Submitted:
11 July 2024
Posted:
12 July 2024
You are already at the latest version
Abstract
The anatomy of mature vertebrate hearts is marked by a looped design, which is roughly characterized by two components: (1), s-shaped (sigmoid) routing of the cardiac flow path(s), and (2,) bilaterally asymmetric (chiral) routing of the cardiac flow path(s). The looped heart design is regarded as a phylogenetically conserved feature among vertebrates and is thought to represent a significant determinant of the cardiac pumping function. It evolves during the embryonic period of development by a morphogenetic process called “cardiac looping”. During the past decades, remarkable progress has been made in the uncovering of the genetic, molecular, and biophysical factors contributing to cardiac looping. Thereby, a special focus was directed on the evolution of a bilaterally asymmetric (chiral) heart configuration, which is said to play a central role in the determination of the alignments and separations of the pulmonary and systemic flow paths in the multi-chambered hearts of tetrapods. In contrast to the recent progress made in uncovering of the biological driver of cardiac looping, our present knowledge about the functional consequences of cardiac looping is at an immature state. This article provides an overview and discussion of the currently available information about the looped design of vertebrate hearts and its functional implications.
Keywords:
embryology
; cardiac looping
; vertebrate hearts
; fish hearts
; comparative anatomy
; functional morphology
; pumping function
1. Introduction
Cardiac looping is a morphogenetic process that occurs during the early stages in development of vertebrate embryonic hearts, when these hearts have the relatively simple design of an actively pulsating tubular blood vessel. Looping morphogenesis leads to the transformation of the initially straight embryonic heart tube into a looped tube whose geometric configuration is described in a simplified, two-dimensional fashion as an ‘S-shaped’ loop since the early 19th century [1,2,3,4,5]. The ‘S-shaped’ configuration of the vertebrate embryonic heart loop principally persists during the whole lifespan of vertebrates since it is the first manifestation of the curved and twisted flow paths found in mature vertebrate hearts. Due to the relatively large body size and easy accessibility of chick embryos, the first historically documented observations of embryonic heart loops were made in these higher vertebrates (for review see [6]). During the past 200 years, however, looping morphogenesis of the embryonic heart tube was observed in a wide spectrum of extant vertebrate species. This spectrum not only includes land-living vertebrates but also phylogenetically ‘primitive’ and ‘modern’ fish species [1,4,7,8,9,10,11,12]. Moreover, it was found that several components of the looping process principally ran the same way in all vertebrate species studied so far [6,13,14,15]. This observation not only provides the explanation for the current usage of zebrafish embryos as models for studying the genetic, cellular, and biophysical control of the early morphogenesis of the vertebrate heart [16,17,18,19]. It also suggests that cardiac looping may fulfill one or more phylogenetically conserved functions.
In the past, the fascination of developmental biologists for cardiac looping mainly has arisen from the fact that the embryonic heart is the first inner organ of vertebrates that adopts a bilaterally asymmetric configuration. Studying the looping embryonic heart may provide central pieces for completion of a still incomplete jigsaw puzzle, which, upon completion, may explain the development of visceral asymmetries in vertebrates. Moreover, it was found that disturbed left-right patterning of vertebrate embryos led to the development of severe congenital heart defects [15,20,21]. It is, therefore, no wonder that the current research on cardiac looping mainly focuses on the biophysical, cellular, and genetic factors driving the bilaterally asymmetric morphogenesis of the early embryonic heart [17,22,23]. Compared to these current ‘mainstream’ studies, only relatively few studies have focused on the functional significance of the looped design of embryonic and mature vertebrate hearts [24,25,26,27,28]. Thus our current knowledge about the functional consequences of this early morphogenetic process seems to lack behind the recent progress made in the uncovering of its biological drivers. The intent of the present article, therefore, is not only to provide an overview and discussion of the currently available information about the looped design of vertebrate hearts and its functional implications but also to stimulate future research in this area. As already mentioned, studies on fishes have significantly contributed to our current understanding of the biological drivers of cardiac looping. Here, I will show that studies on the cardiovascular system of fishes can also provide new insight into the functional consequences of cardiac looping. My article consists of three sections: (1), an overview on the early morphogenesis of vertebrate hearts with a special focus on comparative aspects of cardiac looping among vertebrates. (2), an overview on the comparative anatomy of mature vertebrate hearts, which focuses on the morphological features resulting from cardiac looping. (3), an overview and discussion on functional interpretations of the looped design of embryonic and mature vertebrate hearts.
2. Overview on the Early Morphogenesis of Vertebrate Hearts
In vertebrates, the heart is one of the first biological pumps to form and function during the embryonic period of development. The human embryonic heart, for example, starts its pumping action during the fourth week after fertilization (= sixth gestational week), which corresponds to the second week after the expected onset of the first missed menstrual bleeding [29]. During this early phase of prenatal cardiogenesis, the shape and the pumping action of vertebrate hearts differ markedly from that found in mature hearts. The latter are characterized by the presence of chambers and valves [30], and their mode of generating unidirectional blood flow may be characterized as valve supported pumping [27]. The early embryonic heart of vertebrates, in contrast, has the design of a tubular blood vessel that lacks valves. Its mode of pumping action, therefore, has been characterized as valveless pumping (for review see [31]). The valveless heart tube of vertebrate embryos arises from the union of bilaterally paired areas of the lateral plate mesoderm along the ventral midline of the foregut [32]. These areas of the lateral plate mesoderm are called the left and right heart fields. Their union starts with the formation of a straight heart tube, which is aligned along the body midline and becomes connected to the venous and arterial branches of the embryonic cardio-vascular system at its caudal and cranial ends, respectively. The future atrial and ventricular heart chambers, therefore, have an original caudal (atria) and cranial (ventricles) identity. Subsequent to its establishment, the straight heart tube elongates by the continuous addition of new heart field-derived material to its arterial and venous poles. These heart field-derived materials nowadays are named the anterior and posterior second heart fields, while the materials forming the first portion of the embryonic heart tube are named the first heart field [33]. The elongation of the tubular embryonic heart is accompanied by striking changes in its configuration. The initially straight and bilaterally almost symmetric tube now adopts the configuration of a looped tube. The fully looped heart tube is frequently called the ‘S-shaped’ heart loop since its shape is said to resemble the Latin letter S when viewed in a frontal projection. The S-shaped heart loop is a handed geometrical object. This means that it has a bilaterally asymmetric configuration, which principally can occur in two chiral variants, named the D-loop and the L-loop enantiomorph. In the D-loop enantiomorph, the ventricular (cranial) portion of the S-shaped heart loop is bend toward the right side of the body (dextro-loop), while the ventricular portion of the L-loop enantiomorph is bend toward the left body side (levo-loop). In all vertebrate species studied so far there is normally a very strong bias towards the development of the D-loop enantiomorph, so that the spontaneous occurrence of L-loops is normally a rare event (see Figure 4 in [14]). The D-loop enantiomorph of vertebrate embryonic hearts, therefore, is classified as a fixed or directional asymmetry, what strongly suggests that the direction of asymmetric heart looping is inherited [14].
2.1. Comparative Aspects of Cardiac Looping among Vertebrates
As already mentioned, looping morphogenesis of the tubular embryonic heart was observed in embryos from all classes of extant vertebrates (fishes, amphibia, reptiles, birds, and mammals) studied during the past 200 years. It is, therefore, generally said that cardiac looping is a phylogenetically conserved morphogenetic process among vertebrates. The observations made during the past 200 years, however, have also shown that the morphological phenotype of ‘S-shaped’ heart loops sometimes seems to differ considerably between different species (Figure 1).
To make easier the comparison of cardiac looping between different species, I find it helpful to characterize the complex positional and morphological changes of the looping embryonic heart tube with regard to the main body axes of vertebrates. We, thereby, can distinguish between four different components of cardiac looping:
- (1), positional and morphological changes along the original dorso-ventral heart axis.
- (2), positional and morphological changes along the cranio-caudal body axis.
- (3), positional and morphological changes along the so-called left-right body axis, which lead to a bilaterally asymmetric heart shape. This component may be named “lateral looping” or “chiral looping”.
- (4), positional and morphological changes, which reduce the degree of lateral/chiral looping reached at earlier stages of cardiac looping. This component may be named “final positional adjustments.”
The above mentioned list of four looping components should not be understood as showing the normal developmental sequence of cardiac looping. Some of these changes can occur, in species-specific manners, at the same time. The purpose of the above-described listing of artificially separated events is to make easier the understanding of the early morphogenesis of vertebrate hearts. In the following, the four components of cardiac looping will be described in detail using a simplified geometrical model that consists, from the beginning, of all building blocks of the embryonic heart.
(1), dorso-ventral looping: this component is mainly characterized by a change in the positional relationship of the future atrial and ventricular heart chambers along the original dorso-ventral heart axis. Initially, this axis corresponds to the future dorso-ventral body axis. However, during lateral/chiral looping (see below), which frequently coincides with dorso-ventral looping, the original dorso-ventral polarization of the heart tube becomes obscured by a rightward rotation. The present scheme of dorso-ventral looping (Figure 2), therefore, shows an artificial situation, which omits the changes caused by lateral/chiral looping.
In the pre-looping state, the atrial and ventricular portions of the heart tube are aligned along the cranio-caudal axis with the atrial portion lying caudal to the ventricular portion. The heart tube, therefore, appears as a straight tube. During dorso-ventral looping, the ventricular portion shifts ventrad and the atrial portion shifts dorsad. These positional changes are driven mainly by ventral bending of the ventricular portion of the heart tube. In consequence of dorso-ventral looping, the atrial portion acquires a dorso-caudal position in relation to the ventricular portion, and the entire heart tube and its main flow path start to adopt the shape of the Latin letter S.
(2), cranio-caudal looping: the positional and morphological changes characterizing this component may be described as ascensus of the atrial portion and/or descensus of the ventricular portion of the heart loop along the cranio-caudal body axis (Figure 3).
The atrial ascensus/ventricular descensus brings the atrial portion of the heart tube into its mature topographical relation to the ventricular portion, which is said to be dorsal to the ventricle in most fishes, and dorso-cranial to the ventricle in tetrapods [28,30,34,35]. Atrial ascensus/ventricular descensus, forces the S-shaped deformation of the heart loop and produces the U-shaped ventricular flow path (U-turn loop) usually found in the mature heart of higher vertebrates [28,35]. Observations on sturgeon embryos have shown that the outcome of cranio-caudal looping strongly depends on the growth dynamics of the embryonic pericardial cavity. In this species, cardiac looping is interrupted by a developmental phase in which the already acquired configuration of an S-shaped loop practically disappears due to a stretching of the heart caused by an increase in the cranio-caudal length of the pericardial cavity. The definitive ‘S-shaped’ loop then is formed during a subsequent phase of forced elongation of the heart [11].
(3), lateral or chiral looping: the positional and morphological changes characterizing this component may be described, in a simplified fashion, as resulting from rightward rotation of the heart loop around its original longitudinal (cranio-caudal) axis. Chiral looping differs remarkably between embryonic hearts in which the ventricular portion is connected to the future arterial root only by a vestigial myocardial outflow element (hearts of cyclostomes and modern ray-finned fishes) and those in which the ventricular portion connects to the future arterial root via a substantial myocardial outflow element (hearts of cartilaginous fishes, primitive ray-finned fishes, lobe-finned fishes, and tetrapods). In the first group, rightward rotation affects the entire heart tube. As a consequence, its ventricular portion comes to lie to the right of the body midline and its atrial portion comes to lie to the left of the body midline (Figure 1A and A).
The ‘S-shaped’ heart loop of these animals shows only slight twisting around the atrio-ventricular canal [19] so that these hearts appear as a plane S rather than a three-dimensional geometrical object (A, and A). In the second group (hearts of cartilaginous fishes, etc. … ), only the ventricular portion and its myocardial outflow element become lateralized whereas the atrial portion of the heart loop does not undergo rightward rotation (Figure 4B). The looping hearts of these animals, thus, are subjected to considerable torsion so that their S-shaped configurations tend to resemble helically wound tubes rather than plane geometrical objects. Moreover, due to the fact that the myocardial outflow element (the primordium of the so-called conus arteriosus of hearts of amphibia and some fish species) becomes displaced to the right of the body together with the ventricular bend, the straight outflow element changes its axial orientation from an initially vertical towards a transverse axis (Figure 5B1). The most complex form of helical heart looping is found in lungfish and tetrapods. The embryonic heart loops of these animals do not only develop a counterclockwise helical winding of the ventricular bend and its inflow portion, but, additionally, develop a clockwise helical winding of the ventricular outflow element (B and B2).
(4), final positional adjustments (Figure 6): the positional changes characterizing this component may be described as a kind of back-rotation or untwisting of the heart loop, which leads to a reduction in the degree of lateral/chiral looping reached at earlier stages of cardiogenesis. Untwisting of the heart loop was first observed in teleost fishes (Cyprianus blicca) by the Baltic German scientist Karl Ernst von Baer in 1835 [1]. Compared to lateral/chiral looping, however, this looping component did not receive much attention. It may be no wonder, therefore, that it was later rediscovered several times in diverse teleosts [7,38,39,40]. Untwisting of the developing heart not only was observed in modern ray-finned fishes but also in lungfish [9], and higher vertebrates [6,41,42]. In teleost fishes, untwisting of the developing heart is observed at the transition between the embryonic and larval periods of development [38,39,40], while in higher vertebrates it occurs at embryonic stages [6,42]. At the end of untwisting, the developing heart chambers and great vessels normally have reached an approximation of their definitive topographical relationships.
3. Comparative Anatomy of the Looped Design of Mature Vertebrate Hearts
Comparative anatomical data disclose some striking, species- or class-specific differences in the situs of mature vertebrate hearts. These differences may be explained by differences in the extent to which the above-described looping components have contributed to the definitive positioning of the heart chambers. The present comparative analysis of the definitive heart situs of vertebrates will focus mainly on two aspects. These aspects are, firstly, those positional relationships of the atrial and ventricular chambers that primarily cause the ‘S-shaped’ (sigmoid) configuration of the main course of the cardiac flow path(s), and, secondly, those anatomical features that cause a bilaterally asymmetric morphology of the hearts responsible for a twisting of the sigmoid cardiac flow path(s). The first aspect may be named the “sigmoid routing of the main cardiac blood stream(s)”. Its status reflects mainly the extent to which the dorso-ventral and cranio-caudal looping components have contributed to the definitive heart situs. The second aspect may be named the “bilaterally asymmetric routing” or “chiral routing” of the cardiac blood stream(s). Its status reflects mainly the extent to which lateral/chiral looping and final positional adjustments (untwisting) have contributed to the definitive heart situs.
3.1. Comparative Anatomy of the Sigmoid Routing of the Flow Path(s) of Mature Vertebrate Hearts
Sigmoid routing of the main cardiac flow path(s) was found in the mature heart of all extant vertebrates studied so far [24,30,34,35]. Sigmoid routing of the cardiac blood stream(s), therefore, is a phylogenetically highly conserved feature among vertebrates. The degree of sigmoid routing, however, was found to differ considerably between the species examined. In contemporary articles on the evolution of the vertebrate heart, the degree of sigmoid routing of the cardiac blood stream(s) frequently is said to reflect the position of the species along the phylogenetic tree of vertebrates [30,34,35]. Thereby, the hearts of basal vertebrates (cyclostomes) are shown with the lowest degree of sigmoid routing, mainly resulting from a ‘primitive’ atrial position that is dorso-caudal to the ventricle. The ventricular flow path of these hearts makes roughly a 45° turn from the atrio-ventricular to the arterial valve (ventricular J-turn loop). The hearts of jawed fishes are shown with a moderate degree of sigmoid routing, resulting from an atrial position that is dorsal to the ventricle. The ventricular flow path of these hearts makes roughly a 90° turn from the atrio-ventricular to the arterial valve (ventricular L-turn loop). The hearts of tetrapods (amphibia, reptiles, birds, mammals) usually are shown with the highest degrees of sigmoid routing, resulting from an atrial position that is dorso-cranial to the ventricle(s). The ventricular flow paths of these hearts make roughly 120° to 150° turns from the atrio-ventricular to the arterial valves (ventricular U-turn loop).
I should note here, however, that a critical review of original data, published during the past 200 years, shows that the above-described phylogenetic scenario does not completely match with the reality. With regard to basal vertebrates (cyclostomes), we can note that the atrium of the mature hagfish heart indeed lies in the above-described ‘primitive’ position [43,44,45]. The atrium of the mature heart of lampreys, however, is found at the same cranio-caudal level as the ventricle [44,46,47], which is the position usually assigned to jawed fishes. The atrium of the mature heart of jawed fishes, on the other hand, is not regularly found at the same cranio-caudal level as the ventricle. Among jawed fish species, we can rather find a spectrum of various degrees of sigmoid routing of the main cardiac blood stream (Figure 7).
This spectrum ranges from low degrees, resulting from an atrial position that is dorso-caudal to the ventricle (Figure 7A) (e.g. coelacanths, see [48]; Lophius piscatorius, see [52]), to high degrees, resulting from an atrial position that is dorso-cranial to the ventricle (Figure 7D) (e.g. lepisosteiformis, amiiformis, see [51]; Monopterus albus see [53,54]). Thus, a high degree of sigmoid routing of the main cardiac blood stream(s) is not exclusively found in the mature hearts of tetrapods, which have structurally or functionally separated flow paths for deoxygenated (pulmonary) and oxygenated (systemic) blood. It can also occur in some fish species, whose two-chambered hearts have only a single undivided flow path.
In contrast to the high range of variation in the extent of sigmoid routing of the cardiac blood streams found in fishes, I could not find any report on tetrapod hearts with a low or moderate degree of sigmoid routing of the cardiac flow path(s). A high degree of sigmoid routing of the cardiac blood streams thus seems to be indeed a common feature of the mature heart of tetrapods.
3.2. Comparative Anatomy of the Bilaterally Asymmetric (chiral) Routing of the Flow Path(s) of Mature Vertebrate Hearts
It is generally stated in the biomedical literature that the inner organs of vertebrates display a bilaterally asymmetric anatomy and that the heart is the first inner organ of vertebrates to adopt a bilaterally asymmetric shape during ontogenesis [55,56,57]. Moreover, it is a well-known fact that the bilaterally asymmetric anatomy of the human heart provides the basis for the structural division of its lumen into pulmonary and systemic flow paths. It, therefore, may be no wonder that the mature heart of vertebrates generally is seen as an organ of bilaterally asymmetric anatomy [56,58,59]. While this view is valid for the hearts of tetrapods, the situation in fishes does not match with it. Based on the degree of bilateral asymmetry, the morphological phenotypes of fish hearts can be roughly assigned to three different groups: (1), hearts with a visually conspicuous bilateral asymmetry; (2), hearts with a visually obscured bilateral asymmetry; and (3), hearts with a nearly perfect bilateral symmetry (Figure 8).
Bilaterally asymmetric routing of the cardiac flow path frequently can be noted already at the first visual examination of the outer shape of a fish heart. This is relatively easy in those fish hearts that have a relatively long myocardial outflow element called the conus arteriosus. The conus arteriosus connects the ventricle to the bulbus arteriosus, which is the intrapericardial root of the ventral aorta. The bulbus arteriosus and the ventral aorta are normally aligned in the midsagittal body plane. Alignment of the conus arteriosus in the midsagittal body plane together with the bulbus arteriosus and the ventral aorta is highly suggestive for the presence of bilateral symmetry (Figure 7A3). Deviations of the long axis of the conus arteriosus from the midsagittal body plane, on the other hand, indicate bilaterally asymmetric routing of the cardiac flow path (Figure 7A1).
There is a striking association between the degree of bilateral cardiac asymmetry and the body shape of fishes. Pronounced left-right asymmetry of the heart is frequently found in those fishes that show a flattening of their body along the dorso-ventral body axis (depressed cross-sectional body shape), such as rays and angel sharks. Hearts with a nearly perfect bilateral symmetry, on the other hand, can be found in fishes whose bodies have a fusiform cross section, such as tunas and gars. It is likely that a dorso-ventrally flattened body limits the dorso-ventral dimensions of the pericardial cavity but supports the lateral (left-right) expansion of the pericardial cavity. The establishment of such a spatial condition during the larval stages of development may have forced the post-embryonic heart to retain its embryonic state of bilateral asymmetry. A fusiform body cross-section, on the other hand, limits the lateral (left-right) dimensions of the pericardial cavity but supports the dorso-ventral expansion of the pericardial cavity. The establishment of such a spatial condition during the larval stages of development may have forced the post-embryonic heart to rotate back to its original orientation along the dorso-ventral body axis and thereby to loose its state of bilateral asymmetry reached at the end of the embryonic period of development.
3.2.1. Hearts with a Visually Conspicuous Bilateral Asymmetry
The hearts of cartilaginous fish and some ‘primitive’ ray finned fish, such as sturgeons, show conspicuous bilateral asymmetry (Figure 8A1). The conus arteriosus of these hearts is a relatively long and straight myocardial tube that originates from the right side of the ventricle and runs in an oblique course to the bulbus arteriosus. Moreover, the atrio-ventricular valve opens into the left portion of the ventricle, so that the ventricular inflow and outflow orifices are arranged lateral to one another (Figure 8B1) [61,63,64]. The ventricle of such hearts, therefore, was described as displaying a “looped appearance” [63]. Asymmetric routing of the ventricular flow path can become apparent not only by a lateral displacement of the origin of a long conus arteriosus, but also by a helical deformation of its shape. This situation is found in lungfish [51,65] and is associated with a striking discrepancy between the length of the systolic ventricular flow path and the greatest length of the pericardial cavity (Figure 9).
Apart from an asymmetric arrangement of the ventricle and its outflow tract, a visually conspicuous asymmetry of a mature fish heart can also be characterized by a primitive left-right positioning of the two heart chambers. In cyclostomes, for example, the atrium is frequently said to lie to the left of the ventricle [44,45,46,47]. The mature hearts of these fishes obviously seem to retain their maximum degree of embryonic heart loop lateralization during the whole post-embryonic lifespan of the animals. This suggests that the fourth component of cardiac looping (back-rotation/untwisting) does not significantly contribute to the mature cardiac phenotype in these species. I should note here, however, that there exist some conflicting data about the final left-right positioning of the heart chambers of cyclostomes. Some authors have reported that the left-sided atrium of the embryonic lamprey heart becomes placed more dorsally to the ventricle at later stages of development [66,67] and another group has reported that the mature hagfish heart occupies a midline position in the body and that its atrium lies dorsal and caudal to the ventricle [43]. These findings are at variance to the above-mentioned observations and would suggest that the developing heart of cyclostomes may undergo untwisting. Future studies are needed to clarify the situation.
3.2.2. Hearts with a Visually Obscured Bilateral Asymmetry
In contrast to cartilaginous fishes and ‘primitive’ ray-finned fishes, ‘modern’ ray finned fishes (teleosts) have only a very short conus arteriosus [68,69,70]. Due to its small length, the axis of this ring-shaped outflow element is always aligned in the midsagittal body plane where it connects the ventricle with the bulbus arteriosus (Figure 7A2). Such a situation can obscure the presence of asymmetric routing of the cardiac flow path if the hearts do not display additional, externally visible signs of bilateral asymmetry, such as a left-sided position of the atrium (see above). It, therefore, may be no wonder that the hearts of teleost fishes frequently have been described as bilaterally symmetric structures [44,61,63]. Analysis of the internal anatomy of such hearts, however, frequently discloses an asymmetric position of the central portion of the atrium dorsal and to the left of the ventricle in combination with a left-sided position of the atrio-ventricular orifice (Figure 7B2) [71,72,73].
3.2.3. Hearts with a Nearly Perfect Bilateral Symmetry
In several species of ray-finned fishes (holostei, e.g. lepisosteiformis, amiiformis, as well as teleostei, e.g. tuna), the mature heart occupies a midline position in the body of the animals and appears as a nearly perfect, bilaterally symmetric structure (Figure 7A3). In these hearts, the atrium lies dorsal or dorso-cranial to the ventricle and the atrio-ventricular canal, the ventricular outflow tract, and the ventral aorta, all are aligned in the midsagittal body plane (Figure 7B3) [51,61,74]. We have no embryological data from lepisosteus or tuna fishes that can answer the question as to whether the nearly perfect symmetry of their mature hearts results from a physiological lack of lateral/chiral looping of their embryonic hearts or from the complete elimination of an already adopted bilaterally asymmetric shape during the process of final positional adjustments. Embryological data from Amia calva, however, show that the embryonic hearts of this fish species undergoes S-looping as well as lateral/chiral looping in the same way as in other fishes [75]. This finding suggests that the mature fish hearts with nearly perfect bilateral symmetry have completely lost their embryonic state of lateral/chiral looping, while they have retained their embryonic state of dorso-ventral and cranio-caudal looping. It is likely that the loss of bilateral asymmetry is accomplished by a complete back-rotation of the developing heart during the process of final positional adjustments. In these fishes, the existence of a bilaterally asymmetric heart seems to be a transitory phenomenon that is confined only to the embryonic period of development.
4. The Functional Significance of the Looped Design of Embryonic and Mature Vertebrate Hearts
If we want to clarify the functional significance of the design of an inner organ or a technical device, we should consider at least three different possibilities: (1), a certain feature of the design may make importance contributions to the specific function(s) of the organ/technical device under investigation. Cardiac valves, for example, are important for the main function of the heart, namely the generation of unidirectional blood flow. (2), some aspects of the positioning and shaping of an organ/technical device may not directly contribute to its specific function(s), but merely represent functional solutions of other demands; e.g. a packing problem or the protection of the organ against physical damage. The impressive folding of the surface of our brain, for example, is the solution of the problem of packing a large area of cerebral cortex into the physically confined space of our skull. Transverse or longitudinal mounting of an engine into a vehicle does not have direct impact on the efficiency of force production of the engine but merely can be explained by other functional demands; e.g. transverse mounting can solve the problem of packing an engine into a short car. (3), some organs or some of their form features may not have any function at the time point of investigation. They may represent actually functionless vestiges from earlier phases of individual life (e.g. the belly button) or from earlier phases of phylogeny.
Functional interpretations of the looped design of vertebrate hearts have generally focused on its possible role in the pumping function of the heart. The possibility that a given feature of the looped design may represent a solution of other functional demands of the body, such as a packing problem, or may represent a functionless vestige from earlier stages of ontogenesis or phylogenesis, was largely ignored by the scientific community. The diverse concepts about the functional consequences of cardiac looping, can roughly be assigned to two different functional aspects: (1), the determination of the definitive spatial arrangement of the building units of the heart (chambers, connecting elements, and great blood vessels), which dictates the alignments and separations of the systemic and pulmonary flow paths within the multi-chambered hearts of tetrapods; and (2), the optimization of the cardiac pumping efficiency.
4.1. A Closer View on the Functional Design of Embryonic Vertebrate Hearts
In the past, concepts about the functional significance of the looped design of vertebrate hearts usually focused on the mature heart. The possibility that cardiac looping might have any effect on the pumping function of the valveless embryonic heart tube was ignored by most developmental biologists, since it was thought that the looped design of the vertebrate embryonic heart tube has functional consequences only at advanced stages of development, when the heart has acquired the mature phenotype of a chambered heart [76]. The long-lasting prevalence of the paradigma of late-onset effects of cardiac looping, is astonishing since the looped heart tube shows several form features (e.g. s-shaped curvatures, kinking, helical coiling, torsion) that are known for their potential to significantly change the hemodynamics of circulatory systems at macro-vascular as well as micro-vascular scales (reviewed by [27]). Moreover, the appearance of these form features, during embryonic cardiogenesis, coincides with the onset of a hemodynamically effective blood circulation [6,77,78] and data from zebrafish mutants suggest that failure of heart looping can reduce the pumping efficiency of embryonic hearts [79,80]. Based on these and other arguments, my group has postulated that the looped design of the tubular heart of vertebrate embryos might improve the efficiency of valveless pumping. The physical plausibility of our hypothesis was tested on a valveless pump model in which a unidirectional fluid flow was generated by the so-called Liebau effect, which is a pumping mechanism suspected to generate blood flow in the vertebrate embryonic heart tube [81]. It was found that, under the same conditions of actuation, a looped configuration of our tubular pump generated higher maximum pressure heads and higher average flow rates than the straight configuration [27]. This data principally confirmed the physical plausibility of our hypothesis. Unfortunately, however, we could not identify the mechanism responsible for the improvement of the pumping efficiency. It might be possible that the kinks in our valveless heart loop model, which are typical features of the looped heart tube of vertebrate embryos, have acted in a valve-like manner. Furthermore, I have to note, that at the present time, it is unclear as to whether the valveless heart tubes of vertebrate embryos generate unidirectional blood flow via the above-mentioned Liebau effect or via peristalsis [31]. Thus, our initial simulation experiments should be complemented by future studies aimed to clarify whether a looped design might also improve the efficiency of peristaltic pumps. In view of our initial data from physical models, however, I think that we should no longer ignore the possibility that looping morphogenesis might optimize the pumping function of the valveless heart tube of vertebrate embryos. Moreover, if we assume that the mature heart of vertebrates has phylogenetically evolved from the tubular heart of a vertebrate ancestor [82], similar to the situation found in extant urochordates such as Ciona intestinalis, we can postulate that the phylogenetically oldest function of cardiac looping might have been the improvement of the pumping efficiency of the tubular heart of vertebrate ancestors.
4.2. A Closer View on the Functional Design of Mature Vertebrate Hearts
For the understanding of the functional significance of the looped design of mature vertebrate hearts, it is helpful to roughly distinguish between two different groups of hearts; the two-chambered hearts and the multi-chambered hearts. Two-chambered hearts represent the basic design of the mature vertebrate heart. They are found in all groups of fishes with the exception of lungfish. Two-chambered hearts consist of a single atrial chamber, which receives the blood from a confluence of veins called the sinus venosus, and a single ventricular chamber, which is connected with the atrium via the so-called atrio-ventricular canal and pumps the blood into the ventral aorta via ventriculo-aortic connecting elements (conus arteriosus, bulbus arteriosus). The two-chambered hearts of vertebrates have only a single intra-cardiac flow path, which transports either deoxygenated (gill-breathing fishes) or mixed blood (air breathing fishes) [83]. The group of multi-chambered hearts comprises three- (two atria and one ventricle) and four-chambered (two atria and two ventricles) hearts. The common feature of these hearts is the presence of two intra-cardiac flow paths, which are functionally or structurally separated from each other. One path directs the oxygenated blood from the lungs into the systemic circuit and the other directs the deoxygenated blood from the systemic veins into the pulmonary circuit. Multi-chambered hearts have evolved from two-chambered hearts during vertebrate evolution and, therefore, represent the phylogenetically younger variants of the vertebrate heart design. They are found in lungfish and tetrapods. Proper function of such hearts depends on hemodynamically or structurally correct alignments and separations of their intra-cardiac flow paths.
4.2.1. Looping Dictates the Alignments and Separations of the Systemic and Pulmonary Flow Paths
Since the early 20th century, the major function of the embryonic heart looping is seen in dictating the future alignments and separations of the pulmonary and systemic flow paths within the mature, multi-chambered heart [41,42,57,84,85,86,87]. In this context, especially the lateral/chiral component of cardiac looping is seen as essential for the determination of the atrio-ventricular as well as ventriculo-arterial connections (Figure 10).
The strong and long-lasting focus on this functional consequence of cardiac looping may be explained by a wealth of evidence indicating that abnormal looping of the embryonic heart, especially disturbances in lateral/chiral looping, play a central role in the pathogenesis of complex congenital heart defects with abnormal alignments and separations of the cardiac flow paths [88,89,90,91,92]. I should note, however, that generations of embryologists, comparative morphologists, and pathologists obviously seem to have ignored the fact that this functional interpretation of cardiac looping can be valid only for the multi-chambered hearts of tetrapods and lungfish. The two-chambered hearts of fishes have only a single undivided flow path and their chambers normally are properly connected with each other and with the arterial and venous branches of the circulation before the embryonic heart starts its looping [93]. Dictating the future alignments and separations of the pulmonary and systemic flow paths within the mature, multi-chambered heart of vertebrates, therefore, cannot be regarded as a phylogenetically conserved function among vertebrates (Figure 11) but merely should be regarded only as a phylogenetically, relatively young function of cardiac looping. Therefore, the question is: what are the phylogenetically oldest functions of the looped heart design that are conserved in the two-chambered hearts of fishes as well as in the multi-chambered hearts of lungfish and tetrapods? The answer should be found in the group of concepts that see the functional consequences of the looping process in the optimization of the pumping performance of the mature vertebrate heart.
4.2.2. Looping Might Optimize the Cardiac Pumping Efficiency
Up to the end of the 20th century, several researchers had speculated that the looped design of the vertebrate heart might improve its pumping function. Unfortunately, however, none of these researchers had presented a physically sound concept about the way by which a looped configuration might improve the cardiac pumping function. The scene changed in 2000 when Philip Kilner and co-authors presented, for the first time, physically sound concepts that were based on magnetic resonance phase-velocity mapping of the blood flow patterns in the human heart [24,25,94]. During the past 25 years, Kilner’s concepts were supplemented by further physically sound hypothesis [26,28] so that, at the present time, we can distinguish between five postulated benefits for the pumping function of vertebrate hearts. From these five postulated advantages, only three may be valid in the mature hearts of fishes as well as tetrapods, while two postulated advantages seem to be valid only in tetrapods. The latter two advantages were published by the group of Bruno Marino [26] and the group of Mark Sherrid [28].
Marino and co-workers have noted that lateral/chiral looping does not only seem to dictate the future atrio-ventricular and ventriculo-arterial connections of the mature heart of tetrapods. It, furthermore, causes the ‘spiral’ (helical) arrangement of the ventricular outflow tracts and arteries normally found in the mature heart of higher vertebrates. They have speculated that this chiral form feature may produce a hemodynamically advantageous situation. Computational fluid dynamics simulation and three-dimensional flow imaging indeed have shown that the ‘spiral’ (helical) pattern of the great arteries has fluid dynamic advantages compared to a non-‘spiral’ (linear) pattern typically found in congenital heart defects with transposition of the great arteries [26]. Since the two-chambered hearts of fishes do not show a ‘spiraling’ of their outflow tracts, this functional benefit of cardiac looping cannot be regarded as a phylogenetically old function.
Sherrid and co-workers were focused on the functional significance of the sigmoid routing of the cardiac flow path(s). They were especially interested in indentifying the possible advantages of the ventricular U-turn loop of the human heart, which resulted from the atrial ascensus/ventricular descensus during cardiac looping. They have hypothesized that the cranio-dorsal position of the atriums, which is typical for the multi-chambered hearts of humans and other tetrapods, may provide a favorable hydrostatic position for the systemic venous return as well as pulmonary venous return in land-living vertebrates in which the effects of gravity on the circulating blood are much higher than in water-living fishes [28]. Since the effects of gravity on the circulating blood are largely cancelled out in water [95], the cranio-dorsal position of the atrium found in the two-chambered heart of some fish species (see above) can hardly be interpreted in the same way as in land-living vertebrates. Thus, we may speculate that this feature of the looped design of mature vertebrate hearts may provide more than a single functional advantage. A hydrostatically favorable position then may be regarded as a second, phylogenetically younger functional advantage of the cranio-dorsal position of the atrium that became functional only when the ancestors of land-living vertebrates left their original aquatic habitat. The Asian swamp eel (Monopterus albus) may be regarded as a model for such a scenario. This teleost fish has a two-chambered heart in which the atrium lies in a cranio-dorsal position [53,54]. When this eel-like fish is held, outside of water, in a vertical position with its head upwards, its beating heart is continuously refilled with blood. The beating heart of a true eel (Anguilla anguilla), however, in which the atrium lies dorsal to the ventricle, will rapidly become empty when the fish is held in the same position [53].
Before I will present and discuss the remaining three postulated advantages of a looped heart design, which may be valid for the pumping function of multi-chambered hearts as well as two-chambered hearts of vertebrates, I should point to one difference between the two groups of vertebrate hearts. This difference seems to be of outmost importance for the functional interpretation of a looped heart design but, for unknown reasons, has been completely ignored in previous reflections on the functional significance of cardiac looping. As noted in the section on comparative anatomy, multi-chambered hearts regularly show a bilaterally asymmetric (chiral) anatomy. The functional implications of this feature were discussed in the preceding paragraphs. Two-chambered hearts, on the other hand, do not regularly show a bilaterally asymmetric configuration (Figure 8). Among fishes we can find species with bilaterally asymmetric hearts (e.g. rays, angel sharks, sturgeons) and species with bilaterally symmetric hearts (e.g. tuna, lepisosteiformis, amiiformis). The presence of a nearly perfect bilateral symmetry, especially in the high performance hearts of tuna [96] and other active fishes with a predatory habit, suggests that chiral routing of the cardiac flow path may not significantly contribute to an improvement of the pumping function of two-chambered hearts. Therefore, if we think that the pumping function of two-chambered hearts may profit from a looped design, such a benefit should be attributed to the sigmoid routing of the cardiac flow path, which is a feature found in the two-chambered as well as multi-chambered hearts of vertebrates. The questions then arise: (1), what are the functional implications of chiral looping in fishes; and (2), how can sigmoid routing of the cardiac flow path(s) improve the pumping function of two-chambered as well as multi-chambered hearts?
With regard to the first question, I should refer to the above-mentioned observation that a bilaterally asymmetric heart shape frequently is found in fishes with a dorso-ventrally flattened (‘depressed’) body shape (e.g. rays) while a bilaterally symmetric heart frequently is found in fishes with a laterally ‘compressed’ body shape (e.g. tuna). This finding suggests that a bilaterally asymmetric arrangement of the building units of a two-chambered heart may represent no more than the solution of the problem of packing an s-shaped heart into a body of narrow ventro-dorsal dimensions but wide lateral dimensions. The variability in the positioning of a two-chambered heart within the body of fishes may be compared with the variability in the mounting of an engine into a vehicle, which can be made transverse to the long axis or along the long axis of a car. A striking limitation of the ventro-dorsal dimension of the pericardial cavity not only is found in some fishes but also in early vertebrate embryos where it is regarded as one of the mechanical factors responsible for the chiral/lateral looping of the embryonic heart tube [6]. We therefore may speculate that, during the evolution of vertebrates, the chiral component of the looped design of their hearts originally may have represented no more than the solution of the problem of packing the embryonic as wells as mature s-shaped heart into a physically confined space. The evolution of the genetic control of the left-right patterning of the inner organs of vertebrates then may be primarily interpreted in terms of optimal packing.
With regard to the second question, I can refer to the functional concepts published by Kilner and co-authors [24,25,94]. Based on imaging of the blood flow patterns in the human heart, this group has postulated that, compared to a heart with a linear routing of its flow path, the sigmoid routing of the flow path(s) of vertebrate hearts may confer three interrelated advantages, which are thought to gain significance during strenuous exertion. (1), stabilization of flow. Compared to linear routing, sigmoid routing of the flow path(s) causes a more eccentric (tangential) streaming of the inflowing blood that may allow more stable, less turbulent filling of the heart cavities, and thereby may reduce the dissipation of energy through turbulence. (2), minimizing the loss of momentum of the inflowing blood. Sigmoid routing of the flow path(s) causes eccentric recirculation of the inflowing blood at both atrial and ventricular levels that redirects blood preferentially towards the next chamber or outflow vessel. This may minimize loss of momentum so that the contraction of a chamber only serves to add energy to the blood already moving towards the next chamber (ventricle) or to the arterial stem vessel(s). In a linear heart with a centrally directed flow, the inflow is thought to result in greater instability and turbulence, and would redirect the inflowing blood away from the next chamber or vessel. (3), enhancement of ventriculo-atrial coupling. In a two-chambered heart with a linear arrangement of its building units, such as the heart of the snail Helix pomatia, vigorous ejection of blood during ventricular systole is thought to produce a recoil of the ventricle, like a gun, away from the direction in which the blood is ejected. This is thought to inhibit the reciprocating long-axis displacement of the atrio-ventricular plane (AVP) during the cardiac cycle, which plays an important role in the refilling of the atrial and ventricular chambers of the heart of higher vertebrates [97,98,99,100,101]. Only in the vertebrate heart, with a ventricular U-turn loop, is the postulated systolic recoil of the ventricle thought to be directed so as to enhance rather than inhibit long-axis displacement of the AVP, so supporting atrial filling during the ventricular systole, particularly in the exercising state.
4.2.3. A critical Evaluation of the Proposed Functional Advantages of the Sigmoid Routing of the Cardiac Flow Path(s)
The above-mentioned concepts provide physically sound descriptions of postulated mechanisms by which the sigmoid routing of the flow path(s) of mature vertebrate hearts might optimize the cardiac pumping function. I should note, however, that so far two basic questions still remain unanswered; (1), do these mechanisms really work in vertebrate hearts; and, if so, (2), do these mechanisms really lead to a significant improvement of the pumping function of a looped as opposed to a linear heart?
4.2.3.1. Does Looping Really Minimize the Loss of Momentum of the Inflowing Blood?
Since the first publication of Kilner’s concepts in 2000, only a single study was conducted in which the validity of one of the three postulated advantages was tested by the use of finite element numerical modeling of the hemodynamics of virtual left ventricles [102]. The aim of this study was to compare the hemodynamics of left ventricles with physiologically directed inflow path (towards the apex) and non-physiologically directed inflow path (towards the interventricular septum). It was reported that the physiological flow path did not have an energy-saving effect. This data seem to suggest that sigmoid routing of the cardiac flow path(s) does not improve the efficiency of cardiac pumping by minimizing the loss of momentum of the inflowing blood. It should be noted, however, that the above-mentioned numerical modeling study did not compare the hemodynamics of virtual hearts with a looped and non-looped (linear) arrangement [103]. Thus, answering of the two above-mentioned questions awaits the results of functional studies in which the pumping function of real or virtual hearts with a looped design is compared with the pumping function of non-looped (linear) hearts.
4.2.3.2. Does Looping Really Enhance Ventriculo-Atrial Coupling?
In the human hearts, the reciprocating action of the atrial and ventricular chambers causes a reciprocating displacement of the AVP along the longitudinal axes of the ventricles during the cardiac cycle. During the ventricular systole, the AVP becomes displaced towards the apex while during ventricular diastole the AVP moves back towards the cardiac base (Figure 12A). These movements of the AVP may be compared with the reciprocating movement of a piston unit in a piston pump [104]. It is regarded as the main mechanism for refilling of the atrial and ventricular chambers in the human heart [97,98,99,100,101]. We should, therefore, put a final view on Kilner’s third postulated advantage, which says that a sigmoid routing of the cardiac flow path(s) may produce effects supporting the long-axis displacement of the AVP, while a linear routing of the cardiac flow path(s) may produce effects suppressing the long-axis displacement of the AVP particularly in the exercising state.
Kilner’s concept is based only on reflections about theoretical fluidic and dynamic consequences of a looped versus non-looped (linear) heart design. Although he has noted that a two-chambered heart with a linear design is found in some invertebrates such as the snail Helix pomatia, he has not considered observations on the pumping action of such hearts in his reflections on the postulated disadvantages of a linear heart design. This is astonishing since the action of snail hearts has been studied for more than 100 years. The reported observations consistently show that the pumping action of the two-chambered heart of Helix pomatia and other snails is characterized by an appreciable long-axis displacement of the AVP, which bears striking resemblance to the reciprocating action of a traditional water lifting piston pump (Figure 12B) [105,106,107,108]. Moreover, it has been noted for several times that this movement seems to play a prominent role in the refilling of the atrial and ventricular chamber of the snail heart [106,108]. At first sight, these observations seem to contradict Kilner’s idea that a linear arrangement of the atrial and ventricular heart chambers may hamper the reciprocating long-axis displacement of the AVP during the cardiac cycle. One might argue, however, that Kilner and co-workers had speculated that this postulated disadvantage of a linear heart design might become effective particularly in the exercising state, when an increase in cardiac output leads to a rise in the ventricular recoil resulting from the ejection of blood into the great arteries. Snails are slowly moving animals that do not show a dynamically active behavior. The highest basal heart rates of Helix pomatia, or other land snails reported in the literature, were in the range of 50 to 60 beats per minute [106,109,110]. Most authors, however, reported lower heart rates, which ranged from 20 to 45 beats per minute, even at activity or under stress conditions [107,111,112,113,114]. Activity (locomotion, feeding) or stress (thermal, mechanical) was found to cause a 20 to 80% increase in heart rate, whereby the maximum heart rate did not exceed 40 beats per minute [113,114]. One, therefore, might speculate that the pumping performance of a snail heart normally is so low that, even in an exercising state, recoiling forces resulting from the ejection of blood (hemolymph) into the aorta, cannot fully suppress the long-axis movement of its AVP towards the aorta during ventricular systole. Snail hearts, therefore, may not represent the best biological models for testing the validity of Kilner’s hypothesis. Despite this objection, however, I think that the observations on the pumping action of snail hearts demand a rethinking about the plausibility of Kilner’s theoretical reflections, which use the physics of a firing gun as a model for the postulated behavior of the ventricle of a linearly arranged two-chambered heart during the cardiac cycle. The questions are: (1), is a firing gun really a proper model for a ventricle that ejects blood into the aorta? (2), is it possible that Kilner and co-authors might have neglected one or more functionally important aspects when they made their theoretical reflections on the pumping action of a linearly arranged two-chambered heart? With regard to the first question, I should note that a gun chamber and its outlet (barrel) have rigid walls and therefore do not change their dimensions during shooting. Phenomena typical for the contracting ventricles of vertebrate hearts, such as short-axis shortening and long-axis shortening, do not occur in this kind of launching systems. The design and behavior of a gun, therefore, bears little resemblance to that of the ventricle of a snail or vertebrate heart, which is a chamber that is formed by elastic muscular walls that contract actively and thereby reduce the chamber volume during the ejection of blood. The contracting ventricles of snail and vertebrate hearts rather show strong resemblance to an inflated rubber balloon in which the air is rapidly squeezed out through the opening by the elastic contraction of the balloon. This physical model not only shows short-axis and long-axis shortening. It is, additionally, a simple and well-known teaching device to demonstrate Newton’s third law of motion. I, therefore, think that a balloon rocket is a much better model than a firing gun. With regard to the second question, I should note that Kilner and co-authors have made their theoretical reflections only for the action of hearts that were physically isolated from their normal environment (see Figure 3 in [24]). Such reflections neglect the fact that vertebrate as well as snail hearts normally are fixed at their venous and arterial poles to the wall of a pericardial cavity. The wall of the pericardial cavity, the blood vessels entering and exiting the heart, and the anatomical structures surrounding the pericardial cavity, all together normally act as a frame for fixation of the heart. This frame can absorb the recoil momentum resulting from the systolic ejection of blood, so that the heart and its surrounding structures (pericardium, etc.) together may act as a non-recoil system. The behavior of such a non-recoil system can be demonstrated with the above-mentioned rubber balloon model. If the rubber balloon model is used without physical fixation, the inflated balloon contracts while he flies away into the direction opposite to the opening through which the air streams out off the balloon. If the rubber balloon model is used together with a frame to which the opening of the balloon is physically fixed, the elastic contraction of the balloon will squeeze out the air through the opening but the balloon will remain at its place, where it undergoes short-axis and long-axis shortening.
The pumping action of snail hearts as well as the behavior of the above-described rubber balloon model strongly suggests that, even at strenuous exertion, a linear arrangement of the atrial and ventricular heart chambers may not hamper the long-axis displacement of the AVP. Thus, it appears that, with regard to this functional feature, sigmoid routing of the cardiac flow path(s) may not provide a significant improvement compared to a linear heart design. Moreover, a geometrical analysis shows that, in hearts with an elongated ventricle, an appreciable displacement of the AVP during the cardiac cycle is expected to occur only in those hearts in which the AVP lies perpendicular or almost perpendicular to the ventricular long-axis (Figure 12A, B).
In vertebrates, such a situation is found either in hearts with a low degree of sigmoid routing of the flow path(s) or in hearts with a high degree of sigmoid routing of the flow path(s) (hearts with ventricular V-turn or U-turn loop). The AVP of hearts with a moderate degree of sigmoid routing of the flow path lies perpendicular to the ventricular short axis (hearts with ventricular L-turn loop). In this situation, the extent of AVP displacement does not depend on the extent of long-axis shortening but depends on the degree to which the epicardial short-axis diameter changes during the cardiac cycle (Figure 12C). Compared to the length of the long axis of the ventricle(s), the epicardial short-axis diameter shows only small changes during the cardiac cycle [116]. Thus, AVP displacement will not significantly contribute to the pumping function of hearts with a ventricular L-turn loop, which drive the circulation of blood in many fish species such as the zebrafish.
Compared to hearts with a linear design or hearts with a high degree of sigmoid routing of the flow path(s), hearts with a ventricular L-turn loop show a further morphological feature that holds a potential disadvantage for their pumping function. The atrio-ventricular canal is arranged perpendicular to the outflow path of the ventricle so that, in cases of hearts with long atrio-ventricular valve leaflets, parts of the atrio-ventricular valve apparatus cross the ventricular outflow path (Figure 7B). Such a situation, for example, is found in the fishes Channa argus and Anabas testudines (see Figure 1 in [117], and Figs. 3 and 4 in [118]). This feature may represent a hemodynamically disadvantageous feature that, not only, may disturb the systolic blood stream and thereby may reduce the pumping efficiency of the heart. It may, additionally, predispose to the development of severe outflow tract obstructions in consequence of pathologies of the atrio-ventricular valve. Such a situation seems to resemble the situation found in human heart pathologies with obstruction of the left ventricular outflow tract, in which a reduced angle between the mitral and aortic valves seems to play a prominent pathogenetic role [119]. Thus, it appears that a moderate degree of sigmoid routing of the cardiac flow path does not seem to provide an advantage but rather seems to provide potential disadvantages for the pumping function of mature vertebrate hearts. In view of this situation, it seems that sigmoid routing of the cardiac flow path(s) may not generally provide advantages for the pumping function of mature vertebrate hearts. It is a challenge for the future to clarify the situation by simulation studies facilitating the comparison of the pumping performance between hearts with a linear design and those with various degrees of sigmoid routing of the cardiac flow path.
4.2.3.3. Implications for the Usage of Fishes in Cardiovascular Research
The above-described functional differences between fish hearts with a moderate degree of sigmoid routing (ventricular L-turn loop) and those with a high degree (ventricular u-turn loop) may have practical consequences for biomedical research since they suggest that fish hearts with a moderate degree of sigmoid routing of the flow path, such as the zebra- fish heart, may not represent appropriate models for clarifying the genetic and molecular basis of the pumping function of the left ventricle of the human heart. It might be more suitable to use fish heart models with a high degree of sigmoid routing of the flow path, such as the heart of swamp eels (e.g. Monopterus albus) or the heart of gars (e.g. Lepisosteus osseus). The hearts of the latter two fishes not only show a U-turn loop of their ventricular flow path but, additionally, are characterized by the presence of fibrous continuity between the atrio-ventricular and the arterial valve [51,53].
5. Summary and Conclusions
In this article, I have presented and discussed the currently available information about the looped design of vertebrate hearts and its functional implications. Thereby, emphasis is given to the following observations:
- (1), the looped design of vertebrate hearts is roughly characterized by (a), an s-shaped (sigmoid) routing of the cardiac flow paths, and (b,) a bilaterally asymmetric (chiral) routing of the cardiac flow paths.
- (2), the fully looped heart tube of vertebrate embryos regularly shows sigmoid as well as chiral routing of its flow path.
- (3), data from physical pump models suggest that the looped design of valveless embryonic heart tubes might improve the pumping efficiency.
- (4), among the mature (chambered) hearts of vertebrates, sigmoid as well as chiral routing of the flow path(s) is regularly found only in the multi-chambered hearts of lungfish and tetrapods. Here, the bilaterally asymmetric (chiral) anatomy is regarded as the main determinant of the alignment and functional or structural separation of the systemic and pulmonary flow paths.
- (5), among the two-chambered heart of fishes, only sigmoid routing of the flow path is a regular feature, while a bilaterally asymmetric (chiral) configuration is not a regular feature. Thus, among fishes we can find species with bilaterally asymmetric hearts as well as species with bilaterally symmetric hearts.
- (6), the presence of bilateral symmetry in the mature hearts of some fishes seems to be the consequence of a process of back-rotation of the heart during the post-embryonic (larval) period of development.
- (7), the presence of a nearly perfect bilateral symmetry in the high performance hearts of tuna, and other active fishes, suggests that chiral routing of the cardiac flow path may not significantly improve the pumping function of two-chambered hearts. Thus, if we assume that the pumping function of two-chambered hearts may profit from a looped design, such a benefit should be attributed to the sigmoid routing of the cardiac flow path, which is a feature found in the two-chambered as well as multi-chambered hearts of vertebrates.
- (8), the bilaterally asymmetric (chiral) anatomy of some fish hearts seems to represent no more than the solution of a packing problem.
- (9), it was frequently stated that the evolution of the vertebrate heart was characterized by an increase in the degree of sigmoid routing of the cardiac flow path(s). The lowest degree of sigmoid routing was ascribed to the hearts of basal vertebrates (jawless fishes). A moderate degree was ascribed to the hearts of jawed fishes, while high degrees of sigmoid routing of the cardiac flow path(s) were ascribed to the hearts of lungfish and tetrapods. A review of original data from fishes, however, shows that this phylogenetic scenario does not match with the reality. Among jawed fishes, we can find a spectrum of various degrees of sigmoid routing of the cardiac flow path that ranges from low degrees, resulting from an atrial position that is dorso-caudal to the ventricle, to high degrees, resulting from an atrial position that is dorso-cranial to the ventricle. Thus, a high degree of sigmoid routing of the cardiac flow path(s) is not exclusively found in the mature hearts of tetrapods.
- (10), Kilner an co-authors have postulated that sigmoid routing of the cardiac flow path(s) may produce effects supporting the long-axis displacement of the AVP, while a linear routing of the cardiac flow path(s) may produce effects suppressing the long-axis displacement of the AVP particularly in the exercising state. Data from snail hearts and physical models cast doubt on the validity of this hypothesis. The relation between the looped design and the pumping function of vertebrate hearts remains an enigma.
Funding
This research received no external funding.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Von Baer, K.E. Untersuchungen über die Entwickelungsgeschichte der Fische nebst einem Anhange über die Schwimmblase. F.C.W. Vogel: Leipzig, Germany, 1835.
- Bischoff, T.L.W. Entwickelungsgeschichte des Säugethier- und Menscheneies. L. Voß: Leipzig, Germany, 1842.
- His, W. Anatomie menschlicher Embryonen III.. F.C.W. Vogel: Leipzig, Germany, 1885.
- Sobotta, J. Über die Entwickelung des Blutes, des Herzens und der grossen Gefässstämme der Salmoniden. Anat Hefte 1902, 19, 579–688. [Google Scholar] [CrossRef]
- Bremer, J.L. Part I. An interpretation of the development of the heart. Part II. The left aorta of reptiles. Am J Anat 1928, 42, 307–369. [Google Scholar] [CrossRef]
- Patten, B.M. The formation of the cardiac loop in the chick. Am J Anat 1922, 30, 373–397. [Google Scholar] [CrossRef]
- Senior, H.D. The development of the heart in shad (Alosa Sapadissima, Wilson). Am J Anat 1909, 9, 211–262. [Google Scholar] [CrossRef]
- Tschermak, A. 1909. Physiologische Untersuchungen am embryonalen Fischherzen. Sitzgsber Akad Wiss Wien, Math-naturwiss Kl 3 1909, 118, 17. [Google Scholar]
- Robertson, J.I. The development of the heart and vascular system of Lepidosiren paradoxa. J Cell Sci 1913, 59, 53–132. [Google Scholar] [CrossRef]
- Tschermak, A. Über das Verhalten des embryonalen Fischherzens gegenüber dem konstanten Strom. Z Exper Med 1929, 68, 452–474. [Google Scholar] [CrossRef]
- Icardo, J.M.; Guerrero, A.; Durán, AC.; Domezain, A.; Colvee, E.; Sana-Coma, V. The development of the sturgeon heart. Anat Embryol 2004, 208, 439–449. [Google Scholar] [CrossRef]
- Rodríguez, C,; Sans-Coma, V. ; Grimes, A.C.; Fernández, B.; Arqué, J.M.; Durán, A.C. Embryonic development of the bulbus arteriosus of the primitive heart of jawed vertebrates. Zool Anz 2013, 252, 359–366. [Google Scholar] [CrossRef]
- Cooke, J. Developmental mechanism and evolutionary origin of vertebrate left/right asymmetries. Biol Rev 2004, 79, 377–407. [Google Scholar] [CrossRef]
- Palmer, A.R. Symmetry breaking and the evolution of development. Science 2004, 306, 828–833. [Google Scholar] [CrossRef] [PubMed]
- Ramsdell, A.F. Left-right asymmetry and congenital cardiac defects: getting to the heart of the matter in vertebrate left-right axis determination. Dev Biol 2005, 288, 1–20. [Google Scholar] [CrossRef]
- Stainier, D.Y.R. , Fishman, M.C. The zebrafish as a model system to study cardiovascular development. Trends Cardiovasc Med 1994, 4, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.A. , Uribe, V. Getting to the heart of left-right asymmetry: contributions from the zebrafish model. J Cardiovasc Dev Dis 2021, 8, 64. [Google Scholar] [CrossRef]
- Lombardo, V.A.; Heise, M.; Moghtadaei, M.; Bornhorst, D.; Männer, J.; Abdelilah-Seyfried, S. Morphogenetic control of zebrafish cardia looping by Bmp signaling. Development 2019, 146, dev180091. [Google Scholar] [CrossRef]
- Tessadori, F.; Tsingos, E.; Colizzi, E.S.; Kruse, F.; van den Brink, S.C.; van den Boogaard, M.; Christoffels, V.M.; Merks, R.M.H.; Bakkers, J. Twisting of the zebrafish heart tube during cardiac looping is a tbx5-dependent and tissue-intrinsic process. Elife 2021, 10, e61733. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, G.C.; Lo, C.W. Left-right patterning in congenital heart disease and beyond. Am J Genet C Semin Med Genet 2020, 184, 90–96. [Google Scholar] [CrossRef]
- Nakano, H.; Fajardo, V.M.; Nakano, A. The role of glucose in physiological and pathological heart formation. Dev Biol 2021, 475, 222–233. [Google Scholar] [CrossRef]
- Rahman, T.; Zhang, H.; Fan, J.; Wan, Q. Cell chirality in cardiovascular development and disease. APL Bioeng 2020, 4, 031503. [Google Scholar] [CrossRef]
- Desgrange, A.; Le Garrec, J.F.; Meilac, S.M. Left-right asymmetry in heart development and disease: forming the right loop. Development 2018, 145, dev162776. [Google Scholar] [CrossRef]
- Kilner PJ, Yang GZ, Wilkes AJ, Mohladdin RH, Firmin DN, Yacoub MH. Asymmetric redirection of flow through the heart. Nature 2000, 404, 759–761. [Google Scholar] [CrossRef]
- Kilner, P.J.; Yang, G.Z.; Firmin, D.N. Morphodynamics of flow through sinuous curvatures of the heart. Biorheology 2002, 39, 409–417. [Google Scholar] [PubMed]
- Amodeo, A.; Olivero, M.; Versacci, P.; Marino, B. Spiral shapes in heart and shells: when form and function do matter. Eur J Cardio-Thorac Surg 2012, 41, 473–475. [Google Scholar] [CrossRef] [PubMed]
- Hiermeier, F.; Männer, J. Kinking and torsion can significantly improve the efficiency of valveless pumping in periodically compressed tubular conduits. Implications for understanding of the form-function relationship of embryonic heart tubes. J Cardiovasc Dev Dis 2017, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Sherrid, M.V.; Männer, J.; Swistel, D.G.; Olivotto, I.; Halpern, D.G. On the cardiac loop and its failing: left ventricular outflow tract obstruction. J Am Heart Ass 2020, 9, e014857. [Google Scholar] [CrossRef] [PubMed]
- Männer, J. When does the human embryonic heart start beating? A review of contemporary and historical sources of knowledge about the onset of blood circulation in man. J Cardiovasc Dev Dis 2022, 9, 187. [Google Scholar] [CrossRef] [PubMed]
- Simões-Costa, M.S.; Vasconcelos, M.; Sampaio, A.C.; Cravo, R.M.; Linhares, V.L.; Hochgreb, T.; Yan, C.Y.I.; Davidson, B.; Xavier-Neto, J. The evolutionary origin of cardiac chambers. Dev Biol 2005, 277, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Männer, J.; Wessel, A.; Yelbuz, T.M. How does the tubular embryonic heart work? Looking for the physical mechanism driving unidirectional blood flow in the valveless embryonic heart tube. Dev Dyn 2010, 239, 1035–1046. [Google Scholar] [CrossRef]
- Abu Issa, R.; Kirby, M.L. Heart field: from mesoderm to heart tube. Ann Rev Cell Dev Biol 2007, 23, 45–68. [Google Scholar] [CrossRef]
- Meilhac, S.M.; Lescroart, F.; Blanpain, C.; Buckingham, M.E. Cardiac cell lineages that form the heart. Cold Spring Harb Perspect Med 2014, 4, a013888. [Google Scholar] [CrossRef]
- Burggren, W.W. Cardiac design in lower vertebrates: what can phylogeny reveal about ontogeny? Experentia 1988, 44, 919–930. [Google Scholar] [CrossRef] [PubMed]
- Bettex, D.A.; Prêtre, R.; Chassot, P.G. Is our heart a well- designed pump? The heart along evolution. Eur Heart J 2014, 35, 2322–2332. [Google Scholar] [CrossRef] [PubMed]
- Ziermann, J.M.; Freitas, R.; Diogo, R. Muscle development in the shark Scyliorhinus canicula: implications for the evolution of the gnathostome head and paired appendage musculature. Front Zool 2017, 14, 31. [Google Scholar] [CrossRef] [PubMed]
- Männer, J. On the form problem of embryonic heart loops, its geometrical solutions, and a new biophysical concept of cardiac looping. Ann Anat 2013, 195, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Kunz, Y. Morphologische Studien über die embryonale und postembryonale Entwicklung bei Teleostiern mit besonderer Berücksichtigung des Dottersystems und der Leber. Revue Suisse de Zoologie 1964, 71, 445–525. [Google Scholar] [CrossRef]
- Kunz-Ramsay, Y. Developmental biology of Teleost fishes. Springer: New York, U.S.A., 2013.
- Singelman, C.; Holtzman, N.G. Analysis of postembryonic heart development and maturation in the zebrafish, Danio rerio. Dev Dyn 2012, 241, 1993–2004. [Google Scholar] [CrossRef] [PubMed]
- Pernkopf, E.; Wirtinger, W. Die Transposition der Herzostien – ein Versuch der Erklärung dieser Erscheinung. I. Teil. Die Pheronomie der Herzentwicklung. Z Anat Entwickl Gesch 1933, 100, 563–711. [Google Scholar] [CrossRef]
- Männer, J. The anatomy of cardiac looping: a step towards the understanding of the morphogenesis of several forms of congenital cardiac malformations. Clin Anat 2009, 22, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Icardo, J.M.; Colvee, E.; Schorna, S.; Lauriano, E.R.; Fudge, D.S.; Glover, C.N.; Zaccone, G. Morphological analysis of the hagfish heart I: The ventricle, the arterial connection and the ventral aorta. J Morphol 2015, 277, 326–340. [Google Scholar] [CrossRef]
- Icardo, J.M. Heart morphology and anatomy. In The cardiovascular system: morphology, control and function. Fish Physiology Vol. 36 Part A., Gamperl, A.K.; Gillis, T.E., Farrell, A.P., Eds.; Brauner, C.J. Eds.: Academic Press, 2017; pp. 1–54. [Google Scholar]
- Muramatsu, B.; Suzuki, D.G.; Suzuki, M.; Higashiyama, H. Gross anatomy of the Pacific hagfish, Eptatretus burgeri, with special reference to the coelomic viscera. Anat Rec 2024, 307, 155–171. [Google Scholar] [CrossRef]
- Santer, R.M. 1985. Morphology and innervation of the fish heart. Adv Anat Embryol Cell Biol 1954, 89. [Google Scholar]
- Richardson, M.K.; Admiraal, J.; Wright, G.M. Developmental anatomy of lampreys. Biol Rev 2010, 85, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Anthony, J.; Millot, J.; Robineau, D. Le cœur et l’aorte ventrale de Latimeria chalumnae (Poisson coelacanthidé). CR Acad Sci 1965, 261, 223–226. [Google Scholar]
- Millot, J.; Anthony, J.; Robineau, D. Anatomie de Latimeria chalumnae. Apparell digestif – apparel respiratoire – apparel urogenital – glandes endocrines – apparel circulatoire – teguments – écailles – conclusion générales. CNRS 1978, 3, 198. [Google Scholar]
- Gregory, J.A.; Graham, J.B.; Cech, J.J. Jr.; Dalton, N. Michaels, J. Lai, N.C. Pericardial and peritoneal canal relationships to cardiac function in the white sturgeon (Acipenser transmontanus). Comp Biochem Physiol A 2004, 138, 203–213. [Google Scholar] [CrossRef]
- Boas, J.E.V. Über Herz und Arterienbogen bei Ceratodus und Protopterus. Morphol JB 1880, 6, 321–354. [Google Scholar]
- Home, E. XXVII. Additions to an account of the anatomy of Squalus maximus, contained in a former paper; with observations on the structure of the branchial artery. Phil Trans Royal Soc 1813, 103, 227–241. [Google Scholar]
- Liem, K.F. Tetrapod parallelisms and other features in the functional morphology of the blood vascular system of Fluta alba Zuiew (pisces: teleostei). J Morphol 1961, 108, 131–143. [Google Scholar] [CrossRef]
- Lai, N.C.; Graham, J.B.; Dalton, N.; Shabetai, R.; Bhargava, V. Echocardiographic and hemodynamic determinations of the ventricular filling pattern in some teleost fishes. Physiol Zool 1998, 71, 157–167. [Google Scholar] [CrossRef]
- Ludwig, W. Das Rechts-Links-Problem im Tierreich und beim Menschen. Springer: Berlin, Germany, 1932.
- Mercola, M. Embryological basis for cardiac left-right asymmetry. Sem Cell Dev Biol 1999, 10, 109–116. [Google Scholar] [CrossRef]
- Kathiriya IS, Srivastava D. Left-right asymmetry and cardiac looping: implications for cardiac development and congenital heart disease. Am J Med Genet (Semin Med Genet) 2000, 97, 271–279. [Google Scholar] [CrossRef]
- Fishman, M.C.; Chien, K.R. Fashioning the vertebrate heart: earliest embryonic decisions. Development 1999, 124, 2099–2117. [Google Scholar] [CrossRef] [PubMed]
- Blum, M.; Ott, T. Animal left-right asymmetry. Current Biol 2018, 28, R293–R305. [Google Scholar] [CrossRef] [PubMed]
- Soares, K.D.A.; Toledo-Piza, M. Branching patterns of the afferent branchial arteries and their phylogenetic significance in rays (Batoidea). Sci Rep 2021, 11, 23236. [Google Scholar] [CrossRef] [PubMed]
- Van Mierop, L.H.S.; Kutsche, L.M. Comparative anatomy and embryology of the ventricles and arterial pole of the vertebrate heart. In: Nora, J.J.; Takao, A. Eds. Congenital heart disease. Causes and Processes, Futura, New York, U.S.A. 1984; pp.459-479.
- Zaccone, G.; Grimes, A.C.; Farrell, A.P.; Dabrowski, K.; Marino, F. Morphology, innervation and its phylogenetic step in the heart of the longnose gar Lepisosteus Osseus. Acta Zool 2012, 93, 381–389. [Google Scholar] [CrossRef]
- Tota, B. 1999. Heart. Chapter 10 In: Hamlett, W.C. Ed., Sharks, skated, and rays. The biology of elasmobranch fish. Johns Hopkins University Press, Baltimore, U.S.A. 1999; pp. 238-272.
- Icardo, J.M.; Colvee, E.; Cerra, M.C.; Tota, B. Structure of the conus arteriosus of the sturgeon (Acipenser naccarii) heart I: The conus valves and subendocardium. Anat Rec 2002, 267, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Szidon, J.; Lahiri, S.; Lev, M.; Fishman, A.P. Heart and circulation of the African lungfish. Circulation Res 1969, 25, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Hatta, A.P. Über die Entwicklung des Gefäßsystems des Neunauges, Lampetra mitsukurii Hatta. Zool JB (Anat.) 1923, 44, 1–264. [Google Scholar]
- Piavis, G.W. Embryology. In Biology of lampreys. Hardisty, M.W.; Potter, I.C., Ed.; Academic Press: London, 1971; pp. 361–400. [Google Scholar]
- Icardo, J.M. Conus arteriosus of the teleost heart: dismissed, but not missed. Anat Rec 2006, 288A, 900–908. [Google Scholar] [CrossRef]
- Grimes, A.C.; Kirby, M.L. The outflow tract of the heart of fishes: anatomy, genes, and evolution. J Fish Biol 2009, 74, 983–1036. [Google Scholar] [CrossRef]
- Lorenzale, M.; Lopez-Unzu, M.A.; Rodríguez, C.; Fernández, B.; Durán, A.C.; Sans-Coma, V. The anatomical components of the cardiac outflow tract of chondrichthyans and actinopterygians. Biol Rev 2018, 93, 1604–1619. [Google Scholar] [CrossRef]
- Drechsel, J. Myokardauffaserungsversuche am Fisch- und Vogelherzen. Z Anat Entwickl Gesch 1935, 104, 403–423. [Google Scholar] [CrossRef]
- Victor, S.; Nayak, V.M.; Rajasingh, R. Evolution of the ventricles. Tex Heart Inst 1999, 26, 168–175. [Google Scholar]
- Schäfer, K.J. Untersuchungen zur Frage der bilateral asymmetrischen Morphologie des adulten Zebrafischherzens. Doctoral thesis, UMG, Georg-August-University, Goettingen, May 31 2023.
- Sánchez-Quintana, D.; García-Martínez, V.; Climent, V.; Hurlé, J.M. Myocardial fiber and connective tissue architecture in the fish heart ventricle. J Exp Zool 1996, 275, 112–124. [Google Scholar] [CrossRef]
- Ballard, W.W. Stages and rates of normal development in the holostean fish, Amia calva. J Exp Zool 1986, 238, 337–354. [Google Scholar] [CrossRef]
- Clowes, C; Boylan, M.G.S.; Ridge, L.A.; Barnes, E.; Wright, J.A.; Hentges, K.E. The functional diversity of essential genes required for mammalian cardiac development. Genesis 2014, 52, 713–737. [CrossRef]
- Sabin, F.R. Origin and development of the primitive vessels of the chick and of the pig. Contr Embryol Carnegie Inst 1917, 6, 61–124. [Google Scholar]
- Jones, E.A.V. The initiation of blood flow and flow induced events in the early vascular development. Semin Cell Dev Biol 2011, 22, 1028–1035. [Google Scholar] [CrossRef]
- Kalogirou, S.; Malissovas, N.; Moro, E.; Argenton, F.; Stainier, D.Y.R.; Beis, D. Intracardiac flow dynamics regulate atrioventricular valve morphogenesis. Cardiovasc Res 2014, 104, 49–60. [Google Scholar] [CrossRef]
- Berndt, C.; Poschmann, G.; Stühler, K.; Holmgren, A.; Bräutigam, L. Zebrafish heart development is regulated via glutaredoxin 2 dependent migration and survival of neural crest cells. Redox Biol 2014, 2, 673–678. [Google Scholar] [CrossRef]
- Forouhar, A.S.; Liebling, M.; Hickerson, A.; Nasiraei-Moghaddam, A.; Tsai, H.J.; Hove, J.R.; Fraser, S.E.; Dickinson, M.E.; Gharib, M. The embryonic vertebrate heart tube is a dynamic suction pump. Science 2006, 312, 751–753. [Google Scholar] [CrossRef]
- Bisphoric, N.H. Evolution of the heart from bacteria to man. Ann NY Acad Sci 2005, 1047, 13–29. [Google Scholar] [CrossRef]
- Johansen, K. Air-breathing fishes. Scientific American 1968, 219, 102–111. [Google Scholar] [CrossRef]
- Grant, R.P. The morphogenesis of corrected transposition and other anomalies of cardiac polarity. Circulation 1964, 29, 71–83. [Google Scholar] [CrossRef]
- Meredith, M.A.; Hutchins, G.M.; Moore, G.W. Role of the left interventricular sulcus in formation of the interventricular septum and crista supraventricularis in normal human cardiogenesis. Anat Rec 1979, 194, 417–428. [Google Scholar] [CrossRef]
- Männer, J. Cardiac looping in the chick embryo: a morphological review with special reference to terminological and biomechanical aspects of the looping process. Anat Rec 2000, 259, 248–262. [Google Scholar] [CrossRef]
- Moorman, A.F.M.; Christoffels, V.M. Cardiac chamber formation: development, genes, and evolution. Physiol Rev 2003, 83, 1223–1267. [Google Scholar] [CrossRef]
- Lewis, F.T.; Abbott, M.E. Reversed torsion of the human heart. Anat Rec 1915, 9, 103–105. [Google Scholar] [CrossRef]
- Lewis, F.T.; Abbott, M.E. Reversed torsion of the ventricular bend of the embryonic heart in the explanation of certain forms of cardiac anomaly. Bull Int Assoc Med Mus 1916, 6, 111–115. [Google Scholar]
- De la Cruz, M.V.; Anselmi, G.; Cisneros, F.; Reinhold, M.; Portillo, B.; Espino-Vela, J. An embryologic explanation for the corrected transposition of the great vessels: additional description of the main anatomic features of this malformation and its varieties. Am Heart J 1959, 57, 104–117. [Google Scholar] [CrossRef]
- Van Praagh, R.; Van Praagh, S.; Vlad, P.; Keith, J.D. Anatomic types of congenital dextrocardia: diagnostic and embryologic implications. Am J Cardiol 1964, 13, 510–531. [Google Scholar] [CrossRef]
- Anderson, R.H.; Webb, S.; Brown, N.A. Defective lateralisation in children with congenitally malformed hearts. Cardiol Young 1998, 8, 512–531. [Google Scholar] [CrossRef]
- Stainier, D.Y.R.; Lee, R.K.; Fishman, M.C. Cardiovascular development in the zebrafish. I. Myocardial fate map and heart tube formation. Development 1993, 119, 31–40. [Google Scholar] [CrossRef]
- Yang GZ, Merrifield R, Masood S, Kilner PJ. Flow and myocardial interaction: an imaging perspective. Phil Trans R Soc B 2007, 362, 1329–1341. [Google Scholar] [CrossRef]
- Sandblom, E.; Axelsson, M. The venous circulation: a piscine perspective. Comp Biochem Physiol A 2007, 148, 785–801. [Google Scholar] [CrossRef]
- Farrell, A.P. From hagfish to tuna: a perspective on cardiac function in fish. Physiol Zool 1991, 64, 1137–1164. [Google Scholar] [CrossRef]
- Henein, M.Y.; Gibson, D.G. 1999. Normal long axis function. Heart 1999, 81, 111–113. [Google Scholar] [CrossRef]
- Carlsson, M.; Ugander, M.; Mosen, H.; Buhre, T.; Arheden, H. Atrioventricular plane displacement is the major contributor to left ventricular pumping in healthy, athletes, and patients with dilated cardiomyopathy. AJP Heart Circ Physiol 2007, 292, H1452–H1459. [Google Scholar] [CrossRef]
- Carlsson, M.; Ugander, M.; Heiberg, E.; Arheden, H. The quantitative relationship between longitudinal and radial function in left, right, and total heart pumping in humans. AJP Heart Circ Physiol 2007, 293, H636–H644. [Google Scholar] [CrossRef]
- Steding-Ehrenborg, K.; Carlsson, M.; Stephensen, S.; Arheden, H. Atrial aspiration from pulmonary and caval veins is caused by ventricular contraction and secures 70% of the total stroke volume independent of resting heart rate and heart size. Clin Physiol Funct Imaging 2013, 33, 233–240. [Google Scholar] [CrossRef]
- Arutunyan, A.H. Atrioventricular plane displacement is the sole mechanism of atrial and ventricular refill. AJP Heart Circ Physiol 2015, 308, H1317–H1320. [Google Scholar] [CrossRef]
- Watanabe, H.; Sugiura, S.; Hisada, T. The looped heart does not save energy by maintaining the momentum of blood flowing in the ventricle. AJP Heart Circ Physiol 2008, 294, H2191–H2196. [Google Scholar] [CrossRef]
- Kilner, PJ. 2010. Letter to the editor: “Postulated functional advantages of a looped as opposed to a linearly arranged heart”. AJP Heart Circ Physiol 298:H726. [CrossRef]
- Maksuti, E.; Bjällmark, A.; Broomé, M. Modelling the heart with the atrioventricular plane as a piston unit. Medical Engineering Physics 2015, 37, 87–92. [Google Scholar] [CrossRef]
- Jordan, H.J.; Hirsch, G.C. Übungen aus der vergleichenden Physiologie. Springer: Berlin, Germany; 1927.
- Biering, P. Untersuchungen über das Kreislaufsystem bei den Weichtieren II. Z Vergl Physiol 1929, 10, 465–484. [Google Scholar] [CrossRef]
- Schwartzkopff, J. Über die Leistung des isolierten Herzens der Weinbergschnecke (Helix pomatia L.) im künstlichen Kreislauf. Z Vergl Physiol 1954, 36, 543–594. [Google Scholar] [CrossRef]
- Jones, H.D. Circulatory pressures in Helix pomatia. Comp Biochem Physiol A 1971, 39, 289–295. [Google Scholar] [CrossRef]
- von Skramlik, E. Weichtieren. In Ergebnisse der Biologie. Frisch, K.; Ruhland, W., Koehler, O., Stubbe, H., Eds.; Springer: Berlin, Germany, 1941. [Google Scholar]
- Van Aardt, W.J.; Vosloo, A. Modifications to an optocardiographic method for measuring of heart rate in a range of invertebrate species. S Afr J Zool 1996, 32, 97–100. [Google Scholar]
- Nopp, H. Temperaturbezogene Regulationen des Sauerstoffverbrauchs und der Herzschlagrate bei einigen Pulmonaten. Z Vergl Physiol 1965, 50, 641–659. [Google Scholar] [CrossRef]
- Wünnenberg, W. Diurnal rhythms of the heart rate in the snail Helix pomatia L. Comp Biochem Physiol A 1991, 99, 415–417. [Google Scholar] [CrossRef]
- Romero, S.M.; Hoffmann, A. Heart rate and behavioral patterns of Megalobulimus sanctipauli (mollusca, gastropoda, pulmonata). Braz J Med Biol Res 1991, 24, 223–227. [Google Scholar]
- Renwrantz, L.; Spielvogel, F. Heart rate and hemocyte number as stress indicators in disturbed hibernating vineyard snails, Helix pomatia. Comp Biochem Physiol Part A. [CrossRef]
- Noll, A. Untersuchungen über das Kreislaufsystem bei den Weichtieren III. Z Vergl Physiol 1929, 10, 761–777. [Google Scholar] [CrossRef]
- Ugander, M.; Carlsson, M.; Arheden, H. Short axis epicardial volume change is a measure of cardiac left ventricular short axis function, which is independent of myocardial wall thickess. AJP Heart Circ Physiol 2010, 298, H530–H535. [Google Scholar] [CrossRef] [PubMed]
- Ishimatsu, A.; Itazawa, Y. Differences in blood oxygen levels in the outflow vessels of the heart of an air-breathing fish, Channa argus: do separate blood streams exist in a teleostean heart? J Comp Physiol 1983, 149, 435–440. [Google Scholar] [CrossRef]
- Munshi, J.S.D.; Olson, K.R.; Roy, P.K.; Ghosh, U. Scanning electron microscopy of the heart of the climbing perch. J Fish Biol 2001, 59, 1170–1180. [Google Scholar] [CrossRef]
- Park, M.H.; Imbrie-Moore, A.M.; Zhu, Y.; Wilkerson, R.J.; Wang, H.; Park, G.H.; Wu, C.A.; Pandya, P.K.; Mullis, D.M.; Marin-Cuartas, M.; Woo, Y.J. The critical biomechanics of aortomitral angle and systolic anterior motion: engineering native ex vivo simulation. Ann Biomed Engin 2023, 51, 794–805. [Google Scholar] [CrossRef]
Figure 1.
These scanning electron microscopic images illustrate species-specific differences in the configuration of the ‘S-shaped’ heart loop of vertebrate embryos. Heart loops are shown in frontal views. (A), ‘S-shaped’ heart loop of a zebrafish embryo at 48 hours after fertilization. (B), ‘S-shaped’ heart loop of a mouse embryo at 9.5 days after fertilization. Abbreviations: A = common atrium; AV = atrio-ventricular canal; O = outflow tract; V = ventricular bend.
Figure 1.
These scanning electron microscopic images illustrate species-specific differences in the configuration of the ‘S-shaped’ heart loop of vertebrate embryos. Heart loops are shown in frontal views. (A), ‘S-shaped’ heart loop of a zebrafish embryo at 48 hours after fertilization. (B), ‘S-shaped’ heart loop of a mouse embryo at 9.5 days after fertilization. Abbreviations: A = common atrium; AV = atrio-ventricular canal; O = outflow tract; V = ventricular bend.
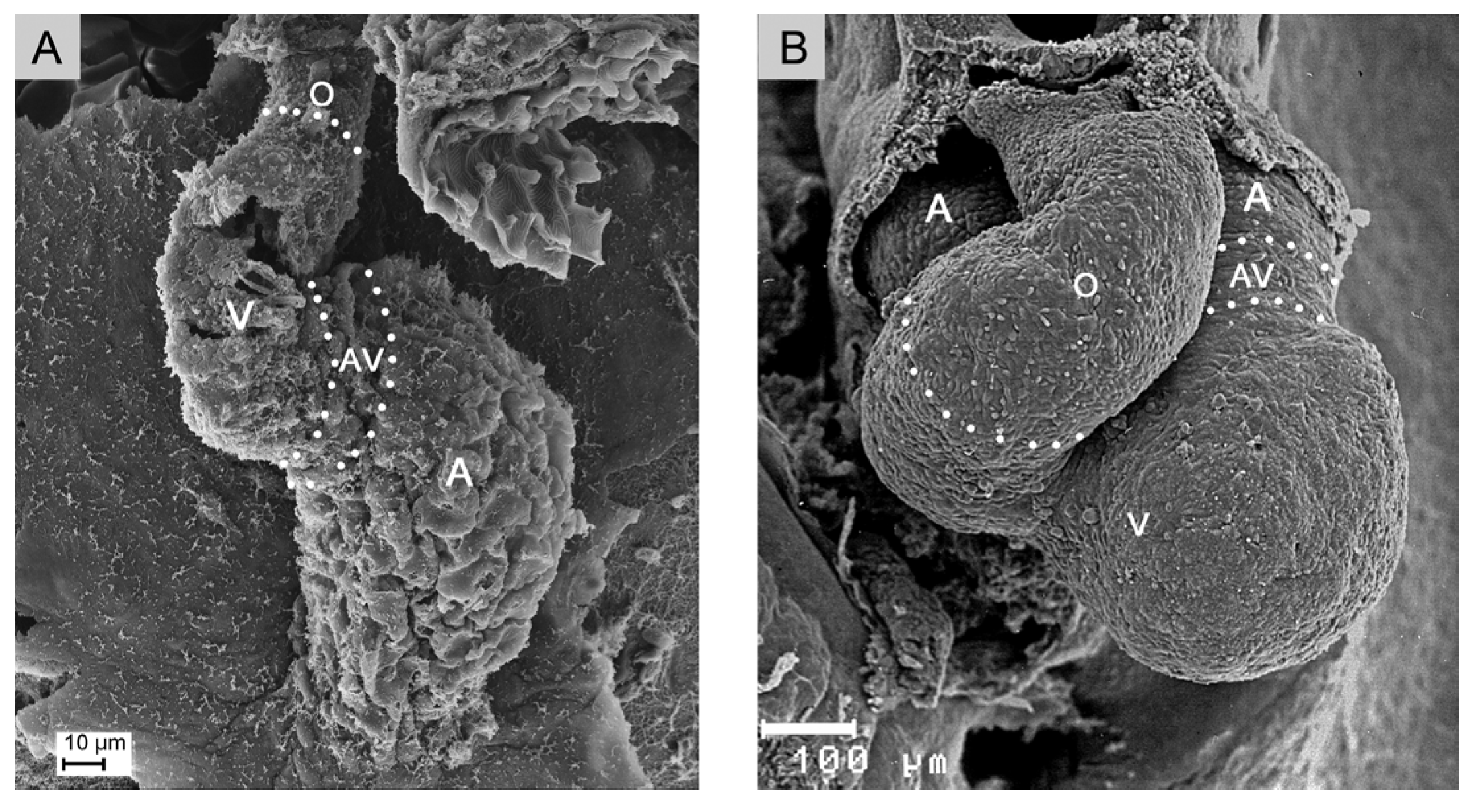
Figure 2.
This schematic drawing illustrates the positional and morphological changes of the looping embryonic heart tube along the original ventro-dorsal heart axis. Color code: blue = confluence of veins; brown = atrial segment; yellow = AV-canal; pink = ventricular segment; red = ventricular outflow element. Abbreviations as used before.
Figure 2.
This schematic drawing illustrates the positional and morphological changes of the looping embryonic heart tube along the original ventro-dorsal heart axis. Color code: blue = confluence of veins; brown = atrial segment; yellow = AV-canal; pink = ventricular segment; red = ventricular outflow element. Abbreviations as used before.
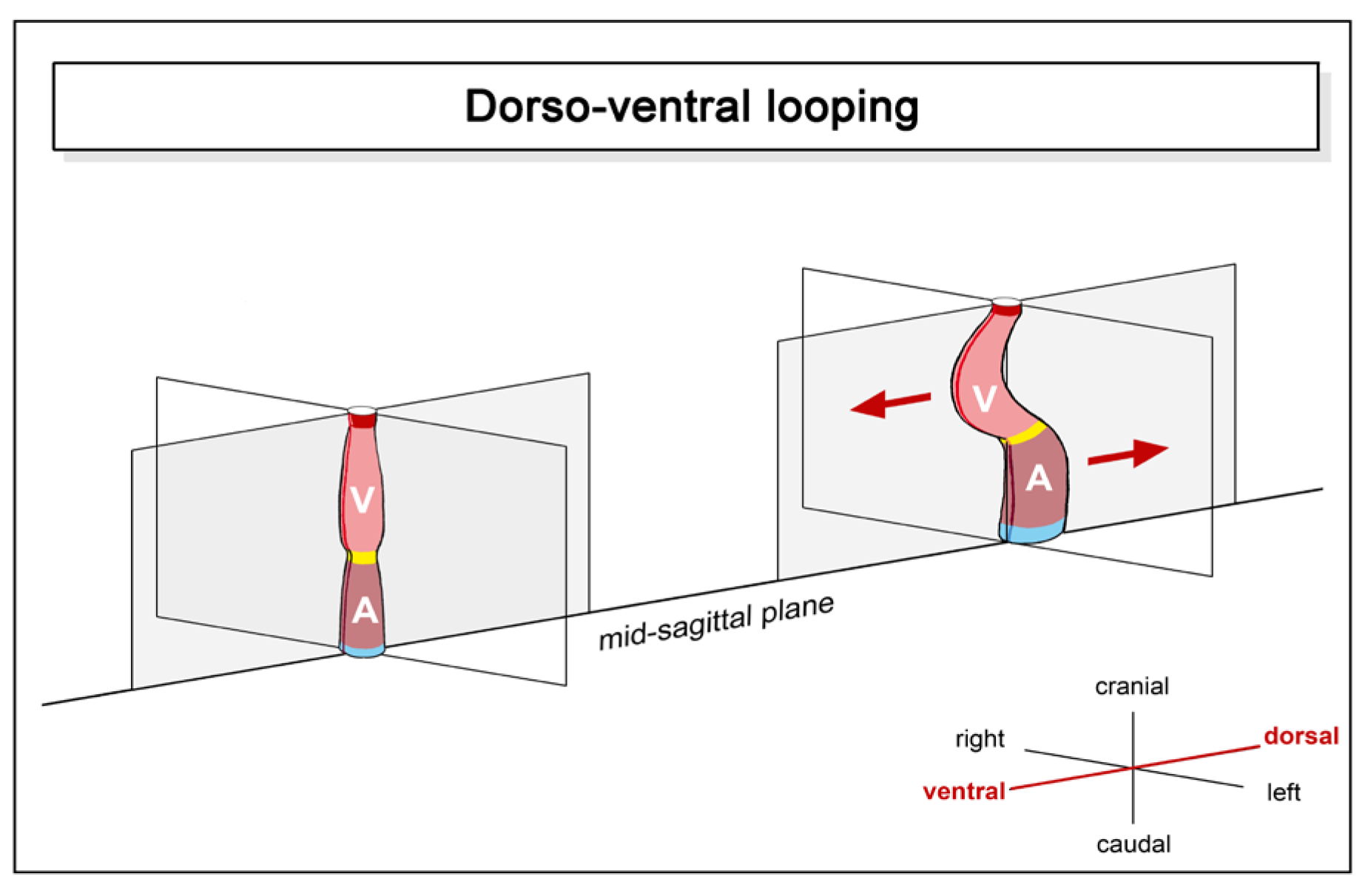
Figure 3.
This schematic drawing illustrates the positional and morphological changes of the looping embryonic heart tube along the original cranio-caudal body axis. Color code and abbreviations as used before.
Figure 3.
This schematic drawing illustrates the positional and morphological changes of the looping embryonic heart tube along the original cranio-caudal body axis. Color code and abbreviations as used before.
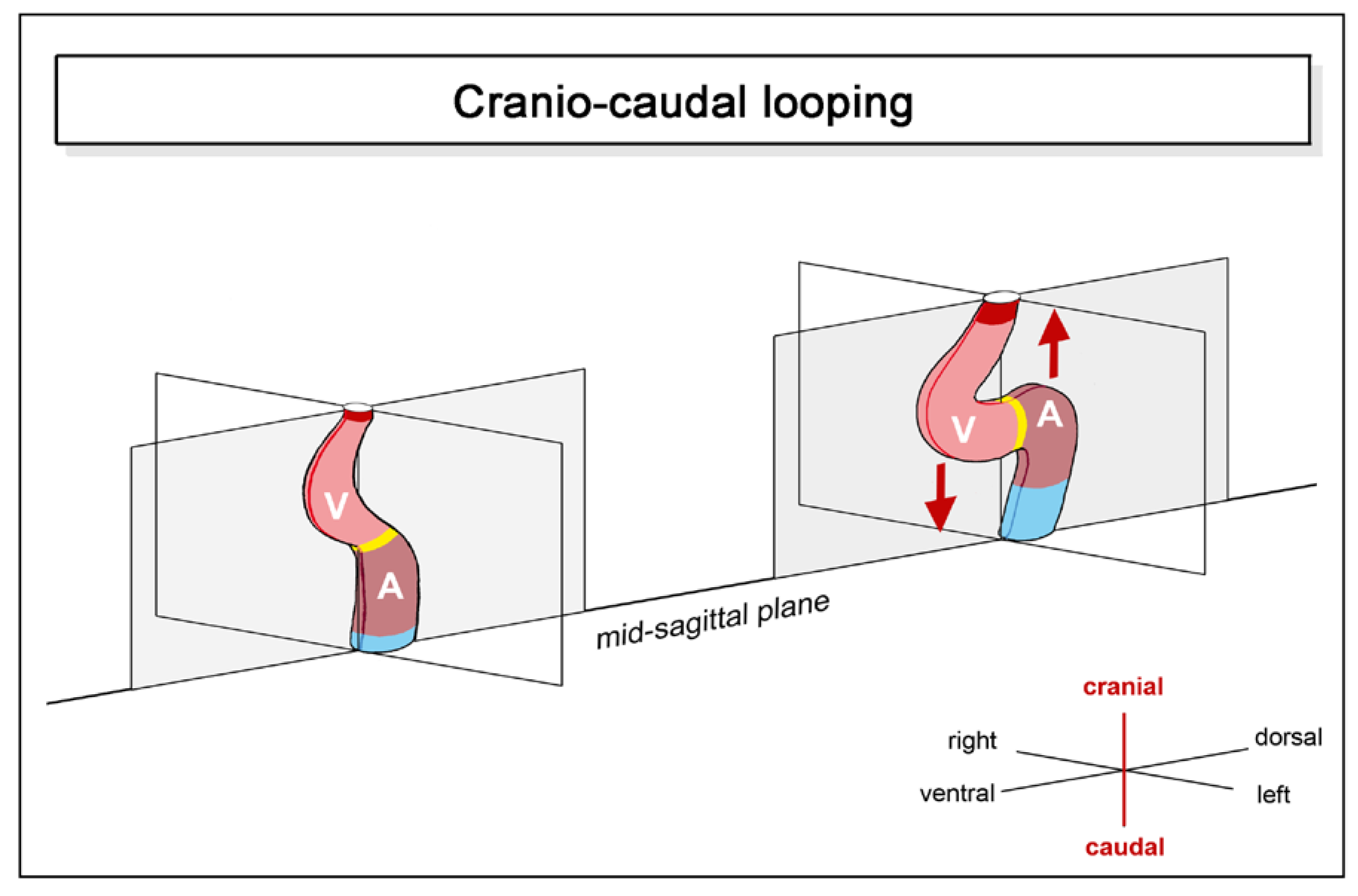
Figure 4.
These schematic drawings illustrate the positional and morphological changes of the looping embryonic heart tube along the left-right body axis; so-called lateral- or chiral looping. Principally two different modes of chiral looping can be found among vertebrates (A, B): The first mode is characterized by rightward rotation of the entire heart around its original longitudinal axis (A). Hearts that loop according to this mode have only a very short outflow element. In the second mode only the ventricular bend and its outflow element undergo rightward rotation (B). Hearts that loop according to this mode have a substantial myocardial outflow element (O). Color code and abbreviations as used before.
Figure 4.
These schematic drawings illustrate the positional and morphological changes of the looping embryonic heart tube along the left-right body axis; so-called lateral- or chiral looping. Principally two different modes of chiral looping can be found among vertebrates (A, B): The first mode is characterized by rightward rotation of the entire heart around its original longitudinal axis (A). Hearts that loop according to this mode have only a very short outflow element. In the second mode only the ventricular bend and its outflow element undergo rightward rotation (B). Hearts that loop according to this mode have a substantial myocardial outflow element (O). Color code and abbreviations as used before.
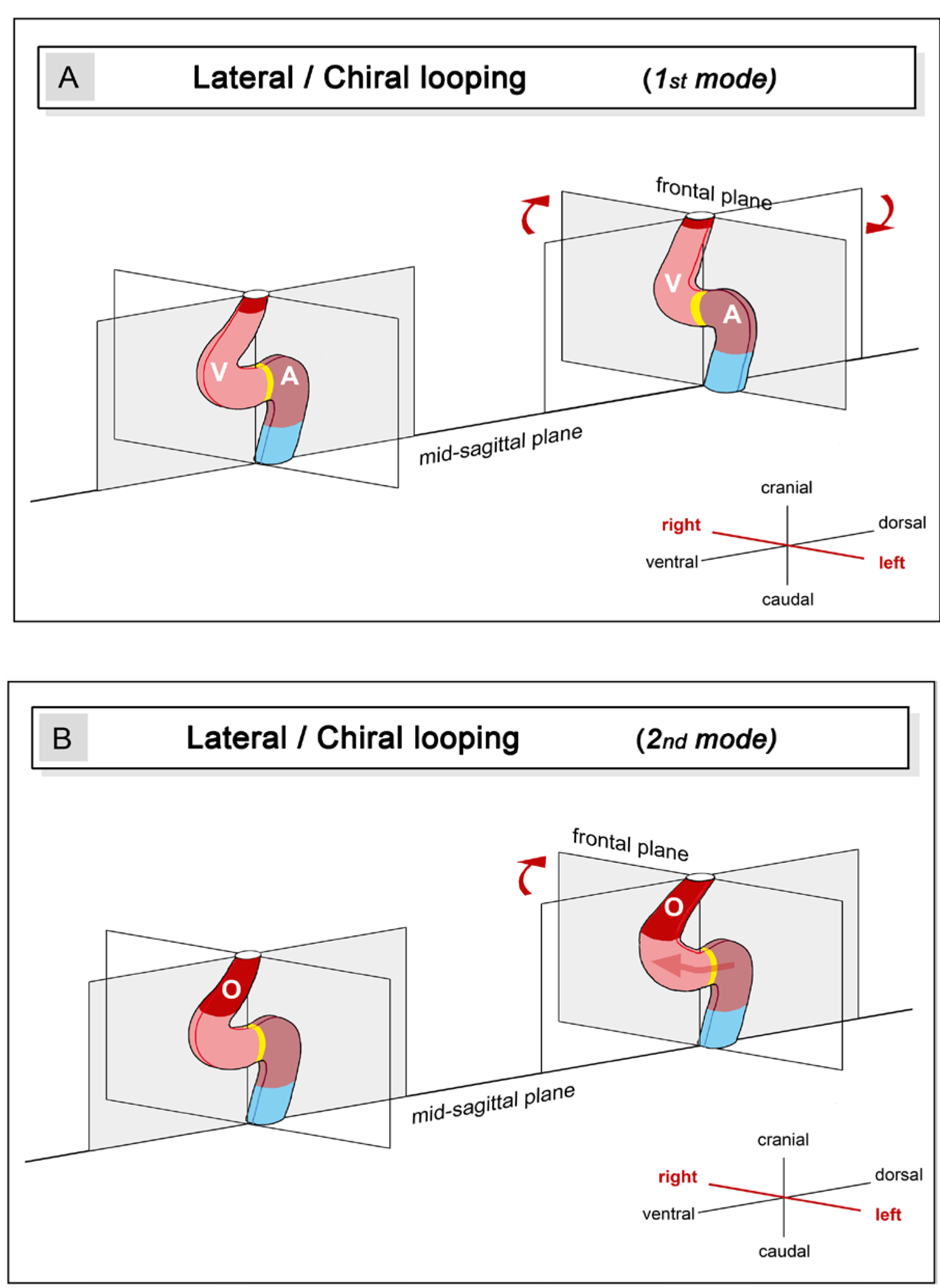
Figure 5.
These schematic drawings depict the three basic types of ‘S-shaped’ heart loops found in vertebrates: (A), ‘plane’ S-loop; (B1), simple helical S-loop; (B2), complex helical S-loop. The blue lines depict the cardiac flow paths. The green lines mark the body midline. Based on own image data from zebrafish (A), and data from [11,36](B1), and [9,37](B2). Color code as used before.
Figure 5.
These schematic drawings depict the three basic types of ‘S-shaped’ heart loops found in vertebrates: (A), ‘plane’ S-loop; (B1), simple helical S-loop; (B2), complex helical S-loop. The blue lines depict the cardiac flow paths. The green lines mark the body midline. Based on own image data from zebrafish (A), and data from [11,36](B1), and [9,37](B2). Color code as used before.
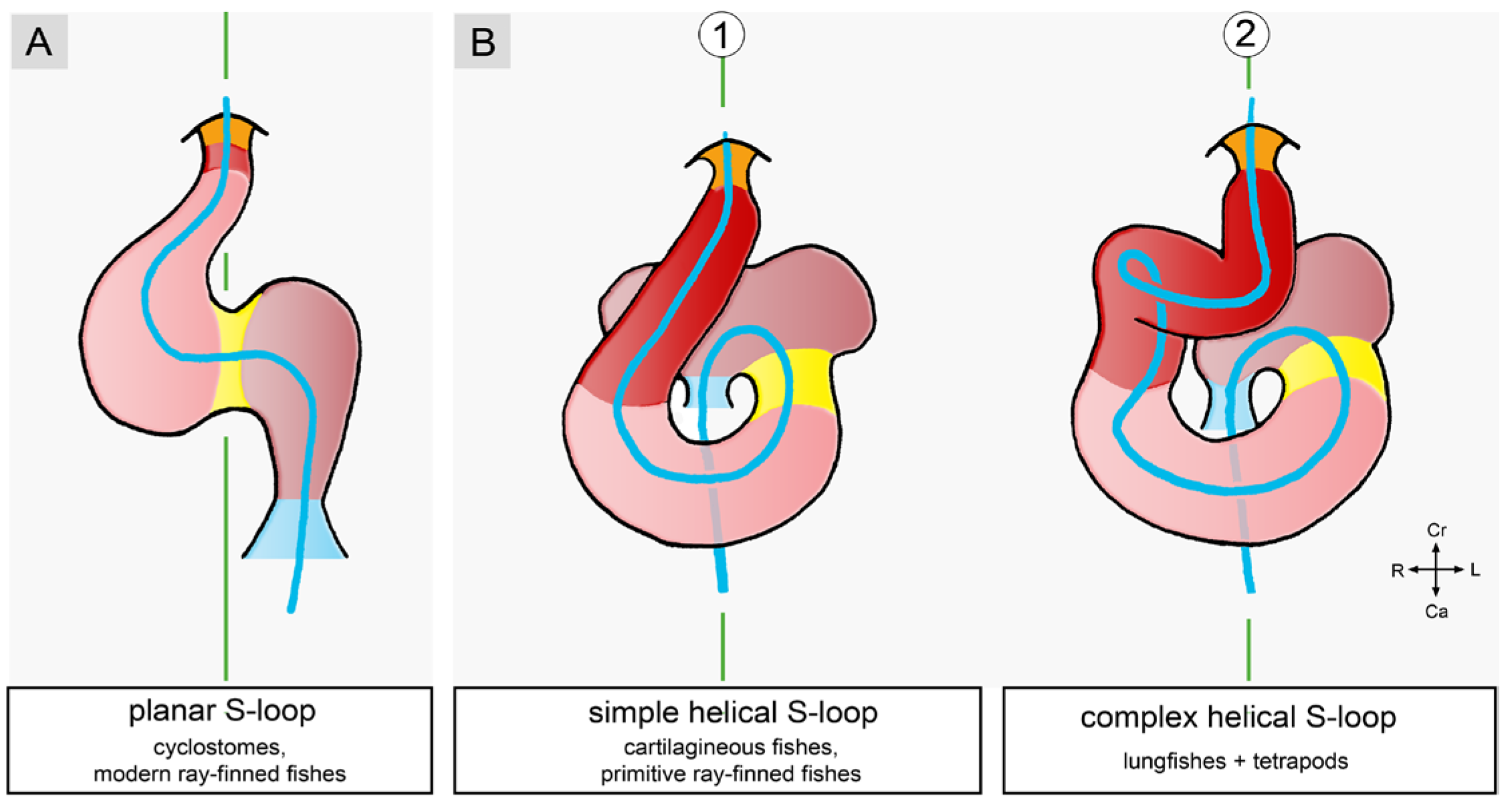
Figure 6.
This schematic drawing illustrates the final positional and morphological changes of the looping embryonic heart tube along the left-right body axis as seen in teleost fishes. The changes may be described as back-rotation around the longitudinal heart axis. Color code and abbreviations as used before.
Figure 6.
This schematic drawing illustrates the final positional and morphological changes of the looping embryonic heart tube along the left-right body axis as seen in teleost fishes. The changes may be described as back-rotation around the longitudinal heart axis. Color code and abbreviations as used before.
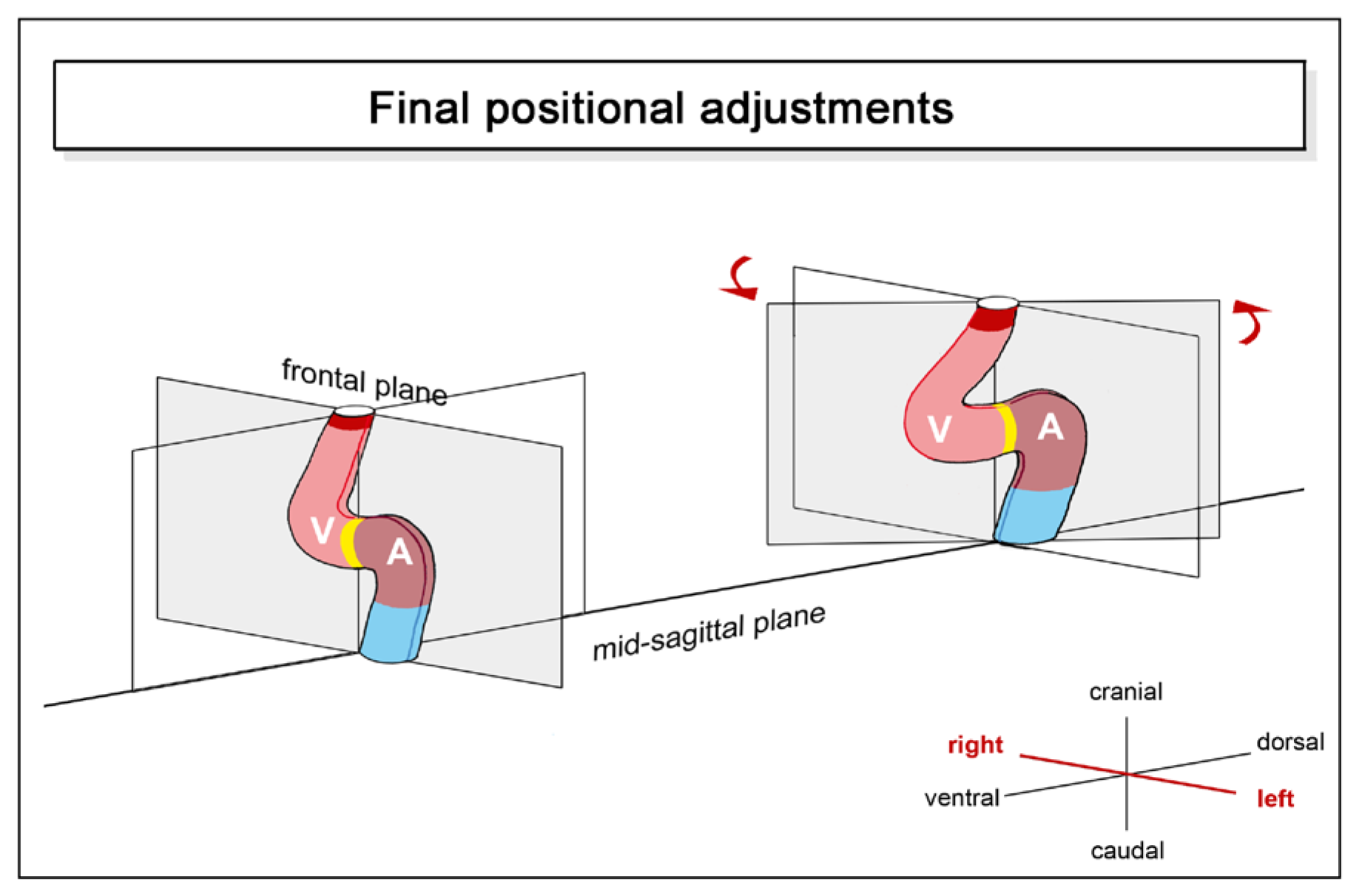
Figure 7.
These semi-schematic drawings illustrate the variation in the degree of sigmoid routing of the cardiac flow path found among the two-chambered hearts of jawed fishes. The spectrum ranges from low degrees (A) to very high degrees of sigmoid routing (D). The situation in the latter hearts resembles the situation found in the left heart of higher vertebrates, such as human beings. The drawings depict hearts from (A) Latimeria columnae, (B) zebrafish, (C) sturgeon, and (D) gar as examples for low, moderate, high, and very high degrees of sigmoid routing of the cardiac flow path, respectively. Further fish species with a similar degree of sigmoid routing of the cardiac flow path are listed in the columns below each example. The angle between the atrio-ventricular plane (AVP) and the cono-ventricular plane (CVP) can be used as a measure for the degree of atrial ascensus. Note that the degree of atrial ascensus does not reflect the position of the species along the phylogenetic tree of vertebrates. Drawings are based on image data from: [48,49](A); own histological specimens (B); [50](C); and [51] (D). Abbreviations: BA= bulbus arteriosus; Co = conus arteriosus; S = sinus venosus; other abbreviations as used before.
Figure 7.
These semi-schematic drawings illustrate the variation in the degree of sigmoid routing of the cardiac flow path found among the two-chambered hearts of jawed fishes. The spectrum ranges from low degrees (A) to very high degrees of sigmoid routing (D). The situation in the latter hearts resembles the situation found in the left heart of higher vertebrates, such as human beings. The drawings depict hearts from (A) Latimeria columnae, (B) zebrafish, (C) sturgeon, and (D) gar as examples for low, moderate, high, and very high degrees of sigmoid routing of the cardiac flow path, respectively. Further fish species with a similar degree of sigmoid routing of the cardiac flow path are listed in the columns below each example. The angle between the atrio-ventricular plane (AVP) and the cono-ventricular plane (CVP) can be used as a measure for the degree of atrial ascensus. Note that the degree of atrial ascensus does not reflect the position of the species along the phylogenetic tree of vertebrates. Drawings are based on image data from: [48,49](A); own histological specimens (B); [50](C); and [51] (D). Abbreviations: BA= bulbus arteriosus; Co = conus arteriosus; S = sinus venosus; other abbreviations as used before.
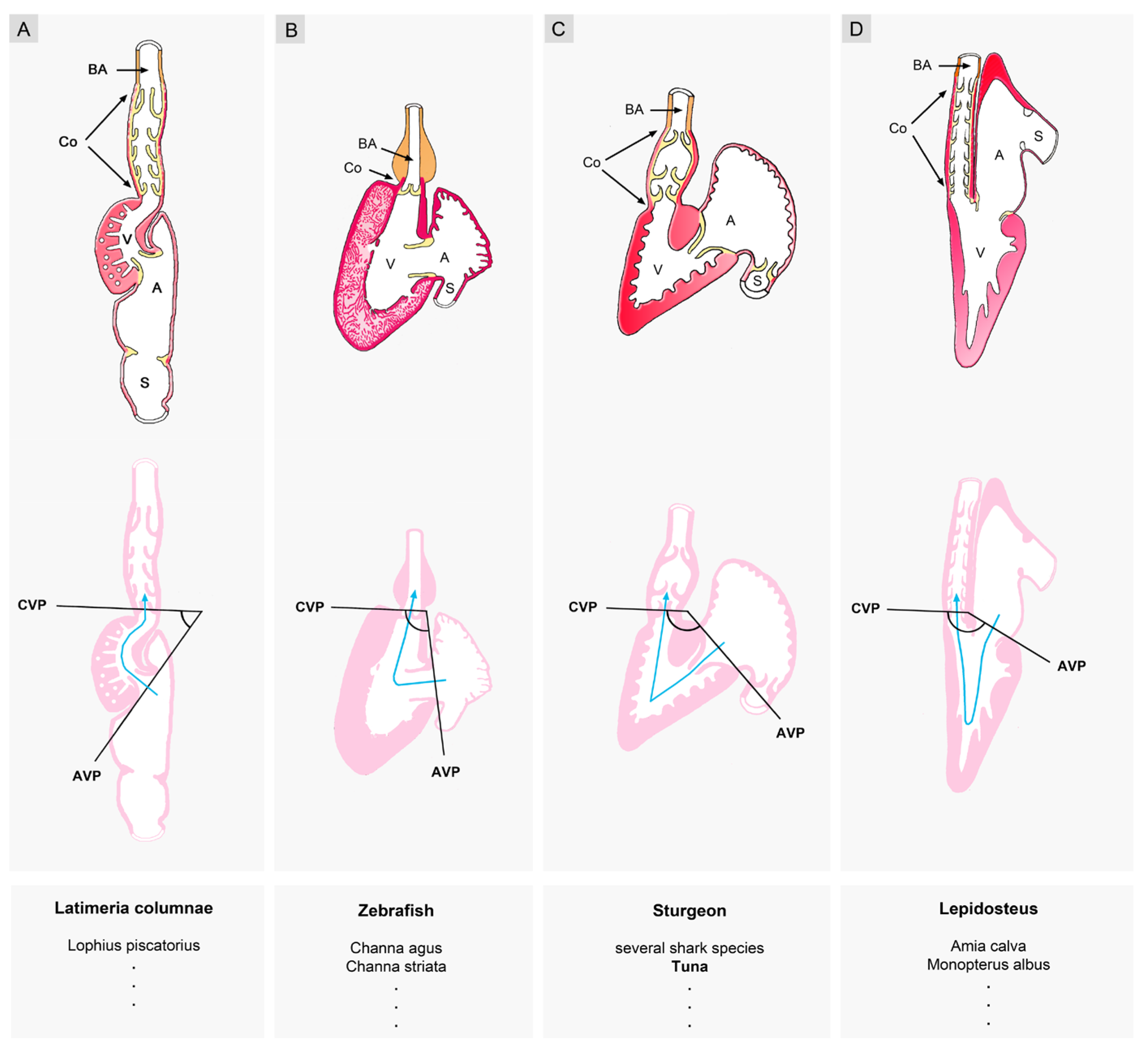
Figure 8.
These semi-schematic drawings illustrate the grouping of fish hearts into three different morphological groups according to the degree of bilateral asymmetry/symmetry: (1), hearts of visually conspicuous asymmetry (A1, B1); (2), hearts of visually obscured asymmetry (A2, B2); and (3), hearts of nearly perfect bilateral symmetry (A3, B3). Panel A depict the hearts in frontal views, while panel B depicts views on the base of the ventricles of the corresponding hearts. The drawings depict hearts from (1), cartilaginous fish (ray); (2), zebrafish; and (3), gar as examples for the respective group. AV marks the mouth of the atrio-ventricular canal; C marks the entrance into the conus arteriosus. The midsagittal body plane is marked by green lines. Drawings are based on image data from [52,60,61,62] (A1, B1, A3, B3), and own specimens (A2, B2).
Figure 8.
These semi-schematic drawings illustrate the grouping of fish hearts into three different morphological groups according to the degree of bilateral asymmetry/symmetry: (1), hearts of visually conspicuous asymmetry (A1, B1); (2), hearts of visually obscured asymmetry (A2, B2); and (3), hearts of nearly perfect bilateral symmetry (A3, B3). Panel A depict the hearts in frontal views, while panel B depicts views on the base of the ventricles of the corresponding hearts. The drawings depict hearts from (1), cartilaginous fish (ray); (2), zebrafish; and (3), gar as examples for the respective group. AV marks the mouth of the atrio-ventricular canal; C marks the entrance into the conus arteriosus. The midsagittal body plane is marked by green lines. Drawings are based on image data from [52,60,61,62] (A1, B1, A3, B3), and own specimens (A2, B2).
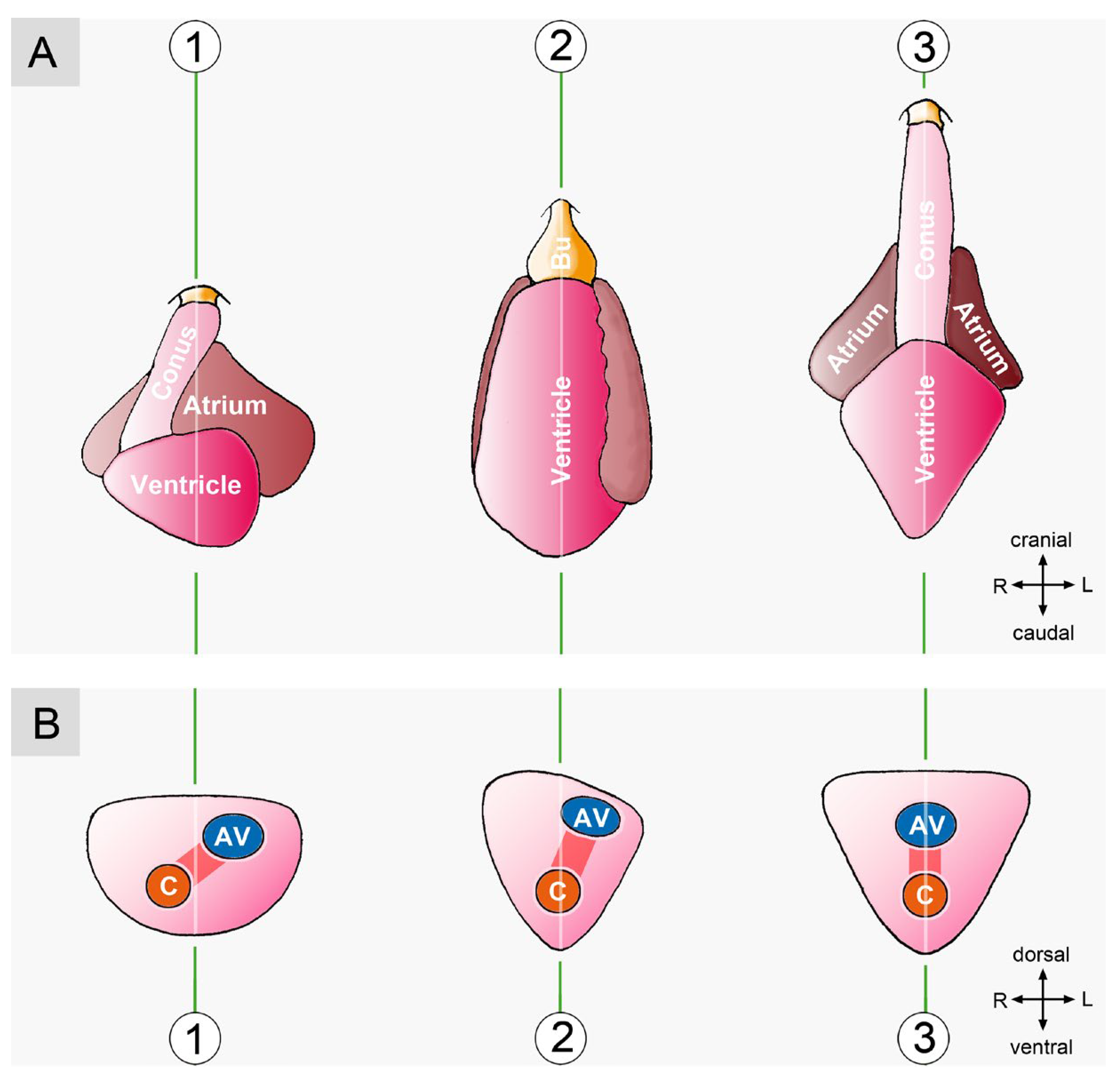
Figure 9.
These drawings depict the bilaterally symmetric heart of a gar (A) and the bilaterally asymmetric heart of a lungfish (B). Note that in the gar, the greatest length of the pericardial cavity (orange arrow) corresponds to the length of the systolic ventricular flow path (blue arrow). In the lungfish, however, there is a striking discrepancy between the greatest length of the pericardial cavity and the length of the systolic ventricular flow path. Hearts are shown at the same relative size. Based on image data from [51,62](A) and [65](B).
Figure 9.
These drawings depict the bilaterally symmetric heart of a gar (A) and the bilaterally asymmetric heart of a lungfish (B). Note that in the gar, the greatest length of the pericardial cavity (orange arrow) corresponds to the length of the systolic ventricular flow path (blue arrow). In the lungfish, however, there is a striking discrepancy between the greatest length of the pericardial cavity and the length of the systolic ventricular flow path. Hearts are shown at the same relative size. Based on image data from [51,62](A) and [65](B).
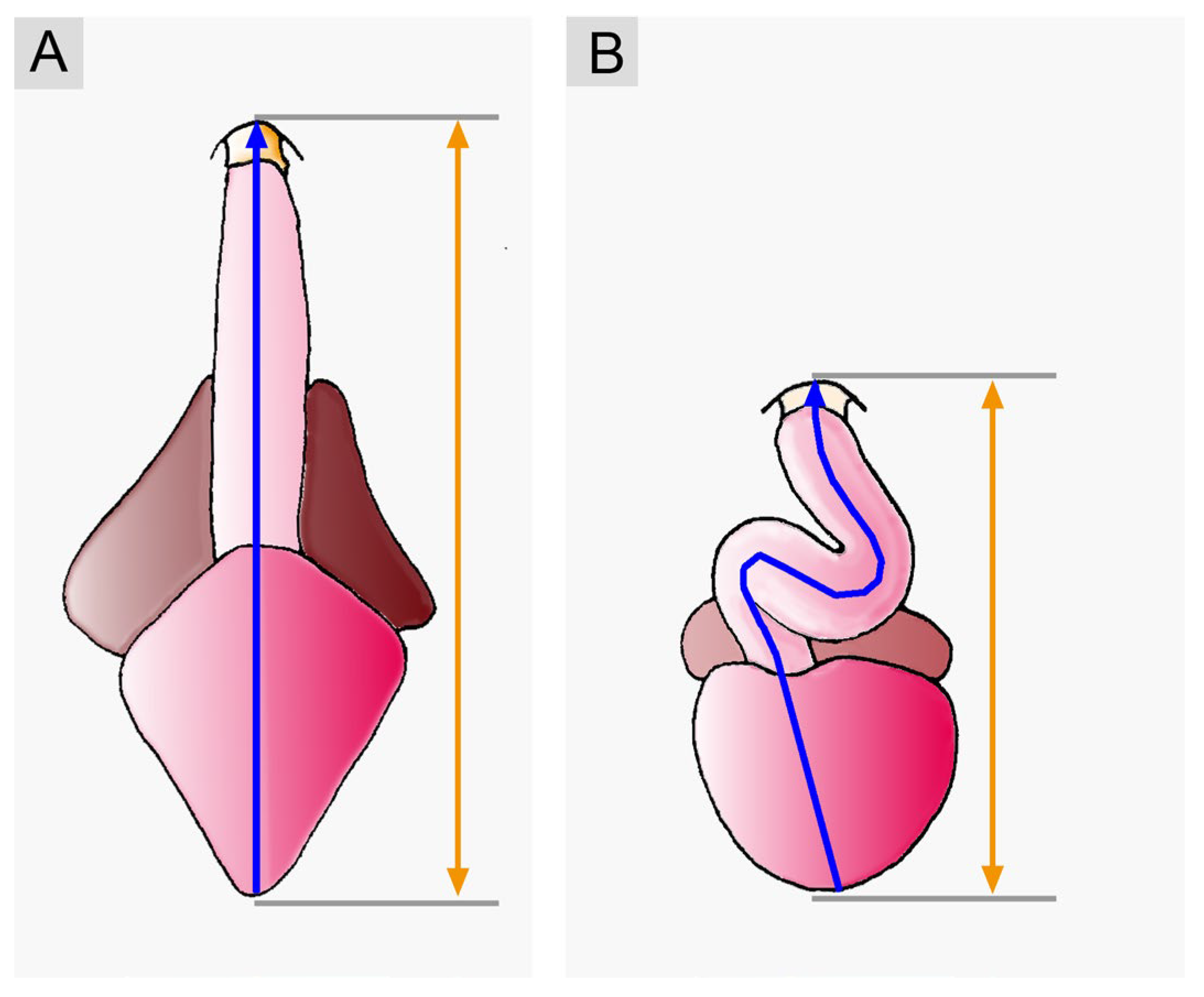
Figure 10.
This scheme illustrates the importance of cardiac looping, especially chiral looping, for the alignment of the flow paths in the multi-chambered hearts of higher vertebrates. In cases of normal left-right patterning of the atrial chambers, rightward rotation of the ventricular bend (ventricular D-loop) sets the scene for correct (concordant) alignment between the atrial and ventricular heart chambers (green background). However, the combination of normal left-right patterning of the atrial chambers with leftward rotation of the ventricular bend (ventricular L-loop) can set the scene for incorrect (discordant) alignment between the atrial and ventricular heart chambers (red background). Abbreviations: AV = atrioventricular; LA = future left atrium; LV = future left ventricle; RA = future right atrium; RV = future right ventricle.
Figure 10.
This scheme illustrates the importance of cardiac looping, especially chiral looping, for the alignment of the flow paths in the multi-chambered hearts of higher vertebrates. In cases of normal left-right patterning of the atrial chambers, rightward rotation of the ventricular bend (ventricular D-loop) sets the scene for correct (concordant) alignment between the atrial and ventricular heart chambers (green background). However, the combination of normal left-right patterning of the atrial chambers with leftward rotation of the ventricular bend (ventricular L-loop) can set the scene for incorrect (discordant) alignment between the atrial and ventricular heart chambers (red background). Abbreviations: AV = atrioventricular; LA = future left atrium; LV = future left ventricle; RA = future right atrium; RV = future right ventricle.
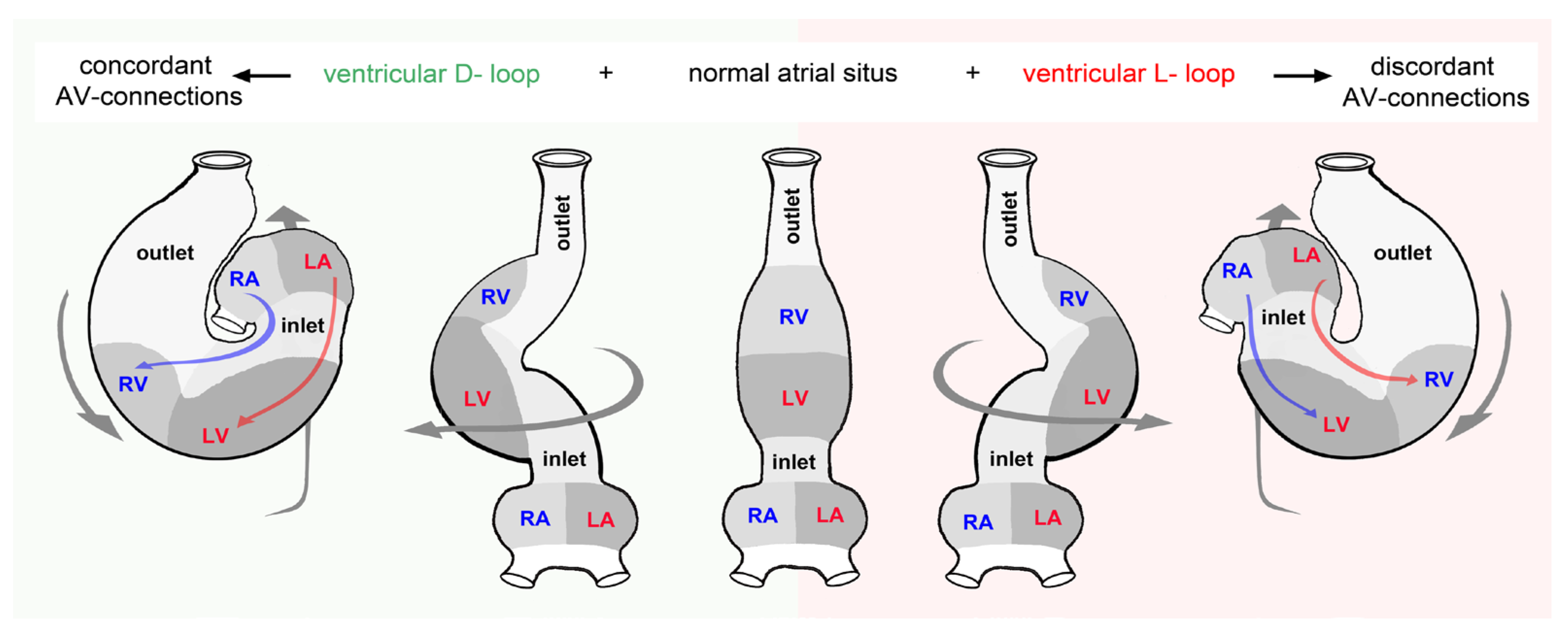
Figure 11.
This scheme illustrates the fact that cardiac looping does not determine the atrio-ventricular as well as ventriculo-arterial alignment in the two-chambered hearts of fishes. Abbreviations and color code as used before.
Figure 11.
This scheme illustrates the fact that cardiac looping does not determine the atrio-ventricular as well as ventriculo-arterial alignment in the two-chambered hearts of fishes. Abbreviations and color code as used before.
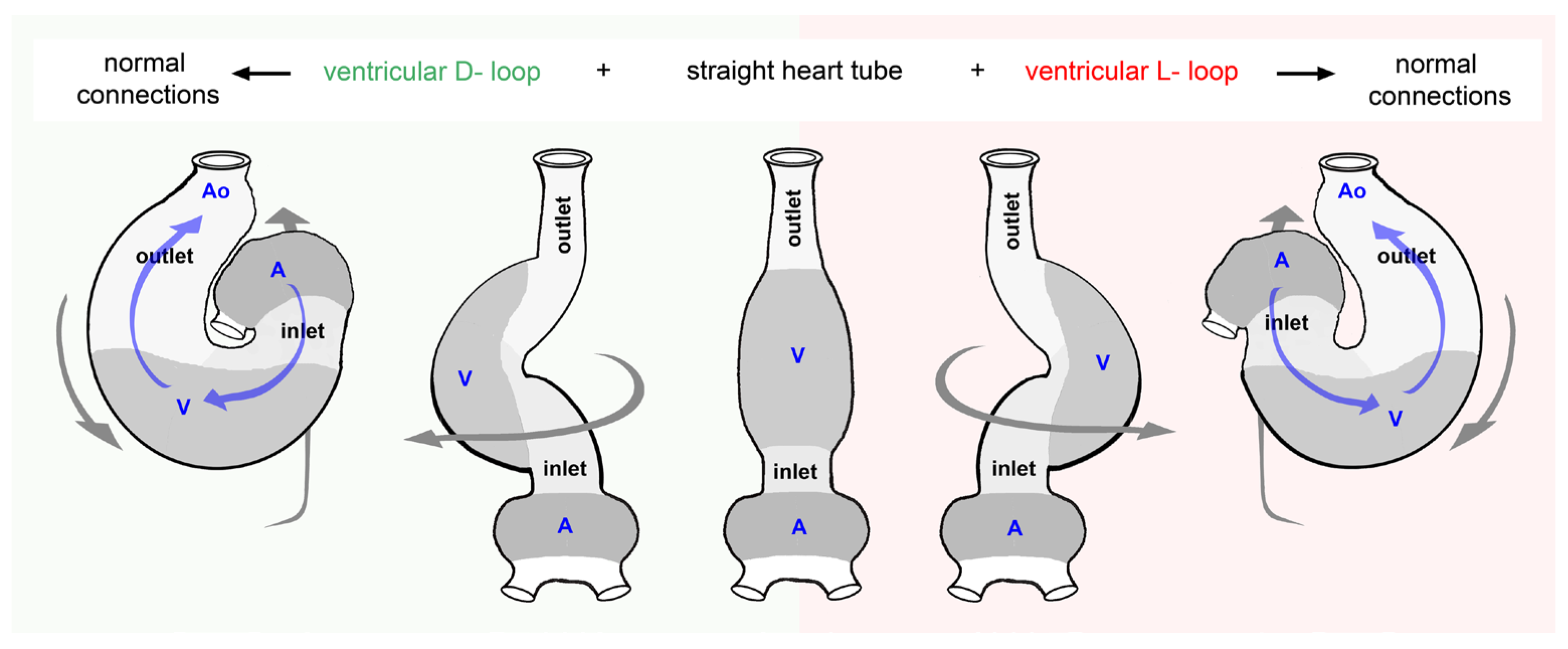
Figure 12.
These drawings depict the relation between the arrangement of the heart chambers and the atrio-ventricular plane displacement (AVPD) during the cardiac cycle. An appreciable AVPD (indicated by transparent green bands) occurs in hearts in which the AVP (marked by green lines) lies perpendicular or almost perpendicular to the ventricular long-axis. Such situations are found in hearts with a very high degree of sigmoid routing of the flow path (ventricular U-turn loop), such as the left heart of human beings (A), or in hearts with a linear arrangement of its chambers, such as the heart of the snail Helix pomatia (B). In hearts with a moderate degree of sigmoid routing (ventricular L-turn loop), such as the zebrafish heart (C), the AVP lies perpendicular to the ventricular short-axis. In this situation, the extent of AVPD is expected to be relatively low since it depends on the degree to which the epicardial short-axis changes during the cardiac cycle (indicated by transparent blue bands). Drawings are based on image data from [98] (A), [105,115](B), and geometric calculations (C). Abbreviations: Ao = aorta; Epi-SAS = epicardial short-axis shortening; LAS = long-axis shortening; other abbreviations as used before.
Figure 12.
These drawings depict the relation between the arrangement of the heart chambers and the atrio-ventricular plane displacement (AVPD) during the cardiac cycle. An appreciable AVPD (indicated by transparent green bands) occurs in hearts in which the AVP (marked by green lines) lies perpendicular or almost perpendicular to the ventricular long-axis. Such situations are found in hearts with a very high degree of sigmoid routing of the flow path (ventricular U-turn loop), such as the left heart of human beings (A), or in hearts with a linear arrangement of its chambers, such as the heart of the snail Helix pomatia (B). In hearts with a moderate degree of sigmoid routing (ventricular L-turn loop), such as the zebrafish heart (C), the AVP lies perpendicular to the ventricular short-axis. In this situation, the extent of AVPD is expected to be relatively low since it depends on the degree to which the epicardial short-axis changes during the cardiac cycle (indicated by transparent blue bands). Drawings are based on image data from [98] (A), [105,115](B), and geometric calculations (C). Abbreviations: Ao = aorta; Epi-SAS = epicardial short-axis shortening; LAS = long-axis shortening; other abbreviations as used before.
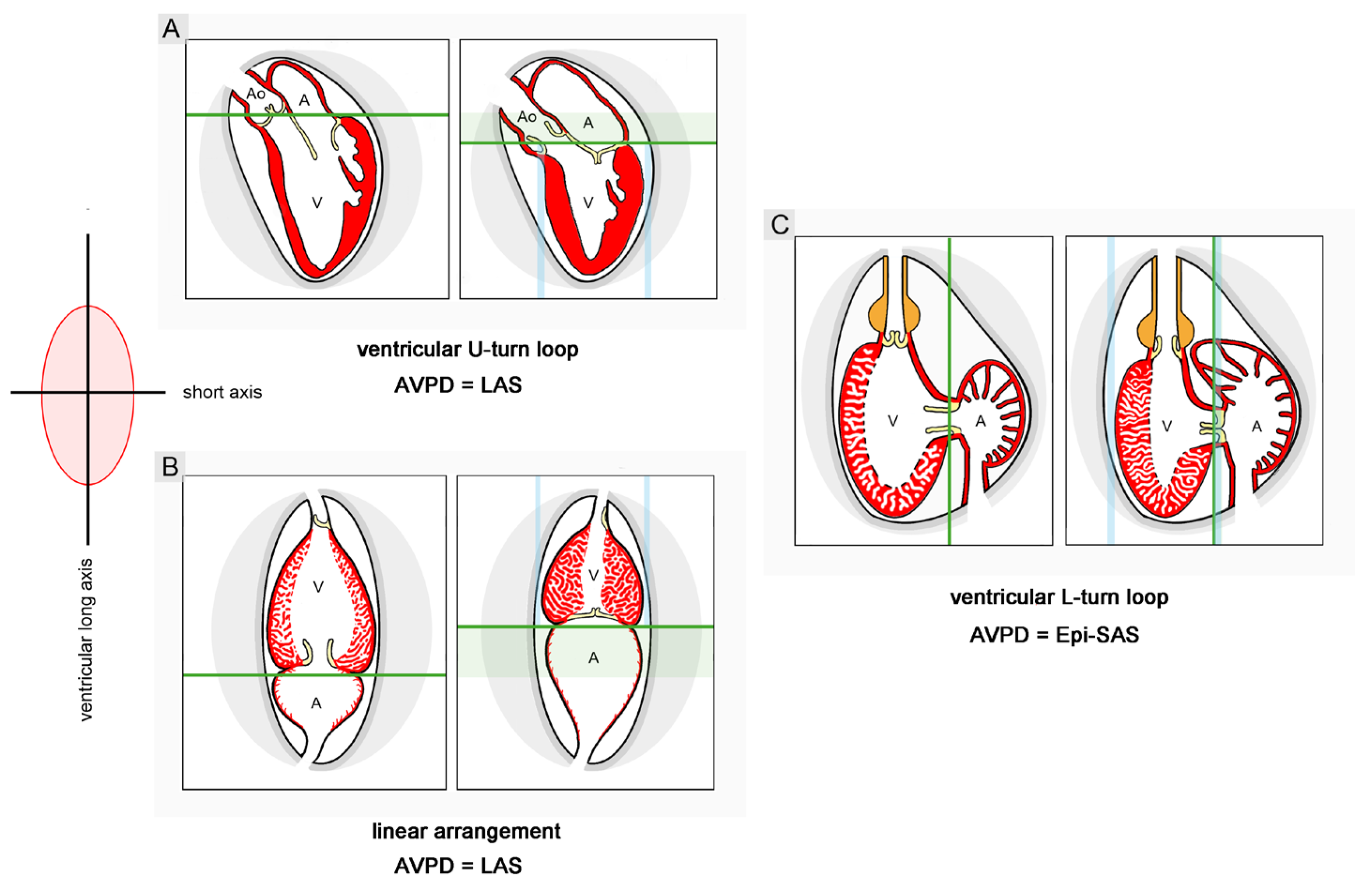
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



