
Submitted:
17 July 2024
Posted:
18 July 2024
You are already at the latest version
Abstract
Cell aging is associated with the increased production of mitochondrial reactive oxygen species (mtROS) due to mitochondrial dysfunction, and various phytonutrients and estrogens have been shown to improve skin health. Thus, the aim of the current study was to examine damage to dermal fibroblasts by chemically-induced mitochondrial dysfunction and to study the mechanism of the protective effects of carotenoids, polyphenols, and estradiol. Rotenone, a Complex I inhibitor, caused mitochondrial dysfunction in human dermal fibroblasts, substantially reducing respiration and ATP levels, followed by increased mitochondrial and cytosolic ROS, which resulted in apoptotic cell death, increased matrix metalloproteinase-1 (MMP1) secretion, and decreased collagen secretion. Pre-treatment with carotenoid-rich tomato extracts, rosemary extract, and estradiol reversed these effects. These protective effects can be partially explained by a cooperative activation of antioxidant response element (ARE/Nrf2) transcriptional activity by the protective compounds and rotenone, which led to upregulation of antioxidant proteins such as NQO1. To determine if ARE/Nrf2 activity is crucial for cell protection, we inhibited it using the Nrf2 inhibitors ML385 and ochratoxin A. This inhibition markedly reduced the protective effects of the test compounds by diminishing their effect to reduce cytosolic ROS. Our study results indicate that phytonutrients and estradiol protect skin cells from damage caused by mitochondria-generated ROS and, thus, may delay skin aging and improve skin health.
Keywords:
mitochondrial dysfunction
; dermal fibroblasts
; rotenone
; reactive oxygen species (ROS)
; matrix metalloproteinase (MMP)
; collagen
; antioxidant response element/Nrf2 (ARE/Nrf2)
; tomato extract
; rosemary extract
; estradiol
1. Introduction
Reactive oxygen species (ROS) play an essential role in regulating various cell functions and biochemical processes. Increased levels of ROS can be generated from environmental sources such as exposure to UV radiation and air pollutants [1], as well as from endogenous sources such as mitochondrial respiration and NADPH oxidases [2]. Oxidative stress, defined as an imbalance between ROS production and antioxidant activity [3], induces damage to nucleic acids, proteins, and lipids and the modulation of signaling pathways [4], activating inflammatory processes that may lead to harmful processes such as neurodegeneration [5] and carcinogenesis [6], and to accelerated aging [7,8].
Mitochondrial respiration and oxidative phosphorylation, which is the main source of ATP in aerobic cells, is a major source of ROS [9]. During energy production through the electron transport chain, some electrons “leak” to oxygen [10], forming superoxide radicals (O2-.), primarily from Complexes I and III [10]. Superoxide radicals are converted to hydrogen peroxide (H2O2) by superoxide dismutases in and out of the mitochondria, and both superoxide and hydrogen peroxide can exit the mitochondria and increase cytosolic ROS [11]. Mitochondrial dysfunction and reduced electron transport activity leads to decreased ATP production and increased production of superoxide radicals [12], which are involved in the pathophysiology of diseases including Parkinson’s [13], as well as in the aging process [12], as suggested by Harman's free radical theory of aging [14].
Mitochondrial dysfunction is involved in both chronological and photo-aging of the skin [15]. Aged skin is characterized by mutations of mitochondrial DNA (mtDNA), damaged mitochondria, and oxidative stress in both the dermal and epidermal layers [16]. Oxidative stress causes skin damage due to stimulation of inflammatory processes [17], induction of cellular apoptosis [18], increased expression of metalloproteinases (MMPs) which degrade extracellular matrix proteins [19,20], and reduced collagen synthesis [20], resulting in decreased skin elasticity and wrinkles. By increasing cellular antioxidant activities, oxidative stress should be lowered, thus, reducing skin damage and skin aging.
Epidemiological and clinical studies have shown that a diet rich in fruits and vegetables promotes skin health [21]. The phytonutrients of carotenoids and polyphenols were found to reduce skin damage [22,23] and to protect dermal fibroblasts by eliminating oxidative stress [24]. It has been suggested that this reduction of ROS is due to activation of the antioxidant defense mechanism [25]. Previous studies demonstrated that carotenoids [26,27] and polyphenols [28,29] activate the antioxidant response element/Nrf2 (ARE/Nrf2) transcription system in several cell types. Additional mechanisms for the protective effects of phytonutrients involve the modulation of signaling pathways which lead to skin damage [30]. Several studies suggest that phytonutrients modulate these signaling pathways, resulting in reduced skin cell death [31] and inflammation [28], and in changes in MMPs and collagen levels [24,30].
In menopause, when estradiol levels decrease, skin health is impaired [32]. This impairment can be partially overcome by estrogens, which are known to increase collagen and elastin levels and reduce MMPs that lead to enhanced skin elasticity [32,33]. Estrogens also increase skin cell viability and proliferation, leading to enhanced skin thickness [33]. These estrogen effects may be related to a reduction of ROS [32], which can result from estrogen-induced expression of antioxidant enzymes [34] or by indirect activation of the ARE/Nrf2 transcription system [35].
The aim of the current study was to investigate the effect of endogenous oxidative stress in primary human dermal fibroblasts that results from rotenone-induced mitochondrial dysfunction, and to examine the protective role of carotenoids, polyphenols, and estradiol. Rotenone is a mitochondrial Complex I inhibitor that disrupts the electron transport chain and reduces mitochondrial respiration, thereby increasing mitochondrial oxidative stress [36]. The effect of rotenone and the treatment compounds was examined on apoptotic cell death, MMP-1 and pro-collagen 1a1 secretions, mitochondrial and cytosolic ROS levels, and mitochondrial function. The activation of the ARE/Nrf2 transcription system by rotenone and by dietary compounds and estradiol, and its inhibition by specific inhibitors were examined to suggest the role this system has in protecting dermal fibroblasts from the damage caused by mitochondria-generated ROS (mtROS).
2. Materials and Methods
2.1. Materials
Tomato extracts (LycomatoTM) and rosemary extract were a gift of Lycored Ltd., Beer Sheva, Israel). The red tomato extract, prepared by ethyl acetate extraction of tomato pulp, contained 6% lycopene, other tomato carotenoids (phytoene and phytofluene above 1%, beta-carotene above 0.2%), and additional fat-soluble tomato components such as natural tocopherols (above 1.5%) and phytosterols (1.1–2.5%). The remainder were triacylglycerols (70–72%), monoacylglycerols (8–9%), and phospholipids (7–8%). Golden tomato extract contained phytoene 6.7%, phytofluene 1.9%, zeta-carotene 2.0%, β-carotene 0.14%, lycopene 0.1%, tocopherols 3.1%, and phytosterols 1.0%. The remainder were triacylglycerols (71%), monoacylglycerols (8%), and phospholipids (6%). Carotenoids were dissolved in tetrahydrofuran (THF, containing 0.025% butylated hydroxytoluene as an antioxidant; Sigma-Aldrich) and solubilized in cell culture medium, and their final concentration was measured as described previously [27,37,38]. Rosemary extract was prepared by extraction with 80% ethanol. Its composition was only partially determined to contain carnosic acid (20.2%) and carnosol (2.5%) as the main polyphenols. 17β-estradiol and rotenone were purchased from Sigma-Aldrich (Rehovot, Israel). The rosemary extract and estradiol were dissolved in ethanol, and rotenone was dissolved in DMSO. Hanks’ solution, and 1M HEPES buffer were purchased from Biological Industries (Beth Haemek, Israel). Dulbecco’s modified Eagle’s medium (DMEM) and dextran-coated charcoal-treated fetal bovine serum (DCC-FBS) were acquired from Capricorn Scientific (Ebsdorfergrund, Germany).
2.2. Cell Culture
Normal human dermal fibroblasts (NHDFs) were purchased from PromoCell GmbH (Heidelberg, Germany). The cells were grown in PromoCell fibroblast growth medium 2, according to the manufacturer’s instructions, in a humidified atmosphere at 37°C in 5% CO2. Before each experiment, cells were depleted of steroid hormones by maintaining them for 3–5 days in phenol red-free DMEM, supplemented with 10% DCC-FBS. This medium was used throughout all of the experiments because it does not contain steroid hormones or any other compound with estrogenic activity, such as phenol red.
2.3. Determination of Cell Number, and Secretion of MMP-1 and Procollagen 1a1
NHDF cells were seeded in 96-well plates at a density of 5x103 cells/well in DMEM-DCC-FBS medium. 24 h later, cells were pre-incubated with phytonutrients or estradiol for another 24 h. Vehicle-treated control cells were incubated with the relevant amounts of solvents used in a particular experiment, which had no effect on the measured parameters. The medium was then replaced with one containing the treatment compounds, with or without rotenone, and incubated for an additional 48 h. Thereafter, the medium was removed and frozen for analysis of secreted proteins, and the cell number was determined by a crystal violet assay (Sigma-Aldrich, Rehovot, Israel), according to the manufacturer’s instructions. MMP-1 and pro-collagen 1a1 protein levels in cell culture supernatants were quantified by ELISA using Human Total MMP-1 DuoSet and Human Pro-Collagen 1a1 DuoSet ELISA kits (R&D Systems, Inc., Minneapolis, MN, USA), according to the manufacturer’s instructions. Optical density was measured using a VERSAmax tunable microplate reader (Molecular Devices, Menlo Park, CA, USA). Results of the cell number, MMP-1, and pro-collagen measurements were calculated as a percentage of the values obtained in control cells, which were treated with a vehicle without rotenone.
2.4. Assessment of Apoptosis
Cells were seeded in 6-well plates at 3 × 105 cells/well. 24 h later, they were pre-incubated with the phytonutrients or estradiol for 24 h. The medium was then replaced with one containing the treatment compounds, with or without rotenone, and incubated for an additional 48 h. Cells were washed in PBS and stained with annexin V using an Annexin V-FITC/7-AAD Apoptosis Detection Kit (Biogems, Cat# 62700-50, Chai Wan, Hong Kong), according to the manufacturer’s protocol. The percentages of apoptotic cells were determined by flow cytometry (Beckman Coulter Inc.). For each analysis, 10,000 events were recorded, and the data were processed using Kaluza software, version 2.1 (Beckman Coulter). As a positive control, cells were incubated with 1.25 µM staurosporin for 24 h. Annexin V-positive cells were considered to be in the apoptotic phase.
2.5. ApoLive-Glo Multiplex Assay for Cell Viability and Apoptosis
Because rotenone is known to affect mitochondrial respiration, we confirmed the results from a crystal violet assay (Sigma-Aldrich) using a method not based on mitochondrial activity. To this end, we used an ApoLive-Glo multiplex assay kit (Promega, Madison, WI) according to the manufacturer’s instructions. The level of the viable cells was measured using a fluorogenic substrate. Furthermore, the apoptosis results obtained by annexin V staining were confirmed by measuring caspase-3/7 activation, a key biomarker of apoptosis, using a luminogenic caspase-3/7 substrate. Both cell viability and caspase-3/7 activity were detected using a SpectraMax Paradigm plate reader (Molecular Devices Co., Sunnyvale, CA, USA).
2.6. Determination of Cytosolic Levels of Reactive Oxygen Species
Cytosolic ROS levels were determined by 2',7'-dichlorofluorescin diacetate staining (DCFH-DA, Sigma-Aldrich). Intracellular ROS oxidizes this probe to a highly fluorescent compound, DCF. Cells were seeded in 96-well plates at 104 cells/well. 24 h later, they were pre-incubated with the phytonutrients or estradiol for another 24 h. The medium was then replaced with one containing the treatment compounds, with or without rotenone, and incubated for an additional 4 h. Cells were washed with Hanks’ solution containing 10 mM HEPES buffer, pH = 7.4 and, subsequently, stained with 40 μM DCFH-DA for 30 min at 37°C in the dark. Fluorescence was detected using a SpectraMax Paradigm plate reader (Molecular Devices Co.). As a positive control, DCFH-DA-loaded cells were treated with 0.5 mM H2O2 for 10 min. Untreated and unstained cells served as a negative control.
2.7. Determination of Mitochondrial Levels of Reactive Oxygen Species
Mitochondrial ROS levels were stained with MitoSOX Red superoxide indicators. Oxidation of the MitoSOX reagent by mitochondrial superoxide produces red fluorescence. Cells were seeded in 96-well plates at 104 cells/well. 24 h later, they were pre-incubated with the phytonutrients or estradiol for 24 h. The medium was then replaced with one containing the treatment compounds, with or without rotenone, and incubated for an additional 90 min. Cells were washed with Hanks’ solution containing 10 mM HEPES buffer, pH = 7.4 and, subsequently, stained with 5 μM MitoSOX for 30 min at 37°C in the dark. Cells were analyzed in a SpectraMax Paradigm plate reader (Molecular Devices Co.). As a positive control, MitoSOX-loaded cells were treated with 5 µM TBHP for 10 min. Untreated and unstained cells served as a negative control.
2.8. RNA Extraction, cDNA Synthesis, and Gene Expression Analysis (RT-qPCR)
Cells were seeded at 105 cells/well. 24 h later, they were pre-incubated with test agents for 24 h. The medium was then replaced with one containing the treatment compounds, with or without rotenone, and incubated for an additional 48 h. An RNA extraction kit (GENEzolTM TriRNA Pure Kit+DNASE I; Geneaid, New Taipei City, Taiwan) was used to extract and purify the total RNA from cell cultures. The RNA was quantified using a micro-volume spectrophotometer (NanoDrop; Wilmington, DE, USA). mRNA was then reversed transcribed to cDNA using a reverse transcriptase kit (qScript cDNA synthesis kit; QUANTA Biosciences; Gaithersburg, MD, USA), and quantitative cDNA amplification was performed by real-time PCR (StepOne Real-Time PCR System; ThermoFisher Scientific; Wilmington, DE, USA) using an advanced master mix (ThermoFisher). A TaqMan assay was used for gene expression analysis of collagen type 1 (COL1A1) (Hs00164004_m1) with actin (Hs99999903_m1) as the housekeeping gene (ThermoFisher). Each treatment was studied with three biological replicates.
2.9. Mitochondrial-Respiration Parameters
Seahorse XF-96 Extracellular Flux (Agilent Technologies, Inc, Santa Clara, CA, USA) was used to measure key parameters of the mitochondrial function by measuring the oxygen consumption rate (OCR) and glycolysis by measuring the extracellular acidification rate (ECAR), according to the manufacturer’s instructions. The modulators included in this assay were oligomycin, carbonyl cyanide-4 (trifluoromethoxy), phenylhydrazone (FCCP), rotenone, and antimycin. Cells were seeded in 96-well XF plates at 104 cells/well. 24 h later, they were pre-incubated with the phytonutrients or estradiol for 24 h. The medium was then replaced with one containing the treatment compounds with rotenone and incubated for an additional 24 h. Each sample was assayed in triplicate. OCR parameters were calculated using Wave software and an Agilent Seahorse XFe Analyzer (version 2.6.3.5). Results were normalized to the living cell number in each well using DAPI staining.
2.10. Cellular ATP Levels
Cells were seeded in 96-well plates at 104 cells/well. 24 h later, they were pre-incubated with test agents for 24 h. The medium was then replaced with one containing the treatment compounds, with or without rotenone, and incubated for an additional 48 h. ATP cellular levels were determined using a CellTiter-Glo Luminescent Cell Viability Assay Kit (Promega, Madison, WI) according to the manufacturer’s instructions. Cells were incubated with ATP assay buffer at room temperature for 10 min. Luminescence was then measured in quadruplicate using a SpectraMax Paradigm plate reader (Molecular Devices Co., Sunnyvale, CA, USA).
2.11. Preparation of Whole Cell Lysates and Western Blotting
Cells were seeded in 6-well plates at 105 cells/well. 24 h later, they were pre-incubated with test agents for 24 h. The medium was then replaced with one containing the treatment compounds, with or without rotenone, and incubated for an additional 48 h. Cells were lysed in a buffer containing 1% (v/v) Triton X-100 at 4°C. 20 μg of each sample were separated by SDS-PAGE, and electroblotted onto nitrocellulose membranes. The membranes were blocked with 5% nonfat milk in Tris-buffered saline containing 0.5% Tween 20 (TBST) for 2 h, and incubated with primary antibodies overnight at 4°C, followed by 1 h incubation with the secondary antibody, Anti-Mouse IgG (H+L) (cat# 5450-0011, Sera Care, Milford, MA, USA). The following primary antibodies were used: NQO1 (Cat# SC-271116, 1:500) with β-actin as the internal loading control (Cat# SC-47778, 1:500), both purchased from Santa Cruz Biotechnology Inc. (Santa Cruz, CA). Protein bands were visualized using Western LightningTM Chemiluminescence Reagent Plus (PerkinElmer Life Sciences, Inc., Boston, MA, USA). The absorbance of each band was determined using an Image Quant LAS 4000 system (GE Healthcare Life Science, Little Chalfont, UK), and the optical density of each band was determined using ImageQuantTM TL software v 8.0 (GE Healthcare Life Science). Optical density values for each band were normalized to the β-actin.
2.12. Transient Transfection and ARE/Nrf2 Reporter Gene Assay
NHDF cells were seeded in 24-well plates at 105 cells/well. 24 h later, they were transfected using jetPEI reagent (Polyplus Transfection, Illkrich, France). Cells were then rinsed once with serum-free medium, followed by the addition of 0.45 mL of medium and 50 µL of a mixture containing DNA and jetPEI reagent at a charge ratio of 1:5. The total amount of DNA was 0.25 µg, containing 0.2 µg 4xARE reporter construct that was kindly provided by Dr. M. Hannink (University of Missouri-Columbia, Columbia, MO, USA) and 0.05 µg Renilla luciferase (P-RL-null) expression vectors, which served as an internal transfection standard, and was purchased from Promega (Madison, WI, USA). These conditions were found to be optimal for the dermal fibroblasts. The cells were then incubated for 6 h at 37°C. Next, the medium was replaced with DMEM-DCC-FBS plus the test compounds, and cells were incubated for an additional 16 h. Cell extracts were prepared for luciferase reporter assay (Dual Luciferase Reporter Assay System, Promega), according to the manufacturer’s instructions, and luminescence was determined using a Turner Biosystems luminometer (Sunnyvale, CA, USA).
2.13. Statistical Analysis
All experiments were conducted three to seven times in triplicates. The number of independent experiments (n) is indicated in the legends to figures. Statistically significant differences between two experimental groups were determined using a one-way ANOVA with Dunnett's multiple comparison post-hoc analysis. A p-value less than 0.05 was considered statistically significant. The statistical analyses were performed using GraphPad Prism 9.0 software (Graph-Pad Software, San Diego, CA).
3. Results
3.1. Mitochondrial and Cytoplasmic ROS Levels in Dermal Fibroblasts Are Increased by Rotenone Treatment and Reduced by Tomato and Rosemary Extracts, and Estradiol
The inhibition of mitochondrial respiration by rotenone should result in oxidative stress. Thus, to confirm that rotenone increases mitochondrial and cytoplasmic oxidative stress, ROS levels were detected using Mitosox and DCFH fluorescent probes, respectively. The average geometric means of fluorescence intensities (MFI)) were measured 2 h after adding rotenone for Mitosox and 4 h for DCFH. Rotenone treatment increased ROS levels by six-fold in the mitochondria (Figure 1a) and four -fold in the cytosol (Figure 1b). The increase in mitochondrial ROS levels was reduced by about 70% when the cells were pre-incubated with tomato and rosemary extracts and estradiol (Figure 1a), and the cytosolic ROS level was reduced by these compounds almost to the basal level (Figure 1b). It was then important to assess if these changes in ROS levels were associated with parallel changes in cell function.
3.2. Rotenone Triggers Apoptotic Cell Death of Dermal Fibroblasts, Which Can Be Reduced by Tomato and Rosemary Extracts and Estradiol
To investigate whether the rotenone-induced rise in ROS level results in apoptotic cell death, dermal fibroblasts were treated with 1 µM rotenone for 48 h, and apoptosis was analyzed by two methods: annexin V fluorescence and caspase 3 activity by an ApoLive-GloTM Multiplex Assay. Typical flow cytometric data of one experiment is shown in Figure 2a. About 10% of the cells were apoptotic in the control cells, whereas the percentage of annexin V-positive cells increased remarkably upon exposure to rotenone, with the average increase about 6-fold (Figure 2b, gray bar). A similar increase of about 5-fold was found by measuring caspase 3 activity—a key biomarker of apoptosis (Figure 2c, gray bar). Cell number and viability were detected by crystal violet staining and also by an ApoLive-Glo Multiplex viability assay. Rotenone treatment for 48 h decreased both cell number and cell viability by 50–60% (Figure 2d and 2e, respectively). Pre-incubation for 24 h with tomato or rosemary extracts or estradiol before exposure to rotenone in the presence of the treatment compounds resulted in a significant reduction of apoptosis (Figure 2b,c) and an increase in cell number and viability (Figure 2d–h). The increase in cell number was dose-dependent for all the treatment compounds (Figure 2f–h).
3.3. Rotenone Increases MMP1 and Reduces Collagen 1a1 Secretion. Carotenoids, Polyphenols, and Estradiol Reverse These Effects
The balance between MMPs and collagen protein level is important for skin elasticity and the prevention of wrinkles. Therefore, MMP-1 and pro-collagen 1a1 secretion from the dermal fibroblasts was measured in the media using specific ELISA assays. While rotenone increased MMP1 secretion by almost 2-fold, tomato and rosemary extracts and estradiol reduced this rotenone-induced rise almost to the basal level (Figure 3a). In contrast, rotenone decreased pro-collagen 1a1 secretion by almost 50%, and the tested compounds increased this secretion (Figure 3b). Collagen 1a1 mRNA expression was measured to determine if the reduction in the pro-collagen level was a result of the reduced expression or only from increased degradation due to the higher MMP1 level. It was found that rotenone reduced collagen 1a1 expression by nearly 60%, and that the phytonutrients and estradiol increased the collagen 1a1 mRNA significantly (Figure 3c).
3.4. Mitochondrial Dysfunction Following Exposure to Rotenone and Recovery by Dietary Compounds and Estradiol
Since the oxidative stress induced by rotenone and the cell damage which followed were partially corrected by the phytonutrients and estradiol, it was important to measure the mitochondrial activity to assess if this was also corrected. Mitochondrial and non-mitochondrial respiration parameters were measured using a Seahorse XF Analyzer. The cells were pre-incubated for 24 h with the treatment compounds or vehicle and then with rotenone, together with the compounds, for 3 h or 24 h, with similar results at both time points. Next, the cells were washed for 1 h according to the Seahorse protocol to remove the rotenone and treatment compounds, and the OCR and ECAR were analyzed. After measuring the basal OCR, cells were exposed sequentially to four modulators of oxidative phosphorylation (oligomycin, FCCP, and antimycin/rotenone), and the OCR was measured after the application of each modulator, enabling sequential measurement of the ATP-linked OCR, maximal respiration, and spare capacity. Treatment with rotenone alone resulted in a substantial reduction in the OCR, which remained low during the subsequent stages of the analysis (Figure 4a). This means that all measured parameters in the rotenone-treated cells were very low, as can be seen in Figure 4b–e. Pre-treatment with the phytonutrients and estradiol protected the mitochondria from the rotenone-induced mitochondrial dysfunction, showing basal OCR values that were significantly higher, reaching 50–60% of the control value (Figure 4b). The effects of the compounds on the ATP-linked OCR were examined after adding oligomycin, where the difference between the basal OCR and the OCR after oligomycin represents the ATP-linked OCR (Figure 4c). The compounds reversed the effect of rotenone to about 50% of the control. The reduction in ATP formation by rotenone was confirmed by directly measuring the ATP cellular level (Figure 4f). In the presence of rotenone alone, this level was reduced to about 40% of the control, while pre-treatment with the protective compounds restored it to about 70% of the control. Evaluation of the effects on glycolysis were measured by the ECAR and showed a significant increase in this rate by rotenone, as the cells were using glycolysis to compensate for the reduced mitochondrial ATP production (Figure 4g). Pre-treatment with the phytonutrients and estradiol, except for rosemary extract, restored the ECAR to the basal level. The enhancing effect of the dietary compounds and estradiol on the maximal mitochondrial respiration measured in the presence of FCCP is shown in Figure 4d, where the maximal respiration was restored to about 80% of the control, indicating improved mitochondrial function. The spare respiratory capacity that reflects the ability of the mitochondria to respond to an increased energy demand was restored to almost 100% of the control (Figure 4e).
3.5. Increased Activation of ARE/Nrf2 by the Combinations of the Phytonutrients or Estradiol with Rotenone
The results reported in the previous sections indicate that during pre-incubation with phytonutrients and estradiol, the cells increase their antioxidant defense capacity, which leads to decreased ROS levels and protection of the dermal fibroblasts from adverse rotenone-induced effects. This increased antioxidant capacity may be caused by activation of the ARE/Nrf2 transcriptional activity, which increases the transcription of antioxidant enzymes, an activity that was determined by a reporter gene assay. As expected, the transcription system was activated by rosemary extract, tomato extracts, and estradiol (Figure 5a) from 3- to 4-fold in the absence of rotenone. Rotenone alone slightly increased the activity by almost 3-fold. In the presence of rotenone, the activation by the phytonutrients and estradiol was significantly higher than that of the compounds or rotenone alone, or even from the sum of the effects of each compound alone (Figure 5a). To clearly present the increased effect of this combination on ARE/Nrf2 transcriptional activity, the sum of the effects of the rotenone and rosemary extract is shown in Figure 5b, where the combined effect is 15-fold and is twice higher than the sum of the effects of rotenone and rosemary extract alone. To verify this transcriptional activity, the protein level of NQO1, one of the antioxidant enzymes induced by this system, was detected using western blot. Indeed, rotenone alone slightly increased the level of the NQO1 protein, and in the presence of the dietary compounds and estradiol, the protein level was significantly higher than that of the rotenone alone (Figure 5c,d).
3.6. Activation of ARE/Nrf2 Is Required for Protection from Rotenone-Induced Damage by Dietary Compounds and Estradiol
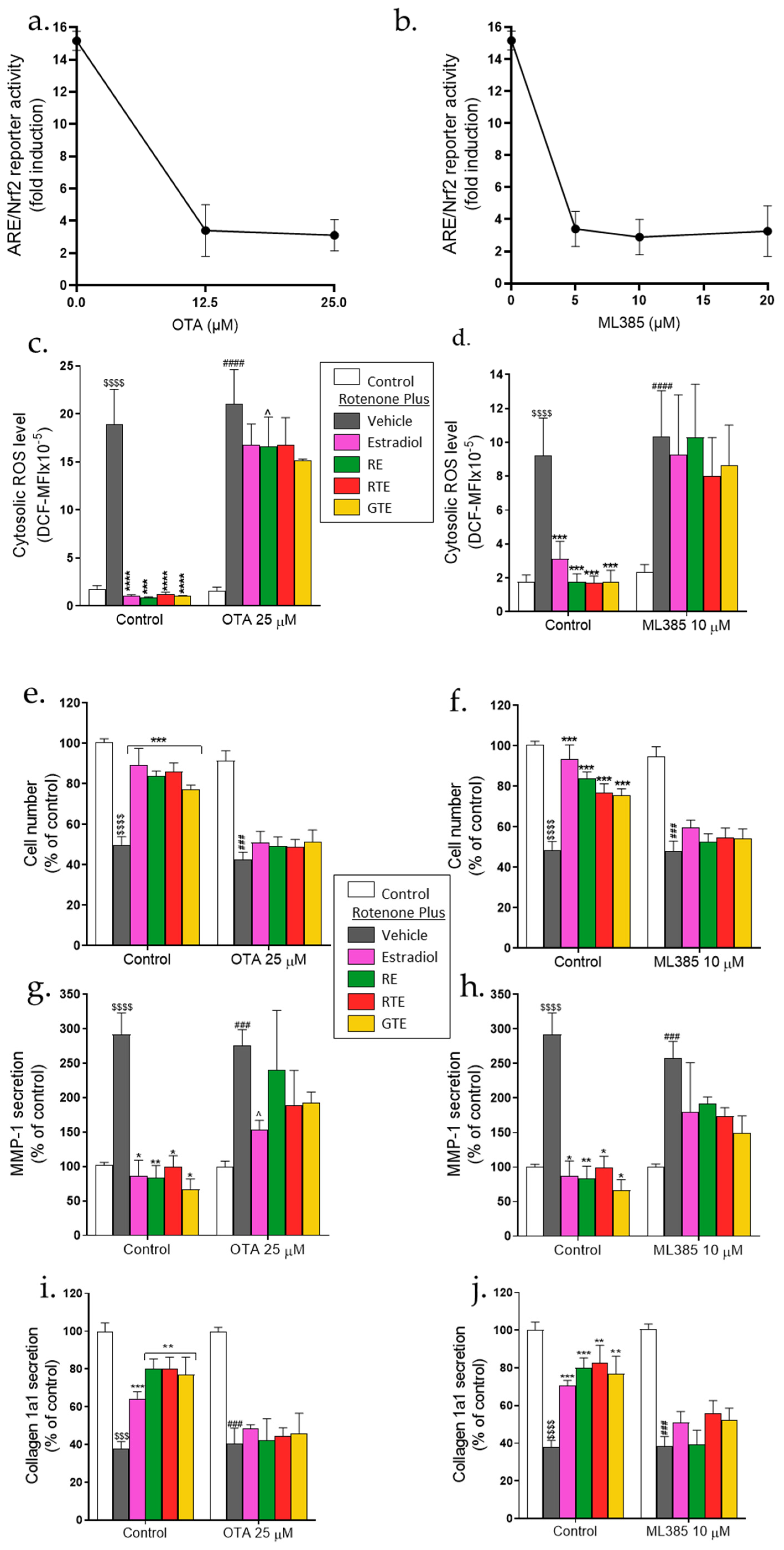
To investigate whether activation of the ARE/Nrf2 transcription system is crucial for protection by the dietary compounds and estradiol, two inhibitors of the ARE/Nrf2 pathway, OTA and ML385, were used. As can be seen in Figure 6a and Figure 6b, OTA or ML385 treatment at different concentrations inhibited the rotenone- and rosemary extract-induced ARE/Nrf2 transcriptional activity. The following results were from experiments performed with 25 µM OTA or 10 µM ML385: The protection, as measured by changes in ROS level, cell number, and MMP-1 and pro-collagen secretion was reduced in the presence of both inhibitors (Figure 6), whereas the effects of the treatment compounds in the absence of the inhibitors were similar to those shown in Figure 1, Figure 2 and Figure 3. In the presence of each inhibitor, only a small reduction in cytosolic ROS was obtained by pre-incubation with the treatment compounds (Figure 6c,d). Moreover, the cell number was not increased by these compounds in the presence of the inhibitors (Figure 6e,f). Only a partial and mostly statistically insignificant reduction in MMP-1 secretion (Figure 6g,h) and no increase in pro-collagen 1a1 secretion (Figure 6i,j) were attained by the phytonutrients and estradiol when cells were treated with the inhibitors.
4. Discussion
In the current study, we examined the harmful effects of endogenously generated oxidative stress in human dermal fibroblasts, which resulted from mitochondrial dysfunction induced by rotenone. Since mitochondrial function is associated with skin health, its dysfunction can lead to skin aging [39], and thus, the use of rotenone may be a good model for this process (see scheme 1a). In the current study, rotenone blocked mitochondrial respiration almost completely, as is evident from a reduction in the OCR, including the ATP-linked OCR. This was associated with reduced cellular ATP, increased mitochondrial and cytosolic ROS, apoptotic cell death, increased MMP1, and reduced collagen secretion. While the reduction in mitochondrial activity by rotenone treatment is much more pronounced than that which is expected during skin cell aging, the resulting robust effect on ROS and cell activity enabled significant effects of the protective compounds to be measured and the mechanisms involved in the damage and in protection to be analyzed. The increase in mitochondrial ROS was due to inhibition of Complex Ι by the rotenone, leading to the production of superoxide radicals [36] (see scheme 1a). The association between mitochondrial ROS and the reduction in mitochondrial metabolism has previously been shown as a reduction in the mitochondrial membrane potential and of oxidative phosphorylation [40]. Mitochondrial dysfunction is known to be caused by deletions in the mtDNA that accumulate with aging, resulting in increased ROS production and apoptosis induction in skin cells [16]. Mitochondria play an important role in the apoptosis cascade, as their dysfunction activates the cascade through cytochrome c release [41].
Wrinkles and loss of skin elasticity, among the disturbing phenotypes of skin aging, result from increased MMPs and reduced collagen production, which cause lower collagen levels [39] (see scheme 1a). In the current study, such changes in collagen metabolism were found following a reduction in mitochondrial activity. These outcomes are supported by a study on mitochondrial disease, myoclonic epilepsy, and ragged red fiber syndrome that occur due to genetic mutations in mtDNA which impair mitochondrial respiration, induce oxidative stress, and, in skin fibroblasts, increase MMP1 expression [42]. Additionally, depletion in mice mtDNA decreased oxidative phosphorylation in skin samples due to instability of the electron transport complexes that reduced expression of the MMP inhibitor TIMP1 and increased expression of MMP2 and MMP9 [43]. In UV-exposed NHDF cells, a decreased production of collagen fibers, due to their lower secretion, was shown to be associated with the low quality of the mitochondria, which resulted in ATP reduction [44]. The lower ATP level that was correlated with increased mitochondrial ROS caused less secretion of type I collagen and fibrillin-1, which is similar to the collagen secretion results in our study.
Although, in the current study, we show the harmful effects of endogenously-generated ROS, it is known that ROS exogenously generated by environmental factors leads to similar effects. In a previous study, we found that oxidative stress induced by H2O2 in dermal fibroblasts caused apoptotic cell death, increased MMP1, and decreased collagen secretion [24]. Additionally, UVB-induced oxidative stress in HaCaT keratinocytes [45] and in dermal fibroblasts [46] led to cell death due to the upregulation of proteins involved in the apoptotic process. The effects of exogenously-generated oxidative stress on the production of MMPs and collagen were also shown in several other studies. For example, exposure of dermal fibroblasts to ROS generated with xanthine and xanthine oxidase caused increased expression of MMP2 and decreased expression of the MMP inhibitor TIMP2 [47]. In in-vivo and in-vitro experiments, it was shown that UVB-induced oxidative stress, associated with disruption of the mitochondrial membrane potential, increased mRNA and the protein expression of MMPs and decreased collagen expression in dermal fibroblasts [48].
Carotenoids, polyphenols, and estradiol protect dermal fibroblasts from the damaging effects of rotenone. These compounds lowered the ROS level, improved mitochondrial function, reduced MMP1 secretion, and increased the secretion of collagen. Estradiol and the dietary compounds increased the activity of the ARE/Nrf2 transcription system cooperatively with rotenone, which suggests that this protection may be explained by activation of this antioxidant defense system (see scheme 1b). We have previously shown that carotenoids, polyphenols, and estradiol activated ARE/Nrf2 in dermal fibroblasts, which was associated with ROS reduction and protection from H2O2-induced damage [24]. In addition to its role in activating this transcription system, it was further shown that estradiol can increase the activity of antioxidant enzymes such as hemeoxygenase-1 via an estrogen receptor α-dependent mechanism [49].
The importance of ARE/Nrf2 for cell protection was proven using the Nrf2 inhibitors, ML385 [50] and ochratoxin A (OTA) [51]. Inhibition of ARE/Nrf2 prevented these protective effects by more than 50%, which indicates that this is one of the main mechanisms for reducing ROS, and leads to reversal of the damaging effects of rotenone (see scheme 1b). In a study on ex-vivo human skin, OTA blocked the anti-apoptotic effect of external DNA and bacterial biofilm, showing the importance of ARE/Nrf2 in preventing UVB-induced damage [52]. The Nrf2 inhibitor, ML385, blocked the protective effect of carnosic acid, the polyphenol found in rosemary extract, against ferroptotic cell death in PC12 cells [53]. In H9C2 myocardial cells exposed to hypoxia/reoxygenation, ginseng polyphenol reversed the process of increasing ROS and apoptotic cell death, and this was also blocked by ML385 [54]. These results show that, similar to skin cells, the mechanism for reducing ROS and preventing cell death in these nerve and myocardial cell lines is the ARE/Nrf2 antioxidant system.
In the current study, the addition of estradiol and phytonutrients improved mitochondrial respiration and increased ATP levels. The positive effect of estradiol to improve mitochondrial respiratory function, by increasing Complex I and III activities, was demonstrated in murine skeletal muscle [55]. In the PC-12 cells and brain mitochondria from both female and male rats, estradiol was found to decrease mitochondrial superoxide by increasing manganese superoxide dismutase activity [56]. Estradiol was also shown to increase the mitochondria membrane potential of human dermal fibroblasts and keratinocytes under H2O2-induced oxidative stress that, like in the current study, was associated with reduced ROS and protection from cell death [32]. Decreased mitochondrial respiration occurs in Friedreich’s ataxia (FRDA) and causes a reduction in ATP production. In skin fibroblasts from a human FRDA donor, that were under oxidative stress, the protective effects of various estrogens were studied [57,58]. The estrogens increased cell viability, oxidative phosphorylation, and intracellular ATP, and prevented collapse of the mitochondrial membrane potential. These protective effects were not related to any estrogen receptor activity and were associated with the phenolic structure and antioxidant properties of estradiol and the other estrogen-like compounds. Thus, it is important to examine the role of estrogen receptors in the protective effects of estradiol, as were found in the current study. It should be noted that the effective concentration of estradiol in these FRDA studies was 100 nM, which is ten times higher than the concentration we used in the current study.
The effects of phytonutrients in improving mitochondrial respiration are supported by previous studies that used other experimental models. For example, astaxanthin improved fatty acid utilization in murine skeletal muscle by increasing the protein amount of mitochondrial oxidative phosphorylation components, and in C2C12 cells, it increased mitochondrial biogenesis and enhanced mitochondrial oxidative respiration [59]. In addition, lycopene prevented testicular lesions induced by aflatoxin in mice by decreasing oxidative stress and increasing mitochondrial biogenesis and membrane potential [60]. In a study on endotoxin-induced damage to primary neonatal rat ventricular myocytes, the diterpene alkaloid, songorine, improved mitochondrial respiration by upregulating the mitochondrial genes involved in the electron transport chain and fatty acid β-oxidation, and prevented endotoxin-induced septic heart injury in mice [61]. The protective effects on mitochondrial respiration and biogenesis were dependent on the activation of ARE/Nrf2, and Nrf2 deficiency diminished this protection. In the current study, the improvement observed in mitochondrial activity, which similarly depends on ARE/Nrf2 activation, may also be associated with increased mitochondrial biogenesis; however, this issue should be examined further.
In this study, phytonutrients and estradiol reduced ROS levels and MMP1 secretion, increased collagen secretion, and decreased cell death by reducing caspase 3 activity and apoptosis. The effect of carotenoids on the apoptosis pathway in human keratinocytes has been previously shown with astaxanthin, which decreased ROS production and reversed the UVB-induced increased expression of apoptosis proteins, including BAX, caspase 3, and PARP, resulting in increased cell viability [62]. Additionally, caspase 3 expression was reduced by topical application of lycopene, which suppressed UVB-induced apoptosis in murine skin [63]. In another study, the polyphenol, resveratrol, inhibited caspase 3 and 9 expression in both HaCaT keratincytes and skin in mice [64]. Other research found that in HaCaT keratinocytes and dermal fibroblasts, estradiol helped protect against oxidative stress by decreasing the ROS level, resulting in increased cell viability and pro-collagen I synthesis [65]. Borrás et al suggested that estradiol has a direct protective effect on the mitochondria as it decreased ROS production by isolated mitochondria, and maintained mitochondrial integrity and membrane potential, preventing leakage of cytochrome c, which should reduce cell apoptosis [66]. Oxidative stress has been found to activate MMP expression [67,68], reduce the expression of pro collagen, and decrease collagen synthesis [69]. Similar to the current study, estradiol was shown to decrease the expression of MMP1 and MMP9 in keratinocytes and dermal fibroblasts [32]. In aged human skin in vivo, topical application of estradiol decreased MMP1 expression and increased that of type I pro collagen mRNA and protein [70]. Several studies have reported that carotenoids [71,72] and polyphenols [73,74,75] reduce the level or expression of MMPs and increase collagen protein expression. These changes in collagen metabolism should lead to increased skin elasticity and reduced wrinkling, and indeed, human studies show that supplementation for 12 weeks with golden tomato extract [76] or 90 days with β-carotene [71] improved skin elasticity and firmness, and reduced fine lines and facial wrinkles. The effects of dietary compounds on MMPs and collagen expression were associated with suppressing the activation of mitogen-activated protein kinases (MAPKs) and activator protein 1 (AP-1) signaling [75,77]. Thus, the involvement of these, and other signaling pathways, in mitigating rotenone-induced damage should be investigated.
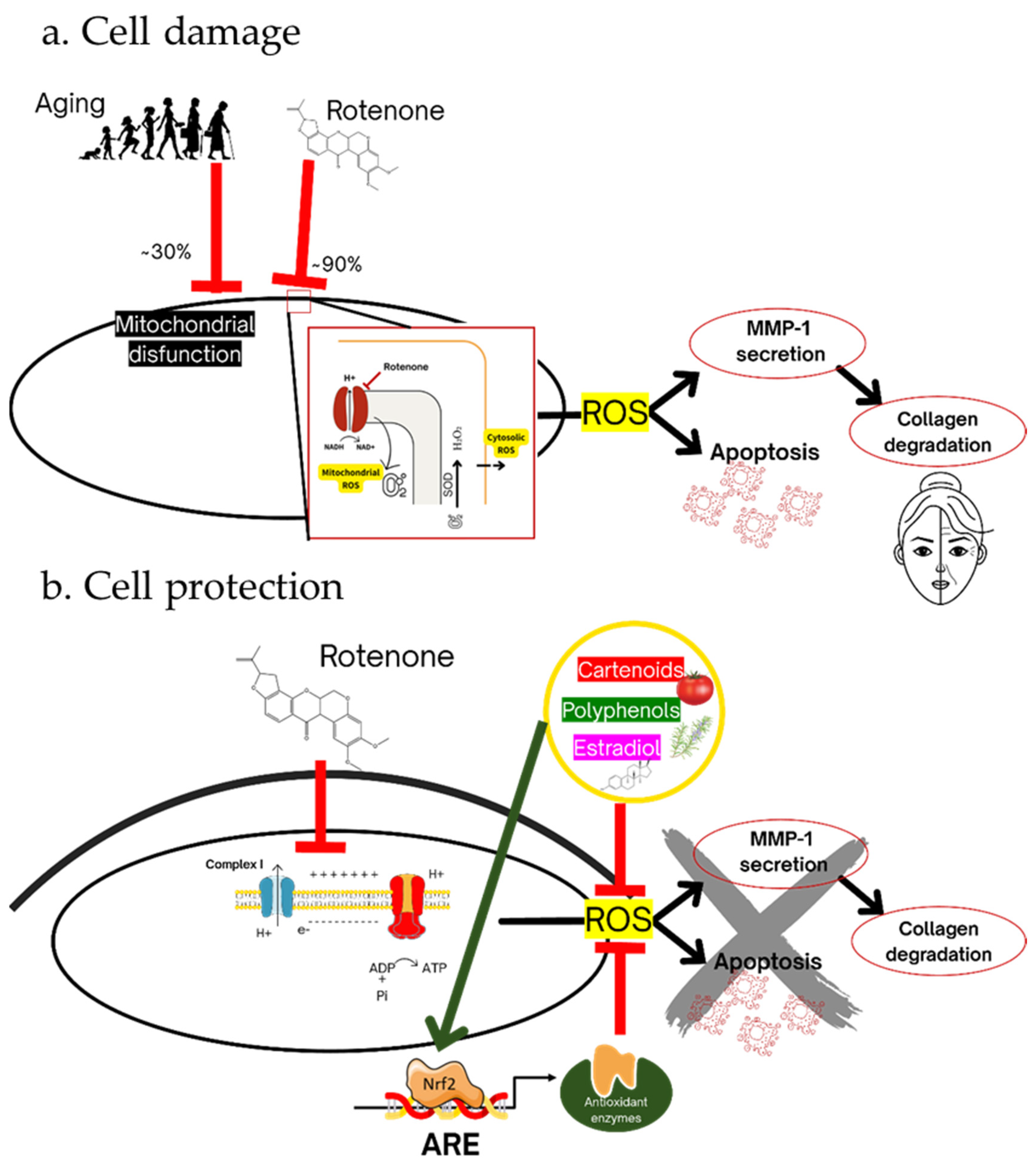
Scheme 1.
(a) Cell damage. Partial mitochondrial dysfunction occurs in skin cell aging and almost complete dysfunction occurs by rotenone treatment with a significant reduction in mitochondrial respiration and ATP level. This dysfunction results in the production of superoxide radical and other ROS, which cause cell damage that is manifested as increased MMP secretion, reduced collagen secretion, and apoptotic cell death. (b) Cell protection. Carotenoids, polyphenols, and the hormone estradiol protect the cell from rotenone-induced damage by activating the ARE/Nrf2 transcription system, resulting in reduced ROS and partial reversal of the harmful effects.
Scheme 1.
(a) Cell damage. Partial mitochondrial dysfunction occurs in skin cell aging and almost complete dysfunction occurs by rotenone treatment with a significant reduction in mitochondrial respiration and ATP level. This dysfunction results in the production of superoxide radical and other ROS, which cause cell damage that is manifested as increased MMP secretion, reduced collagen secretion, and apoptotic cell death. (b) Cell protection. Carotenoids, polyphenols, and the hormone estradiol protect the cell from rotenone-induced damage by activating the ARE/Nrf2 transcription system, resulting in reduced ROS and partial reversal of the harmful effects.
5. Conclusions
The mitochondrial theory of aging suggests that dysfunction of the mitochondria induces cell aging; thus, in the current study, we used rotenone to establish a model of these processes. Our findings indicate that carotenoids, polyphenols, and estradiol protect dermal fibroblasts from mitochondrial dysfunction-induced damage. Therefore, the consumption of these and similar dietary compounds can delay skin aging. Moreover, activation of the ARE/Nrf2 transcription system was shown to be an important mechanism for protecting skin cells from a reduction in mitochondrial activity. However, the results obtained in the presence of ARE/Nrf2 inhibitors indicate that this is not the only protective mechanism, and thus, the role of other signaling pathways in cell damage and for skin protection should be analyzed.
Author Contributions
Conceptualization: A.D. and Y.S.; methodology: A.D.; formal analysis: A.D. and A.T.; investigation: A.D.; writing—original draft preparation: A.D. and Y.S.; writing—review and editing: A.D., A.T., and Y.S.; visualization: A.D., A.T., and Y.S.; supervision: Y.S.; project administration: Y.S.; funding acquisition: Y.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Lycored Ltd., Beer Sheva, Israel (to Y.S.), grant numbers 87872611, 81101211 and 81184811.
Acknowledgments
We thank Dr. M. Hannink (University of Missouri-Columbia, Columbia, MO) for donating the 4xARE reporter construct used in this study, and Robin Miller for English editing.
Conflicts of Interest
Y.S. is a consultant for Lycored Ltd, Beer Sheva, Israel, and received research funding from this company. All other authors declare no conflicts of interest. Lycored is a supplier to the dietary supplement and functional food industries worldwide. It had no role in the design of the study; in the collection, analyses, or interpretation of the data; in the writing of the manuscript; or in the decision to publish the results.
References
- Bickers, D.R.; Athar, M. Oxidative stress in the pathogenesis of skin disease. Journal of Investigative Dermatology 2006, 126, 2565-2575. [CrossRef]
- Dan Dunn, J.; Alvarez, L.A.J.; Zhang, X.; Soldati, T. Reactive oxygen species and mitochondria: A nexus of cellular homeostasis. Redox Biology 2015, 6, 472-485. [CrossRef] [PubMed]
- Betteridge, D.J. What is oxidative stress? Metabolism 2000, 49, 3-8. [CrossRef]
- Ray, P.D.; Huang, B.-W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cellular Signalling 2012, 24, 981-990. [CrossRef] [PubMed]
- Shukla, V.; Mishra, S.K.; Pant, H.C. Oxidative stress in neurodegeneration. Advances in Pharmacological Sciences 2011, 2011, 572634. [CrossRef] [PubMed]
- Sarmiento-Salinas, F.L.; Perez-Gonzalez, A.; Acosta-Casique, A.; Ix-Ballote, A.; Diaz, A.; Treviño, S.; Rosas-Murrieta, N.H.; Millán-Perez-Peña, L.; Maycotte, P. Reactive oxygen species: Role in carcinogenesis, cancer cell signaling and tumor progression. Life Sciences 2021, 284, 119942. [CrossRef] [PubMed]
- Haigis, M.C.; Yankner, B.A. The aging stress response. Mol Cell 2010, 40, 333-344. [CrossRef]
- Bratic, A.; Larsson, N.-G. The role of mitochondria in aging. The Journal of Clinical Investigation 2013, 123, 951-957. [CrossRef]
- Angelova, P.R.; Abramov, A.Y. Functional role of mitochondrial reactive oxygen species in physiology. Free Radical Biology and Medicine 2016, 100, 81-85. [CrossRef]
- Jastroch, M.; Divakaruni, A.S.; Mookerjee, S.; Treberg, J.R.; Brand, M.D. Mitochondrial proton and electron leaks. Essays in Biochemistry 2010, 47, 53-67. [CrossRef]
- Sena, Laura A.; Chandel, Navdeep S. Physiological roles of mitochondrial reactive oxygen species. Molecular Cell 2012, 48, 158-167. [CrossRef]
- Reddy, P.H. Mitochondrial Medicine for Aging and neurodegenerative diseases. NeuroMolecular Medicine 2008, 10, 291-315. [CrossRef] [PubMed]
- Dias, V.; Junn, E.; Mouradian, M.M. The role of oxidative stress in Parkinson's disease. J Parkinsons Dis 2013, 3, 461-491. [CrossRef] [PubMed]
- HARMAN, D. The biologic clock: The mitochondria? Journal of the American Geriatrics Society 1972, 20, 145-147. [CrossRef] [PubMed]
- Krutmann, J.; Schroeder, P. Role of mitochondria in photoaging of human skin: The defective powerhouse model. Journal of Investigative Dermatology Symposium Proceedings 2009, 14, 44-49. [CrossRef] [PubMed]
- Sreedhar, A.; Aguilera-Aguirre, L.; Singh, K.K. Mitochondria in skin health, aging, and disease. Cell Death & Disease 2020, 11, 444. [CrossRef]
- Natarelli, N.; Gahoonia, N.; Aflatooni, S.; Bhatia, S.; Sivamani, R.K. Dermatologic manifestations of mitochondrial dysfunction: A review of the literature. International Journal of Molecular Sciences 2024, 25, 3303. [CrossRef] [PubMed]
- Zhou, B.-r.; Yin, H.-b.; Xu, Y.; Wu, D.; Zhang, Z.-h.; Yin, Z.-q.; Permatasari, F.; Luo, D. Baicalin protects human skin fibroblasts from ultraviolet A radiation-induced oxidative damage and apoptosis. Free Radical Research 2012, 46, 1458-1471. [CrossRef] [PubMed]
- Rinnerthaler, M.; Bischof, J.; Streubel, M.K.; Trost, A.; Richter, K. Oxidative stress in aging human Skin. Biomolecules 2015, 5, 545-589. [CrossRef]
- Wang, L.; Lee, W.; Jayawardena, T.U.; Cha, S.H.; Jeon, Y.J. Dieckol, an algae-derived phenolic compound, suppresses airborne particulate matter-induced skin aging by inhibiting the expressions of pro-inflammatory cytokines and matrix metalloproteinases through regulating NF-κB, AP-1, and MAPKs signaling pathways. Food Chem Toxicol 2020, 146, 111823. [CrossRef]
- Fam, V.W.; Charoenwoodhipong, P.; Sivamani, R.K.; Holt, R.R.; Keen, C.L.; Hackman, R.M. Plant-based foods for skin health: A Narrative Review. J Acad Nutr Diet 2021. [CrossRef]
- Michalak, M.; Pierzak, M.; Kręcisz, B.; Suliga, E. Bioactive compounds for skin health: A review. Nutrients 2021, 13. [CrossRef]
- Sies, H.; Stahl, W. Nutritional protection against skin damage from sunlight. Annual Review of Nutrition 2004, 24, 173-200. [CrossRef]
- Darawsha, A.; Trachtenberg, A.; Levy, J.; Sharoni, Y. The protective effect of carotenoids, polyphenols, and estradiol on dermal fibroblasts under oxidative stress. Antioxidants 2021, 10, 2023. [CrossRef] [PubMed]
- Park, C.; Lee, H.; Noh, J.S.; Jin, C.-Y.; Kim, G.-Y.; Hyun, J.W.; Leem, S.-H.; Choi, Y.H. Hemistepsin A protects human keratinocytes against hydrogen peroxide-induced oxidative stress through activation of the Nrf2/HO-1 signaling pathway. Archives of Biochemistry and Biophysics 2020, 691, 108512. [CrossRef] [PubMed]
- Ben-Dor, A.; Steiner, M.; Gheber, L.; Danilenko, M.; Dubi, N.; Linnewiel, K.; Zick, A.; Sharoni, Y.; Levy, J. Carotenoids activate the antioxidant response element transcription system. Mol Cancer Ther 2005, 4, 177-186. [CrossRef]
- Linnewiel, K.; Ernst, H.; Caris-Veyrat, C.; Ben-Dor, A.; Kampf, A.; Salman, H.; Danilenko, M.; Levy, J.; Sharoni, Y. Structure activity relationship of carotenoid derivatives in activation of the electrophile/antioxidant response element transcription system. Free Radical Biology & Medicine 2009, 47, 659-667.
- Calniquer, G.; Khanin, M.; Ovadia, H.; Linnewiel-Hermoni, K.; Stepensky, D.; Trachtenberg, A.; Sedlov, T.; Braverman, O.; Levy, J.; Sharoni, Y. Combined effects of carotenoids and polyphenols in balancing the response of skin cells to UV irradiation. Molecules 2021, 26. [CrossRef]
- Zhou, Y.; Jiang, Z.; Lu, H.; Xu, Z.; Tong, R.; Shi, J.; Jia, G. Recent advances of natural polyphenols activators for Keap1-Nrf2 signaling pathway. Chemistry & Biodiversity 2019, 16, e1900400. [CrossRef]
- Heo, H.; Lee, H.; Yang, J.; Sung, J.; Kim, Y.; Jeong, H.S.; Lee, J. Protective activity and underlying mechanism of ginseng seeds against UVB-induced damage in human fibroblasts. Antioxidants (Basel) 2021, 10. [CrossRef]
- Lim, S.-R.; Kim, D.-W.; Sung, J.; Kim, T.H.; Choi, C.-H.; Lee, S.-J. Astaxanthin inhibits autophagic cell death induced by bisphenol A in human dermal fibroblasts. antioxidants 2021, 10, 1273. [CrossRef] [PubMed]
- Savoia, P.; Raina, G.; Camillo, L.; Farruggio, S.; Mary, D.; Veronese, F.; Graziola, F.; Zavattaro, E.; Tiberio, R.; Grossini, E. Anti-oxidative effects of 17 β-estradiol and genistein in human skin fibroblasts and keratinocytes. J Dermatol Sci 2018, 92, 62-77. [CrossRef] [PubMed]
- Lephart, E.D. A review of the role of estrogen in dermal aging and facial attractiveness in women. J Cosmet Dermatol 2018, 17, 282-288. [CrossRef] [PubMed]
- Jackson, R.L.; Greiwe, J.S.; Schwen, R.J. Ageing skin: oestrogen receptor β agonists offer an approach to change the outcome. Exp Dermatol 2011, 20, 879-882. [CrossRef] [PubMed]
- Chen, C.S.; Tseng, Y.T.; Hsu, Y.Y.; Lo, Y.C. Nrf2-Keap1 antioxidant defense and cell survival signaling are upregulated by 17β-estradiol in homocysteine-treated Dopaminergic SH-SY5Y cells. Neuroendocrinology 2013, 97, 232-241. [CrossRef] [PubMed]
- Sherer, T.B.; Betarbet, R.; Testa, C.M.; Seo, B.B.; Richardson, J.R.; Kim, J.H.; Miller, G.W.; Yagi, T.; Matsuno-Yagi, A.; Greenamyre, J.T. Mechanism of toxicity in rotenone models of Parkinson's disease. The Journal of Neuroscience 2003, 23, 10756-10764. [CrossRef] [PubMed]
- Cooney, R.V.; Joseph Kappock, T.; Pung, A.; Bertram, J.S. Solubilization, cellular uptake, and activity of β-carotene and other carotenoids as inhibitors of neoplastic transformation in cultured cells. In Methods in Enzymology; Academic Press: 1993; Volume 214, pp. 55-68.
- Levy, J.; Bosin, E.; Feldman, B.; Giat, Y.; Miinster, A.; Danilenko, M.; Sharoni, Y. Lycopene is a more potent inhibitor of human cancer cell proliferation than either alpha-carotene or beta-carotene. Nutr Cancer 1995, 24, 257-266. [CrossRef] [PubMed]
- Stout, R.; Birch-Machin, M. Mitochondria’s role in skin ageing. Biology 2019, 8, 29. [CrossRef] [PubMed]
- Rottenberg, H.; Hoek, J.B. The path from mitochondrial ROS to aging runs through the mitochondrial permeability transition pore. Aging Cell 2017, 16, 943-955. [CrossRef]
- Liu, X.; Kim, C.N.; Yang, J.; Jemmerson, R.; Wang, X. Induction of apoptotic program in cell-free extracts: requirement for dATP and cytochrome c. Cell 1996, 86, 147-157. [CrossRef]
- MA, Y.-S.; CHEN, Y.-C.; LU, C.-Y.; LIU, C.-Y.; WEI, Y.-H. Upregulation of matrix metalloproteinase 1 and disruption of mitochondrial network in skin fibroblasts of patients with MERRF syndrome. Annals of the New York Academy of Sciences 2005, 1042, 55-63. [CrossRef] [PubMed]
- Singh, B.; Schoeb, T.R.; Bajpai, P.; Slominski, A.; Singh, K.K. Reversing wrinkled skin and hair loss in mice by restoring mitochondrial function. Cell Death & Disease 2018, 9, 735. [CrossRef]
- Katsuyama, Y.; Yamawaki, Y.; Sato, Y.; Muraoka, S.; Yoshida, M.; Okano, Y.; Masaki, H. Decreased mitochondrial function in UVA-irradiated dermal fibroblasts causes the insufficient formation of type I collagen and fibrillin-1 fibers. Journal of Dermatological Science 2022, 108, 22-29. [CrossRef] [PubMed]
- Wang, Y.; Li, W.; Xu, S.; Hu, R.; Zeng, Q.; Liu, Q.; Li, S.; Lee, H.; Chang, M.; Guan, L. Protective skin aging effects of cherry blossom extract (Prunus Yedoensis) on oxidative stress and apoptosis in UVB-irradiated HaCaT cells. Cytotechnology 2019, 71, 475-487. [CrossRef] [PubMed]
- Gary, A.-S.; Rochette, P.J. Apoptosis, the only cell death pathway that can be measured in human diploid dermal fibroblasts following lethal UVB irradiation. Scientific Reports 2020, 10, 18946. [CrossRef] [PubMed]
- Kawaguchi, Y.; Tanaka, H.; Okada, T.; Konishi, H.; Takahashi, M.; Ito, M.; Asai, J. The effects of ultraviolet A and reactive oxygen species on the mRNA expression of 72-kDa type IV collagenase and its tissue inhibitor in cultured human dermal fibroblasts. Arch Dermatol Res 1996, 288, 39-44. [CrossRef] [PubMed]
- Mu, J.; Ma, H.; Chen, H.; Zhang, X.; Ye, M. Luteolin prevents UVB-induced skin photoaging damage by modulating SIRT3/ROS/MAPK Signaling: An in vitro and in vivo studies. Frontiers in Pharmacology 2021, 12. [CrossRef] [PubMed]
- Baruscotti, I.; Barchiesi, F.; Jackson, E.K.; Imthurn, B.; Stiller, R.; Kim, J.-H.; Schaufelberger, S.; Rosselli, M.; Hughes, C.C.W.; Dubey, R.K. Estradiol stimulates capillary formation by human endothelial progenitor cells. Hypertension 2010, 56, 397-404, doi:. [CrossRef]
- Singh, A.; Venkannagari, S.; Oh, K.H.; Zhang, Y.Q.; Rohde, J.M.; Liu, L.; Nimmagadda, S.; Sudini, K.; Brimacombe, K.R.; Gajghate, S.; et al. Small molecule inhibitor of NRF2 selectively intervenes therapeutic resistance in KEAP1-deficient NSCLC tumors. ACS Chem Biol 2016, 11, 3214-3225. [CrossRef]
- Limonciel, A.; Jennings, P. A review of the evidence that ochratoxin A is an Nrf2 inhibitor: Implications for nephrotoxicity and renal carcinogenicity. Toxins (Basel) 2014, 6, 371-379. [CrossRef]
- Ron-Doitch, S.; Frušić-Zlotkin, M.; Soroka, Y.; Duanis-Assaf, D.; Amar, D.; Kohen, R.; Steinberg, D. eDNA-mediated cutaneous protection against UVB damage conferred by staphylococcal epidermal colonization. Microorganisms 2021, 9, 788. [CrossRef] [PubMed]
- Cheng, J.; Xu, T.; Xun, C.; Guo, H.; Cao, R.; Gao, S.; Sheng, W. Carnosic acid protects against ferroptosis in PC12 cells exposed to erastin through activation of Nrf2 pathway. Life Sciences 2021, 266, 118905. [CrossRef] [PubMed]
- Sun, J.; Yu, X.; Huangpu, H.; Yao, F. Ginsenoside Rb3 protects cardiomyocytes against hypoxia/reoxygenation injury via activating the antioxidation signaling pathway of PERK/Nrf2/HMOX1. Biomedicine & Pharmacotherapy 2019, 109, 254-261. [CrossRef]
- Torres, M.J.; Kew, K.A.; Ryan, T.E.; Pennington, E.R.; Lin, C.-T.; Buddo, K.A.; Fix, A.M.; Smith, C.A.; Gilliam, L.A.; Karvinen, S.; et al. 17β-estradiol directly lowers mitochondrial membrane microviscosity and improves bioenergetic function in skeletal Muscle. Cell Metabolism 2018, 27, 167-179.e167. [CrossRef] [PubMed]
- Razmara, A.; Duckles, S.P.; Krause, D.N.; Procaccio, V. Estrogen suppresses brain mitochondrial oxidative stress in female and male rats. Brain Research 2007, 1176, 71-81. [CrossRef] [PubMed]
- Richardson, T.E.; Yang, S.-H.; Wen, Y.; Simpkins, J.W. Estrogen protection in Friedreich's ataxia skin fibroblasts. Endocrinology 2011, 152, 2742-2749. [CrossRef] [PubMed]
- Richardson, T.E.; Yu, A.E.; Wen, Y.; Yang, S.-H.; Simpkins, J.W. Estrogen prevents oxidative damage to the mitochondria in Friedreich's ataxia skin fibroblasts. PLOS ONE 2012, 7, e34600. [CrossRef] [PubMed]
- Nishida, Y.; Nawaz, A.; Kado, T.; Takikawa, A.; Igarashi, Y.; Onogi, Y.; Wada, T.; Sasaoka, T.; Yamamoto, S.; Sasahara, M.; et al. Astaxanthin stimulates mitochondrial biogenesis in insulin resistant muscle via activation of AMPK pathway. Journal of Cachexia, Sarcopenia and Muscle 2020, 11, 241-258. [CrossRef] [PubMed]
- Huang, W.; Cao, Z.; Cui, Y.; Huo, S.; Shao, B.; Song, M.; Cheng, P.; Li, Y. Lycopene ameliorates aflatoxin B1-induced testicular lesion by attenuating oxidative stress and mitochondrial damage with Nrf2 activation in mice. Ecotoxicology and Environmental Safety 2023, 256, 114846. [CrossRef]
- Li, Y.; Feng, Y.-F.; Liu, X.-T.; Li, Y.-C.; Zhu, H.-M.; Sun, M.-R.; Li, P.; Liu, B.; Yang, H. Songorine promotes cardiac mitochondrial biogenesis via Nrf2 induction during sepsis. Redox Biology 2021, 38, 101771. [CrossRef]
- Chung, B.Y.; Park, S.H.; Yun, S.Y.; Yu, D.S.; Lee, Y.B. Astaxanthin protects ultraviolet b-induced oxidative stress and apoptosis in human keratinocytes via intrinsic apoptotic pathway. Ann Dermatol 2022, 34, 125-131. [CrossRef] [PubMed]
- Fazekas, Z.; Gao, D.; Saladi, R.N.; Lu, Y.; Lebwohl, M.; Wei, H. Protective effects of lycopene against ultraviolet B-induced photodamage. Nutrition and Cancer 2003, 47, 181-187. [CrossRef] [PubMed]
- Cui, B.; Wang, Y.; Jin, J.; Yang, Z.; Guo, R.; Li, X.; Yang, L.; Li, Z. Resveratrol treats UVB-induced photoaging by anti-MMP expression, through anti-inflammatory, antioxidant, and antiapoptotic properties, and treats photoaging by Upregulating VEGF-B expression. Oxidative Medicine and Cellular Longevity 2022, 2022, 6037303. [CrossRef] [PubMed]
- Bottai, G.; Mancina, R.; Muratori, M.; Di Gennaro, P.; Lotti, T. 17β-estradiol protects human skin fibroblasts and keratinocytes against oxidative damage. Journal of the European Academy of Dermatology and Venereology 2013, 27, 1236-1243. [CrossRef] [PubMed]
- Borrás, C.; Gambini, J.; López-Grueso, R.; Pallardó, F.V.; Viña, J. direct antioxidant and protective effect of estradiol on isolated mitochondria. Biochimica et Biophysica Acta - Molecular Basis of Disease 2009, 1802, 205. [CrossRef] [PubMed]
- Fisher, G.J.; Datta, S.C.; Talwar, H.S.; Wang, Z.-Q.; Varani, J.; Kang, S.; Voorhees, J.J. Molecular basis of sun-induced premature skin ageing and retinoid antagonism. Nature 1996, 379, 335-339. [CrossRef] [PubMed]
- Kim, D.J.; Iwasaki, A.; Chien, A.L.; Kang, S. UVB-mediated DNA damage induces matrix metalloproteinases to promote photoaging in an AhR- and SP1-dependent manner. JCI Insight 2022, 7. [CrossRef] [PubMed]
- Siwik, D.A.; Pagano, P.J.; Colucci, W.S. Oxidative stress regulates collagen synthesis and matrix metalloproteinase activity in cardiac fibroblasts. American Journal of Physiology-Cell Physiology 2001, 280, C53-C60. [CrossRef]
- Son, E.D.; Lee, J.Y.; Lee, S.; Kim, M.S.; Lee, B.G.; Chang, I.S.; Chung, J.H. Topical application of 17β-estradiol increases extracellular matrix protein synthesis by stimulating TGF-β signaling in aged human skin in vivo. Journal of Investigative Dermatology 2005, 124, 1149-1161. [CrossRef]
- Cho, S.; Lee, D.H.; Won, C.-H.; Kim, S.M.; Lee, S.; Lee, M.-J.; Chung, J.H. Differential effects of low-dose and high-dose beta-carotene supplementation on the signs of photoaging and type i procollagen gene expression in human Skin in vivo. Dermatology 2010, 221, 160-171. [CrossRef]
- Yang, T.-H.; Lai, Y.-H.; Lin, T.-P.; Liu, W.-S.; Kuan, L.-C.; Liu, C.-C. Chronic exposure to rhodobacter sphaeroides extract lycogen™ prevents UVA-induced malondialdehyde accumulation and procollagen i down-regulation in human dermal Fibroblasts. International Journal of Molecular Sciences 2014, 15, 1686-1699. [CrossRef] [PubMed]
- Galicka, A.; Sutkowska-Skolimowska, J. The beneficial effect of rosmarinic acid on benzophenone-3-induced alterations in human skin fibroblasts. International Journal of Molecular Sciences 2021, 22, 11451. [CrossRef] [PubMed]
- Philips, N.; Conte, J.; Chen, Y.-J.; Natrajan, P.; Taw, M.; Keller, T.; Givant, J.; Tuason, M.; Dulaj, L.; Leonardi, D.; et al. Beneficial regulation of matrixmetalloproteinases and their inhibitors, fibrillar collagens and transforming growth factor-β by Polypodium leucotomos, directly or in dermal fibroblasts, ultraviolet radiated fibroblasts, and melanoma cells. Archives of Dermatological Research 2009, 301, 487-495. [CrossRef] [PubMed]
- Oh, J.H.; Joo, Y.H.; Karadeniz, F.; Ko, J.; Kong, C.-S. Syringaresinol inhibits UVA-Induced MMP-1 expression by suppression of MAPK/AP-1 signaling in HaCaT keratinocytes and human dermal fibroblasts. International Journal of Molecular Sciences 2020, 21, 3981. [CrossRef] [PubMed]
- Tarshish, E.; Hermoni, K.; Sharoni, Y.; Muizzuddin, N. Effect of Lumenato oral supplementation on plasma carotenoid levels and improvement of visual and experiential skin attributes. Journal of Cosmetic Dermatology 2022, 21, 4042-4052. [CrossRef]
- Choi, S.-I.; Han, H.-S.; Kim, J.-M.; Park, G.; Jang, Y.-P.; Shin, Y.-K.; Ahn, H.-S.; Lee, S.-H.; Lee, K.-T. Eisenia bicyclis extract repairs uvb-induced skin photoaging in vitro and in vivo: Photoprotective effects. Marine Drugs 2021, 19, 693. [CrossRef]
Figure 1.
Tomato and rosemary extracts and estradiol reduce rotenone-induced mitochondrial and cytosolic ROS. Cells were seeded in 96-well plates (5 × 103 cells/well) and pre-incubated for 24 h with rosemary extract (RE; 10 μM carnosic acid), red tomato extract (RTE; 10 μM lycopene), golden tomato extract (GTE; 40 μM phytoene), or estradiol (10 nM). Then the medium was replaced with one containing the treatment compounds plus 1 µM rotenone and incubated for 90 min or 4 h to detect mitochondrial ROS (Mitosox fluorescence) or cytosolic ROS (DCF fluorescence), respectively. (a) Mitochondrial ROS. (b) Cytosolic ROS. Values are averaged geometric means of fluorescence intensities (MFI) of Mitosox or DCF, and are the means ± SEM of four experiments, each performed in triplicate. $$$–$$$$, p < 0.001–0.0001, significant difference between the control and rotenone. *-****, p < 0.05–0.0001), significant difference between the vehicle and other treatments in the presence of rotenone.
Figure 1.
Tomato and rosemary extracts and estradiol reduce rotenone-induced mitochondrial and cytosolic ROS. Cells were seeded in 96-well plates (5 × 103 cells/well) and pre-incubated for 24 h with rosemary extract (RE; 10 μM carnosic acid), red tomato extract (RTE; 10 μM lycopene), golden tomato extract (GTE; 40 μM phytoene), or estradiol (10 nM). Then the medium was replaced with one containing the treatment compounds plus 1 µM rotenone and incubated for 90 min or 4 h to detect mitochondrial ROS (Mitosox fluorescence) or cytosolic ROS (DCF fluorescence), respectively. (a) Mitochondrial ROS. (b) Cytosolic ROS. Values are averaged geometric means of fluorescence intensities (MFI) of Mitosox or DCF, and are the means ± SEM of four experiments, each performed in triplicate. $$$–$$$$, p < 0.001–0.0001, significant difference between the control and rotenone. *-****, p < 0.05–0.0001), significant difference between the vehicle and other treatments in the presence of rotenone.
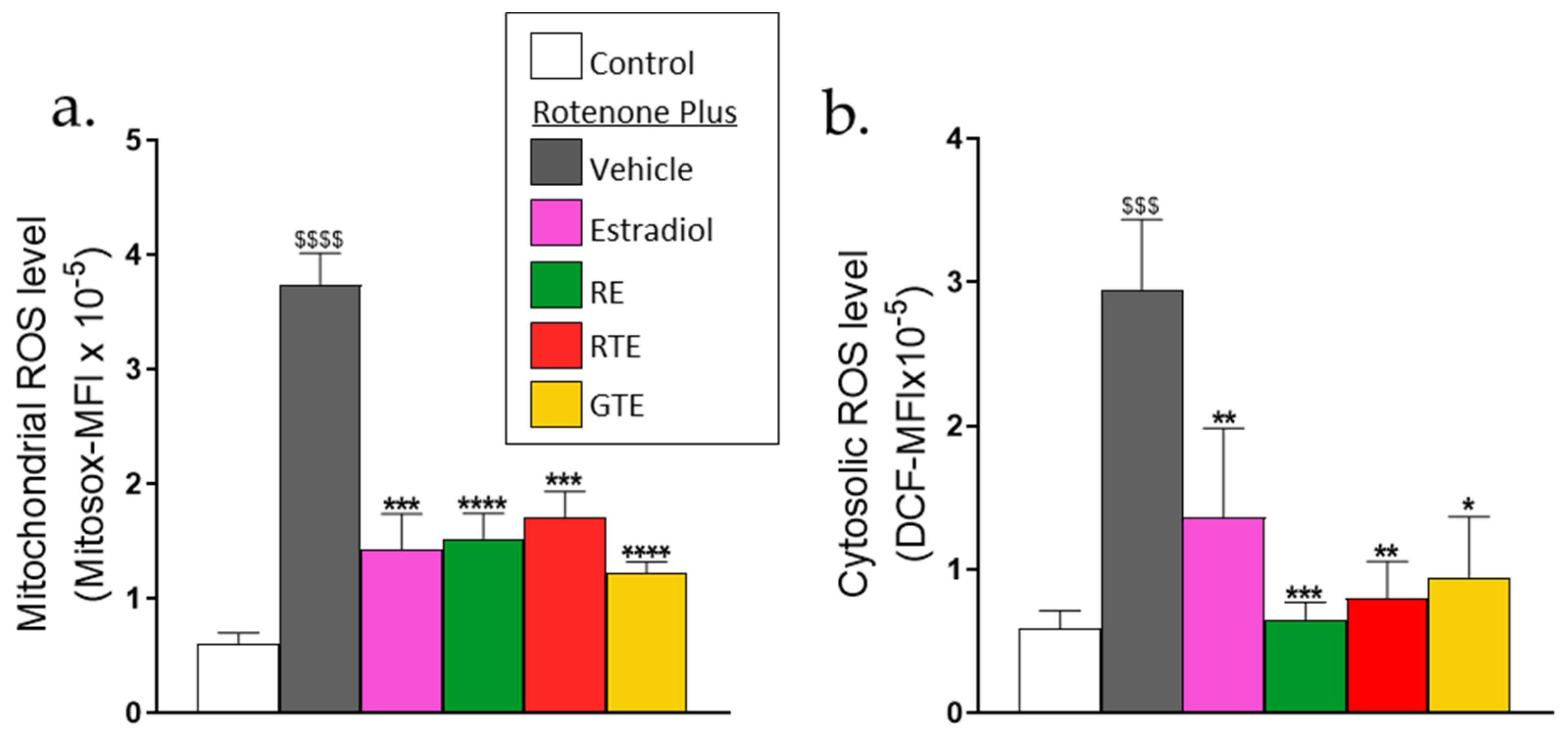
Figure 2.
Rotenone triggers apoptotic cell death of dermal fibroblasts, which can be reduced by tomato and rosemary extracts and estradiol. (a and b). Cell apoptosis was determined by annexin V flow cytometry. Cells were seeded in 6-well plates (a,b) (3 × 105 cells/well) and pre-incubated for 24 h with estradiol (10 nM), rosemary extract (RE; 10 μM carnosic acid), red tomato extract (RTE; 10 μM lycopene), or golden tomato extract (GTE; 40 μM phytoene). Then the medium was replaced with one containing the treatment compounds plus 1 µM rotenone and incubated for an additional 48 h. (a) Typical flow cytometric data of annexin V fluorescence. (b) Percent apoptotic cells. Results are the means ± SEM of five experiments, each performed in duplicate. (c) Apoptosis was analyzed by measuring caspase 3/7 activity using an ApoLive-GloTM Multiplex Assay. Cells were seeded in 96-well plates (5 × 103 cells/well) and treated as described above. Results are presented as a percent of the control. (d–h) Cell number and viability analysis. Cells were seeded in 96-well plates (5 × 103 cells/well) and treated as described above for a and b. At the end of the experiments, the media were removed for further analysis (see Figure 3), and viability or cell number was determined. (d,e) Comparison of cell death measured by cell number analysis with crystal violet (d) and cell viability by ApoLive-GloTM Multiplex Assay (e). (f,g) cell number was determined by crystal violet staining at the indicated concentrations of (f) rosemary extract, (g) red tomato extract, and (h) estradiol. Results are presented as percent of the control without rotenone and the treatment compounds. 100% of the cell number was 22,080 ± 3,153 cells/well. Values are the means ± SEM of four experiments, each performed in triplicate. $–$$$$, p < 0.05–0.0001, significant difference between the control and rotenone. *–****, p < 0.05–0.0001, significant difference between the vehicle and other treatments in the presence of rotenone.
Figure 2.
Rotenone triggers apoptotic cell death of dermal fibroblasts, which can be reduced by tomato and rosemary extracts and estradiol. (a and b). Cell apoptosis was determined by annexin V flow cytometry. Cells were seeded in 6-well plates (a,b) (3 × 105 cells/well) and pre-incubated for 24 h with estradiol (10 nM), rosemary extract (RE; 10 μM carnosic acid), red tomato extract (RTE; 10 μM lycopene), or golden tomato extract (GTE; 40 μM phytoene). Then the medium was replaced with one containing the treatment compounds plus 1 µM rotenone and incubated for an additional 48 h. (a) Typical flow cytometric data of annexin V fluorescence. (b) Percent apoptotic cells. Results are the means ± SEM of five experiments, each performed in duplicate. (c) Apoptosis was analyzed by measuring caspase 3/7 activity using an ApoLive-GloTM Multiplex Assay. Cells were seeded in 96-well plates (5 × 103 cells/well) and treated as described above. Results are presented as a percent of the control. (d–h) Cell number and viability analysis. Cells were seeded in 96-well plates (5 × 103 cells/well) and treated as described above for a and b. At the end of the experiments, the media were removed for further analysis (see Figure 3), and viability or cell number was determined. (d,e) Comparison of cell death measured by cell number analysis with crystal violet (d) and cell viability by ApoLive-GloTM Multiplex Assay (e). (f,g) cell number was determined by crystal violet staining at the indicated concentrations of (f) rosemary extract, (g) red tomato extract, and (h) estradiol. Results are presented as percent of the control without rotenone and the treatment compounds. 100% of the cell number was 22,080 ± 3,153 cells/well. Values are the means ± SEM of four experiments, each performed in triplicate. $–$$$$, p < 0.05–0.0001, significant difference between the control and rotenone. *–****, p < 0.05–0.0001, significant difference between the vehicle and other treatments in the presence of rotenone.
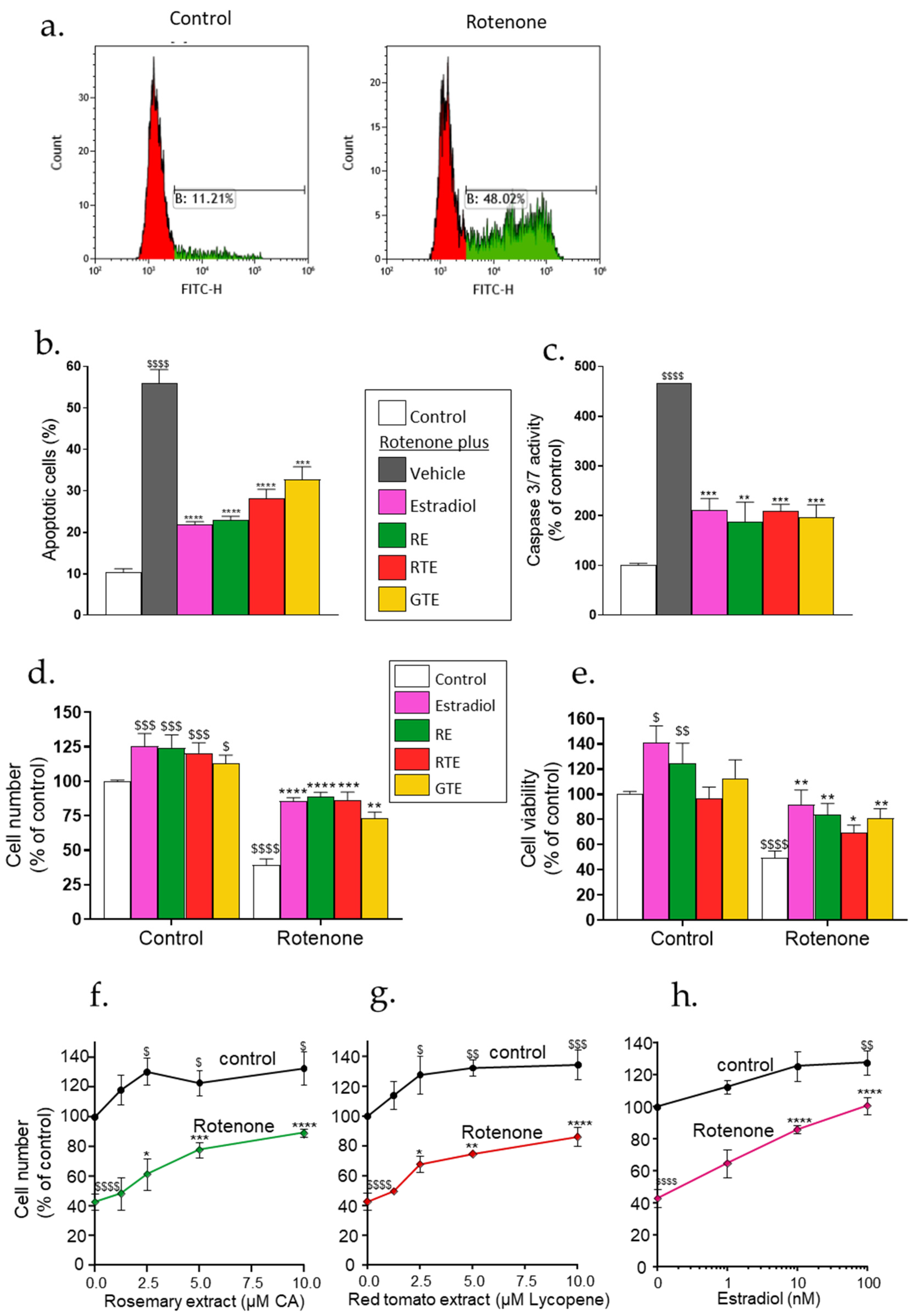
Figure 3.
Rotenone increases MMP1 secretion, and reduces collagen 1a1 secretion and mRNA expression, whereas tomato (RTE, GTE) and rosemary (RE) extracts and estradiol reverse these effects. Secretion of MMP-1 and pro-collagen 1a1 were determined in the media collected in the experiments described in Figure 2, using ELISA as described in the Methods section. (a) MMP-1 level. (b) Pro-collagen 1a1 level. Results are presented as percent of the control values. 100% of the MMP-1 secretion was 42.7 ± 6.4 pmol/1,000 cells. 100% of the pro-collagen 1a1 secretion was 80.9 ± 14.6 pmol/1,000 cells. (c) Collagen 1a1 mRNA expression was determined by RT-qPCR as described in the Methods section. Values are the means ± SEM of four experiments, each performed in triplicate. $$$$, p < 0.0001, significant difference between the control and rotenone. **–****, p < 0.01–0.0001), significant difference between the vehicle and other treatments in the presence of rotenone.
Figure 3.
Rotenone increases MMP1 secretion, and reduces collagen 1a1 secretion and mRNA expression, whereas tomato (RTE, GTE) and rosemary (RE) extracts and estradiol reverse these effects. Secretion of MMP-1 and pro-collagen 1a1 were determined in the media collected in the experiments described in Figure 2, using ELISA as described in the Methods section. (a) MMP-1 level. (b) Pro-collagen 1a1 level. Results are presented as percent of the control values. 100% of the MMP-1 secretion was 42.7 ± 6.4 pmol/1,000 cells. 100% of the pro-collagen 1a1 secretion was 80.9 ± 14.6 pmol/1,000 cells. (c) Collagen 1a1 mRNA expression was determined by RT-qPCR as described in the Methods section. Values are the means ± SEM of four experiments, each performed in triplicate. $$$$, p < 0.0001, significant difference between the control and rotenone. **–****, p < 0.01–0.0001), significant difference between the vehicle and other treatments in the presence of rotenone.
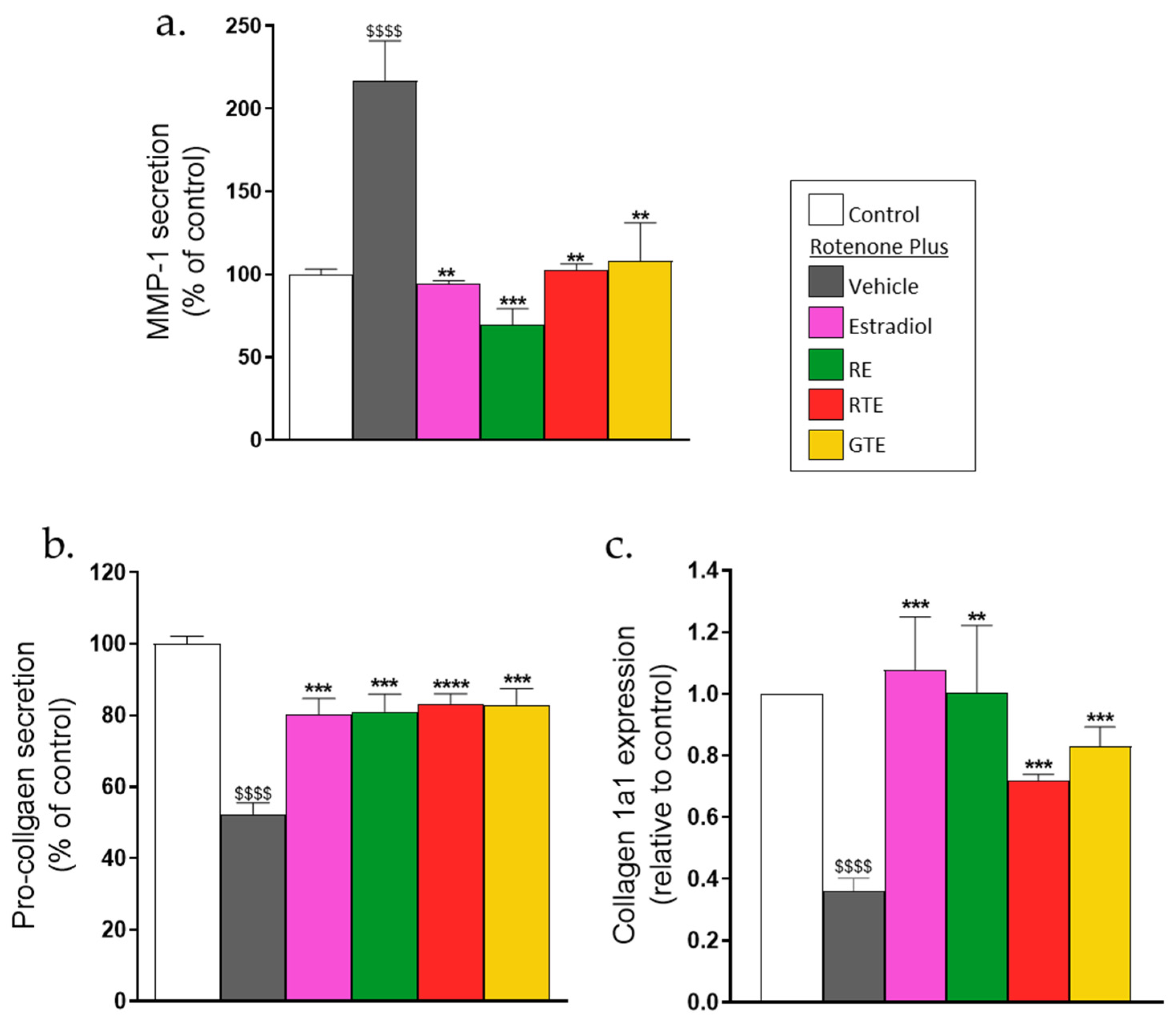
Figure 4.
Mitochondrial dysfunction following exposure to rotenone and recovery by dietary compounds and estradiol. Cells were seeded in XF 96-well plates (1 × 104 cells/well) and treated with the treatment compounds as described in Figure 1. 24 h after addition of rotenone, cells were washed for 1 h, and OCR and ECAR were measured by an Agilent SeahorseXFe96 analyzer according to the manufacturer’s protocol. OCR and ECAR were normalized to nuclei DAPI staining. (a) A representative OCR recording at basal conditions and after injection of oligomycin (1.5 μM), FCCP (1 μM), and antimycin A plus rotenone (1 μM). (b) Basal OCR results are expressed as pmol/minx1000 cells. Results of the following parameters (c–f) are expressed in the percentage of control (c) ATP-linked OCR. 100% = 8.04±0.46 pmol/minx1000 cells, (d) maximal respiration, 100% = 12.54±0.76 pmol/minx1000 cells, (e) spare respiratory capacity 100% = 4.53±0.49 pmol/minx1000 cells, and (f) ATP cellular level measured using the CellTiter-Glo Luminescent Cell Viability Assay Kit. (g) ECAR, results are expressed as mpH/minx1000 cells. Results are means ± SEM of four experiments, each in six replicates. $–$$$$, p < 0.05–0.0001, significant difference between the control and rotenone. ****, p < 0.0001), significant difference between the vehicle and other treatments in the presence of rotenone.
Figure 4.
Mitochondrial dysfunction following exposure to rotenone and recovery by dietary compounds and estradiol. Cells were seeded in XF 96-well plates (1 × 104 cells/well) and treated with the treatment compounds as described in Figure 1. 24 h after addition of rotenone, cells were washed for 1 h, and OCR and ECAR were measured by an Agilent SeahorseXFe96 analyzer according to the manufacturer’s protocol. OCR and ECAR were normalized to nuclei DAPI staining. (a) A representative OCR recording at basal conditions and after injection of oligomycin (1.5 μM), FCCP (1 μM), and antimycin A plus rotenone (1 μM). (b) Basal OCR results are expressed as pmol/minx1000 cells. Results of the following parameters (c–f) are expressed in the percentage of control (c) ATP-linked OCR. 100% = 8.04±0.46 pmol/minx1000 cells, (d) maximal respiration, 100% = 12.54±0.76 pmol/minx1000 cells, (e) spare respiratory capacity 100% = 4.53±0.49 pmol/minx1000 cells, and (f) ATP cellular level measured using the CellTiter-Glo Luminescent Cell Viability Assay Kit. (g) ECAR, results are expressed as mpH/minx1000 cells. Results are means ± SEM of four experiments, each in six replicates. $–$$$$, p < 0.05–0.0001, significant difference between the control and rotenone. ****, p < 0.0001), significant difference between the vehicle and other treatments in the presence of rotenone.
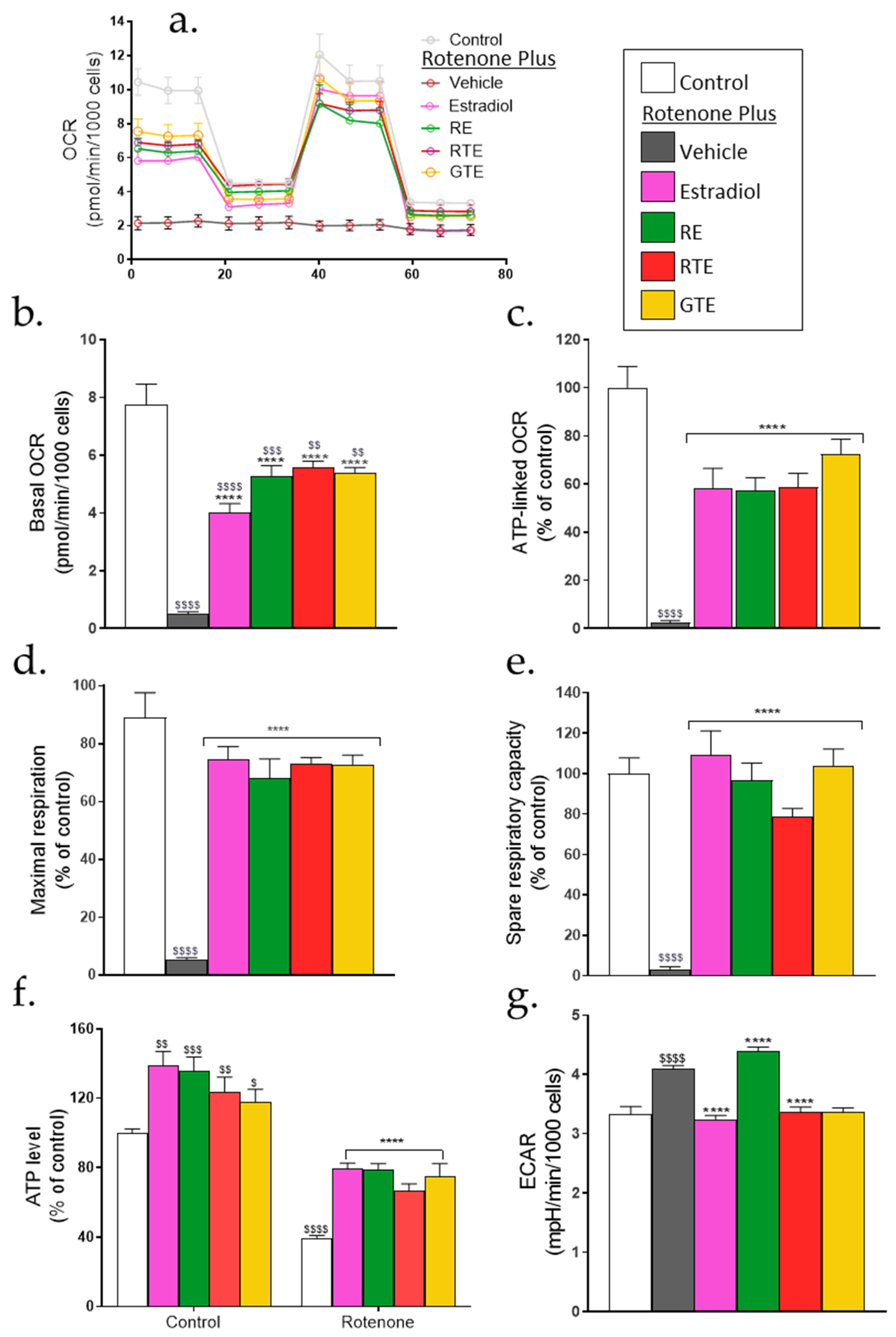
Figure 5.
Increased activation of ARE/Nrf2 by the combinations of rotenone with the phytonutrients or estradiol. (a,b) ARE/Nrf2 transcriptional activity. Cells were seeded in 24-well plates (105 cells/well), and 24 h later were transfected with the ARE/Nrf2 reporter gene. After transfection, cells were incubated for 16 h with rosemary extract (RE; 10 μM carnosic acid), red tomato extract (RTE; 10 μM lycopene), golden tomato extract (GTE; 40 μM phytoene), or estradiol (10 nM). ARE/Nrf2 transcriptional activity was determined using a luciferase reporter assay as described in the Methods section. Values (fold induction) are the means ± SEM of four experiments, each performed in triplicate. (c,d) NQO1 protein levels. Cells were seeded in 6-well plates (3 × 105 cells/well) and were treated as described in Figure 2. NQO-1 protein level was determined by western blot. (c) Quantitation of NQO1 level. (d) Representative western blot gel. Values (fold of control) are the means ± SEM of four experiments. $$$–$$$$, p < 0.001–0.0001, significant difference between the control and with rotenone. **–****, p < 0.01–0.0001), significant difference between the vehicle and other treatments in the presence of rotenone.
Figure 5.
Increased activation of ARE/Nrf2 by the combinations of rotenone with the phytonutrients or estradiol. (a,b) ARE/Nrf2 transcriptional activity. Cells were seeded in 24-well plates (105 cells/well), and 24 h later were transfected with the ARE/Nrf2 reporter gene. After transfection, cells were incubated for 16 h with rosemary extract (RE; 10 μM carnosic acid), red tomato extract (RTE; 10 μM lycopene), golden tomato extract (GTE; 40 μM phytoene), or estradiol (10 nM). ARE/Nrf2 transcriptional activity was determined using a luciferase reporter assay as described in the Methods section. Values (fold induction) are the means ± SEM of four experiments, each performed in triplicate. (c,d) NQO1 protein levels. Cells were seeded in 6-well plates (3 × 105 cells/well) and were treated as described in Figure 2. NQO-1 protein level was determined by western blot. (c) Quantitation of NQO1 level. (d) Representative western blot gel. Values (fold of control) are the means ± SEM of four experiments. $$$–$$$$, p < 0.001–0.0001, significant difference between the control and with rotenone. **–****, p < 0.01–0.0001), significant difference between the vehicle and other treatments in the presence of rotenone.
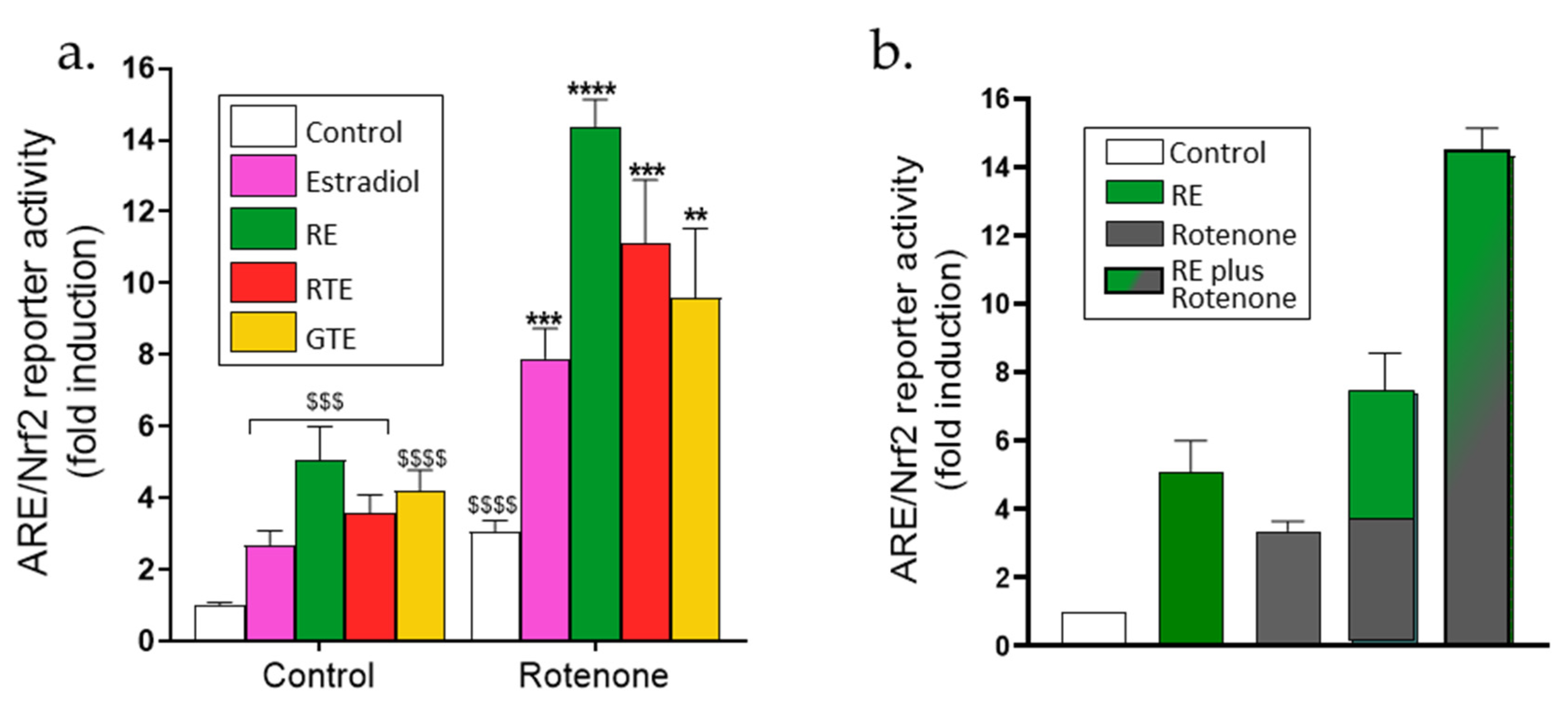
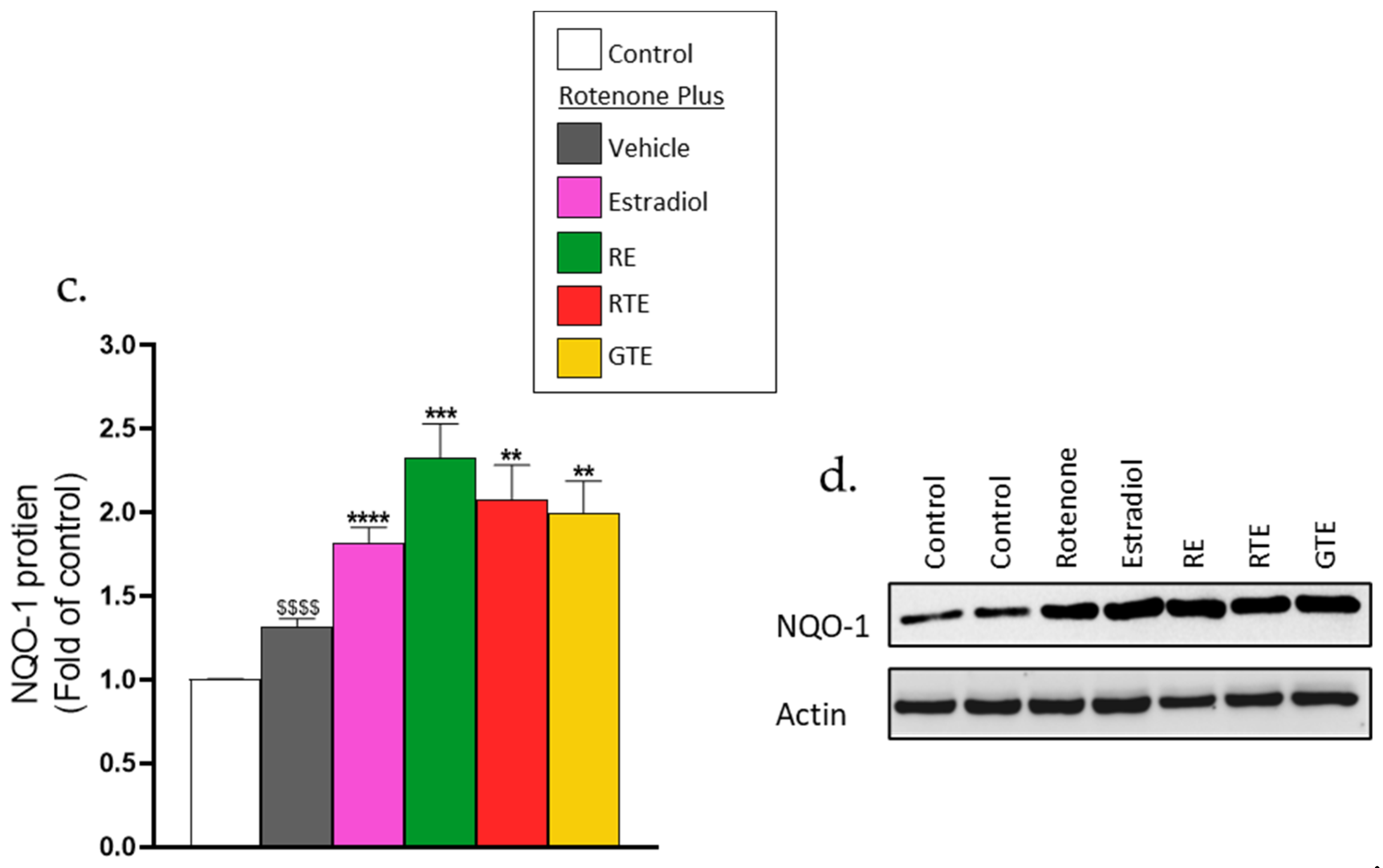
Figure 6.
Protection from rotenone-induced damage by dietary compounds and estradiol is markedly reduced in the presence of ARE/Nrf2 inhibitors. Cells were seeded in 96-well plates (5 × 103 cells/well) and treated essentially as described in Figures 1 to 3, but incubation was either with or without OTA (25 μM) or ML385 (10 μM). (a,b) ARE/Nrf2 transcriptional activity. This activity was measured in the presence of rotenone and rosemary extract (RE) at the indicated inhibitor concentrations. (c,d) Cytosolic ROS. Values are the means ± SEM of four experiments of averaged geometric means of fluorescence intensities (MFI) of DCF. (e,f) Cell number. (g,h) MMP-1 level. (i,j) Pro-collagen 1a1 level. Values (% of control) are the means ± SEM of four experiments. $$$$, p < 0.0001, significant difference between the control and rotenone; *-****, p < 0.05-0.0001), significant difference between the vehicle and other treatments in the presence of rotenone. ###–###, p < 0.001–0.0001), significant difference between the control and rotenone in the presence of the inhibitors. ^, p < 0.05, significant difference between the vehicle and other treatments in the presence of rotenone and the inhibitors.
Figure 6.
Protection from rotenone-induced damage by dietary compounds and estradiol is markedly reduced in the presence of ARE/Nrf2 inhibitors. Cells were seeded in 96-well plates (5 × 103 cells/well) and treated essentially as described in Figures 1 to 3, but incubation was either with or without OTA (25 μM) or ML385 (10 μM). (a,b) ARE/Nrf2 transcriptional activity. This activity was measured in the presence of rotenone and rosemary extract (RE) at the indicated inhibitor concentrations. (c,d) Cytosolic ROS. Values are the means ± SEM of four experiments of averaged geometric means of fluorescence intensities (MFI) of DCF. (e,f) Cell number. (g,h) MMP-1 level. (i,j) Pro-collagen 1a1 level. Values (% of control) are the means ± SEM of four experiments. $$$$, p < 0.0001, significant difference between the control and rotenone; *-****, p < 0.05-0.0001), significant difference between the vehicle and other treatments in the presence of rotenone. ###–###, p < 0.001–0.0001), significant difference between the control and rotenone in the presence of the inhibitors. ^, p < 0.05, significant difference between the vehicle and other treatments in the presence of rotenone and the inhibitors.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



