
Submitted:
01 July 2024
Posted:
03 July 2024
You are already at the latest version
Abstract
Keywords: Chronic Obstructive Pulmonary Disease (COPD), Long Non-Coding RNAs (lncRNAs), Molecular Pathogenesis, Inflammation, Lung Cancer, Diagnostic Biomarkers
Keywords:
Chronic Obstructive Pulmonary Disease (COPD)
; Long Non-Coding RNAs (lncRNAs)
; Molecular Pathogenesis
; Inflammation
; Lung Cancer
; Diagnostic Biomarkers
Introduction
COPD represents a formidable global health challenge, characterized by a pro-gressive and irreversible decline in airflow, caused by chronic inflammation, and other types of respiratory symptoms [1]. Principally associated with prolonged exposure to noxious particles, notably tobacco smoke, COPD imposes a substantial burden on individuals and healthcare systems worldwide [2]. Recent years have witnessed heightened scrutiny into the intricate molecular mechanisms underpinning COPD pathogenesis. Among the diverse array of factors contributing to its etiology, lncRNAs have emerged as pivotal regulators of gene expression and cellular processes [3]. Defined as RNA molecules exceeding 200 nucleotides in length with limited protein-coding potential, lncRNAs exert multifaceted effects on chromatin structure, transcriptional regulation, and post-transcriptional processes [4]. Dysregulated expression of lncRNAs in the context of COPD has been intricately linked to processes of inflammation, oxidative stress, and tissue remodeling, underscoring their potential as diagnostic and prognostic markers of disease progression [5].
The intricate interplay between COPD and lung cancer further complicates the clinical landscape, presenting overlapping risk factors such as tobacco smoke exposure and independently amplifying the risk of lung cancer development [6]. This intricate relationship involves a convergence of molecular pathways, creating a microenvironment conducive to malignant transformation [7]. The present review embarks on a comprehensive overview of COPD's multifaceted background, accentuating clinical challenges and the pressing need for deeper molecular insights. Central to this discourse is the pivotal role of lncRNAs in COPD pathophysiology and their potential as both biomarkers and modulators of key signaling cascades for disease progression [8]. Moreover, this review summarizes the intricate linkages between COPD and lung cancer progression, shedding light on common molecular signatures and their potential therapeutic interventions.
1. Chronological Progression of COPD
COPD is a complex and multifactorial respiratory condition characterized by persistent airflow limitation and respiratory symptoms, including cough, sputum production, and dyspnea [9]. Its progression often takes several decades and is marked by distinct stages, each contributing to the gradual deterioration of respiratory function [10]. This review delves into the mode and mechanism of the progression of COPD and explores current approaches to its diagnosis and monitoring.
1.1. Disease Progression in COPD
The pathogenesis of COPD comprises various stages, involving both structural and functional changes of the respiratory system. It typically occurs after prolonged exposure to noxious substances, such as cigarette smoke, that cause chronic inflammation, mucus hypersecretion, and alterations in ciliary function [11]. This initial insult triggers a cascade of events, including oxidative stress and protease-antiprotease imbalances, resulting in the destruction of lung parenchyma and shortness of breath (emphysema) [12].
As COPD advances, the airway is remodeled by elevating smooth muscle mass, fibrosis, and narrowing lumens. Chronic bronchitis may manifest with persistent cough and sputum production. The lung function, measured as the forced expiratory volume in one second (FEV1), gradually declines, marking a critical juncture of the disease progression [13]. In later stages, respiratory functions may deteriorate, contributing to worsened disease morbidity and mortality [14]. Furthermore, COPD-associated cachexia, manifested as weight loss and muscle dysfunction, exacerbates the overall impact of the disease [15]. Understanding the disease progression, as well as identification of individuals at risk and mitigation of exposure and inflammation, are essential for COPD management and prevention [16].
1.2. The Major Causative Factors of COPD
The development and progression of COPD are influenced by a myriad of factors, including genetic predisposition, environmental exposures, and individual lifestyle behaviors (Figure 1). Understanding the interplay between these factors is essential for elucidating the pathogenesis of COPD and developing effective prevention and management strategies [17]. We will introduce several major causative factors of COPD below.
Air Pollution: Outdoor and indoor air pollution represent significant risk factors for COPD [18]. Inhalation of fine particulate matter, including nitrogen dioxide, sulfur dioxide, ozone from vehicle emissions, industrial activities, and biomass combustion, increases the risk of respiratory symptoms such as COPD. Urbanization, industrialization, and climate change exacerbate air pollution levels, raising public concerns about COPD [19]. Implementation of clean air policies, sustainable transportation strategies, and new energy sources is critical for reducing air pollution and protecting respiratory health [20].
Environmental Exposures: Exposure to environmental pollutants, occupational hazards, and indoor air contaminants contributes to the development and exacerbation of COPD [21]. Inhalation of particulate matter, chemical fumes, and noxious gases from industrial processes, biomass burning, and indoor cooking fuels leads to airway inflammation, oxidative stress, and lung damage in susceptible individuals. Environmental regulations, workplace safety measures, and indoor air quality improvements are essential for reducing environmental exposures and mitigating the burden of COPD in the general population [22].
Cigarette Smoking: Cigarette smoking stands as the primary causative factor for the development of COPD. The inhalation of toxic substances present in cigarette smoke, such as tar, nicotine, and carbon monoxide, triggers inflammation and damage to the airways and lung parenchyma [23]. Chronic exposure to cigarette smoke leads to the progressive destruction of lung tissue, airflow limitation, and respiratory symptoms characteristic of COPD. Moreover, smoking cessation remains the most effective intervention to slow disease progression and reduce morbidity and mortality associated with COPD [24].
Gender: Gender differences play a significant role in the prevalence and clinical manifestations of COPD [25]. Historically, COPD has been more prevalent among males due to higher rates of smoking. However, recent studies have shown an increasing burden of COPD among females, reflecting changes in smoking behaviors as well as the occurrence of environmental irritants that preferentially afflict women [26].
Differences in Smoking Patterns: Historically, COPD has been more strongly associated with males due to higher rates of smoking among men. However, smoking patterns have changed over time, with an increasing number of females smoking cigarettes. In recent years, smoking prevalence among females has risen, contributing to a higher incidence of COPD in women [27].
Biological Differences: Biological factors play a significant role in the development and progression of COPD. Generally, females have narrower airways compared to males, which may contribute to increased susceptibility to COPD. Additionally, hormonal differences between males and females, such as estrogen levels, may influence inflammatory responses to pollutants [28].
Exposure to Indoor Air Pollutants: Women may be more exposed to indoor air pollutants such as biomass fuel smoke, cooking fumes, and secondhand smoke, particularly in regions where indoor cooking with solid fuels is common. Prolonged exposure to these pollutants can increase the risk of developing COPD [29].
Occupational Exposures: Certain occupational exposures, such as working in industries with high levels of airborne pollutants or dust, may disproportionately affect females and contribute to the development of COPD. For example, women working in women-dominant occupations such as house cleaning and textile manufacturing, may be exposed to respiratory hazards that increase their risk of COPD [30].
Socioeconomic Factors: Socioeconomic factors, including income level, education, and access to healthcare, influence the risk of COPD development and disease outcomes. Individuals from lower socioeconomic backgrounds are disproportionately affected by COPD due to higher rates of smoking, occupational exposures, and limited access to healthcare services [31:34]. Socioeconomic disparities contribute to delayed diagnosis, inadequate treatment, and poorer prognosis in COPD patients from disadvantaged communities [35]. Addressing socioeconomic inequalities is essential for reducing the burden of COPD among vulnerable populations [36].
Aging: Aging is a major risk factor for the development of COPD. Age-related lung structure and function changes, including decreased lung elasticity, reduced mucociliary clearance, and impaired immune responses, predispose older adults to COPD and exacerbations [37]. Moreover, cumulative exposure to environmental pollutants, respiratory infections, and other comorbidities further exacerbates the decline of lung functions and disease progression in elderly individuals. Comprehensive geriatric assessment and management strategies are essential for improving COPD care and outcomes in older patients [38].
Respiratory Infections: Respiratory infections, particularly viral and bacterial infections, contribute to the pathogenesis and exacerbations of COPD. Acute respiratory infections, such as influenza and pneumonia, trigger airway inflammation, mucous production, and exacerbation in COPD patients [39]. Moreover, recurrent respiratory infections accelerate the decline of lung functions and worsen the clinical outcomes of COPD patients. Vaccination and appropriate antimicrobial therapies play crucial roles in preventing and managing COPD [40].
Genetic Factors: Genetic predisposition plays a significant role in COPD susceptibility and disease heterogeneity [41]. Variations in genes involved in lung development, inflammation, and immunological defense influence susceptibility to COPD and response to environmental exposures, such as cigarette smoke and air pollution [42]. For example, alpha-1 antitrypsin deficiency, an inherited disorder, is a well-established genetic risk factor for early-onset COPD. Understanding the genetic basis of COPD facilitates early identification of at-risk individuals and personalized treatment based on genetic profiling [43].
1.3. Diagnosis and Monitoring of COPD
Accurate and timely diagnosis of COPD is fundamental for effective management and intervention. Diagnosis is typically established through a combination of clinical assessment, spirometry, and consideration of risk factors such as smoking history and environmental exposures [44]. Spirometry, measuring the FEV1 and the ratio of FEV1 to forced vital capacity (FVC), remains the gold standard for diagnosing airflow limitation [45]. Monitoring disease progression involves regular assessments of symptoms, exacerbation history, and lung function. The global initiative for chronic obstructive lung disease (GOLD) classification provides a framework for categorizing disease severity based on spirometric measurements [46]. However, it is increasingly recognized that a comprehensive assessment of COPD should extend beyond spirometry, encompassing symptoms, exacerbation risk, and impact on patient health status [47]. Novel approaches to COPD monitoring involve the exploration of biomarkers and imaging techniques to provide a more nuanced understanding of disease activity. Blood biomarkers, such as C-reactive protein (CRP) and fibrinogen, have shown promise in reflecting systemic inflammation and predicting exacerbation risk [48]. Imaging modalities, including computed tomography (CT) and magnetic resonance imaging (MRI), offer insights into structural lung changes and phenotypic variations, aiding in personalized therapeutic strategies [49].
2. Prognostic and Diagnostic Significance of LncRNA in COPD
LncRNAs have emerged as crucial regulators of gene expression, exerting significant influence on various biological processes [50]. Unlike their protein-coding counterparts, lncRNAs do not encode proteins but instead orchestrate diverse cellular functions through mechanisms such as chromatin remodeling, transcriptional regulation, and post-transcriptional processing [51]. In the context of COPD, the dysregulation of lncRNAs has been implicated in disease pathogenesis and progression, offering promising avenues for prognostic and diagnostic applications [52]. COPD is a multifactorial respiratory disorder characterized by persistent airflow limitation and progressive decline in lung function [53]. Despite advances in therapeutic strategies, COPD remains a leading cause of morbidity and mortality worldwide. The identification of reliable prognostic and diagnostic biomarkers is therefore imperative for early disease detection, prognostic assessment, and personalized treatment strategies [54]. In this regard, lncRNAs have garnered considerable attention as potential biomarkers due to their tissue-specific expression patterns, stability in various biological fluids, and functional relevance in COPD pathophysiology [55].
2.1. Identification and Profiling of COPD-Associated LncRNAs
The validated COPD-associated lncRNAs and lung cancer-associated lncRNAs have been thoroughly reviewed from the existing literature (Figure 2). These lncRNAs, along with miRNAs, are known to interact with mRNAs, thereby modulating the progression of COPD and/or cancer through competitive endogenous RNA (ceRNA) networks. the ceRNA hypothesis suggests that lncRNAs can act as molecular sponges for miRNAs, thus regulating the expression of miRNA target genes. This interaction framework opens new avenues for research into how lncRNAs influence COPD (Supplementary Excel files 1) and lung cancer, providing a deeper understanding of their roles in disease modulation and progression (Supplementary Excel files 2).
Identifying and profiling COPD-associated lncRNAs represents a critical step toward understanding the molecular mechanisms underlying the disease's pathogenesis and progression. Genome-wide expression profiling studies have consistently revealed dysregulated expression patterns of lncRNAs in COPD patients compared to healthy controls, underscoring their potential utility as diagnostic and prognostic biomarkers [56]. Such dysregulation highlights the importance of lncRNAs in the disease process and their potential role in the development of therapeutic strategies [56].
Recent advancements in high-throughput sequencing technologies and bioinformatics tools have facilitated the systematic characterization of lncRNA expression profiles in COPD [57]. Transcriptomic analyses utilizing next-generation sequencing platforms have enabled the comprehensive identification of novel lncRNAs and the elucidation of their regulatory networks in COPD pathophysiology [58].
One of the challenges in lncRNA profiling studies is the heterogeneity of COPD phenotypes and disease severity, which necessitates careful selection of patient cohorts and appropriate control groups for comparative analyses [59]. Integrative approaches combining transcriptomic data with clinical parameters, imaging findings, and functional assays are essential for prioritizing candidate lncRNAs and elucidating their biological significance in COPD [60]. A comprehensive understanding of the regulatory mechanisms governing lncRNA expression in COPD is crucial for deciphering their functional roles and clinical implications. Emerging evidence suggests that lncRNAs are subject to dynamic regulation by transcription factors, epigenetic modifiers, and signaling pathways implicated in COPD pathogenesis [61].
2.2. Master Regulators of Gene Expression in Lung Cancers Progression
LncRNAs are pivotal regulators of gene expression in lung cancer progression. They control gene expression through various mechanisms, including epigenetic modifications, competition with microRNAs (miRNAs), and direct interaction with mRNA transcripts [62]. Through these diverse pathways, lncRNAs exert precise control over gene expression programs implicated in lung cancer progression (Figure 3), underscoring their importance in the disease's molecular landscape [63].
Transcriptional Regulation: LncRNAs are known to play crucial roles in transcriptional regulation by influencing gene expression at the chromatin level [64]. One such example is the MALAT1, which has been extensively studied in the context of lung cancer metastasis. MALAT1 functions as a key regulator of gene transcription by interacting with transcription factors and chromatin-modifying proteins [65]. Through these interactions, MALAT1 modulates the expression of genes involved in various aspects of lung cancer progression, including metastasis. For instance, MALAT1 has been shown to promote the expression of metastasis-associated genes by facilitating chromatin remodeling and transcriptional activation [66].
Epigenetic Regulation: Epigenetic regulation mediated by lncRNAs is a fundamental mechanism governing gene expression in lung diseases. An illustrative example is the HOTAIR, which has been implicated in epigenetic modifications associated with lung cancer progression [67]. HOTAIR functions as a scaffold for chromatin-modifying complexes, such as polycomb repressive complex 2 (PRC2), leading to alterations in histone methylation patterns. Specifically, HOTAIR interacts with PRC2 to promote the deposition of repressive histone marks, such as histone H3 lysine 27 trimethylation (H3K27me3), at target gene promoters [68].
Post-transcriptional Regulation: Post-transcriptional regulation by lncRNAs plays a critical role in modulating gene expression levels and mRNA processing in lung diseases. A notable example is the GAS5, which functions as a competing endogenous RNA (ceRNA) in idiopathic pulmonary fibrosis (IPF) [69]. GAS5 has been shown to sequester miR-21, a known regulator of fibrosis-related genes, thereby relieving its inhibitory effect on target mRNAs. By acting as a molecular sponge for miR-21, GAS5 promotes the expression of its target gene PTEN (Phosphatase and Tensin Homolog), leading to decreased cell proliferation and fibrosis progression in IPF [70].
Signaling Pathway Regulation: LncRNAs are involved in the regulation of signaling pathways that govern cellular processes in lung diseases, offering potential targets for therapeutic intervention. An exemplary lncRNA in this context is H19, which modulates the transforming growth factor-beta (TGF-β) signaling pathway in lung cancer [71]. H19 has been shown to enhance TGF-β signaling by promoting the phosphorylation and nuclear translocation of Smad2/3, key mediators of the TGF-β pathway. This enhanced signaling cascade induces epithelial-mesenchymal transition (EMT) and promotes metastasis in lung cancer cells [72].
RNA-RNA Interactions: Interactions between lncRNAs and other RNA molecules contribute to the complex regulatory networks governing gene expression in lung diseases. A notable example is the lncRNA XIST, which interacts with the mRNA of the tumor suppressor gene RBM5 (RNA Binding Motif Protein 5) in lung cancer cells [73]. XIST-mediated sequestration of RBM5 mRNA leads to decreased protein expression levels, resulting in enhanced proliferation and survival of lung cancer cells [74]. This RNA-RNA interaction exemplifies the intricate crosstalk between lncRNAs and mRNAs, highlighting their role in shaping the molecular landscape of lung diseases.
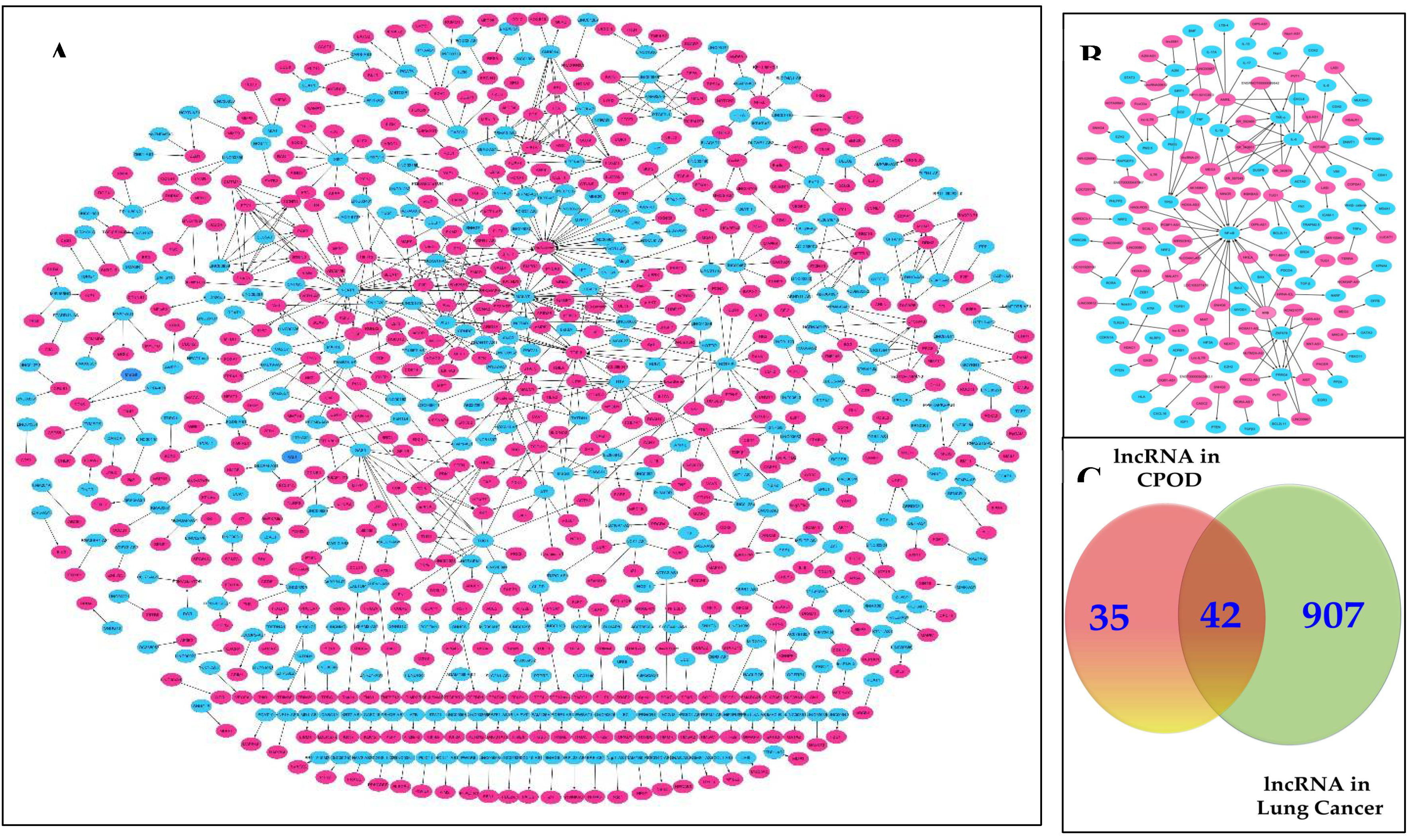
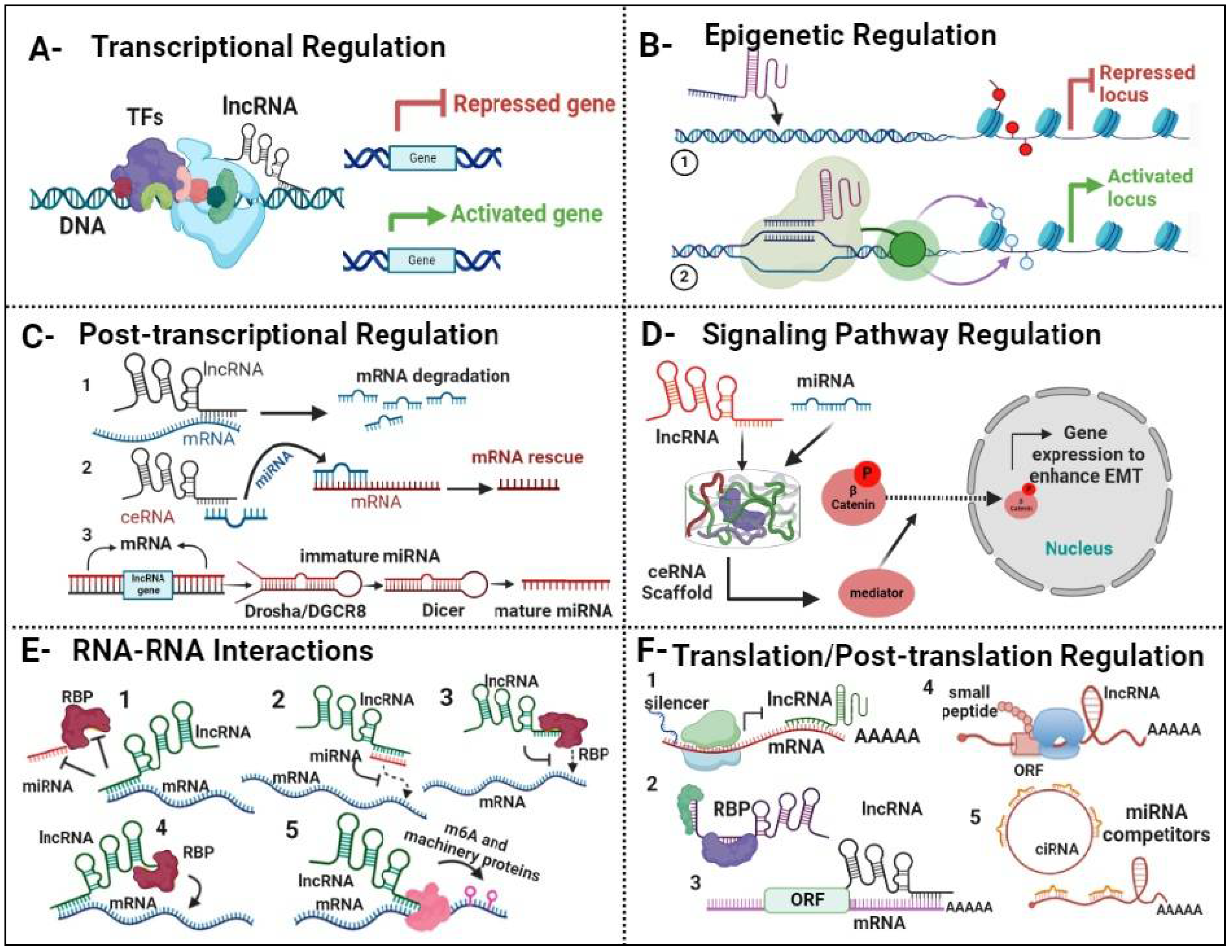
Figure 2.
Profiling of LncRNA in COPD and Lung Cancer. A. Representative Network of interacting lncRNAs/mRNAs in COPD This network elucidates the intricate relationships between these RNA molecules, shedding light on their regulatory roles and potential implications in COPD pathogenesis. Through systematic analysis, this representation aims to provide insights into the underlying molecular mechanisms driving COPD progression. B. Representative Network of interacting lncRNAs/mRNAs in lung cancer, this network visualization offers a comprehensive view of the intricate interactions between these RNA species, highlighting their regulatory roles and potential significance in the context of lung cancer. By mapping out these interactions, this representation aims to unravel the complex molecular mechanisms underlying lung cancer development and progression C. Venn Diagram to illustrate the shared lnRNAs between COPD and lung cancer, This visual representation showcases the overlapping lncRNA profiles between the two respiratory conditions, offering valuable insights into potential common molecular pathways and regulatory mechanisms, blue color indicate for lncRNAs and red color indicate for mRNAs.
Figure 2.
Profiling of LncRNA in COPD and Lung Cancer. A. Representative Network of interacting lncRNAs/mRNAs in COPD This network elucidates the intricate relationships between these RNA molecules, shedding light on their regulatory roles and potential implications in COPD pathogenesis. Through systematic analysis, this representation aims to provide insights into the underlying molecular mechanisms driving COPD progression. B. Representative Network of interacting lncRNAs/mRNAs in lung cancer, this network visualization offers a comprehensive view of the intricate interactions between these RNA species, highlighting their regulatory roles and potential significance in the context of lung cancer. By mapping out these interactions, this representation aims to unravel the complex molecular mechanisms underlying lung cancer development and progression C. Venn Diagram to illustrate the shared lnRNAs between COPD and lung cancer, This visual representation showcases the overlapping lncRNA profiles between the two respiratory conditions, offering valuable insights into potential common molecular pathways and regulatory mechanisms, blue color indicate for lncRNAs and red color indicate for mRNAs.
Figure 3.
LncRNA Regulators of Gene Expression in Lung Cancer. A. Transcriptional regulation: lncRNA interacts either with transcription factors (TFs) or directly with DNA, resulting in either repression or enhancement of gene expression. B. Epigenetic regulation: lncRNAs regulate histone proteins through both direct and indirect mechanisms, thereby influencing gene expression by either repressing or activating target genes. C. Post-Transcriptional regulation: 1. lncRNA) directly binds to mRNA, leading to its degradation (1), lncRNA can also act as a competitive inhibitor with miRNA, forming a scaffold that binds to mRNA, thereby rescuing it from degradation (2), Additionally, lncRNA plays a role in facilitating the maturation process of miRNA (3). D. Signaling Pathway regulation: lncRNA and miRNAs collaborate to create a scaffold complex, orchestrating the beta-catenin signaling pathway. This complex regulates gene expression to promote the epithelial-mesenchymal transition (EMT) process. E. RNA-RNA interaction: lncRNAs play a pivotal role in modulating mRNA stability through various mechanisms. These include sequestering miRNAs or RNA-binding proteins (RBPs) to prevent their interaction with mRNA molecules (1), directly binding to miRNAs (2) or RBPs (3), and binding to mRNA via RBPs (4). Additionally, lncRNA have the capacity to regulate RNA modification by interacting with m6A machinery proteins (5). F. Translation/post-translation regulation: Cytoplasmic LncRNA compete for ribosomal protein to mRNA to regulate expression by regulating mRNA stability (1), lncRNA binds to RBPs (2), mRNA translation (3), few lncRNA contain small open reading frames (ORFs) that can be translated in biological active small peptides (4). In addition, lncRNA competes for miRNA binding (5).
Figure 3.
LncRNA Regulators of Gene Expression in Lung Cancer. A. Transcriptional regulation: lncRNA interacts either with transcription factors (TFs) or directly with DNA, resulting in either repression or enhancement of gene expression. B. Epigenetic regulation: lncRNAs regulate histone proteins through both direct and indirect mechanisms, thereby influencing gene expression by either repressing or activating target genes. C. Post-Transcriptional regulation: 1. lncRNA) directly binds to mRNA, leading to its degradation (1), lncRNA can also act as a competitive inhibitor with miRNA, forming a scaffold that binds to mRNA, thereby rescuing it from degradation (2), Additionally, lncRNA plays a role in facilitating the maturation process of miRNA (3). D. Signaling Pathway regulation: lncRNA and miRNAs collaborate to create a scaffold complex, orchestrating the beta-catenin signaling pathway. This complex regulates gene expression to promote the epithelial-mesenchymal transition (EMT) process. E. RNA-RNA interaction: lncRNAs play a pivotal role in modulating mRNA stability through various mechanisms. These include sequestering miRNAs or RNA-binding proteins (RBPs) to prevent their interaction with mRNA molecules (1), directly binding to miRNAs (2) or RBPs (3), and binding to mRNA via RBPs (4). Additionally, lncRNA have the capacity to regulate RNA modification by interacting with m6A machinery proteins (5). F. Translation/post-translation regulation: Cytoplasmic LncRNA compete for ribosomal protein to mRNA to regulate expression by regulating mRNA stability (1), lncRNA binds to RBPs (2), mRNA translation (3), few lncRNA contain small open reading frames (ORFs) that can be translated in biological active small peptides (4). In addition, lncRNA competes for miRNA binding (5).
2.3. Tobacco Smoke Exposure and Expression Profiles of LncRNA in COPD
Exposure to tobacco smoke is a major risk factor for the development and progression of COPD, a complex and debilitating respiratory condition characterized by chronic inflammation, airway obstruction, and irreversible lung damage (Figure 4) [75]. Tobacco smoke contains thousands of toxic compounds, including carcinogens and reactive oxygen species, which can induce widespread genomic alterations and dysregulate various cellular processes in the lung microenvironment [76]. Emerging evidence suggests that exposure to tobacco smoke profoundly influences the landscape of lncRNAs in COPD, contributing to disease pathogenesis and progression. This reflects the complex interplay between environmental stimuli, genetic predisposition, and epigenetic modifications in disease pathophysiology [77]. High-throughput sequencing technologies and genome-wide expression profiling have facilitated the identification of tobacco smoke-responsive lncRNAs associated with COPD susceptibility, severity, and treatment outcomes. These dysregulated lncRNAs exhibit altered expression patterns in response to tobacco smoke exposure, modulating key cellular processes involved in inflammation, oxidative stress, and tissue remodeling in the lung [78].
Functional characterization of tobacco smoke-responsive lncRNAs provides in-sights into their roles as critical regulators of gene expression networks and signaling pathways implicated in COPD pathogenesis [79]. Tobacco smoke-induced lncRNAs, such as HOTAIR, MALAT1, and NEAT1, modulate inflammatory responses, and fibrotic processes by interacting with chromatin-modifying complexes, transcription factors, and RNA-binding proteins [80]. Dysregulated expression of these lncRNAs contributes to exacerbated airway inflammation, oxidative stress, and tissue damage in COPD patients [81]. Furthermore, tobacco smoke exposure alters the epigenetic landscape of the lung, leading to aberrant DNA methylation patterns, histone modifications, and chromatin remodeling events that regulate lncRNA expression and function in COPD [82]. Such epigenetic changes may reprogram the transcriptional profiles of lncRNAs involved in immune response, cell proliferation, and apoptosis, driving disease progression and exacerbation in COPD patients. Understanding the molecular mechanisms for the effects of tobacco smoke exposure on the landscape of lncRNAs in COPD is essential for identifying novel biomarkers and therapeutic targets for disease intervention and personalized treatment approaches [83].
The intricate landscape of lncRNAs has emerged as a crucial player in COPD pathogenesis, orchestrating diverse cellular processes implicated in disease onset and progression. In response to tobacco smoke exposure, aberrant expression patterns of lncRNAs contribute to mitochondrial dysfunction, chronic inflammation, mucus dysregulation, epigenetic alterations, and cellular senescence, which are key pathological hallmarks of COPD [84].
Mitochondrial dysfunction represents a central feature of COPD pathogenesis, contributing to oxidative stress, impaired energy metabolism, and cellular damage in the lung microenvironment [85]. This phenomenon is in part attributed to dysregulated expression of lncRNAs, such as HOTAIR and MALAT1, alters mitochondrial function and biogenesis. Through interactions with regulatory proteins and signaling pathways, a group of lncRNAs play pivotal roles in maintaining mitochondrial homeostasis and cellular resilience in response to tobacco smoke exposure [86].
Chronic inflammation is a hallmark of COPD, driven by dysregulated immune responses and pro-inflammatory pathways in the lung microenvironment. LncRNAs, including NEAT1 and GAS5, regulate inflammatory signaling and cytokine production for modulating immune cell function and tissue inflammation [87,88]. Dysregulated expression of these lncRNAs in COPD patients amplifies inflammatory responses and exacerbates tissue damage after being exposed to tobacco smoke, highlighting their potential as therapeutic targets.
Mucus dysregulation is a common feature of COPD, characterized by excessive mucus production, impaired clearance mechanisms, and airway obstruction [89]. LncRNAs, such as H19 and XIST, regulate mucin gene expression and mucociliary clearance pathways, influencing mucus production and secretion in airways. Dysregulated expression of these lncRNAs disrupts airway homeostasis and exacerbates mucus hypersecretion in response to tobacco smoke exposure, contributing to airflow limitation and respiratory symptoms in COPD patients [90].
Epigenetic alterations play a crucial role in COPD pathogenesis through modulation of gene expression patterns and cellular phenotypes in response to environmental stimuli, such as tobacco smoke exposure [91]. LncRNAs, such as ANRIL and MEG3, regulate epigenetic modifications and chromatin remodeling, influencing gene transcription and cellular responses. They do so by guiding the recruitment of chromatin modifiers or transcriptional activators at specific loci. Dysregulated expression of these lncRNAs alters the epigenetic landscape of COPD airways, promoting aberrant gene expression profiles and disease progression in response to tobacco smoke exposure [92].
Lastly, cellular senescence is a key feature of COPD pathophysiology, characterized by irreversible growth arrest, senescence-associated secretory phenotype (SASP, and tissue remodeling [93]. LncRNAs, including TUG1 and FENDRR, regulate cellular senescence and senescence-associated inflammatory responses. Dysregulated expression of these lncRNAs accelerates cellular senescence and exacerbates tissue damage in COPD patients exposed to tobacco smoke, contributing to disease progression and ex-acerbation [94].
2.4. Correlation Between LncRNA Expression and COPD Severity
The correlation between lncRNA expression and COPD severity is a topic of significant interest in respiratory medicine, offering insights into disease progression and clinical outcomes [95]. Several studies have reported associations between aberrant lncRNA expression profiles and various parameters of COPD severity, including a decline of lung function, exacerbation frequency, and radiological features [96].
MALAT1, one of the most extensively studied lncRNAs in COPD, has been positively correlated with disease severity metrics such as FEV1 and the BODE index (Body mass index, airflow Obstruction, Dyspnea, and Exercise capacity) [97]. Elevated MALAT1 expression levels have been associated with accelerated lung function decline, increased risk of exacerbations, and poorer prognosis in COPD patients [98].
Similarly, HOTAIR expression levels have also been positively correlated with COPD severity scores and radiological evidence of emphysema and airway remodeling. The dysregulation of HOTAIR-mediated signaling pathways, including TGF-β (Transforming Growth Factor-β) and Wnt/β-catenin, has been implicated in the pathogenesis of COPD and the progression of emphysema [99]. The prognostic value of lncRNAs in predicting COPD outcomes extends beyond their individual expression levels to encompass their interactions with other molecular mediators and cellular pathways. Integrative analyses incorporating lncRNA signatures with other clinical predictors and biomarkers of COPD hold promise for refining risk stratification models and optimizing therapeutic strategies for the disease [100].
2.5. Prognostic and Diagnostic Value of LncRNA in Predicting COPD Outcomes
The prognostic value of lncRNAs in predicting COPD outcomes is a burgeoning area of research, with implications for disease monitoring, treatment response prediction, and personalized patient care [101]. Longitudinal cohort studies have identified specific lncRNA signatures associated with disease progression, exacerbation risk, and mortality in COPD patients [102].
GAS5, a well-characterized lncRNA involved in regulation of apoptosis, has emerged as a good prognostic biomarker for COPD outcomes. Decreased GAS5 expression levels are correlated with increased risk of exacerbations, hospitalizations, and mortality among COPD patients [103]. Conversely, NEAT1, a nuclear-enriched lncRNA involved in regulation of chromatin organization and gene expression, has be linked to worse prognosis of COPD patients. Elevated NEAT1 expression levels are associated with accelerated decline of lung functions, poorer response to bronchodilator therapy, and increased mortality [104]. Similarly, elevated expression levels of MALAT1 and NEAT1 are correlated with adverse clinical outcomes and poorer prognosis. These observations strongly suggest that these luncRNAs serve as prognostic indicators for risk stratification and treatment optimization [105], as well as diagnostic biomarkers for COPD, offering potential alternatives or complementary tools to traditional clinical assessments [106]. Furthermore, the expression levels of certain lncRNAs such as MALAT1, HOTAIR, and GAS5 predict COPD phenotypes and the degrees of disease severity and exacerbation risk [107].
Beyond their diagnostic and prognostic implications, lncRNAs also hold promise as therapeutic targets and predictors of disease progression and therapeutic response for COPD, enabling personalized treatment regimens tailored to individual patient profiles [108]. The dysregulation of lncRNAs contributes to key pathophysiological processes underlying COPD, including inflammation, oxidative stress, and airway remodeling. Targeted modulation of such dysregulated lncRNAs through pharmacological interventions or gene therapy represents a promising strategy for mitigating disease progression and improving patient outcomes [109].
3. Regulatory Functions of LncRNAs in COPD
LncRNAs regulate gene expression through various mechanisms at different stages of gene expression, including transcription, RNA processing, and translation. These mechanisms involve interactions with chromatin-modifying complexes, transcription factors, miRNAs, and other RNA-binding proteins [110].
Chromatin Remodeling: LncRNAs can influence chromatin structure and accessibility by recruiting chromatin-modifying complexes to specific genomic loci. For example, lncRNAs can interact with histone-modifying enzymes such as histone methyltransferases or histone deacetylases to promote or inhibit histone modifications that regulate gene expression [111]. By altering chromatin structure, lncRNAs can control the accessibility of transcriptional machinery to target genes [112]. Transcriptional Regulation: LncRNAs can directly influence transcriptional activity by interacting with transcription factors or RNA polymerase complexes. Some lncRNAs act as co-activators or co-repressors of transcription by binding to transcription factors and modulating their activity or recruitment to target gene promoters [113]. Others serve as scaffolds for the assembly of transcriptional complexes, bringing together regulatory proteins and DNA elements to regulate gene expression [114]. RNA Processing and Stability: LncRNAs can regulate RNA processing events, such as alternative splicing, RNA editing, and RNA stability. By binding to pre-mRNA transcripts or RNA processing factors, lncRNAs can influence splice site selection or exon inclusion/exclusion patterns, leading to the production of different mRNA isoforms with distinct functions [115]. Additionally, lncRNAs can stabilize or destabilize target mRNAs by forming RNA duplexes or competing for binding with miRNAs, thereby modulating mRNA turnover and translation efficiency [116]. Epigenetic Regulation: LncRNAs can regulate gene expression in a heritable manner by influencing epigenetic modifications such as DNA methylation and histone modification patterns. Some lncRNAs act as guides or scaffolds for recruiting chromatin-modifying enzymes to specific genomic loci, leading to changes in DNA methylation status or histone acetylation/methylation patterns that affect gene expression. These epigenetic changes can be stably inherited through cell divisions and play important roles in cell fate determination and differentiation [117]. Subcellular Localization: LncRNAs can localize to specific subcellular compartments, such as the nucleus or cytoplasm, where they exert distinct regulatory functions [118]. Nuclear lncRNAs often regulate transcriptional processes by interacting with chromatin or transcriptional machinery, while cytoplasmic lncRNAs may modulate post-transcriptional events such as mRNA stability, translation, or protein localization. Overall, lncRNAs play diverse and complex roles in the regulation of gene expression, acting at multiple levels to fine-tune cellular processes and maintain cellular homeostasis [119].
3.1. LncRNAs Control Gene Expression in COPD
LncRNAs can regulate the expression of various genes involved in the pathogenesis of COPD. These genes participate in processes such as inflammation, oxidative stress, tissue remodeling, and immune response, all of which contribute to the development and progression of COPD. Here are a few examples of genes that could be regulated by lncRNAs in COPD [120].
Interleukin 8 (IL-8): IL-8 is a pro-inflammatory cytokine that plays a crucial role in recruiting neutrophils to the site of inflammation in COPD airways. Dysregulated expression of IL-8 contributes to chronic inflammation and airway remodeling in COPD [121]. Certain lncRNAs, such as NEAT1, have been shown to regulate the expression of IL-8 by modulating chromatin remodeling and transcriptional regulation mechanisms [122]. Tumor Necrosis Factor Alpha (TNF-α): TNF-α is another pro-inflammatory cytokine that is elevated in the lungs of COPD patients and contributes to airway inflammation and tissue damage. Several lncRNAs, including HOTAIR and GAS5, have been implicated in the regulation of TNF-α expression through various mechanisms, such as interaction with transcription factors or modulation of miRNA activity [123]. Matrix Metalloproteinases (MMPs): MMPs are a family of enzymes involved in tissue remodeling and extracellular matrix degradation. MMPs are upregulated in COPD and contribute to the destruction of lung parenchyma and emphysema development [124]. LncRNAs such as MALAT1 and H19 have been shown to regulate the expression of MMPs by influencing transcriptional activity or post-transcriptional processing [125]. Nuclear Factor-kappa B (NF-κB): NF-κB is a key transcription factor that regulates the expression of genes involved in inflammation, immune response, and cell survival. Dysregulated NF-κB signaling is implicated in the pathogenesis of COPD. LncRNAs like MEG3 and ANRIL can modulate NF-κB activity by interacting with NF-κB regulatory elements or regulating upstream signaling pathways [126]. Fibroblast Growth Factor (FGF) Family: The FGF family members are involved in cell proliferation, angiogenesis, and tissue repair. Dysregulated FGF signaling has been implicated in COPD pathogenesis, particularly in the context of tissue repair and remodeling. LncRNAs such as HOTAIR and TUG1 have been shown to influence FGF signaling pathways by interacting with transcription factors or modulating chromatin structure [127].
3.2. LncRNA-Mediated Immune Response in COPD
COPD is characterized by persistent inflammation and immune dysregulation in the airways and lung parenchyma, driven by complex interactions between immune cells, cytokines, and regulatory molecules. LncRNAs have emerged as critical regulators of the immune response in COPD, modulating inflammatory signaling pathways, immune cell differentiation, and cytokine production in the lung microenvironment [128].
Several lncRNAs have been implicated in the modulation of immune responses in COPD, including NEAT1, GAS5, MALAT1, TUG1, and HOTAIR. These lncRNAs interact with RNA-binding proteins, miRNAs, and transcription factors to regulate the expression of immune-related genes and signaling pathways involved in COPD pathogenesis [129].
NEAT1: NEAT1 is a nuclear-retained lncRNA that forms the structural scaffold for the formation of nuclear bodies known as paraspeckles [130]. In COPD, dysregulated expression of NEAT1 alters inflammatory cytokine production and immune cell function, contributing to disease progression. NEAT1 interacts with RNA-binding proteins and transcription factors to regulate the expression of immune-related genes involved in COPD pathogenesis [131]. GAS5: GAS5 is a stress-induced lncRNA that regulates the proliferation and apoptosis of immune cells, including T cells and macrophages. Dysregulated expression of GAS5 in COPD affects immune cell function and inflammatory cytokine production, exacerbating airway inflammation and tissue damage. GAS5 modulates the activity of key signaling pathways involved in COPD pathogenesis, including NF-κB and JAK-STAT signaling [132]. HOTAIR: HOTAIR is an oncogenic lncRNA implicated in the regulation of immune responses in COPD. Dysregulated expression of HOTAIR alters the expression of immune-related genes and cytokines, contributing to chronic inflammation and tissue damage in COPD patients. HOTAIR interacts with chromatin-modifying complexes and transcription factors to regulate the expression of genes involved in immune cell activation and cytokine signaling [133]. MALAT1: MALAT1 is a highly conserved lncRNA that regulates immune responses and inflammatory signaling pathways in COPD. Dysregulated expression of MALAT1 affects immune cell infiltration, cytokine production, and tissue remodeling processes in COPD patients. MALAT1 interacts with RNA-binding proteins and miRNAs to modulate the expression of genes involved in inflammatory responses and immune cell function [134]. TUG1: TUG1 is a lncRNA implicated in the regulation of immune responses and inflammatory signaling pathways in COPD. Dysregulated expression of TUG1 alters the expression of immune-related genes and cytokines, contributing to airway inflammation and tissue remodeling in COPD patients. TUG1 interacts with chromatin-modifying complexes and transcription factors to regulate the expression of genes involved in immune cell activation and inflammatory responses [135].
3.3. Complexities of Inflammatory Mechanisms in COPD
Recent studies have implicated lncRNAs as key regulators of inflammatory pathways in various diseases, including COPD, dysregulated expression of lncRNAs has been associated with aberrant inflammatory responses, airway remodeling, and disease progression [136]. Several lncRNAs have been identified as critical regulators of inflammatory signaling pathways in COPD pathogenesis. Moreover, emerging evidence suggests that lncRNAs may also modulate the expression of key mediators involved in COPD-associated inflammation, including chemokines, growth factors, and proteases. By influencing the expression and activity of these mediators, lncRNAs can modulate the recruitment and activation of inflammatory cells, exacerbating the inflammatory milieu within the airways [137].
COPD is a multifaceted respiratory condition characterized by persistent airflow limitation and inflammation within the airways. This section critically examines the inflammatory mechanisms underpinning COPD pathogenesis, delineated by two seminal texts. Firstly, cigarette smoke and various irritants emerge as primary instigators, igniting inflammatory cascades within the respiratory tract [138]. In response to these insults, activated macrophages liberate potent neutrophil chemotactic factors like interleukin-8 (IL-8) and leukotriene B4 (LTB4), priming the groundwork for cellular recruitment and activation. These neutrophils, once mobilized, unleash proteases that inflict collateral damage upon the lung parenchyma, fostering emphysematous changes and exacerbating mucus hypersecretion. Moreover, the delicate balance between proteases and inhibitors, including alpha-1-antitrypsin, secretory leukocyte protease inhibitor (SLPI), and tissue inhibitor of metalloproteinases (TIMP), underscores the dynamic interplay governing tissue homeostasis [139]. Concurrently, cytotoxic T cells (CD8+) and fibroblasts are implicated as additional protagonists, orchestrating alveolar wall destruction and fibrotic remodeling under the influence of growth factors released by macrophages and epithelial cells [140].
Expanding upon this intricate landscape, the second discourse delves into the intricate web of inflammation permeating COPD pathology. Inflammation, as portrayed, is a complex phenomenon orchestrated by a myriad of activated inflammatory and structural cells. Lipid mediators such as LTB4, alongside chemokines including monocyte chemotactic protein-1 (MCP-1) and macrophage inflammatory protein-1 alpha (MIP-1α), emerge as pivotal in orchestrating leukocyte recruitment and activation [141]. The triumvirate of interleukin-8 (IL-8), growth-related oncogene alpha (GRO-α), and interferon-gamma-inducible protein-10 (IP-10) further potentiates this cellular influx, with the latter also attracting cytotoxic T cells alongside reactive oxygen species (ROS) and nitric oxide (NO) [142]. Notably, the pro-inflammatory cytokine tumor necrosis factor-alpha (TNF-α) assumes a central role in amplifying inflammation, gene expression, and mediating systemic sequelae of COPD [143]. Additionally, endothelin and transforming growth factor-beta (TGF-β) emerge as pivotal mediators in fibrotic remodeling, epitomizing the intricate interplay between inflammation and tissue remodeling in COPD pathogenesis (Figure 5) [144]. Furthermore, the release of proteases, including neutrophil elastase, proteinase C, cathepsins, and MMPs, underscores the proteolytic milieu perpetuating elastolysis and mucus hypersecretion, thus perpetuating the hallmark pathophysiology of COPD [145].
3.4. Modulation of Cellular Processes by LncRNA in COPD
LncRNAs play pivotal roles in modulating cellular processes implicated in COPD, contributing to disease pathogenesis and progression. Through diverse mechanisms, including regulation of gene expression, modulation of signaling pathways, and interaction with cellular components, lncRNAs influence key cellular processes in COPD [146].
Epithelial-to-Mesenchymal Transition (EMT): EMT is a cellular process characterized by the transition of epithelial cells into mesenchymal cells, leading to increased cell motility, invasiveness, and fibrosis [147]. Dysregulated expression of lncRNAs such as HOTAIR and MALAT1 has been implicated in promoting EMT in COPD. These lncRNAs regulate the expression of genes involved in EMT-related pathways, including TGF-β signaling and matrix metalloproteinases, facilitating airway remodeling and fibrosis in COPD patients [148].
Cell Proliferation and Apoptosis: Dysregulated cell proliferation and apoptosis contribute to tissue remodeling, airway obstruction, and emphysema in COPD [149]. LncRNAs such as GAS5 and TUG1 modulate cell cycle progression, apoptosis, and senescence in COPD by regulating the expression of genes involved in cell growth and survival pathways. Dysregulated expression of these lncRNAs alters the balance between cell proliferation and apoptosis, contributing to disease progression and exacerbation in COPD patients [150].
Inflammatory Signaling Pathways: Chronic inflammation is a hallmark feature of COPD, characterized by increased production of pro-inflammatory cytokines and chemokines in the lung microenvironment [151]. LncRNAs such as NEAT1 and XIST regulate inflammatory signaling pathways in COPD by modulating the expression of genes involved in cytokine production, immune cell infiltration, and tissue remodeling. Dysregulated expression of these lncRNAs contributes to sustained inflammation and tissue damage in COPD patients, exacerbating disease severity and progression [152].
Oxidative Stress Response: Oxidative stress plays a central role in COPD pathogenesis, contributing to airway inflammation, oxidative damage, and impaired lung function [153]. lncRNAs such as H19 and MEG3 regulate oxidative stress response pathways in COPD by modulating the expression of antioxidant enzymes, stress-responsive genes, and redox signaling molecules. Dysregulated expression of these lncRNAs disrupts cellular homeostasis and exacerbates oxidative damage in COPD patients, contributing to disease progression and exacerbation [154].
Fibrotic Processes: Pulmonary fibrosis is a common complication of COPD, characterized by excessive deposition of extracellular matrix proteins and tissue remodeling in the lung parenchyma [155]. LncRNAs such as FENDRR and HOTAIR regulate fibrotic processes in COPD by modulating the expression of genes involved in fibroblast activation, collagen synthesis, and tissue remodeling. Dysregulated expression of these lncRNAs promotes aberrant fibroblast proliferation and extracellular matrix deposition, leading to progressive fibrosis and loss of lung function in COPD patients [156]. Understanding the molecular mechanisms by which lncRNAs modulate cellular processes in COPD offers insights into disease pathogenesis and potential therapeutic targets for COPD management. Targeting dysregulated lncRNAs may offer novel strategies for inhibiting airway remodeling, inflammation, oxidative stress, and fibrosis in COPD patients, ultimately improving disease outcomes and quality of life [157].
4. Modulatory Mechanisms of COPD-Associated LncRNA in Lung Cancer Progression
Lung cancer remains a significant global health challenge, with high mortality rates and limited treatment options. LncRNAs have emerged as critical regulators of lung cancer progression, offering potential insights into novel therapeutic strategies and prognostic markers [158]. This review explores the intricate modulatory mechanisms of lncRNAs in lung cancer progression, focusing on the interaction between COPD-associated lncRNAs and lung cancer pathways, the regulatory functions of lncRNA in lung cancer development from COPD, and the implications of lncRNA dysregulation in lung cancer metastasis [159].
4.1. Interaction Between COPD-Associated LncRNA and Lung Cancer Pathways
COPD and lung cancer are two major respiratory diseases that share common risk factors, including cigarette smoking and exposure to environmental pollutants [160]. Emerging evidence suggests that dysregulated lncRNAs play pivotal roles in both COPD pathogenesis and lung cancer progression [161]. This review delves into the intricate relationship between COPD-implicated lncRNAs and lung cancer progression, highlighting the molecular mechanisms underlying their interplay and potential implications for disease management
4.1.1. Shared Molecular Pathways
COPD-implicated lncRNAs and lung cancer progression often converge on shared molecular pathways involved in inflammation, oxidative stress, and cellular proliferation [162]. For instance, lncRNA H19, initially identified as a regulator of embryonic development and imprinting, exhibits dysregulated expression in COPD and lung cancer tissues [163]. H19 promotes lung cancer progression by modulating pathways associated with cell proliferation, invasion, and metastasis. Its overexpression correlates with advanced tumor stage and poor prognosis in lung cancer patients, highlighting its potential as a prognostic biomarker and therapeutic target [164].
Similarly, lncRNA MALAT1 has been implicated in COPD pathogenesis and lung cancer metastasis. MALAT1 expression is upregulated in COPD and lung cancer tissues, where it promotes tumor invasion and metastasis by regulating alternative splicing events and gene expression programs associated with metastatic progression [165]. Its aberrant expression correlates with disease aggressiveness and poor clinical outcomes in lung cancer patients, underscoring its potential as a therapeutic target for metastatic disease [166].
4.1.2. Regulatory roles of COPD-Associated LncRNA in Lung Cancer
COPD-implicated lncRNAs exert diverse regulatory functions in lung cancer progression, including modulation of gene expression, epigenetic regulation, and interaction with signaling pathways implicated in tumor development and metastasis [167]. For instance, ANRIL (antisense non-coding RNA in the INK4 locus) is upregulated in COPD and lung cancer tissues, where it promotes tumor growth and metastasis by interacting with chromatin remodeling complexes and transcription factors involved in oncogenic signaling pathways [168].
Moreover, lncRNA TUG1 exhibits dysregulated expression in COPD and lung cancer tissues, contributing to disease aggressiveness and treatment resistance [169]. TUG1 promotes lung cancer progression by modulating epithelial-to-mesenchymal transition (EMT) and metastasis through its interaction with miRNAs and protein-coding genes involved in tumor initiation and progression. Its aberrant expression serves as a prognostic marker for poor survival outcomes in lung cancer patients with a history of COPD, highlighting its potential as a therapeutic target for preventing disease progression [170].
4.2. Dysregulated Expression of COPD-Associated LncRNAs in Lung Cancer
COPD is a prevalent respiratory condition characterized by airflow limitation and chronic inflammation, primarily caused by exposure to tobacco smoke and environmental pollutants. It significantly increases the risk of developing lung cancer, the leading cause of cancer-related mortality worldwide. Recent studies have highlighted the dysregulated expression of lncRNAs in both COPD and lung cancer, suggesting their potential roles as biomarkers and therapeutic targets in disease progression [171]. This review aims to explore the dysregulated expression of COPD-associated lncRNAs in lung cancer, elucidating their molecular mechanisms and clinical implications.
ANRIL: ANRIL, also known as CDKN2B-AS1, is a well-characterized lncRNA implicated in COPD and lung cancer pathogenesis [172]. Its dysregulated expression contributes to aberrant cell cycle progression, apoptosis, and cellular senescence, promoting tumor growth and metastasis. In COPD, ANRIL upregulation correlates with disease severity and airflow limitation, suggesting its potential as a biomarker for COPD progression. Similarly, ANRIL overexpression in lung cancer tissues is associated with advanced tumor stage, lymph node metastasis, and poor prognosis in patients [173]. Mechanistically, ANRIL regulates the expression of tumor suppressor genes, oncogenes, and cell cycle regulators, orchestrating a complex network of molecular interactions that drive oncogenic processes in lung cancer cells [174]. H19: H19 is a maternally expressed lncRNA involved in embryonic development and genomic imprinting, whose dysregulated expression has been implicated in COPD and lung cancer progression. In COPD patients, H19 upregulation correlates with airway inflammation, oxidative stress, and lung function decline, suggesting its potential as a diagnostic and prognostic biomarker [175]. In lung cancer, H19 promotes tumor growth, angiogenesis, and metastasis through its interactions with miRNAs, transcription factors, and signaling pathways involved in tumor progression. Its aberrant expression in lung cancer tissues is associated with tumor aggressiveness, treatment resistance, and poor clinical outcomes in patients [176]. MALAT1: MALAT1 is a highly conserved lncRNA implicated in COPD pathogenesis and lung cancer metastasis. Its dysregulated expression contributes to disease progression by modulating alternative splicing events, gene expression programs, and signaling pathways associated with tumor invasion and metastasis [177]. In COPD, MALAT1 upregulation correlates with airway remodeling, mucus hypersecretion, and disease severity, suggesting its potential as a therapeutic target for COPD management. In lung cancer, MALAT1 promotes metastatic spread, EMT, and resistance to chemotherapy and targeted therapies, highlighting its prognostic value and therapeutic potential in advanced-stage disease [178]. TUG1: TUG1 is a conserved lncRNA implicated in COPD pathogenesis and lung cancer progression, whose dysregulated expression correlates with disease severity and poor clinical outcomes. In COPD patients, TUG1 upregulation is associated with airway inflammation, oxidative stress, and lung function decline, indicating its potential as a biomarker for disease progression and exacerbation risk [179]. In lung cancer, TUG1 promotes tumor growth, metastasis, and resistance to therapy by modulating the expression of oncogenes, tumor suppressors, and signaling pathways involved in cell proliferation and survival. Its aberrant expression serves as a prognostic marker for poor survival outcomes in lung cancer patients, highlighting its therapeutic potential as a target for personalized treatment approaches [180]. HOTAIR: HOTAIR is a well-characterized lncRNA implicated in COPD and lung cancer pathogenesis, whose dysregulated expression correlates with disease severity and progression. In COPD patients, HOTAIR upregulation is associated with airway inflammation, fibrosis, and lung function decline, suggesting its potential as a biomarker for disease exacerbation and progression risk. In lung cancer, HOTAIR promotes tumor invasion, metastasis, and angiogenesis by modulating chromatin remodeling complexes, transcription factors, and signaling pathways involved in tumor progression [181]. Its aberrant expression is associated with advanced tumor stage, lymph node metastasis, and poor prognosis in patients, highlighting its prognostic value and therapeutic potential in advanced-stage disease [182]. UCA1: UCA1 is an oncogenic lncRNA implicated in COPD pathogenesis and lung cancer progression, whose dysregulated expression contributes to disease aggressiveness and poor clinical outcomes [183]. In COPD patients, UCA1 upregulation correlates with airway inflammation, mucus hypersecretion, and disease severity, indicating its potential as a biomarker for disease progression and exacerbation risk. In lung cancer, UCA1 promotes tumor growth, metastasis, and resistance to therapy by modulating cell proliferation, apoptosis, and DNA repair mechanisms. Its aberrant expression serves as a prognostic marker for poor survival outcomes in lung cancer patients, highlighting its potential as a therapeutic target for personalized treatment approaches [184]. FENDRR (Fetal-lethal Non-coding Developmental RNA): FENDRR is a developmentally regulated lncRNA implicated in COPD and lung cancer pathogenesis, whose dysregulated expression correlates with disease severity and progression. In COPD patients, FENDRR upregulation is associated with airway inflammation, oxidative stress, and lung function decline, suggesting its potential as a biomarker for disease exacerbation and progression risk [185]. In lung cancer, FENDRR functions as a tumor suppressor by inhibiting cell proliferation, invasion, and metastasis through its interactions with miRNAs and transcription factors involved in tumor progression. Its downregulation is associated with advanced tumor stage, lymph node metastasis, and poor prognosis in patients, highlighting its potential as a prognostic biomarker and therapeutic target in lung cancer [186]. AK098656: AK098656 is a less studied lncRNA but has been implicated in both COPD and lung cancer. Its dysregulated expression suggests potential roles in disease pathogenesis and progression, although further research is needed to elucidate its molecular mechanisms and clinical implications in COPD and lung cancer [187].
5. LncRNA in Body Fluids for COPD Diagnosis and Therapy
LncRNAs have emerged as promising biomarkers and therapeutic targets for COPD. Detection of lncRNA biomarkers in body fluids such as blood and urine offer non-invasive diagnostic tools for early disease detection and monitoring [188]. Moreover, the therapeutic potential of lncRNAs in COPD management holds promise for personalized treatment approaches and disease intervention strategies.
5.1. Detection of LncRNA Biomarkers in Blood Samples
Blood-based biomarkers have emerged as indispensable tools for diagnosing and monitoring COPD owing to their accessibility and minimally invasive nature. LncRNAs detected in peripheral blood samples have demonstrated considerable promise as diagnostic and prognostic biomarkers for COPD [189]. Among these, NEAT1, MALAT1, and HOTAIR have surfaced as notable candidates, showcasing dysregulated expression patterns in COPD patients compared to healthy controls. Specifically, the circulating levels of these lncRNAs exhibit variations correlated with disease severity, exacerbation risk, and response to therapeutic interventions, underscoring their clinical relevance in the context of COPD diagnosis and prognosis [190].
In recent years, the advent of high-throughput sequencing technologies and sophisticated bioinformatics analyses has accelerated the identification of novel lncRNA biomarkers in blood samples of COPD patients [191]. By leveraging multi-omics data integration and machine learning algorithms, researchers have endeavored to develop robust predictive models for assessing COPD risk and monitoring disease progression. These endeavors hold promise in enhancing the precision and efficacy of COPD management strategies. However, it is imperative to conduct comprehensive validation studies in large patient cohorts to ascertain the diagnostic accuracy and clinical utility of blood-based lncRNA biomarkers in COPD management [192].
Furthermore, the landscape of blood-based biomarkers in COPD extends beyond individual lncRNAs to encompass a spectrum of molecular entities, including miRNAs, messenger RNAs, and proteins. For instance, miR-21 and miR-146a have emerged as key regulators implicated in COPD pathogenesis, displaying dysregulated expression patterns reflective of disease severity and exacerbation risk [193]. Likewise, the expression profiles of certain protein biomarkers, such as C-reactive protein (CRP), interleukin-6 (IL-6), and tumor necrosis factor-alpha (TNF-alpha), exhibit dynamic alterations associated with COPD progression and exacerbation events. The integration of diverse molecular biomarkers holds the potential to furnish clinicians with comprehensive insights into COPD pathophysiology and patient-specific disease trajectories [194].
Moreover, the advent of precision medicine approaches underscores the importance of leveraging blood-based biomarkers to tailor therapeutic interventions according to individual patient profiles. By elucidating the intricate molecular signatures underpinning COPD pathogenesis, blood-based biomarkers pave the way for personalized and targeted therapeutic strategies aimed at mitigating disease progression and improving patient outcomes [195]. The exploration of lncRNA biomarkers in blood samples represents a pivotal avenue in the quest for refined diagnostic and prognostic tools in COPD management. Through concerted efforts in biomarker discovery, validation, and clinical translation, researchers aspire to harness the full potential of blood-based biomarkers to usher in a new era of precision medicine tailored to the intricate nuances of COPD pathophysiology.
5.2. Urinary LncRNA as a Diagnostic Tool for COPD
Urine-based biomarkers stand at the forefront of non-invasive and readily accessible samples for diagnosing and monitoring COPD, a prevalent respiratory condition posing significant public health challenges. LncRNAs, discerned within urinary exosomes and cell-free fractions, have emerged as promising candidates for enhancing COPD diagnostic precision [196]. Notably, the dysregulated expression profiles of urinary lncRNAs, including UCA1, TUG1, and H19, have exhibited correlations with the severity of COPD manifestations, declines in pulmonary function, and heightened risks of exacerbations among afflicted individuals. Such associations underscore the potential utility of urinary lncRNAs as indispensable diagnostic biomarkers, offering insights into the dynamic interplay between molecular signatures and disease progression [197].
Delving deeper into the molecular landscape, the characterization of urinary lncRNA profiles in COPD unveils intricate mechanistic underpinnings and unveils potential therapeutic targets. Mechanistic inquiries have shed light on the pivotal roles of urinary lncRNAs in orchestrating inflammatory cascades, modulating oxidative stress responses, and orchestrating tissue remodeling processes within the pulmonary microenvironment. For instance, investigations have delineated how dysregulated urinary lncRNAs, such as UCA1, influence the activation of inflammatory signaling pathways, exacerbating pulmonary inflammation and perpetuating tissue damage characteristic of COPD pathogenesis [198]. Moreover, elucidating the contributions of urinary lncRNAs, like TUG1 and H19, to oxidative stress dynamics underscores their potential as modulators of redox homeostasis, thereby influencing disease progression and exacerbation susceptibility [199].
Furthermore, the exploration of urinary lncRNA signatures unveils novel avenues for personalized treatment strategies and targeted interventions in COPD management. By discerning aberrant expression patterns of urinary lncRNAs, clinicians can tailor therapeutic modalities to address individualized disease phenotypes and molecular profiles [200]. For instance, therapeutic targeting of dysregulated urinary lncRNAs holds promise for mitigating inflammatory responses, ameliorating oxidative stress burdens, and attenuating tissue remodeling processes within the pulmonary microenvironment. Integrating insights from mechanistic studies with clinical observations paves the way for innovative therapeutic approaches aimed at disrupting pathological pathways underpinning COPD progression [201].
In essence, urinary lncRNAs represent invaluable diagnostic tools and therapeutic targets in the multifaceted landscape of COPD. Their intricate roles in mediating inflammatory cascades, modulating oxidative stress dynamics, and orchestrating tissue remodeling processes underscore their significance in unraveling the molecular complexities of COPD pathogenesis [202]. Characterizing the molecular signatures of urinary lncRNAs in COPD provides insights into disease pathogenesis and potential therapeutic targets. Mechanistic studies have elucidated the role of urinary lncRNAs in modulating inflammatory responses, oxidative stress, and tissue remodeling processes in the lung microenvironment. Targeting dysregulated urinary lncRNAs may offer novel strategies for personalized treatment approaches and disease intervention in COPD patients [203]. Harnessing the diagnostic and therapeutic potential of urinary lncRNAs not only enhances our understanding of COPD pathophysiology but also offers unprecedented opportunities for personalized disease management and intervention strategies tailored to individual patient needs [204].
5.3. Pharmacological Actions of LncRNA Molecules as Potential Therapeutics for COPD
LncRNAs have emerged as promising targets for pharmacological intervention in COPD, offering potential therapeutic strategies for mitigating disease progression and improving patient outcomes. Several lncRNAs have been identified as key regulators of inflammatory responses, airway remodeling, exacerbation risk, vascular remodeling, and mucous hypersecretion in COPD (Figure 6) [205]. Targeting dysregulated lncRNA molecules holds promise for developing novel therapeutics with anti-inflammatory, anti-airway remodeling, anti-exacerbation, anti-vascular remodeling, and anti-mucous hypersecretion properties [206].
Anti-inflammatory Actions: In COPD, chronic inflammation plays a central role in disease pathogenesis, contributing to airway obstruction, tissue damage, and decline in lung function [207]. Dysregulated expression of lncRNAs such as NEAT1, MEG3, H19, MALAT1, and GAS5 has been implicated in modulating inflammatory signaling pathways and cytokine production in COPD patients. Targeting these lncRNAs with pharmacological agents such as antisense oligonucleotides or small interfering RNAs may attenuate inflammatory responses, reduce immune cell infiltration, and mitigate tissue inflammation in COPD [208]. Anti-Airway Remodeling Effects: Airway remodeling is a hallmark feature of COPD, characterized by structural changes in the airway epithelium, smooth muscle hypertrophy, and extracellular matrix deposition [209]. LncRNAs such as GAS5, HOTAIR, TUG1, and FENDRR regulate genes involved in airway remodeling processes, including TGF-β signaling, matrix metalloproteinases, and collagen synthesis. Pharmacological targeting of these lncRNAs may inhibit airway remodeling, restore airway structure and function, and improve lung function in COPD patients [210]. Anti-Exacerbation Properties: Exacerbations are acute worsening of COPD symptoms, often triggered by respiratory infections, air pollution, or environmental factors [211]. Dysregulated expression of lncRNAs such as RP11-713B14.1, XIST, UCA1, and H19 has been associated with exacerbation risk and disease progression in COPD patients. Targeting these lncRNAs with pharmacological interventions may reduce exacerbation frequency, attenuate symptom severity, and improve the quality of life in COPD patients [212]. Anti-Vascular Remodeling Effects: Vascular remodeling contributes to pulmonary hypertension, right heart failure, and cardiovascular complications in COPD patients [213]. lncRNAs such as ANRIL, MEG3, HOTAIR, and MALAT1 regulate vascular smooth muscle cell proliferation, endothelial dysfunction, and angiogenesis in COPD. Pharmacological modulation of these lncRNAs may inhibit vascular remodeling, reduce pulmonary vascular resistance, and improve pulmonary hemodynamics in COPD patients [214]. Anti-Mucous Hypersecretion Actions: Mucous hypersecretion is a common feature of COPD, contributing to airway obstruction, impaired mucociliary clearance, and recurrent infections. lncRNAs such as TUG1, GAS5, MALAT1, and NEAT1 regulate mucin gene expression, goblet cell hyperplasia, and mucous production in COPD. Pharmacological targeting of these lncRNAs may reduce mucous hypersecretion, improve airway clearance, and reduce the risk of respiratory infections in COPD patients [215].
5.4. Potential Therapeutic Applications of LncRNA in COPD
Therapeutic modulation of lncRNA expression holds promise for COPD management, offering novel approaches for disease intervention and personalized treatment strategies. Targeting dysregulated lncRNAs involved in COPD pathogenesis, such as HOTAIR, MALAT1, and GAS5, may attenuate airway inflammation, oxidative stress, and tissue remodeling in COPD patients [216].
Various strategies have been explored for lncRNA-based therapeutics, including antisense oligonucleotides, small interfering RNAs, and CRISPR-based genome editing. These approaches enable targeted modulation of lncRNA expression and activity, restoring cellular homeostasis and attenuating disease progression in COPD [217]. Moreover, nanoparticle-mediated delivery systems offer efficient and targeted delivery of lncRNA therapeutics to the lung microenvironment, minimizing off-target effects and enhancing therapeutic efficacy [218].
Clinical translation of lncRNA-based therapeutics requires rigorous preclinical validation and safety assessments in animal models and human clinical trials. Optimization of delivery strategies, dosage regimens, and treatment protocols is essential for maximizing therapeutic efficacy and minimizing potential adverse effects. Furthermore, elucidating the molecular mechanisms underlying lncRNA-mediated therapeutic effects in COPD provides insights into disease pathogenesis and potential biomarkers for treatment response monitoring [219].
Conclusions and Future Directions:
The exploration of COPD-associated lncRNAs reveals a complex regulatory network implicated in disease pathogenesis, progression, and therapeutic interventions. From MALAT1 to HOTAIR and GAS5, dysregulated lncRNA expression profiles intertwine with COPD severity metrics, offering insights into disease prognosis and treatment response. Longitudinal cohort studies unravel distinct lncRNA signatures associated with exacerbation risk, lung function decline, and mortality, laying the foundation for personalized therapeutic regimens. While diagnostic and prognostic implications abound, lncRNAs also emerge as promising therapeutic targets for mitigating airway inflammation, oxidative stress, and tissue remodeling in COPD. The translation of lncRNA-based interventions into clinical practice necessitates rigorous validation studies and mechanistic elucidation to optimize treatment strategies and refine biomarker discovery. In essence, the intricate interplay between lncRNAs and COPD unveils novel avenues for precision medicine, promising improved patient outcomes and enhanced quality of life in COPD management.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/XXXXXX, Supplementary Excel files 1. LncRNA/miRNA/mRNA implicated in COPD with their functions during the disease progression. Each entry includes detailed information on the specific functions of these RNA molecules and their roles in the progression of COPD. The table highlights how these molecular components interact and contribute to the underlying mechanisms of the disease, offering insights into their potential as therapeutic targets and biomarkers. Supplementary Excel files 2. LncRNA/miRNA/mRNA is involved in lung carcinogenesis and modulatory role during lung cancer progression. The table provides an in-depth overview of how these RNA molecules influence cancer development and progression, emphasizing their potential in diagnosing, prognosis, and treating lung cancer.
Funding
The research endeavors of S.F. were made possible through the generous backing of the Metro Health Medical Center's startup fund, the American Cancer Society Research Scholar Grant (RSG-18-238-01-CSM), and the National Cancer Institute Research Grants (R01CA248304 and R21CA288449).
Authors Contributions
O.S. and S.F. contributed to the conception and design of the review study, conducted the literature search and review, and drafted the original manuscript. R.M., T.T., M.A., A.A., E.Z., Y.C., V.T., E.C., and M.A. made significant contributions to the reviewing and editing of the manuscript through their dedicated and strenuous efforts. O.S., R.M., and S.F. provided critical feedback on the manuscript and revised the manuscript. All authors have read and approved the final version of the manuscript for publication.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
This article contains the data presented in the study as well as supplementary Excel files for reference.
Acknowledgments
Heartfelt thanks are extended to all authors whose studies underwent comprehensive examination, enriching our manuscript with invaluable feedback and astute suggestions. The authors affirm a complete absence of conflicts of interest regarding this review. To ensure the research's impartiality and integrity, every necessary precaution has been meticulously undertaken by the authors. Special thanks to Neb Burke Zell for editing, revising the manuscript, and adjusting the grammar.
References
- Global Initiative for Chronic Obstructive Lung Disease (GOLD). Global Strategy for the Diagnosis, Management, and Prevention of Chronic Obstructive Pulmonary Disease; 2021. Available online: https://goldcopd.org/gold-reports/ (accessed on 18 January 2024).
- Soriano, J.B.; Kendrick, P.J.; Paulson, K.R.; et al. Prevalence and Attributable Health Burden of Chronic Respiratory Diseases, 1990–2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet Respir Med. 2020, 8, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Gurer, D.C.; Erdogan, İ.; Ahmadov, U.; Basol, M.; Sweef, O.; Cakan-Akdogan, G.; Akgül, B. Transcriptomics Profiling Identifies Cisplatin-Inducible Death Receptor 5 Antisense Long Non-coding RNA as a Modulator of Proliferation and Metastasis in HeLa Cells. Front Cell Dev Biol. 2021, 9, 688855. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Quinn, J.J.; Chang, H.Y. Unique Features of Long Non-Coding RNA Biogenesis and Function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Ding, J.W.; Wang, X.A.; et al. Dysregulation of lncRNAs and mRNAs Expression in the Development of Chronic Obstructive Pulmonary Disease. J. Cell. Biochem. 2018, 119, 4600–4610. [Google Scholar] [CrossRef]
- de-Torres, J.P.; Wilson, D.O.; Sanchez-Salcedo, P.; et al. Lung Cancer in Patients with Chronic Obstructive Pulmonary Disease: Development and Validation of the COPD Lung Cancer Screening Score. Am. J. Respir. Crit. Care Med. 2015, 191, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Durham, A.L.; Adcock, I.M. The Relationship between COPD and Lung Cancer. Lung Cancer 2015, 90, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Erdoğan, İ.; Sweef, O.; Akgül, B. Long Noncoding RNAs in Human Cancer and Apoptosis. Curr Pharm Biotechnol. 2023, 24, 872–888. [Google Scholar] [CrossRef] [PubMed]
- Agusti, A.; Edwards, L.D.; Rennard, S.I.; et al. Persistent systemic inflammation is associated with poor clinical outcomes in COPD: a novel phenotype. PLoS One 2012, 7, e37483. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Inflammatory mechanisms in patients with chronic obstructive pulmonary disease. J. Allergy Clin. Immunol. 2016, 138, 16–27. [Google Scholar] [CrossRef]
- Vogelmeier, C.F.; Criner, G.J.; Martinez, F.J.; et al. Global Strategy for the Diagnosis, Management, and Prevention of Chronic Obstructive Lung Disease 2017 Report. GOLD Executive Summary. Am. J. Respir. Crit. Care Med. 2017, 195, 557. [Google Scholar] [CrossRef]
- Wedzicha, J.A.; Seemungal, T.A. COPD exacerbations: defining their cause and prevention. Lancet 2007, 370, 786–796. [Google Scholar] [CrossRef] [PubMed]
- Celli, B.R.; MacNee, W. ; ATS/ERS Task Force. Standards for the diagnosis and treatment of patients with COPD: a summary of the ATS/ERS position paper. Eur. Respir. J. 2004, 23, 932–946. [Google Scholar] [CrossRef] [PubMed]
- Global Initiative for Chronic Obstructive Lung Disease (GOLD). Global Strategy for the Diagnosis, Management, and Prevention of Chronic Obstructive Pulmonary Disease; 2021. Available online: https://goldcopd.org/gold-reports/ (accessed on 15 April 2024).
- Pellegrino, R.; Viegi, G.; Brusasco, V.; et al. Interpretative strategies for lung function tests. Eur. Respir. J. 2005, 26, 948–968. [Google Scholar] [CrossRef] [PubMed]
- Celli, B.R.; Cote, C.G.; Marin, J.M.; et al. The Body-Mass Index, Airflow Obstruction, Dyspnea, and Exercise Capacity Index in Chronic Obstructive Pulmonary Disease. N. Engl. J. Med. 2004, 350, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Sin, D.D.; Miller, B.E.; Duvoix, A.; et al. Serum PARC/CCL-18 concentrations and health outcomes in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2011, 183, 1187–1192. [Google Scholar] [CrossRef] [PubMed]
- Sweef, O.; Yang, C.; Wang, Z. The Oncogenic and Tumor Suppressive Long Non-Coding RNA-microRNA-Messenger RNA Regulatory Axes Identified by Analyzing Multiple Platform Omics Data from Cr(VI)-Transformed Cells and Their Implications in Lung Cancer. Biomedicines 2022, 10, 2334. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Scoditti, E.; Massaro, M.; Garbarino, S.; Toraldo, D.M. Role of Diet in Chronic Obstructive Pulmonary Disease Prevention and Treatment. Nutrients 2019, 11, 1357. [Google Scholar] [CrossRef] [PubMed]
- Byanova, K.L.; Abelman, R.; North, C.M.; Christenson, S.A.; Huang, L. COPD in People with HIV: Epidemiology, Pathogenesis, Management, and Prevention Strategies. Int. J. Chron. Obstruct. Pulmon. Dis. 2023, 18, 2795–2817. [Google Scholar] [CrossRef] [PubMed]
- Schleich, F.; Bougard, N.; Moermans, C.; Sabbe, M.; Louis, R. Cytokine-targeted therapies for asthma and COPD. Eur. Respir. Rev. 2023, 32, 220193. [Google Scholar] [CrossRef] [PubMed]
- Kaur, M.; Chandel, J.; Malik, J.; Naura, A.S. Particulate matter in COPD pathogenesis: an overview. Inflamm. Res. 2022, 71, 797–815. [Google Scholar] [CrossRef]
- Sin, D.D.; Doiron, D.; Agusti, A.; Anzueto, A.; Barnes, P.J.; Celli, B.R.; Criner, G.J.; Halpin, D.; Han, M.K.; Martinez, F.J.; et al. Air pollution and COPD: GOLD 2023 committee report. Eur. Respir. J. 2023, 61, 2202469. [Google Scholar] [CrossRef] [PubMed]
- Neşe, A.; Samancıoğlu Bağlama, S. The Effect of Progressive Muscle Relaxation and Deep Breathing Exercises on Dyspnea and Fatigue Symptoms of COPD Patients: A Randomized Controlled Study. Holist. Nurs. Pract. 2022, 36, E18–E26. [Google Scholar] [CrossRef] [PubMed]
- Ntritsos, G.; Franek, J.; Belbasis, L.; Christou, M.A.; Markozannes, G.; Altman, P.; Fogel, R.; Sayre, T.; Ntzani, E.E.; Evangelou, E. Gender-specific estimates of COPD prevalence: a systematic review and meta-analysis. Int. J. Chron. Obstruct. Pulmon. Dis. 2018, 13, 1507–1514. [Google Scholar] [CrossRef] [PubMed]
- Segal, L.N.; Martinez, F.J. Chronic obstructive pulmonary disease subpopulations and phenotyping. J. Allergy Clin. Immunol. 2018, 141, 1961–1971. [Google Scholar] [CrossRef] [PubMed]
- Duffy, S.P.; Criner, G.J. Chronic Obstructive Pulmonary Disease: Evaluation and Management. Med. Clin. North Am. 2019, 103, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Bagdonas, E.; Raudoniute, J.; Bruzauskaite, I.; Aldonyte, R. Novel aspects of pathogenesis and regeneration mechanisms in COPD. Int. J. Chron. Obstruct. Pulmon. Dis. 2015, 10, 995–1013. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.S.; Liu, Z.; Sweef, O.; Saeed, A.F.; Kluz, T.; Costa, M.; Shroyer, K.R.; Kondo, K.; Wang, Z.; Yang, C. Hexavalent chromium exposure activates the non-canonical nuclear factor kappa B pathway to promote immune checkpoint protein programmed death-ligand 1 expression and lung carcinogenesis. Cancer Lett. 2024, 589, 216827. [Google Scholar] [CrossRef] [PubMed]
- Murgia, N.; Gambelunghe, A. Occupational COPD-The most under-recognized occupational lung disease? Respirology 2022, 27, 399–410. [Google Scholar] [CrossRef] [PubMed]
- GBD 2019 Chronic Respiratory Diseases Collaborators. Global burden of chronic respiratory diseases and risk factors, 1990-2019: an update from the Global Burden of Disease Study 2019. EClinicalMedicine 2023, 59, 101936. [CrossRef]
- Rabe, K.F.; Watz, H. Chronic obstructive pulmonary disease. Lancet 2017, 389, 1931–1940. [Google Scholar] [CrossRef]
- Lakshmanan, S.; Jankowich, M.; Wu, W.C.; Blackshear, C.; Abbasi, S.; Choudhary, G. Gender Differences in Risk Factors Associated With Pulmonary Artery Systolic Pressure, Heart Failure, and Mortality in Blacks: Jackson Heart Study. J. Am. Heart Assoc. 2020, 9, e013034. [Google Scholar] [CrossRef] [PubMed]
- Yang, I.A.; Jenkins, C.R.; Salvi, S.S. Chronic obstructive pulmonary disease in never-smokers: risk factors, pathogenesis, and implications for prevention and treatment. Lancet Respir. Med. 2022, 10, 497–511. [Google Scholar] [CrossRef] [PubMed]
- Mannino, D.M.; Buist, A.S. Global burden of COPD: risk factors, prevalence, and future trends. Lancet 2007, 370, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Salvi, S.S.; Barnes, P.J. Chronic obstructive pulmonary disease in non-smokers. Lancet 2009, 374, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Easter, M.; Bollenbecker, S.; Barnes, J.W.; Krick, S. Targeting Aging Pathways in Chronic Obstructive Pulmonary Disease. Int. J. Mol. Sci. 2020, 21, 6924. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Sun, S.; Tang, R.; Qiu, H.; Huang, Q.; Mason, T.G.; Tian, L. Major air pollutants and risk of COPD exacerbations: a systematic review and meta-analysis. Int. J. Chron. Obstruct. Pulmon. Dis. 2016, 11, 3079–3091. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.S.; Liu, Z.; Sweef, O.; Xie, J.; Chen, J.; Zhu, H.; Zeidler-Erdely, P.C.; Yang, C.; Wang, Z. Long noncoding RNA ABHD11-AS1 interacts with SART3 and regulates CD44 RNA alternative splicing to promote lung carcinogenesis. Environ Int. 2024, 185, 108494. [Google Scholar] [CrossRef] [PubMed]
- Bekkat-Berkani, R.; Wilkinson, T.; Buchy, P.; Dos Santos, G.; Stefanidis, D.; Devaster, J.M. Seasonal influenza vaccination in patients with COPD: a systematic literature review. BMC Pulm. Med. 2017, 17, 79. [Google Scholar] [CrossRef] [PubMed]
- Odimba, U.; Senthilselvan, A.; Farrell, J.; Gao, Z. Current Knowledge of Asthma-COPD Overlap (ACO) Genetic Risk Factors, Characteristics, and Prognosis. COPD 2021, 18, 585–595. [Google Scholar] [CrossRef]
- Silverman, E.K. Genetics of COPD. Annu. Rev. Physiol. 2020, 82, 413–431. [Google Scholar] [CrossRef]
- Huang, X.; Mu, X.; Deng, L.; Fu, A.; Pu, E.; Tang, T.; Kong, X. The etiologic origins for chronic obstructive pulmonary disease. Int. J. Chron. Obstruct. Pulmon. Dis. 2019, 14, 1139–1158. [Google Scholar] [CrossRef] [PubMed]
- Kahnert, K.; Jörres, R.A.; Behr, J.; Welte, T. The Diagnosis and Treatment of COPD and Its Comorbidities. Dtsch. Arztebl. Int. 2023, 120, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Şerifoğlu, İ.; Ulubay, G. The methods other than spirometry in the early diagnosis of COPD. Tuberk Toraks 2019, 67, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Huprikar, N.A.; Skabelund, A.J.; Bedsole, V.G.; Sjulin, T.J.; Karandikar, A.V.; Aden, J.K.; Morris, M.J. Comparison of Forced and Slow Vital Capacity Maneuvers in Defining Airway Obstruction. Respir. Care 2019, 64, 786–792. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; He, X.; Xu, Q.F.; Wang, H.Y.; Casaburi, R. Increased difference between slow and forced vital capacity is associated with reduced exercise tolerance in COPD patients. BMC Pulm. Med. 2014, 14, 16. [Google Scholar] [CrossRef] [PubMed]
- Christenson, S.A.; Smith, B.M.; Bafadhel, M.; Putcha, N. Chronic obstructive pulmonary disease. Lancet 2022, 399, 2227–2242. [Google Scholar] [CrossRef] [PubMed]
- van den Bemt, L.; Schermer, T.; Smeele, I.; Bischoff, E.; Jacobs, A.; Grol, R.; van Weel, C. Monitoring of patients with COPD: a review of current guidelines' recommendations. Respir. Med. 2008, 102, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Jarroux, J.; Morillon, A.; Pinskaya, M. History, Discovery, and Classification of lncRNAs. Adv. Exp. Med. Biol. 2017, 1008, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Fu, H.; Wu, Y.; Zheng, X. Function of lncRNAs and approaches to lncRNA-protein interactions. Sci. China Life Sci. 2013, 56, 876–885. [Google Scholar] [CrossRef]
- Bridges, M.C.; Daulagala, A.C.; Kourtidis, A. LNCcation: lncRNA localization and function. J. Cell Biol. 2021, 220, e202009045. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, D.; Li, D.; Zheng, Z.; Xu, J.; Liang, X.; Zhang, C.; Wang, S.; Wang, J.; Lu, W. Long non-coding RNA expression patterns in lung tissues of chronic cigarette smoke induced COPD mouse model. Sci. Rep. 2018, 8, 7609. [Google Scholar] [CrossRef] [PubMed]
- Negewo, N.A.; Gibson, P.G.; McDonald, V.M. COPD and its comorbidities: Impact, measurement and mechanisms. Respirology 2015, 20, 1160–1171. [Google Scholar] [CrossRef] [PubMed]
- Uwagboe, I.; Adcock, I.M.; Lo Bello, F.; Caramori, G.; Mumby, S. New drugs under development for COPD. Minerva Med. 2022, 113, 471–496. [Google Scholar] [CrossRef] [PubMed]
- Raherison, C.; Girodet, P.O. Epidemiology of COPD. Eur. Respir. Rev. 2009, 18, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Ko, F.W.; Chan, K.P.; Hui, D.S.; Goddard, J.R.; Shaw, J.G.; Reid, D.W.; Yang, I.A. Acute exacerbation of COPD. Respirology 2016, 21, 1152–1165. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhu, Y.; Wang, R. Long noncoding RNAs in respiratory diseases. Histol. Histopathol. 2018, 33, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Poulet, C.; Njock, M.S.; Moermans, C.; Louis, E.; Louis, R.; Malaise, M.; Guiot, J. Exosomal Long Non-Coding RNAs in Lung Diseases. Int. J. Mol. Sci. 2020, 21, 3580. [Google Scholar] [CrossRef] [PubMed]
- Soni, D.K.; Biswas, R. Role of Non-Coding RNAs in Post-Transcriptional Regulation of Lung Diseases. Front. Genet. 2021, 12, 767348. [Google Scholar] [CrossRef] [PubMed]
- Booton, R.; Lindsay, M.A. Emerging role of MicroRNAs and long noncoding RNAs in respiratory disease. Chest 2014, 146, 193–204. [Google Scholar] [CrossRef]
- Sweef, O.; Zaabout, E.; Bakheet, A.; Halawa, M.; Gad, I.; Akela, M.; Tousson, E.; Abdelghany, A.; Furuta, S. Unraveling Therapeutic Opportunities and the Diagnostic Potential of microRNAs for Human Lung Cancer. Pharmaceutics. 2023, 15, 2061. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fang, D.; Ou, X.; Sun, K.; Zhou, X.; Li, Y.; Shi, P.; Zhao, Z.; He, Y.; Peng, J.; Xu, J. m6A modification-mediated lncRNA TP53TG1 inhibits gastric cancer progression by regulating CIP2A stability. Cancer Sci. 2022, 113, 4135–4150. [Google Scholar] [CrossRef] [PubMed]
- Herman, A.B.; Tsitsipatis, D.; Gorospe, M. Integrated lncRNA function upon genomic and epigenomic regulation. Mol. Cell 2022, 82, 2252–2266. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Sarkissyan, M.; Ogah, O.; Kim, J.; Vadgama, J.V. Expression of MALAT1 Promotes Trastuzumab Resistance in HER2 Overexpressing Breast Cancers. Cancers 2020, 12, 1918. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Piao, H.L.; Kim, B.J.; Yao, F.; Han, Z.; Wang, Y.; Xiao, Z.; Siverly, A.N.; Lawhon, S.E.; Ton, B.N.; et al. Long noncoding RNA MALAT1 suppresses breast cancer metastasis. Nat. Genet. 2018, 50, 1705–1715. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; Wang, Y.; Brzoska, P.; Kong, B.; Li, R.; West, R.B.; van de Vijver, M.J.; Sukumar, S.; Chang, H.Y. Long Non-Coding RNA HOTAIR Reprograms Chromatin State to Promote Cancer Metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Shen, Y.; Chen, B.; Wu, Y.; Jia, L.; Li, Y.; Zhu, Y.; Yan, Y.; Li, M.; Chen, R.; Guo, L.; Chen, X.; Chen, Q. H3K27me3 Induces Multidrug Resistance in Small Cell Lung Cancer by Affecting HOXA1 DNA Methylation via Regulation of the lncRNA HOTAIR. Ann. Transl. Med. 2018, 6, 440. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Shaker, O.G.; Khalifa, A.A.; Ezzat, E.M.; Elghobary, H.A.; Abdel Mawla, T.S.; Elkhateeb, A.F.; Elebiary, A.M.A.; Elamir, A.M. LncRNAs NEAT1, HOTAIR, and GAS5 Expression in Hypertensive and Non-Hypertensive Associated Cerebrovascular Stroke Patients, and Its Link to Clinical Characteristics and Severity Score of the Disease. Noncoding RNA Res. 2022, 8, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ren, X.; Yuan, Y.; Yuan, B.S. Downregulated lncRNA GAS5 and Upregulated miR-21 Lead to Epithelial-Mesenchymal Transition and Lung Metastasis of Osteosarcomas. Front. Cell Dev. Biol. 2021, 9, 707693. [Google Scholar] [CrossRef]
- Hao, X.; Wei, H. LncRNA H19 alleviates sepsis-induced acute lung injury by regulating the miR-107/TGFBR3 axis. BMC Pulm. Med. 2022, 22, 371. [Google Scholar] [CrossRef]
- Zhang, R.; Pan, T.; Xiang, Y.; Zhang, M.; Xie, H.; Liang, Z.; Chen, B.; Xu, C.; Wang, J.; Huang, X.; et al. Curcumenol triggered ferroptosis in lung cancer cells via lncRNA H19/miR-19b-3p/FTH1 axis. Bioact. Mater. 2021, 13, 23–36. [Google Scholar] [CrossRef]
- Rojas-Cruz, A.F.; Bermúdez-Santana, C.I. Computational Prediction of RNA-RNA Interactions between Small RNA Tracks from Betacoronavirus Nonstructural Protein 3 and Neurotrophin Genes during Infection of an Epithelial Lung Cancer Cell Line: Potential Role of Novel Small Regulatory RNA. Viruses 2023, 15, 1647. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Shen, Q.; Zhang, X.; Yang, C.; Cui, S.; Sun, Y.; Wang, L.; Fan, X.; Xu, S. The Long Non-Coding RNA XIST Controls Non-Small Cell Lung Cancer Proliferation and Invasion by Modulating miR-186-5p. Cell Physiol. Biochem. 2017, 41, 2221–2229. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Tang, Y.; Wang, T.; Yang, Y.; Zhang, Y.; Wang, R.; Zhang, Y.; Li, Y.; Wu, M.; Tang, M.; et al. Chronic pulmonary bacterial infection facilitates breast cancer lung metastasis by recruiting tumor-promoting MHCIIhi neutrophils. Signal Transduct. Target. Ther. 2023, 8, 296. [Google Scholar] [CrossRef] [PubMed]
- Warren, G.W.; Cummings, K.M. Tobacco and lung cancer: risks, trends, and outcomes in patients with cancer. Am. Soc. Clin. Oncol. Educ. Book 2013, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Lin, H.; Su, Q.; Li, C. Cuproptosis-related lncRNA predict prognosis and immune response of lung adenocarcinoma. World J. Surg. Oncol. 2022, 20, 275. [Google Scholar] [CrossRef] [PubMed]
- Desind, S.Z.; Iacona, J.R.; Yu, C.Y.; Mitrofanova, A.; Lutz, C.S. PACER lncRNA regulates COX-2 expression in lung cancer cells. Oncotarget 2022, 13, 291–306. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Jiang, X.; Duan, L.; Xiong, Q.; Yuan, Y.; Liu, P.; Jiang, L.; Shen, Q.; Zhao, S.; Yang, C.; et al. LncRNA PKMYT1AR promotes cancer stem cell maintenance in non-small cell lung cancer via activating Wnt signaling pathway. Mol. Cancer 2021, 20, 156. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Dang, H.X.; Lim, D.A.; Feng, F.Y.; Maher, C.A. Long noncoding RNAs in cancer metastasis. Nat. Rev. Cancer 2021, 21, 446–460. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Xu, A.; Li, M.; Xia, X.; Li, P.; Han, R.; Fei, G.; Zhou, S.; Wang, R. Effect of Methylation Status of lncRNA-MALAT1 and MicroRNA-146a on Pulmonary Function and Expression Level of COX2 in Patients With Chronic Obstructive Pulmonary Disease. Front. Cell Dev. Biol. 2021, 9, 667624. [Google Scholar] [CrossRef]
- Sundar, I.K.; Yao, H.; Rahman, I. Oxidative stress and chromatin remodeling in chronic obstructive pulmonary disease and smoking-related diseases. Antioxid. Redox Signal. 2013, 18, 1956–1971. [Google Scholar] [CrossRef]
- Zhou, D.; Wang, J.; Liu, X. Development of six immune-related lncRNA signature prognostic model for smoking-positive lung adenocarcinoma. J. Clin. Lab. Anal. 2022, 36, e24467. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Gao, F.; Mao, Z.; Xiao, Y.; Yuan, L.; Huang, Z.; Lv, Q.; Qin, C.; Du, M.; Zhang, Z.; et al. LncRNA BCCE4 Genetically Enhances the PD-L1/PD-1 Interaction in Smoking-Related Bladder Cancer by Modulating miR-328-3p-USP18 Signaling. Adv. Sci. 2023, 10, e2303473. [Google Scholar] [CrossRef] [PubMed]
- Maremanda, K.P.; Sundar, I.K.; Rahman, I. Role of inner mitochondrial protein OPA1 in mitochondrial dysfunction by tobacco smoking and in the pathogenesis of COPD. Redox Biol. 2021, 45, 102055. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, P.; Wu, C.W.; Pham, A.; Zeki, A.A.; Royer, C.M.; Kodavanti, U.P.; Takeuchi, M.; Bayram, H.; Pinkerton, K.E. Animal models and mechanisms of tobacco smoke-induced chronic obstructive pulmonary disease (COPD). J. Toxicol. Environ. Health B Crit. Rev. 2023, 26, 275–305. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Ding, Y.; Shi, T.; He, W.; Feng, J.; Mei, Z.; Chen, X.; Feng, X.; Zhang, X.; Jie, Z. Long noncoding RNA GAS5 attenuates cigarette smoke-induced airway remodeling by regulating miR-217-5p/PTEN axis. Acta Biochim. Biophys. Sin. (Shanghai) 2022, 54, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Li, R.; Piao, T.H.; Wang, C.L.; Wu, X.L.; Cai, H.Y. Pathological Mechanism and Targeted Drugs of COPD. Int. J. Chron. Obstruct. Pulmon. Dis. 2022, 17, 1565–1575. [Google Scholar] [CrossRef] [PubMed]
- Lewis, A.; Lee, J.Y.; Donaldson, A.V.; Natanek, S.A.; Vaidyanathan, S.; Man, W.D.; Hopkinson, N.S.; Sayer, A.A.; Patel, H.P.; Cooper, C.; et al. Increased expression of H19/miR-675 is associated with a low fat-free mass index in patients with COPD. J. Cachexia Sarcopenia Muscle 2016, 7, 330–344. [Google Scholar] [CrossRef] [PubMed]
- Forder, A.; Zhuang, R.; Souza, V.G.P.; Brockley, L.J.; Pewarchuk, M.E.; Telkar, N.; Stewart, G.L.; Benard, K.; Marshall, E.A.; Reis, P.P.; et al. Mechanisms Contributing to the Comorbidity of COPD and Lung Cancer. Int. J. Mol. Sci. 2023, 24, 2859. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Geng, S.; Jiang, H. Long noncoding RNAs antisense noncoding RNA in the INK4 locus (ANRIL) correlates with lower acute exacerbation risk, decreased inflammatory cytokines, and mild GOLD stage in patients with chronic obstructive pulmonary disease. J. Clin. Lab. Anal. 2019, 33, e22678. [Google Scholar] [CrossRef] [PubMed]
- Araya, J.; Tsubouchi, K.; Sato, N.; Ito, S.; Minagawa, S.; Hara, H.; Hosaka, Y.; Ichikawa, A.; Saito, N.; Kadota, T.; et al. PRKN-regulated mitophagy and cellular senescence during COPD pathogenesis. Autophagy 2019, 15, 510–526. [Google Scholar] [CrossRef]
- Poulet, C.; Njock, M.S.; Moermans, C.; Louis, E.; Louis, R.; Malaise, M.; Guiot, J. Exosomal Long Non-Coding RNAs in Lung Diseases. Int. J. Mol. Sci. 2020, 21, 3580. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.J.; Huang, H.B.; Shen, H.B.; Chen, W.; Yang, Z.H. Role of long non-coding RNA MALAT1 in chronic obstructive pulmonary disease. Exp. Ther. Med. 2020, 20, 2691–2697. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Sen, C.; Peters, K.; Frizzell, R.A.; Biswas, R. Comparative analyses of long non-coding RNA profiles in vivo in cystic fibrosis lung airway and parenchyma tissues. Respir. Res. 2019, 20, 284. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Tang, F. The potency of lncRNA MALAT1/miR-155/CTLA4 axis in altering Th1/Th2 balance of asthma. Biosci. Rep. 2020, 40, BSR20190397. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.P.; Liu, Z.B.; Wu, M.; Ge, Y.P.; Zhang, Q. Effect of lncRNA MALAT1 expression on survival status of elderly patients with severe pneumonia. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 3959–3964. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Philp, A.M.; Corte, T.; Travis, M.A.; Schilter, H.; Hansbro, N.G.; Burns, C.J.; Eapen, M.S.; Sohal, S.S.; Burgess, J.K.; et al. Therapeutic targets in lung tissue remodelling and fibrosis. Pharmacol. Ther. 2021, 225, 107839. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, A.; Sangiorgi, C.; Gnemmi, I.; Casolari, P.; Brun, P.; Ricciardolo, F.L.M.; Contoli, M.; Papi, A.; Maniscalco, P.; Ruggeri, P.; et al. TGF-β Signaling Pathways in Different Compartments of the Lower Airways of Patients With Stable COPD. Chest 2018, 153, 851–862. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, A.; Sangiorgi, C.; Gnemmi, I.; Casolari, P.; Brun, P.; Ricciardolo, F.L.M.; Contoli, M.; Papi, A.; Maniscalco, P.; Ruggeri, P.; Girbino, G.; Cappello, F.; Pavlides, S.; Guo, Y.; Chung, K.F.; Barnes, P.J.; Adcock, I.M.; Balbi, B.; Caramori, G. TGF-β Signaling Pathways in Different Compartments of the Lower Airways of Patients with Stable COPD. Chest 2018, 153, 851–862. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhang, H.; Tang, J.; Wang, R. Long non-coding RNA FAM230B is a novel prognostic and diagnostic biomarker for lung adenocarcinoma. Bioengineered 2022, 13, 7919–7925. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Huang, Q.; He, F. Correlation of small nucleolar RNA host gene 16 with acute respiratory distress syndrome occurrence and prognosis in sepsis patients. J. Clin. Lab. Anal. 2022, 36, e24516. [Google Scholar] [CrossRef]
- Mo, R.; Li, J.; Chen, Y.; Ding, Y. lncRNA GAS5 promotes pyroptosis in COPD by functioning as a ceRNA to regulate the miR 223 3p/NLRP3 axis. Mol. Med. Rep. 2022, 26, 219. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Zhang, C.F.; Guo, J.L.; Su, J.L.; Guo, Z.K.; Li, H.Y. Involvement of NEAT1/PINK1-mediated mitophagy in chronic obstructive pulmonary disease induced by cigarette smoke or PM2. 5. Ann. Transl. Med. 2022, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Jen, J.; Tang, Y.A.; Lu, Y.H.; Lin, C.C.; Lai, W.W.; Wang, Y.C. Oct4 transcriptionally regulates the expression of long non-coding RNAs NEAT1 and MALAT1 to promote lung cancer progression. Mol. Cancer 2017, 16, 104. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Kelava, L.; Zhang, L.; Kiss, I. Microarray data analysis to identify miRNA biomarkers and construct the lncRNA-miRNA-mRNA network in lung adenocarcinoma. Medicine (Baltimore) 2022, 101, e30393. [Google Scholar] [CrossRef] [PubMed]
- Tokgun, O.; Tokgun, P.E.; Inci, K.; Akca, H. lncRNAs as Potential Targets in Small Cell Lung Cancer: MYC-dependent Regulation. Anticancer Agents Med. Chem. 2020, 20, 2074–2081. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.F.; Deng, J.M. Emerging Role of Long Noncoding RNAs in Asthma. Comb. Chem. High Throughput Screen. 2023, 26, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Liu, Y.; Li, M.; Zhou, H.; Cao, L.; Zhang, X.; Li, Y. Senescence associated long non-coding RNA 1 regulates cigarette smoke-induced senescence of type II alveolar epithelial cells through sirtuin-1 signaling. J. Int. Med. Res. 2021, 49, 300060520986049. [Google Scholar] [CrossRef] [PubMed]
- Ezegbunam, W.; Foronjy, R. Posttranscriptional control of airway inflammation. Wiley Interdiscip. Rev. RNA 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Min, L.; Qiu, X.; Wu, X.; Liu, C.; Ma, J.; Zhang, D.; Zhu, L. Biological Function of Long Non-coding RNA (lncRNA) Xist. Front. Cell Dev. Biol. 2021, 9, 645647. [Google Scholar] [CrossRef]
- Huang, M.; Wang, H.; Hu, X.; Cao, X. lncRNA MALAT1 binds chromatin remodeling subunit BRG1 to epigenetically promote inflammation-related hepatocellular carcinoma progression. Oncoimmunology 2018, 8, e1518628. [Google Scholar] [CrossRef]
- Dai, J.; Qu, T.; Yin, D.; Cui, Y.; Zhang, C.; Zhang, E.; Guo, R. LncRNA LINC00969 promotes acquired gefitinib resistance by epigenetically suppressing of NLRP3 at transcriptional and posttranscriptional levels to inhibit pyroptosis in lung cancer. Cell Death Dis. 2023, 14, 312. [Google Scholar] [CrossRef] [PubMed]
- Xing, C.; Sun, S.G.; Yue, Z.Q.; Bai, F. Role of lncRNA LUCAT1 in cancer. Biomed. Pharmacother. 2021, 134, 111158. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Chen, L.; Piao, S.; Wang, Y.; Li, Z.; Lin, Y.; Tang, X.; Zhang, H.; Zhang, H.; Wang, X. M6A RNA methylation-mediated RMRP stability renders proliferation and progression of non-small cell lung cancer through regulating TGFBR1/SMAD2/SMAD3 pathway. Cell Death Differ. 2023, 30, 605–617. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, S.Q.; Zhu, J.B.; Wang, L.N.; Lin, H.; Li, L.F.; Cheng, Y.D.; Duan, C.J.; Zhang, C.F. LncRNA CALML3-AS1 modulated by m6A modification induces BTNL9 methylation to drive non-small-cell lung cancer progression. Cancer Gene Ther. 2023, 30, 1649–1662. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, E.; Sznajder, J.I. Epigenetic regulation of muscle phenotype and adaptation: a potential role in COPD muscle dysfunction. J. Appl. Physiol. (1985) 2013, 114, 1263–1272. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Huang, K.; Chang, C.; Chu, X.; Zhang, K.; Li, B.; Yang, T. Serum Proteomic Profiling in Patients with Chronic Obstructive Pulmonary Disease. Int. J. Chron. Obstruct. Pulmon. Dis. 2023, 18, 1623–1635. [Google Scholar] [CrossRef] [PubMed]
- Salvato, I.; Ricciardi, L.; Nucera, F.; Nigro, A.; Dal Col, J.; Monaco, F.; Caramori, G.; Stellato, C. RNA-Binding Proteins as a Molecular Link between COPD and Lung Cancer. COPD 2023, 20, 18–30. [Google Scholar] [CrossRef] [PubMed]
- An, L.; Xia, H.; Zheng, W.; Hua, L. Comparison of cell subsets in chronic obstructive pulmonary disease and controls based on single-cell transcriptome sequencing. Technol. Health Care 2023, 31, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zheng, H.; Zhang, H.; Ma, W.; Wang, F.; Liu, C.; He, S. Increased interleukin (IL)-8 and decreased IL-17 production in chronic obstructive pulmonary disease (COPD) provoked by cigarette smoke. Cytokine 2011, 56, 717–725. [Google Scholar] [CrossRef]
- Liu, F.; Zhang, X.; Du, W.; Du, J.; Chi, Y.; Sun, B.; Song, Z.; Shi, J. Diagnosis values of IL-6 and IL-8 levels in serum and bronchoalveolar lavage fluid for invasive pulmonary aspergillosis in chronic obstructive pulmonary disease. J. Investig. Med. 2021, 69, 1344–1349. [Google Scholar] [CrossRef]
- Matera, M.G.; Calzetta, L.; Cazzola, M. TNF-alpha inhibitors in asthma and COPD: we must not throw the baby out with the bath water. Pulm. Pharmacol. Ther. 2010, 23, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Belvisi, M.G.; Bottomley, K.M. The role of matrix metalloproteinases (MMPs) in the pathophysiology of chronic obstructive pulmonary disease (COPD): a therapeutic role for inhibitors of MMPs? Inflamm. Res. 2003, 52, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Wu, Z.; Wu, T.; Huang, Y.; Cheng, Z.; Li, X.; Sun, T.; Xie, X.; Zhou, Y.; Du, Z. Tumor-suppressive function of long noncoding RNA MALAT1 in glioma cells by downregulation of MMP2 and inactivation of ERK/MAPK signaling. Cell Death Dis. 2016, 7, e2123. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Li, H.; Qiu, T.; Dai, J.; Zhang, Y.; Chen, J.; Cai, H. Loss of PTEN induces lung fibrosis via alveolar epithelial cell senescence depending on NF-κB activation. Aging Cell 2019, 18, e12858. [Google Scholar] [CrossRef] [PubMed]
- Guzy, R. Fibroblast Growth Factor Inhibitors in Lung Fibrosis: Friends or Foes? Am. J. Respir. Cell Mol. Biol. 2020, 63, 273–274. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.Y.; Oliver, B. Innate Immune Reprogramming in Chronic Obstructive Pulmonary Disease: New Mechanisms for Old Questions. Am. J. Respir. Cell Mol. Biol. 2023, 68, 470–471. [Google Scholar] [CrossRef] [PubMed]
- Park, E.G.; Pyo, S.J.; Cui, Y.; Yoon, S.H.; Nam, J.W. Tumor immune microenvironment lncRNAs. Brief Bioinform. 2022, 23, bbab504. [Google Scholar] [CrossRef]
- Lv, X.; Zhang, X.Y.; Zhang, Q.; Nie, Y.J.; Luo, G.H.; Fan, X.; Yang, S.; Zhao, Q.H.; Li, J.Q. lncRNA NEAT1 aggravates sepsis-induced lung injury by regulating the miR-27a/PTEN axis. Lab Invest. 2021, 101, 1371–1381. [Google Scholar] [CrossRef] [PubMed]
- Mayama, T.; Marr, A.K.; Kino, T. Differential Expression of Glucocorticoid Receptor Noncoding RNA Repressor Gas5 in Autoimmune and Inflammatory Diseases. Horm. Metab. Res. 2016, 48, 550–557. [Google Scholar] [CrossRef]
- Mayama, T.; Marr, A.K.; Kino, T. Differential Expression of Glucocorticoid Receptor Noncoding RNA Repressor Gas5 in Autoimmune and Inflammatory Diseases. Horm. Metab. Res. 2016, 48, 550–557. [Google Scholar] [CrossRef]
- Chen, L.; Huang, S.; Huang, J.; Chen, Q.; Zhuang, Q. Role and Mechanism of Exosome-Derived Long Noncoding RNA HOTAIR in Lung Cancer. ACS Omega 2021, 6, 17217–17227. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Zhou, Q.; Li, J.; Yang, X.; Wang, R.; Wang, X.; Xu, M.; Han, L.; Wu, W.; Wang, S. Solamargine enhanced gefitinib antitumor effect via regulating MALAT1/miR-141-3p/Sp1/IGFBP1 signaling pathway in non-small cell lung cancer. Carcinogenesis 2023, 44, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Zhou, Q.; Li, J.; Yang, X.; Wang, R.; Wang, X.; Xu, M.; Han, L.; Wu, W.; Wang, S. Solamargine enhanced gefitinib antitumor effect via regulating MALAT1/miR-141-3p/Sp1/IGFBP1 signaling pathway in non-small cell lung cancer. Carcinogenesis 2023, 44, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Lan, Y.; Chen, Y.; Zuo, F.; Gong, Y.; Luo, G.; Peng, Y.; Yuan, Z. LncRNA GAS5 suppresses inflammatory responses by inhibiting HMGB1 release via miR-155-5p/SIRT1 axis in sepsis. Eur. J. Pharmacol. 2023, 942, 175520. [Google Scholar] [CrossRef]
- Li, J.; Liu, S. LncRNA GAS5 suppresses inflammatory responses and apoptosis of alveolar epithelial cells by targeting miR-429/DUSP1. Exp. Mol. Pathol. 2020, 113, 104357. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, J.; Sun, Y.; Zhou, Q.; Ding, X.; Chen, X. Chinese herbal compound Huangqin Qingrechubi capsule reduces lipid metabolism disorder and inflammatory response in gouty arthritis via the LncRNA H19/APN/PI3K/AKT cascade. Pharm. Biol. 2023, 61, 541–555. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xie, Y.; He, Q.; Geng, Y.; Xu, J. LncRNA-Cox2 regulates macrophage polarization and inflammatory response through the CREB-C/EBPβ signaling pathway in septic mice. Int. Immunopharmacol. 2021, 101, 108347. [Google Scholar] [CrossRef] [PubMed]
- Xue, Q.; Wang, Y.; Zheng, Q.; Chen, L.; Jin, Y.; Shen, X.; Li, Y. Construction of a prognostic immune-related lncRNA model and identification of the immune microenvironment in middle- or advanced-stage lung squamous carcinoma patients. Heliyon 2022, 8, e09521. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.; Xie, L.; Zhao, K.; Zhang, Q.; Zhou, M.; He, P. Cigarette smoke-induced lung inflammation in COPD mediated via LTB4/BLT1/SOCS1 pathway. Int. J. Chron. Obstruct. Pulmon. Dis. 2015, 11, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Barta, I.; Paska, C.; Antus, B. Sputum Cytokine Profiling in COPD: Comparison Between Stable Disease and Exacerbation. Int. J. Chron. Obstruct. Pulmon. Dis. 2022, 17, 1897–1908. [Google Scholar] [CrossRef]
- Zhang, L.; Gu, H.; Gu, Y.; Zeng, X. Association between TNF-α -308 G/A polymorphism and COPD susceptibility: a meta-analysis update. Int. J. Chron. Obstruct. Pulmon. Dis. 2016, 11, 1367–1379. [Google Scholar] [CrossRef] [PubMed]
- Saito, A.; Horie, M.; Nagase, T. TGF-β Signaling in Lung Health and Disease. Int. J. Mol. Sci. 2018, 19, 2460. [Google Scholar] [CrossRef] [PubMed]
- Christopoulou, M.E.; Papakonstantinou, E.; Stolz, D. Matrix Metalloproteinases in Chronic Obstructive Pulmonary Disease. Int. J. Mol. Sci. 2023, 24, 3786. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.W.; Cai, Q.Q.; Yang, Y.; Dong, S.; Liu, Y.Y.; Chen, Z.Y.; Kang, C.L.; Qi, B.; Dong, Y.W.; Wu, W.; Zhuang, L.P.; Shen, Y.H.; Meng, Z.Q.; Wu, X.Z. LncRNA BC promotes lung adenocarcinoma progression by modulating IMPAD1 alternative splicing. Clin. Transl. Med. 2023, 13, e1129. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Jose, C.C.; Cuddapah, S. Epithelial-mesenchymal transition: Insights into nickel-induced lung diseases. Semin. Cancer Biol. 2021, 76, 99–109. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Ji, X.; Cao, L.; Wang, Z.; Wang, X.; Li, X.M.; Miao, M. Medicine Targeting Epithelial-Mesenchymal Transition to Treat Airway Remodeling and Pulmonary Fibrosis Progression. Can. Respir. J. 2023, 2023, 3291957. [Google Scholar] [CrossRef] [PubMed]
- Qian, G.; Liao, Q.; Li, G.; Yin, F. miR-378 associated with proliferation, migration and apoptosis properties in A549 cells and targeted NPNT in COPD. PeerJ 2022, 10, e14062. [Google Scholar] [CrossRef] [PubMed]
- Demedts, I.K.; Demoor, T.; Bracke, K.R.; Joos, G.F.; Brusselle, G.G. Role of apoptosis in the pathogenesis of COPD and pulmonary emphysema. Respir. Res. 2006, 7, 53. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.C.; Lin, T.L.; Chen, T.W.; Kuo, Y.L.; Chang, C.J.; Wu, T.R.; Shu, C.C.; Tsai, Y.H.; Swift, S.; Lu, C.C. Gut microbiota modulates COPD pathogenesis: role of anti-inflammatory Parabacteroides goldsteinii lipopolysaccharide. Gut 2022, 71, 309–321. [Google Scholar] [CrossRef]
- Wang, S.M.; Liu, G.Q.; Xian, H.B.; Si, J.L.; Qi, S.X.; Yu, Y.P. LncRNA NEAT1 alleviates sepsis-induced myocardial injury by regulating the TLR2/NF-κB signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 4898–4907. [Google Scholar] [CrossRef]
- Wiegman, C.H.; Li, F.; Ryffel, B.; Togbe, D.; Chung, K.F. Oxidative Stress in Ozone-Induced Chronic Lung Inflammation and Emphysema: A Facet of Chronic Obstructive Pulmonary Disease. Front. Immunol. 2020, 11, 1957. [Google Scholar] [CrossRef]
- Liu, K.; Hua, S.; Song, L. PM2.5 Exposure and Asthma Development: The Key Role of Oxidative Stress. Oxid. Med. Cell. Longev. 2022, 2022, 3618806. [Google Scholar] [CrossRef] [PubMed]
- Salazar, L.M.; Herrera, A.M. Fibrotic response of tissue remodeling in COPD. Lung 2011, 189, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wu, D.; Chen, J.; Chen, J.; Qiu, F.; Li, Y.; Liu, L.; Cao, Y.; Yang, B.; Zhou, Y.; Lu, J. A functional CNVR_3425. 1 damping lincRNA FENDRR increases lifetime risk of lung cancer and COPD in Chinese. Carcinogenesis 2018, 39, 347–359. [Google Scholar] [CrossRef] [PubMed]
- Kume, H.; Yamada, R.; Sato, Y.; Togawa, R. Airway Smooth Muscle Regulated by Oxidative Stress in COPD. Antioxidants 2023, 12, 142. [Google Scholar] [CrossRef]
- Groot, M.; Zhang, D.; Jin, Y. Long Non-Coding RNA Review and Implications in Lung Diseases. JSM Bioinform. Genom. Proteom. 2018, 3, 1033. [Google Scholar] [CrossRef]
- Xiao, J.H.; Hao, Q.Y.; Wang, K.; Paul, J.; Wang, Y.X. Emerging Role of MicroRNAs and Long Noncoding RNAs in Healthy and Diseased Lung. Adv. Exp. Med. Biol. 2017, 967, 343–359. [Google Scholar] [CrossRef] [PubMed]
- de Groot, P.; Munden, R.F. Lung cancer epidemiology, risk factors, and prevention. Radiol. Clin. North Am. 2012, 50, 863–876. [Google Scholar] [CrossRef] [PubMed]
- Ginn, L.; Shi, L.; Montagna, M.; Garofalo, M. LncRNAs in Non-Small-Cell Lung Cancer. Noncoding RNA 2020, 6, 25. [Google Scholar] [CrossRef]
- Wang, C.; Zhou, J.; Wang, J.; Li, S.; Fukunaga, A.; Yodoi, J.; Tian, H. Progress in the mechanism and targeted drug therapy for COPD. Signal Transduct. Target. Ther. 2020, 5, 248. [Google Scholar] [CrossRef]
- Zhao, Y.; Feng, C.; Li, Y.; Ma, Y.; Cai, R. LncRNA H19 promotes lung cancer proliferation and metastasis by inhibiting miR-200a function. Mol. Cell. Biochem. 2019, 460, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Roh, J.; Kim, B.; Im, M.; Jang, W.; Chae, Y.; Kang, J.; Youn, B.; Kim, W. MALAT1-regulated gene expression profiling in lung cancer cell lines. BMC Cancer 2023, 23, 818. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, S.; Zhao, M.; Chen, F. LncRNA MALAT1 accelerates non-small cell lung cancer progression via regulating miR-185-5p/MDM4 axis. Cancer Med. 2020, 9, 9138–9149. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Mo, J.; Xie, Y.; Wang, C. Ultrasound microbubbles-mediated miR-216b affects MALAT1-miRNA axis in non-small cell lung cancer cells. Tissue Cell 2022, 74, 101703. [Google Scholar] [CrossRef] [PubMed]
- Attaway, A.H.; Bellar, A.; Welch, N.; Sekar, J.; Kumar, A.; Mishra, S.; Hatipoğlu, U.; McDonald, M.L.; Regan, E.A.; Smith, J.D.; et al. Gene polymorphisms associated with heterogeneity and senescence characteristics of sarcopenia in chronic obstructive pulmonary disease. J. Cachexia Sarcopenia Muscle 2023, 14, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Zhang, F.; Liu, L.; Shen, H.; Liu, T.; Jin, J.; Yu, N.; Wan, Z.; Wang, H.; Hu, X.; et al. LncRNA ANRIL promotes HR repair through regulating PARP1 expression by sponging miR-7-5p in lung cancer. BMC Cancer 2023, 23, 130. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Niu, H.; Wang, Y.; Li, R.; Zhao, Y.; Liu, C.; Cao, H.; Chen, H.; Xie, R.; Zhuang, L. LncRNA TUG1 contributes to the tumorigenesis of lung adenocarcinoma by regulating miR-138-5p-HIF1A axis. Int. J. Immunopathol. Pharmacol. 2021, 35, 20587384211048265. [Google Scholar] [CrossRef] [PubMed]
- Qi, F.; Lv, Z.D.; Huang, W.D.; Wei, S.C.; Liu, X.M.; Song, W.D. LncRNA TUG1 promotes pulmonary fibrosis progression via up-regulating CDC27 and activating PI3K/Akt/mTOR pathway. Epigenetics 2023, 18, 2195305. [Google Scholar] [CrossRef] [PubMed]
- Entezari, M.; Ghanbarirad, M.; Taheriazam, A.; Sadrkhanloo, M.; Zabolian, A.; Goharrizi, M.A.S.B.; Hushmandi, K.; Aref, A.R.; Ashrafizadeh, M.; Zarrabi, A.; et al. Long non-coding RNAs and exosomal lncRNAs: Potential functions in lung cancer progression, drug resistance and tumor microenvironment remodeling. Biomed. Pharmacother. 2022, 150, 112963. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Hashimoto, R.F. Dynamical Analysis of a Boolean Network Model of the Oncogene Role of lncRNA ANRIL and lncRNA UFC1 in Non-Small Cell Lung Cancer. Biomolecules 2022, 12, 420. [Google Scholar] [CrossRef]
- Fisher, L. Retraction: Long noncoding RNA ANRIL knockdown increases sensitivity of non-small cell lung cancer to cisplatin by regulating the miR-656-3p/SOX4 axis. RSC Adv. 2021, 11, 6255. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.H.; Kim, D.; Jin, E.J. Down-regulation of Phospholipase D Stimulates Death of Lung Cancer Cells Involving Up-regulation of the Long ncRNA ANRIL. Anticancer Res. 2015, 35, 2795–2803. Available online: https://pubmed.ncbi.nlm.nih.gov/25964559/. [PubMed]
- Zhang, H.; Li, X.; Jia, M.; Ji, J.; Wu, Z.; Chen, X.; Yu, D.; Zheng, Y.; Zhao, Y. Roles of H19/miR-29a-3p/COL1A1 axis in COE-induced lung cancer. Environ. Pollut. 2022, 313, 120194. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.T.; Xing, W.; Zhao, R.S.; Tan, Y.; Wu, X.F.; Ao, L.Q.; Li, Z.; Yao, M.W.; Yuan, M.; Guo, W.; et al. HDAC2 inhibits EMT-mediated cancer metastasis by downregulating the long noncoding RNA H19 in colorectal cancer. J. Exp. Clin. Cancer Res. 2020, 39, 270. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhao, M. High expression of long non-coding RNA MALAT1 correlates with raised acute respiratory distress syndrome risk, disease severity, and increased mortality in sepstic patients. Int. J. Clin. Exp. Pathol. 2019, 12, 1877–1887. Available online: https://wwwncbinlmnihgov/pmc/articles/PMC6947113/. [PubMed]
- Bhat, A.A.; Afzal, O.; Afzal, M.; Gupta, G.; Thapa, R.; Ali, H.; Hassan Almalki, W.; Kazmi, I.; Alzarea, S.I.; Saleem, S.; et al. MALAT1: A key regulator in lung cancer pathogenesis and therapeutic targeting. Pathol. Res. Pract. 2024, 253, 154991. [Google Scholar] [CrossRef] [PubMed]
- Qiu, N.; Xu, X.; He, Y. LncRNA TUG1 alleviates sepsis-induced acute lung injury by targeting miR-34b-5p/GAB1. BMC Pulm. Med. 2020, 20, 49. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Shen, J.; Chan, M.T.; Wu, W.K. TUG1: A Pivotal Oncogenic Long Non-Coding RNA of Human Cancers. Cell Prolif. 2016, 49, 471–475. [Google Scholar] [CrossRef]
- Loewen, G.; Jayawickramarajah, J.; Zhuo, Y.; Shan, B. Functions of lncRNA HOTAIR in lung cancer. J. Hematol. Oncol. 2014, 7, 90. [Google Scholar] [CrossRef]
- Ren, M.M.; Xu, S.; Wei, Y.B.; Yang, J.J.; Yang, Y.N.; Sun, S.S.; Li, Y.J.; Wang, P.Y.; Xie, S.Y. Roles of HOTAIR in lung cancer susceptibility and prognosis. Mol. Genet. Genomic Med. 2020, 8, e1299. [Google Scholar] [CrossRef]
- Tang, L.; Wang, S.; Wang, Y.; Li, K.; Li, Q. LncRNA-UCA1 regulates lung adenocarcinoma progression through competitive binding to miR-383. Cell Cycle 2023, 22, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Ling, L.; Li, C.; Wu, R.; Zhang, M.; Shao, F.; Wang, Y. LncRNA UCA1 promoted cisplatin resistance in lung adenocarcinoma with HO1 targets NRF2/HO1 pathway. J. Cancer Res. Clin. Oncol. 2023, 149, 1295–1311. [Google Scholar] [CrossRef] [PubMed]
- Szafranski, P.; Gambin, T.; Karolak, J.A.; Popek, E.; Stankiewicz, P. Lung-specific distant enhancer cis regulates expression of FOXF1 and lncRNA FENDRR. Hum. Mutat. 2021, 42, 694–698. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Merchan, A.; Cuadros, M.; Rodriguez, M.I.; Rodriguez, S.; Torres, R.; Estecio, M.; Coira, I.F.; Loidi, C.; Saiz, M.; Carmona-Saez, P.; Medina, P.P. The value of lncRNA FENDRR and FOXF1 as a prognostic factor for survival of lung adenocarcinoma. Oncotarget 2017, 11, 1172–1185. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Shirvani-Farsani, Z.; Hussen, B.M.; Taheri, M.; Samsami, M. The key roles of non-coding RNAs in the pathophysiology of hypertension. Eur. J. Pharmacol. 2022, 931, 175220. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Ma, Y.; Chen, Y. Extracellular vesicles and COPD: foe or friend? J. Nanobiotechnol. 2023, 21, 147. [Google Scholar] [CrossRef] [PubMed]
- Fanale, D.; Taverna, S.; Russo, A.; Bazan, V. Circular RNA in Exosomes. Adv. Exp. Med. Biol. 2018, 1087, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Sarfi, M.; Abbastabar, M.; Khalili, E. Long noncoding RNAs biomarker-based cancer assessment. J. Cell Physiol. 2019, 234, 16971–16986. [Google Scholar] [CrossRef] [PubMed]
- Reggiardo, R.E.; Maroli, S.V.; Kim, D.H. LncRNA Biomarkers of Inflammation and Cancer. Adv. Exp. Med. Biol. 2022, 1363, 121–145. [Google Scholar] [CrossRef]
- Chi, Y.; Wang, D.; Wang, J.; Yu, W.; Yang, J. Long Non-Coding RNA in the Pathogenesis of Cancers. Cells 2019, 8, 1015. [Google Scholar] [CrossRef]
- Chen, B.B.; Li, Z.H.; Gao, S. Circulating miR-146a/b Correlates with Inflammatory Cytokines in COPD and Could Predict the Risk of Acute Exacerbation COPD. Medicine 2018, 97, e9820. [Google Scholar] [CrossRef] [PubMed]
- Hoult, G.; Gillespie, D.; Wilkinson, T.M.A.; Thomas, M.; Francis, N.A. Biomarkers to Guide the Use of Antibiotics for Acute Exacerbations of COPD (AECOPD): A Systematic Review and Meta-Analysis. BMC Pulm. Med. 2022, 22, 194. [Google Scholar] [CrossRef] [PubMed]
- Walcher, L.; Kistenmacher, A.K.; Suo, H.; Kitte, R.; Dluczek, S.; Strauß, A.; Blaudszun, A.R.; Yevsa, T.; Fricke, S.; Kossatz-Boehlert, U. Cancer Stem Cells-Origins and Biomarkers: Perspectives for Targeted Personalized Therapies. Front. Immunol. 2020, 11, 1280. [Google Scholar] [CrossRef] [PubMed]
- Vogelmeier, C.F.; Román-Rodríguez, M.; Singh, D.; Han, M.K.; Rodríguez-Roisin, R.; Ferguson, G.T. Goals of COPD Treatment: Focus on Symptoms and Exacerbations. Respir. Med. 2020, 166, 105938. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Inflammatory Endotypes in COPD. Allergy 2019, 74, 1249–1256. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Liang, H.; Cui, R.; Ji, J.; Liu, H.; Liu, X.; Shen, P.; Wang, H.; Wang, X.; Song, Z.; Jiang, Y. Construction of a Risk Model and Prediction of Prognosis and Immunotherapy Based on Cuproptosis-Related LncRNAs in the Urinary System Pan-Cancer. Eur. J. Med. Res. 2023, 28, 198. [Google Scholar] [CrossRef] [PubMed]
- Brisotto, G.; Guerrieri, R.; Colizzi, F.; Steffan, A.; Montico, B.; Fratta, E. Long Noncoding RNAs as Innovative Urinary Diagnostic Biomarkers. Methods Mol. Biol. 2021, 2292, 73–94. [Google Scholar] [CrossRef] [PubMed]
- Terracciano, D.; Ferro, M.; Terreri, S.; Lucarelli, G.; D'Elia, C.; Musi, G.; de Cobelli, O.; Mirone, V.; Cimmino, A. Urinary Long Noncoding RNAs in Nonmuscle-Invasive Bladder Cancer: New Architects in Cancer Prognostic Biomarkers. Transl. Res. 2017, 184, 108–117. [Google Scholar] [CrossRef]
- Wang, Y.; Fu, X.; Yu, B.; Ai, F. Long Non-Coding RNA THRIL Predicts Increased Acute Respiratory Distress Syndrome Risk and Positively Correlates with Disease Severity, Inflammation, and Mortality in Sepsis Patients. J. Clin. Lab. Anal. 2019, 33, e22882. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Xie, D.; Pan, L.; Lou, Y.; Shi, M. Urinary Exosomal Long Noncoding RNAs Serve as Biomarkers for Early Detection of Non-Small Cell Lung Cancer. Biosci. Rep. 2021, 41, BSR20210908. [Google Scholar] [CrossRef]
- Zhang, Y.; Luo, G.; Zhang, Y.; Zhang, M.; Zhou, J.; Gao, W.; Xuan, X.; Yang, X.; Yang, D.; Tian, Z.; Ni, B.; Tang, J. Critical Effects of Long Non-Coding RNA on Fibrosis Diseases. Exp. Mol. Med. 2018, 50, e428. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Feng, Y.; Zheng, X.; Xu, X. The Diagnostic and Therapeutic Role of snoRNA and LincRNA in Bladder Cancer. Cancers 2023, 15, 1007. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, S.Q.; Wang, L.; Lin, H.; Zhu, J.B.; Chen, R.; Li, L.F.; Cheng, Y.D.; Duan, C.J.; Zhang, C.F. m6A Methyltransferase METTL3-Induced LncRNA SNHG17 Promotes Lung Adenocarcinoma Gefitinib Resistance by Epigenetically Repressing LATS2 Expression. Cell Death Dis. 2022, 13, 657. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, L.; Liu, J.; Wan, Z.; Zhou, L.; Liao, H.; Wan, R. LncRNA ITGB2-AS1 Promotes Cisplatin Resistance of Non-Small Cell Lung Cancer by Inhibiting Ferroptosis via Activating the FOSL2/NAMPT Axis. Cancer Biol. Ther. 2023, 24, 2223377. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Hu, T.; Tao, W.; Tong, J.; Han, Z.; Herndler-Brandstetter, D.; Wei, Z.; Liu, R.; Zhou, T.; Liu, Q.; Xu, X.; Zhang, K.; Zhou, R.; Cho, J.H.; Li, H.B.; Huang, H.; Flavell, R.A.; Zhu, S. A LncRNA from an Inflammatory Bowel Disease Risk Locus Maintains Intestinal Host-Commensal Homeostasis. Cell Res. 2023, 33, 372–388. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.N.; Li, J.; Tang, L.B.; Chen, W.T.; Zhang, L.; Xiong, L.X. MiRNAs and LncRNAs: Dual Roles in TGF-β Signaling-Regulated Metastasis in Lung Cancer. Int. J. Mol. Sci. 2020, 21, 1193. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Wang, B.; Tan, F.; Wu, Y.; Chen, J.; Zhao, F.; Liu, M.; Zhou, G.; Yuan, C. The Regulatory Role of LncRNA HCG18 in Various Cancers. J. Mol. Med. (Berl.) 2023, 101, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Hansel, N.N.; McCormack, M.C.; Kim, V. The Effects of Air Pollution and Temperature on COPD. COPD 2016, 13, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Egranov, S.D.; Lin, C.; Yang, L. Long Noncoding RNA Loss in Immune Suppression in Cancer. Pharmacol. Ther. 2020, 213, 107591. [Google Scholar] [CrossRef]
- Chen, C.; Ding, P.; Yan, W.; Wang, Z.; Lan, Y.; Yan, X.; Li, T.; Han, J. Pharmacological Roles of LncRNAs in Diabetic Retinopathy with a Focus on Oxidative Stress and Inflammation. Biochem. Pharmacol. 2023, 214, 115643. [Google Scholar] [CrossRef]
- Zeng, L.; Zhou, M.; Wang, X.; Long, X.; Ye, M.; Yuan, Y.; Tan, W. Aberrant LncRNA Expression in Patients with Proliferative Diabetic Retinopathy: Preliminary Results from a Single-Center Observational Study. BMC Ophthalmol. 2023, 23, 94. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, H.M.F.; Abdelghany, A.A.; Al Ageeli, E.; Kattan, S.W.; Hassan, R.; Toraih, E.A.; Fawzy, M.S.; Mokhtar, N. Long Non-Coding RNAs Gene Variants as Molecular Markers for Diabetic Retinopathy Risk and Response to Anti-VEGF Therapy. Pharmgenomics Pers. Med. 2021, 14, 997–1014. [Google Scholar] [CrossRef] [PubMed]
- Foronjy, R. Commentary on: The Potency of LncRNA MALAT1/miR-155 in Altering Asthmatic Th1/Th2 Balance by Modulation of CTLA4. Biosci. Rep. 2020, 40, BSR20190768. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Baskoro, H.; Rennard, S.I.; Seyama, K.; Takahashi, K. MicroRNAs as Therapeutic Targets in Lung Disease: Prospects and Challenges. Chronic Obstr. Pulm. Dis. 2015, 3, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Castaldi, P.J.; Guo, F.; Qiao, D.; Du, F.; Naing, Z.Z.C.; Li, Y.; Pham, B.; Mikkelsen, T.S.; Cho, M.H.; Silverman, E.K.; Zhou, X. Identification of Functional Variants in the FAM13A Chronic Obstructive Pulmonary Disease Genome-Wide Association Study Locus by Massively Parallel Reporter Assays. Am. J. Respir. Crit. Care Med. 2019, 199, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Vij, N. Nano-based theranostics for chronic obstructive lung diseases: challenges and therapeutic potential. Expert Opin. Drug Deliv. 2011, 8, 1105–1109. [Google Scholar] [CrossRef]
- Pleasants, R.A.; Hess, D.R. Aerosol Delivery Devices for Obstructive Lung Diseases. Respir. Care 2018, 63, 708–733. [Google Scholar] [CrossRef]
Figure 1.
The Causative Factors of COPD. Foremost among these is cigarette smoking, a primary contributor to COPD pathology, compounded by environmental exposures and the deleterious effects of air pollution. Genetic predispositions, respiratory infections, and the inexorable process of aging further exacerbate susceptibility. Additionally, socioeconomic disparities and gender differentials intricately intersect, shaping the complex landscape of COPD development. Moreover, the presence of risk factors can activate immune cells, prompting them to initiate airway inflammation, instigate remodeling processes, and inflict lasting lung damage, amplifying the intricate pathogenesis of COPD.
Figure 1.
The Causative Factors of COPD. Foremost among these is cigarette smoking, a primary contributor to COPD pathology, compounded by environmental exposures and the deleterious effects of air pollution. Genetic predispositions, respiratory infections, and the inexorable process of aging further exacerbate susceptibility. Additionally, socioeconomic disparities and gender differentials intricately intersect, shaping the complex landscape of COPD development. Moreover, the presence of risk factors can activate immune cells, prompting them to initiate airway inflammation, instigate remodeling processes, and inflict lasting lung damage, amplifying the intricate pathogenesis of COPD.
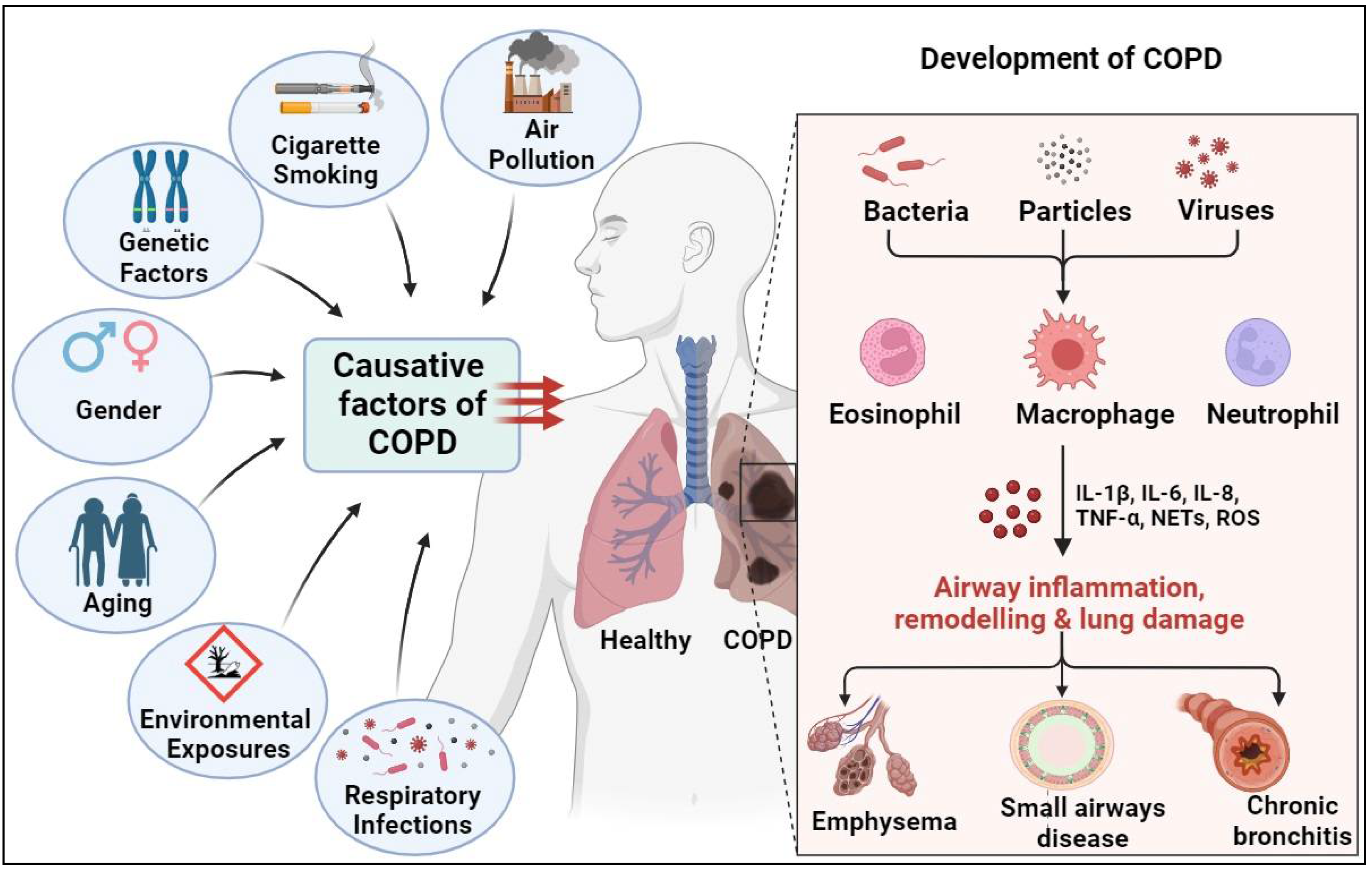
Figure 4.
Overview Highlighting the Role of Tobacco Smoke in Modulating LncRNAs During COPD Pathogenesis. Exposure to tobacco smoke induces significant alterations in the expression and functions of lncRNAs, precipitating a cascade of augmented COPD pathologies characterized by mitochondrial dysfunction, chronic inflammation, mucus dysregulation, epigenetic modifications, and cellular senescence. Noteworthy lncRNAs implicated in this multifaceted process include ANRIL, HOTAIR, LIPCAR, M-AS1, MALAT1, MANTIS, MEG3, MUC5B-AS1, PINT, SAL-RNA, SNHG-16, Sox2ot, and XIST, each intricately woven into the intricate tapestry of COPD-associated molecular dysregulation.
Figure 4.
Overview Highlighting the Role of Tobacco Smoke in Modulating LncRNAs During COPD Pathogenesis. Exposure to tobacco smoke induces significant alterations in the expression and functions of lncRNAs, precipitating a cascade of augmented COPD pathologies characterized by mitochondrial dysfunction, chronic inflammation, mucus dysregulation, epigenetic modifications, and cellular senescence. Noteworthy lncRNAs implicated in this multifaceted process include ANRIL, HOTAIR, LIPCAR, M-AS1, MALAT1, MANTIS, MEG3, MUC5B-AS1, PINT, SAL-RNA, SNHG-16, Sox2ot, and XIST, each intricately woven into the intricate tapestry of COPD-associated molecular dysregulation.
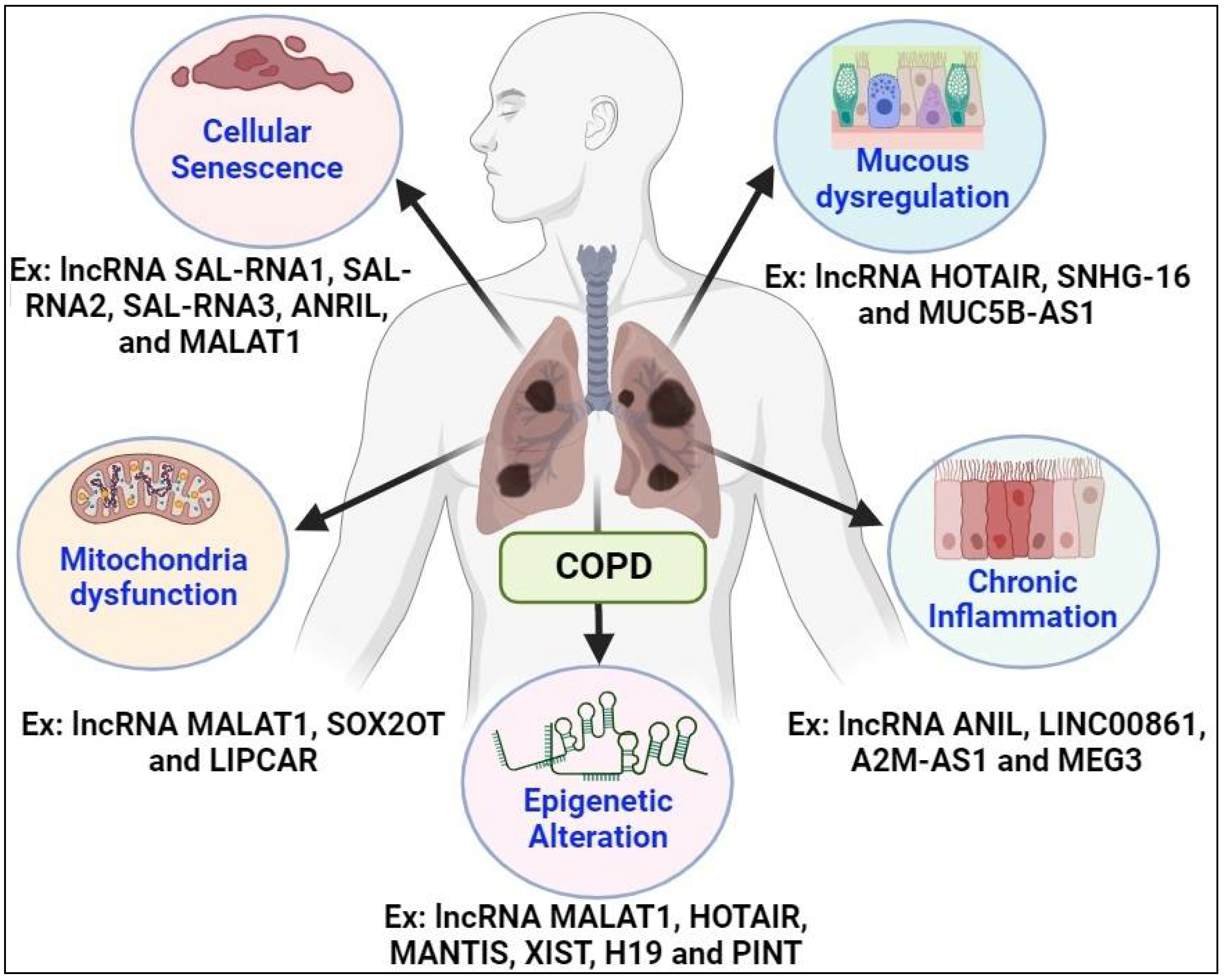
Figure 5.
The Multifaceted Landscape of Inflammation in COPD: A. Symphony of mediators and cells A. Inflammatory Immune Response in COPD. The fiery cascade of inflammatory mechanisms in COPD ignites with the activation of macrophages by the major risk factor of COPD (cigarette smoke and other irritants), unleashing a torrent of neutrophil chemotactic factors like IL-8 and LTB4. These chemical messengers spur the release of proteases, which ruthlessly dismantle connective tissue in the lung parenchyma, paving the path to emphysema while fueling mucus hypersecretion. Against this onslaught, protease inhibitors like α-1-antitrypsin, SLPI, and TIMP stand as frail barriers. Yet, cytotoxic T cells (CD8+) lurk, potentially exacerbating alveolar wall destruction, while fibroblasts, fueled by growth factors from macrophages and epithelial cells, stand poised for activation, amplifying the relentless assault on lung integrity. B. Inflammation complexity in COPD. The intricate tapestry of inflammation in COPD unfurls with myriad activated inflammatory and structural cells, orchestrating a symphony of mediators. Among them, lipid mediators like LTB4 beckon neutrophils, while chemokines such as MCP-1 and MIP-1α allure monocytes. IL-8 and GRO-α extend their call to both neutrophils and monocytes, while IP-10 beckons CD8+ cells alongside ROS and NO. GM-CSF prolongs the survival of neutrophils, while TNF-α ignites inflammation by activating multiple inflammatory genes and may underscore systemic manifestations of the disease. Meanwhile, endothelin and TGF-β set the stage for fibrosis. Concurrently, a host of proteinases including neutrophil elastase, proteinase C, cathepsins, and MMPs orchestrate elastolysis, ushering in the hallmark pathophysiology of COPD characterized by inflammatory cell activation, elastolysis, and mucus hypersecretion.
Figure 5.
The Multifaceted Landscape of Inflammation in COPD: A. Symphony of mediators and cells A. Inflammatory Immune Response in COPD. The fiery cascade of inflammatory mechanisms in COPD ignites with the activation of macrophages by the major risk factor of COPD (cigarette smoke and other irritants), unleashing a torrent of neutrophil chemotactic factors like IL-8 and LTB4. These chemical messengers spur the release of proteases, which ruthlessly dismantle connective tissue in the lung parenchyma, paving the path to emphysema while fueling mucus hypersecretion. Against this onslaught, protease inhibitors like α-1-antitrypsin, SLPI, and TIMP stand as frail barriers. Yet, cytotoxic T cells (CD8+) lurk, potentially exacerbating alveolar wall destruction, while fibroblasts, fueled by growth factors from macrophages and epithelial cells, stand poised for activation, amplifying the relentless assault on lung integrity. B. Inflammation complexity in COPD. The intricate tapestry of inflammation in COPD unfurls with myriad activated inflammatory and structural cells, orchestrating a symphony of mediators. Among them, lipid mediators like LTB4 beckon neutrophils, while chemokines such as MCP-1 and MIP-1α allure monocytes. IL-8 and GRO-α extend their call to both neutrophils and monocytes, while IP-10 beckons CD8+ cells alongside ROS and NO. GM-CSF prolongs the survival of neutrophils, while TNF-α ignites inflammation by activating multiple inflammatory genes and may underscore systemic manifestations of the disease. Meanwhile, endothelin and TGF-β set the stage for fibrosis. Concurrently, a host of proteinases including neutrophil elastase, proteinase C, cathepsins, and MMPs orchestrate elastolysis, ushering in the hallmark pathophysiology of COPD characterized by inflammatory cell activation, elastolysis, and mucus hypersecretion.
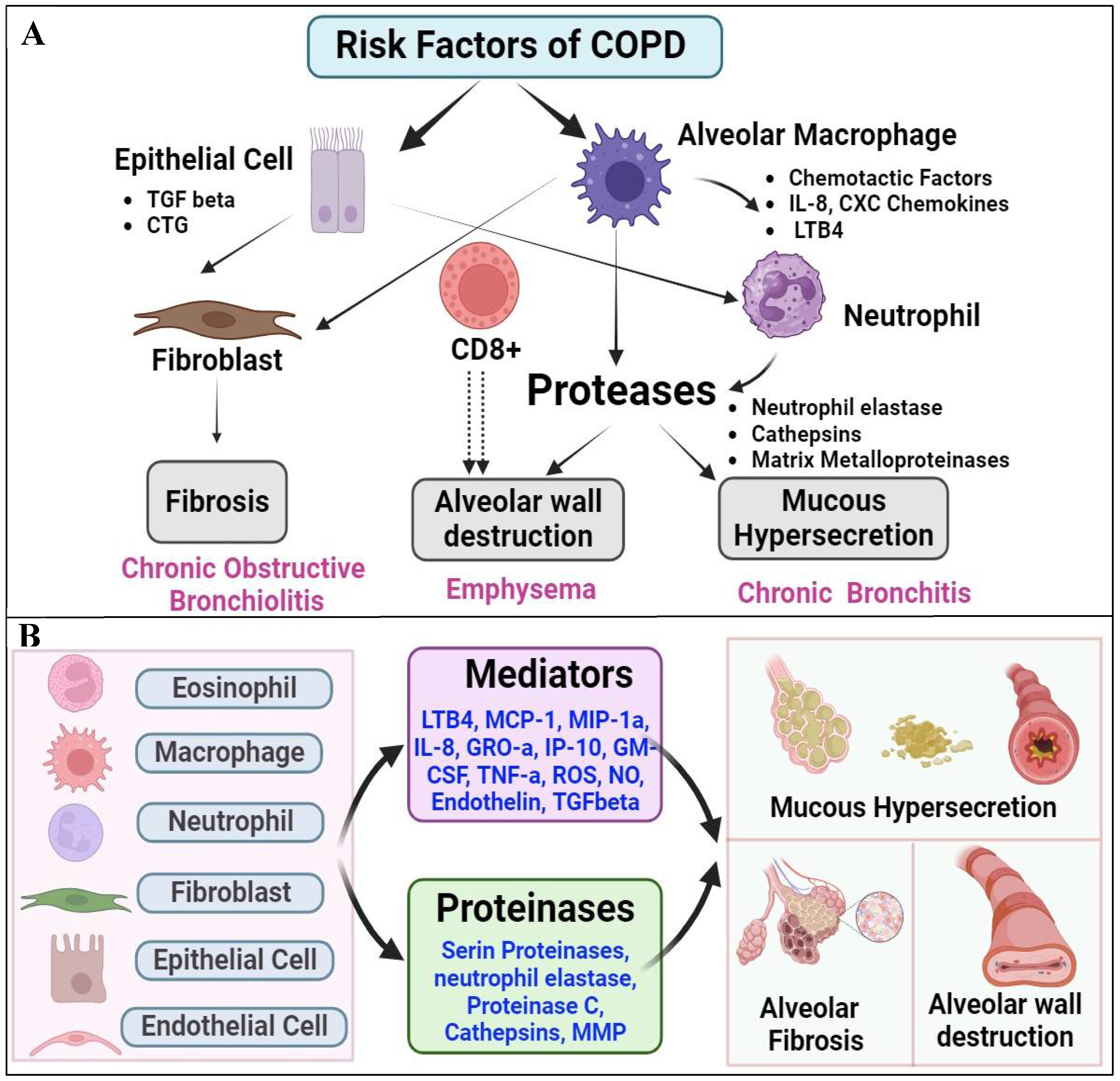
Figure 6.
Representative Diagram Illustrating Smoke Signals: LncRNAs Driving COPD Pathophysiology. Tobacco smoke exposure profoundly reshapes the landscape of lncRNAs, instigating a cascade of events leading to exacerbated COPD pathologies. This alteration encompasses mitochondrial dysfunction, chronic inflammation, mucus dysregulation, epigenetic modifications, and cellular senescence.
Figure 6.
Representative Diagram Illustrating Smoke Signals: LncRNAs Driving COPD Pathophysiology. Tobacco smoke exposure profoundly reshapes the landscape of lncRNAs, instigating a cascade of events leading to exacerbated COPD pathologies. This alteration encompasses mitochondrial dysfunction, chronic inflammation, mucus dysregulation, epigenetic modifications, and cellular senescence.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



