagy could possess a considerable therapeutic activity [57,58]. Autophagosomal activation by the treatment of trehalose could also remove the cytotoxic properties of stuffs including hyper-phosphorylated tau [58,59]. Notably, the experimental alteration of autophagy/mitophagy could commonly hamper the neuroprotective effect of trehalose [60]. In addition, it has been revealed that the dose-dependent effects of appropriate autophagy in the hippocampus and/or frontal cortex could improve the cognitive impairment as well as behavioral disturbances [61]. Now, autophagy/mitophagy has been identified as a strategic player and/or target in the pathology of cognitive dysfunctions or neurotoxicity.
In general, autophagy/mitophagy is developed to clear misfolded proteins and damaged mitochondria thereby supporting cellular homeostasis and survival [62], which may be regulated by the adenosine monophosphate-activated protein kinase (AMPK) and mammalian/mechanistic target of rapamycin (mTOR) cascade [63]. Remarkably, the activation of GLP-1 receptor could expedite the autophagy in the relevant signaling pathways [64]. (Figure 2) Interventions of GLP-1 mediated modulation of autophagy/mitophagy might be applied in the clinical practice to reduce the incidence of type 2 diabetes mellitus for the protection of islet beta-cells [65]. It is known that amyloid-β could interact with the neuronal NMDA receptor, resulting in the excessive entry of calcium thereby causing excito-toxicity for neuronal cells [66]. (Figure 3) Memantine, an FDA-approved drug usually prescribed for patients with Alzheimer’s disease, is a non-competitive inhibitor of the NMDA receptor, which could work in the regulation of calcium entry into cytoplasm [67]. Additionally, memantine has been also shown to stimulate autophagy reducing the accumulation of amyloid-β with the amended survival of neurons [68,69]. Another antagonist for NMDA receptor with calcium channel-blocking might also exhibit to slowdown the cognitive decline [70]. Because modulation of NMDA receptor activity could change the excitability of neuronal routes, however, slight differences in the mechanisms of action of NMDA receptor antagonists could fervently impact on their clinical effects [70] (Figure 4).
5. Clinical Translation
As for the clinical translation, the effectiveness with less adverse events would be critical applied in any medical science. Phenomenon of an excito-toxicity with glutamate stimulation may have guided to the discovery of new molecules that could be employed to treat neurodegenerative diseases by obstructing NMDA receptors. Among the NMDA receptor inhibitors, memantine has been permitted for the treatment of Alzheimer's disease and/or dementia, however, no other inhibitor of NMDA receptors has approved for clinical trials [71]. Memantine, an NMDA receptor channel blocker, could prevent glutamine hyper-activity in Alzheimer's disease by modulating autophagy, which has been used as well for the treatment of mild cognitive dysfunctions [72,73]. Memantine could bind the NMDA receptor within the entrance of the ion channel, which may help to close the ion channel gate, actually blocking ion permeation [73]. Excess activation of superfluous NMDA receptors could bring about the death of glial cells/neurons that is deliberated as a relevant factor in the development of Alzheimer's disease, which could be at least in part avoided by memantine [74]. Another receptor antagonist ifenprodil has been found to be an effective negative modulator that could also inhibit NMDA receptors including the GluN2B subunit [75]. Arrangement of memantine with the ifenprodil or a selective serotonin reuptake inhibitor (SSRI) has been more proficient in improving cognitive significances [75,76]. Interestingly, it has been shown that citalopram, an SSRI, could also activate autophay/mitophagy in patients with Alzheimer’s disease [77]. On the other hand, GLP-1 receptor agonists may also prevent the neurotoxicity associated with Alzheimer's disease, while presently encouraged for the treatment of type 2 diabetes or obesity. For example, one of the GLP-1 receptor agonists, semaglutide, has been clinically studied for the reduction of neuroinflammation as well as amyloid-β and/or cortical tau protein in the brain with Alzheimer's disease [78]. Exenatide, another GLP-1 receptor agonist may be an available therapeutic agent also presently used in the treatment of diabetes or obesity, which might have additional and favorable effects on cognitive function in human patients [79]. Furthermore, the impact of GLP-1 from dipeptidyl peptidase-4 inhibitors has also been evaluated on cognitive function in patients with type 2 diabetes mellitus [80].
Parenthetically, preceding investigations have emphasized that spermidine has a favorable ability to initiate the process of disappearing amyloid-beta plaques by the mechanism of autophagy [81]. In addition, polyphenols such as anthocyanins could also amend the memory, learning, and cognitive function by modifying autophagy [82]. Interestingly, mild fasting might also stimulate autophagy for the beneficial effects in the brain function [83]. In line with this, a recent study has suggested that a form of recurrent mild fasting within a restricted time frame may contribute to the treatment of neurodegenerative diseases, which might improve autophagy, decrease oxidative stress, and expand cognitive function [84]. Similarly, it has been shown that a diet restricted to the circadian rhythm may stimulate the autophagy, producing an increased level of brain-derived neurotrophic factor (BDNF) in the forebrain area, and increasing synaptic plasticity, thus enhancing cognitive function [85]. In these ways, clinical studies have currently shown that appropriate autophagy/mitophagy can significantly improve cognitive function, although the mechanisms are not completely understood.
6. Future Directions
Dementia, cognitive disorders, and/or depression are now notorious owing to several limitations for the therapy. Strategies that slow or prevent the clinical progression of these disorders have mostly remained elusive, until recently. Therefore, further investigations involving validated tools for the therapy are required. Many competitive or noncompetitive agonists/antagonists of GLP-1 and/or NMDA receptors might have been manifested by undesirable adverse effects, which are connected to the receptors-binding affinity and/or allosteric modulation of receptors function. Through NMDA ionotropic receptor, certain amino acids could modify intracellular Ca2+ dynamics allowing the cell proliferation and/or apoptosis in neurulation. Glutamate synthesis depends on the mitochondrial enzymes such as glutaminase 1 (GLS1) that is widely expressed in brain, which may contribute to the neurogenic processes [86]. Interestingly, it has been shown that the NMDA receptor antagonism with the GLP-1 receptor agonism may effectively reverse the hyper-glycemia, dyslipidemia and/or obesity in animal model of metabolic diseases [87]. These modulators may offer potential in addressing various psychiatric and/or neurodegenerative disorders. Therefore, a more comprehensive exploration of the structure and/or function of these receptors would have led to the development of some specific modulators with satisfactory side-effect profiles. For example, pharmacological NMDA receptor-blockade could attenuate the food intake decrease resulting from the stimulation of astrocytes, which could also modulate the fasting or anorexic effect of GLP-1 [88]. As food intake behavior may be under the strict control of CNS, the signaling actions of NMDA receptor with GLP-1 receptor might be encouraging for the safe modulation of CNS. Nevertheless, you can’t see the forest for the trees. The ideal approach would be to utilize agonists of GLP-1 receptors and/or antagonists of NMDA receptors to halt the neuron-damaging process for the neuroprotection. Currently, these approach for the treatment of cognitive dysfunctions does have only its limitations due to detrimental adverse effects. However, these limitations might have given researchers further insight on how to augment the efficacy to enable precise adjustment of autophagy/mitophagy and enhance the crossfire of therapeutics at the caveolae site. (Figure 4) The use of biomarkers that indicate pathological alterations suggesting the development of cognitive dysfunctions may significantly contribute not only to the effort to identify the disease as early as feasible but also to the evaluation for the precise therapeutic efficacy during the disease process with the treatment. To probe the minimal essential dose for the effective treatment is necessary for the safety of therapy. Concurrently, clinical trials should eventually be evaluating the impact of biomarkers that theoretically slow the progression of cognitive dysfunctions.
Exosomes are kinds of extracellular vesicles released upon the exocytosis characterized by a bilayer membrane structure. (Figure 1) Exosomes in the CNS could transmit messages across the brain via the cerebrospinal fluid (CSF) [89]. Exosomes, containing specific cargoes such as lipids, proteins, and nucleic acid molecules including non-coding RNAs or DNA and other bioactive compounds, are transported from donor cells to distant recipient cells through mechanisms like endocytosis/phagocytosis, exocytosis, and/or plasma membrane fusion, facilitating an information transmission and/or a material movement. Therefore, exosomes may participate in the cell to cell communication, which may play a key role in the development of several tissues/organs including CNS. Interestingly, it has been shown that exosomes with mitochondrial DNA may possess potential as an indicator of cognitive consequences [90]. Recently, the increased study of exosomes has led to an increasing attention of their involvement in neurodegenerative disorders including Alzheimer’s disease. While therapeutic treatments with nanotechnology might currently be effective in treating the cognitive dysfunctions, those exosomes should be applied to the evaluation for the precise therapeutic efficacy during the disease progression/suppression with treatments. Again, there is an urgent necessity for diagnostics and/or therapeutic options against cognitive dysfunctions. Additionally, certain exosomes may have a key role in Alzheimer's disease by contributing to alleviate the pathological progression of the disease [91]. For example, some exosomes have been detected to regulate specific microRNAs including miRNA-17-5p, miRNA-21, and miRNA-126-3p, encouraging their therapeutic potential [91]. By modulating the autophagy, some exosomes may potentially boost neurogenesis contributing to improve learning and/or memory, decreasing Aβ accumulation, then opposing Alzheimer’s disease [92]. In fact, it has been recognized a novel mechanism linking some exosomes and synaptic susceptibility for the innovative treatment of cognitive dysfunctions [93]. At last, it is worth mentioning that brain can discharge several exosomes to communicate with peripheral along with distant cells/organs, which might also assist the homeostasis of brain cognitive action.
Author Contributions
Conceptualization, MN, SY, NS, and SM; original draft preparation and editing, MN, and SM; visualization, MN and SM; supervision, SM. Each author (MN, NS, SY, and SM) has participated sufficiently in this work of drafting the article and/or revising the article for the important rational content. Then, all authors gave final approval of the version to be submitted. Finally, all authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare that they have no competing financial interests.
Abbreviations
| AMPK | adenosine monophosphate-activated protein kinase |
| BDNF | brain-derived neurotrophic factor |
| CSF | cerebrospinal fluid |
| CNS | central nervous system |
| GLS1 | glutaminase 1 |
| GLP-1 | glucagon-like peptide-1 |
| CREB | cAMP-response element binding protein |
| LTD | long-term depression |
| LTP | long-term potentiation |
| mTOR | mammalian/mechanistic target of rapamycin |
| NMDA | N-methyl-d-aspartate |
| ROS | reactive oxygen species |
| SSRI | selective serotonin reuptake inhibitor |
References
- Rong L, et al. Effects of ketogenic diet on cognitive function of patients with Alzheimer's disease: a systematic review and meta-analysis. J Nutr Health Aging. 2024;28(8):100306.
- Carotenuto, A.; Andreone, V.; Amenta, F.; Traini, E. Effect of Treatment of the Cholinergic Precursor Choline Alphoscerate in Mild Cognitive Dysfunction: A Randomized Controlled Trial. Medicina 2024, 60, 925. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, T.; Chouliaras, L.; Morrell, R.; Rubio, D.; Radford, D.; Marchant, N.L.; Walker, Z. The criteria used to rule out mild cognitive impairment impact dementia incidence rates in subjective cognitive decline. Alzheimer's Res. Ther. 2024, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Livingston, G.; Huntley, J.; Sommerlad, A.; Ames, D.; Ballard, C.; Banerjee, S.; Brayne, C.; Burns, A.; Cohen-Mansfield, J.; Cooper, C.; et al. Dementia prevention, intervention, and care: 2020 report of the Lancet Commission. Lancet 2020, 396, 413–446. [Google Scholar] [CrossRef] [PubMed]
- Self, W.K.; Holtzman, D.M. Emerging diagnostics and therapeutics for Alzheimer disease. Nat. Med. 2023, 29, 2187–2199. [Google Scholar] [CrossRef] [PubMed]
- Mauch, D.H.; Nägler, K.; Schumacher, S.; Göritz, C.; Müller, E.-C.; Otto, A.; Pfrieger, F.W. CNS Synaptogenesis Promoted by Glia-Derived Cholesterol. Science 2001, 294, 1354–1357. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Li, Y.; Yang, B.; Wang, H.; Lu, C.; Chang, A.K.; Huang, X.; Zhang, X.; Lu, Z.; Lu, X.; et al. Suppression of neuronal cholesterol biosynthesis impairs brain functions through insulin-like growth factor I-Akt signaling. Int. J. Biol. Sci. 2021, 17, 3702–3716. [Google Scholar] [CrossRef] [PubMed]
- Lentini, D.; Guzzi, F.; Pimpinelli, F.; Zaninetti, R.; Cassetti, A.; Coco, S.; Maggi, R.; Parenti, M. Polarization of caveolins and caveolae during migration of immortalized neurons. J. Neurochem. 2007, 104, 514–523. [Google Scholar] [CrossRef] [PubMed]
- Shin JW, Lee JC. Roles of microglial membranes in Alzheimer's disease. Curr Top Membr. 2020;86:301-314.
- Mattera, V.; Occhiuzzi, F.; Correale, J.; Pasquini, J.M. Remyelinating effect driven by transferrin-loaded extracellular vesicles. Glia 2023, 72, 338–361. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.W.; Hnasko, R.; Schubert, W.; Lisanti, M.P. Role of Caveolae and Caveolins in Health and Disease. Physiol. Rev. 2004, 84, 1341–1379. [Google Scholar] [CrossRef]
- Parton, R.G.; Simons, K. The multiple faces of caveolae. Nat. Rev. Mol. Cell Biol. 2007, 8, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Head, B.P.; Hu, Y.; Finley, J.C.; Saldana, M.D.; Bonds, J.A.; Miyanohara, A.; Niesman, I.R.; Ali, S.S.; Murray, F.; Insel, P.A.; et al. Neuron-targeted Caveolin-1 Protein Enhances Signaling and Promotes Arborization of Primary Neurons. J. Biol. Chem. 2011, 286, 33310–33321. [Google Scholar] [CrossRef] [PubMed]
- Mandyam, C.D.; Schilling, J.M.; Cui, W.; Egawa, J.; Niesman, I.R.; Kellerhals, S.E.; Staples, M.C.; Busija, A.R.; Risbrough, V.B.; Posadas, E.; et al. Neuron-Targeted Caveolin-1 Improves Molecular Signaling, Plasticity, and Behavior Dependent on the Hippocampus in Adult and Aged Mice. Biol. Psychiatry 2015, 81, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Francesconi, A.; Kumari, R.; Zukin, R.S. Regulation of Group I Metabotropic Glutamate Receptor Trafficking and Signaling by the Caveolar/Lipid Raft Pathway. J. Neurosci. 2009, 29, 3590–3602. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Yang, L.; Zhang, K.; Guo, Y.; Liu, S.; Wu, Y.; Li, X.; Song, Q.; Zhuo, M.; Zhao, M. Increased coupling of caveolin-1 and estrogen receptor α contributes to the fragile X syndrome. Ann. Neurol. 2015, 77, 618–636. [Google Scholar] [CrossRef] [PubMed]
- Luo L, et al. Caveolin-1-Mediated Cholesterol Accumulation Contributes to Exaggerated mGluR-Dependent Long-Term Depression and Impaired Cognition in Fmr1 Knockout Mice. Mol Neurobiol. 2023;60(6):3379-3395.
- Gaudreault, S.B.; Chabot, C.; Gratton, J.-P.; Poirier, J. The Caveolin Scaffolding Domain Modifies 2-Amino-3-hydroxy-5-methyl-4-isoxazole Propionate Receptor Binding Properties by Inhibiting Phospholipase A2 Activity. J. Biol. Chem. 2004, 279, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Ramsakha, N.; Ojha, P.; Pal, S.; Routh, S.; Citri, A.; Bhattacharyya, S. A vital role for PICK1 in the differential regulation of metabotropic glutamate receptor internalization and synaptic AMPA receptor endocytosis. J. Biol. Chem. 2023, 299, 104837. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.M.; Lisanti, M.P. The caveolin proteins. Genome Biol. 2004, 5, 214–214. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.M.; Bastiani, M.; Luetterforst, R.; Kirkham, M.; Kirkham, A.; Nixon, S.J.; Walser, P.; Abankwa, D.; Oorschot, V.M.; Martin, S.; et al. PTRF-Cavin, a Conserved Cytoplasmic Protein Required for Caveola Formation and Function. Cell 2008, 132, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Carman, C.V.; Lisanti, M.P.; Benovic, J.L. Regulation of G Protein-coupled Receptor Kinases by Caveolin. J. Biol. Chem. 1999, 274, 8858–8864. [Google Scholar] [CrossRef] [PubMed]
- Panetta, D.; Biedi, C.; Repetto, S.; Cordera, R.; Maggi, D. IGF-I regulates caveolin 1 and IRS1 interaction in caveolae. Biochem. Biophys. Res. Commun. 2004, 316, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Tang W, Li Y, Li Y, Wang Q. Caveolin-1, a novel player in cognitive decline. Neurosci Biobehav Rev. 2021;129:95-106.
- Wu, J.; Zhou, S.-L.; Pi, L.-H.; Shi, X.-J.; Ma, L.-R.; Chen, Z.; Qu, M.-L.; Li, X.; Nie, S.-D.; Liao, D.-F.; et al. High glucose induces formation of tau hyperphosphorylation via Cav-1-mTOR pathway: A potential molecular mechanism for diabetes-induced cognitive dysfunction. Oncotarget 2017, 8, 40843–40856. [Google Scholar] [CrossRef] [PubMed]
- Gupta A, Sharma A, Kumar A, Goyal R. Alteration in memory cognition due to activation of caveolin-1 and oxidative damage in a model of dementia of Alzheimer's type. Indian J Pharmacol. 2019;51(3):173-180.
- Ikezu, T.; Trapp, B.D.; Song, K.S.; Schlegel, A.; Lisanti, M.P.; Okamoto, T. Caveolae, Plasma Membrane Microdomains for α-Secretase-mediated Processing of the Amyloid Precursor Protein. J. Biol. Chem. 1998, 273, 10485–10495. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Singh, M.; Sharma, P.L. Ameliorative effect of daidzein: a caveolin-1 inhibitor in vascular endothelium dysfunction induced by ovariectomy. . 2012, 50, 28–34. [Google Scholar] [PubMed]
- Ajmani, P.; Yadav, H.N.; Singh, M.; Sharma, P.L. Possible involvement of caveolin in attenuation of cardioprotective effect of ischemic preconditioning in diabetic rat heart. BMC Cardiovasc. Disord. 2011, 11, 43–43. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Li, Y.; He, S.; Jiang, T.; Wang, N.; Du, M.; Cheng, B.; Gao, W.; Li, Y.; Wang, Q. Caveolin-1 Alleviates Diabetes-Associated Cognitive Dysfunction Through Modulating Neuronal Ferroptosis-Mediated Mitochondrial Homeostasis. Antioxidants Redox Signal. 2022, 37, 867–886. [Google Scholar] [CrossRef] [PubMed]
- Puddu, A.; Maggi, D. Emerging Role of Caveolin-1 in GLP-1 Action. Front. Endocrinol. 2021, 12. [Google Scholar] [CrossRef]
- Brubaker PL, Drucker DJ. Structure-function of the glucagon receptor family of G protein-coupled receptors: the glucagon, GIP, GLP-1, and GLP-2 receptors. Recept Channels. 2002;8(3-4):179–188.
- Syme, C.A.; Zhang, L.; Bisello, A. Caveolin-1 Regulates Cellular Trafficking and Function of the Glucagon-Like Peptide 1 Receptor. Mol. Endocrinol. 2006, 20, 3400–3411. [Google Scholar] [CrossRef] [PubMed]
- Del Galdo F, Lisanti MP, Jimenez SA. Caveolin-1, transforming growth factor-beta receptor internalization, and the pathogenesis of systemic sclerosis. Curr Opin Rheumatol. 2008;20(6):713–719.
- Hamoudane M, Maffioli S, Cordera R, Maggi D, Salani B. Caveolin-1 and polymerase I and transcript release factor: new players in insulin-like growth factor-I receptor signaling. J Endocrinol Invest. 2013;36(3):204–208.
- Lee H, et al. Caveolin-1 mutations (P132L and null) and the pathogenesis of breast cancer: caveolin-1 (P132L) behaves in a dominant-negative manner and caveolin-1 (-/-) null mice show mammary epithelial cell hyperplasia. Am J Pathol. 2002;161(4):1357–1369.
- Jones, B.; Buenaventura, T.; Kanda, N.; Chabosseau, P.; Owen, B.M.; Scott, R.; Goldin, R.; Angkathunyakul, N.; Jr, I.R.C.; Bosco, D.; et al. Targeting GLP-1 receptor trafficking to improve agonist efficacy. Nat. Commun. 2018, 9, 1–17. [Google Scholar] [CrossRef]
- Thompson A, Kanamarlapudi V. Agonist-induced internalisation of the glucagon-like peptide-1 receptor is mediated by the Galphaq pathway. Biochem Pharmacol. 2015;93(1):72–84.
- Kutlu MD, Kose S, Akillioglu K.GLP-1 agonist Liraglutide prevents MK-801-induced schizophrenia-like behaviors and BDNF, CREB, p-CREB, Trk-B expressions in the hippocampus and prefrontal cortex in Balb/c mice. Behav Brain Res. 2023;445:114386.
- Abubakar MD, et al. GLP-1/GIP Agonist as an Intriguing and Ultimate Remedy for Combating Alzheimer's Disease through its Supporting DPP4 Inhibitors: A Review. Curr Top Med Chem. 2024.
- Ikeda, Y.; Nagase, N.; Tsuji, A.; Kitagishi, Y.; Matsuda, S. Neuroprotection by dipeptidyl-peptidase-4 inhibitors and glucagon-like peptide-1 analogs via the modulation of AKT-signaling pathway in Alzheimer’s disease. World J. Biol. Chem. 2021, 12, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Hsu, T.M.; E Noble, E.; Liu, C.M.; Cortella, A.M.; Konanur, V.R.; Suarez, A.N.; Reiner, D.J.; Hahn, J.D.; Hayes, M.R.; E Kanoski, S. A hippocampus to prefrontal cortex neural pathway inhibits food motivation through glucagon-like peptide-1 signaling. Mol. Psychiatry 2017, 23, 1555–1565. [Google Scholar] [CrossRef] [PubMed]
- Abraham WC, Jones OD, Glanzman DL. Is plasticity of synapses the mechanism of long-term memory storage? NPJ Sci Learn. 2019;4:9.
- Li, W.; Kutas, M.; Gray, J.A.; Hagerman, R.H.; Olichney, J.M. The Role of Glutamate in Language and Language Disorders - Evidence from ERP and Pharmacologic Studies. Neurosci. Biobehav. Rev. 2020, 119, 217–241. [Google Scholar] [CrossRef]
- Wu E, Zhang J, Zhang J, Zhu S. Structural Insights into Gating Mechanism and Allosteric Regulation of NMDA Receptors. Curr. Opin. Neurobiol. 2023;83:102806.
- Tsai, Y.-C.; Huang, S.-M.; Peng, H.-H.; Lin, S.-R.; Chin, T.-Y.; Huang, S.-M. Imbalance of synaptic and extrasynaptic NMDA receptors induced by the deletion of CRMP1 accelerates age-related cognitive decline in mice. Neurobiol. Aging 2024, 135, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Tajima, N. Structural insights into NMDA receptor pharmacology. Biochem. Soc. Trans. 2023, 51, 1713–1731. [Google Scholar] [CrossRef] [PubMed]
- Mony, L.; Paoletti, P. Mechanisms of NMDA receptor regulation. Curr. Opin. Neurobiol. 2023, 83, 102815. [Google Scholar] [CrossRef]
- Morris, P.G.; Mishina, M.; Jones, S. Altered Synaptic and Extrasynaptic NMDA Receptor Properties in Substantia Nigra Dopaminergic Neurons From Mice Lacking the GluN2D Subunit. Front. Cell. Neurosci. 2018, 12, 354. [Google Scholar] [CrossRef]
- Olivero, G.; Grilli, M.; Marchi, M.; Pittaluga, A. Metamodulation of presynaptic NMDA receptors: New perspectives for pharmacological interventions. Neuropharmacology 2023, 234, 109570. [Google Scholar] [CrossRef] [PubMed]
- Perluigi, M.; Di Domenico, F.; Barone, E.; Butterfield, D. mTOR in Alzheimer disease and its earlier stages: Links to oxidative damage in the progression of this dementing disorder. Free. Radic. Biol. Med. 2021, 169, 382–396. [Google Scholar] [CrossRef]
- Su, H.; Wang, X. Autophagy and p62 in cardiac protein quality control. Autophagy 2011, 7, 1382–1383. [Google Scholar] [CrossRef]
- Mary, A.; Eysert, F.; Checler, F.; Chami, M. Mitophagy in Alzheimer’s disease: Molecular defects and therapeutic approaches. Mol. Psychiatry 2022, 28, 202–216. [Google Scholar] [CrossRef] [PubMed]
- Babygirija, R.; Sonsalla, M.M.; Mill, J.; James, I.; Han, J.H.; Green, C.L.; Calubag, M.F.; Wade, G.; Tobon, A.; Michael, J.; et al. Protein restriction slows the development and progression of pathology in a mouse model of Alzheimer’s disease. Nat. Commun. 2024, 15, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Mustapha, M.; Ibrahim, N.; Yap, K.H.; Azmin, S.; Makpol, S.; Damanhuri, H.; Hamzah, J. Profiling neuroprotective potential of trehalose in animal models of neurodegenerative diseases: a systematic review. Neural Regen. Res. 2023, 18, 1179–1185. [Google Scholar] [CrossRef]
- Rusmini, P.; Cortese, K.; Crippa, V.; Cristofani, R.; Cicardi, M.E.; Ferrari, V.; Vezzoli, G.; Tedesco, B.; Meroni, M.; Messi, E.; et al. Trehalose induces autophagy via lysosomal-mediated TFEB activation in models of motoneuron degeneration. Autophagy 2018, 15, 631–651. [Google Scholar] [CrossRef] [PubMed]
- Khalifeh, M.; Read, M.I.; Barreto, G.E.; Sahebkar, A. Trehalose against Alzheimer's Disease: Insights into a Potential Therapy. BioEssays 2020, 42, e1900195. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Barkhordarian, H.; Emadi, S.; Park, C.B.; Sierks, M.R. Trehalose differentially inhibits aggregation and neurotoxicity of beta-amyloid 40 and 42. Neurobiol. Dis. 2005, 20, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Tien, N.T.; Karaca, I.; Tamboli, I.Y.; Walter, J. Trehalose Alters Subcellular Trafficking and the Metabolism of the Alzheimer-associated Amyloid Precursor Protein. J. Biol. Chem. 2016, 291, 10528–10540. [Google Scholar] [CrossRef] [PubMed]
- Pupyshev AB, et al. Combined induction of mTOR-dependent and mTOR-independent pathways of autophagy activation as an experimental therapy for Alzheimer’s disease-like pathology in a mouse model. Pharmacol. Biochem. Behav. 2022;217:173406.
- Pupyshev, A.B.; Akopyan, A.A.; Tenditnik, M.V.; Ovsyukova, M.V.; Dubrovina, N.I.; Belichenko, V.M.; Korolenko, T.A.; Zozulya, S.A.; Klyushnik, T.P.; Tikhonova, M.A. Alimentary Treatment with Trehalose in a Pharmacological Model of Alzheimer’s Disease in Mice: Effects of Different Dosages and Treatment Regimens. Pharmaceutics 2024, 16, 813. [Google Scholar] [CrossRef] [PubMed]
- Pi, H.; Li, M.; Tian, L.; Yang, Z.; Yu, Z.; Zhou, Z. Enhancing lysosomal biogenesis and autophagic flux by activating the transcription factor EB protects against cadmium-induced neurotoxicity. Sci. Rep. 2017, 7, srep43466. [Google Scholar] [CrossRef]
- Querfurth, H.; Lee, H.-K. Mammalian/mechanistic target of rapamycin (mTOR) complexes in neurodegeneration. Mol. Neurodegener. 2021, 16, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Złotek M, Kurowska A, Herbet M, Piątkowska-Chmiel I. GLP-1 analogs, SGLT-2, and DPP-4 inhibitors: a triad of Hope for Alzheimer's disease therapy. Biomedicine. 2023;11(11):3035.
- Xia W, Yu H, Wen P. Meta-analysis on GLP-1 mediated modulation of autophagy in islet beta-cells: Prospectus for improved wound healing in type 2 diabetes. Int Wound J. 2024;21(4):e14841.
- Shankar, G.M.; Bloodgood, B.L.; Townsend, M.; Walsh, D.M.; Selkoe, D.J.; Sabatini, B.L. Natural Oligomers of the Alzheimer Amyloid-β Protein Induce Reversible Synapse Loss by Modulating an NMDA-Type Glutamate Receptor-Dependent Signaling Pathway. J. Neurosci. 2007, 27, 2866–2875. [Google Scholar] [CrossRef] [PubMed]
- Skeberdis, V.A.; Chevaleyre, V.; Lau, C.G.; Goldberg, J.H.; Pettit, D.L.; O Suadicani, S.; Lin, Y.; Bennett, M.V.L.; Yuste, R.; E Castillo, P.; et al. Protein kinase A regulates calcium permeability of NMDA receptors. Nat. Neurosci. 2006, 9, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jiang, B.; Luo, W. Memantine ameliorates oxaliplatin-induced neurotoxicity via mitochondrial protection. Bioengineered 2022, 13, 6688–6697. [Google Scholar] [CrossRef] [PubMed]
- Lee JR, Jeong KW. N-retinylidene-N-retinylethanolamine degradation in human retinal pigment epithelial cells via memantine- and ifenprodil-mediated autophagy. Korean J Physiol Pharmacol. 2023;27(5):449-456.
- Johnson, J.W.; Kotermanski, S. Mechanism of action of memantine. Curr. Opin. Pharmacol. 2005, 6, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Winkler D, Leyhe T. [Alzheimer's disease - State of the art, and emerging diagnostics and therapeutics]. Ther Umsch. 2018;75:432–437.
- Companys-Alemany J, et al. NMDA receptor antagonists reduce amyloid-β deposition by modulating calpain-1 signaling and autophagy, rescuing cognitive impairment in 5XFAD mice. Cell Mol Life Sci. 2022;79:408.
- Song, X.; Jensen, M.; Jogini, V.; Stein, R.A.; Lee, C.-H.; Mchaourab, H.S.; Shaw, D.E.; Gouaux, E. Mechanism of NMDA receptor channel block by MK-801 and memantine. Nature 2018, 556, 515–519. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Reddy, P.H. Role of Glutamate and NMDA Receptors in Alzheimer’s Disease. J. Alzheimer's Dis. 2017, 57, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Harris, L.D.; Regan, M.C.; Myers, S.J.; Nocilla, K.A.; Akins, N.S.; Tahirovic, Y.A.; Wilson, L.J.; Dingledine, R.; Furukawa, H.; Traynelis, S.F.; et al. Novel GluN2B-Selective NMDA Receptor Negative Allosteric Modulator Possesses Intrinsic Analgesic Properties and Enhances Analgesia of Morphine in a Rodent Tail Flick Pain Model. ACS Chem. Neurosci. 2023, 14, 917–935. [Google Scholar] [CrossRef] [PubMed]
- Lavretsky, H.; Laird, K.T.; Krause-Sorio, B.; Heimberg, B.F.; Yeargin, J.; Grzenda, A.; Wu, P.; Thana-Udom, K.; Ercoli, L.M.; Siddarth, P. A Randomized Double-Blind Placebo-Controlled Trial of Combined Escitalopram and Memantine for Older Adults With Major Depression and Subjective Memory Complaints. Am. J. Geriatr. Psychiatry 2019, 28, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.P.; Sawant, N.; Morton, H.; Kshirsagar, S.; E Bunquin, L.; Yin, X.; Reddy, P.H. Selective serotonin reuptake inhibitor citalopram ameliorates cognitive decline and protects against amyloid beta-induced mitochondrial dynamics, biogenesis, autophagy, mitophagy and synaptic toxicities in a mouse model of Alzheimer’s disease. Hum. Mol. Genet. 2021, 30, 789–810. [Google Scholar] [CrossRef] [PubMed]
- Koychev, I.; I Adler, A.; Edison, P.; Tom, B.; E Milton, J.; Butchart, J.; Hampshire, A.; Marshall, C.; Coulthard, E.; Zetterberg, H.; et al. Protocol for a double-blind placebo-controlled randomised controlled trial assessing the impact of oral semaglutide in amyloid positivity (ISAP) in community dwelling UK adults. BMJ Open 2024, 14, e081401. [Google Scholar] [CrossRef] [PubMed]
- Carr, R.D.; Larsen, M.O.; Jelic, K.; Lindgren, O.; Vikman, J.; Holst, J.J.; Deacon, C.F.; Ahrén, B. Secretion and Dipeptidyl Peptidase-4-Mediated Metabolism of Incretin Hormones after a Mixed Meal or Glucose Ingestion in Obese Compared to Lean, Nondiabetic Men. J. Clin. Endocrinol. Metab. 2010, 95, 872–878. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Yan, R.; Zhang, C.; Bai, X.; Yang, X.; Yang, Y.; Feng, T.; Liu, X. Dipeptidyl peptidase-4 inhibitors alleviate cognitive dysfunction in type 2 diabetes mellitus. Lipids Heal. Dis. 2023, 22, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pekar T, et al. The positive effect of spermidine in older adults suffering from dementia : First results of a 3-month trial. Wien Klin Wochenschr. 2021;133(9-10):484-491.
- Rosli, H.; Shahar, S.; Rajab, N.F.; Din, N.C.; Haron, H. The effects of polyphenols-rich tropical fruit juice on cognitive function and metabolomics profile – a randomized controlled trial in middle-aged women. Nutr. Neurosci. 2021, 25, 1577–1593. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Lopez, N.; Tarabra, E.; Toledo, M.; Garcia-Macia, M.; Sahu, S.; Coletto, L.; Batista-Gonzalez, A.; Barzilai, N.; Pessin, J.E.; Schwartz, G.J.; et al. System-wide Benefits of Intermeal Fasting by Autophagy. Cell Metab. 2017, 26, 856–871. [Google Scholar] [CrossRef] [PubMed]
- Wells, R.G.; Neilson, L.E.; McHill, A.W.; Hiller, A.L. Dietary fasting and time-restricted eating in Huntington’s disease: therapeutic potential and underlying mechanisms. Transl. Neurodegener. 2024, 13, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Peng, J.; Tang, W.; Xia, Y.; Song, P. A circadian rhythm-restricted diet regulates autophagy to improve cognitive function and prolong lifespan. Biosci. Trends 2023, 17, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Benavides-Rivas, C.; Tovar, L.M.; Zúñiga, N.; Pinto-Borguero, I.; Retamal, C.; Yévenes, G.E.; Moraga-Cid, G.; Fuentealba, J.; Guzmán, L.; Coddou, C.; et al. Altered Glutaminase 1 Activity During Neurulation and Its Potential Implications in Neural Tube Defects. Front. Pharmacol. 2020, 11, 900. [Google Scholar] [CrossRef] [PubMed]
- Petersen J, et al. GLP-1 directed NMDA receptor antagonism for obesity treatment. Nature. 2024;629(8014):1133-1141.
- Mishra, D.; Richard, J.E.; Maric, I.; Shevchouk, O.T.; Börchers, S.; Eerola, K.; Krieger, J.-P.; Skibicka, K.P.; B, S. Lateral parabrachial nucleus astrocytes control food intake. Front. Endocrinol. 2024, 15, 1389589. [Google Scholar] [CrossRef] [PubMed]
- Caruso Bavisotto C, et al. Extracellular vesicle-mediated cell⁻cell communication in the nervous system: focus on neurological diseases. Int J Mol Sci. 2019;20(2):434.
- Mendes-Silva, A.P.; Nikolova, Y.S.; Rajji, T.K.; Kennedy, J.L.; Diniz, B.S.; Gonçalves, V.F.; Vieira, E.L. Exosome-associated mitochondrial DNA in late-life depression: Implications for cognitive decline in older adults. J. Affect. Disord. 2024, 362, 217–224. [Google Scholar] [CrossRef]
- Sun, M.; Chen, Z. Unveiling the Complex Role of Exosomes in Alzheimer’s Disease. J. Inflamm. Res. 2024, ume 17, 3921–3948. [Google Scholar] [CrossRef]
- Ebrahim N, et al. Exploring the molecular mechanisms of MSC-derived exosomes in Alzheimer's disease: Autophagy, insulin and the PI3K/Akt/mTOR signaling pathway. Biomed Pharmacother. 2024;176:116836.
- Micci, M.-A.; Krishnan, B.; Bishop, E.; Zhang, W.-R.; Guptarak, J.; Grant, A.; Zolochevska, O.; Tumurbaatar, B.; Franklin, W.; Marino, C.; et al. Hippocampal stem cells promotes synaptic resistance to the dysfunctional impact of amyloid beta oligomers via secreted exosomes. Mol. Neurodegener. 2019, 14, 25. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
An illustrative representation of caveolae associated with caveolin proteins. Caveolae may be a kind of platforms for various signaling molecules including GLP-1 and NMDA receptors as well as even for the development of certain exosomes, which may be interrelated to the scaffold domain of caveolins. Note that some critical pathways for the development of various disease-related signaling have been omitted for clarity. Abbreviation: GLP-1, glucagon-like peptide-1; NMDA, N-methyl-d-aspartate.
Figure 1.
An illustrative representation of caveolae associated with caveolin proteins. Caveolae may be a kind of platforms for various signaling molecules including GLP-1 and NMDA receptors as well as even for the development of certain exosomes, which may be interrelated to the scaffold domain of caveolins. Note that some critical pathways for the development of various disease-related signaling have been omitted for clarity. Abbreviation: GLP-1, glucagon-like peptide-1; NMDA, N-methyl-d-aspartate.
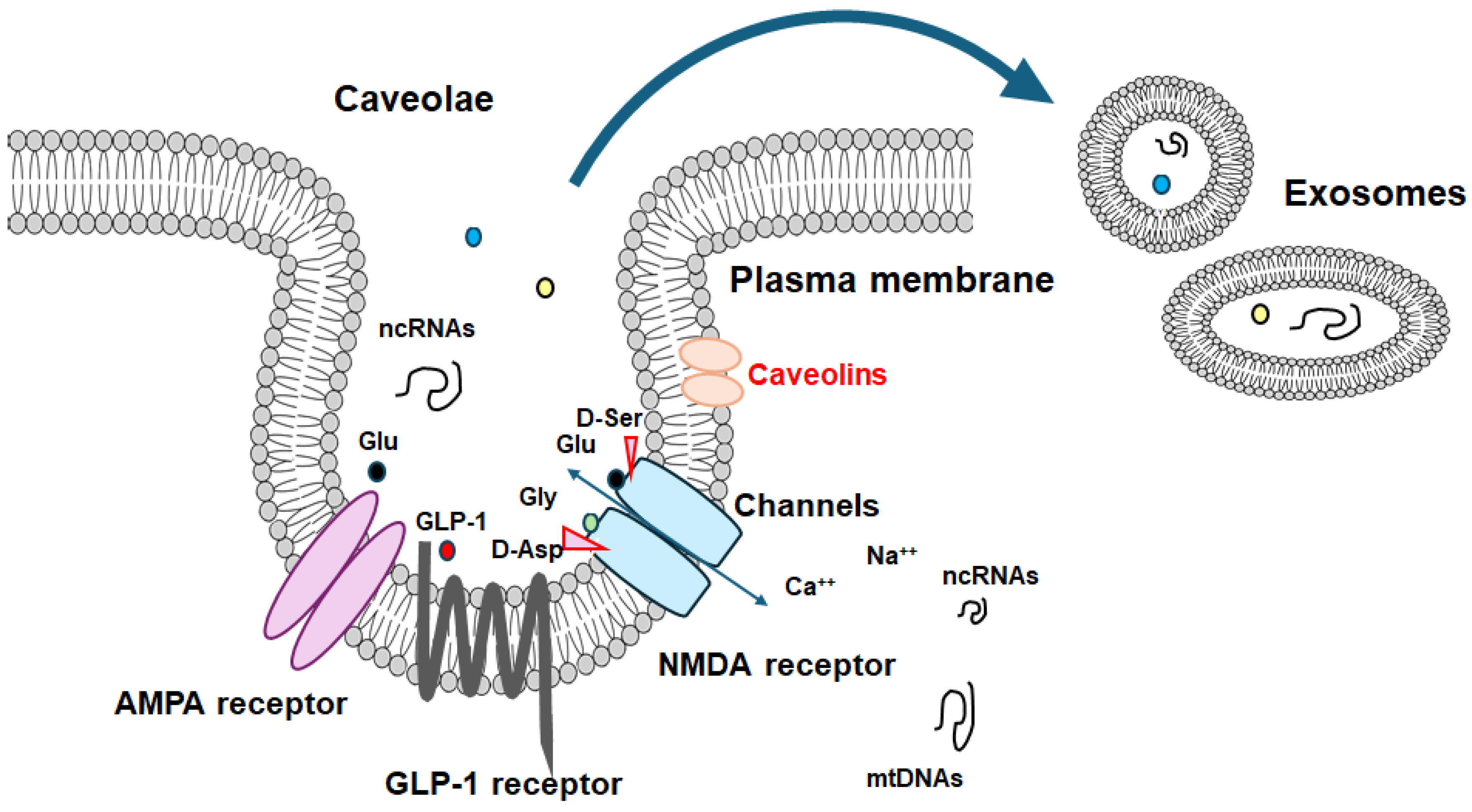
Figure 2.
Several modulator molecules linked to the signaling of GLP-1 receptor within the PI3K/AKT/mTOR pathway are demonstrated. Example molecules known to act on an AMPK/mTOR pathway are also shown. The activation of GLP-1 receptor results in the stimulation of adenylyl cyclase (AC) that transforms adenosine triphosphate (ATP) into 3 -5 -cyclic adenosine monophosphate (cAMP), which then activates the cAMP-dependent protein kinase (PKA) as well as the following AMPK for the modification of autophagy/mitophagy. Some example factors including fasting, starvation, and/or metformin known to act on the autophagy/mitophagy signaling are also shown. Note that some critical pathway such as for insulin-induction has been omitted for clarity. Arrowhead means stimulation whereas hammerhead represents inhibition. Abbreviation: GLP-1, glucagon-like peptide-1; AMPK, adenosine monophosphate-activated protein kinase; mTOR, mammalian/mechanistic target of rapamycin; PI3K, phosphoinositide-3 kinase; PKA, protein kinase A; PTEN, phosphatase and tensin homologue deleted on chromosome 10.
Figure 2.
Several modulator molecules linked to the signaling of GLP-1 receptor within the PI3K/AKT/mTOR pathway are demonstrated. Example molecules known to act on an AMPK/mTOR pathway are also shown. The activation of GLP-1 receptor results in the stimulation of adenylyl cyclase (AC) that transforms adenosine triphosphate (ATP) into 3 -5 -cyclic adenosine monophosphate (cAMP), which then activates the cAMP-dependent protein kinase (PKA) as well as the following AMPK for the modification of autophagy/mitophagy. Some example factors including fasting, starvation, and/or metformin known to act on the autophagy/mitophagy signaling are also shown. Note that some critical pathway such as for insulin-induction has been omitted for clarity. Arrowhead means stimulation whereas hammerhead represents inhibition. Abbreviation: GLP-1, glucagon-like peptide-1; AMPK, adenosine monophosphate-activated protein kinase; mTOR, mammalian/mechanistic target of rapamycin; PI3K, phosphoinositide-3 kinase; PKA, protein kinase A; PTEN, phosphatase and tensin homologue deleted on chromosome 10.
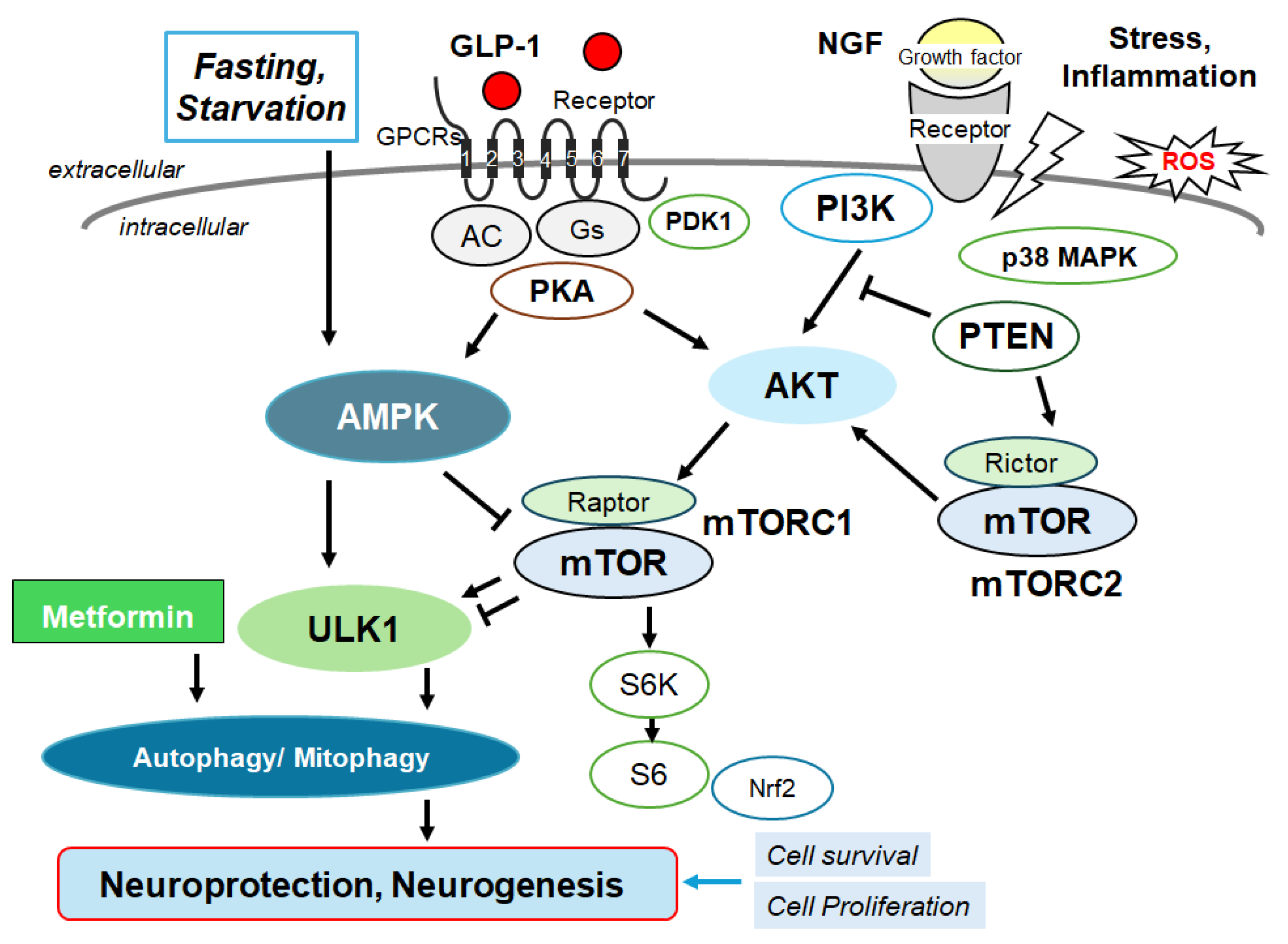
Figure 3.
Schematic depiction of NMDA receptor signaling pathway along with a GTP-binding protein-coupled receptor (GPCR) signaling pathway for autophagy/mitophagy. NMDA receptors may require the binding of several amino acid molecules of glycine, glutamate, and/or aspartate, which could modulate the receptor function. For example, glutamate is in the glutamate-binding site and glycine is in the glycine-binding site within the NMDA receptor subunit. The stimulation of NMDA receptors may allow Ca2+ entry into the cytoplasm of neurons, which may contribute to long-term potentiation (LTP) or long-term depression (LTD) as well as to a phase of excitotoxicity and/or apoptosis. Stimulatory effects are indicated with arrows.
Figure 3.
Schematic depiction of NMDA receptor signaling pathway along with a GTP-binding protein-coupled receptor (GPCR) signaling pathway for autophagy/mitophagy. NMDA receptors may require the binding of several amino acid molecules of glycine, glutamate, and/or aspartate, which could modulate the receptor function. For example, glutamate is in the glutamate-binding site and glycine is in the glycine-binding site within the NMDA receptor subunit. The stimulation of NMDA receptors may allow Ca2+ entry into the cytoplasm of neurons, which may contribute to long-term potentiation (LTP) or long-term depression (LTD) as well as to a phase of excitotoxicity and/or apoptosis. Stimulatory effects are indicated with arrows.
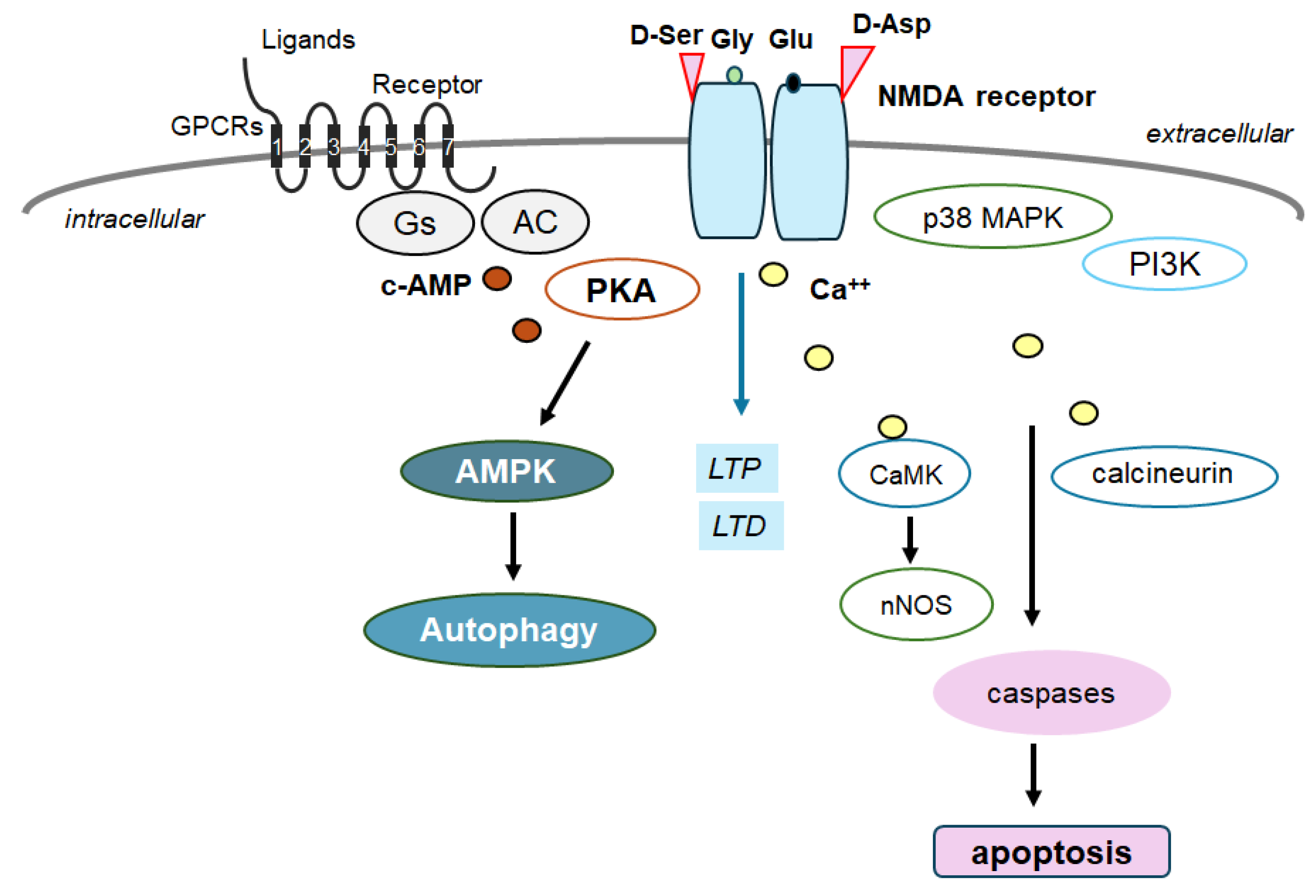
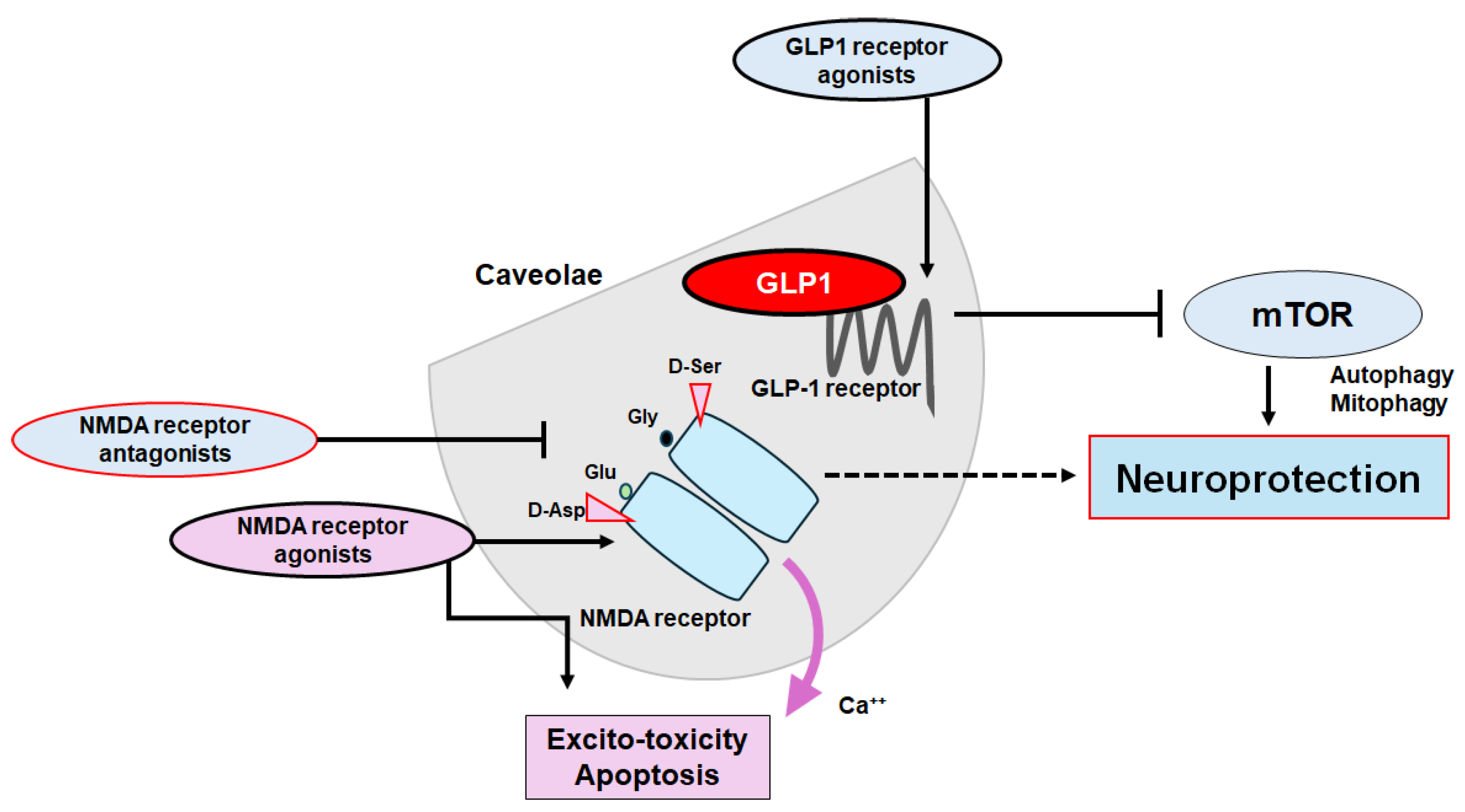
Figure 4.
The plausible tactics for the neuroprotection and/or against the cognitive impairment have been shown. The ideal approach might be to utilize agonists of GLP-1 receptors and/or antagonists of NMDA receptors to halt the neuron-damaging process for the neuroprotection. Note that several important activities such as inflammatory reaction, autophagy initiation, and reactive oxygen species (ROS) production have been omitted for clarity. Stimulatory effects are indicated with arrows; inhibitory effects with a line ending in a hammerhead. Broken lines indicate possible mechanism of action. “?” means for author speculation.
Figure 4.
The plausible tactics for the neuroprotection and/or against the cognitive impairment have been shown. The ideal approach might be to utilize agonists of GLP-1 receptors and/or antagonists of NMDA receptors to halt the neuron-damaging process for the neuroprotection. Note that several important activities such as inflammatory reaction, autophagy initiation, and reactive oxygen species (ROS) production have been omitted for clarity. Stimulatory effects are indicated with arrows; inhibitory effects with a line ending in a hammerhead. Broken lines indicate possible mechanism of action. “?” means for author speculation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



