Submitted:
04 July 2024
Posted:
05 July 2024
You are already at the latest version
Abstract
The process of oxygenic photosynthesis is primarily driven by two multiprotein complexes known as photosystem II (PSII) and photosystem I (PSI). PSII facilitates the light–induced reactions of water–splitting and plastoquinone reduction, while PSI functions as the light–driven plastocyanin-ferredoxin oxidoreductase. In contrast to the highly conserved structure of PSII among all oxygen–evolving photosynthetic organisms, the structures of PSI exhibit remarkable variations, especially for photosynthetic organisms that grow in special environments. In this review, we make a concise overview of the recent investigations of PSI from photosynthetic microorganisms including prokaryotic cyanobacteria and eukaryotic algae from the perspective of structural biology. All known PSI complexes contain a highly conserved heterodimeric core; however, their pigment compositions and peripheral light-harvesting proteins are substantially flexible. This structural plasticity of PSI reveals the dynamic adaptation to environmental changes for photosynthetic organisms.
Keywords:
photosystem I
; LHCI
; cyanobacteria
; algae
; environmental adaptation
1. Introduction
Photosystem I (PSI) is a large pigment-protein complex that catalyzes the reactions including the light-induced electron transfer and the reduction of ferredoxin (Fd) at the early stage of oxygenic photosynthesis [1]. The basic functional unit of PSI consists of a core complex and its peripheral light-harvesting antenna. The core complex harbors three pairs of chlorophylls (Chls), two phylloquinones and three [4Fe-4S] clusters, which constitute the central electron transfer chain (ETC) [2]. Once the solar energy is absorbed by antenna pigments, it is transferred to the core complex and the excited energy is trapped at P700, a special pair of Chls, where charge separation happens. The generated free electron is quickly transferred to one acceptor Chl (A0) and subsequently to the terminal electron acceptor Fd through cofactors along the ETC. Simultaneously, the oxidized P700 (P700+) is replenished with an electron from a soluble electron donor of either a plastocyanin or a cytochrome. The reduced Fd provides the reducing power for the generation of ATP and NADPH, both of which are used in the subsequent CO2 fixation reactions [3].
Due to its crucial role, the structure and function of PSI have been extensively studied [4]. The first atomic structure of cyanobacterial PSI revealed by X–ray crystallography is composed of 12 protein subunits and 127 cofactors [5]. With the rapid development of structural biology, especially the advancements of cryo-electron microscopy (cryoEM) technology, a large number of new studies have revealed the structural plasticity of PSI, which is important for the self-regulation mechanisms of photosynthetic organisms to cope with various environmental challenges such as light intensity fluctuations, far red light, iron- deficiency stress and salt stress [6,7,8,9]. In spite of the conserved function of PSI in different oxygen-evolving photosynthetic organisms, the structural organization of PSI exhibits significant differences [6,7,8,9]. In particular, prokaryotic cyanobacteria have PSI core complexes in the form of monomers, dimers, trimers, or tetramers, while eukaryotes mainly have monomer PSI associated with various light–harvesting proteins [10,11,12,13,14,15].
This review focuses on the structural variations of PSI in photosynthetic microorganisms, including prokaryotic cyanobacteria and various eukaryotic algae, and discusses the molecular mechanisms of high–efficiency energy and electron transfer within different PSI complexes. Through an in–depth exploration of these variations, we aim to inspire new thinking about the key role of PSI and the adaptive mechanisms by which different photosynthetic organisms thrive in different ecological niches.
2. Structural Variations of PSI Complexes in Cyanobacteria
2.1. Oligomers of PSI Complexes in Cyanobacteria
In most Cyanobacteria, the PSI monomers assemble into a stable trimeric structure with a 3-fold rotational symmetry [5] (Figure 1). Each PSI monomer consists of a highly conserved heterodimeric core (PsaA/B) and variable peripheral subunits (PsaC/D/E/F/I /J /K/L/M/X), depending on the species [10]. The majority of the cofactors are coordinated by the dimeric PsaA/B core, and the molecular mass of a PSI trimer exceeds 1M Da [5]. Recently, owing to advancements of cryo–EM, structures of PSI monomers, tetramers and some heterogeneous complexes from Cyanobacteria have been determined at near atomic resolutions [15,16,17,18] (Figure 1).
The tetrameric PSI is widespread among heterocyst-forming cyanobacteria such as the mesophilic filamentous heterocyst–forming cyanobacterium Anabaena sp. PCC 7120 and the thermophilic non-heterocyst-forming cyanobacterium Chroococcidiopsis sp. TS–821, and their close relatives [18,19,20]. Physiological studies have shown that factors such as nitrogen source, temperature stress or salinity have no effect on the formation of PSI trimer or tetramers; however, under high-light conditions, tetrameric PSI formation is favored and is accompanied by an increased content of novel carotenoids (Cars) like myxoxanthophyll, canthaxanthin, and echinenone [20]. Compared with the trimeric PSI, the four monomers within the PSI tetramer are structurally organized as a dimer of dimers with a C2 symmetry, forming two distinct interfaces (Figure 1). The two attached PSI dimers are stabilized to a large extent by electrostatic interactions of amino acid residues at the interfaces, as well as by the nearby Chls and specific lipids [21] (Figure 1).
The dimeric cyanobacterial PSI complexes can be identified by biochemical methods in Thermosynechococcus elongatus and Anabaena sp. PCC 7120 [13,18]; however, only the structure of Anabaena PSI dimer had been reported, which is same as one of the dimers in a PSI tetramer [15] (Figure 1). Low–resolution structures of monomeric PSI complexes have only recently been reported [11,22] (Figure 1). The PSI monomer shows no significant difference in NADP+ reduction rates compared to the PSI trimer [22]; however, the fluorescence spectrum of monomeric PSI exhibits a significant blue shift, which is caused by the loss of several peripheral red Chls [11]. Another notable difference lies in the disruption of the short C-terminal α-helix of PsaL after monomerization [13,22]. This short α-helix is believed to function as a critical factor in the formation and stabilization of trimeric and tetrameric PSI complexes [17,23,24,25].
2.2. PSI-IsiA in Iron–Deficient Environment
Over the long period of evolution, cyanobacteria have developed special antenna systems that enable them to thrive under diverse environmental pressures, notably low iron, intense light, and oxidative stress [26,27,28,29,30]. These antenna systems primarily comprise two types: membrane-bound iron-stress-induced protein A (IsiA) and the prochlorophyte Chl a/b–binding (Pcb) protein family, and the water-soluble phycobilisomes [31,32]. Both of the IsiA/Pcb families possess six transmembrane helices and exhibit similar structural features with the CP47 and CP43 subunits of PSII [33,34,35] (Figure 2). Under normal growth conditions, in order to achieve efficient absorption and utilization of light, cyanobacteria form giant complexes of PSI–phycobilisome and PSII–phycobilisome, whose structures have recently been resolved [36,37,38].
In cyanobacterium Synechocystis sp. PCC 6803, IsiA proteins can congregate in clusters of up to 18 copies surrounding a trimeric PSI core, forming a PSI3–IsiA18 supercomplex [39,40] (Figure 2). The binding IsiA complexes enhance the absorption cross section, thereby compensating for the diminished PSI levels within the membrane [41,42,43]. The IsiA protein was primarily proposed to serve as an excess energy quencher and/or a reservoir for Chls, facilitating its subsequent integration into the photosystems [44,45,46]. However, high-resolution structure and time–resolved fluorescence spectra of the PSI–IsiA complex from a thermophilic cyanobacterium Thermosynechococcus vulcanus showed clear excitation-energy transfer from IsiA to PSI, strongly indicating that the IsiA protein mainly functions as an energy donor, but not an energy quencher within the complex [30]. When cultured in iron deprivation environments, the protein Flavodoxin (Fld) takes over the role of Fd, accepting electrons from one of the [4Fe-4S] clusters coordinated by the PSI subunit PsaC [47]. The structure of the PSI–IsiA18–Fld3 complex from Synechococcus sp. PCC 7942 reveals that three Fld molecules bind symmetrically to the trimeric PSI core [26]. Within each PSI monomer, Fld utilizes its surface acidic residues to form a stable interaction with a positively charged pocket on PSI [26].
The number of isiA genes varies among different species of cyanobacteria. Cyanobacterium Anabaena sp. PCC 7120 has four types of isiA genes: isiA1, isiA2, isiA3, and isiA5 [28]. However, the structures of the IsiA proteins identified in the PSI–IsiA complexes from Synechocystis sp. PCC 6803, Synechococcus elongatus PCC 7942, and Thermosynechococcus vulcanus NIES–2134 are similar to that of IsiA1 from Anabaena [26,27,30]. In Anabaena, the PSI–monomer–IsiA complex consists of six IsiA subunits, five of which contain six transmembrane helices that bind Chls and Cars [29] (Figure 2). The remaining IsiA subunit (IsiA2) adjacent to PsaD/I has nine transmembrane helices and exhibits a remarkable structural correspondence with PsaL, particularly in the C–terminal domain which may substitute the role of PsaL in the Anabaena PSI tetramer [29] (Figure 2).
2.3. PSI Complexes from Chls d/f–Containing Cyanobacteria
Chlorophylls are crucial for energy capture, transfer, and charge separation in photosynthesis [48,49]. Photosynthetic organisms have evolved a variety of Chls with varied molecular structures and absorption ranges to achieve efficient light utilization [50]. Most oxygenic photosynthesis organisms possess Chls a in their photosynthetic cores complex as well as their light-harvesting antennas [51]. However, other types of Chls, such as Chls b, c, d, and f, are found in specific lineages, which enables organisms to occupy unique ecological niches [49,52,53]. Unlike Chls a, b and c, which mainly absorb higher–energy light [54], Chls d and f absorb lower–energy light [55,56,57]. A mixture of different types of Chls broaden the absorption spectrum and promote the light utilization efficiency [51].
Acaryochloris marina (A. marina) is a unique cyanobacterial species which uses d–type Chls as its dominant photosynthetic pigments, and is capable of using far–red light to drive oxygenic photosynthesis [58,59,60]. The structure of A. marina PSI has been determined with resolutions of 2.58 Å and 3.3 Å, respectively [61,62]. The overall structure of A. marina PSI resembles the PSI trimers from other cyanobacteria; however, PsaI and PsaX are missing in A. marina PSI. A novel subunit, Psa27, identified in A. marina PSI exhibits similar structure and location with PsaI of Thermosynechococcus elongatus, indicating their same functions in stabilizing the PSI trimer [62]. Uniquely, the paired Chls (known as P740) along the ETC of A. marina PSI is a dimer of Chl d and its epimer Chl d′, and the primary electron acceptor A0 is composed of two pheophytin a, rather than Chl a, which is found in other known PSI structures [61,62] (Figure 3).
Chl f–containing cyanobacteria demonstrate a remarkable capacity of photosynthesis in the far–red and near–infrared regions [57,63,64]. However, Chl f is only induced under far–red light conditions and accounts approximately 10% of the total Chls, indicating their specific roles within the photosystem [65,66]. The structures of PSI complexes from Halomicronema hongdechloris (H. hongdechloris) and Fischerella thermalis PCC 7521 grown under far–red light have been resolved [67,68]. The H. hongdechloris PSI binds 83 Chls a and 7 Chls f, with all Chls f located at the periphery of PSI and excluded from the electron transfer chain, which suggests that Chls f function to harvest far-red light and enhance uphill energy transfer [68]. Further studies demonstrate that far-red light induces extensive remodeling of the photosynthetic apparatus in H. hongdechloris by altering the expression of genes encoding PSI core subunits, and by modifying the types of pigments associated in PSI [69,70,71]. Indeed, differences in the sequences of several core subunits, including PsaA, PsaB, PsaI, and PsaL, have been observed in the structure of PSI under far–red light compared to white light [68].
3. Structural Variations of Algal PSI-LHCI Complexes
3.1. PSI-LHCI Complexes of Chlamydomonas reinhardtii
The unicellular eukaryotic green alga Chlamydomonas reinhardtii (C. reinhardtii) is a model organism for studying photosynthesis, and the structure and function of its photosystem have been extensively studied [72]. The overall structure of C. reinhardtii PSI–LHCI complex is similar to that of plant PSI–LHCI, which is composed of a core complex and a peripheral antenna system [73]. However, C. reinhardtii PSI combines more peripheral antenna subunits, and the 10 Lhca proteins are distributed in three belts: one inner belt (Lhca1a/Lhca8/Lhca7/Lhca3), one outer belt (Lhca1b/Lhca4/Lhca6/Lhca5) and a Lhca2–Lhca9 heterodimer [74] (Figure 4). The Lhca2/Lhca9 heterodimer loosely attaches to the PSI core at the opposite side from the other LHCI belts, which may provide a docking site for the cytochrome b6f complex and enhance the photosynthetic cyclic electron flow [74]. All the C. reinhardtii Lhca subunits have a conserved transmembrane structure with other LHC family members, while showing differences at the N and C terminals [74]. The huge light-harvesting system and its complicated pigment network enable C. reinhardtii cells to adapt to the changing light environment, which is the result of long-term evolution.
To optimize the photosynthetic efficiency in fluctuating light conditions, photosynthetic organisms like plants and algae have developed a short–term light adaptation mechanism called state transitions [75]. This process ensures a balanced distribution of excitation energy between the two photosystems, PSI and PSII, through dynamic relocation of the light-harvesting antenna complex II (LHCII) [76,77]. The state transitions are regulated by the redox state of the plastoquinone (PQ) pool: in state 1, when the PQ pool is oxidized, LHCII remains primarily associated with PSII, forming the PSII-LHCII complex; conversely, in state 2, upon reduction of the PQ pool, protein kinases (such as STN7 in higher plants or STT7 in green algae) are activated via the cytochrome b6f complex [78,79]. These kinases then phosphorylate LHCIIs, promoting their partial dissociation from PSII and migration to PSI, which triggers the formation of PSI–LHCI–LHCII supercomplex, enhancing PSI’s light–harvesting capacity [80,81,82,83]. High–resolution structure of the PSI–LHCI–LHCII supercomplex from C. reinhardtii has been elucidated, and all four types of LHCII are found to associate with PSI under state 2 conditions [82] (Figure 4). Two LHCII trimers are associated with PSI–LHCI at the PsaO–PsaL–PsaH–Lhca2 side: one (LHCII–1) attaches to the PSI core by PsaO, PsaH, and PsaL, the other one (LHCII–2) attaches to the Lhca2 and LHCII–1 subunits [82] (Figure 4). As PSII is generally considered to be more susceptible to oxidative damage than PSI under high–light conditions, the transfer of LHCII to PSI helps to reduce the excessive accumulation of excitation energy in PSII, thereby reducing the oxidative damage of PSII [81,84,85,86]. However, if the energy absorbed by LHCII is excessively funneled to the PSI core, the resulting overactivation of PSI can trigger the production of reactive oxygen species (ROS), ultimately diminishing the overall photosynthetic efficiency [84].
Recently, PSI–LHCI dimer is isolated from C. reinhardtii cells grown in low–light and anoxic conditions [87]. This unique oligomerization of PSI is formed by two PSI–LHCI monomers arranged head-to-head, containing 40 protein subunits and more than 600 cofactors [87]. Unlike the dimeric formation observed in cyanobacterial PSI tetramers, which is mainly due to the movement of the stromal helices of PsaL [17], C. reinhardtii PSI–LHCI dimer is formed by the interactions of four subunits: Lhca9, PsaI, PsaL, PsaG, and their associated pigments [87]. The PsaH and Lhca2 subunits observed in PSI–LHCI monomer are absent in the C. reinhardtii PSI–LHCI dimer, instead, a second Lhca9 subunit occupies the corresponding space for PsaH and Lhca2, and is attached the PsaG, PsaL and PsaI subunits of two PSI-LHCI monomers (Figure 4) [87]. This unique oligomeric state of PSI–LHCI in C. reinhardtii reflects the diversity of its adaptation mechanisms to different environmental conditions [87].
3.2. PSI-LHCI Complexes of Red Algae
In addition to state transition, an alternative strategy that algae employed to regulate the antenna size of PSI is to change the number of associated LHCI subunits [88]. For red algae Cyanidioschyzon merolae (C. merolae), even when grown under optimal laboratory conditions, two PSI–LHCI complexes with antenna systems of different sizes can be isolated [89] (Figure 5). The smaller form (PSI–3Lhcr) binds three antenna subunits which form an LHCR band and interact with PsaF, PsaJ, PsaA, and PsaK. The larger form (PSI–5Lhcr) binds two additional Lhcr proteins that are associated with the surface of PsaL, PsaI, PsaM, and PsaB, forming an additional LHCR* belt [89]. The major structural difference between PSI–5Lhcr and PSI–3Lhcr lies in the additional LHCR* belt, the counterpart core subunits and three Lhcr subunits of the canonical LHCR belt are identical [89]. Biochemical and spectroscopic data have revealed a close correlation between the ratio of these two forms and the light intensities in red algae [90]. Similarly, in the unicellular green algae C. reinhardtii and Bryopsis corticulans [91], the PSI core complex can associate with either 8 to 10 LHCI subunits, indicating that the binding state of LHCI and PSI in algae is flexible with environmental conditions [88]. Uniquely, the red alga Porphyridium purpureum PSI–LHCI contained seven LHCI subunits and one chlorophyll a/b-binding- like protein (RedCAP) and another red alga Cyanidium caldarium RK–1 belonging to the Cyanidiophyceae possesses seven LHCI subunits (Figure 5).
3.3. A Minimal PSI from Salt-Tolerant Green Alga Dunaliella salina
Dunaliella salina (D. salina) represents a unicellular green alga which can adapt to hypersaline environments and light stress. There exist two forms of PSI-LHCI in D. salina:
a mini PSI–LHCI and a large PSI–LHCI [92]. The mini D. salina PSI–LHCI contains only seven PSI core subunits (PsaA–F and PsaJ) and four LHCI proteins (Lhca1–4) which are positioned at similar positions as the counterparts in plant PSI-LHCI [73,93] (Figure 6). The Large D. salina PSI–LHCI contains 13 core subunits and 6 LHCI proteins (Lhca1–6), exhibiting a similar structure when compared to PSI–LHCI complexes derived from red algae and green algae [88,91]. However, the large D. salina PSI–LHCI lacks the second round of light-harvesting antenna (Figure 6). The different forms of D. salina PSI–LHCI complexes reveal different regulatory mechanisms of reducing the association of antenna proteins or forming distinct subunit interactions under certain physiological conditions.
3.4. PSI–LHCI in Desert Algae Chlorella ohadii
In the highly light–tolerant green algae Chlorella ohadii (C. ohadii), it has been observed that even when exposed to light intensities four times higher than those required for saturating CO2 fixation, the algae do not suffer from photodamage [94,95]. A comparative analysis of the PSI–LHCI structures from C. ohadii cells grown in low-light (LL) and high-light (HL) environments reveals that, to minimize photodamage, part of LHCI and the PSI core subunit, PsaO, are eliminated in PSIHL [96]. The devoid of the PsaO subunit indicate that the state transition is not triggered in C. ohadii under high light conditions (Figure 6). Another remarkable difference between PSILL and PSIHL lies in the pigment composition and their number in LHCIs: approximately 50% of the Chls b in LHCIHL are replaced by Chls a [96]. Notably, the highest substitution rates occur in the first LHCIHL belt and the LHCIHL dimer, both of which surround the PSI core complex [96]. This pigment substitution may lead to higher electron transfer rates within C. ohadii PSIHL as PSIHL exhibits a higher efficiency of photocurrent induction [96].
3.5. Diatom PSI-FCPI Complex
Diatoms are a large group of eukaryotic algae which account for about 20% of the global primary carbon fixation [97]. The PSI of diatoms is distinguished by a large number of light–harvesting subunits called fucoxanthin–Chl proteins (FCPIs), which bind Chl a/c instead of Chl b and fucoxanthin instead of lutein [98]. The PSI-FCPI complex from diatom Chaetoceros gracilis shows an asymmetrical heart-shaped structure, consisting of 12 core subunits and 24 FCPIs, and associating with 326 Chls a, 34 Chls c, 102 fucoxanthins, 35 diadinoxanthins, 18 β–carotenes and other cofactors [99] (Figure 7). The structure of diatom PSI core is conserved with that of cyanobacterial PSI, however, the subunit of PsaK is missing and two new subunits (PsaR and PsaS) are present [99]. The large FCPI antenna around the PSI core could be divided into three layers: an innermost layer, a semi-ring middle layer and an outermost layer [99]. Each FCPI subunit exhibits distinctive structures with different pigment compositions, and these FCPI subunits interweave with the PSI core to form a sophisticated pigment-protein network for the efficient light capture and energy transfer [99].
3.6. Tetrameric PSI from Glaucophyte Algae
Glaucophyte alga is a unique photosynthetic eukaryote that has plastid-like organelles termed cyanelles [100]. The cryo–EM structure of PSI from the glaucophyte alga Cyanophora paradoxa is determined as a tetramer, however, it is remarkably different from the previously observed tetrameric PSI from cyanobacteria in subunit composition and organization [101]. The Cyanophora PSI tetramers are composed of two types of structurally similar PSI monomers termed as monomer 1 and monomer 2, the latter of which lacks the PsaK subunit [101]. A monomer 1 attaches to a monomer 2, creating a monomer 1-monomer 2 heterodimer, and two such heterodimers further give rise to a tetramer in an inverse parallel manner [101]. Due to the unique assembly, the monomer–monomer interactions as well as the excitation energy transfer among Chls in Cyanophora PSI tetramers are entirely different from that in cyanobacteria PSI tetramers [101]. The distinctive structural features of Cyanophora PSI highlight a vital evolutionary transition in photosynthetic machineries, illustrating an intermediary phase in the transformation from oligomeric forms to monomeric units within this early eukaryotic alga [101].
3.7. PSI-ACPI Complex in Cryptophytes
Cryptophytes (also called Cryptomonads or Cryptophyceae) are ancestral photosynthetic organisms that evolved through secondary endosymbiosis possibly between a red alga-like organism and a heterotrophic host [102,103,104]. Cryptophyte cells perform efficient oxygenic photosynthesis through their membrane-imbedded alloxanthin–Chl a/c binding proteins (ACPs) and soluble phycobiliproteins as light harvesting antennas [105]. The PSI-ACPI supercomplex from the cryptophyte Chroomonas placoidea consists of a monomeric PSI core and a huge peripheral antenna [106]. The PSI core contains 14 subunits, including 12 red algae-originated subunits, one diatom PsaR homolog, and one loosely associated extrinsic subunit (Unk1) [106]. The outer antenna surrounding the PSI core is made up of 14 ACPI subunits which are distributed in two layers: the inner layer has 11 ACPIs, and the outer layer has 3 ACPIs [106]. The association and energy transfer between the outer and inner ACPIs are mediated by a 20.6–kDa pigment-binding subunit termed ACPI-S [106]. However, the structure and pigment binding sites of ACPI–S are different from the typical ACPI subunits [106]. Specifically, all ACPI apoproteins contain three major TM helices (αA, αB, αC) and an additional amphipathic helix (αD or αE). By contrast, ACPI–S has only one transmembrane helix and an amphipathic helix, but contains several long–terminal loops [106].
3.8. PSI-ACPPCI Complex in Symbiotic Dinoflagellates
Dinoflagellates constitute a significant proportion of unicellular eukaryotes, inhabiting diverse aquatic habitats [107,108,109]. Many dinoflagellate species could form symbionts with invertebrates such as corals, or with algae like diatoms [110,111,112]. The well-known symbiosis between corals and photosynthetic dinoflagellates of the Symbiodiniaceae family is instrumental in the development and sustenance of coral reefs [110]. The PSI–LHCI supercomplex from Symbiodinium comprises a core complex and a unique peripheral antenna system, showing similar structural features with red algal PSI–LHCR, cryptophyte PSI–ACPI, and diatom PSI–FCPI, but exhibiting specific characteristics in the protein organization [113,114,115]. In particular, the PSI core consists of 13 subunits including 2 new-found extrinsic subunits, PsaT and PsaU [115]. The overall structure of Symbiodinium PSI core is similar to that of the diatom PSI core, but the PsaK and PsaO subunits are missing [115]. Remarkable differences emerge as the modifications to the extrinsic loop regions of PsaA and PsaB subunits, and the alterations in the C-terminal regions of several subunits including PsaD/E/I/J/L/M/R [114,115]. The peripheral antenna system of Symbiodinium PSI is composed of 13~14 peridinin–Chl a/c–binding light-harvesting antenna proteins (AcpPCIs), which are distributed in two layers around the PSI core [114,115]. Most of the pigment binding sites in Symbiodinium PSI–AcpPCI are conserved with those in diatom PSI–FCPI, but there are some notable differences. Although Symbiodinium PSI–AcpPCI contains a small number of pigments and antenna subunits, the extended end domains of the PSI core and antenna subunits enable efficient protein interactions and intermolecular energy transfer [114,115]. Interestingly, in a red tidal dinoflagellate Amphidinium carterae, both the structures of PsaA/B subunits exhibit substantial shortenings and have more short loops, leading to a reduction of over 20 pigment-binding sites compared with that of diatom PsaA/B subunits [114]. However, the other core subunits, including PsaD/ F/I/J/L/M/R, show significant elongations and additional pigment-binding sites compared with those in diatom PSI [114]. Additionally, the Amphidinium carterae PSI core is associated with 18 AcpPCIs that bind a large number of xanthophyll cycle Cars, which may compensate for the smaller PsaA/B subunits [114].
4. Perspective
The precise structural determination of photosynthetic protein complexes is crucial for revealing the mechanisms of their efficient working modes. As more and more PSI structures from different species are resolved, we are able to gain a more complete understanding of how this complex protein machine works. Here, we discussed the structural and functional characteristics of various PSI complexes and their associated light-harvesting proteins in cyanobacteria and algae, reflecting their remarkable plasticity of environmental adaptation under different conditions.
Given its pivotal role in oxygenic photosynthesis, the structural diversity of PSI is a consequence of billions of years of biological evolution. For eukaryotic photosynthetic organisms (such as algae and plants), they tend to evolve more complex light-harvesting systems and finer energy regulation mechanisms (such as the state transition mechanism), while in prokaryotic photosynthetic organisms (such as cyanobacteria), there are different aggregation forms of reaction centers. Across all oxygen-evolving photosynthetic organisms, PSI is characterized by a structurally conserved heterodimeric core. This core serves as a critical binding platform for the majority of PSI's cofactors, establishing the foundational architecture essential for the complex processes of charge separation and electron transport within the photosynthetic machinery.
Recent structural analyses of photosystems from non-oxygenic phototrophic bacteria, including Heliobacteria, green sulfur bacteria, and Acidobacteria, reveal that their reaction centers embody homodimeric type I photosystems. These photosystems exhibit a striking structural conservation with the PSI core of oxygenic photosynthetic organisms, albeit notably devoid of the intricate membrane-bound light-harvesting protein subunits characteristic of the latter. Instead, these organisms have evolved an extracellular light-harvesting apparatus—such as the chlorosome in green sulfur bacteria—to efficaciously capture light energy, compensating for the absence of complex intracellular light-trapping mechanisms. In recent years, the structures of photosystems from non-oxygen-producing photosynthetic bacteria, including heliobacteria, green sulfur bacteria and Acidobacteria, have been resolved [116,117,118]. The photosynthetic reaction centers of these bacteria are in the forms of homodimers, which are structurally highly conserved with the PSI core of oxygen-producing photosynthetic organisms, but lack complex light-trapping protein subunits within the membrane. Instead, efficient absorption of light energy is mainly achieved through specific extracellular light-trapping systems, such as chlorosome.
Author Contributions
L.T. and J.-H.C. wrote the manuscript. J.-H.C. finalized the manuscript. L.T. and J.-H.C. provided the funding. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Zhejiang Provincial Natural Science Foundation of China grant LR22C010001, and National Natural Science Foundation of China grants 32100202 and 32100189.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Nelson, N.; Ben-Shem, A. The complex architecture of oxygenic photosynthesis. Nat. Rev. Mol. Cell Biol. 2004, 5, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Chitnis, P.R. Photosystem I: function and physiology. Annu. Rev. Plant Biol. 2001, 52, 593–626. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.F. Photosynthesis of ATP-electrons, proton pumps, rotors, and poise. Cell 2002, 110, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Wang, Y. Function and Structure of Cyanobacterial Photosystem I. In Photosynthesis: Structures, Mechanisms, and Applications; Chapter 7; Hou, H., Najafpour, M., Moore, G., Allakhverdiev, S., Eds.; Springer: Cham, Switzerland, 2017; pp. 111–168. ISBN 978-3-319-48873-8. [Google Scholar]
- Jordan, P.; Fromme, P.; Witt, H.T.; Klukas, O.; Saenger, W.; Krauss, N. Three-dimensional structure of cyanobacterial photosystem I at 2.5 Å resolution. Nature 2001, 411, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Hippler, M.; Nelson, N. The plasticity of photosystem I. Plant Cell Physiol. 2021, 62, 1073–1081. [Google Scholar] [CrossRef]
- Bai, T.; Guo, L.; Xu, M.; Tian, L. Structural diversity of photosystem I and its light-harvesting system in eukaryotic algae and plants. Front. Plant Sci. 2021, 12, 781035. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.-R. Structure, function, and variations of the photosystem I-antenna supercomplex from different photosynthetic organisms, in Macromolecular Protein Complexes IV. Subcellular Biochemistry, Harris J. R., Marles-Wright J., Eds. 2022, 99, 351–377. [Google Scholar]
- Suga, M.; Shen, J.-R. Structural variations of photosystem I-antenna supercomplex in response to adaptations to different light environments. Curr. Opin. Struct. Biol. 2020, 63, 10–17. [Google Scholar] [CrossRef]
- Grotjohann, I.; Fromme, P. Structure of cyanobacterial photosystem I. Photosynth. Res. 2005, 85, 51–72. [Google Scholar] [CrossRef]
- Çoruh, O.; Frank, A.; Tanaka, H.; Kawamoto, A.; El-Mohsnawy, E.; Kato, T.; Namba, K.; Gerle, C.; Nowaczyk, M.M.; Kurisu, G. Cryo-EM structure of a functional monomeric Photosystem I from Thermosynechococcus elongatus reveals red chlorophyll cluster. Commun. Biol. 2021, 4, 304. [Google Scholar] [CrossRef]
- Nelson, N. Investigating the balance between structural conservation and functional flexibility in Photosystem I. Int. J. Mol. Sci. 2024, 25, 5073. [Google Scholar] [CrossRef] [PubMed]
- Rögner, M.; Mühlenhoff, U.; Boekema, E.J.; Witt, H.T. Mono-, di- and trimeric PS I reaction center complexes isolated from the thermophilic cyanobacterium Synechococcus sp.: Size, shape and activity. Biochim. Biophys. Acta. 1990, 1015, 415–424. [Google Scholar] [CrossRef]
- Chen, M.; Liu, X.; He, Y.; Li, N.; He, J.; Zhang, Y. Diversity among cyanobacterial Photosystem I oligomers. Front. Microbiol. 2022, 12, 781826. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Nagao, R.; Jiang, T.Y.; Ueno, Y.; Yokono, M.; Chan, S.K.; Watanabe, M.; Ikeuchi, M.; Shen, J.R.; Akimoto, S.; et al. Structure of a cyanobacterial photosystem I tetramer revealed by cryo-electron microscopy. Nat. Commun. 2019, 10, 4929. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Li, Y.; Li, X.; Zhong, Q.; Li, N.; Zhang, K.; Zhang, Y.; Chu, H.; Ma, C.; Li, G.; et al. Structural and functional insights into the tetrameric photosystem I from heterocyst-forming cyanobacteria. Nat. Plants 2019, 5, 1087–1097. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Perez-Boerema, A.; Zhang, L.; Li, Y.; Yang, M.; Li, S.; Amunts, A. Distinct structural modulation of photosystem I and lipid environment stabilizes its tetrameric assembly. Nat. Plants. 2020, 6, 314–320. [Google Scholar] [CrossRef]
- Watanabe, M.; Kubota, H.; Wada, H.; Narikawa, R.; Ikeuchi, M. Novel supercomplex organization of photosystem I in Anabaena and Cyanophora paradoxa. Plant Cell Physiol. 2011, 52, 162–168. [Google Scholar] [CrossRef]
- Li, M.; Semchonok, D.A.; Boekema, E.J.; Bruce, B.D. Characterization and evolution of tetrameric photosystem I from the thermophilic cyanobacterium Chroococcidiopsis sp TS-821. Plant Cell. 2014, 26, 1230–1245. [Google Scholar] [CrossRef]
- Li, M.; Calteau, A.; Semchonok, D.A.; Witt, T.A.; Nguyen, J.T.; Sassoon, N.; Boekema, E.J.; Whitelegge, J.; Gugger, M.; Bruce, B.D. Physiological and evolutionary implications of tetrameric photosystem I in cyanobacteria. Nat. Plants 2019, 5, 1309–1319. [Google Scholar] [CrossRef]
- Semchonok, D.A.; Mondal, J.; Cooper, C.J.; Schlum, K.; Li, M.; Amin, M.; Sorzano, C.O.S.; Ramı´rez-Aportela, E.; Kastritis, P.L.; Boekema1, E.J.; Guskov, A.; Bruce, B.D. Cryo-EM structure of a tetrameric photosystem I from Chroococcidiopsis TS-821, a thermophilic, unicellular, non-heterocyst-forming cyanobacterium. Plant Commun. 2022, 3, 100248. [Google Scholar] [CrossRef]
- Netzer-El, S.Y.; Caspy, I.; Nelson, N. Crystal structure of Photosystem I monomer from Synechocystis PCC 6803. Front. Plant Sci. 2019, 9, 1865. [Google Scholar] [CrossRef]
- Malavath, T.; Caspy, I.; Netzer-El, S.Y.; Klaiman, D.; Nelson, N. Structure and function of wild-type and subunit-depleted photosystem I in Synechocystis. Biochim. Biophys. Acta. 2018, 1859, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Chitnis, V.P.; Chitnis, P.R. PsaL subunit is required for the formation of Photosystem I trimers in the cyanobacterium Synechocystis sp. PCC 6803. FEBS Lett. 1993, 336, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Kłodawska, K.; Kovács, L.; Vladkova, R.; Rzaska, A.; Gombos, Z.; Laczkó-Dobos, H.; Malec, P. Trimeric organization of photosystem I is required to maintain the balanced photosynthetic electron flow in cyanobacterium Synechocystis sp. PCC 6803. Photosynth. Res. 2020, 143, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Cao, P.; Cao, D.; Si, L.; Su, X.; Tian, L.; Chang, W.; Liu, Z.; Zhang, X.; Li, M. Structural basis for energy and electron transfer of the photosystem I–IsiA–flavodoxin supercomplex. Nat. Plants. 2020, 6, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Toporik, H.; Li, J.; Williams, D.; Chiu, P.-L.; Mazor, Y. The structure of the stress-induced photosystem I–IsiA antenna supercomplex. Nat. Struct. Mol. Biol. 2019, 26, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Nagao, R.; Yokono, M.; Ueno, Y.; Suzuki, T.; Kato, K.; Kato, K.; Tsuboshita, N.; Jiang, T.; Dohmae, N.; Shen, J.-R.; Ehira, S.; Akimoto, S. Molecular organizations and function of iron-stress-induced-A protein family in Anabaena sp. PCC 7120. Biochim. Biophys. Acta. 2021, 862, 148327. [Google Scholar] [CrossRef]
- Nagao, R.; Kato, K.; Hamaguchi, T.; Ueno, Y.; Tsuboshita, N.; Shimizu, S.; Furutani, M.; Ehira, S.; Nakajima, Y.; Kawakami, K.; et al. Structure of a monomeric photosystem I core associated with iron-stress-induced-A proteins from Anabaena sp. PCC 7120. Nat. Commun. 2023, 14, 920. [Google Scholar] [CrossRef]
- Akita, F.; Nagao, R.; Kato, K.; Nakajima, Y.; Yokono, M.; Ueno, Y.; Suzuki, T.; Dohmae, N.; Shen, J.-R.; Akimoto, S.; et al. Structure of a cyanobacterial photosystem I surrounded by octadecameric isiA antenna proteins. Commun. Biol. 2020, 3, 232. [Google Scholar] [CrossRef]
- Chen, H.-Y.S.; Bandyopadhyay, A.; Pakrasi, H.B. Function, regulation and distribution of IsiA, a membrane-bound chlorophyll a-antenna protein in cyanobacteria. Photosynthetica 2018, 56, 322–333. [Google Scholar] [CrossRef]
- Adir, N.; Bar-Zvi, S.; Harris, D. The amazing phycobilisome. Biochim. Biophys. Acta. 2020, 1861, 148047. [Google Scholar] [CrossRef]
- Harris, D.; Toporik, H.; Schlau-Cohen, G.S.; Mazor, Y. Energetic robustness to large scale structural fluctuations in a photosynthetic supercomplex. Nat. Commun. 2023, 14, 4650. [Google Scholar] [CrossRef] [PubMed]
- Umena, Y.; Kawakami, K.; Shen, J.-R.; Kamiya, N. Crystal structure of oxygen-evolving photosystem II at a resolution of 1.9 Å. Nature 2011, 473, 55–60. [Google Scholar] [CrossRef]
- La Roche, J.; Van der Staay, G.W.M.; Partensky, F.; Ducret, A.; Aebersold, R.; Li, R.; Golden, S. S.; Hiller, R. G.; Wrench, P. M.; Larkum, A.W.; Green, B.R. Independent evolution of the prochlorophyte and green plant chlorophyll a/b light-harvesting proteins. Proc. Natl. Acad. Sci. USA. 1996, 93, 15244–15248. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Semchonok, D.A.; Webber-Birungi, M.T.; Ehira, S.; Kondo, K.; Narikawa, R.; Ohmori, M.; Boekema, E.J.; Ikeuchi, M. Attachment of phycobilisomes in an antenna–photosystem I supercomplex of cyanobacteria. Proc. Natl. Acad. Sci. USA. 2014, 111, 2512–2517. [Google Scholar] [CrossRef] [PubMed]
- You, X.; Zhang, X.; Cheng, J.; Xiao, Y.; Ma, J.; Sun, S.; Zhang, X.; Wang, H.W.; Sui, S.F. In situ structure of the red algal phycobilisome–PSII–PSI–LHC megacomplex. Nature 2023, 616, 199–206. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, J.; Liu, D.; Qin, S.; Sun, S.; Zhao, J.; Sui, S.-F. Structure of phycobilisome from the red alga Griffitshia pacifica. Nature 2017, 551, 57–63. [Google Scholar] [CrossRef]
- Boekema, E.J.; Hifney, A.; Yakushevska, A.E.; Piotrowski, M.; Keegstra, W.; Berry, S.; Michel, K.P.; Pistorius, E.K.; Kruip, J. A giant chlorophyll-protein complex induced by iron deficiency in cyanobacteria. Nature 2001, 412, 745–748. [Google Scholar] [CrossRef]
- Bibby, T.S.; Nield, J.; Barber, J. Iron defciency induces the formation of an antenna ring around trimeric photosystem I in cyanobacteria. Nature 2001, 412, 743–745. [Google Scholar] [CrossRef]
- Melkozernov, A.N.; Bibby, T.S.; Lin, S.; Barber, J.; Blankenship, R.E. Time-resolved absorption and emission show that the CP43′ antenna ring of iron-stressed Synechocystis sp. PCC6803 is efficiently coupled to the Photosystem I reaction center core. Biochemistry 2003, 42, 3893–3903. [Google Scholar] [CrossRef]
- Andrizhiyevskaya, E.G.; Frolov, D.; Van Grondelle, R.; Dekker, J.P. Energy transfer and trapping in the Photosystem I complex of Synechococcus PCC 7942 and in its supercomplex with IsiA. Biochim. Biophys. Acta. 2004, 1656, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Ryan-Keogh, T.J.; Macey, A.I.; Cockshutt, A.M.; Moore, C.M.; Bibby, T.S. The cyanobacterial chlorophyll-binding-protein isiA acts to increase the in vivo effective absorption cross-section of PSI under iron limitation. J Phycol. 2011, 48, 145–154. [Google Scholar] [CrossRef]
- Sandström, S.; Park, Y.-I.; Öquist, G.; Gustafsson, P. CP43′, the IsiA gene product, functions as an excitation energy dissipator in the cyanobacterium Synechococcus Sp. PCC 7942. Photochem. Photobiol. 2007, 74, 431–437. [Google Scholar] [CrossRef]
- Ihalainen, J.A.; D’Haene, S.; Yeremenko, N.; van Roon, H.; Arteni, A.A.; Boekema, E.J.; van Grondelle, R.; Matthijs, H.C.P.; Dekker, J.P. Aggregates of the chlorophyll-binding protein IsiA (CP43′ ) dissipate energy in cyanobacteria. Biochemistry 2005, 44, 10846–10853. [Google Scholar] [CrossRef] [PubMed]
- Yeremenko, N.; Kouřil, R.; Ihalainen, J.A.; D'haene, S.; Van Oosterwijk, N.; Andrizhiyevskaya, E.G.; Keegstra, W.; Dekker, H.L.; Hagemann, M.; Boekema, E.J.; et al. Supramolecular organization and dual function of the isia chlorophyll-binding protein in cyanobacteria. Biochemistry 2004, 43, 10308–10313. [Google Scholar] [CrossRef] [PubMed]
- Goñi, G.; Herguedas, B.; Hervás, M.; Peregrina, J.R.; De La Rosa, M.A.; Gómez-Moreno, C.; Navarro, J.A.; Hermoso, J.A.; Martínez-Júlvez, M.; Medina, M. Flavodoxin: A compromise between efficiency and versatility in the electron transfer from Photosystem I to Ferredoxin-NADP+ reductase. Biochim. Biophys. Acta. 2009, 1787, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Deng, J. Photosynthetic pigments: A study of photosynthetic pigments; John McCrae Secondary School, 2018. [Google Scholar]
- Chen, M.; Schliep, M.; Willows, R.D.; Cai, Z.L.; Neilan, B.A.; Scheer, H. A red-shifted chlorophyll. Science 2010, 329, 1318–1319. [Google Scholar] [CrossRef] [PubMed]
- Melkozernov, A.N.; Blankenship, R.E. Photosynthetic Functions of Chlorophylls. In Chlorophylls and Bacteriochlorophylls. Advances in Photosynthesis and Respiration; Grimm, B., Porra, R.J., Rüdiger, W., Scheer, H., Eds.; Springer: Dordrecht, 2006; pp. 397–412. ISBN 978-1-4020-4516-5. [Google Scholar]
- Motten, A.F. Diversity of Photosynthetic Pigments. in: Tested studies for laboratory teaching, Volume 16 (C. A. Goldman, Editor). Proceedings of the 16thWorkshop/Conference of the Association for Biology Laboratory Education (ABLE), 1995; pp. 81–98.
- Gisriel, C.J. Recent structural discoveries of photosystems I and II acclimated to absorb far-red light. Biochim. Biophys. Acta. 2024, 1865, 149032. [Google Scholar] [CrossRef] [PubMed]
- Büchel, C. Light harvesting complexes in chlorophyll c-containing algae. Biochim. Biophys. Acta. 2020, 1861, 148027. [Google Scholar] [CrossRef]
- Croce, R.; van Amerongen, H. Light harvesting in oxygenic photosynthesis: Structural biology meets spectroscopy. Science 2020, 369, eaay2058. [Google Scholar] [CrossRef]
- Gan, F.; Zhang, S.; Rockwell, N.C.; Martin, S.S.; Lagarias, J.C.; Bryant, D.A. Extensive remodeling of a cyanobacterial photosynthetic apparatus in far-red light. Science 2014, 345, 1312–1317. [Google Scholar] [CrossRef]
- Airs, R.L.; Temperton, B.; Sambles, C.; Farnham, G.; Skill, S.C.; Llewellyn, C.A. Chlorophyll f and chlorophyll d are produced in the cyanobacterium Chlorogloeopsis fritschii when cultured under natural light and near–infrared radiation. FEBS Lett. 2014, 588, 3770–3777. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, L.; Brejnrod, A.; Schliep, M.; Sørensen, S.J.; Larkum, A.W.; Kühl, M. Chlorophyll f-driven photosynthesis in a cavernous cyanobacterium. ISME J. 2015, 9, 2108–2111. [Google Scholar] [CrossRef]
- Miyashita, H.; Ikemoto, H.; Kurano, N.; Adachi, K.; Chihara, M.; Miyachi, S. Chlorophyll d as a major pigment. Nature 1996, 383, 402. [Google Scholar] [CrossRef]
- Miyashita, H.; Ohkubo, S.; Komatsu, H.; Sorimachi, Y.; Fukayama, D.; Fujinuma, D.; Akutsu, S.; Kobayashi, M. Discovery of chlorophyll d in Acaryochloris marina and chlorophyll f in a unicellular cyanobacterium, Strain KC1, isolated from Lake Biwa. J Phys Chem Biophys 2014, 4, 149. [Google Scholar] [CrossRef]
- Loughlin, P.; Lin, Y.; Chen, M. Chlorophyll d and Acaryochloris marina: current status. Photosynth. Res. 2013, 116, 277–293. [Google Scholar] [CrossRef]
- Hamaguchi, T.; Kawakami, K.; Shinzawa-Itoh, K.; Inoue-Kashino, N.; Itoh, S.; Ifuku, K.; Yamashita, E.; Maeda, K.; Yonekura, K.; Kashino, Y. Structure of the far-red light utilizing photosystem I of Acaryochloris marina. Nat. Commun. 2021, 12, 2333. [Google Scholar] [CrossRef]
- Xu, C.; Zhu, Q.; Chen, J.; Shen, L.; Yi, X.; Huang, Z.; Wang, W.; Chen, M.; Kuang, T.; Shen, J.; et al. A unique photosystem I reaction center from a chlorophyll d-containing cyanobacterium Acaryochloris marina. J. Integr. Plant Biol. 2021, 63, 1740–1752. [Google Scholar] [CrossRef]
- Chen, M.; Li, Y.; Birch, D.; Willows, R.D. A cyanobacterium that contains chlorophyll f–a red-absorbing photopigment. FEBS Lett. 2012, 586, 3249–3254. [Google Scholar] [CrossRef]
- Kühl, M.; Trampe, E.; Mosshammer, M.; Johnson, M.; Larkum, A.W.; Frigaard, N.U.; Koren, K. Substantial near-infrared radiation-driven photosynthesis of chlorophyll f-containing cyanobacteria in a natural habitat. eLife 2020, 9, e50871. [Google Scholar] [CrossRef]
- Shen, G.; Canniffe, D.P.; Ho, M.Y.; Kurashov, V.; van der Est, A.; Golbeck, J.H.; Bryant, D.A. Characterization of chlorophyll f synthase heterologously produced in Synechococcus sp. PCC 7002. Photosynth. Res. 2019, 140, 77–92. [Google Scholar] [CrossRef]
- Nürnberg, D.J.; Morton, J.; Santabarbara, S.; Telfer, A.; Joliot, P.; Antonaru, L.A.; Ruban, A.V.; Cardona, T.; Krausz, E.; Boussac, A.; Fantuzzi, A.; Rutherford, A.W. Photochemistry beyond the red limit in chlorophyll f-containing photosystems. Science 2018, 360, 1210–1213. [Google Scholar] [CrossRef] [PubMed]
- Gisriel, C.; Shen, G.; Kurashov, V.; Ho, M.-Y.; Zhang, S.; Williams, D.; Golbeck, J.H.; Fromme, P.; Bryant, D.A. The structure of Photosystem I acclimated to far-red light illuminates an ecologically important acclimation process in photosynthesis. Sci. Adv. 2020, 6, eaay6415. [Google Scholar] [CrossRef]
- Kato, K.; Shinoda, T.; Nagao, R.; Akimoto, S.; Suzuki, T.; Dohmae, N.; Chen, M.; Allakhverdiev, S.I.; Shen, J.-R.; Akita, F.; et al. Structural basis for the adaptation and function of chlorophyll f in photosystem I. Nat. Commun. 2020, 11, 238. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.-Y.; Shen, G.; Canniffe, D.P.; Zhao, C.; Bryant, D.A. Light-dependent chlorophyll f synthase is a highly divergent paralog of PsbA of photosystem II. Science 2016, 353, aaf9178. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Gan, F.; Shen, G.; Bryant, D.A. RfpA, RfpB, and RfpC are the master control elements of far-red light photoacclimation (FaRLiP). Front. Microbiol. 2015, 6, 1303. [Google Scholar] [CrossRef]
- Chen, M.; Hernandez-Prieto, M.A.; Loughlin, P.C.; Li, Y.; Willows, R.D. Genome and proteome of the chlorophyll f-producing cyanobacterium Halomicronema hongdechloris: adaptative proteomic shifts under different light conditions. BMC Genomics. 2019, 20, 207. [Google Scholar] [CrossRef]
- Dupuis, S.; Merchant, S.S. Chlamydomonas reinhardtii: a model for photosynthesis and so much more. Nat Methods 2023, 20, 1441–1442. [Google Scholar] [CrossRef]
- Qin, X.; Suga, M.; Kuang, T.; Shen, J.-R. Structural basis for energy transfer pathways in the plant PSI-LHCI supercomplex. Science 2015, 348, 989–995. [Google Scholar] [CrossRef]
- Suga, M.; Ozawa, S.-I.; Yoshida-Motomura, K.; Akita, F.; Miyazaki, N.; Takahashi, Y. Structure of the green algal photosystem I supercomplex with a decameric light-harvesting complex I. Nat. Plants 2019, 5, 626–636. [Google Scholar] [CrossRef]
- Mullineaux, C.W. State transitions: an example of acclimation to low-light stress. J. Exp. Botany 2004, 56, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.F. Protein phosphorylation in regulation of photosynthesis. Biochim. Biophys. Acta. 1992, 1098, 275–335. [Google Scholar] [CrossRef] [PubMed]
- Nawrocki, W.J.; Santabarbara, S.; Mosebach, L.; Wollman, F.-A.; Rappaport, F. State transitions redistribute rather than dissipate energy between the two photosystems in Chlamydomonas. Nat. Plants 2016, 2, 16031. [Google Scholar] [CrossRef] [PubMed]
- Bellafiore, S.; Barneche, F.; Peltier, G.; Rochaix, J.-D. State transitions and light adaptation require chloroplast thylakoid protein kinase STN7. Nature 2005, 433, 892–895. [Google Scholar] [CrossRef] [PubMed]
- Wollman, F.A. State-transitions-reveal-the-dynamics-and-flexibility-of-the-photosynthetic-apparatus. EMBO J. 2001, 20, 3623–3630. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Chen, S.; Wang, C.; Lin, W.; Huang, C.; Fan, C.; Han, D.; Lu, D.; Xu, X.; Sui, S.; Zhang, L. Regulatory dynamics of the higher-plant PSI–LHCI supercomplex during state transitions. Mol. Plant. 2023, 16, 1937–1950. [Google Scholar] [CrossRef] [PubMed]
- Shang, H.; Li, M.; Pan, X. Dynamic regulation of the light-harvesting system through state transitions in land plants and green algae. Plants 2023, 2023. 12, 1173. [Google Scholar] [CrossRef]
- Huang, Z.; Shen, L.; Wang, W.; Mao, Z.; Yi, X.; Kuang, T.; Shen, J.-R.; Zhang, X.; Han, G. Structure of photosystem I-LHCI-LHCII from the green alga Chlamydomonas reinhardtii in State 2. Nat. Commun. 2021, 12, 1100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Tang, K.; Yan, Q.; Li, X.; Shen, L.; Wang, W.; He, Y.-K.; Kuang, T.; Han, G.; Shen, J.R.; Zhang, X. Structural insights into a unique PSI–LHCI–LHCII–Lhcb9 supercomplex from moss Physcomitrium patens. Nat. Plants 2023, 9, 832–846. [Google Scholar] [CrossRef]
- Lima-Melo, Y.; Kiliç, M.; Aro, E.-M.; Gollan, P.J. Photosystem I inhibition, protection and signalling: knowns and unknowns. Front. Plant Sci. 2021, 12, 791124. [Google Scholar] [CrossRef]
- Zavafer, A.; Mancilla, C. Concepts of photochemical damage of Photosystem II and the role of excessive excitation. J Photoch. Photobio. C. 2021, 47, 100421. [Google Scholar] [CrossRef]
- Su, J.; Jiao, Q.; Jia, T.; Hu, X. The photosystem-II repair cycle: updates and open questions. Planta 2023, 259, 20. [Google Scholar] [CrossRef]
- Naschberger, A.; Mosebach, L.; Tobiasson, V.; Kuhlgert, S.; Scholz, M.; Perez-Boerema, A.; Ho, T.T.H.; Vidal-Meireles, A.; Takahashi, Y.; Hippler, M.; et al. Algal photosystem I dimer and high-resolution model of PSI-plastocyanin complex. Nat. Plants 2022, 8, 1191–1201. [Google Scholar] [CrossRef]
- Su, X.; Ma, J.; Pan, X.; Zhao, X.; Chang, W.; Liu, Z.; Zhang, X.; Li, M. Antenna arrangement and energy transfer pathways of a green algal photosystem-I–LHCI supercomplex. Nat. Plants 2019, 5, 273–281. [Google Scholar] [CrossRef]
- Pi, X.; Tian, L.; Dai, H.-E.; Qin, X.; Cheng, L.; Kuang, T.; Sui, S.-F.; Shen, J.-R. Unique organization of photosystem I–light-harvesting supercomplex revealed by Cryo-EM from a red alga. Proc. Natl. Acad. Sci. USA. 2018, 115, 4423–4428. [Google Scholar] [CrossRef]
- Chang, L.; Tian, L.; Ma, F.; Mao, Z.; Liu, X.; Han, G.; Wang, W.; Yang, Y.; Kuang, T.; Pan, J.; et al. Regulation of photosystem I-light-harvesting complex I from a red alga Cyanidioschyzon Merolae in response to light intensities. Photosynth. Res. 2020, 146, 287–297. [Google Scholar] [CrossRef]
- Qin, X.; Pi, X.; Wang, W.; Han, G.; Zhu, L.; Liu, M.; Cheng, L.; Shen, J.-R.; Kuang, T.; Sui, S.-F. Structure of a green algal photosystem I in complex with a large number of light-harvesting complex I subunits. Nat. Plants 2019, 5, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Perez-Boerema, A.; Klaiman, D.; Caspy, I.; Netzer-El, S.Y.; Amunts, A.; Nelson, N. Structure of a minimal photosystem I from the green alga Dunaliella salina. Nat. Plants 2020, 6, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Mazor, Y.; Borovikova, A.; Caspy, I.; Nelson, N. Structure of the plant photosystem I supercomplex at 2.6 Å resolution. Nat. Plants 2017, 3, 17014. [Google Scholar] [CrossRef] [PubMed]
- Treves, H.; Raanan, H.; Kedem, I.; Murik, O.; Keren, N.; Zer, H.; Berkowicz, S.M.; Giordano, M.; Norici, A.; Shotland, Y.; et al. The mechanisms whereby the green alga Chlorella ohadii, isolated from desert soil crust, exhibits unparalleled photodamage resistance. New Phytol. 2016, 210, 1229–1243. [Google Scholar] [CrossRef]
- Kedem, I.; Milrad, Y.; Kaplan, A.; Yacoby, I. Juggling lightning: how Chlorella ohadii handles extreme energy inputs without damage. Photosynth. Res. 2021, 147, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Caspy, I.; Neumann, E.; Fadeeva, M.; Liveanu, V.; Savitsky, A.; Frank, A.; Kalisman, Y.L.; Shkolnisky, Y.; Murik, O.; Treves, H.; et al. Cryo-EM photosystem I structure reveals adaptation mechanisms to extreme high light in Chlorella ohadii. Nat. Plants 2021, 7, 1314–1322. [Google Scholar] [CrossRef] [PubMed]
- Malviya, S.; Scalco, E.; Audic, S.; Vincent, F.; Veluchamy, A.; Poulain, J.; Wincker, P.; Iudicone, D.; De Vargas, C.; Bittner, L.; Zingone, A.; Bowler, C. Insights into global diatom distribution and diversity in the world’s ocean. Proc. Natl. Acad. Sci. USA 2016, 113, 1516–1525. [Google Scholar] [CrossRef] [PubMed]
- Büchel, C. Light-Harvesting Complexes of Diatoms: Fucoxanthin-Chlorophyll Proteins. In Photosynthesis in Algae: Biochemical and Physiological Mechanisms. Advances in Photosynthesis and Respiration; Larkum, A., Grossman, A., Raven, J., Eds.; Springer: Cham, Switzerland, 2020; pp. 441–457. ISBN 978-3-030-33397-3. [Google Scholar]
- Xu, C.; Pi, X.; Huang, Y.; Han, G.; Chen, X.; Qin, X.; Huang, G.; Zhao, S.; Yang, Y.; Kuang, T.; et al. Structural basis for energy transfer in a huge diatom PSI-FCPI supercomplex. Nat. Commun. 2020, 11, 5081. [Google Scholar] [CrossRef]
- Löffelhardt, W.; Bohnert, H.J.; Bryant, D.A.; Hagemann, R. The cyanelles of Cyanophora paradoxa. Crit. Rev. Plant Sci. 1997, 16, 393–413. [Google Scholar] [CrossRef]
- Kato, K.; Nagao, R.; Ueno, Y.; Yokono, M.; Suzuki, T.; Jiang, T.-Y.; Dohmae, N.; Akita, F.; Akimoto, S.; Miyazaki, N.; et al. Structure of a tetrameric photosystem I from a glaucophyte alga Cyanophora paradoxa. Nat. Commun. 2022, 13, 1679. [Google Scholar] [CrossRef] [PubMed]
- Stiller, J.W.; Schreiber, J.; Yue, J.; Guo, H.; Ding, Q.; Huang, J. The evolution of photosynthesis in chromist algae through serial endosymbioses. Nat. Commun. 2014, 5, 5764. [Google Scholar] [CrossRef]
- Zimorski, V.; Ku, C.; Martin,W. F.; Gould, S.B. Endosymbiotic theory for organelle origins. Curr. Opin. Microbiol. 2014, 22, 38–48. [Google Scholar] [CrossRef]
- Kim, J.I.; Moore, C.E.; Archibald, J.M.; Bhattacharya, D.; Yi, G.; Yoon, H.S.; Shin, W. Evolutionary dynamics of cryptophyte plastid genomes. Genome Biol. Evol. 2017, 9, 1859–1872. [Google Scholar] [CrossRef]
- Hoffman, G.E.; Sanchez-Puerta, M.V.; Delwiche, C.F. Evolution of light harvesting complex proteins from Chl c-containing algae. BMC Evol. Biol. 2011, 11, 101. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, P.; Li, K.; Zhang, Q.; He, F.; Li, C.; Su, H.; Chen, X.; Liu, L.; Zhang, Y. Structural basis and evolution of the photosystem I–light-harvesting supercomplex of cryptophyte algae. Plant Cell 2023, 35, 2449–2463. [Google Scholar] [CrossRef] [PubMed]
- Cohen, N.R.; McIlvin, M.R.; Moran, D.M.; Held, N.A.; Saunders, J.K.; Hawco, N.J.; Brosnahan, M.; DiTullio, G.R.; Lamborg, C.; McCrow, J.P.; et al. Dinoflagellates alter their carbon and nutrient metabolic strategies across environmental gradients in the central Pacific Ocean. Nat Microbiol. 2021, 6, 173–86. [Google Scholar] [CrossRef]
- Stephens, T.G.; Ragan, M.A.; Bhattacharya, D.; Chan, C.X. Core genes in diverse dinoflagellate lineages include a wealth of conserved dark genes with unknown functions. Sci. Rep. 2018, 8, 17175. [Google Scholar] [CrossRef] [PubMed]
- Janouškovec, J.; Gavelis, G.S.; Burki, F.; Dinh, D.; Bachvaroff, T.R.; Gornik,S. G.; Bright,K. J.; Imanian, B.; Strom, S.L.; Delwiche, C.F.; et al. Major transitions in dinoflagellate evolution unveiled by phylotranscriptomics. Proc Natl Acad Sci USA. 2017, 114, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Aranda, M.; Li, Y.; Liew, Y.J.; Baumgarten, S.; Simakov, O.; Wilson, M.C.; Piel, J.; Ashoor, H.; Bougouffa, S.; Bajic, V.B.; et al. Genomes of coral dinoflagellate symbionts highlight evolutionary adaptations conducive to a symbiotic lifestyle. Sci. Rep. 2016, 6, 39734. [Google Scholar] [CrossRef]
- Jacobovitz, M.R.; Hambleton, E.A.; Guse, A. Unlocking the complex cell biology of coral–dinoflagellate symbiosis: a model systems approach. Annu. Rev. Genet. 2023, 57, 411–434. [Google Scholar] [CrossRef] [PubMed]
- Hehenberger, E.; Burki, F.; Kolisko, M.; Keeling, P.J. Functional relationship between a dinoflagellate host and its diatom endosymbiont. Mol. Biol. Evol. 2016, 33, 2376–2390. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Wu, S.; He, J.; Wang, X.; Grossman, A.R. Shining light on dinoflagellate photosystem I. Nat Commun, 2024, 15, 3337. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, Z.; Wang, F.; Zhao, S.; Xu, C.; Mao, Z.; Duan, J.; Feng, Y.; Yang, Y.; Shen, L.; et al. Structures and organizations of PSI–AcpPCI supercomplexes from red tidal and coral symbiotic photosynthetic dinoflagellates. Proc. Natl. Acad. Sci. USA 2024, 121, e2315476121. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, N.; Li, K.; Li, C.; Guo, J.; He, F.; Liu, G.; Chen, X.; Gao, J.; Liu, L.; et al. Architecture of symbiotic dinoflagellate photosystem I–light-harvesting supercomplex in Symbiodinium. Nat. Commun. 2024, 15, 2392. [Google Scholar] [CrossRef]
- Gisriel, C.; Sarrou, I.; Ferlez, B.; Golbeck, J.H.; Redding, K.E.; Fromme, R. Structure of a symmetric photosynthetic reaction center–photosystem. Science 2017, 357, 1021–1025. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-H.; Wu, H.; Xu, C.; Liu, X.-C.; Huang, Z.; Chang, S.; Wang, W.; Han, G.; Kuang, T.; Shen, J.-R.; et al. ,Architecture of the photosynthetic complex from a green sulfur bacterium. Science, 2020, 370, eabb6350. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Huang, G.; Wang, C.; Wang, J.; Sui, S.-F.; Qin, X. Structure of the Acidobacteria homodimeric reaction center bound with cytochrome c. Nat. Commun. 2022, 13, 7745. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structural diversity of cyanobacterial PSI. (a,b) The monomeric (PDB ID: 6LU1) and trimeric PSI complexes (PDB ID: 1JB0) from Thermosynechococcus elongatus share the similar subunit components; however, the PSI monomer lacks two peripheral subunits, PsaK and PsaX, which are colored in red. (c) The tetrameric PSI is mainly observed in heterocyst-forming cyanobacteria. A PSI tetramer (PDB ID: 6JEO) in Anabaena sp. PCC 7120 is organized with two PSI dimers with a C2 symmetry, forming two different interfaces between the neighboring PSI monomers.
Figure 1.
Structural diversity of cyanobacterial PSI. (a,b) The monomeric (PDB ID: 6LU1) and trimeric PSI complexes (PDB ID: 1JB0) from Thermosynechococcus elongatus share the similar subunit components; however, the PSI monomer lacks two peripheral subunits, PsaK and PsaX, which are colored in red. (c) The tetrameric PSI is mainly observed in heterocyst-forming cyanobacteria. A PSI tetramer (PDB ID: 6JEO) in Anabaena sp. PCC 7120 is organized with two PSI dimers with a C2 symmetry, forming two different interfaces between the neighboring PSI monomers.
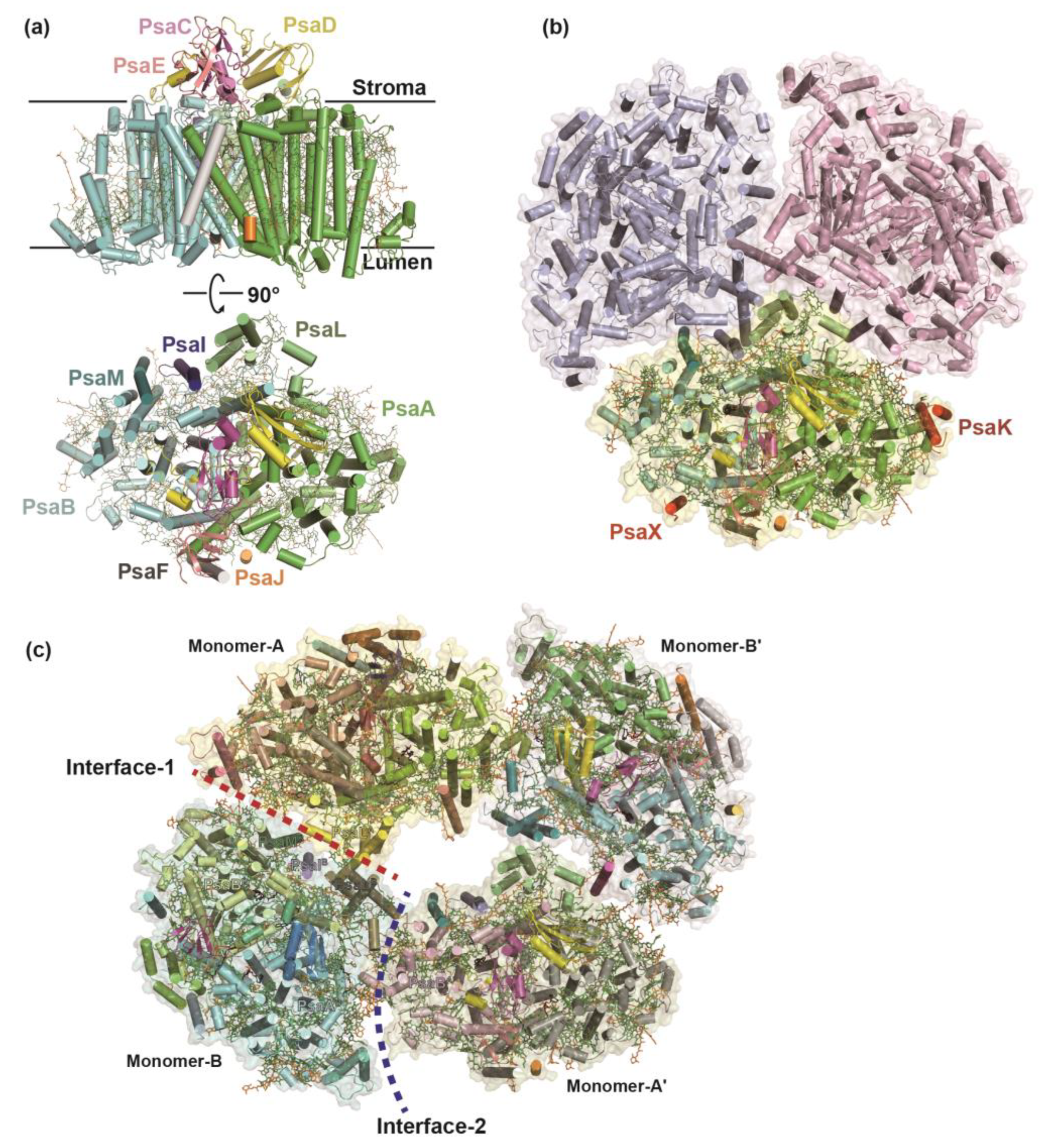
Figure 2.
Two structures of cyanobacterial PSI–IsiA complexes under iron-deficient condition. (a) The mesophilic cyanobacterium Synechocystis sp. PCC 6803 forms PSI3–IsiA18 supercomplex (PDB ID: 6K33) with threefold rotational symmetry. (b) The monomer–PSI–IsiA6 complex of Anabaena sp. PCC 7120 (PDB ID: 7Y3F) associates six IsiA subunits. The C–terminal PsaL–like domain of IsiA2-1 occupies the corresponding position of PsaL in the Anabaena PSI tetramer.
Figure 2.
Two structures of cyanobacterial PSI–IsiA complexes under iron-deficient condition. (a) The mesophilic cyanobacterium Synechocystis sp. PCC 6803 forms PSI3–IsiA18 supercomplex (PDB ID: 6K33) with threefold rotational symmetry. (b) The monomer–PSI–IsiA6 complex of Anabaena sp. PCC 7120 (PDB ID: 7Y3F) associates six IsiA subunits. The C–terminal PsaL–like domain of IsiA2-1 occupies the corresponding position of PsaL in the Anabaena PSI tetramer.
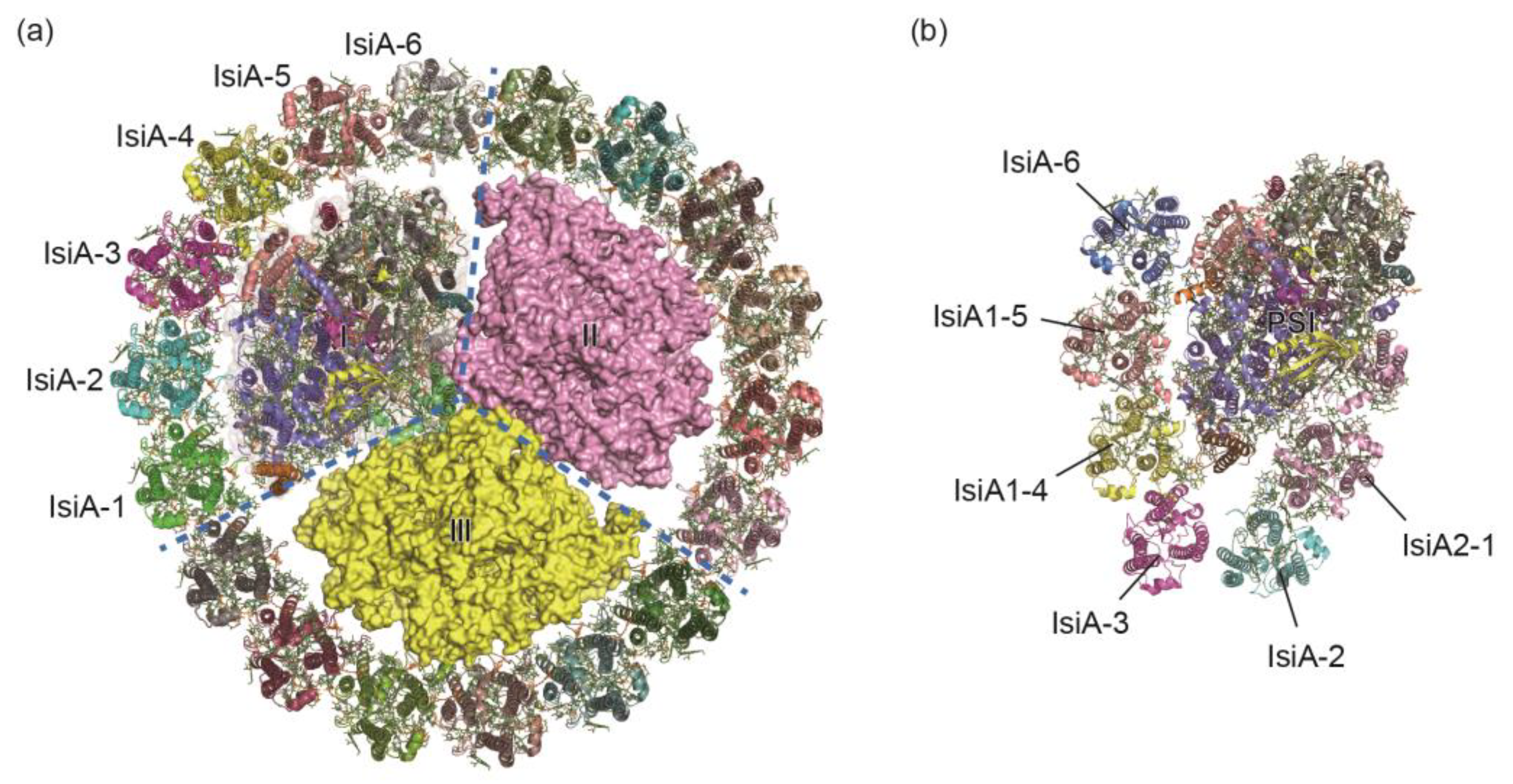
Figure 3.
The cofactors arranged along the electron transfer chain (ETC) of A. marina PSI. The cofactors of P740, Acc, A0, and A1 of the ETC are arranged in two separated branches (branch–A and branch–B). Phylloquinones and Fe4S4 clusters are labeled as PhyQ, FX, FA, and FB, respectively. Uniquely, the Chls of A0 in A. marina PSI are identified as two pheophytins a, instead of the typical Chls a in other cyanobacterial PSI complexes.
Figure 3.
The cofactors arranged along the electron transfer chain (ETC) of A. marina PSI. The cofactors of P740, Acc, A0, and A1 of the ETC are arranged in two separated branches (branch–A and branch–B). Phylloquinones and Fe4S4 clusters are labeled as PhyQ, FX, FA, and FB, respectively. Uniquely, the Chls of A0 in A. marina PSI are identified as two pheophytins a, instead of the typical Chls a in other cyanobacterial PSI complexes.
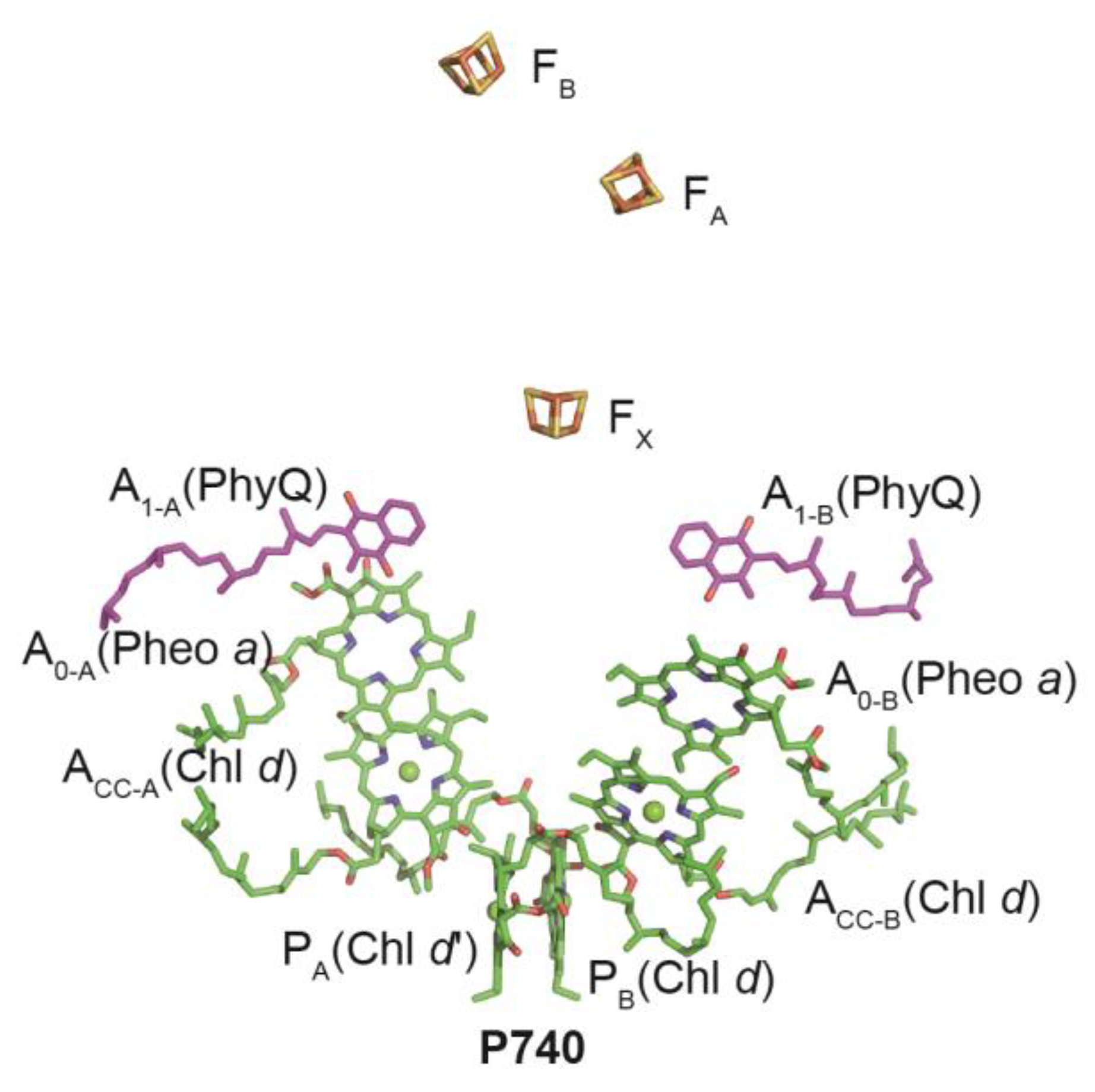
Figure 4.
Structural variations of C. reinhardtii PSI–LHCI complexes. (a) The structures of C. reinhardtii PSI–LHCI complexes binding with eight and ten Lhca subunits. The antenna subunits are distributed as three belts: two crescent–shaped belts including one inner belt and one outer belt which are associated to one side of the PSI core (PDB ID: 6JO6), and an additional small belt made up of the Lhca2 and Lhca9 heterodimer on the opposite side (PDB ID: 6JO5). (b) During the state transitions (state 2), phosphorylated LHCIIs dissociate from PSII and migrate to PSI, forming the PSI–LHCI–LHCII supercomplex (PDB ID: 7D0J). (c) Under low–light and anoxic conditions, PSI–LHCI dimer is formed two PSI–LHCI monomers that are arranged in a “head–to–head” manner. The PsaH and Lhca2 subunits are absent, instead, two Lhca9 subunits interact with the subunits of PsaI, PsaL, PsaG from two PSI–LHCI monomers (PDB ID: 7ZQD).
Figure 4.
Structural variations of C. reinhardtii PSI–LHCI complexes. (a) The structures of C. reinhardtii PSI–LHCI complexes binding with eight and ten Lhca subunits. The antenna subunits are distributed as three belts: two crescent–shaped belts including one inner belt and one outer belt which are associated to one side of the PSI core (PDB ID: 6JO6), and an additional small belt made up of the Lhca2 and Lhca9 heterodimer on the opposite side (PDB ID: 6JO5). (b) During the state transitions (state 2), phosphorylated LHCIIs dissociate from PSII and migrate to PSI, forming the PSI–LHCI–LHCII supercomplex (PDB ID: 7D0J). (c) Under low–light and anoxic conditions, PSI–LHCI dimer is formed two PSI–LHCI monomers that are arranged in a “head–to–head” manner. The PsaH and Lhca2 subunits are absent, instead, two Lhca9 subunits interact with the subunits of PsaI, PsaL, PsaG from two PSI–LHCI monomers (PDB ID: 7ZQD).
Figure 5.
Comparison of the PSI-LHCI complexes from different red algae. (a,b) C. merolae contains two different forms of PSI-LHCI complexes, with one binding three Lhcr subunits and the other one binding five Lhcr subunits. (c) Porphyridium purpureum PSI–LHCI associates seven LHCI subunits and one chlorophyll a/b-binding- like protein (RedCAP). (d) Cyanidium caldarium PSI-LHCI complex contains five LHCI subunits which are distributed as two separated clusters.
Figure 5.
Comparison of the PSI-LHCI complexes from different red algae. (a,b) C. merolae contains two different forms of PSI-LHCI complexes, with one binding three Lhcr subunits and the other one binding five Lhcr subunits. (c) Porphyridium purpureum PSI–LHCI associates seven LHCI subunits and one chlorophyll a/b-binding- like protein (RedCAP). (d) Cyanidium caldarium PSI-LHCI complex contains five LHCI subunits which are distributed as two separated clusters.
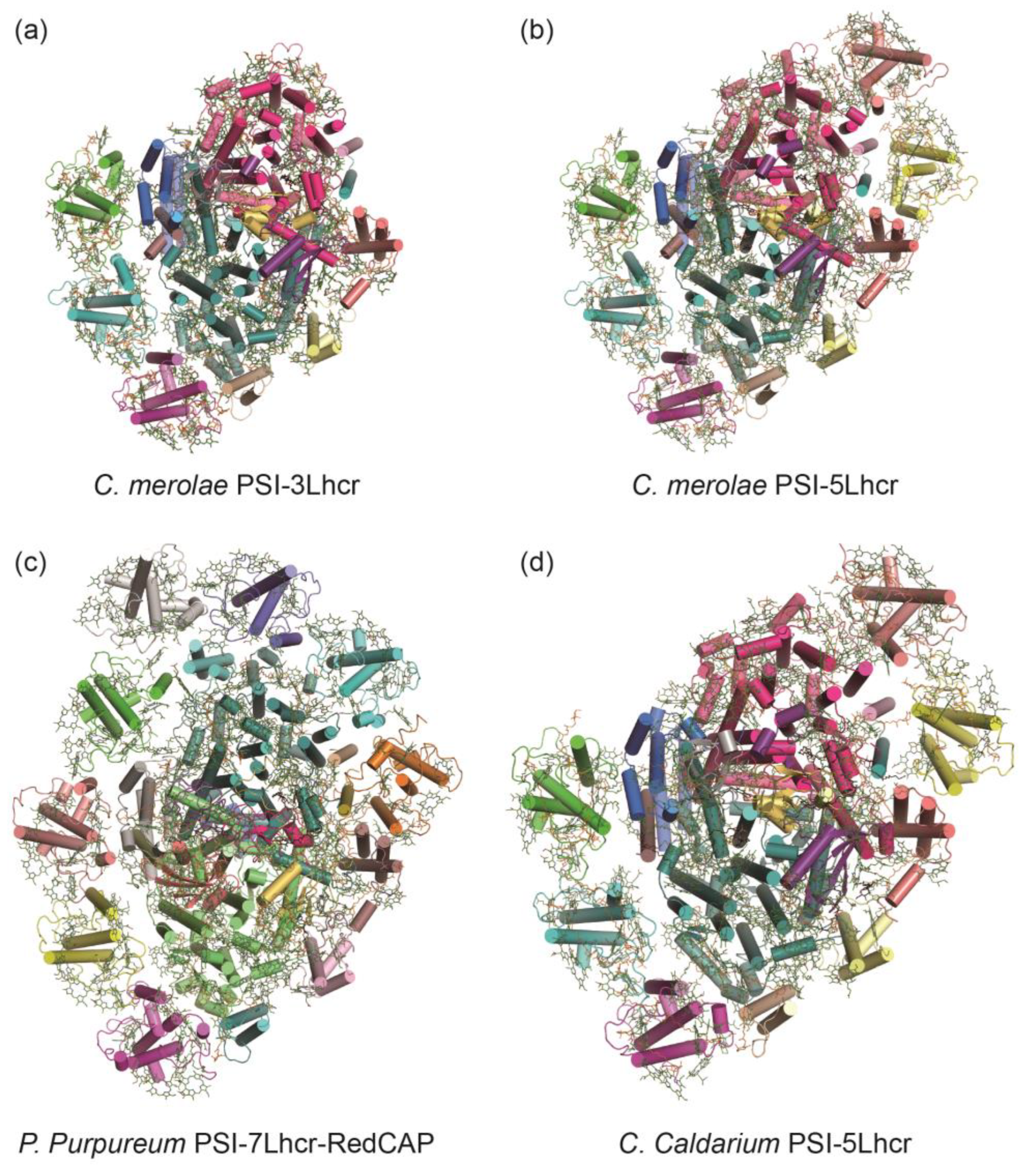
Figure 6.
Structures of PSI–LHCI complexes from green algae D. salina (a,b) and C. ohadii (c).
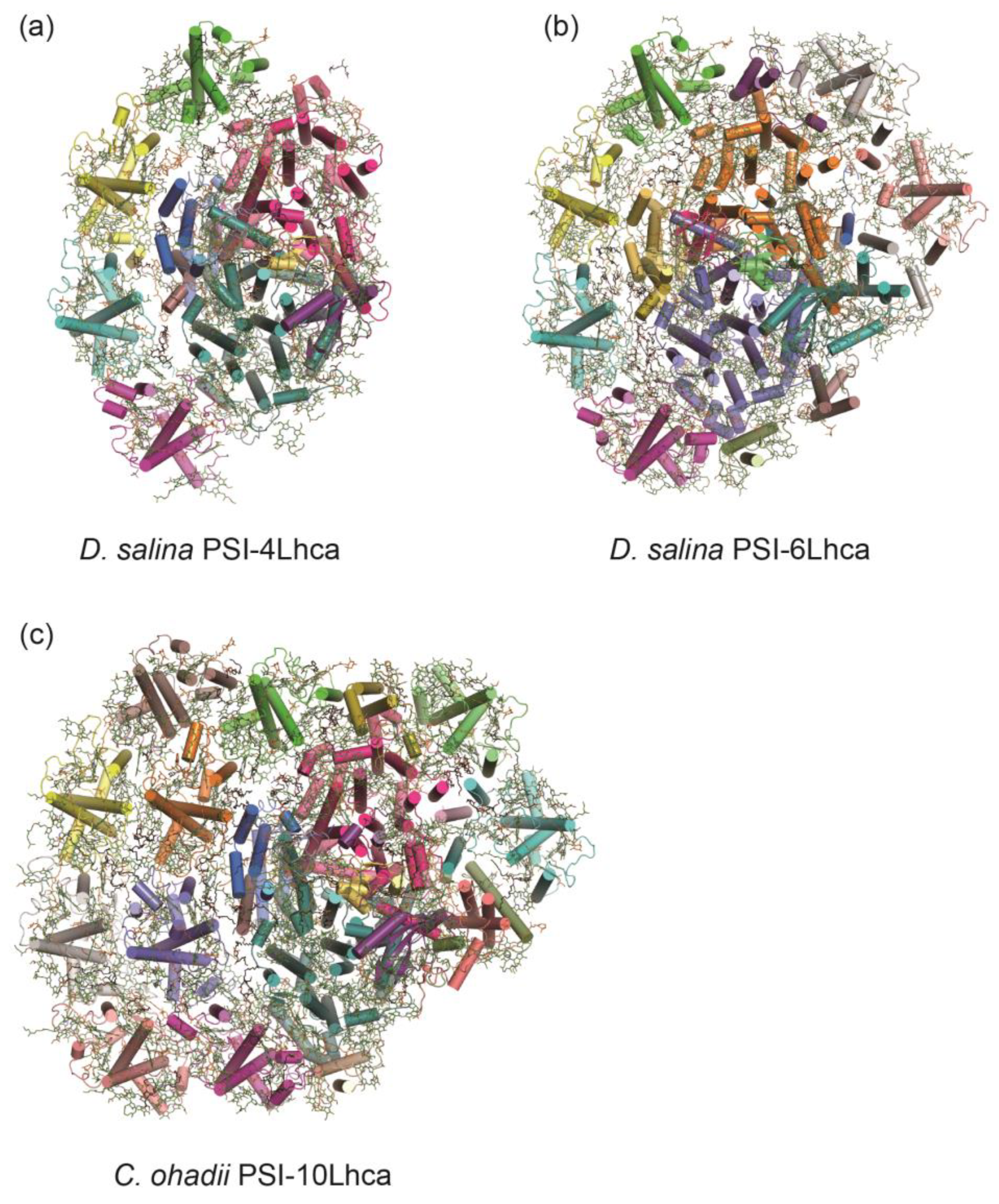
Figure 7.
Structures of PSI–FCPI complexes from diatom C. gracilis. Subunit-organization (a) and pigment network (b) of diatom C. gracilis PSI–FCPI complexes.
Figure 7.
Structures of PSI–FCPI complexes from diatom C. gracilis. Subunit-organization (a) and pigment network (b) of diatom C. gracilis PSI–FCPI complexes.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



