Submitted:
11 July 2024
Posted:
11 July 2024
You are already at the latest version
Abstract
Histone changes that are adjustable and transmitted without altering the DNA sequence are part of epigenetic control. In many disorders, most famously malignancy, abnormal genetic information is seen as a result of mismanagement of typical epigenetic mechanisms. A paradigm change in the diagnosis and treatment of cancer has been brought about by recent discoveries about the processes of epigenetic modifications, chromatin remodeling, and noncoding RNAs implicated in the changed expression patterns of tumor cells. There is a significant increase in the search for drugs that could alter the epigenetic environment altered by tumor cells and benefit from their therapeutic potential. Phytochemicals with a high concentration of naturally produced phenol molecules known as flavonoids can regulate genomic processes. Understanding the precise epigenetic alterations brought on by flavonoid-mediated mechanisms is necessary to develop combination therapy techniques and genomic therapies for cancer. This article aims to thoroughly examine the epigenetic modifications of flavonoids, including their anticancer properties.
Keywords:
cancer
; Chemotherapy
; DNA methylation
; Epigenetic
; Flavonoids
1. Introduction
Epigenetics studies how cells control gene activity without changing the DNA sequence itself. Various factors, including environment, behavior, and age, can influence these modifications. With no alterations to the base DNA code, epigenetic modifications are created even during the initial phases of cell diversification. They are permanently inherited across successive dividing cells to produce diverse cellular characteristics [1,2]. As depicted in many studies, chromatin remodeling is a component of the genomic control of gene expression. It, therefore, is controlled by processes including gene methylation, histone changes, and the actions of noncoding RNA. Although these epigenetic alterations are necessary for organisms to mature normally, their dysregulation can result in some pathological conditions, such as malignancy [3]. The epigenetics in cancer is presented in Figure 1.
Biologists generally agree that disease can be viewed as just a genetic inevitable. As humans grow, genetic mutations increase in our cells. The changes may occur naturally as a result of mistakes in the replication of DNA, or either chemicals or mechanical gene mutations may bring them on. Thus, the ecological or exogenous variables produce alterations in 90–95% of total cancers, while in the other 10% of the overall cases, the genetic component of malignancy is endogenous [4]. It is fascinating to consider that several gene abnormalities in malignancies are caused by epigenetic modifications that alter the expression pattern of these genes rather than changes in their sequence [5,6]. New advances in epigenetics have brought attention to widespread epigenetic aberrations that can emerge in cancerous cells in the initial phases of cell cycle growth. Epigenetic alterations can be used as new pharmacological targets for such cancer therapy because they are programmable and appear at the onset of cancer development in healthy cells [7,8]. At the same time, the number of new instances of cancer worldwide is 2 million in the year 2024. with approximately 10 million fatalities as a consequence, the number of cancer-related fatalities has been falling since 1991, and the mortality rate has decreased since 2021 [9,10]. The development of targeted therapies is responsible for increased patient survival rates. Chemotherapy drugs, radiation treatment, and immunotherapy are frequently employed for cancer treatment, so these therapies demonstrate great potential [11]. Histone deacetylase antagonists, other epigenetic stimulators, and specific cutting-edge techniques, such as chimeric antigen receptor-engineered immune cells (CART cells), have already been utilized to treat cancers with abnormal epigenetic changes [12]. Unfortunately, the majority of cancer treatments, including immune therapeutics, are linked to a variety of adverse reactions [13].
The search for alternative medicines with minimal side effects has, therefore, grown in light of the negative impact that radiotherapy and chemotherapy drugs have on patients. The antitumor activities of phytonutrients are investigated because they best meet this requirement. Anti-carcinogenic components in plant-based products have been shown to prevent cancer through several methods, including immunomodulatory and antioxidant actions. Fascinating reports suggest that phytonutrients can influence how histone deacetylases (HDACs) and DNA methyltransferases (DNMTs) regulate epigenetic activities [14]. An essential family of phytonutrients called flavonoids is linked to altering cancer-related gene expression patterns, as presented in Figure 1 [15]. The role of epigenetic abnormalities in malignancy is first covered in this paper, after which a summary of the potential of flavonoids as chemotherapy and anti-carcinogenic agents is provided. The statistics of epigenetic modulators are presented in Figure 2.
2. Cancer Epigenetics
Nucleosomes are formed in eukaryotes by wrapping DNA around the core of histones. These nucleosomes can either disappear or permanently assume a compressed state by wrapping themselves over. Various genetic regulation processes regulate how much a gene is transcribed within every cell, most of which disrupt chromatin structure. Refer Figure 3.
Genes become hetero-chromatinized and dormant due to the tight packaging of nucleosomes that occurs during nuclear condensation. In contrast, chromatin de-condensation results in nucleosome opening and enhanced gene function. Normal cell transition into cancerous cells includes epigenetic changes and is typically accompanied by genetic variations. DNA changes (such as methylation), histone changes (such as phosphorylation, ubiquitination, and deacetylation), chromatin position, and noncoding RNAs (such as siRNA, lncRNA, and miRNA) are a few of the significant systems occurring in epigenetic control, as presented in Figure 2. Cell equilibrium is significantly impacted by these pathways [16,17]. These changes are often implicated in human pathologies, including solid tumors and hematological neoplasms [18]. The epigenetic deregulation in cancer resulted into the alteration of transcription along with the translation of oncogenic gene targets as presented in Figure 4.
1.1. DNA Methylation
DNA methylation or hydroxy-methylation is a crucial epigenetic process for controlling the expression of genes in cells [19], and DNA methylation sequences help to create epigenetic recollection. A family of DNA methyltransferase catalysethe transfer of a methyl group from S-adenyl methionine (SAM) to the fifth carbon of cytosine residue to form 5-methylcytosine. So, SAM is a key donoe of methyl group in this process. [20]. However, the placement of modified bases regarding the coding sections of a protein being regulated affects the relationship between DNA methylation and gene function, which is often negatively associated [21].
Most frequently, cytosine methylation occurs at CpG sites and is found throughout the entire genomic DNA [22]. Whenever an area of a CpG site has a size of much more than 200 bp, a GC concentration of much more than 55%, and an identified CpG ratio of >60%, it is referred to as a CpG island [23]. Over 70% of mammalian promoter regions contain CpG islands, which are especially common there. When CpG locations are found in the enhancer or promoter parts of genes, their methylation suppresses the expression of genes; nevertheless, if CpG spots are found in the programming sections of genes, their methylation increases gene transcription [24]. In contrast to typical cells, cancerous cells have a different trend of CpG methylation [25]. Additionally, hypermethylation on their promotor gene is highly expressed in the case of human telomerase gene (hTERT) and TERT hypermethylated oncogenic region (THOR) associated with telomerase activation stable under various epigenetic drugs [26]. Whereas most CpG locations in the DNA stay methylated in healthy cells, the CpG islands that precede regulators are unmethylated, permitting effective transcription. CpG islands are often hypomethylated, and CpG dinucleotides exhibit approximately 50% lower methylation in cancerous cells than in healthy cells [27]. Typically, oncogene regulators in malignancy are hypomethylated and triggered, whereas those for tumor suppressors become hypermethylated, limiting their activity. Anomalous DNA methylation causes dysregulation of genes that regulate DNA repair, cell growth, motility, and death in tumor tissue [28]. DNMTs, made up of the three isoenzymes DNMT1, DNMT3, and DNMT3a, are responsible for this genetic DNA methylation activity. The enzyme Ten-elevan translocation (TET) catalyses the oxidation of 5-methylcytosine, leading to sequential removal of methyl group [29].
2.1. Histone Modifications
Positively charged molecules called histones are involved in the packing and compressing DNA to nucleosomes within the nuclei. Euchromatin, an open chromosome architecture, is linked to gene transcription, while heterochromatin, an enclosed nucleosome structure, is linked to transcriptional regulation. The methylation, acetylation, and phosphorylation of histones, in particular, could control chromatin conformational changes that affect the expression of genes[30,31]. The DNA methylation and histone modifications are presented in Figure 5. Acetyl CoA is synthesize within the cell mainly by glycolysis, fatty acid oxidation and amino acid catabolism pathways. Then Acetyl-CoA is transported from the cytoplasm into nucleus, and inside the nucleus, acetyl CoA serve as substrate for histone acetyltransferase (HAT), which add an acetyl group to lysine groups found in the histone tail at a time of histone acetyltransferase (HAT) process. The DNA-histone connection is weakened by this alteration, which ultimately causes chromatin to decondense and transcriptional expression to rise. Lactylation is Post translation modification of histons, which involves the addition of a lactyl group to lysine resudues. This modification has been identified as a significant factor in the regulation of gene expression, which causes supression of immune system, therapy resistance, and metastasis inhibition of antitumor immune response. As lactate is produced as a consequence of anaerobic glycolysis, lactylation starts. Because of the Warburg effect or hypoxia, cancer cells generate a lot of lactate. Lactyl-CoA, the donor molecule for the lactyl group, can be produced from lactate. The ε-amino group of lysine residues on histone proteins receives the lactyl group from lactyl-CoA. Gene expression and chromatin structure can be impacted by histone lactylation. Depending on the situation and the particular lysine residues involved, lactylation can either stimulate or repress the transcription of a certain gene by altering histones [32].
In contrast, HDACs are a significant type of enzyme that deacetylates e-amino compounds on lysine residues, which causes nucleosomes to condense and reduces the expression of genes. Researchers have discovered that benign and cancerous cells exhibit strikingly distinct patterns of histone acetylation [33]. Tumors have lower levels of monoacetylation of Lys20 and contain three methods of Lys16 on histone H4 [34]. Moreover, reduced acetylation of histone modifications H3 and H4 is associated with the development of cancer [35]. Another epigenetic alteration that controls the expression of genes in malignancy is histone methylation. Histone demethylases (HDMs) and histone methyltransferases (HMTs) of the appropriate amino acids in histone proteins catalyse these alterations. The control of histone methylation is frequently linked to the transcriptional patterns of particular cancer genes [36]. For example, a reduction in trimethylation on histone H3 lysine 4 (H3K4me3) linked to malignancy is accompanied by a rise in mono-methylation on lysine 9 of that same histone H3, which has an impact on the expression of genes [37]. Another histone posttranslational change caused by kinases relevant to a cell is activation [38]. Genomic instability eventually causes cancer due to the phosphorylation of serine at the C-terminus of the H2A histone close relative X (H2AX), a genome double-strand break indicator [39]. RNAs that do not encode noncoding RNAs, which have been demonstrated to impact gene transcription significantly, constitute the following critical epigenetic process determining how cells function [40,41]. MicroRNAs are small nucleotide molecules that often link with the 3' domain of the mRNA to alter mRNA stabilization and translation. Numerous miRNAs can impact the activity of a single gene, or just one miRNA might control the function of numerous genes [42].
2.2. Non-Coding RNAs
MiRNAs have been shown to regulate the transcription of approximately 60% of human genes that encode proteins [43]. Many miRNAs have indeed been linked to the control of oncogenes, implicating them in cancer development [44]. For example, miR-16Let-7 and miR-15 are the most prevalent onco-miRNAs (miRNAs linked to cancer) and are promising possibilities for the treatment of cancer [45]. A tumor suppressor gene role for miR-125b1 has additionally been demonstrated, and its decline has been linked to prostatic as well as ovarian tumors [46]. A large portion of the newly discovered information about cancers is influenced by microscopic RNAs' functions in tumor growth [47]. Generic changes, promoter hypermethylation, and other epigenetic alterations all control how miRNAs are expressed, which enhances community cell changes and the cancerous process [48,49,50]. Polyadenylated RNAs with a size exceeding 200 nucleotides are known as longer noncoding RNAs (lncRNAs), and they can attach to peptides, RNA, and DNA. One of the most prevalent techniques for controlling the expression of genes via lncRNAs is epigenetic modification, which is frequently linked to genetic suppression. According to research, lncRNAs could indeed behave as tumor suppressor genes or oncogenes through several processes, such as interacting with the polycomb repressive complex (PRC), which controls the manufacturing, stability, and transformation of transcripts; interacting with miRNAs, which are genomic boosters and regulatory proteins; and interacting with transcriptional regulators, which regulate the generation and transfer of transcripts [51,52,53,54,55]. Tumor patients have higher levels of lncRNAs, such as HOTAIR and MALAT1, which are linked to metastases, and lower levels of lncRNAs, such as MEG3 and PTENP1, which prevent the growth of cells and their movement [56]. Telomeric repeat-containing RNAs (TERRA) are small RNA molecules synthesized from telomeric regions, TERRA, naturally occurring ligands of telomerase and their genetic and epigenetic regulations in cancer aging-associated diseases. It plays a role in specific osteosarcomas and liposarcomas [57]. Long noncoding RNA (lncRNA) H19 leads to the progression of liver cancer by affecting biological processes, cell proliferation, apoptosis, invasion, and epigenetic modification [58]. Soy-isoflavone genistein mediated downregulation of miR-155 and inhibit mammary tumor growth and metastasis in highly metastatic MDA-MB-435 cancer cells in immunocompromised mice. Genistein produces its effect without affecting nonmetastatic MCF-7 breast cancer cell viability [59].
3. Flavonoids As Epigenetic Modulators
With a fundamental benzo-c-pyrone composition, flavonoids are a significant group of organically produced low-molecular-weight polyphenolic chemicals. Such secondary metabolites of plants are prevalently found in various vegetables, fruits, cereals, and nuts, especially in several drinks (tea and coffee). Various well-being products are associated with flavonoids, which also play a significant role in several pharmacological, nutritional, cosmetic, and medical uses. Flavonols, flavan-3-ols, flavanones, flavones, anthocyanidins, and isoflavones are some of the subclasses of flavonoids according to their biochemical compositions. In addition, flavonoids have various advantageous medicinal effects, such as hepatoprotective, antibacterial, antiviral, cytostatic, analgesic, antiestrogenic, estrogenic, anti-allergic, and apoptotic effects [60,61]. Suppression of kinases, modulation of transcriptional activation, effects on cell growth, and epigenetics are just a few of the molecular pathways believed to be linked to these diverse medicinal operations of flavones. New research indicates flavonoids may restore the typical epigenetic modifications altered during cancer [62,63,64,65,66,67,68]. Such phytochemical compounds typically stop the growth of tumors by attacking important signaling transducers, which restore tumor inhibitor genes and prevent the activation of oncogenes. These modifications and the ensuing antitumor effects are frequently caused by flavonoids' epigenetic stimulatory actions, which change epigenetic catalysts such as HDACs, DNMTs, and HATs.
Moreover, flavonoids are effective at modifying the transcription of lncRNAs and miRNAs, which are altered throughout the disease. Chemotherapy medicines can save lives when used to fight cancer, but one drawback of such medications is that they may have toxic effects on healthy cells. Therefore, it is crucial to have good options free of adverse reactions. Nevertheless, more extensive studies are required to assess their safety and side effects. Inside this area, flavonoids offer encouraging findings that many anti-carcinogenic flavonoids exhibit comparatively reduced cytotoxicity against healthy cells [64,68,69,70,71]. Additionally, when ingested at higher levels, they have been shown to have prooxidant effects on healthy cells, which can result in the generation of free radicals that are genetic mutations and inhibitory effects on necessary enzymes engaged in hormonal metabolic activities [72]. Nevertheless, flavonoids that alter gene expression, such as those used in combinational therapy, may be desirable for future cancer treatments.
4. Cancer Prevention And Therapy By Epigenetically Active Flavonoids
4.1. Flavan-3-Ols/Flavanols/Catechins
Green tea contains a potent polyphenolic component called epigallocatechin gallate (EGCG), a member of the catechin family of flavonoids. Epicatechin 3-gallate, epigallocatechin, and epicatechin make up the remaining ingredients in herbal tea. Several in vitro, in vivo, and human-based studies have suggested that EGCG may have proapoptotic, antiproliferative, anti-invasive, and antiangiogenic characteristics. A wealth of research reveals how green tea determines cancer epigenetic regulation. For the first time, the ability of EGG to suppress DNA hypermethylation of CpG islands by interfering with DNMTs was discovered by Fang et al. [67,73,74,75,76]. The potential actions of EGCG on adult cells with prostate cancer have been demonstrated in another investigation. In patients with prostate cancer, drinking green tea increases the amount of acetylated histone H3 in the total cellular nucleus, which induces epigenetic changes in p21/waf1 and Bax reactivation, causing arrest of the cell cycle and apoptotic cell death. In androgen-dependent cancer cells of the prostate stimulated with green tea catechins, Lee et al. measured the activities of histone acetyltransferase to investigate the control of adrenoceptor acetylation (epigallocatechin-3-gallate, epigallocatechin and epicatechin). These catechins result in the suppression of agonist-dependent androgen receptor (AR) excitation as well as AR-regulated transcriptional regulation in prostate cancer cells [77,78]. In a separate study, the combination of clofarabine as well as EGCG or genistein was reported to inhibit the development of breast tumor cells (MCF7 and MDA-MB231) and cause cell death, which was accompanied by RARB hypomethylation, leading to a massive rise in the concentrations of the transcripts for CDKN1A, PTEN, and RARB. In breast cancer cells from humans, this mixture of flavonoids as epigenetic modulation techniques induces death. It restores tumor suppressor genes repressed by DNA methylation with unusually aggressive potential. Negatively affecting histone deacetylases and DNA methyltransferases causes EGCG to change the transcription of several cancer genes found in human cervical carcinoma cell lines. Additionally, due to noticeable changes in the methylation of a gene promoter of these genes, time-dependent treatment with EGCG caused the stimulation of very well cancer genes (TSGs) in such cells. Recently, Ciesielski et al investigated how EGCG affected human endothelial cell nucleosome remodeling and the histone posttranslational alteration mechanism (HMEC-1 and HUVEC origin). The findings demonstrated that EGCG enhances histone acetylation (H3K9/14ac, H3ac) and modification, including both functional (H3K4me3) and inhibitory (H3K9me3) histone marks. These findings demonstrated the extensive epigenetic capability of EGCG concerning the regulation and activity of epigenome stimulators such as HDAC7, HDAC5, KMT2A, CREBP, LSD1, and p300 [79,80,81]. Additional research demonstrates that EGCG's anticancer activity was achieved in HeLa cells via synchronized transcription alteration of various biological targets via various signaling pathways. Many chromatin modifiers, particularly histone substitutions as well as DNA methyltransferases (DNMT3A, DNMT3B, AURKA, DNMT1, AURKC, PRMT6, PRMT7, KDM4A, HDAC5, KDM5C, HDAC7, HDAC6, UBE2B and HDAC11), were shown to have their transcriptional activity modulated in this work. Several pro-inflammatory molecules, such as CCNB2, CCNB1, PIK3C2B, TERT, IL6, MMP7, MMP2, MAPK8, and PIK3CA, as well as essential signaling molecules of the Wnt, PI3K, and MAPK networks, were also downregulated. Kang et al. demonstrated that EGCG may effectively prevent IR-induced damage to mice healthy hepatocytes (AML-12) but significantly increase the radiosensitivity of mice hepatic tissue H22 to 60Co. Researchers have also demonstrated that EGCG promotes the miR34a/Sirt1/p53 signaling mechanism, essential for the radiosensitization of H22 cells. According to Deb et al., exposure to green tea polyphenols (GTPs) and their primary component, epigallocatechin-3-gallate (EGCG), promoted the transcription of TIMP3 through epigenetic pathways in adult cells with prostate cancer (DUPro and LNCaP). Additionally, clinical research on EGCG-consuming prostatectomy patients revealed increased plasma TIMP3 concentrations. Nutritional flavonoids may influence miRNAs and other noncoding RNAs in malignancy. Oral treatment with EGCG reduces the miR483-3p-induced metastases of hepatic cancer, according to a current in vivo model by Kang et al. [82,83,84,85,86,87,88]. When used to treat stomach cancer, EGCG targets the LINC00511/miR-29b/KDM2A axis to modify noncoding RNAs and slow tumor development. Transcription of the tumor suppressor miRNAs miR-34a and let-7a was dramatically enhanced by EGCG-capped metal nanoparticles [89,90].
4.2. Flavonols
The most common flavonoids within the diet are called flavonols (3-hydroxyflavones). One of the most prevalent plant flavonols in a wide range of vegetables and fruits, including apples, onions, and strawberries, includes myricetin, quercetin, fisetin, and kaempferol. From an epigenetic standpoint, quercetin's protective advantages for human wellness are assisted by its multifaceted, pleiotropic action. In cervical cancer cell carcinoma (HeLa) cells, quercetin modifies the transcription of several chromatin moderators and decreases the expression of HMTs, DNMTs, and HDACs dose-dependently. Additionally, it reduced the promoter methylation levels for the evaluated tumor suppressor in a sharp dose-dependent manner with recovery of its expression. This effect was shown in both a time-based and dose-dependent manner. Together with BET blockers, quercetin increases death and reduces pancreatic cancer cells' capacity for cell division and spherical formation [91,92,93,94,95,96]. Additionally, it was demonstrated that quercetin exerts certain antitumor activities with the aid of hnRNPA1, a nuclear protein recognized for regulating antiapoptotic gene mRNA synthesis and mRNA exportation [97,98,99]. Nwaeburu et al. found that quercetin also increased let-7c, inhibiting pancreatic cancer growth by indirectly inhibiting Notch and posttranscriptionally activating Numbl. Zheng et al. found that quercetin nanoliposomes, in combination with butyric acid, tunable aberrant epigenetic changes in Eca9706 molecules through epigenetic-NF-B signaling. In this research, p16INK4 and caspase-3 translations were ready, while opposite expression of global HDAC1, NF-B p65, Cyclin D1, and DNMT1 was downregulated. Moreover, quercetin modulates miR-197/IGFBP5, miR-16-5p/WEE1, miR-1-3p/TAGLN2, miR-16/HOXA10, miR-22/WNT1/β-catenin, miR15a/16, TP53/miR-15/miR-16, miR-145, miR-146a and miR-200b-3p in different types of cancer. Possible HDAC blockers and the anticancer drug kaempferol (3,4′,5,7-tetrahydroxyflavone) are effective against various malignancies. The unique epigenetic action of kaempferol, which HDACs inhibit, was originally described by Berger et al. Kaempferol falls into ligand binding of HDAC2, 4, 7, or 8, and in vitro screening among all preserved adult HDACs of categories I, II, and IV showed that this truly suppressed every HDAC that has been tested.
Moreover, HepG2 and Hep3B (hepatocellular carcinoma cell types) and HCT-116 (intestinal carcinoma carcinoma cells) exhibit hyperacetylation of histones in response to kaempferol. In gastric cancer cells, kaempferol causes the death of autophagic cells by blocking the HDAC/G9a axis, including IRE1-JNK-CHOP signaling. Kaempferol promotes the miR-340 production in lung A549 cells, which is associated with death and limits cell growth in NSCLC. Fisetin, a flavonol, is a potent antitumor drug used to block the activities of the Bcl-2 protein family in numerous cancerous cell lines, promote death, slow cellular proliferation, stop the advancement of cell growth, and suppress different phases of cancerous cells. Additionally, it decreases the amount of the oncogene securin, blocks the stimulation of ROS/PKC/p38 MAPK and ERK1/2 signal transduction, and activates NF-B less. Fisetin suppresses pancreatic cancer growth by causing damage to DNA through RFXAP/KDM4A-dependent histone H3K36 demethylation, according to research published earlier by Ding et al. [100,101,102,103,104,105,106,107,108,109,110,111,112].
4.3. Flavones
Flavones, a subgroup of flavonoids with the chemical formula 2-phenyl chromen-4-one (2-phenyl-1-benzopyran-4-one), were frequently discovered in plants such as celery as well as parsley as well as in practically every kind of nutritious grain. Some popular flavones are luteolin, tangeretin, apigenin, chrysin, baicalein, tricin, 6-hydroxy flavone, and rhoifolin. According to Pandey et al., apigenin suppresses class I HDACs, specifically HDAC1 and HDAC3, altering chromatin to cause inhibition and death in adult cancer cells from the prostate. Apigenin reduced MDA-MB-231 cancer cellular growth and tumor formation by inducing G2/M pause and histone H3 acetylation-mediated p21 transcription. By regulating the miR-520b/ATG7 axis, apigenin increases the levels of miR-16 and miRNA215-5p, which suppresses the growth of gliomas and intestinal cancers, respectively, and chemosensitizes doxorubicin-resistant hepatic cancerous cells [113,114,115,116,117]. Wu et al. recently discovered that luteolin hindered triple-negative breast cancer cells with adrenoceptor positivity in their ability to proliferate and metastasize by altering the genomic regulation of MMP9 transcription by decreasing the concentrations of the histone modifications and H3K27Ac and H3K56Ac that stimulate AKT/mTOR.
Additionally, it was previously discovered that luteolin inhibits the spread of triple-negative cancer and reduces the production of ß-catenin to reverse epithelial-to-mesenchymal transitions. Luteolin causes death in colorectal cancer cells by suppressing the expression of calpain, DNMT1, and UHRF1. By controlling the chromatin regulator UHRF1, this study also implies that calpain may be engaged in genomic code inheritance. Via a cooperative method involving EGFR-associated kinase activity, gefitinib and luteolin control the cell growth cycle genes (CCNE2, CDC25A, CCNA2, PLK-1, and CDKN1B) in human prostate (PC-3) cells. The author's group previously demonstrated the interaction of luteolin with type II complex formation on histone H4 and how these phytonutrients likely impact the epigenetic expression of genes.
Furthermore, Farooqi et al. and Mishan et al. carefully analyzed how effectively luteolin can alter the transcription of miRNA in different malignancies. A tangeretin derivative suppresses the growth of tumor stem-like cells and stops the progression of biological cells with prostate cancer by epigenetic changes repairing p21 gene transcription. Breast, human prostate, myeloma, and T24 bladder cancer cells are affected by baicalein (5,6,7-trihydroxyflavone), which inhibits the growth of cancer cells, arrests the replication process, and triggers death. Baicalin hydrate suppresses the proliferation of NPC cells both in vivo and in vitro, according to research by Lai et al. on the genetic effect of this substance in nasopharyngeal cancer (NPC). Baicalin hydrate also boosted m6A RNA methylation rather than DNA methylation and facilitated Suv39H1 gene translation. Additionally, it has been shown that baicalin increases DNA methylation, inhibits apoptotic cell death, increases HSP70 transcription, and enhances mouse organogenesis in vitro. Baicalein modulates the transcription of miR-183, miR-139-3p, miR-25, and miR-196b-5p in multiple myeloma, according to many current studies [118,119,120,121,122,123,124,125].
4.4. Flavanones
Citrus fruits mostly contain fragrant, colorless ketones called flavanones. Other major flavanones include isosakuranetin, hesperetin, naringenin, naringin, eriodictyol, iso-sakuranetin, and their specific glycosides. Frequent citrus flavanone hesperetin promotes DOT1L breakdown and reduces histone H3K79 activation to stop the spread of stomach cancer, demonstrating its epigenetic function. Silibinin, a native flavonolignan, is silymarin's (A milk thistle medicinal plant) most potent naturally occurring chemical [126]. It significantly inhibits the proliferation of many cancerous cells and is effective on its own or in conjunction with various chemotherapy and epigenetic treatments [127,128]. It induced the release of e-cadherin in concert with DNA methyltransferase and histone deacetylase blockers. It prevented the proliferation and spread of adult non-small cell cancer cells in the lungs. These findings are incredibly significant because the failure of E-cadherin, as well as the spread of the disease through epithelial-to-mesenchymal conversion, is linked to a poor outcome and high mortality rates in these tumor tissues.
EZH2 gene expression is decreased by silibinin in adult prostatic tumor (DU145 and PC3) cells, whereas H3K27me3 levels are elevated. Such reactions relied on lowered expression of activated EZH2 and Akt (ser21) (ser473). Other evidence that it alters the epigenome of human prostatic tumor tissues includes decreasing the transcription of histone deacetylases 1-2 (HDACs1-2) while increasing the action of complete DNA methyltransferase (DNMT). According to Hossainzadeh et al., polymersome nanoparticles that contain silibinin suppress the production of carcinogenic miR-182 miRNAs and miR-125b. Citrus fruits contain naringin in the aglycone method known as naringenin (4,5,7 trihydroxyflavanone). It synergistically increased transamidation efficiency, and suberoylanilide hydroxamic acids (HDAC inhibitor) caused toxicity in neuroblastoma cells, although it had no cytotoxicity on healthy nonmalignant molecules. It suggests that naringenin has powerful histone deacetylase inhibiting action, but additional research is required to fully comprehend its possibility for altering epigenetic processes [129,130,131,132,133].
4.5. Isoflavones
Phytoestrogens are isoflavonoids that naturally occur and are primarily present in soybeans, legumes, and soy-based foods. They have a range of strong therapeutic actions, including anti-inflammatory, oxidative, antibacterial, and anticancer properties. These have clear evidence of having estrogenic and/or antiestrogenic effects. Isoflavones are utilized in a variety of complementary treatments for a broad range of menstrual irregularities, including many cancers, osteoporosis, menstrual issues, and heart disease. There seem to be conflicting claims that isoflavones interfere with endocrine function, although most adverse effects are modest and manifest in the digestive organs [134,135,136,137]. Isoflavones include substances such as genistein, daidzein, glycitein, and genistin. The strongest and most physiologically active isoflavone is genistein, which has shown various in vitro and in vivo antitumor and antiproliferative actions on various human malignancies. Soy phytoestrogens (daidzein and genistein) decrease DNA methylation at EPHB2, BRCA1, and GSTP1 promoters in prostatic cell cultures (DU-145 and PC-3). Genomic sequence DNA methylation conditions in prostatic malignancy are modulated by phytoestrogens (genistein and daidzein), according to research by Karsli-Cebioglu et al. In prostatic tumor DU-145 and LNCaP cell cultures, they discovered that daidzein and genistein treatments altered the methylation patterns of 58 genes. Genistein therapy significantly changed the MAD1L1, hTERT, TRAF7, and KDM4B gene modification rates. In cancer malignancy, genistein regulates the production of miRNAs. Genistein activates the miR-34a/RTCB axis in cancers of the head and neck, which causes ROS-associated apoptosis, a reduction in stemness, and inhibition of EMT. Imai-Sumida et al. [138,139,140,141] have discovered that genistein inhibits renal tumors by suppressing chromatin and HOTAIR modeling mechanisms. On the other hand, according to Allred et al., nutritional genistin and genistein could boost the proliferation of estrogen-dependent breast cancer cells in situ. To fully comprehend the characterized nature of such nutritional chemicals, including their ability to influence epigenetics, toxicity, and antitumor qualities, in-depth molecular studies are required in light of such contradictory data [142,143,144,145].
4.6. Anthocyanidins
The sugar-free version of anthocyanins, known as anthocyanidins, is a group of water-soluble vital plant pigments. Delphinidin is a powerful and prevalent anthocyanin flavonoid in colored veggies and fruit, particularly blueberries. According to Kuo et al., delphinidin inhibits the malignant conversion of mouse epidermis JB6 P+ cells by epigenetically reactivating the Nrf2-ARE pathway. Inside the mouse Nrf2 gene promoter between nucleotides -1,226 and -863 from the start of transcription, 15 CpG spots were demethylated, and this action was linked to the stimulation of the Nrf2-ARE axis. In agreement with observed reductions in the protein production of DNA methyltransferase 1 (DNMT3a) and class I/II chromatin deacetylases (HDACs), the CpG activation percentage in the Nrf2 promoter region dropped. The primary anthocyanin molecule delphinidin was found by Jeong et al. to have epigenetic transceivers that cause adult prostatic tumor cells to undergo apoptosis [146,147,148]. Delphinidin was used to decreased the growth of cancerous cells, and a rise in the histone deacetylase function of caspase-3, -7, and -8 activity was observed. Delphinidin was the only substance that could prevent the function of HDAC3, the most prevalent class I HDAC.
Furthermore, the death induced by delphinidin depended on the breakdown of HDAC3 by caspases, which stabilized and acetylated p53. Researchers also noticed that anthocyanidin significantly downregulated many anti-apoptotic genes while efficiently upregulating pro-apoptotic genes controlled by p53. Han et al. found that delphinidin affects the HOT AIR/miR-34a pathway to reduce breast tumorigenesis and suppress colon cancer migration by upregulating the production of miR-204-3p [149,150]. For chemical structure of flavonoids refer Figure 6.
5. Applications of Flavanoids in Cancer
Flavonoids have been found to have a broad range of anticancer properties, including the ability to control reactive oxygen species (ROS)-scavenging enzyme activities, engage in cell cycle arrest, induce apoptosis and autophagy, and reduce cancer cell proliferation and invasiveness. Flavonoids have a dual activity regarding ROS homeostasis; under normal circumstances, they operate as antioxidants, but in cancer cells, they are potent pro-oxidants, activating apoptotic pathways and downregulating pro-inflammatory signaling pathways. Table 1 summarizes the application of various flavonoids in cancer management. Flavonoids exhibit anti-cancer effects by various mechanisms and pathways. It exhibits anti-oxidant activity and neutralizes free radicals, induces apoptosis in cancer cells, inhibits angiogenesis, and deprives tumors of the blood supply. It also induces cell cycle arrest and modulates enzyme activity. In this way, it exhibits effective anti-cancer effects [151].
6. Conclusions
According to the body of research on the pathogenic mechanisms of malignancy, genes, and epigenetics are crucial to the development and spread of the disease. Epigenetic modifications are changeable and can be used to improve treatment plans. Naturally occurring substances such as phytochemicals can control or suppress epigenetic modifiers, including DNMTs, HDACs, HMTs, HATs, and others. Many flavonoids, including flavonols, flavanones, and flavones, were addressed in this overview, along with their ability to treat and fight diseases by altering epigenetic processes. It is common practice to fight cancer with a variety of plant-based medications, such as vinca alkaloids (such as vincristine and vinblastine), taxanes (such as paclitaxel as well as docetaxel), camptothecins, and many others. Many phytochemicals, such as flavonoids, have strong anticancer effects and are now through different preliminary research and clinical testing phases. Its poor bioavailability, poor water solubility, fast absorption by healthy cells, poor pharmacological index, and unfavorable impact on the hepatic system constitute a few of their drawbacks. These problems can be solved by creating unique techniques such as nanomedicines. The importance of phytonutrients is growing in the cure and avoidance of malignancy. They provide essential weapons in the fight against cancer because of their ability to reverse epigenetic changes made by cancerous cells. Combinatorial medicines that combine conventional chemotherapy and precision medicine with plant-based chromatin modifiers can improve treatment strategies and lessen adverse effects. Significant research is needed to identify and characterize anti-carcinogenic phytonutrients and their corresponding mechanisms of action. Given the rapid technical advancement, this looks like a viable endeavour in our effort to combat cancer.
Declarations
Author Contributions
All authors contributed to the study conception, design, data collection, and analysis. All authors read and approved the final manuscript.
Funding
The authors declare that no funds, grants, or other support were received during the preparation of this manuscript.
Competing Interests
The authors have no relevant financial or non-financial interests to disclose.
Data Availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Ethics Approval
Not applicable
Consent to Participate
Not applicable
Consent to Publish
Not applicable
References
- Weinhold B. Epigenetics: The Science of Change. Environ Health Perspect 2006;114. [CrossRef]
- Cheedipudi S, Genolet O, Dobreva G. Epigenetic inheritance of cell fates during embryonic development. Front Genet 2014;5. [CrossRef]
- Egger G, Liang G, Aparicio A, Jones PA. Epigenetics in human disease and prospects for epigenetic therapy. Nature 2004;429:457–63. [CrossRef]
- Anand P, Kunnumakara AB, Sundaram C, Harikumar KB, Tharakan ST, Lai OS, et al. Cancer is a Preventable Disease that Requires Major Lifestyle Changes. Pharm Res 2008;25:2097–116. [CrossRef]
- Cavalli G, Heard E. Advances in epigenetics link genetics to the environment and disease. Nature 2019;571:489–99. [CrossRef]
- Taby R, Issa J-PJ. Cancer Epigenetics. CA Cancer J Clin 2010;60:376–92. [CrossRef]
- Bennett RL, Licht JD. Targeting Epigenetics in Cancer. Annu Rev Pharmacol Toxicol 2018;58:187–207. [CrossRef]
- Dawson MA, Kouzarides T. Cancer Epigenetics: From Mechanism to Therapy. Cell 2012;150:12–27. [CrossRef]
- Siegel RL, Miller KD, Fuchs HE, Jemal A. Cancer Statistics, 2021. CA Cancer J Clin 2021;71:7–33. [CrossRef]
- Siegel RL, Giaquinto AN, Jemal A. Cancer statistics, 2024. CA Cancer J Clin 2024;74:12–49. [CrossRef]
- Yu W-D, Sun G, Li J, Xu J, Wang X. Mechanisms and therapeutic potentials of cancer immunotherapy in combination with radiotherapy and/or chemotherapy. Cancer Lett 2019;452:66–70. [CrossRef]
- Wang X, Waschke BC, Woolaver RA, Chen SMY, Chen Z, Wang JH. HDAC inhibitors overcome immunotherapy resistance in B-cell lymphoma. Protein Cell 2020;11:472–82. [CrossRef]
- Lacouture M, Sibaud V. Toxic Side Effects of Targeted Therapies and Immunotherapies Affecting the Skin, Oral Mucosa, Hair, and Nails. Am J Clin Dermatol 2018;19:31–9. [CrossRef]
- Shukla S, Meeran SM, Katiyar SK. Epigenetic regulation by selected dietary phytochemicals in cancer chemoprevention. Cancer Lett 2014;355:9–17. [CrossRef]
- Busch C, Burkard M, Leischner C, Lauer UM, Frank J, Venturelli S. Epigenetic activities of flavonoids in the prevention and treatment of cancer. Clin Epigenetics 2015;7:64. [CrossRef]
- Brower V. Epigenetics: Unravelling the cancer code. Nature 2011;471:S12–3. [CrossRef]
- Zhang L, Lu Q, Chang C. Epigenetics in Health and Disease, 2020, p. 3–55. [CrossRef]
- Ilango S, Paital B, Jayachandran P, Padma PR, Nirmaladevi R. Epigenetic alterations in cancer. Front Biosci Landmark Ed 2020;25:1058–109. [CrossRef]
- Gao F, Das SK. Epigenetic regulations through DNA methylation and hydroxymethylation: clues for early pregnancy in decidualization. Biomol Concepts 2014;5:95–107. [CrossRef]
- Bird A. DNA methylation patterns and epigenetic memory. Genes Dev 2002;16:6–21. [CrossRef]
- Blake LE, Roux J, Hernando-Herraez I, Banovich NE, Perez RG, Hsiao CJ, et al. A comparison of gene expression and DNA methylation patterns across tissues and species. Genome Res 2020;30:250–62. [CrossRef]
- Jones PA. Functions of DNA methylation: islands, start sites, gene bodies and beyond. Nat Rev Genet 2012;13:484–92. [CrossRef]
- Takai D, Jones PA. Comprehensive analysis of CpG islands in human chromosomes 21 and 22. Proc Natl Acad Sci 2002;99:3740–5. [CrossRef]
- Greenberg MVC, Bourc’his D. The diverse roles of DNA methylation in mammalian development and disease. Nat Rev Mol Cell Biol 2019;20:590–607. [CrossRef]
- Esteller M. Cancer epigenomics: DNA methylomes and histone-modification maps. Nat Rev Genet 2007;8:286–98. [CrossRef]
- Chebly A, Ropio J, Peloponese J-M, Poglio S, Prochazkova-Carlotti M, Cherrier F, et al. Exploring hTERT promoter methylation in cutaneous T-cell lymphomas. Mol Oncol 2022;16:1931–46. [CrossRef]
- Herman JG, Baylin SB. Gene Silencing in Cancer in Association with Promoter Hypermethylation. N Engl J Med 2003;349:2042–54. [CrossRef]
- Cheung H-H, Lee T-L, Rennert OM, Chan W-Y. DNA methylation of cancer genome. Birth Defects Res Part C Embryo Today Rev 2009;87:335–50. [CrossRef]
- Moore LD, Le T, Fan G. DNA Methylation and Its Basic Function. Neuropsychopharmacology 2013;38:23–38.
- Bannister AJ, Kouzarides T. Regulation of chromatin by histone modifications. Cell Res 2011;21:381–95. [CrossRef]
- Karlić R, Chung H-R, Lasserre J, Vlahoviček K, Vingron M. Histone modification levels are predictive for gene expression. Proc Natl Acad Sci 2010;107:2926–31. [CrossRef]
- De La Cruz-López KG, Castro-Muñoz LJ, Reyes-Hernández DO, García-Carrancá A, Manzo-Merino J. Lactate in the Regulation of Tumor Microenvironment and Therapeutic Approaches. Front Oncol 2019;9:1143. [CrossRef]
- Di Cerbo V, Schneider R. Cancers with wrong HATs: the impact of acetylation. Brief Funct Genomics 2013;12:231–43. [CrossRef]
- Fraga MF, Ballestar E, Villar-Garea A, Boix-Chornet M, Espada J, Schotta G, et al. Loss of acetylation at Lys16 and trimethylation at Lys20 of histone H4 is a common hallmark of human cancer. Nat Genet 2005;37:391–400. [CrossRef]
- Audia JE, Campbell RM. Histone Modifications and Cancer. Cold Spring Harb Perspect Biol 2016;8:a019521. [CrossRef]
- Varier RA, Timmers HTM. Histone lysine methylation and demethylation pathways in cancer. Biochim Biophys Acta BBA - Rev Cancer 2011;1815:75–89. [CrossRef]
- Richon VM, Sandhoff TW, Rifkind RA, Marks PA. Histone deacetylase inhibitor selectively induces p21 WAF1 expression and gene-associated histone acetylation. Proc Natl Acad Sci 2000;97:10014–9. [CrossRef]
- Rossetto D, Avvakumov N, Côté J. Histone phosphorylation. Epigenetics 2012;7:1098–108. [CrossRef]
- Bonner WM, Redon CE, Dickey JS, Nakamura AJ, Sedelnikova OA, Solier S, et al. γH2AX and cancer. Nat Rev Cancer 2008;8:957–67. [CrossRef]
- Kurokawa R, Rosenfeld MG, Glass CK. Transcriptional regulation through noncoding RNAs and epigenetic modifications. RNA Biol 2009;6:233–6. [CrossRef]
- Statello L, Guo C-J, Chen L-L, Huarte M. Gene regulation by long non-coding RNAs and its biological functions. Nat Rev Mol Cell Biol 2021;22:96–118. [CrossRef]
- Mohr A, Mott J. Overview of MicroRNA Biology. Semin Liver Dis 2015;35:003–11. [CrossRef]
- Friedman RC, Farh KK-H, Burge CB, Bartel DP. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res 2009;19:92–105. [CrossRef]
- Peng Y, Croce CM. The role of MicroRNAs in human cancer. Signal Transduct Target Ther 2016;1:15004. [CrossRef]
- Esquela-Kerscher A, Slack FJ. Oncomirs — microRNAs with a role in cancer. Nat Rev Cancer 2006;6:259–69. [CrossRef]
- Soto-Reyes E, González-Barrios R, Cisneros-Soberanis F, Herrera-Goepfert R, Pérez V, Cantú D, et al. Disruption of CTCF at the miR-125b1 locus in gynecological cancers. BMC Cancer 2012;12:40. [CrossRef]
- Ali Syeda Z, Langden SSS, Munkhzul C, Lee M, Song SJ. Regulatory Mechanism of MicroRNA Expression in Cancer. Int J Mol Sci 2020;21:1723. [CrossRef]
- Baer C, Claus R, Plass C. Genome-Wide Epigenetic Regulation of miRNAs in Cancer. Cancer Res 2013;73:473–7. [CrossRef]
- Liu X, Chen X, Yu X, Tao Y, Bode AM, Dong Z, et al. Regulation of microRNAs by epigenetics and their interplay involved in cancer. J Exp Clin Cancer Res 2013;32:96. [CrossRef]
- Cammaerts S, Strazisar M, De Rijk P, Del Favero J. Genetic variants in microRNA genes: impact on microRNA expression, function, and disease. Front Genet 2015;6. [CrossRef]
- Marín-Béjar O, Marchese FP, Athie A, Sánchez Y, González J, Segura V, et al. Pint lincRNA connects the p53 pathway with epigenetic silencing by the Polycomb repressive complex 2. Genome Biol 2013;14:R104. [CrossRef]
- Vance KW, Ponting CP. Transcriptional regulatory functions of nuclear long noncoding RNAs. Trends Genet 2014;30:348–55. [CrossRef]
- Zhang X, Wang W, Zhu W, Dong J, Cheng Y, Yin Z, et al. Mechanisms and Functions of Long Non-Coding RNAs at Multiple Regulatory Levels. Int J Mol Sci 2019;20:5573. [CrossRef]
- Hou Y, Zhang R, Sun X. Enhancer LncRNAs Influence Chromatin Interactions in Different Ways. Front Genet 2019;10. [CrossRef]
- Marchese FP, Raimondi I, Huarte M. The multidimensional mechanisms of long noncoding RNA function. Genome Biol 2017;18:206. [CrossRef]
- Huarte M. The emerging role of lncRNAs in cancer. Nat Med 2015;21:1253–61. [CrossRef]
- Sinha S, Shukla S, Khan S, Farhan M, Kamal MA, Meeran SM. Telomeric Repeat Containing RNA (TERRA): Aging and Cancer. CNS Neurol Disord Drug Targets 2015;14:936–46. [CrossRef]
- Wang Y, Zeng J, Chen W, Fan J, Hylemon PB, Zhou H. Long Noncoding RNA H19: A Novel Oncogene in Liver Cancer. Non-Coding RNA 2023;9:19. [CrossRef]
- de la Parra C, Castillo-Pichardo L, Cruz-Collazo A, Cubano L, Redis R, Calin GA, et al. Soy Isoflavone Genistein-Mediated Downregulation of miR-155 Contributes to the Anticancer Effects of Genistein. Nutr Cancer 2016;68:154–64. [CrossRef]
- Panche AN, Diwan AD, Chandra SR. Flavonoids: an overview. J Nutr Sci 2016;5:e47. [CrossRef]
- Kumar S, Pandey AK. Chemistry and Biological Activities of Flavonoids: An Overview. Sci World J 2013;2013:1–16. [CrossRef]
- Woo H-H, Jeong BR, Hawes MC. Flavonoids: from cell cycle regulation to biotechnology. Biotechnol Lett 2005;27:365–74. [CrossRef]
- Pietta P-G. Flavonoids as Antioxidants. J Nat Prod 2000;63:1035–42. [CrossRef]
- Miron A, Aprotosoaie AC, Trifan A, Xiao J. Flavonoids as modulators of metabolic enzymes and drug transporters. Ann N Y Acad Sci 2017;1398:152–67. [CrossRef]
- Jucá MM, Cysne Filho FMS, de Almeida JC, Mesquita D da S, Barriga JR de M, Dias KCF, et al. Flavonoids: biological activities and therapeutic potential. Nat Prod Res 2020;34:692–705. [CrossRef]
- Khan H, Belwal T, Efferth T, Farooqi AA, Sanches-Silva A, Vacca RA, et al. Targeting epigenetics in cancer: therapeutic potential of flavonoids. Crit Rev Food Sci Nutr 2021;61:1616–39. [CrossRef]
- Jiang W, Xia T, Liu C, Li J, Zhang W, Sun C. Remodeling the Epigenetic Landscape of Cancer—Application Potential of Flavonoids in the Prevention and Treatment of Cancer. Front Oncol 2021;11. [CrossRef]
- Carlos-Reyes Á, López-González JS, Meneses-Flores M, Gallardo-Rincón D, Ruíz-García E, Marchat LA, et al. Dietary Compounds as Epigenetic Modulating Agents in Cancer. Front Genet 2019;10. [CrossRef]
- Abbas A, Hall JA, Patterson WL, Ho E, Hsu A, Al-Mulla F, et al. Sulforaphane modulates telomerase activity via epigenetic regulation in prostate cancer cell lines. Biochem Cell Biol 2016;94:71–81. [CrossRef]
- Abbas A, Patterson W, Georgel PT. The epigenetic potentials of dietary polyphenols in prostate cancer management. Biochem Cell Biol 2013;91:361–8. [CrossRef]
- Galati G, O’Brien PJ. Potential toxicity of flavonoids and other dietary phenolics: significance for their chemopreventive and anticancer properties. Free Radic Biol Med 2004;37:287–303. [CrossRef]
- Skibola CF, Smith MT. Potential health impacts of excessive flavonoid intake. Free Radic Biol Med 2000;29:375–83. [CrossRef]
- Singh BN, Shankar S, Srivastava RK. Green tea catechin, epigallocatechin-3-gallate (EGCG): Mechanisms, perspectives and clinical applications. Biochem Pharmacol 2011;82:1807–21. [CrossRef]
- Henning SM, Wang P, Carpenter CL, Heber D. Epigenetic effects of green tea polyphenols in cancer. Epigenomics 2013;5:729–41. [CrossRef]
- Giudice A, Montella M, Boccellino M, Crispo A, D’Arena G, Bimonte S, et al. Epigenetic Changes Induced by Green Tea Catechins a re Associated with Prostate Cancer. Curr Mol Med 2018;17. [CrossRef]
- Fang MZ, Wang Y, Ai N, Hou Z, Sun Y, Lu H, et al. Tea polyphenol (-)-epigallocatechin-3-gallate inhibits DNA methyltransferase and reactivates methylation-silenced genes in cancer cell lines. Cancer Res 2003;63:7563–70.
- Gupta S. Green tea polyphenols increase p53 transcriptional activity and acetylation by suppressing class�I histone deacetylases. Int J Oncol 2012. [CrossRef]
- Yoon H-G. EGCG suppresses prostate cancer cell growth modulating acetylation of androgen receptor by anti-histone acetyltransferase activity. Int J Mol Med 2012. [CrossRef]
- Lubecka K, Kaufman-Szymczyk A, Cebula-Obrzut B, Smolewski P, Szemraj J, Fabianowska-Majewska K. Novel Clofarabine-Based Combinations with Polyphenols Epigenetically Reactivate Retinoic Acid Receptor Beta, Inhibit Cell Growth, and Induce Apoptosis of Breast Cancer Cells. Int J Mol Sci 2018;19:3970. [CrossRef]
- KHAN MA, HUSSAIN A, SUNDARAM MK, ALALAMI U, GUNASEKERA D, RAMESH L, et al. (−)-Epigallocatechin-3-gallate reverses the expression of various tumor-suppressor genes by inhibiting DNA methyltransferases and histone deacetylases in human cervical cancer cells. Oncol Rep 2015;33:1976–84. [CrossRef]
- Ciesielski O, Biesiekierska M, Balcerczyk A. Epigallocatechin-3-gallate (EGCG) Alters Histone Acetylation and Methylation and Impacts Chromatin Architecture Profile in Human Endothelial Cells. Molecules 2020;25:2326. [CrossRef]
- Kedhari Sundaram M, Haque S, Somvanshi P, Bhardwaj T, Hussain A. Epigallocatechin gallate inhibits HeLa cells by modulation of epigenetics and signaling pathways. 3 Biotech 2020;10:484. [CrossRef]
- Kang Q, Zhang X, Cao N, Chen C, Yi J, Hao L, et al. EGCG enhances cancer cells sensitivity under 60Coγ radiation based on miR-34a/Sirt1/p53. Food Chem Toxicol 2019;133:110807. [CrossRef]
- Deb G, Shankar E, Thakur VS, Ponsky LE, Bodner DR, Fu P, et al. Green tea-induced epigenetic reactivation of tissue inhibitor of matrix metalloproteinase-3 suppresses prostate cancer progression through histone-modifying enzymes. Mol Carcinog 2019;58:1194–207. [CrossRef]
- Ahmed Youness R, Amr Assal R, Mohamed Ezzat S, Zakaria Gad M, Abdel Motaal A. A methoxylated quercetin glycoside harnesses HCC tumor progression in a TP53/miR-15/miR-16 dependent manner. Nat Prod Res 2020;34:1475–80. [CrossRef]
- Ahmed F, Ijaz B, Ahmad Z, Farooq N, Sarwar MB, Husnain T. Modification of miRNA Expression through plant extracts and compounds against breast cancer: Mechanism and translational significance. Phytomedicine 2020;68:153168. [CrossRef]
- Yang L, Zhang W, Chopra S, Kaur D, Wang H, Li M, et al. The Epigenetic Modification of Epigallocatechin Gallate (EGCG) on Cancer. Curr Drug Targets 2020;21:1099–104. [CrossRef]
- Singh S, Raza W, Parveen S, Meena A, Luqman S. Flavonoid display ability to target microRNAs in cancer pathogenesis. Biochem Pharmacol 2021;189:114409. [CrossRef]
- Zhao Y, Chen X, Jiang J, Wan X, Wang Y, Xu P. Epigallocatechin gallate reverses gastric cancer by regulating the long noncoding RNA LINC00511/miR-29b/KDM2A axis. Biochim Biophys Acta BBA - Mol Basis Dis 2020;1866:165856. [CrossRef]
- MOSTAFA SM, GAMAL-ELDEEN AM, MAKSOUD NA EL, FAHMI AA. Epigallocatechin gallate-capped gold nanoparticles enhanced the tumor suppressors let-7a and miR-34a in hepatocellular carcinoma cells. An Acad Bras Ciênc 2020;92. [CrossRef]
- Zheng N-G, Wang J-L, Yang S-L, Wu J-L. Aberrant Epigenetic Alteration in Eca9706 Cells Modulated by Nanoliposomal Quercetin Combined with Butyrate Mediated via Epigenetic-NF-κB Signaling. Asian Pac J Cancer Prev 2014;15:4539–43. [CrossRef]
- Kedhari Sundaram M, Hussain A, Haque S, Raina R, Afroze N. Quercetin modifies 5′CpG promoter methylation and reactivates various tumor suppressor genes by modulating epigenetic marks in human cervical cancer cells. J Cell Biochem 2019;120:18357–69. [CrossRef]
- Nwaeburu CC, Abukiwan A, Zhao Z, Herr I. Quercetin-induced miR-200b-3p regulates the mode of self-renewing divisions in pancreatic cancer. Mol Cancer 2017;16:23. [CrossRef]
- Pham TND, Stempel S, Shields MA, Spaulding C, Kumar K, Bentrem DJ, et al. Quercetin Enhances the Anti-Tumor Effects of BET Inhibitors by Suppressing hnRNPA1. Int J Mol Sci 2019;20:4293. [CrossRef]
- Zheng N-G, Wang J-L, Yang S-L, Wu J-L. Aberrant Epigenetic Alteration in Eca9706 Cells Modulated by Nanoliposomal Quercetin Combined with Butyrate Mediated via Epigenetic-NF-κB Signaling. Asian Pac J Cancer Prev 2014;15:4539–43. [CrossRef]
- Wang Y, Chen X, Li J, Xia C. Quercetin Antagonizes Esophagus Cancer by Modulating miR-1-3p/TAGLN2 Pathway-Dependent Growth and Metastasis. Nutr Cancer 2022;74:1872–81. [CrossRef]
- Russo GL, Ungaro P. Epigenetic Mechanisms of Quercetin and Other Flavonoids in Cancer Therapy and Prevention. Epigenetics Cancer Prev., Elsevier; 2019, p. 187–202. [CrossRef]
- Kedhari Sundaram M, Hussain A, Haque S, Raina R, Afroze N. Quercetin modifies 5′CpG promoter methylation and reactivates various tumor suppressor genes by modulating epigenetic marks in human cervical cancer cells. J Cell Biochem 2019;120:18357–69. [CrossRef]
- Pham TND, Stempel S, Shields MA, Spaulding C, Kumar K, Bentrem DJ, et al. Quercetin Enhances the Anti-Tumor Effects of BET Inhibitors by Suppressing hnRNPA1. Int J Mol Sci 2019;20:4293. [CrossRef]
- Hu S, Cheng J, Zhao W, Zhao H. Quercetin induces apoptosis in meningioma cells through the miR-197/IGFBP5 cascade. Environ Toxicol Pharmacol 2020;80:103439. [CrossRef]
- Wang Z, Chen H. Genistein increases gene expression by demethylation of WNT5a promoter in colon cancer cell line SW1116. Anticancer Res 2010;30:4537–45.
- Zhang C, Hao Y, Sun Y, Liu P. Quercetin suppresses the tumorigenesis of oral squamous cell carcinoma by regulating microRNA-22/WNT1/β-catenin axis. J Pharmacol Sci 2019;140:128–36. [CrossRef]
- Zhao Y, Chen X, Jiang J, Wan X, Wang Y, Xu P. Epigallocatechin gallate reverses gastric cancer by regulating the long noncoding RNA LINC00511/miR-29b/KDM2A axis. Biochim Biophys Acta BBA - Mol Basis Dis 2020;1866:165856. [CrossRef]
- Ramos YAL, Souza OF, Novo MCT, Guimarães CFC, Popi AF. Quercetin shortened survival of radio-resistant B-1 cells in vitro and in vivo by restoring miR15a/16 expression. Oncotarget 2021;12:355–65. [CrossRef]
- Imran M, Saeed F, Gilani SA, Shariati MA, Imran A, Afzaal M, et al. Fisetin: An anticancer perspective. Food Sci Nutr 2021;9:3–16. [CrossRef]
- Berger A, Venturelli S, Kallnischkies M, Böcker A, Busch C, Weiland T, et al. Kaempferol, a new nutrition-derived pan-inhibitor of human histone deacetylases. J Nutr Biochem 2013;24:977–85. [CrossRef]
- Kim TW, Lee SY, Kim M, Cheon C, Ko S-G. Kaempferol induces autophagic cell death via IRE1-JNK-CHOP pathway and inhibition of G9a in gastric cancer cells. Cell Death Dis 2018;9:875. [CrossRef]
- Han X, Liu C-F, Gao N, Zhao J, Xu J. RETRACTED: Kaempferol suppresses proliferation but increases apoptosis and autophagy by up-regulating microRNA-340 in human lung cancer cells. Biomed Pharmacother 2018;108:809–16. [CrossRef]
- Fernandez S, Risolino M, Mandia N, Talotta F, Soini Y, Incoronato M, et al. miR-340 inhibits tumor cell proliferation and induces apoptosis by targeting multiple negative regulators of p27 in non-small cell lung cancer. Oncogene 2015;34:3240–50. [CrossRef]
- Imran M, Saeed F, Gilani SA, Shariati MA, Imran A, Afzaal M, et al. Fisetin: An anticancer perspective. Food Sci Nutr 2021;9:3–16. [CrossRef]
- Pal HC, Pearlman RL, Afaq F. Fisetin and Its Role in Chronic Diseases, 2016, p. 213–44. [CrossRef]
- Ding G, Xu X, Li D, Chen Y, Wang W, Ping D, et al. Fisetin inhibits proliferation of pancreatic adenocarcinoma by inducing DNA damage via RFXAP/KDM4A-dependent histone H3K36 demethylation. Cell Death Dis 2020;11:893. [CrossRef]
- Pandey M, Shukla S, Gupta S. Promoter demethylation and chromatin remodeling by green tea polyphenols leads to re-expression of GSTP1 in human prostate cancer cells. Int J Cancer 2010:NA-NA. [CrossRef]
- Gao A-M, Zhang X-Y, Hu J-N, Ke Z-P. Apigenin sensitizes hepatocellular carcinoma cells to doxorubic through regulating miR-520b/ATG7 axis. Chem Biol Interact 2018;280:45–50. [CrossRef]
- Tseng T-H, Chien M-H, Lin W-L, Wen Y-C, Chow J-M, Chen C-K, et al. Inhibition of MDA-MB-231 breast cancer cell proliferation and tumor growth by apigenin through induction of G2/M arrest and histone H3 acetylation-mediated p21 WAF1/CIP1 expression. Environ Toxicol 2017;32:434–44. [CrossRef]
- Chen X-J, Wu M-Y, Li D-H, You J. Apigenin inhibits glioma cell growth through promoting microRNA-16 and suppression of BCL-2 and nuclear factor-κB/MMP-9. Mol Med Rep 2016;14:2352–8. [CrossRef]
- Cheng Y, Han X, Mo F, Zeng H, Zhao Y, Wang H, et al. Apigenin inhibits the growth of colorectal cancer through down-regulation of E2F1/3 by miRNA-215-5p. Phytomedicine 2021;89:153603. [CrossRef]
- Wu H-T, Lin J, Liu Y-E, Chen H-F, Hsu K-W, Lin S-H, et al. Luteolin suppresses androgen receptor-positive triple-negative breast cancer cell proliferation and metastasis by epigenetic regulation of MMP9 expression via the AKT/mTOR signaling pathway. Phytomedicine 2021;81:153437. [CrossRef]
- Lin D, Kuang G, Wan J, Zhang X, Li H, Gong X, et al. Luteolin suppresses the metastasis of triple-negative breast cancer by reversing epithelial-to-mesenchymal transition via downregulation of β-catenin expression. Oncol Rep 2017;37:895–902. [CrossRef]
- Krifa M, Leloup L, Ghedira K, Mousli M, Chekir-Ghedira L. Luteolin Induces Apoptosis in BE Colorectal Cancer Cells by Downregulating Calpain, UHRF1, and DNMT1 Expressions. Nutr Cancer 2014;66:1220–7. [CrossRef]
- Markaverich BM, Vijjeswarapu M, Shoulars K, Rodriguez M. Luteolin and gefitinib regulation of EGF signaling pathway and cell cycle pathway genes in PC-3 human prostate cancer cells. J Steroid Biochem Mol Biol 2010;122:219–31. [CrossRef]
- Shoulars K, Rodriguez MA, Thompson T, Markaverich BM. Regulation of cell cycle and RNA transcription genes identified by microarray analysis of PC-3 human prostate cancer cells treated with luteolin. J Steroid Biochem Mol Biol 2010;118:41–50. [CrossRef]
- Farooqi AA, Butt G, El-Zahaby SA, Attar R, Sabitaliyevich UY, Jovic JJ, et al. Luteolin mediated targeting of protein network and microRNAs in different cancers: Focus on JAK-STAT, NOTCH, mTOR and TRAIL-mediated signaling pathways. Pharmacol Res 2020;160:105188. [CrossRef]
- Mohr A, Mott J. Overview of MicroRNA Biology. Semin Liver Dis 2015;35:003–11. [CrossRef]
- Lai W, Jia J, Yan B, Jiang Y, Shi Y, Chen L, et al. Baicalin hydrate inhibits cancer progression in nasopharyngeal carcinoma by affecting genome instability and splicing. Oncotarget 2018;9:901–14. [CrossRef]
- Bijak M. Silybin, a Major Bioactive Component of Milk Thistle (Silybum marianum L. Gaernt.)—Chemistry, Bioavailability, and Metabolism. Mol J Synth Chem Nat Prod Chem 2017;22:1942. [CrossRef]
- Barreca D, Mandalari G, Calderaro A, Smeriglio A, Trombetta D, Felice MR, et al. Citrus Flavones: An Update on Sources, Biological Functions, and Health Promoting Properties. Plants 2020;9:288. [CrossRef]
- Khan H, Belwal T, Efferth T, Farooqi AA, Sanches-Silva A, Vacca RA, et al. Targeting epigenetics in cancer: therapeutic potential of flavonoids. Crit Rev Food Sci Nutr 2021;61:1616–39. [CrossRef]
- Wang S, Sheng H, Zheng F, Zhang F. Hesperetin promotes DOT1L degradation and reduces histone H3K79 methylation to inhibit gastric cancer metastasis. Phytomedicine 2021;84:153499. [CrossRef]
- Mateen S, Raina K, Agarwal C, Chan D, Agarwal R. Silibinin Synergizes with Histone Deacetylase and DNA Methyltransferase Inhibitors in Upregulating E-cadherin Expression Together with Inhibition of Migration and Invasion of Human Non-small Cell Lung Cancer Cells. J Pharmacol Exp Ther 2013;345:206–14. [CrossRef]
- Anestopoulos I, Sfakianos A, Franco R, Chlichlia K, Panayiotidis M, Kroll D, et al. A Novel Role of Silibinin as a Putative Epigenetic Modulator in Human Prostate Carcinoma. Molecules 2016;22:62. [CrossRef]
- Hossainzadeh S, Ranji N, Naderi Sohi A, Najafi F. Silibinin encapsulation in polymersome: A promising anticancer nanoparticle for inducing apoptosis and decreasing the expression level of miR-125b/miR-182 in human breast cancer cells. J Cell Physiol 2019;234:22285–98. [CrossRef]
- Ling D, Marshall GM, Liu PY, Xu N, Nelson CA, Iismaa SE, et al. Enhancing the anticancer effect of the histone deacetylase inhibitor by activating transglutaminase. Eur J Cancer 2012;48:3278–87. [CrossRef]
- Panche AN, Diwan AD, Chandra SR. Flavonoids: an overview. J Nutr Sci 2016;5:e47. [CrossRef]
- Taku K, Melby MK, Nishi N, Omori T, Kurzer MS. Soy isoflavones for osteoporosis: An evidence-based approach. Maturitas 2011;70:333–8. [CrossRef]
- Vitale DC, Piazza C, Melilli B, Drago F, Salomone S. Isoflavones: estrogenic activity, biological effect and bioavailability. Eur J Drug Metab Pharmacokinet 2013;38:15–25. [CrossRef]
- Qin Y, Niu K, Zeng Y, Liu P, Yi L, Zhang T, et al. Isoflavones for hypercholesterolaemia in adults. Cochrane Database Syst Rev 2013. [CrossRef]
- Sathyapalan T, Aye M, Rigby AS, Thatcher NJ, Dargham SR, Kilpatrick ES, et al. Soy isoflavones improve cardiovascular disease risk markers in women during the early menopause. Nutr Metab Cardiovasc Dis 2018;28:691–7. [CrossRef]
- Křížová L, Dadáková K, Kašparovská J, Kašparovský T. Isoflavones. Molecules 2019;24:1076. [CrossRef]
- Gómez-Zorita S, González-Arceo M, Fernández-Quintela A, Eseberri I, Trepiana J, Portillo MP. Scientific Evidence Supporting the Beneficial Effects of Isoflavones on Human Health. Nutrients 2020;12:3853. [CrossRef]
- Spagnuolo C, Russo GL, Orhan IE, Habtemariam S, Daglia M, Sureda A, et al. Genistein and Cancer: Current Status, Challenges, and Future Directions. Adv Nutr 2015;6:408–19. [CrossRef]
- Adjakly M, Bosviel R, Rabiau N, Boiteux J-P, Bignon Y-J, Guy L, et al. DNA methylation and soy phytoestrogens: quantitative study in DU-145 and PC-3 human prostate cancer cell lines. Epigenomics 2011;3:795–803. [CrossRef]
- Karsli-Ceppioglu S, Ngollo M, Adjakly M, Dagdemir A, Judes G, Lebert A, et al. Genome-Wide DNA Methylation Modified by Soy Phytoestrogens: Role for Epigenetic Therapeutics in Prostate Cancer? OMICS J Integr Biol 2015;19:209–19. [CrossRef]
- Tanaka MI-SPDPKMSYHVSSMY, Yamamura RDS. Genistein Represses HOTAIR/Chromatin Remodeling Pathways to Suppress Kidney Cancer. Cell Physiol Biochem 2020;54:53–70. [CrossRef]
- Allred CD. Dietary genistin stimulates growth of estrogen-dependent breast cancer tumors similar to that observed with genistein. Carcinogenesis 2001;22:1667–73. [CrossRef]
- Khoo HE, Azlan A, Tang ST, Lim SM. Anthocyanidins and anthocyanins: colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr Res 2017;61:1361779. [CrossRef]
- Kuo H-CD, Wu R, Li S, Yang AY, Kong A-N. Anthocyanin Delphinidin Prevents Neoplastic Transformation of Mouse Skin JB6 P+ Cells: Epigenetic Re-activation of Nrf2-ARE Pathway. AAPS J 2019;21:83. [CrossRef]
- Jeong M-H, Ko H, Jeon H, Sung G-J, Park S-Y, Jun WJ, et al. Delphinidin induces apoptosis via cleaved HDAC3-mediated p53 acetylation and oligomerization in prostate cancer cells. Oncotarget 2016;7:56767–80. [CrossRef]
- Han B, Peng X, Cheng D, Zhu Y, Du J, Li J, et al. Delphinidin suppresses breast carcinogenesis through the HOTAIR /micro RNA -34a axis. Cancer Sci 2019;110:3089–97. [CrossRef]
- Huang C-C, Hung C-H, Hung T-W, Lin Y-C, Wang C-J, Kao S-H. Dietary delphinidin inhibits human colorectal cancer metastasis associating with upregulation of miR-204-3p and suppression of the integrin/FAK axis. Sci Rep 2019;9:18954. [CrossRef]
- Kopustinskiene DM, Jakstas V, Savickas A, Bernatoniene J. Flavonoids as Anticancer Agents. Nutrients 2020;12. [CrossRef]
- Chae H-S, Xu R, Won J-Y, Chin Y-W, Yim H. Molecular Targets of Genistein and Its Related Flavonoids to Exert Anticancer Effects. Int J Mol Sci 2019;20:2420. [CrossRef]
- BANERJEE S, LI Y, WANG Z, SARKAR FH. MULTI-TARGETED THERAPY OF CANCER BY GENISTEIN. Cancer Lett 2008;269:226–42. [CrossRef]
- Sanaei M, Kavoosi F, Atashpour S, Haghighat S. Effects of Genistein and Synergistic Action in Combination with Tamoxifen on the HepG2 Human Hepatocellular Carcinoma Cell Line. Asian Pac J Cancer Prev APJCP 2017;18:2381–5. [CrossRef]
- Konstantinou EK, Gioxari A, Dimitriou M, Panoutsopoulos GI, Panagiotopoulos AA. Molecular Pathways of Genistein Activity in Breast Cancer Cells. Int J Mol Sci 2024;25:5556. [CrossRef]
- Donald G, Hertzer K, Eibl G. Baicalein – An Intriguing Therapeutic Phytochemical in Pancreatic Cancer. Curr Drug Targets 2012;13:1772–6.
- Lai W, Jia J, Yan B, Jiang Y, Shi Y, Chen L, et al. Baicalin hydrate inhibits cancer progression in nasopharyngeal carcinoma by affecting genome instability and splicing. Oncotarget 2018;9:901–14. [CrossRef]
- Yan X, Qi M, Li P, Zhan Y, Shao H. Apigenin in cancer therapy: anti-cancer effects and mechanisms of action. Cell Biosci 2017;7:50. [CrossRef]
- Xu Y, Xin Y, Diao Y, Lu C, Fu J, Luo L, et al. Synergistic Effects of Apigenin and Paclitaxel on Apoptosis of Cancer Cells. PLoS ONE 2011;6:e29169. [CrossRef]
- Tseng T, Chien M, Lin W, Wen Y, Chow J, Chen C, et al. Inhibition of MDA-MB-231 breast cancer cell proliferation and tumor growth by apigenin through induction of G2/M arrest and histone H3 acetylation-mediated p21 WAF1/CIP1 expression. Environ Toxicol 2017;32:434–44. [CrossRef]
- Seelinger G, Merfort I, Wölfle U, Schempp CM. Anti-carcinogenic Effects of the Flavonoid Luteolin. Molecules 2008;13:2628–51. [CrossRef]
- Jang CH, Moon N, Lee J, Kwon MJ, Oh J, Kim J-S. Luteolin Synergistically Enhances Antitumor Activity of Oxaliplatin in Colorectal Carcinoma via AMPK Inhibition. Antioxidants 2022;11:626. [CrossRef]
- Zuo Q, Wu R, Xiao X, Yang C, Yang Y, Wang C, et al. The dietary flavone luteolin epigenetically activates the Nrf2 pathway and blocks cell transformation in human colorectal cancer HCT116 cells. J Cell Biochem 2018;119:9573–82. [CrossRef]
- Yoshimizu N, Otani Y, Saikawa Y, Kubota T, Yoshida M, Furukawa T, et al. Anti-tumour effects of nobiletin, a citrus flavonoid, on gastric cancer include: antiproliferative effects, induction of apoptosis and cell cycle deregulation. Aliment Pharmacol Ther 2004;20 Suppl 1:95–101. [CrossRef]
- Ohnishi H, Asamoto M, Tujimura K, Hokaiwado N, Takahashi S, Ogawa K, et al. Inhibition of cell proliferation by nobiletin, a dietary phytochemical, associated with apoptosis and characteristic gene expression, but lack of effect on early rat hepatocarcinogenesis in vivo. Cancer Sci 2004;95:936–42. [CrossRef]
- Morley KL, Ferguson PJ, Koropatnick J. Tangeretin and nobiletin induce G1 cell cycle arrest but not apoptosis in human breast and colon cancer cells. Cancer Lett 2007;251:168–78. [CrossRef]
- Wu X, Song M, Qiu P, Rakariyatham K, Li F, Gao Z, et al. Synergistic chemopreventive effects of nobiletin and atorvastatin on colon carcinogenesis. Carcinogenesis 2017;38:455–64. [CrossRef]
- Wang J, Jian W, Li Y, Zhang J. Nobiletin promotes the pyroptosis of breast cancer via regulation of miR -200b/ JAZF1 axis. Kaohsiung J Med Sci 2021;37:572–82. [CrossRef]
- Mirazimi SMA, Dashti F, Tobeiha M, Shahini A, Jafari R, Khoddami M, et al. Application of Quercetin in the Treatment of Gastrointestinal Cancers. Front Pharmacol 2022;13.
- Kundur S, Prayag A, Selvakumar P, Nguyen H, McKee L, Cruz C, et al. Synergistic anticancer action of quercetin and curcumin against triple-negative breast cancer cell lines. J Cell Physiol 2019;234:11103–18. [CrossRef]
- Wang P, Phan T, Gordon D, Chung S, Henning SM, Vadgama JV. Arctigenin in combination with quercetin synergistically enhances the antiproliferative effect in prostate cancer cells. Mol Nutr Food Res 2015;59:250–61. [CrossRef]
- Ponte LGS, Pavan ICB, Mancini MCS, da Silva LGS, Morelli AP, Severino MB, et al. The Hallmarks of Flavonoids in Cancer. Molecules 2021;26:2029. [CrossRef]
- Qattan MY, Khan MI, Alharbi SH, Verma AK, Al-Saeed FA, Abduallah AM, et al. Therapeutic Importance of Kaempferol in the Treatment of Cancer through the Modulation of Cell Signalling Pathways. Molecules 2022;27:8864. [CrossRef]
- Kim TW, Lee SY, Kim M, Cheon C, Ko S-G. Kaempferol induces autophagic cell death via IRE1-JNK-CHOP pathway and inhibition of G9a in gastric cancer cells. Cell Death Dis 2018;9:875. [CrossRef]
- Javed Z, Khan K, Herrera-Bravo J, Naeem S, Iqbal MJ, Raza Q, et al. Myricetin: targeting signaling networks in cancer and its implication in chemotherapy. Cancer Cell Int 2022;22:239. [CrossRef]
- Phillips PA, Sangwan V, Borja-Cacho D, Dudeja V, Vickers SM, Saluja AK. Myricetin induces pancreatic cancer cell death via the induction of apoptosis and inhibition of the phosphatidylinositol 3-kinase (PI3K) signaling pathway. Cancer Lett 2011;308:181–8. [CrossRef]
- Aggarwal V, Tuli HS, Thakral F, Singhal P, Aggarwal D, Srivastava S, et al. Molecular mechanisms of action of hesperidin in cancer: Recent trends and advancements. Exp Biol Med 2020;245:486–97. [CrossRef]
- Shahbazi R, Cheraghpour M, Homayounfar R, Nazari M, Nasrollahzadeh J, Davoodi SH. Hesperidin inhibits insulin-induced phosphoinositide 3–kinase/Akt activation in human pre-B cell line NALM-6. J Cancer Res Ther 2018;14:503–8. [CrossRef]
- Sohel M, Sultana H, Sultana T, Al Amin Md, Aktar S, Ali MdC, et al. Chemotherapeutic potential of hesperetin for cancer treatment, with mechanistic insights: A comprehensive review. Heliyon 2022;8:e08815. [CrossRef]
- Sajeev A, Hegde M, Girisa S, Devanarayanan TN, Alqahtani MS, Abbas M, et al. Oroxylin A: A Promising Flavonoid for Prevention and Treatment of Chronic Diseases. Biomolecules 2022;12:1185. [CrossRef]
- Huang H, Cai H, Zhang L, Hua Z, Shi J, Wei Y. Oroxylin A inhibits carcinogen-induced skin tumorigenesis through inhibition of inflammation by regulating SHCBP1 in mice. Int Immunopharmacol 2020;80:106123. [CrossRef]
- Tai MC, Tsang SY, Chang LYF, Xue H. Therapeutic Potential of Wogonin: A Naturally Occurring Flavonoid. CNS Drug Rev 2005;11:141–50. [CrossRef]
- Yang D, Guo Q, Liang Y, Zhao Y, Tian X, Ye Y, et al. Wogonin induces cellular senescence in breast cancer via suppressing TXNRD2 expression. Arch Toxicol 2020;94:3433–47. [CrossRef]
- Im N-K, Jang WJ, Jeong C-H, Jeong G-S. Delphinidin Suppresses PMA-Induced MMP-9 Expression by Blocking the NF-κB Activation Through MAPK Signaling Pathways in MCF-7 Human Breast Carcinoma Cells. J Med Food 2014;17:855–61. [CrossRef]
- Sharma A, Choi H-K, Kim Y-K, Lee H-J. Delphinidin and Its Glycosides’ War on Cancer: Preclinical Perspectives. Int J Mol Sci 2021;22:11500. [CrossRef]
- Deep G, Agarwal R. Anti-metastatic Efficacy of Silibinin: Molecular Mechanisms and Therapeutic Potential against Cancer. Cancer Metastasis Rev 2010;29:447–63. [CrossRef]
- Du G-J, Zhang Z, Wen X-D, Yu C, Calway T, Yuan C-S, et al. Epigallocatechin Gallate (EGCG) Is the Most Effective Cancer Chemopreventive Polyphenol in Green Tea. Nutrients 2012;4:1679–91. [CrossRef]
- Eom D-W, Lee JH, Kim Y-J, Hwang GS, Kim S-N, Kwak JH, et al. Synergistic effect of curcumin on epigallocatechin gallate-induced anticancer action in PC3 prostate cancer cells. BMB Rep 2015;48:461–6. [CrossRef]
- Henning SM, Wang P, Carpenter CL, Heber D. Epigenetic Effects of Green Tea Polyphenols in Cancer. Epigenomics 2013;5:729–41. [CrossRef]
- Maasomi ZJ, Soltanahmadi YP, Dadashpour M, Alipour S, Abolhasani S, Zarghami N. Synergistic Anticancer Effects of Silibinin and Chrysin in T47D Breast Cancer Cells. Asian Pac J Cancer Prev APJCP 2017;18:1283–7. [CrossRef]
- Pal-Bhadra M, Ramaiah MJ, Reddy TL, Krishnan A, Pushpavalli S, Babu KS, et al. Plant HDAC inhibitor chrysin arrest cell growth and induce p21 WAF1 by altering chromatin of STAT response element in A375 cells. BMC Cancer 2012;12:180. [CrossRef]
- Ko Y-C, Choi HS, Liu R, Kim J-H, Kim S-L, Yun B-S, et al. Inhibitory Effects of Tangeretin, a Citrus Peel-Derived Flavonoid, on Breast Cancer Stem Cell Formation through Suppression of Stat3 Signaling. Molecules 2020;25:2599. [CrossRef]
- Wei G-J, Chao Y-H, Tung Y-C, Wu T-Y, Su Z-Y. A Tangeretin Derivative Inhibits the Growth of Human Prostate Cancer LNCaP Cells by Epigenetically Restoring p21 Gene Expression and Inhibiting Cancer Stem-like Cell Proliferation. AAPS J 2019;21:86. [CrossRef]
- Porras G, Bacsa J, Tang H, Quave CL. Characterization and Structural Analysis of Genkwanin, a Natural Product from Callicarpa americana. Crystals 2019;9:491. [CrossRef]
- Wei C, Lu J, Zhang Z, Hua F, Chen Y, Shen Z. Genkwanin attenuates lung cancer development by repressing proliferation and invasion via phosphatidylinositol 3-kinase/protein kinase B pathway. Mater Express 2021;11:319–25.
- Curti V, Di Lorenzo A, Rossi D, Martino E, Capelli E, Collina S, et al. Enantioselective Modulatory Effects of Naringenin Enantiomers on the Expression Levels of miR-17-3p Involved in Endogenous Antioxidant Defenses. Nutrients 2017;9:215. [CrossRef]
- Effat H, Abosharaf HA, Radwan AM. Combined effects of naringin and doxorubicin on the JAK/STAT signaling pathway reduce the development and spread of breast cancer cells. Sci Rep 2024;14:2824. [CrossRef]
Figure 1.
Summary of cancer epigenetics involving promoter hypermethylation, genome-wide hypomethylation resulted into the abnormal methylation which induces abnormal chromatin structure due to loss of function mutation of switch/sucrose nonfermenting (SWI/SNF).
Figure 1.
Summary of cancer epigenetics involving promoter hypermethylation, genome-wide hypomethylation resulted into the abnormal methylation which induces abnormal chromatin structure due to loss of function mutation of switch/sucrose nonfermenting (SWI/SNF).
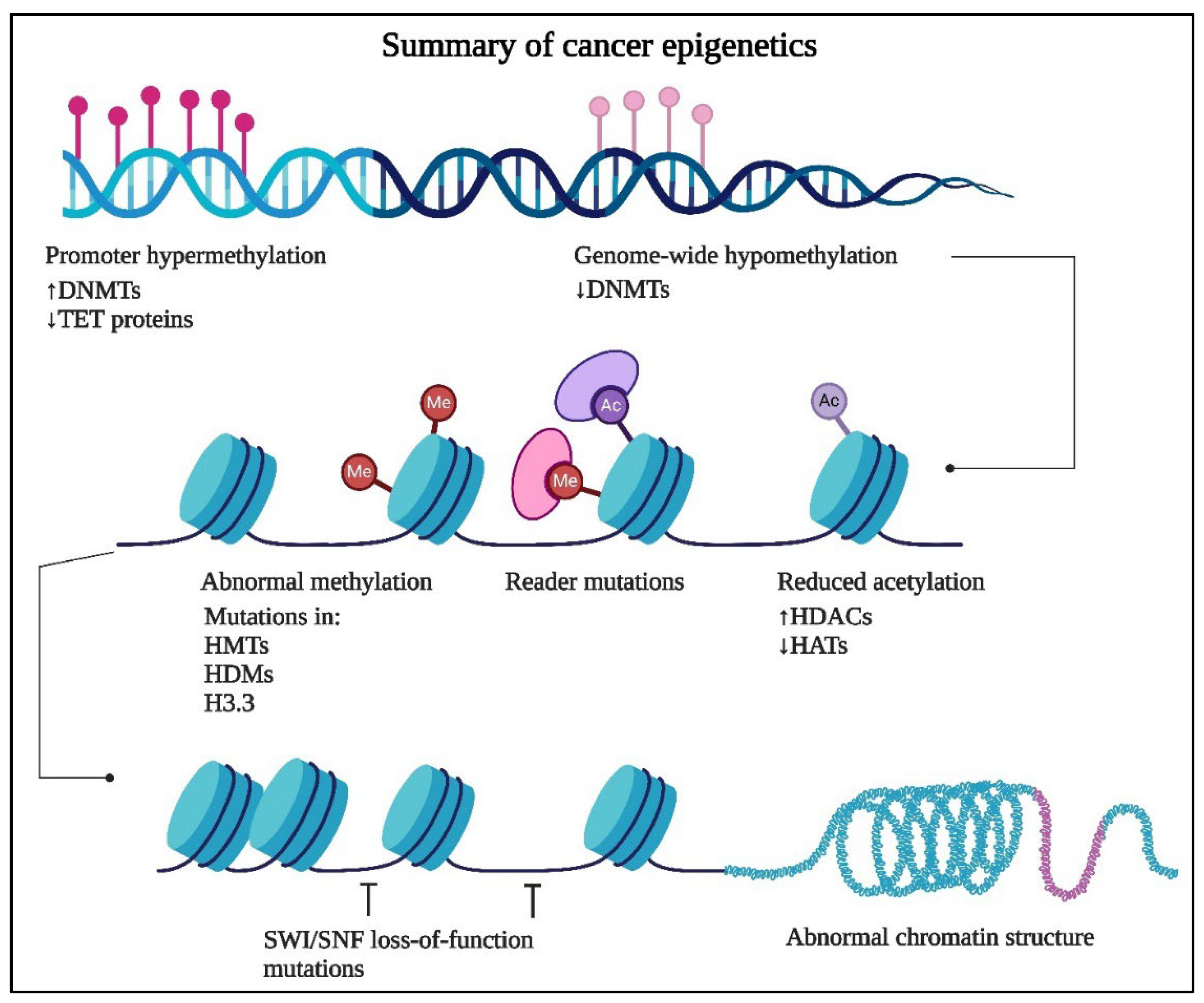
Figure 2.
Summary of statistics for epigenetic modulators from the year 2000 to 2024 including number of review articles and books published, number of clinical trials and meta-analysis conducted and gender vise available data.
Figure 2.
Summary of statistics for epigenetic modulators from the year 2000 to 2024 including number of review articles and books published, number of clinical trials and meta-analysis conducted and gender vise available data.
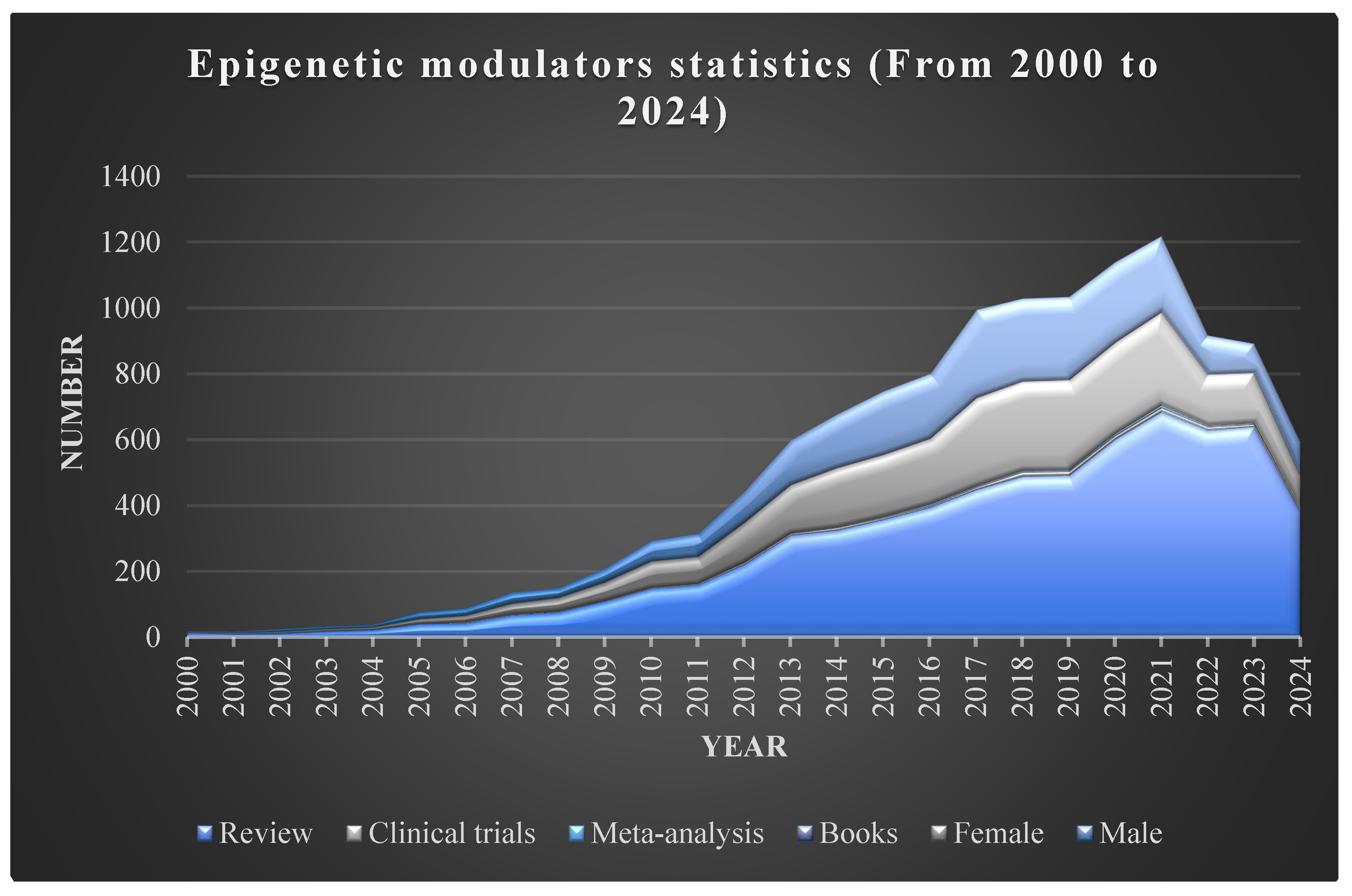
Figure 3.
Epigenetic variation of genomic DNA. Genes from cells undergo hereditary variations in gene transcription with no alteration in genomic DNA called epigenetics. These epigenetic modifications provide no variations in the DNA base code and can be present in the early phase of cell diversification. Such events indicate permanently inherited successive dividing cells, resulting in diverse cellular characteristics. The dysregulation induces an altered expression pattern of the genes, which results in epigenetic aberrations. These aberrations generate cancer, which is treated with conventional chemotherapy and advanced drug treatment containing histone deacetylase antagonists, epigenetic stimulators, and CAR-T cells. Such therapy contributes more side effects, and hence, these are addressed by flavonoid formulations for better protection against cancer cells. (Created using Biorender.com).
Figure 3.
Epigenetic variation of genomic DNA. Genes from cells undergo hereditary variations in gene transcription with no alteration in genomic DNA called epigenetics. These epigenetic modifications provide no variations in the DNA base code and can be present in the early phase of cell diversification. Such events indicate permanently inherited successive dividing cells, resulting in diverse cellular characteristics. The dysregulation induces an altered expression pattern of the genes, which results in epigenetic aberrations. These aberrations generate cancer, which is treated with conventional chemotherapy and advanced drug treatment containing histone deacetylase antagonists, epigenetic stimulators, and CAR-T cells. Such therapy contributes more side effects, and hence, these are addressed by flavonoid formulations for better protection against cancer cells. (Created using Biorender.com).
Figure 4.
The summary of epigenetic deregulation in cancer including chromatin remodeling, DNA methylation and non-coding RNA resulted into alteration in transcription as well as translation of the oncogenic gene targets.
Figure 4.
The summary of epigenetic deregulation in cancer including chromatin remodeling, DNA methylation and non-coding RNA resulted into alteration in transcription as well as translation of the oncogenic gene targets.
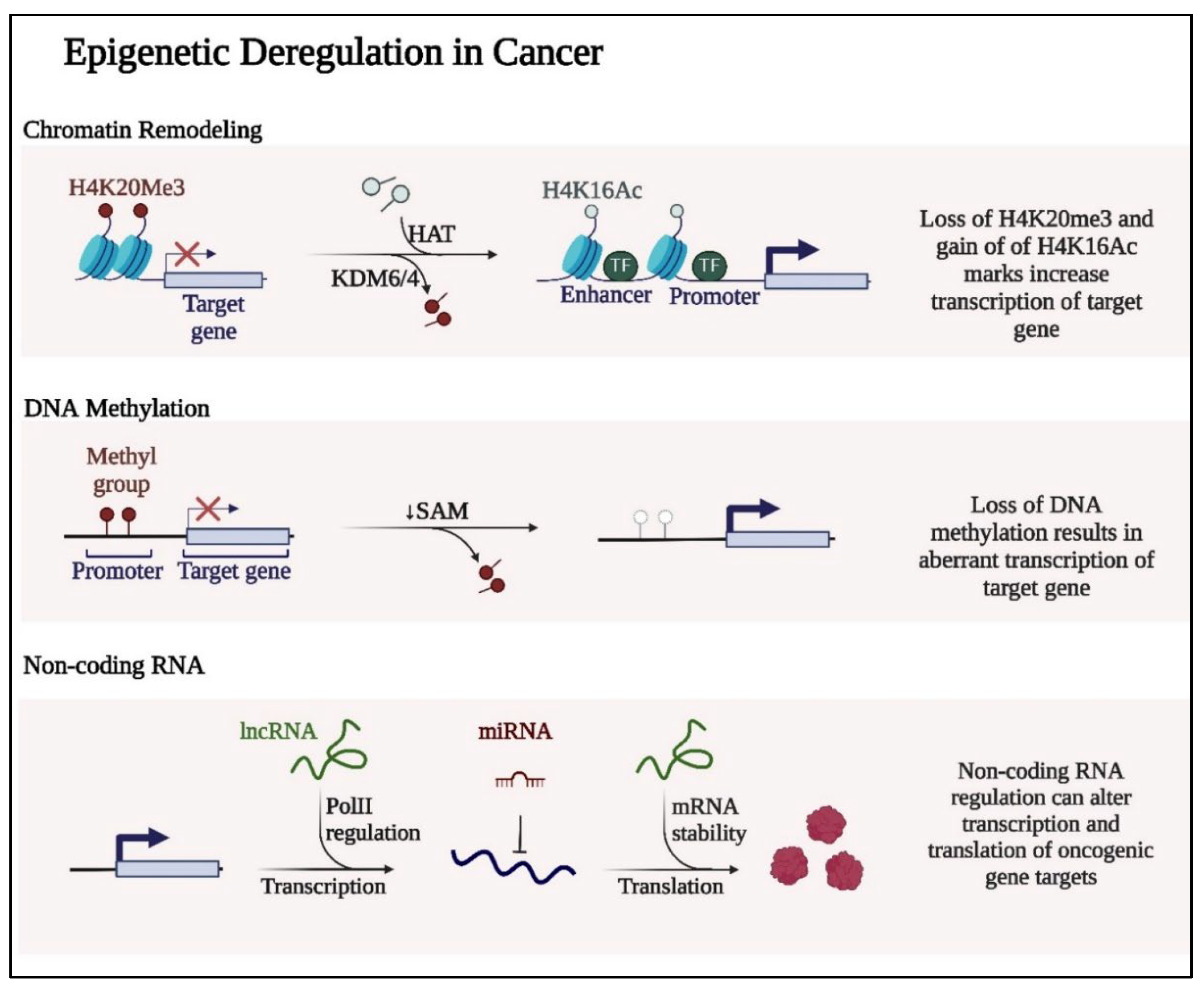
Figure 5.
The DNA methylation in closed chromatin and histone modification involving acetylation, ubiquitination, and phosphorylation.
Figure 5.
The DNA methylation in closed chromatin and histone modification involving acetylation, ubiquitination, and phosphorylation.
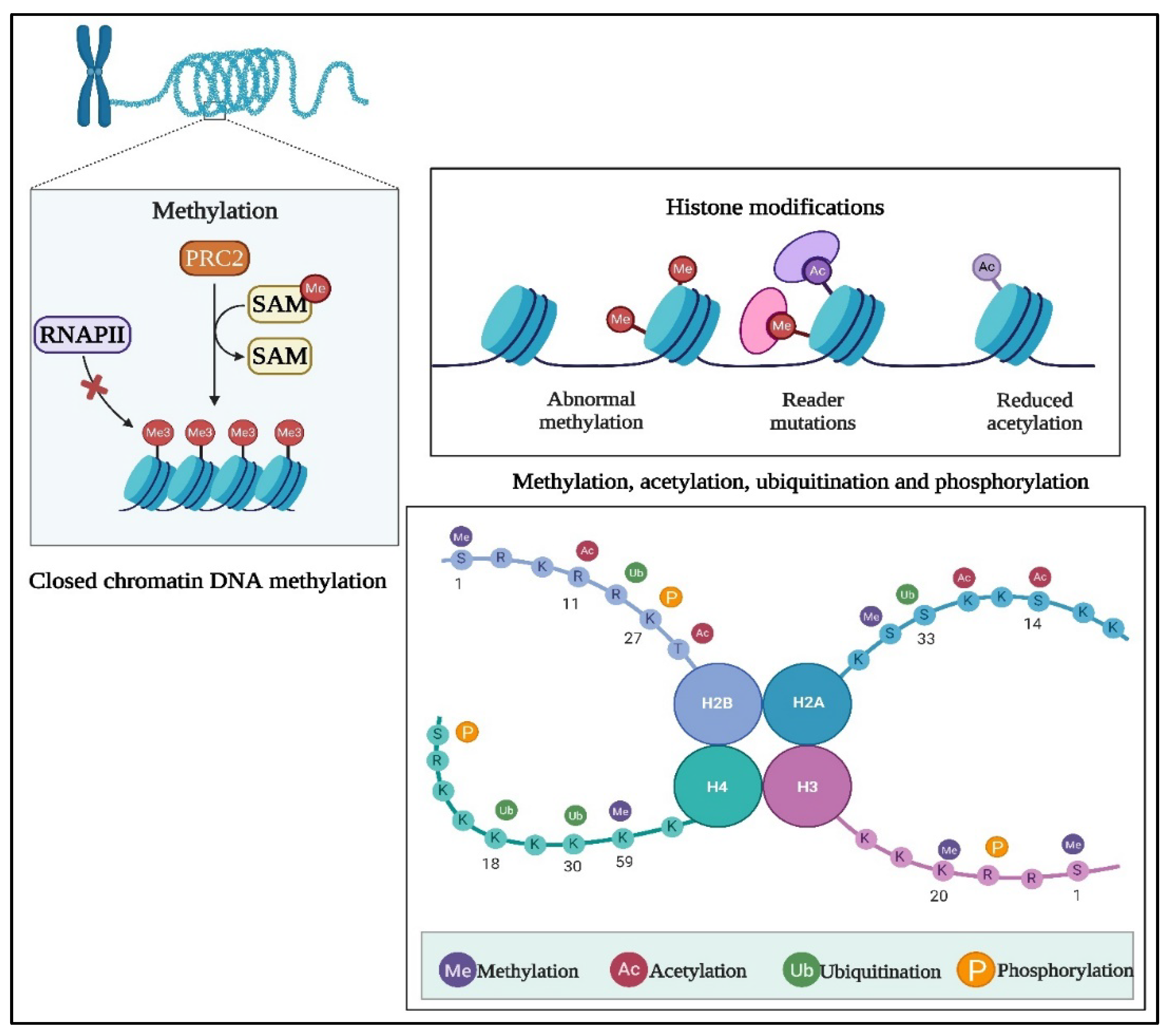
Figure 6.
Chemical Structure of Flavonoids. Created using Chemsketch.com.
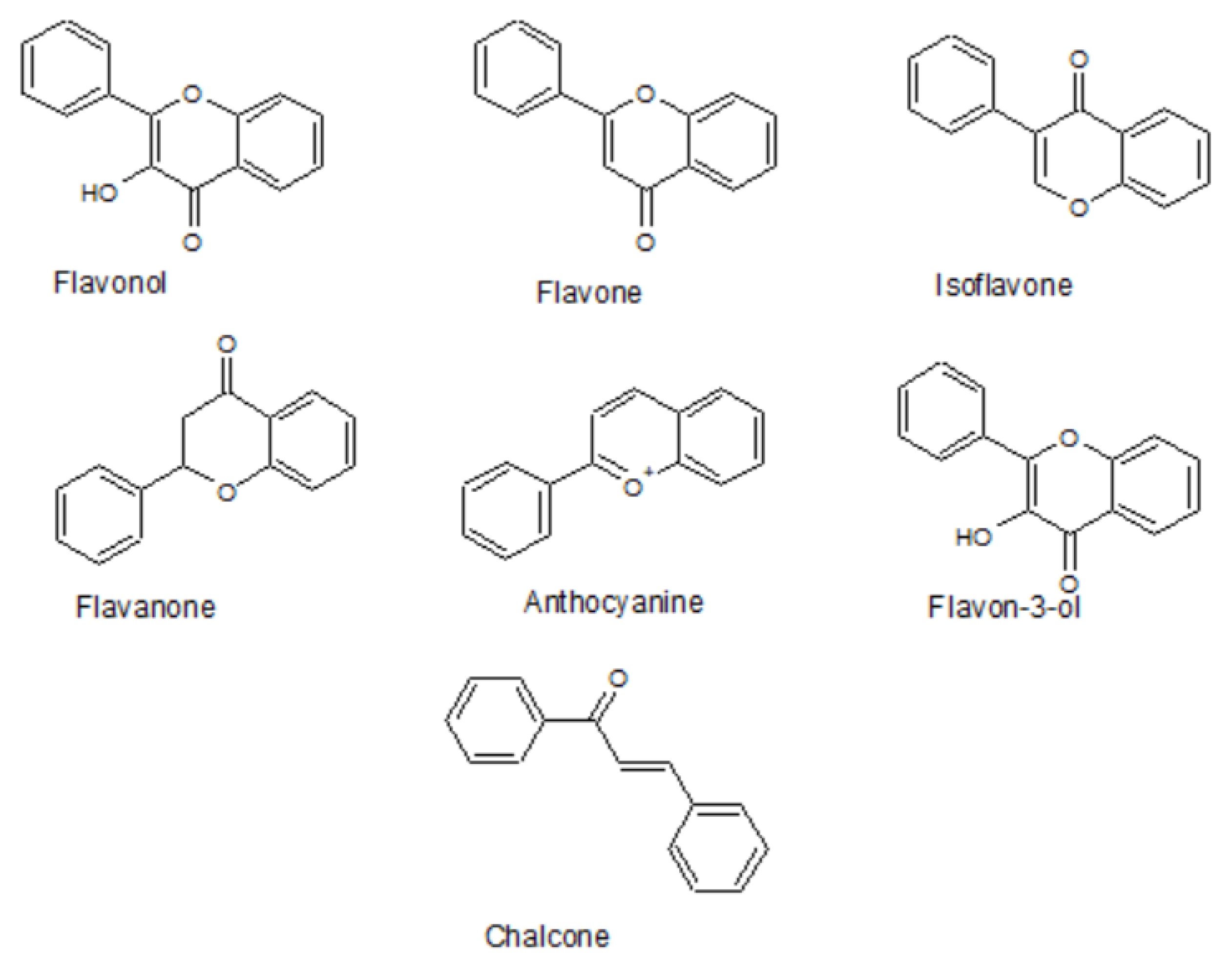

Table 1.
Flavonoids in cancer management with their molecular targets.
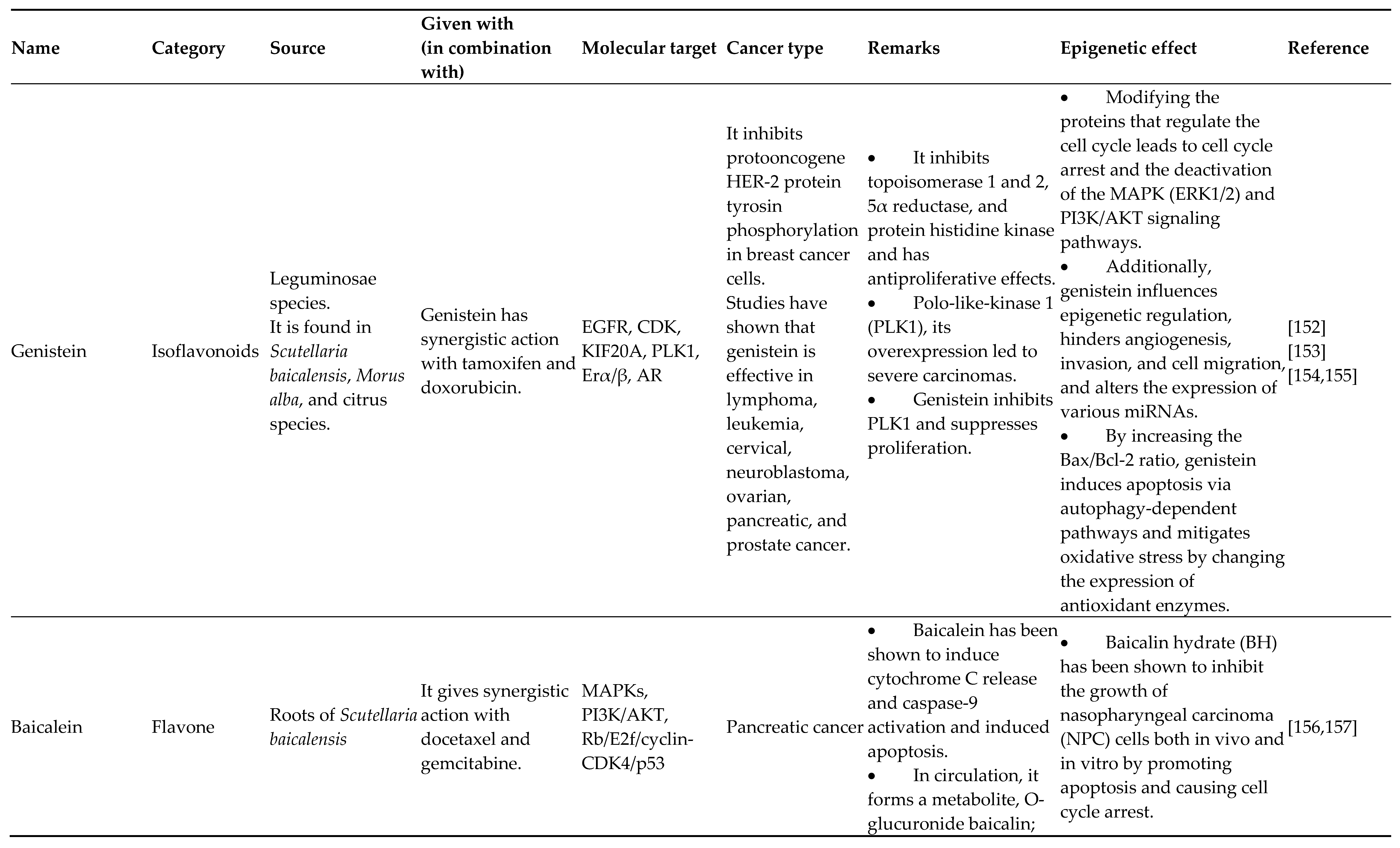
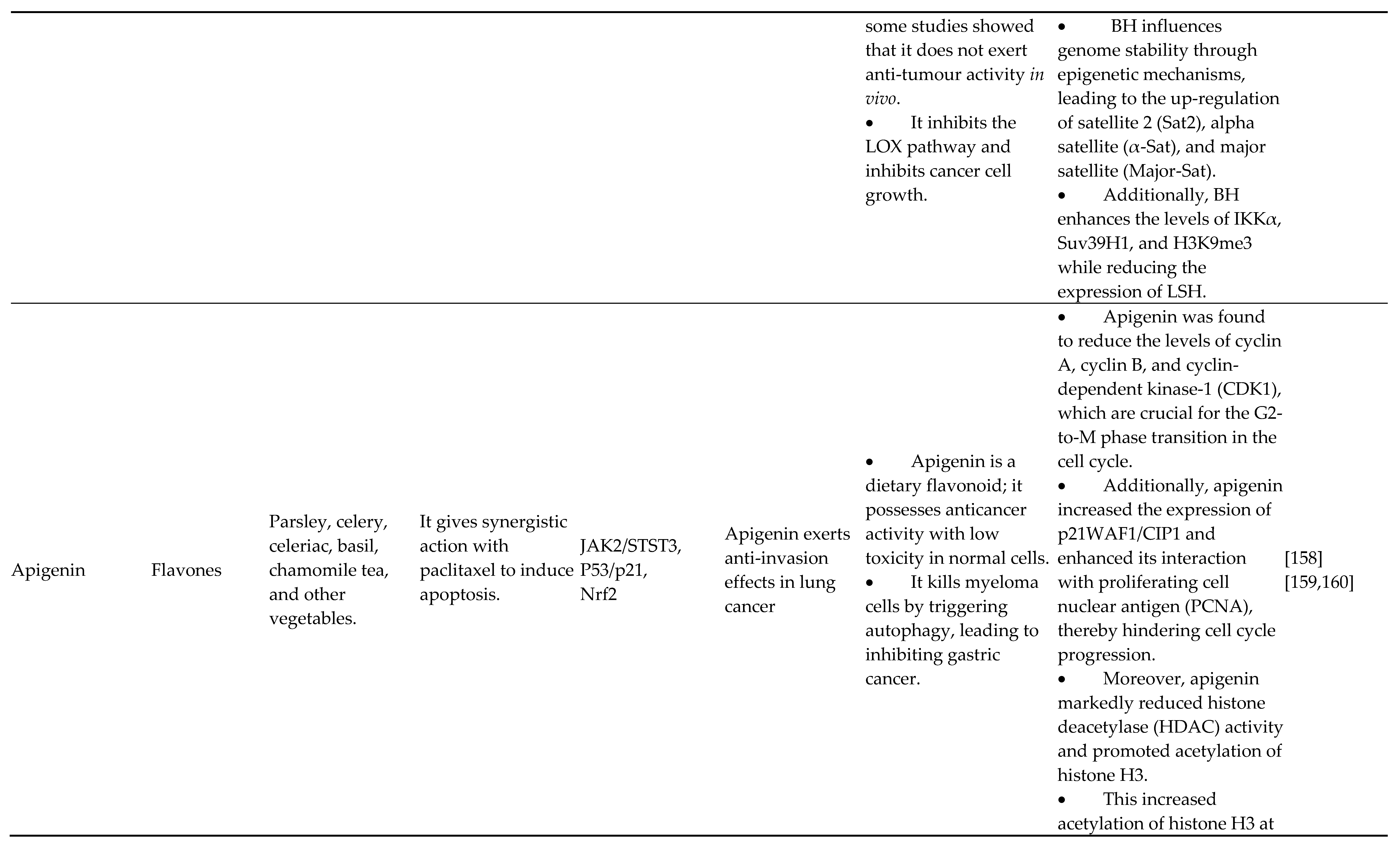
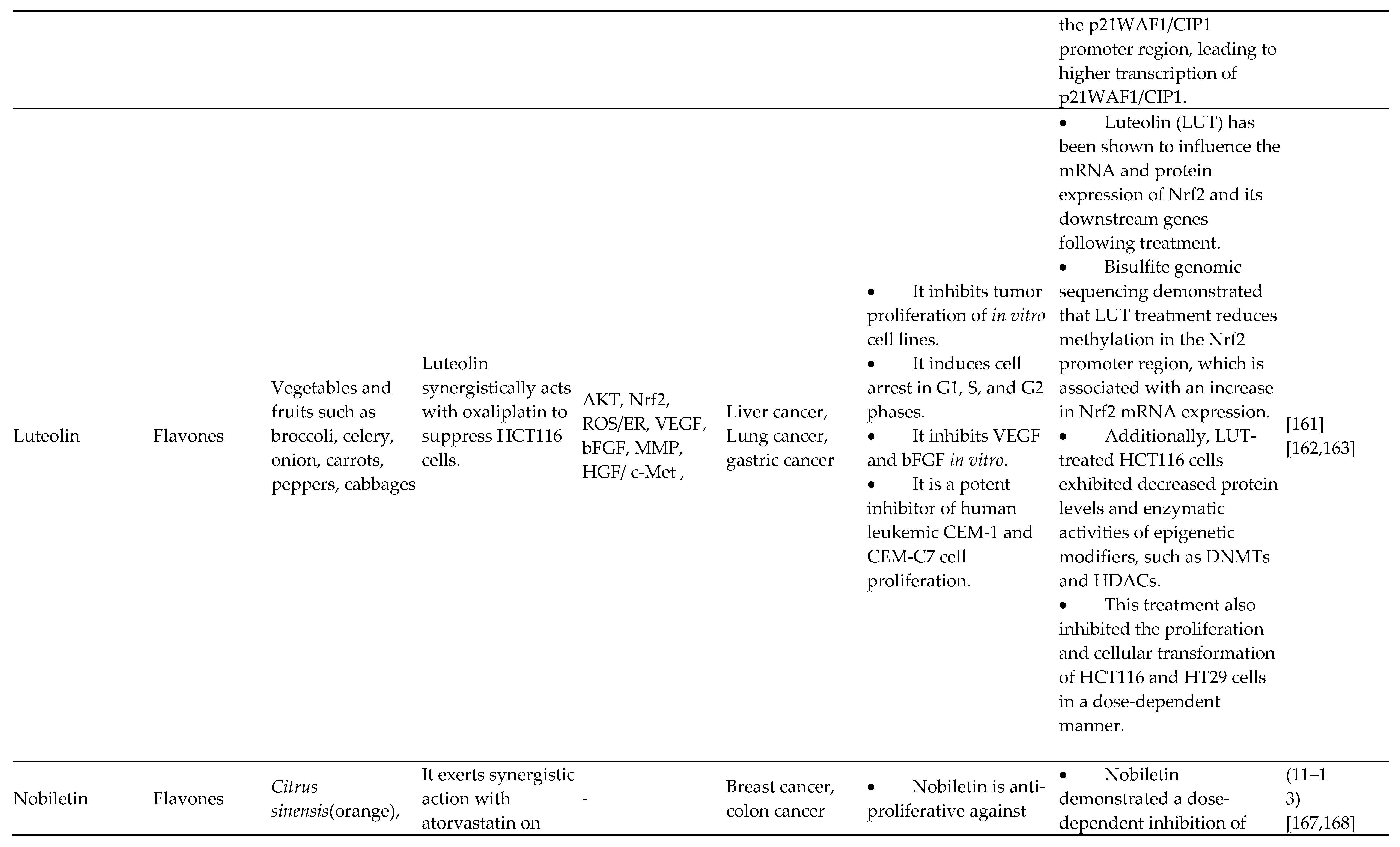


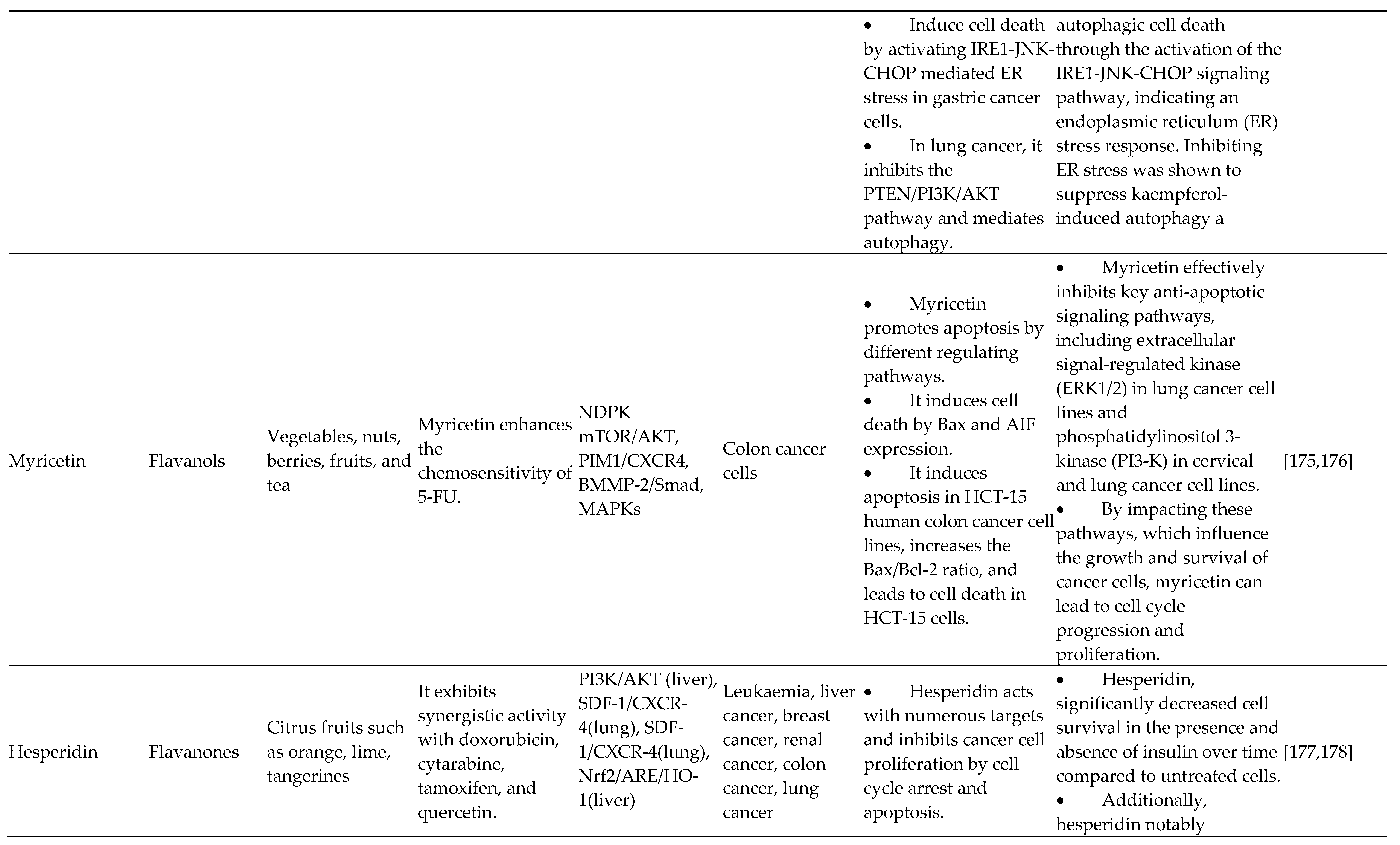

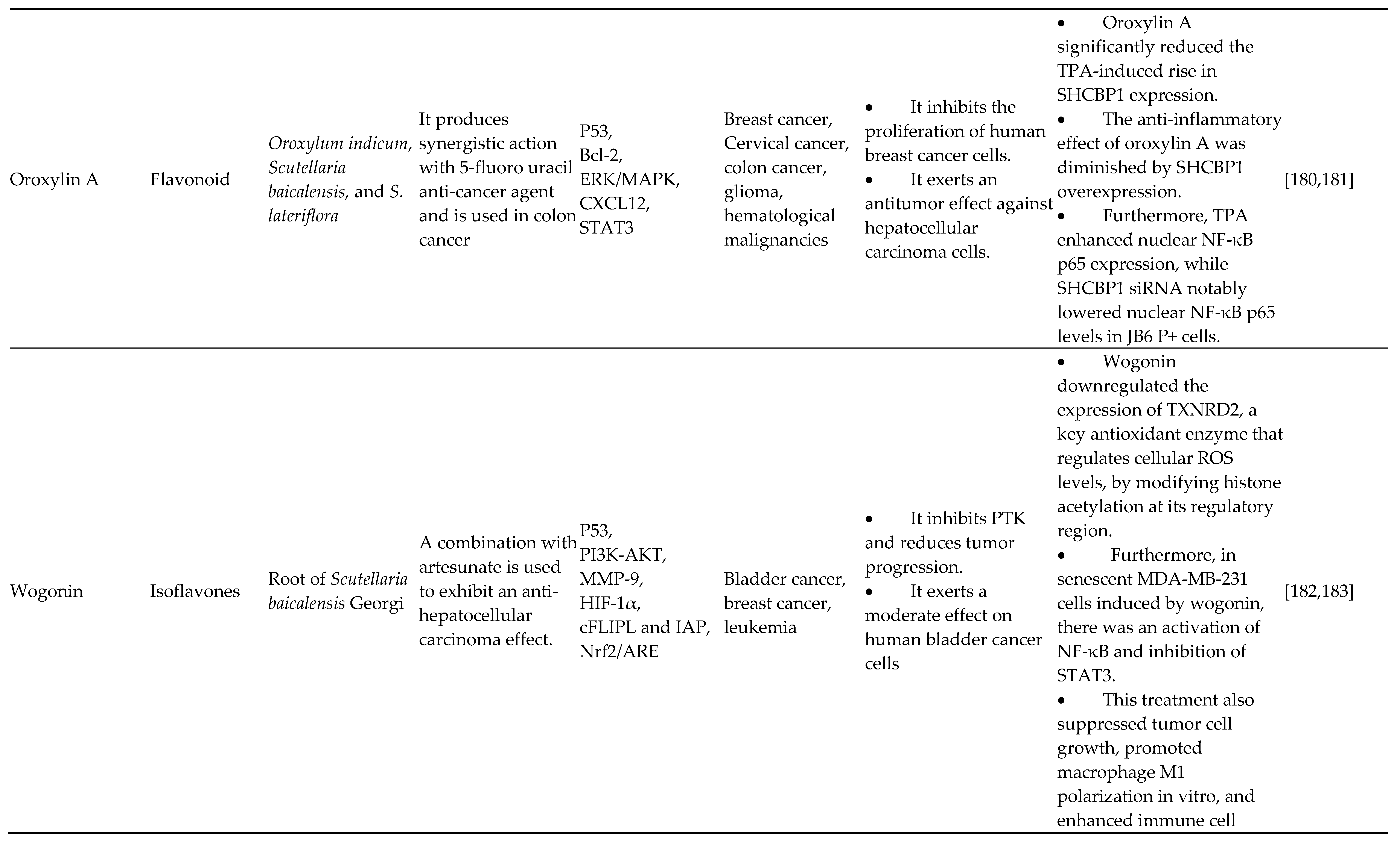
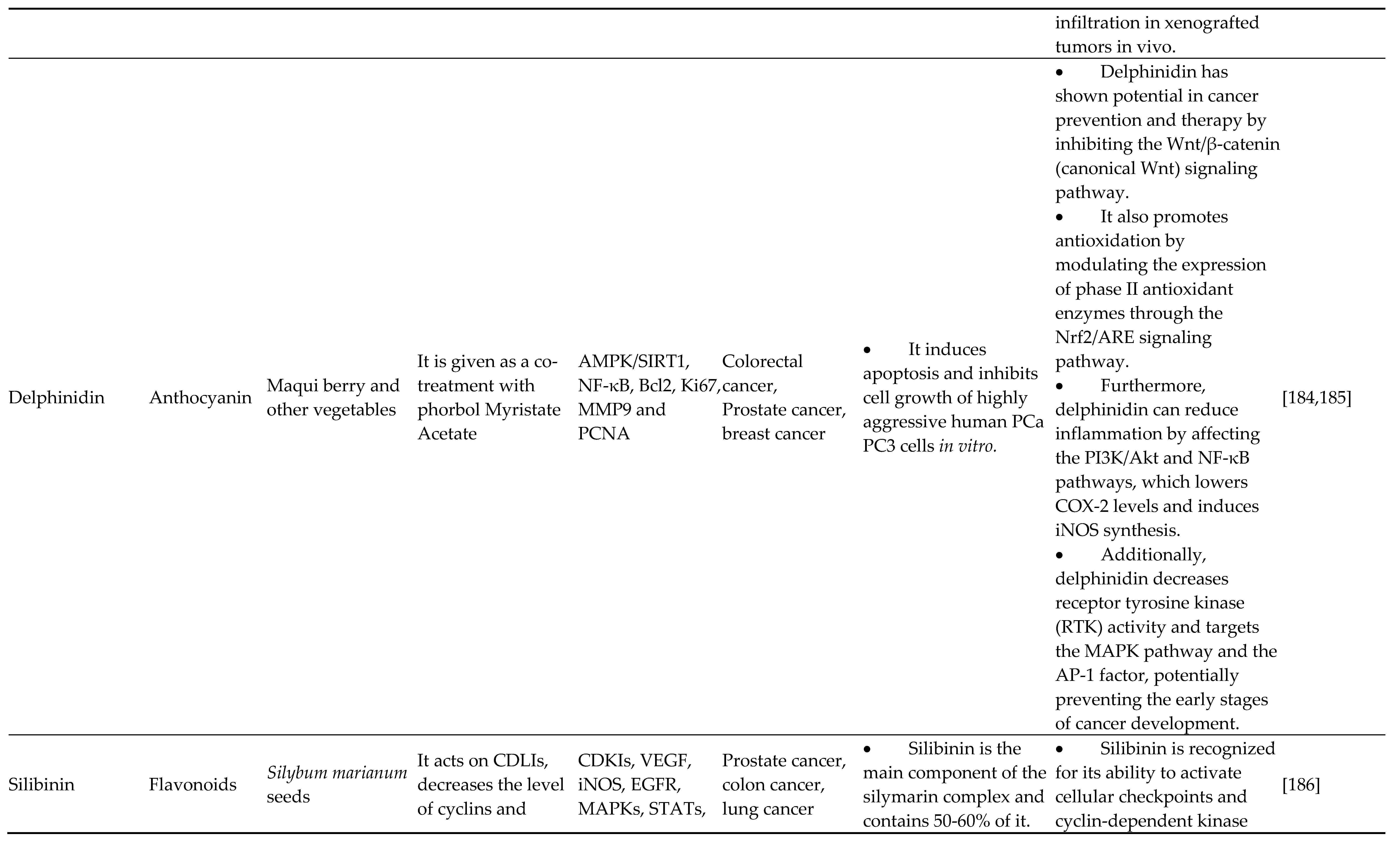

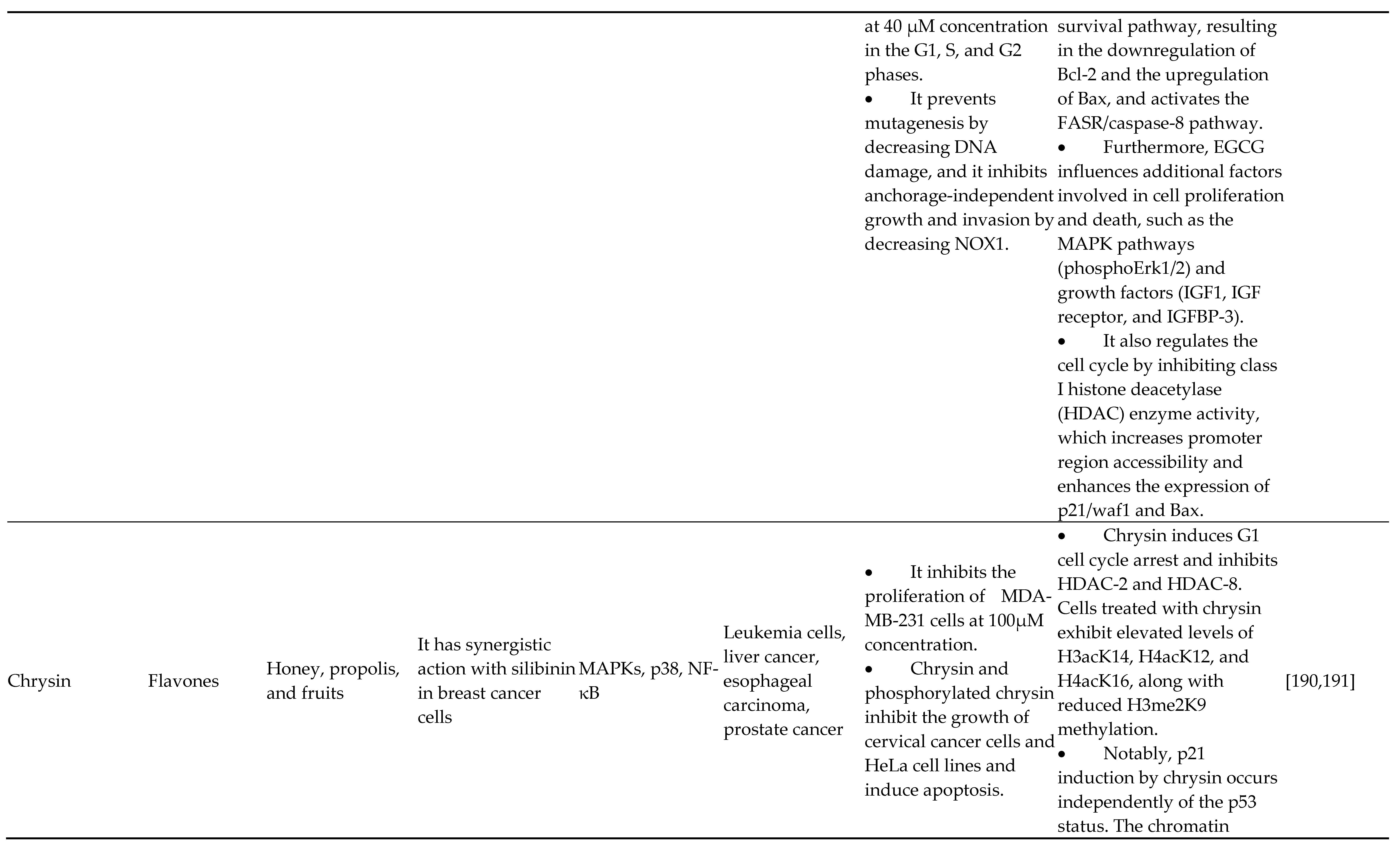

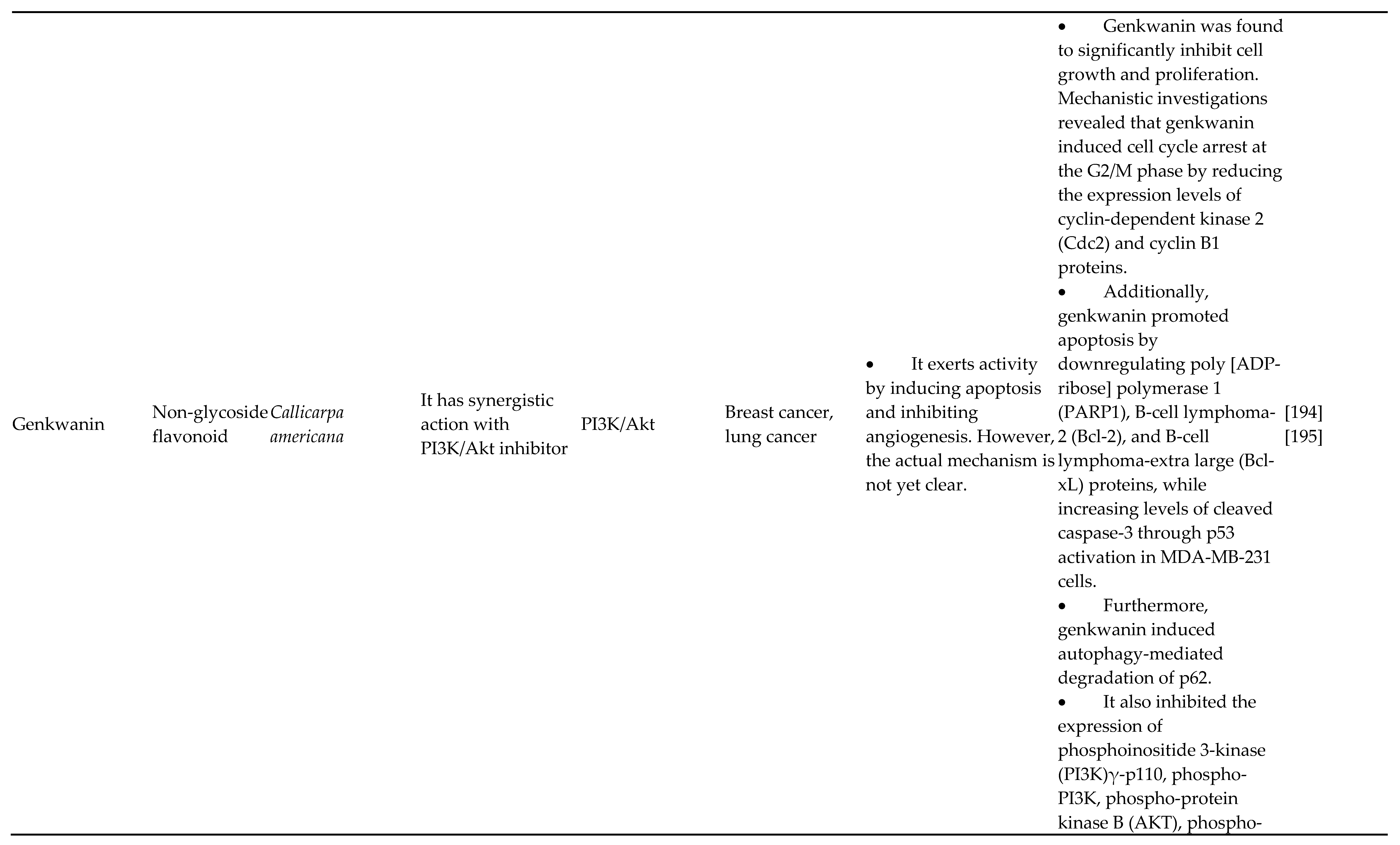
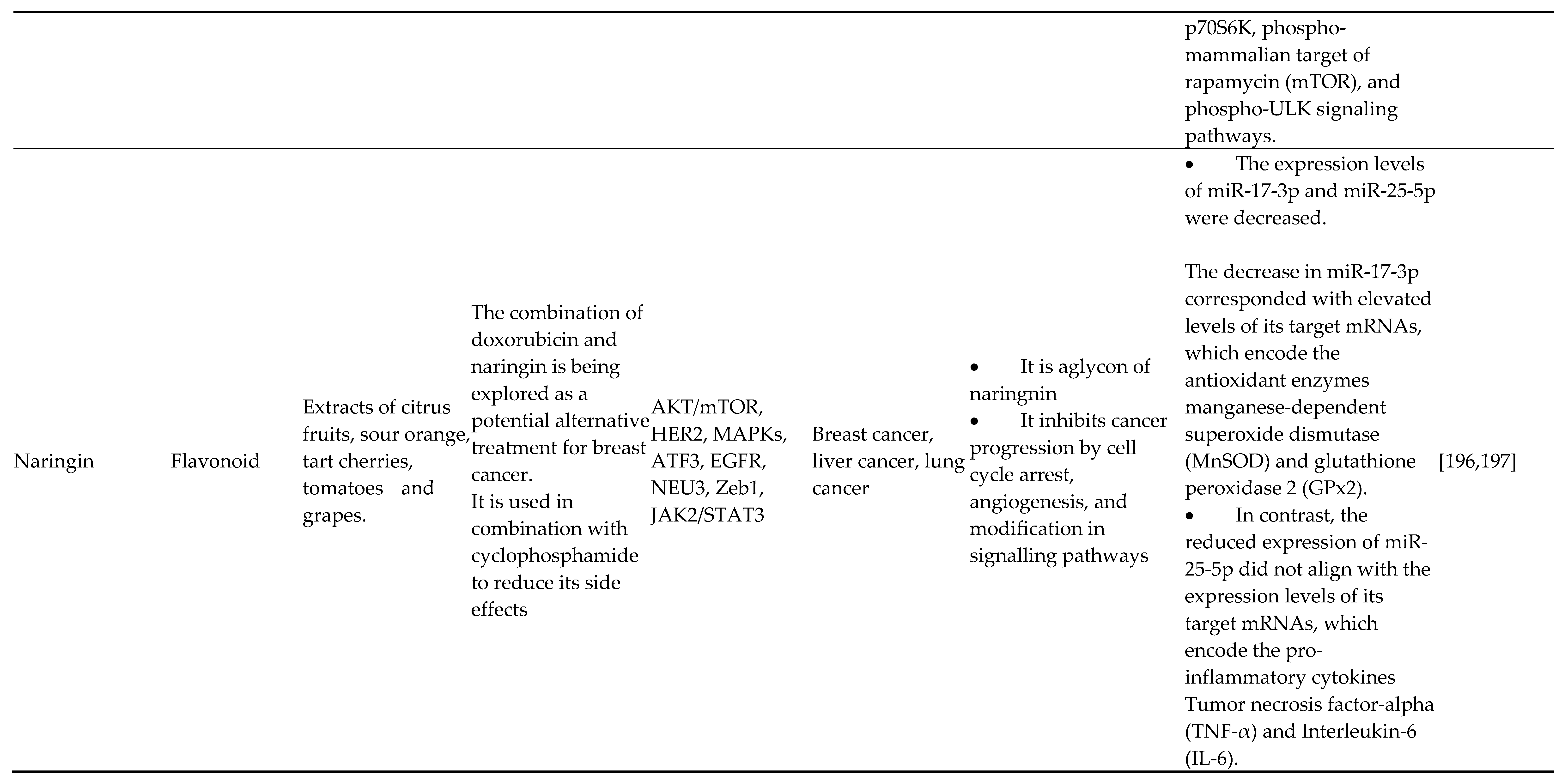
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



