Submitted:
22 June 2024
Posted:
24 June 2024
You are already at the latest version
Abstract
Abstract: Palynomorph assemblages and mineral microfossils from five southern Red Sea coastal environments were studied to demonstrate their distribution pattern and suggest their source. The environments studied were 1. mangrove swamp 2. middle Holocene paleochannel 3. algal mat, 4. Sabia Island coral reef, and intertidal zone. The samples studied were from the top 3 cm semi-consolidated sediments from various environments. Standard palynological maceration technique was used to isolate palynomorphs. These sediment samples are mainly clay and fine sand and yielded low numbers but high diversity of palynomorph assemblages. They are grouped into pollen and spores, dinoflagellate cysts, and algal remains, fungal spores, hyphae and fruit bodies, protists, and invertebrates. A few Ascidian and sponge spicules, and phytoliths (mineral microfossils) occur in the Sabia Island coral reef environment. The protists and invertebrate remain include microforaminifera, thecamoebians, tintinnomorphs, crustacean and annelid palynomorphs. The assemblage also includes six form taxa belonging to the form genera Katora and Mangrovia. These palynomorphs belong to both marine and terrestrial environments and are of autochthonous and allochthonous origins. The source and genetic relationship of each palynomorphs is discussed.
Keywords:
Red Sea coastal environments
; non-pollen palynomorphs (NPP)
; Palynology
; Foraminiferal palynomorphs
; Tintinnomorphs
; Rotifers
; Ascidian and sponge spicules
; Phytoliths
1. Introduction
Since non-pollen palynomorphs (NPP) are sourced from a wider range of environments, they have become an important part of palynological research. Primarily NPP are microscopic remains belonging to a variety of plants, algae, fungi, protists, and invertebrates or to a lesser extent the whole organisms like dinoflagellate cysts. During the past few decades their identifications and possible sources have been studied. Initially palynological research mainly concentrated on pollen, spores, and dinoflagellate cysts, and generally NPP was overlooked. Although NPP studies were initiated from freshwater sediments in Europe [1,2,3,4]. However, other important NPP studies are from the Black Sea corridor [5], the Caspian-Black Sea-Mediterranean corridor [6], and Holocene sediments of offshore Adélie Land, East Antarctica [7]. An extensive discussion on a variety of marine NPP was recently published [8,9].
In the Arabian Peninsula region, there are very few studies reporting on NPP. The earliest publications from this region were recent dinoflagellate cysts from the Persian Gulf, Gulf of Oman, and northwestern Arabian Sea [10,11]. Palynological report on two offshore Oman samples [12] and palynological studies from Egypt [13,14] were other publications reporting presence of NPP from this region. Kholief [15] reported on palynology of mangrove sediments from the northern Red Sea coastal environments that reported few NPP. The present author published on palynology of the southern Red Sea coastal environments [16,17] reporting on in addition to pollen and spores, a wide range of NPP that included 1. dinoflagellate cysts and other algal remains, 2. fungal spores, hyphae, and fruit bodies, 3. microforaminiferal test linings, 4. thecamoebians, 5. tintinnomorphs and worms, and 6. crustacean and annelid palynomorphs. Additionally mineral microfossils like Ascidian and sponge spicules, and phytoliths were reported as well. Subsequently, new form taxa of NPP were proposed [18], followed by a morphometric study on foraminiferal palynomorphs [19], and a detailed morphological and taxonomic study of fungal remains were also published [20]. The present study compiles all the previously published palynomorph assemblages from five coastal environments, describes their proportional composition in various environments, discusses their sources, and environmental and palaeoecological implications. To avoid duplication of microphotographs of all the palynomorphs reported in this study, no photo plates are added to this manuscript. They have been illustrated in several photo plates in the previous publications [16,17,18,19,20].
Another objective of this study is to provide comprehensive palynological baseline data about the southern Red Sea coastal environments. Since the discovery of oil in the Arabian region during the 1920s the economic explosion continues to this day [21,22]. Extensive infrastructure projects, both on and offshore, are progressing on the eastern and western coastal regions of the Arabian Peninsula. It is resulting in adverse environmental consequences to the coastal ecosystem where mangroves and offshore sea-grass communities are being negatively impacted [23,24]. Baseline data about the coastal environments and their flora and fauna is required to define the present health of the coastal ecosystem. Study of palynomorphs occurring as fossils or subfossils in various coastal sedimentary settings represent local and regional climate, and environments. The present area of study has not yet been significantly influenced by any anthropogenic activity, except for a road and a few small buildings. Thus, the present study becomes significant as it provides baseline data for undisturbed parts of southern coastal Red Sea environments.
This study presents a palynological analysis of sediments of various coastal environments of the Wadi Hali area of the southern Red Sea coast of Saudi Arabia (Figure 1 and Figure 2), and discusses vegetation types, geological and environmental factors that control the distribution of palynomorphs in various coastal environments of Wadi Hali. The study describes environmental parameters controlling palynomorph distribution is also discussed. Finally, this research presents a palynological database that could be utilized for future environmental monitoring in this region.
2. Study Area and Its Environment
The Red Sea is a long and narrow body of water, its surface water temperatures are constant year-round at 210-250C [25]. Its salinity ranges between ~ 36‰ (3.6% on the Practical Salinity Scale or PSU, meaning actual dissolved salts) in the southern region and ~ 41‰ (4.0% PSU) in the northern region. The world average is ~ 35‰ or 3.5% PSU [26]. Mangroves, seagrasses, coastal vegetation and associated flora, and terrestrial mammals are common around the coastal zones. Latitudinal trends indicate increased abundance of mangroves, seagrasses and coastal flora towards the southern Red Sea, and a decrease in the number of coral reefs [27]. The littoral area of Red Sea sediments is related to a complex system of fringing and barrier reefs and alluvial fans. They belong to the arid regions covered by Quaternary terraced alluvial deposits, eolian sands, river-bed alluvium and sabkhas (salt pans). Mangrove swamps and wadis (dry riverbeds) are scattered all along the coast. Environmental changes along the Red Sea coast are influenced by anthropogenic activities, such as tourism, new towns, and port development. The natural causes for coastal changes are either wadi run-off or variations in longshore drift and erosion events [21].
The present study covers the Wadi Hali (also spelled Haliy) fan that lies on the Red Sea coast of Saudi Arabia, south of Jeddah and north of Jizan (Figure 1). This is a small region covering an area of around 50 km N-S and 40 km E-W. This area has several distinct environments like aeolian, fluvial, coastal, intertidal, brackish-water ponds with mangrove stands and saline lagoons. Estuarine clastic sediments have been deposited in this region since Quaternary [28]. Wadi Hali now is an ephemeral stream; however, it must have been active since at least Quaternary [28]. This area was semi-arid, having intense rainfall during the humid phase (Late Pliocene to Early Pleistocene and Late Pleistocene – Early Holocene). It is likely that in the past Wadi Hali was a permanent stream and during the intervening arid phase it became an ephemeral stream during the past 4 kyr phase of the present aridity. Massive flooding events are common in the SW regions of the Arabian Peninsula due to periodic intense cloud bursts. This wadi has been eroding the Arabian Shield and transporting sediments to the Red Sea. A brief description of coastal flora and vegetation of the southern Red Sea coast was provided by the author [16].
The study area covers Wadi Hali and its surroundings. Wadi Hali is a short ephemeral stream that originates in the hilly regions east of the coastal region (Figure 2). The area covers marginal marine to marine environments; that includes intertidal environments (Figure 3), scattered patches of mangrove stands and their muddy environments (Figure 4A,B,C), algal (microbial) mats and mud flats (Figure 4D,E), Sabia Island coral reef environment (Figure 5A), and A mid Holocene paleochannel section is also included in this study because it contains palynomorphs belonging to the extant plants and animals (Figure 5B).
Such studies are significant in understanding the ecology of various life forms that inhabit such environments. This type of data set has palaeoecological significance as well since it may also be used in the study of fossil sediments representing similar environments. The present paper describes palynomorph assemblages from mangrove swamps, a middle Holocene paleochannel deposit, algal mat and Sabia Island coral reef and discusses their sources and environmental significance. This study also provides comprehensive palynological baseline data about the southern Red Sea coastal environments. Baseline data about the coastal environments and their flora and fauna is required to define the present health of the coastal ecosystem. Study of palynomorphs occurring as fossils or subfossils in various coastal sedimentary settings represent local and regional climate, and environments.
3. Materials and Methods
This study is based on 25 samples collected on March 1 and 2, 2011, from five different localities representing five different coastal environments around the Wadi Hali area (180 49′35.27″N: 410 22′44.23″E) and its surroundings in the southern Red Sea coast of Saudi Arabia (Figure 3, Figure 4 and Figure 5). The samples and their locations are as follows.
Locality 1: Two surface (top one cm) mud samples (M1 and M2).
Locality 2A: Two surface (top one cm) mud samples (M3 and M4).
Locality 2B: Three surface (top one cm) mud samples (M5, M6 and M7).
This is a small outcrop at the base of one of the channels of Wadi Hali; it is located about 2 km inland from the coastline.
Locality 3: Paleochannel outcrop is 1.5 m thick. Seven clay to fine sand samples (WH1 through WH7) were collected from bottom to top at 20 to 25 cm interval, except the samples WH3 and WH4 which are about 40 cm apart. Samples represent lithological variations in the outcrop.
Locality 5: Two surface (top one cm) mud samples (AM1 and AM2) from the algal mats of the hyper-saline pools on the tidal flats.
Locality 4: Two surface calcareous sand samples (SI1 and SI2) from the coral reefs around the island. These are grab samples collected by diving in the shallow water.
a. Close to the shore in one m water depth (SI1)
b. Further offshore in two m water depth (SI2)
Location 6: Seven surface samples (L1-L7). Four clay to fine sand samples (L1 through L4) from tidal flats close to the shoreline and two clay samples (L6 and L7) from a small pond with abundant algae. The sample L5 is subtidal mud collected 100 m away from the coastline under 1.5 m water depth.
Sediment samples were macerated following the techniques described in Kumar [16]. One cubic centimeter (cm3) of dry sample was used in the maceration process using the following method. (1) 10 % HCl treatment for 15 minutes to remove carbonates. (2) Overnight cold HF (40 %) treatment to remove silicate minerals. (3) Commercial grade HNO3 (sp. gr. 1.35) treatment for two hours at 800 C for oxidation of organic matter to remove unwanted organic debris. (4) 10% KOH treatment for 15 minutes followed by rotation at 1,000 rpm for 5 minutes to remove humic matter. (5) Treatment of solution with 5 % HCl to neutralize pH. (6) Residue sieved through 10 μm mesh size sieve to remove clay minerals. (7) Residue with fraction larger than 10 µm fraction mixed with 3 drops of polyvinyl alcohol, mixed thoroughly, and smeared over coverslip and dried at 600 C over hot plate. (8) Mounted with Canada Balsam and kept for 20 minutes over hot plate at 800 C. (9) Four slides of each sample were made. There is a possibility of fungal contamination after the samples were collected and sealed in a sample bag since fungus can grow and may contaminate them.
The slides were studied under an OMAX Optical Microscope (MD827S30L Series) using transmitted light. Each slide was scanned under ×400 magnification and palynomorphs were photographed at ×400 and ×1,000 (oil immersion) using an inbuilt camera system in the microscope. A few large palynomorphs were photographed at ×100 as well. Since palynomorph numbers are generally low (see tables), all palynomorphs in each slide were counted (except in few slides where numbers of one form exceeded 500).
4. Results
The palynomorphs recovered in this study are classified into the following groups: pollen and spores, dinoflagellate cysts and algal remains, fungal spores, hyphae and fruit bodies, protists, and invertebrates. Mineral microfossils (ascidian spicules, sponge spicules and phytoliths) are not palynomorphs because are not organic, however, since they occur in palynological preparations, they are included in the assemblage. Fragments of a variety of cuticles, tracheid, wood and charcoal occur commonly in almost all samples. Palynomorph preservation ranges from moderate to good. Several palynomorphs belonging to various groups could not be identified, thus labelled as indeterminate. The distribution of palynomorphs recorded from various environments is shown in Table 1, Table 2, Table 3, Table 4, Table 5 and Table 6 (Table 1; pollen and spores, Table 2; dinoflagellate cysts and other algal remains, Table 3; fungal taxa, Table 4. microforaminifera, thecamoebians and tintinnomorphs, and form taxa, Table 5. numerical distribution of palynomorphs and foraminiferal palynomorphs in various environments, Table 6. invertebrate remains and mineral microfossils. Table 7 summarizes distribution of palynomorphs and palynomorph groups in various environments. The relative abundance of palynomorphs are denoted as Rare (R): 1-4 specimens; Few (F): 5-10 specimens; Common (C ): 11-25 specimens; Very Common (VC): 26-50 specimens; Abundant (A): 51 or more specimens. The occurrences of various groups of palynomorphs and their possible source is discussed.
4.1. Pollen and Spores
Pollen and spores form a minor component of the palynomorph assemblages, only rare occurrences of individual pollen and spore were observed. A total of 25 taxa of pollen (21 angiosperms and 4 gymnosperms) and 6 taxa of spores were observed in this study (Table 1). The pollen and spores identified were Acacia sp. (Leguminosae), Artemesia sp. (Asteraceae), Avicennia marina (Avicenniaceae), cf. Cyperaceae pollen, Euphorbia sp. (Euphorbiaceae), Inaperturate pollen, Juniperus sp. (Cupressaceae), Nypa sp. (Arecaceae), Palmaepollenites sp. (Arecaceae), Palmidites sp. (Arecaceae), cf. Potamogeton sp. (Potamogetonaceae), Proxapertites sp. (Araceae/Arecaceae), Rannunculaceae pollen, Rhizophora mucronata (Rhizophoraceae), Trichotosulcites sp., and Zea mays (Graminae; cultivated).
There is no published pollen atlas of Saudi Arabian plants. However, there are few publications on the palynology of Quaternary and Recent sediments in and around the Arabian Peninsula describing both pollen and NPP. They describe and illustrate the morphology of common recent pollen and spores [15,30,31,32,33]. Few other publications also describe and illustrate pollen [12,13,34]. These papers helped in identifying certain pollen and spores. Most importantly floral composition of various vegetation types in Saudi Arabia [35] was used to discuss the provenance of pollen types identified. A 12-month long aerobiological survey of pollen in Riyadh, the capital city of Saudi Arabia located in the east-central region of the country, recorded several types of pollen taxa including Palmae/Arecaceae (36). In order from the most to least common taxa were Chenopodiaceae, grasses, plantain, Artemisia, and Ambrosia. Grasses and Chenopodiaceae released pollen 11 to 12 months of the year [36]. This study shows the presence of airborne pollen in the central part of Saudi Arabia and most of them are reported in the present study. However, Palmae (Arecaceae) pollen dominates the coastal assemblages because mostly they inhabit the coastal regions.
Among the angiosperms, the monocotyledonous family Arecaceae (syn. Palmae) is represented by the largest number of taxa; they are Neocouperipollis sp., Palmaepollenites sp., Palmidites sp. Spinizonocolpites sp. (Nypa palm), Trichotomosulcites sp. and Verrualetes sp. According to Thomas [35] four palm species occur in Saudi Arabia; they are Hyphaene thebaica, Phoenix caespitosa, Phoenix dactylifera, and Phoenix reclinata. The date palm (Phoenix dactylifera) is an important crop in Saudi Arabia. This family also contains commercial species such as coconuts, areca nuts and date palms, and many indoor and ornamental species as well. Presence of Arecaceae pollen taxa reported in this assemblage probably belong to these species of palms transported by wind (sand and dust storms) and water (flash floods and rains) from the neighboring regions and beyond since various species of palms occur widely in the Arabian region and northeast Africa. The genus Proxapertites sp. (Araceae/Arecaceae) too is sourced from one of the palm species found in the region.
Acaciapollenites sp., Euphorbia sp., unidentified tricolpate pollen and Chenopodiaceae/ Amaranthaceae pollen are other significant pollen taxa of the assemblage. Most of them were derived from the vegetation of the coastal plains, sabkhas and wadis characterized by low sand dunes and saline marshes. These environments are inhabited by open, drought-deciduous thorn woodlands, mangroves, halophytes, open xeromorphic and thorn woodlands. Avicennia marina and Rhizophora mucronata are mangrove pollen found in this assemblage.
Cyperaceae pollen and pollen of Artemisia sp. and Calligonum sp. are probably derived from open xeromorphic grasslands where common shrubs of the sand-dunes are Artemisia monosperma and Calligonum comosum. All these shrubs co-occur with perennial herbs and grasses such as Cyperus conglomeratus.
Presence of a single pollen of cf. Casuarina sp. is problematic because it is not native to this area, but Casuarina trees planted near Dammam along the Arabian Gulf coast of Saudi Arabia were observed (A. Kumar, personal observation). Most likely the Casuarina pollen may have been transported during thunderstorms or dust storms which are common in this region.
The presence of cf. Potomogeton pollen in this assemblage is problematic because it is a pond weed that occurs in submerged aquatic habitats. At present there are only rare ponds along the Tihama Plain in this desert environment where Potomogeton could grow. Although Saudi Arabia does not have rivers or perennial streams, its seasonal streams and ponds do contain a few species. The aquatic flora in Saudi Arabia contains more than forty aquatic or semi aquatic species, of which Potamogetonaceae has the maximum number of species [35]. Formation of ponds occurs during flooding events caused by thunderstorms in which heavy to very heavy rainfall occurs within a short period of time [37]. I suggest that a species of Potomogeton might have grown in one such pond contributing to the pollen assemblage.
Several plant species belonging to the family Ranunclaceae occur in Saudi Arabia as well. In the southwestern region of Saudi Arabia plant species belonging to this family are Clematis hirsute, Clematis simensis, Ranunculus multifidus and Ranunculus forskoehlii [35]. There is a possibility that a Rannunculaceae pollen reported in this study might be sourced from one of these plants.
Among the gymnospermous pollen Juniperus sp., Pinuspollenites sp. and Podocarpus sp. were observed. The Juniperus sp. pollen is related to Juniperus procera that inhabits higher altitudes in the southwestern parts of the Arabian Peninsula. Pinus and Podocarpus are not known in the Arabian region. However, Pinus and Podocarpus pollen were reported from Pleistocene and Holocene sediments of the Nile Cone, Egypt [14]. Pinus is a genus of the Pinaceae family; it has a characteristic bisaccate pollen. This family has worldwide distribution including North Africa. Pinus radiata was introduced for timber production in sub-Saharan Africa and Podocarpus is known from the tropical highlands of Africa. Tropical cyclones originating in the Arabian Sea are known to impact the Arabian Peninsula, mainly in the southern regions such as Yemen and Oman. Pollen of Pinus and Podocarpus are most likely wind transported from East Africa by these cyclones or the southwest monsoon, when the summer wind blows across the Arabian Sea from northeast Africa to India with a branch that veers north towards the Arabian Peninsula.
The monoporate pollen Zea mays (Graminae; cultivated) is known from the Arabian region and have been reported in a few studies from Iraq [32,34,38]. Monoporate pollen could also be derived from the family Poaceae/Gramineae which are common in the Asir Mountains, Jizan and in the wadis of southwestern regions of Saudi Arabia [35].
Six taxa of pteridophytic spores were observed belonging to the fern families Lycopodiaceae and Polypodiaceae. In Saudi Arabia there are twenty-seven species of Pteridophytes. They occur in most terrestrial habitats and in some aquatic communities. The observed spore taxa must be related to these plants and most likely transported by both wind and water to the site of deposition. Rare specimens of Hypnaceae spore (Moss) were observed in algal mat samples. The genus Hypnum has a cosmopolitan distribution occurring on all continents except Antarctica. They are typically found in moist forest areas, while some species are aquatic. They can also be found living on soil, rocks, and live trees.
4.2. Dinoflagellate Cysts and Algal Remains
4.2.1. Dinoflagellate Cysts
Four known and fourteen unknown dinoflagellate cyst taxa were observed in this study (Table 2). The known taxa are cf. Brigantedinium simplex, ?Operculodinium sp., Peridinium sp., and Spiny round brown cyst. The following six unknown taxa were informally described as dinoflagellate cyst types A, B, C, D, E and F [16], and further eight more were described as dinoflagellate cyst types G, H, I, J, K, L, M and N [17]. Rare specimens of acritarchs were observed as well, usually they are indeterminate, and a known form identified as Micrhystridium sp.
These morphotypes are commonly thin walled and lack complete dinoflagellate cyst like paratabulation characters except for their shape, thin and generally psilate autophragm with indistinct archaeopyle. Their comparison with any known dinoflagellate cyst is difficult because of their poor preservation and incomplete morphological features. Such morphotypes types were found only as one or two specimens and appear to have morphological affinities with fresh and brackish water dinoflagellate cysts. Their low abundance may be due to treatment with HNO3 and KOH which may have destroyed such thin-walled cysts.
Dinoflagellate cysts taxa are a minor part of the overall palynomorph assemblage. They appear to be an assortment of fresh or brackish water forms mixed with very few marine forms that are most likely derived from the coastal lagoons of the Red Sea. Fresh water dinoflagellates are characteristically thin walled and often the archaeopyle is not visible because it might be hidden by folds in the autophragm or due to poor preservation. Furthermore, paratabulation features are either not well developed or not identifiable [39]. Fresh to brackish water dinoflagellate cysts typically are smaller, have thin autophragm and have simple proximate shapes. They are often associated with tidal channel and tidal mudflat facies that provide evidence of a possible upper estuarine setting [40]. There are approximately 350 freshwater dinoflagellate species, and 84 resting cyst species. Their shapes, wall ornamentations, archeopyles and colors are important morphological characteristics for identification at the generic level [41].
The dinoflagellate cyst assemblage observed in this study is marine (cf. Brigantedinium simplex, ?Operculodinium sp.; Spiny Round Brown Cyst (SRBC) with theropylic archaeopyle but also has some freshwater forms especially Peridinium sp. Resting cysts attributed to the freshwater dinoflagellate genus Peridinium sp. have been described from the lakes of North America [42,43,44].
4.2.2. Algal Remains
There are no published accounts of any algal remains from the Red Sea coastal sediments. However, there are few such studies from Egypt [14,45]. As with pollen and spores and dinoflagellate cysts, algal remains are minor constituents of the palynomorph assemblages; only rare occurrences were observed in various samples (Table 2).
Long slender filaments, generally septate, branched, or unbranched are found commonly in most samples. Thin-walled spherical cysts with psilate or scabrate, intact or broken wall, usually ranging between 25-40 μm are common in few samples. They have been attributed to algal cell types A and B and algal cysts (unidentified). Botryococcus, Concentricystis and Spirogyra zygospore, Lecaniella (Spirogyra zygospore), Cyanobacteria Gleotrichia sp. and Rivularia are the other algal remains in the present assemblages. A total of 13 algal forms were recorded in this study (Table 2).
Botryococcus is a colonial green alga of the order Chlorococcales that tolerates variable salinities, thus inhabits brackish or freshwater environments including ponds and temporary pools and found in temperate to tropical oligotrophic lakes and estuaries [46]. Concentricystis is another algal spore found in freshwater habitats. These are zygospores of spheroid to obovoid shape, have hyaline, ornamented wall with concentric rings in polar view. It is indicative of freshwater marshy environments. It is also known as Pseudoschizaea [47].
Cyanobacteria or the blue-green algae normally occur in lakes and ponds. Gloeotrichia is a Cyanobacteria (blue-green algae) and their filaments [6] occur in the assemblage indicates fresh-water input in the brackish water condition [47,48], thus indicates brackish water, littoral environment. Rivularia is a modern cyanobacteria belonging to the family Rivulariaceae, order Nostocales, grows on plants and could also be free-floating and photosynthetic, and become abundant when there is an incursion of salt-water, thus, their abundance could be a proxy for brackish water, littoral environment [47,48].
Lecaniella is a Spirogyra zygospore. Spirogyra is a filamentous chlorophyte green algae of the order Zygnematales and is commonly found in freshwater habitats. According to Hoshaw and McCourt [49] “Zygospores (resting, encysted zygotes of some chlorophyte algae) are often produced during seasonal dessication of the habitat, and changes in water chemistry (nitrogen depletion and rise in pH) and light regime seem to be responsible for triggering zygospore formation.” Seasonal desiccation of habitat and changes in water chemistry in the Red Sea coastal ponds is an annual occurrence when Spirogyra zygospore might have formed. Additional fresh-water algal remains that occur in the present assemblages are Zygnema-type spore [5] and zygospore of Debraya glyptosperma [50]; they belong to a family of fresh-water algae Zygnemataceae. Oospores of Oedogonium sp., which is a fresh water green alga, also occurs in these assemblages [51]. Many broken pieces of marine diatoms were also observed in the Sabia Island coral reef samples.
Several spherical to subspherical cells of variable diameters having psilate, scabrate or granulate walls were observed in this study. All such forms are grouped as algal cells, their morphological characters were described earlier [17]. A typical algal cell is oval to ovoidal in shape, light grey to light brown, wall thick, unevenly granulate, grana size variable.
Thus, it is evident from the dinoflagellate cysts and algal remains that these assemblages contain a mixture of freshwater or brackish and marine forms. This interpretation is also confirmed by the presence of microforaminifera and thecamoebians in these assemblages.
4.3. Fungal Spores, Hyphae, and Fruit Bodies
Fungal remains are a significant part of the palynomorph assemblages in most samples. Several taxa of fungal spores, fruit bodies and hyphae were observed (Table 3). Fungal palynomorphs were tentatively identified, described, and illustrated in earlier study [16,17]. Later, a morphological and taxonomic restudy of these fungal palynomorphs was carried out and these morphotypes were placed in various genera and species [52]. The generic identifications are based on comprehensive treatises on fossil fungi by Kalgutkar and Jansonius [53], and Saxena and Tripathi [59]. Fungal remains were represented by 23 genera and 27 named species [52]. In addition, there are several specimens which could be identified only up to generic level and have been described as sp. 1, sp. 2, etc. The assemblage consists mainly of fungal spores whereas sporocarps are poorly represented by only three genera. The extant relationship of fungal sporocarps, spores, mycelia and other fungal remains is presented. Eight new species were proposed, they are: Dicellaesporites bisariae, Dicellaesporites foveolatus, Inapertisporites choudharyi, Inapertisporites kharei, Inapertisporites foveolatus, Inapertisporites microverrucatus, Monoporisporites rotundus and Pluricellaesporites mishrae (Table 3).
Several specimens of Glomus sp. were observed in many samples [16,17]. Glomus sp. (family Glomeraceae) is an endomycorrhizal fungus showing a symbiotic relationship with roots of higher plants. These are globose chlamydospores, aseptate, inaperturate and are of variable size (18-138 µm). They are formed below the soil surface and are not normally transported [54]. They are indicative of soil erosion in the catchment area and provide a useful line of evidence in palaeoecology [55].
Fungal fruit bodies (Microthyriaceae) are multicellular, epiphyllous forms; they occur most likely on the coastal mangrove stands and are indicative of tropical to subtropical moist humid climate and heavy rainfall. Fungal hyphae are unicellular to multicellular, branched, or unbranched forms that occur primarily in freshwater marsh/flood plain environments and are indicative of high organic input and heavy rainfall. Fungal spores vary from unicellular to multi-cellular, septate or aseptate, with or without pores and occur in the present assemblages.
Since fungi lack chlorophyll, thus are heterotrophs and survive as epiphytes, saprophytes, parasites or in symbiotic associations on or in living or dead plants and animals. They rarely produce hard resistant tissues thus not prone to fossilization, however their spores, hyphae and fruit bodies are preserved as organic matter and can be isolated using palynological maceration techniques [53]. A comprehensive account of the diverse aspects of Palaeomycology which includes study of fossil fungal palynomorphs their morphology, descriptive terminology, identification and nomenclature, geological history and paleoenvironmental and paleoclimatic significance, etc. is discussed [53,56]. Fungi provides evidence for relative humidity, atmospheric pressure, and precipitation rates. The assemblage of fungal spores in higher diversity and numbers are usually considered to be indicative of warm and humid climatic conditions [47,48].
Fossil fungal remains have been used as proxies for environmental perturbations and vegetation changes caused by climate change, as well as floristic changes, and the sequence of vegetational modifications during the past. Microclimatic and edaphic interactions of fossil fungi with geological soils have also been investigated by Kalgutkar and Jansonius [53]. A useful review of the relationship between fungi and their environment in the geological past is provided by Stubblefield and Taylor [57] and Taylor [58]. Saxena and Tripathi [59] suggested that it is advisable to take into consideration the complete palynological assemblage for paleoenvironmental interpretations rather than just based on fungal remains.
In these marginal marine environments, there is free mixing of fresh water. Many of the fungal palynomorphs were transported from land in these environments by wind and water. Conidia of Tetraploa are found generally associated with grasses indicating grass supporting environment nearby. Similarly, Alternaria spores indicate the presence of herbaceous flora. It is well known that to survive, Alternaria needs a moist warm environment. This fungus lives in seeds and seedlings and is spread by spores. Nigrospora (being represented by Exesisporites) grows in soil, air, and plants as a leaf pathogen. Microthyriaceous ascocarps, viz. Callimothallus, Spinosporonites and Trichothyrites are also epiphyllous fungi growing on leaves. In general, the assemblage indicates warm and moist conditions in the area as indicated by dry, hot, and brackish coastal environments that periodically become wet and humid due to short term excessive rainfall during tropical storms.
4.4. Protists and Invertebrates
Foraminifers, thecamoebians and tintinnids are three groups of Protists that commonly occur in these assemblages. Benthic foraminifers are quite ubiquitous in brackish/marine environments ranging from intertidal zone to deep oceans and from polar regions to the tropics. Thus microforaminiferal test linings are found in all these environments. Thecamoebians, also known as agglutinated rhizopods or testate amoebae are benthic microfaunal groups that occur worldwide in bodies of fresh and brackish water. Tintinnids are present as tintinnomorphs that are part of the microzooplankton inhabiting primarily marine, but brackish and freshwaters as well.
4.4.1. Microforaminiferal Test Linings
Microforaminiferal test linings are the inner organic remnants of the benthic microforaminifera, commonly observed in palynomorph assemblages of marine and brackish water sediments. They are acid-resistant, chitinoid linings, usually <150 μm size. Mathison and Chmura [60] developed a qualitative classification of the physical condition of microforaminiferal test linings based on whether individual chambers were intact or damaged (split or torn), and if they were closely spaced (proximate) or the spacing of the individual chamber was open. Based on these criteria, sixteen microforaminiferal lining types were described as lining types 1 through 16 [16,17]. None of the specimens have any torn or split chambers in the present assemblages. Their distribution is shown in Table 4 and Table 5. Microforaminiferal test linings generally occur as rare or few in most samples, however, occasionally they are common to very common in few samples (Table 4, Table 5 and Table 7).
Microforaminiferal linings have been used as proxies for water pollution, ocean productivity and paleoenvironmental studies [61]. Monga et al. [62] used various groups of microforaminiferal linings to delineate marine and terrestrial realms in the Paleogene sequences and facilitated better interpretation of the depositional environment. Mudie and Yanko-Holmbach [63] carried out concurrent palynological and micropaleontological studies to investigate the relationship between the morphology and abundance of these linings and benthic foraminiferal assemblages along the transects off the Danube River Delta, NW Black Sea. Al-Dubai et al. [64] published on the diversity and distribution of benthic foraminifera in the Al-Kharrar Lagoon, eastern Red Sea coast of Saudi Arabia. They defined five assemblages characterizing various sub-environments in the lagoon including intertidal assemblage Quinqueloculina seminula - Q. laevigata and Affinetrina quadrilateralis-Neorotalia calcar and a few other forms. It is not possible to relate these benthic foraminifers to the microforaminiferal linings of the present study.
A further study of microforaminiferal palynomorph assemblages was carried out to investigate palaeoenvironmental and palaeoecological utility of foraminiferal palynomorphs in absence of any foraminiferal data in marine and brackish water environments [19]. The study demonstrated that there are significant differences in relative abundances among informally described morphotypes of foraminiferal palynomorphs in various environments. In the intertidal and mangrove environments smaller benthic calcareous foraminifers were common. However, in the coral reef environments smaller benthic, both calcareous and porcelaneous forms were observed. Common occurrence of foraminiferal palynomorphs related to small-sized opportunistic Ammonia has palaeoecological significance due to its abundance in the mangrove sediments in proximity to roots of mangrove plant Avicennia marina. No foraminiferal palynomorphs were observed in the algal mat environment. Thus, in absence of data on foraminiferal assemblages, relative abundances of foraminiferal palynomorph morphotypes may be used for distinguishing various coastal (brackish and marine) environments.
4.4.2. Thecamoebians
There are only two published accounts of thecamoebians from the Red Sea coast of the Arabian Peninsula [16,17]. Further, a report on the acid-resistant Cretaceous thecamoebian tests from the Arabian Peninsula [65] is the only other publication on thecamoebian studies from this region. A useful illustrated key for the identification of Holocene lacustrine thecamoebian taxa [67] is widely used for thecamoebian studies. There is an extensive review on testate rhizopod (Thecamoebian) research that discusses the several aspects of studies on thecamoebians and their usefulness in paleoenvironmental and (paleo)limnological research [68].
Thecamoebian occurrence ranges between rare to common in these samples (Table 4). The taxa observed were Arcella sp., Centropyxis aculeata, Centropyxis sp., Cyclopyxis sp., Difflugia sp., Euglepha sp., Geopyxella sylvicola, Thecamoebian types A, B, F, and ?Thecamoebian [17]. Centropyxis aculeata occurs in almost every lacustrine environment; however, it prefers warmer waters above thermocline. Its morphology ranges from spiny to spineless. The occurrence of Geopyxella sylvicola, a soil testate amoeba was reported from the tropical coastal lowlands of southwest India [47]. Thecamoebian type A resembles Arcella megastoma [65] which is known from fresh-water lakes, coastal wetlands, mouth of the rivers, and the sewage drainage system in India. Thecamoebian type B resembles Cyclopyxis sp. [65] which is known from Nilarevu mouth of Godavari River, on the eastern coast of peninsular India [66]. Thecamoebian type F resembles Centropyxis laevigata.
4.4.3. Tintinnomorphs
Tintinnids are protists of complex eukaryotic organisms and are part of the microzooplankton (size ranges between 20 and 200 µm) that inhabit primarily marine, but brackish and freshwaters as well [69]. They produce a chitinous shell, called lorica with an oral opening at the anterior end, and the posterior end may be conical, globular, or tubular. The shape of the lorica is species specific. However, there are considerable intraspecific morphological variations related to the life cycle stages of these ciliates [70].
van Waveren [71] introduced the term 'Tintinnomorphs' to emphasize that palynomorphs resembling tintinnids cannot always be separated into loricae or cysts and may even represent cysts of other protozoans. Da Silva et al. [69] expanded the concept of ‘Tintinnomorphs’ for the group of palynomorphs resembling organic remains of tintinnids, which are not always identifiable as true lorica, cysts or pouch, and may even represent other structures of distinct organisms, such as rotifers and turbellarians. Many loricae and cysts occur in palynological preparations, especially in modern marine and marginal marine sediments (see references in 69) and were classified as zoomorphs [72].
van Waveren [71] described, illustrated, and categorized tintinnomorphs in an informal morphological system. Tintinnomorphs resemble organic remains of tintinnids, but are not necessarily identifiable as true lorica, cysts or pouch. In the present study, 34 tintinnomorphs types 1 through 34 [16,17] were described using the terminology of van Waveren [71]. Later most of these morphotypes were transferred to the newly proposed form taxa [18] except for Tintinnomorph types 9, 11, 12, 13, 14 and 16 described from the intertidal sediments (Table 4).
There is a possibility that tintinnomorphs described here might be Neorhabdocoela oocytes [73], because there is a morphological resemblance between them. Tintinnomorphs, as stated earlier, is a collective term for loricae, cysts and stalked pouches of Tintinnids and some of them appear morphologically like oocytes (egg capsules) of aquatic flatworms of the order Neorhabdocoela [73]. Neorhabdocoela oocytes are typically elliptical to spherical and of dark brown color, relatively larger in size (60 - 600 µm), but tintinnomorphs in contrast are usually cup-shaped, oval and of diverse shapes, pale yellow and of relatively smaller size. Neorhabdocoela oocytes occur predominantly in fresh-water environments, however, present assemblage is reported from the intertidal environment.
A high morphological diversity of tintinnomorphs indicates high diversity of Tintinnids in the southern Red Sea zooplankton; however, their numerical presence is usually rare in the intertidal sediments. Abu Zaid and Hellal [74] published an assemblage of modern tintinnids from the Hurghada coast of Red Sea in Egypt. However, it is difficult to assign a species or genus name to the titinnomorphs just based on the shape of lorica or cyst.
4.4.4. New Form Taxa (18)
Palynological slides of Quaternary-Recent marine and brackish water sediments invariably comprise a variety of vase-shaped palynomorphs. They are of variable sizes, could be hyaline or agglutinated and their colour ranges between yellowish to dark brown, and sometimes greyish. Biological affinities of such palynomorphs are debatable, generally palynologists relate them to tintinnid loricae and cysts, thus known as tintinnomorphs. However, similar morphotypes are known from lacustrine sediments as well and considered to be oocytes or resting eggs (6-200 µm) of Neorhabdocoela, a small, soft-bodied flatworm belonging to class Turbellaria, phylum Platyhelminthes, found in freshwater environments all over the world. Sometimes their affinity has also been related to testate amoebae. Palynological literature reports occurrences of many such palynomorphs that were described primarily based on a variety of their shapes, such as vase, urn, flask, funnel, tube, spherical and shapes that could be morphological variants among them. Identification of such forms is confusing because there is no standard nomenclature to describe them. Although attempts to informally describe them in various morphological groups were made, confusion prevails in their identifications because they do not have any formal taxonomic identity as genus and species.
Kumar (18) proposed new form genera and form species from a large morphologically diverse population of tintinnomorphs recovered from the southern Red Sea coastal environments of Saudi Arabia [16,17]. From these assemblages two new form genera Katora, and Mangrovia, and six new form species K. arabiaca, K. elongata, K. oblonga, K. twinmorpha, M. redseaensis, and M. hallii were proposed. They occur in all the environments studied and their abundance ranges between rare to common (Table 4).
4.4.5. Crustacean and Annelid Palynomorphs
The crustacean and annelid remain found in these palynomorph assemblages are diverse and occur in all samples; though their absolute numbers range between rare to few. A total of 17 forms including 12 crustacean and seven annelid occur in these samples (Table 6). A neoichnological study reported several biogenic traces that occur in the intertidal and supratidal flats of the same locality as the present study [75]. They were interpreted to have been formed by gastropods, land hermit crabs Coenobita clypeatus, ghost crab Ocypode quadrata, insects, beetles, sand wasps, and annelid species Arenicola marina, a large marine worm of the phylum Annelida; it is also known as lugworm or sandworm. Copepods (Subphylum Crustacea; Subclass Copepoda) inhabit the area of the present study as well. They are microcrustaceans (0.5-2 mm in size), normally occur in nearly every freshwater and saltwater habitat as planktic or benthic organisms. They have two pairs of antennae and a single red compound eye; are mostly crawlers, walkers, and burrowers in terrestrial environments, but still live where water is generally available [76].
It is suggested that chitinous remains of crustaceans and annelids observed in this study should most likely belong to these invertebrate animals. At present it is not always possible to relate such palynomorphs to their parent organisms. More work needs to be done on this matter.
4.4.6. Miscellaneous and Indeterminate Palynomorphs
1. Miscellaneous palynomorphs
Miscellaneous palynomorphs include tracheid, stomata; leaf epidermis showing stomata; charcoal; and phytoliths. They are rare or few in their occurrences and do not occur in all samples (Table 7). Leaf epidermis, probably of grass with stomata as well as phytoliths indicate the presence of family Poaceae in the region, but no Poaceae pollen was observed in these assemblages. The presence of charcoal indicates either natural forest fires or possibly wood burning due to human activity.
2. Indeterminate palynomorphs
A total of seven indeterminate forms were identified and informally named as A, B, C, D, E, F, and G. Most of these indeterminate palynomorphs occur rarely in the samples (Table 7). A tentative suggestion about their affinity is given wherever possible, within brackets. They are Indeterminate form A (may be worm oocytes with operculum); indeterminate form B (Algal spore?); indeterminate form C (Cyanobacteria?); indeterminate form D (Pseudoschizaea/Concentricystis sp.?); indeterminate form E (Crustacean egg?); indeterminate form F (Algal cyst?) and indeterminate form G (Fungal sporangium?).
4.4.7. Ascidian and Sponge Spicules
Ascidian spicules are mineral microfossils (calcareous) thus cannot be considered as palynomorphs. However, they occur in palynological slides along with other palynomorphs. Ascidians (Phylum Chordata, Class Ascidiacea), also known as sea squirts, are sac-like marine invertebrate filter feeders characterized by a tough outer "tunic" made of the polysaccharide cellulose. They are sessile animals that remain attached to their substratum, such as rocks and shells. They are found in all marine habitats from shallow water to the deep sea. Ascidians generate calcareous spicules (usually between 15 µm and 100 µm ) of characteristic shapes that accumulate in their tunic or internal tissues. Spicule morphology is quite diverse, thus is significant in ascidian taxonomy [77,78]. A brief overview of the fossil record of ascidian spicules is provided [79,80].
The present assemblage contains forms that range in size between 26 µm and 38 µm. This size range compares well with the forms described from Quaternary piston core of muddy sediments deposited by turbidity currents from offshore Argentina [80] where average size ranges between 25 µm and 30 µm. The ascidian spicules described from Argentina are from shallow marine environments that were transported to deeper marine environments by turbidity currents.
Numbers of ascidian spicules in the present assemblage range between rare to few (Table 6). They are not identifiable with published forms; informal names were used to describe them [17]. Ascidian spicules were observed only in the Sabia Island coral reef assemblage. They were informally described using the terminology of Lukowiak [79] and Lukowiak et al. [81]. Ascidian spicule types 1 through 5 belong to the Didemnidae Family and the ascidian spicule type 6 belongs to the Polycitoridae Family [77].
Ascidian spicules type 1 has close morphological resemblance with Didemnum-like spicule A described from Miocene of Moldova [81]. However, the Moldovian specimens are significantly larger; their size ranges between 90 µm to 100 µm and has only 10 rays. Ascidian spicules type 2 has close morphological resemblance with Trididemnum-like spicule B described from Miocene of Moldova [81]. However, the Moldovian specimens are significantly larger; their size is 100 µm and has only 8 to10 rays. Ascidian spicules type 3 resembles Didemnid spicule type II described from Eocene of Australia [79]. However, the Australian specimens are larger measuring 60 µm. Ascidian spicules type 4 resembles Didemnid spicule type III described from Eocene of Australia [79]. However, the Australian specimens are larger measuring 100 µm. Ascidian spicules type 5 is triangular, Y-shaped form, with three long, slender, and conical rays with pointed ends. Length of each ray varies between 76-36 µm and maximum width at the center is 23.6 µm. Surface of rays is smooth. Ascidian spicules type 6 resembles Cystodytes cf. dellechiajei described [79] from Eocene of Australia. However, the Australian specimens are 100-120 µm in diameter with a smooth central part. The present specimen is significantly smaller and has a granular internal surface.
One specimen of sponge spicule was observed in sample SI2.
4.4.8. Polychaete Chaetae
Rare specimens of setae of polychaetes were observed in the Sabia Island coral reef assemblage. chaeta (also spelled cheta or seta; plural chaetae) is a chitinous bristle found in insects, arthropods, and annelid worms. The Polychaeta or the bristle worms are generally marine annelid worms having a segmented body. Each segment has a pair of fleshy protrusions called parapodia that bear chitinous bristles called chaetae. Common Polychaetes are lugworm (Arenicola marina) and sand worm that were reported from this area in a neoichnological study by the author [75]. Chaetae shape and size vary considerably and are often species-specific; main types include capillary, compound, pseudo compound, hooked and many more [82].
5. Discussion
This study describes palynomorph assemblages and their source from five different coastal environments of southern Red Sea, Saudi Arabia; they are mangrove swamps, Mid Holocene wadi Hali paleochannel section, algal mat, and Sabia Island coral reef, and intertidal environment. Numerical distribution of palynomorphs from these coastal environments is presented in Table 1, Table 2, Table 3, Table 4, Table 5, Table 6 and Table 7, and percentage representation of palynomorph groups is shown in Figure 6, Figure 7, Figure 8 and Figure 9. Provenance of various palynomorphs and palynomorph groups is discussed.
5.1. Mangrove Swamp Environment
Mangrove swamps in this region cover small patches of mangrove stands and the areas covered by them vary considerably. They are distributed in restricted, quieter environments, occasionally they also inhabit higher energy intertidal environments. The southern Red Sea coast has a relatively denser growth of mangroves than the northern coast. This is because of the tropical climate, more wadis, higher rainfall and nutrients, and less saline waters along this part of the Red Sea coast [83].
Palynomorph assemblages in the mangrove swamp environment show higher numerical abundance and diversity than the Wadi Hali paleochannel section and show closer similarity to the assemblages obtained from the intertidal samples [16]. Rare occurrences pollen and spores, which are mostly of angiosperm affinity except for the only gymnosperm Juniperus pollen were observed. The occurrence of dinoflagellate cysts is rare; however, four new types of dinoflagellate cyst types G, H, I and J were informally described. Only rare specimens of the new dinoflagellate cysts were observed. Algal remains are more or less similar to the assemblage described from intertidal environments, except that several algal remains could not be identified (indeterminate) and their representation in the assemblage ranges between few to common. This indicates a relatively higher diversity of dinoflagellate cysts and algal remains in the mangrove swamps than in the paleochannel section. The diversity and numerical abundance of fungal remains is higher and ranges between rare to common. Although the diversity of fungal remains is as high in the intertidal samples, their numerical abundance is lower and ranges between rare to few.
Generally, low numerical abundances and higher taxonomic diversity among the protists was observed in the mangrove swamp samples that included several types of thecamoebians, microforaminiferal test linings, and tintinnomorphs. Thecamoebian taxa found in these samples are like the intertidal samples, but five new types were informally described; they are Thecamoebian types A, B, C, D, and E. All these new forms occur as rare, but numerical abundance of thecamoebian type C ranges from rare to common. Several microforaminiferal test linings described from the intertidal samples occur in mangrove samples, and their numerical abundance ranges between rare to few.
The six new form species K. arabica, K. elongata, K. oblonga, K. twinmorpha, M. redseaensis, and M. hallii occur commonly in this environment (Table 4). Numerical abundance of invertebrate remains is significantly higher than that of other groups of palynomorphs. These can be grouped into annelid, and crustacean remains. Their numerical abundance ranges between common to abundant. Rare remains of an ostracod species cf. Leptocythere sp. were observed in a few samples.
5.2. Wadi Hali Paleochannel Section
This section is possibly a mid-Holocene paleochannel deposit, is located about 2 km inland from the present-day coastline. There is no age control on these samples. However, geological, paleoenvironmental and paleoclimatic evidence provide certain clues to its age. In the Wadi Hali Quadrangle extensive flood plain deposits are present along the lower parts of major wadis. These deposits consist of silt, fine grained sand, and clay [29]. These clastic sediments were most likely deposited during the wetter conditions 6-9 ka BP in southern Saudi Arabia [84]. The middle Holocene wet phase has been dated to be 6.0 ± 0.6 ka BP [85,86]. Thus, it is suggested that the paleochannel deposit is most likely of middle Holocene age.
Wadi Hali is an example of an intermittent river [87]. Such rivers are dry most of the time, but they do discharge water during flooding events when they erode, transport and deposit huge amounts of sediment. The Wadi Hali paleochannel section may represent such a fluvial deposit. Mud cracks were observed on the surface of the outcrop along with traces of fossil plant roots in the section [75].
The percentage distribution of various palynomorph groups is shown in Figure 6A and Figure 8A. The assemblage contains pollen grains, spores, algal and fungal remains, protists like thecamoebians and tintinnomorphs along with crustacean and worm remains. Except for crustacean remains, all other palynomorphs occur as rare to few. Only crustacean antennae are abundant in all samples. Pollen, spores, algal and fungal remains are allochthonous that were transported by wind and water in this fluvial deposit. No dinoflagellate cysts and only one specimen each of microforaminiferal test linings and Katora arabica were observed.
The occurrence of thecamoebian taxa like Arcella sp., Centropyxis aculeata, Difflugia sp., Euglepha sp. and Geopyxella sylvicola in this section is problematic. Since these forms are benthic protists, they require a quiet standing body of water to survive. They are not expected to inhabit an intermittent river as described because they are dry most of the time and water moves extremely fast due to storm related flooding events [87]. Thus, it is suggested that during the Middle Holocene wetter climatic conditions, the Wadi Hali paleochannel section probably represented an isolated body of water such as a pond close to one of the channels of Wadi Hali. Few different taxa of thecamoebians inhabited this pond along with crustaceans and worms. The occurrence of abundant crustacean antennae in this section probably is due to their accumulation at a certain place because of sorting by waves within the suggested body of water. Isolated specimens of Katora arabica and microforaminiferal test linings are contaminants in this assemblage, and probably were brought in from the intertidal sediments during periodic storms that impacted this region [37]. A variety of tintinnomorphs and microforaminiferal test linings were reported from the intertidal sediments in this area [16].
5.3. Algal Mat Environment
An algal mat is a sheet-like accumulation of microbial layer made of filamentous algae and/or cyanobacteria on marine or fresh water soft bottoms normally in the warmer regions. Usually, they occur in shallow marine subtidal to supratidal environments, as well as in lakes and swamps [87]. The numerical distribution of palynomorphs in this environment is shown in Table 1, Table 2, Table 3, Table 4, Table 5, Table 6 and Table 7. Figure 7A and Figure 8C show proportional representation of palynomorph groups in this environment.
Palynological slides of these samples are characterized by the presence of abundant green filamentous, branched alga and numerous fungal filaments which are branched and unbranched, septate and aseptate. Pollen and spores occur rarely. Likewise, dinoflagellate cysts and algal remains occur rarely but there are a few occurrences of algal cells (indeterminate). The occurrence of form taxa ranges from rare to common, especially Katora arabica, K. oblonga, and Mangrovia redseaensis are common to very common. Diversity and numerical abundance of fungal remains vary between rare to common, and Callimothallus type fruit body and Glomus sp. are prominent among the fungal remains. The occurrence of Protists range between rare to common and only thecamoebians were observed. Crustacean remains are a significant part of the algal mat samples, and they range between few to abundant. Crustacean antennae and exoskeleton are major contributors to the palynomorphs of invertebrate affinity. Unidentified worm remains are important in the assemblage as well.
5.4. Sabia Island Coral Reef Environment
This small island in the Red Sea, is about 20 km away from the nearest coast of Saudi Arabia and is surrounded by coral reefs. Present status of Red Sea coral reefs is discussed [88,89]. Palynomorph distribution is shown in the tables and Figure 7B and Figure 8D show proportional representation of palynomorph groups in this environment.
Since these two samples represent the marine environment, unlike other samples that represented marginal marine environments, the palynomorph assemblages are quite distinct in them. As expected, rare specimens of only three pollen were observed, that must have been blown-in by wind from the mainland. No spores were observed. Dinoflagellate cysts and acritarchs occur rarely, but isolated specimens of informally described new dinoflagellate cysts were observed; they are dinoflagellate cysts K, L, M and N. Dinoflagellate cyst D earlier described from the intertidal samples [16] occur in this assemblage as well. Algal remains are a prominent group dominated by various types of algal cells and broken specimens of marine diatoms. Rare occurrences of fungal remains were observed, and their composition is like algal mat samples.
Protists are represented by a variety of microforaminiferal test linings. Their numerical abundances and diversity dominate the assemblage. There are no tintinnomorphs, but Katora arabica, K. oblonga, and K. elongata occur commonly. Thecamoebians occur rarely; that includes Centropyxis aculeata, Geopyxella sylvicola and an informally described form is thecamoebian type F. Presence of Centropyxis aculeata and Geopyxella sylvicola in this assemblage raises questions, because they are not known from the marine environments, instead they inhabit fresh to brackish water environments. Centropyxis aculeata is known to occur in a wide range of environments ranging from fresh to brackish water and prefers warmer waters above the thermocline [68]. Geopyxella sylvicola, is a soil testate amoeba found in tropical coastal lowlands in southwest India [47]. Thecamoebians were also reported from the Godineau River Estuary, Gulf of Praia, off the island of Trinidad [90], but none from the marine environments. It seems that the thecamoebian taxa that occur in these samples got contaminated and most likely were transported by storm winds from the intertidal environments.
The palynomorphs having affinity with invertebrates are once again dominated by crustacean remains, mainly antennae and skeleton, and their occurrence ranges between rare to abundant. However, annelid remains occur rarely. The most striking addition to this assemblage is the occurrence of ascidian spicules. Six types of ascidian spicules from type1 through type 6 were informally described [17]. Rare occurrence of sponge spicules and phytoliths were also recorded.
5.5. Intertidal Environment
Distribution of palynomorphs in these samples are presented in the tables. Palynomorph assemblage varies considerably among the samples. Generally, the occurrence of pollen and spores, dinoflagellates and algal remains, and fungal remains range from rare to very common in different samples. Protists like tintinnomorphs, thecamoebians, and microforaminiferal linings rarely occur. Crustacean remains are abundant and annelid palynomorphs are common to very common in various samples. Katora spp. occur as rare to common in these samples.
Sample L1 has many microscopic remains of protists and invertebrates. Crustacean antennae and other remains are abundant in this sample. Pollen and spores, algal and fungal remains have only rare to few specimens. Palynomorph composition in samples L2 and L3 are very similar to sample L1. The assemblage of this sample is dominated by a few protists and invertebrates such as crustacean and annelid palynomorphs. Rare specimens of microforaminiferal linings occur in most of the samples. Palynomorph assemblages of samples L4, L5 and L6 have rare fungal hyphae, spores, and unidentifiable forms. Predominant palynomorphs in all these samples are derived protists and invertebrates. Crustacean antennae are either abundant or very common in these samples. Sample L7 is almost barren, has few cuticles, trachieds, fungal remains, and rare fragments of charcoal.
6. Conclusions
Table 7 shows the summary of relative distribution of palynomorphs and palynomorph groups in all the five southern Red Sea coastal environments. All five groups of palynomorphs occur in all these environments, however their relative abundances vary significantly among them (Figure 6, Figure 7, Figure 8 and Figure 9). The intertidal environment is characterized by mostly rare occurrences of pollen, spores, dinoflagellate cysts and other algal remains (Figure 9). The same is true for fungal remains, except for fungal fruit bodies and few indeterminate forms that are common to very common. Palynomorphs having affinity with protists and invertebrates dominate the assemblage. The presence of tintinnomorphs, crustacean and annelid palynomorphs range between few to very common. The Mid Holocene Wadi Hali paleochannel section has an impoverished assemblages and has rare occurrences of pollen, spores, fungal remains, algal remains and no dinoflagellate cysts. Among protists and invertebrates, only crustacean remains are common to abundant. Mangrove swamp environment has a relatively richer composition of palynomorphs compared with the intertidal assemblage. Pollen, spores, algal remains and dinoflagellate cysts occur rarely, but unlike intertidal assemblages, mangrove assemblages have a higher diversity and numerical abundance of palynomorphs. Distribution of protists and invertebrates is like the intertidal environment, where thecamoebians, tintinnomorphs and microforaminifera range between few to common. Crustacean and annelid remains are common to abundant. Algal mat environment is characterized by an abundance of filamentous green algae where fungal hyphae are the second most common form. It has rare to few occurrences of pollen and spores, fungal remains and dinoflagellate cysts and other algal remains. Protists and invertebrate palynomorphs are rare to few but crustacean remains are common to abundant. Sabia Island coral reef environment mostly has rare presence of pollen, spores, fungal remains and dinoflagellate cysts and algal remains. Protists and invertebrates dominate the assemblage; tintinnomorphs, microforaminifera and crustacean remains are few to abundant. No crustacean antennae were observed. Ascidian spicules occur only in this environment and are common to very common. A few specimens of phytoliths were also observed in this assemblage. Katora spp. occur as rare to very common in all the environments except for the paleochannel section. Mangrovia spp. occurs only in mangrove and algal mat environments. Few charcoal fragments occur in most samples.
This study provides comprehensive palynological baseline data about the southern Red Sea coastal environments indicating their flora and fauna that defines the present health of the coastal ecosystem. Study of palynomorphs occurring as fossils or subfossils in various coastal sedimentary settings represent local and regional climate, and environments.
Acknowledgments
I thank my former colleagues from King Fahad University of Petroleum and Minerals (KFUPM), Saudi Arabia: Lamidi Babalola, Michael Kaminski, and Khalid Ramadan, who joined me during the field trip and helped in sample collection. Asif Khan prepared the location map and Osman Abdullatif joined us to discuss various aspects of this project. I thank KFUPM for financial support for the field trip. I am thankful to Dr. Peta Mudie (Geological Survey of Canada), Dr. Francine McCarthy (Brock University, Canada), Dr. Kazumi Matsuoka (Kyoto, Japan), and Dr. Kenneth Mertens (Institut Français de Recherche pour l'Exploitation de la Mer, France) for helping me in few identifications.
References
- van Geel, B. 1972. Palynology of a section from the raised peat bog ‘‘Wietmarscher Moor’’ with special reference to fungal remains. Acta Botanica Nederland, 21: 261–284. [CrossRef]
- van Geel, B. 1976. Fossil spores of Zygnemataceae in ditches of a pre-historic settlement in Hoogkarspel (The Netherlands). Rev. Palaeobot. Palynol., 22: 337–344. [CrossRef]
- van Geel, B. 1986. Application of fungal and algal remains and other microfossils in palynological analyses. In: Bergland BE, (Ed.) Handbook of Holocene palaeoecology and palaeohydrology. New York: Wiley, :497–505.
- van Geel, B. 2001. Non-pollen palynomorphs. In: Smol JP, Birks HJB, Last WM, (Eds.). Tracking environmental change using lake sediments, Volume 3: Terrestrial, algal and siliceous indicators. Dordrecht: Kluwer Academic Publishers, :99–119.
- Mudie PJ, Marret F, Rochon R, Aksu AE. 2010. Non-pollen palynomorphs in the Black Sea corridor. Vegetation History and Archaeology, 19: 531-544. [CrossRef]
- Mudie PJ, Leroy SAG, Marret F, Gerasimenko N, Kholeif SEA, Sapelko T, Filipova-Marinova M. 2011. Non-pollen palynomorphs: Indicators of salinity and environmental change in the Caspian-Black Sea-Mediterranean corridor. The Geological Society of America Special Paper 473: 1-27.
- Hartman JD, Bijl PK, Sangiorgi F. 2018. A review of the ecological affinities of marine organic microfossils from a Holocene record offshore of Ad´elie Land (East Antarctica). Journal of Micropalaeontology, 37 (2): 445–497. [CrossRef]
- Mudie PJ, Marret F, Gurdebeke PR, Hartman JD, Reid PC. 2021a. Marine dinocysts, acritarchs and less well-known NPP: tintinnids, ostracod and foraminiferal linings, copepod and worm remains. In: Marret, F., O’Keefe, J., Osterloff, P., Pound, M. and Shumilovskikh, L. (Eds) Applications of Non-Pollen Palynomorphs: from Palaeoenvironmental Reconstructions to Biostratigraphy. Geological Society, London, Special Publications, 511. [CrossRef]
- Mudie PJ, Yanko-Hombach VV, Mudryk I. 2021b. Palynomorphs in surface sediments of the North-Western Black Sea as indicators of environmental conditions. [CrossRef]
- Bradford, MR. 1975. New dinoflagellate cyst genera from the recent sediments of the Persian Gulf; Canadian Journal of Botany, 53: 3064-3074. [CrossRef]
- Bradford MR, WALL DA. 1984 The distribution of Recent organic-walled dinoflagellate cysts in the Persian Gulf, Gulf of Oman, and northwestern Arabian Sea; Palaeontographica, Abteilung B, 192: 16-84, pl. 1-6.
- Matsuoka K, Yurimoto T, Chong VC, Man A. 2017. Marine Palynomorphs Dominated by Heterotrophic Organism Remains in the Tropical Coastal Shallow-Water Sediment; the Case of Selangor Coast and the Estuary of the Manjung River in Malaysia. Paleontological Research, 21(1):14-26. [CrossRef]
- Kholeif SEA. 2004. Palynology and paleovegetation reconstruction in late Quaternary sediments in the southern Suez Isthmus, Egypt. Journal of African Earth Sciences, 22 (1): 93-112.
- Kholeif SEA, Mudie PJ. 2009. Palynological records of climate and oceanic conditions in the late Pleistocene and Holocene of the Nile Cone, southeastern Mediterranean, Egypt. Palynology 33:1–24. [CrossRef]
- Kholeif SEA. 2007. Palynology of mangrove sediments in the Hamata Area, Red Sea coast, Egypt: vegetation and restoration overview. In: Isermann, M., and Keihl, K. (Eds). Restoration of Coastal ecosystems. Coastline Reports, 7:5-16. Palynology and paleovegetation reconstruction in late Quaternary sediments in the southern Suez Isthmus, Egypt.
- Kumar, A. 2020. Palynology of the recent intertidal sediments of the Southern Red Sea Coast of Saudi Arabia. Palynology, 45(1): 143-163. [CrossRef]
- Kumar, A. 2021. Palynomorph assemblages and mineral microfossils from various Southern Red Sea coastal environments of Saudi Arabia. Earth Science India, 14(2): 41-97. [CrossRef]
- Kumar, A. 2023a. New form taxa of non-pollen palynomorphs (NPP) from southern Red Sea coastal sediments of Saudi Arabia. Geophytology, 53(1): 1–24.
- Kumar, A. 2023b. Foraminiferal palynomorphs from the marine and brackish water sediments of the southern Red Sea coast of Saudi Arabia, their palaeoenvironmental and palaeoecological implications. Geophytology, 53(2): 109–132.
- Saxena RK, Kumar A. 2023. Fungal palynomorphs from southern Red Sea coastal sediments of Saudi Arabia. Geophytology, 53(2): 151–170.
- Alharbi OA, Phillips MR, Williams AT, Thomas T, Hakami M, Kerbe J, Niang AJ, El-Sayed H, Al-Ghamdi K. 2017. Temporal shoreline change and infrastructure influences along the southern Red Sea coast of Saudi Arabia. [CrossRef]
- Tarawneh QY, Faraj TK. 2020. The effect of anthropogenic activity on the extreme climate events and solar irradiation in Saudi Arabia. Arabia. [CrossRef]
- Kumar, A. 2009. Reclaimed islands and new offshore townships in the Arabian Gulf: Potential natural hazards. Current Science, 96(4): 480-485. [CrossRef]
- Khan AM, Kumar A. 2009. Impact of “urban development” on mangrove forests along the west coast of the Arabian Gulf. Earth Science India, 2(III): 159-173.
- Jado AR, Zölt JG.1984. Quaternary Period in Saudi Arabia. Vol. 2. Springer-Verlag, 360p.
- Hanauer, E. 1988. The Egyptian Red Sea: A Diver's Guide. Aqua Quest Publications, Inc. ISBN 9780922769049.
- Price ARG, Jobbins G, Shepherd ARD, Ormond RFG. 1998. An integrated environmental assessment of the Red Sea coast of Saudi Arabia. Environmental Conservation 25 (1): 65–76. [CrossRef]
- Jado AR, Hotzl H, Roscher B. 1990. Development of sedimentation along the Saudi Arabian Red Sea coast. J. K. A. U.: Earth Sciences, Vol. 3, Special Issue: 1st Saudi Symposium on Earth Sciences, Jeddah, 1989: 47-62. [CrossRef]
- Prinz, WC. 1984. Explanatory notes to the Geological Map of the Wadi Haliy Quadrangle, sheet 18E, Kingdom of Saudi Arabia. Ministry of Petroleum and Mineral Resources, Jeddah, Saudi Arabia. 13 p.
- El-Naggar SM, El-Husseini N. 2001. Pollen Atlas of the Flora of Egypt. 2. Species of Polygonaceae. Taeckholmia, 21(1):143-151.
- Al-Dabbas MA, Abba MA, Al-Khafaji RM. 2010. Dust storm loads analyses- Iraq. Arabian Journal of Geoscience. [CrossRef]
- Awadh SM, Ali MO, Ali RA. 2010. Mineralogy and palynology of the Mesopotamian plain sediments, Central Iraq. Arabian Journal of Geoscience. [CrossRef]
- Shamso EM, Toshiyuki F. 2012. The Pollen Flora of Faiyum, Egypt I Archichlamydeae. Taeckholmia, 32: 58-70.
- Al-Ameri TK, Jassim SY. 2009. Environmental changes in the wetlands of southern Iraq based on palynological studies. Arabian Journal of Geoscience. [CrossRef]
- Thomas, J. 2011. Plant Diversity in Saudi Arabia. Herbarium, Department of Botany and Microbiology, King Saud University, Riyadh, Saudi Arabia. http://plantdiversityofsaudiarabia.info/index.htm Last accessed on July 25, 2020.
- Al-Frayh A, Reilly H, Harfi HA, Hasnain SM, Thorogood R, Wilson JD. 1989. A 12-Month Aerobiological Survey of Pollen in Riyadh. Annals of Saudi Medicine, 9(5): 443-447. [CrossRef]
- Kumar, A. 2013. Invited Paper: Natural hazards of the Arabian Peninsula; their causes and possible remediation. In: Sinha, R, Ravindra, R (Eds.) Earth System Processes and Disaster Management, Society of Earth Scientists Series 1. Springer-Verlag, Berlin, Heidelberg, 155-180. [CrossRef]
- Ali, MO. 2014. Palynological evidence on paleoclimate and paleoenvironmental changes during Holocene of Al-Hussaynia District, Central Iraq. [CrossRef]
- Cole, JB. 1992. Fresh water dinoflagellate cysts and acritarchs from Neogene and Oligocene sediments of the South China Sea and adjacent areas. In: Head, MJ, Wrenn JH. (Eds.), 1992, Neogene and Quaternary Dinoflagellate Cysts and Acritarchs: American Association of Stratigraphic Palynologists Foundation, 181-196.
- Wainman CC, Mantle DJ, Carey HC, McCabe PJ. 2019. Possible freshwater dinoflagellate cysts and colonial algae from the Upper Jurassic strata of the Surat Basin, Australia, Palynology, 43(3): 411-422. [CrossRef]
- Mertens KN, Rengefor K, Moestrup Ø, Ellegard M. 2012. A review of recent freshwater dinoflagellate cysts: taxonomy, phylogeny, ecology and palaeocology. Phycologia, 51: 612–619. [CrossRef]
- McCarthy FMG, Mertens KN, Ellegaard M, Sherman K, Pospelova V, Ribeiro S, Vercauteren D. 2011. Resting cysts of freshwater dinoflagellates in southeastern Georgian Bay (Lake Huron) as proxies of cultural eutrophication. Rev. Palaeobot. Palynol., 166(1-2): 46-62. [CrossRef]
- McCarthy FMG, Riddick NL, Volika O, Danesh DC, Krueger AM. 2018. Algal palynomorphs as proxies of human impact on freshwater resources in the Great Lakes region. Anthropocene, 21:16–31. [CrossRef]
- Krueger AM, McCarthy FMG. 2016. Great Canadian Lagerstätten 5. Crawford Lake – A Canadian Holocene Lacustrine Konservat-Lagerstätte with Two-Century-Old Viable Dinoflagellate Cysts. Geoscience Canada, 43:123–132. [CrossRef]
- Elshanawany R, Zonneveld K, Ibrahim MI, Kholeif SEA. 2010. Distribution patterns of recent organic-walled dinoflagellate cysts in relation to environmental parameters in the Mediterranean Sea, Palynology, 34: 233-260. [CrossRef]
- de Vernal, A. 2009. Marine palynology and its use for studying nearshore environments. Deepsea to Coastal Zones: Methods and Techniques for Studying Paleoenvironments. IOP Conf. Series: Earth and Environmental Science 5, 012002. [CrossRef]
- Limaye RB, Kumaran KPN, Nair KM, Padamlal D. 2007. Non-pollen palynomorphs as potential palaeoenvironmental indicators in the Late Quaternary sediments of the southwest coast of India. Current Science, 92(10): 1370-1382.
- Kumaran KPN, Limaye RB, Nair KM, Padamlal D. 2008. Palaeoecological and palaeoclimate potential of subsurface palynological data from the Late Quaternary sediments of South Kerala Sedimentary Basin, southwest India. Current Science, 95(4): 515-526.
- Hoshaw RW, McCourt RM. 1988. The Zygnemataceae (Chlorophyta): a twenty-year update of research. Phycologia, 27: 511-548. [CrossRef]
- Head, MJ. 1993. Dinoflagellates, sporomorphs, and other palynomorphs from the Upper Piocene St. Earth Beds of Cornwall, southwestern England. The Paleontological Society Memoir 31, Supplement to Journal of Paleontology, 67(3) Part III of III, 1-62.
- Zippi, PA. 1998. Fresh water algae from the Mattagami Formation (Albian), Ontario: Paleoecology, botanical affinities, and systematic taxonomy. Micropaleontology, 44 (suppl. 1): 1-78. [CrossRef]
- Saxena RK, Kumar A. 2023. Fungal palynomorphs from southern Red Sea coastal sediments of Saudi Arabia. Geophytology, 53(2): 151–170.
- Kalgutkar RM, Jansonius J. 2000. Synopsis of fungal spores, mycelia and fructifications. A.A.S.P. Contribution Series 39:1–423.
- Cook EJ, van Geel B, van der Kaars S, van Arkel J. 2011. A review of the use of non-pollen palynomorphs in palaeoecology with examples from Australia. Palynology, 35 (2):155–178. [CrossRef]
- Anderson RS, Homola RA, Davis RB, Jacobson Jr. GL.1984. Fossil remains of the mycorrhizal fungal Glomus fasciculatum complex in post glacial lake sediments from Marine. Can. J. Botany, 62: 2325-2328. [CrossRef]
- Elsik, WC. 1996. Fungi-Chapter 10; in Palynology: principles and applications. Jansonius, J and McGregor, DC (Eds.); American Association of Stratigraphic Palynologists Foundation, Publishers Press, Salt Lake City, Utah, USA, 1: 293-306.
- Stubblefield SP, Taylor TN. 1988. Tansley Review No. 12. Recent advances in Paleoemycology. New Phytology, 108: 3-25.
- Taylor, TN. 1990. Fungal associations in terrestrial paleoecosystem. Trends in Ecology and Evolution, 5:21-25.
- Saxena RK, Tripathi SKM. 2011. Indian Fossil Fungi. The Palaeobotanist, 60 (1): 1-208. [CrossRef]
- Mathison SW, Chmura GL. 1995. Utility of microforaminiferal test lining in palynological preparations. Palynology, 19: 77–84. [CrossRef]
- Stancliffe, RPW. 1996. Chapter 13. Microforaminiferal linings. In: Jansonius, J., and McGregor, D.C., eds., Palynology: Principles and Applications, p. 373–379. Dallas, Texas, American Association of Stratigraphic Palynologists Foundation, 1:373-379.
- Monga P, Kumar M, Joshi Y. 2015. Morphological variations and depositional processes of microforaminiferal linings in the early Tertiary sediments of northeastern and northwestern India. The Palaeobotanist, 64(2): 129–138. [CrossRef]
- Mudie JP, Yanko-Holmbach V. 2019. Microforaminiferal linings as proxies for paleosalinity and pollution: Danube Delta example. Micropaleontology, 65 (1): 1-19.
- Al-Dubai TA, Abu-Zied RH, Basaham AS. 2018. Diversity and distribution of benthic foraminifera in the Al-Kharrar Lagoon, eastern Red Sea coast, Saudi Arabia. Micropaleontology, 63 (5): 275–303. [CrossRef]
- Kumar, A. 2011. Acid-resistant Cretaceous thecamoebian tests from the Arabian Peninsula: a suggestion for study of agglutinated rhizipods in palynological slides. Journal of Micropalaeontology, 30: 1-5. [CrossRef]
- Farooqui A, Naidu TY. 2010. Thecamoebians and palynological assemblage in Gautami-Godavari River mouth, India: Environment and sea level since 3000 years. J Geol Soc India 75, 841–850. [CrossRef]
- Kumar A, Dalby AP. 1998. Identification key for Holocene lacustrine arcellacean (thecamoebian) taxa. Palaeontologia Electronica, 1(1): 1-39. [CrossRef]
- Patterson RT, Kumar A. 2002. A review of current testate rhizopod (Thecamoebian) research in Canada. Paleogeography, Paleoecology, Paleoclimatology, 180: 225-251. [CrossRef]
- da Silva WG, de Souza PA, van Waveren I. 2017. New Insights on the systematic classification of certain palynological taxa (tintinnomorphs) from Holocene deposits of the Coastal Plain of Southern Brazil. Revista Brasileira de Paleontologia, 20(3):321-332. [CrossRef]
- Agatha S, Laval-Peuto M, Simon P. 2013. The Tintinnid Lorica, in: The Biology and Ecology of Tintinnid Ciliates: Models for Marine Plankton, In: Dolan JR et al. (Eds.) The biology and ecology of tintinnid ciliates: models for marine plankton, p. 42–84. [CrossRef]
- van Waveren, IM. 1994. Tintinnomorphs from the deep-sea sediments of the Banda Sea (Indonesia). Scripta Geologica, 105: 27-51.
- Traverse, A. 1994. Sedimentation of palynomorphs and palynodebris: an introduction. In: A. Traverse (ed.) Sedimentation of organic particles, Cambridge University Press, p. 1–8.
- Haas, JN. 1996. Neorhabdocoela oocytes — palaeoecological indicators found in pollen preparations from Holocene freshwater lake sediments. Review of Palaeobotany and Palynology, 91: 371-382. [CrossRef]
- Abu Zaid MM, Hellal AM. 2012. Tintinnids (Protozoa: Ciliata) from the coast of Hurghada Red Sea, Egypt. Egyptian Journal of Aquatic Research, 38: 249–268. [CrossRef]
- Kumar, A. 2017. Recent biogenic traces from the coastal environments of the southern Red Sea coast of Saudi Arabia. Arabian Journal of Geoscience 10 (22): 3-11. [CrossRef]
- Glime JM. 2017. Arthropods: Crustacea – Copepoda and Cladocera. Chapt. 10-1. In: Glime, J. M. Bryophyte Ecology. Volume 2. Bryological Interaction. E book sponsored by Michigan Technological University and the International Association of Bryologists. Last updated 21 April 2017 and available at .
- Varol O, Houghton S. 1996. A review and classification of fossil didemnid ascidian spicules. J Micropaleontol, 15: 135–149. [CrossRef]
- Shenkar N, Swalla BJ. 2011. Global Diversity of Ascidiacea. PLoS ONE 6(6): e20657. [CrossRef]
- Lukowiak, M. 2012. First record of late Eocene Ascidians (Ascidiacea, Tunicata) from southwestern Australia. Journal of Paleontology, 86(3): 521–526. [CrossRef]
- Ferreira EP, Alves CF, Sanjinés AES, Alves MC. 2019. Ascidian spicules of Quaternary sediments from the lower slope of the Campos Basin (Brazil). Quaternary International, 508 (1) :116-124. [CrossRef]
- Lukowiak M, Dumitriu SD, Ionesi V. 2016. First fossil record of early Sarmatian didemnid ascidian spicules (Tunicata) from Moldova. Geobios, 49 (3): 201-209. [CrossRef]
- Hausen, H. 2005. "Chaetae and chaetogenesis in polychaetes (Annelida)". Hydrobiologia. 535-536: 37–52. [CrossRef]
- Khan, MA, Kumar A, Muqtadir A. 2010. Distribution of mangroves along the Red Sea coast of the Arabian Peninsula: Part-2: the southern coast of western Saudi Arabia. Earth Science India, 3 (3): 154-162.
- McClure, HA. 1976. Radiocarbon chronology of late Quaternary lakes in the Arabian desert. Nature, 263: 755-756. [CrossRef]
- Juyal N, Singhvi AK, Glennie, KW. 1998. Chronology and paleoenvironmental significance of Quaternary desert sediment in southeastern Arabia. In: Alsharhan AS, Glennie KW, Whittle GL, Kendall CGSC (Eds). Quaternary Deserts and Climate Change, Balkema, Rotterdam, p. 393-409.
- Glennie KW, Singhvi AK. 2002. Event stratigraphy, paleoenvironment and chronology of SE Arabian deserts. Quaternary Science Reviews, 21: 853-869. [CrossRef]
- Friedman GM, Sanders JE.1978. Principles of Sedimentology. John Wiley & Sons, New York. 792p.
- Wilkinson, C. 2008. Status of coral reefs of the world: 2008. Global Coral Reef Monitoring Network and Reef and Rainforest Research Centre, Townsville, Australia, 296 p.
- Rasul NMA, Stewart ICF. 2015. The Red Sea: The Formation, Morphology, Oceanography and Environment of a Young Ocean Basin. Springer Earth System Sciences. 620 p.
- Laut L, Clemente I, Martins MVA, Frontalini F, Raposo D, Belart P, Habib R, Fortes R, Lorini ML. 2017. Benthic Foraminifera and Thecamoebians of Godineau River Estuary, Gulf of Paria, Trinidad Island. Anuár i o do Inst i tut o de Geoc iênc ias, 40 (2):118-143. [CrossRef]
Figure 1.
Topographical map of the Arabian Peninsula showing locations of various countries and Red Sea. Location of study is marked by L. (modified after https://commons.wikimedia.org/wiki/File:Saudi_Arabia_Topography.png).
Figure 1.
Topographical map of the Arabian Peninsula showing locations of various countries and Red Sea. Location of study is marked by L. (modified after https://commons.wikimedia.org/wiki/File:Saudi_Arabia_Topography.png).
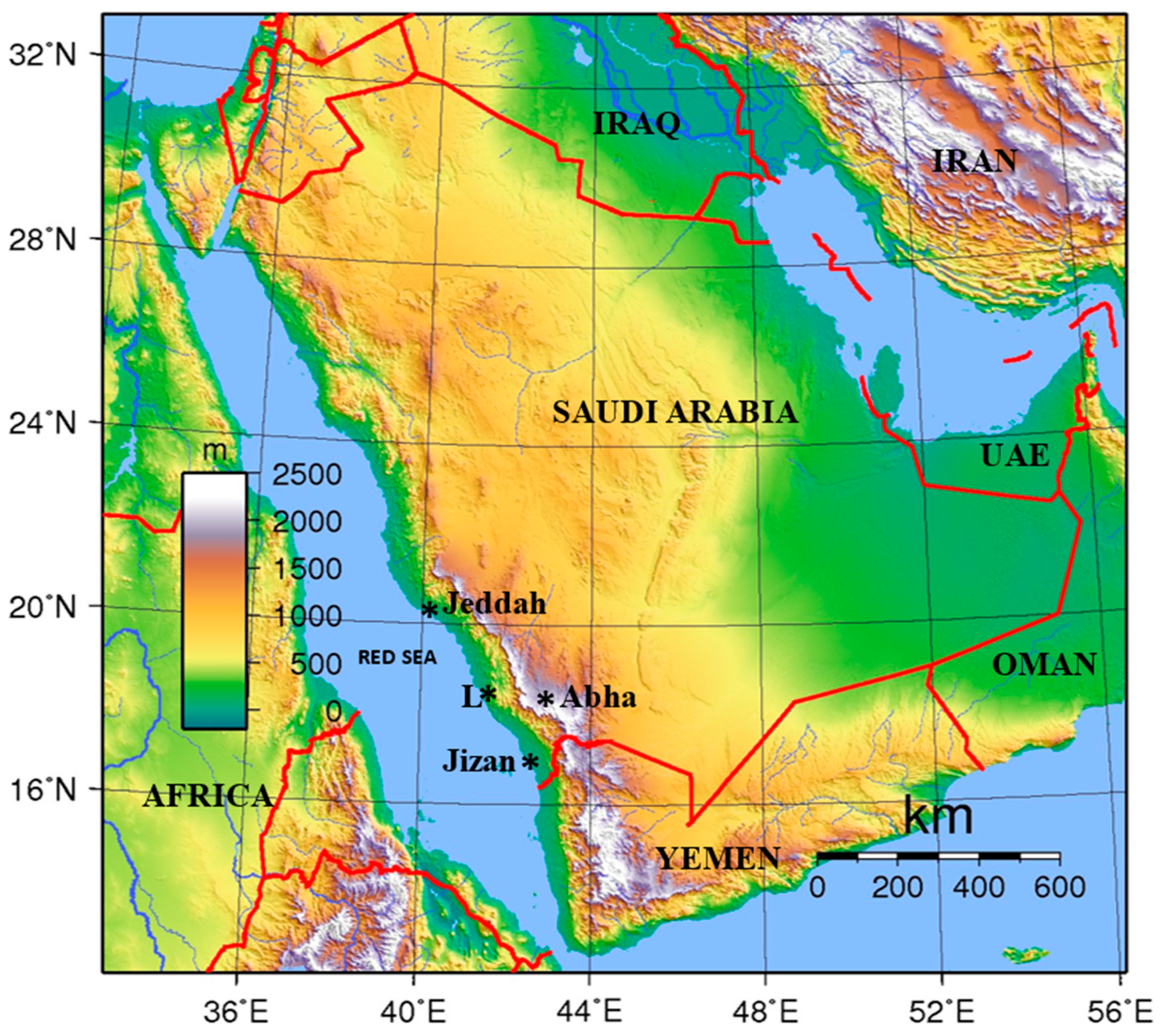
Figure 2.
A part of the Geological Map of the Wadi Hali Quadrangle, sheet 18E, Kingdom of Saudi Arabia [29]. The map shows offshore islands, coral reefs and surficial deposits that include two fans that were formed by ancient streams now wadis (dry riverbeds). The region is covered with Holocene-Recent flood plain deposits, sabkhas (salt plains), scattered patches of mangrove swamps and eolian sand fields. Sample locations (1 through 6) are as follows. Mangrove Swamp Environment: Locations 1, 2A and 2B; Paleochannel Section: Location 3; Sabia Island Coral Reef Environment: Location 4; Algal Mat Environment: Location 5; Intertidal Environment: Location 6.
Figure 2.
A part of the Geological Map of the Wadi Hali Quadrangle, sheet 18E, Kingdom of Saudi Arabia [29]. The map shows offshore islands, coral reefs and surficial deposits that include two fans that were formed by ancient streams now wadis (dry riverbeds). The region is covered with Holocene-Recent flood plain deposits, sabkhas (salt plains), scattered patches of mangrove swamps and eolian sand fields. Sample locations (1 through 6) are as follows. Mangrove Swamp Environment: Locations 1, 2A and 2B; Paleochannel Section: Location 3; Sabia Island Coral Reef Environment: Location 4; Algal Mat Environment: Location 5; Intertidal Environment: Location 6.
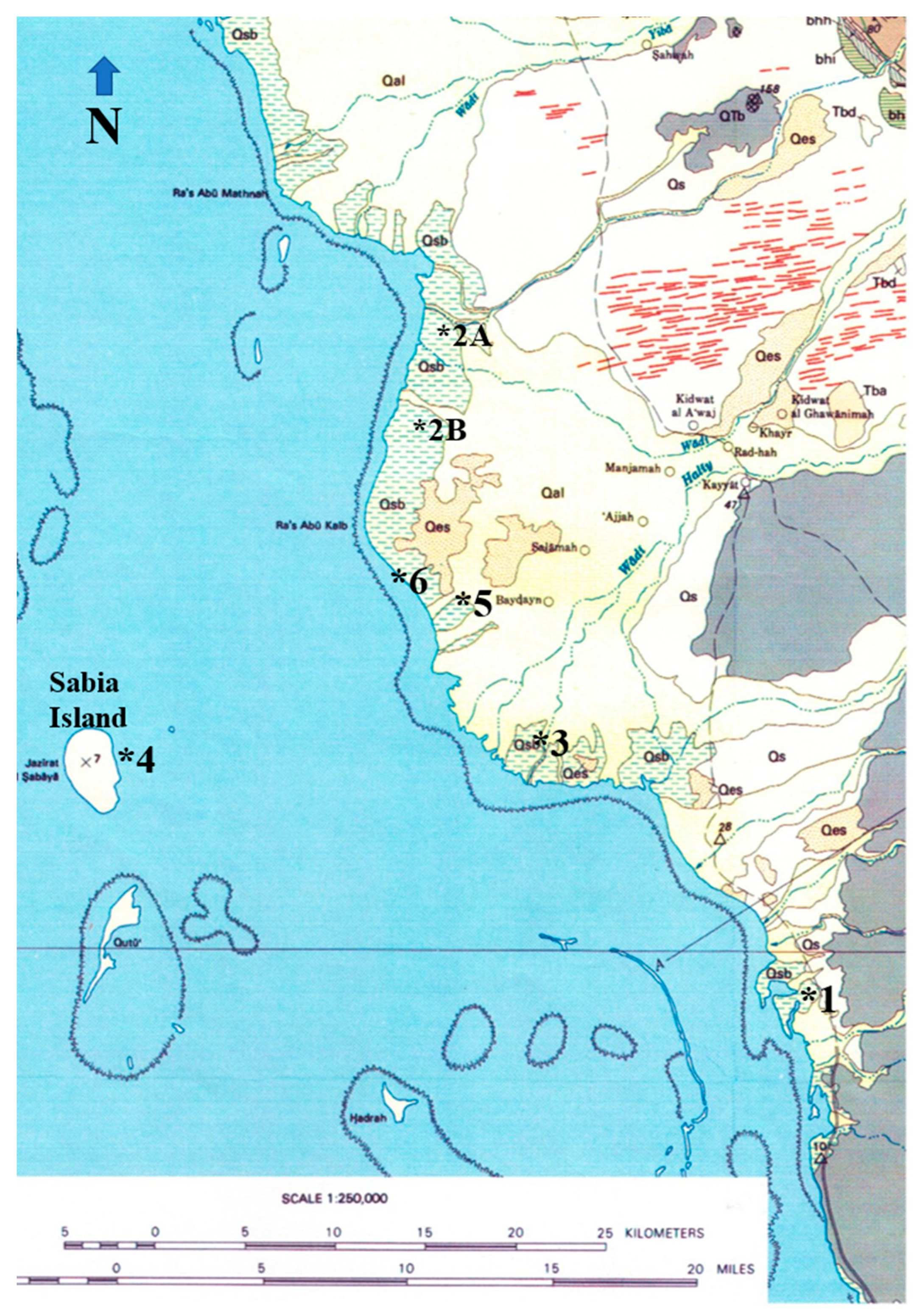
Figure 3.
A. Coastal and terrestrial environments around the area of study. Satellite image of the area showing marine and coastal environments. Mangroves are shown in green (modified after Khan et al. 2010). B. close-up of the area where samples (L1 through L7) were collected showing sample localities.
Figure 3.
A. Coastal and terrestrial environments around the area of study. Satellite image of the area showing marine and coastal environments. Mangroves are shown in green (modified after Khan et al. 2010). B. close-up of the area where samples (L1 through L7) were collected showing sample localities.
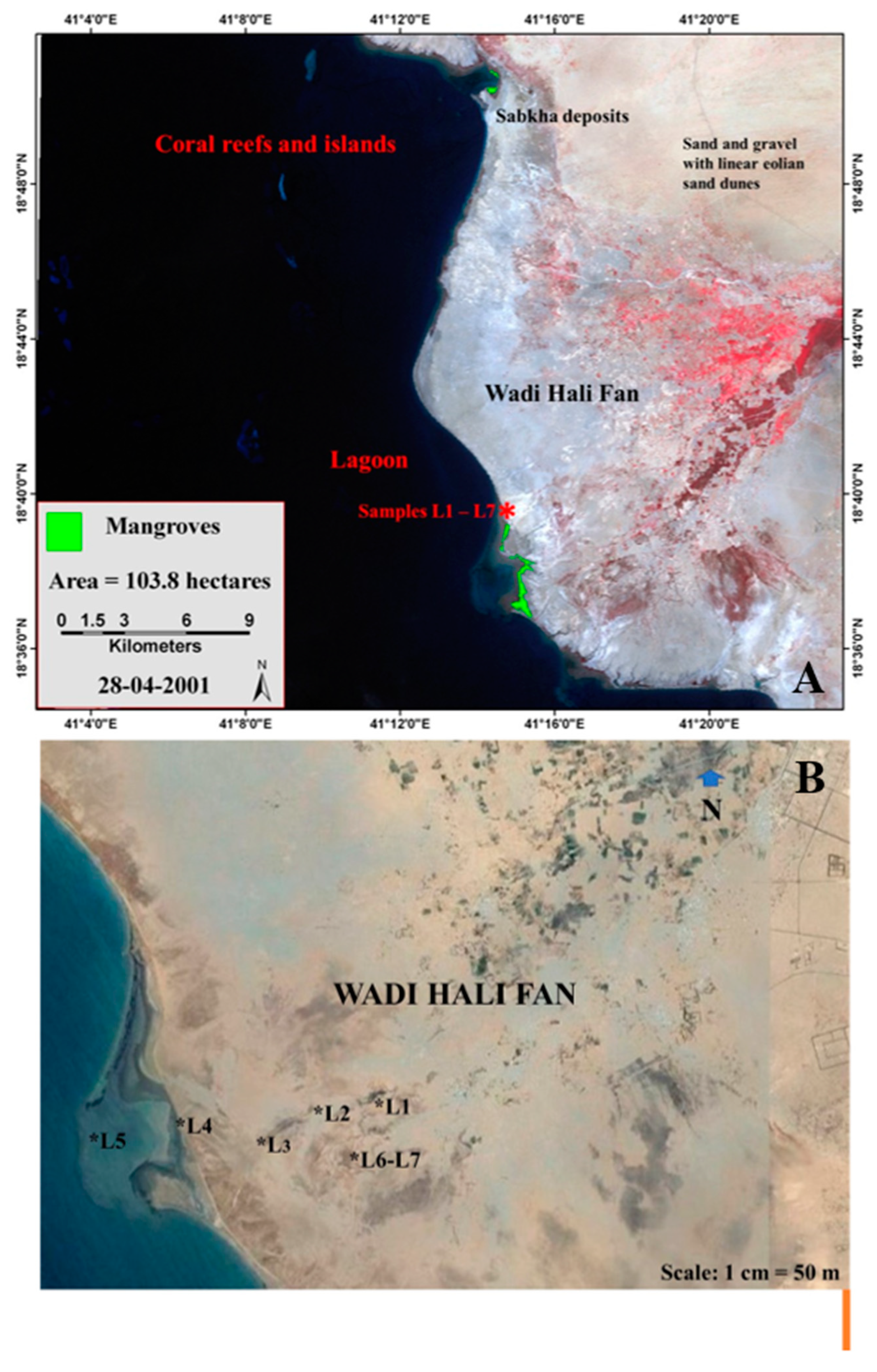
Figure 4.
Various localities of samples collected for this study. A. Two clay samples (M1 and M2) from the mangrove swamp (Figure 2, Location 1) B. Two clay samples (M3 and M4) from the mangrove swamp (Figure 2, Location 2A) C. Three clay samples (M5, M6 and M7) from the mangrove swamp (Figure 2, Location 2B) D. One sample (AM1) from algal mat (water depth 50 cm or less; Figure 2, Location 5) E. One sample (AM2) from dried part of the algal mat (Figure 2, Location 5).
Figure 4.
Various localities of samples collected for this study. A. Two clay samples (M1 and M2) from the mangrove swamp (Figure 2, Location 1) B. Two clay samples (M3 and M4) from the mangrove swamp (Figure 2, Location 2A) C. Three clay samples (M5, M6 and M7) from the mangrove swamp (Figure 2, Location 2B) D. One sample (AM1) from algal mat (water depth 50 cm or less; Figure 2, Location 5) E. One sample (AM2) from dried part of the algal mat (Figure 2, Location 5).

Figure 5.
The locations of samples collected for this study. A. Two fine sand samples (SI1 and SI2) from offshore Sabia Island (location 4) B. Seven clay and fine sand samples (WH1 through WH7) from the paleochannel section (location 3).
Figure 5.
The locations of samples collected for this study. A. Two fine sand samples (SI1 and SI2) from offshore Sabia Island (location 4) B. Seven clay and fine sand samples (WH1 through WH7) from the paleochannel section (location 3).

Figure 6.
Percentage distribution of palynomorph groups. A. Wadi Hali paleochannel assemblage (crustacean antennae excluded), B. Mangrove swamp assemblage (crustacean antennae excluded).
Figure 6.
Percentage distribution of palynomorph groups. A. Wadi Hali paleochannel assemblage (crustacean antennae excluded), B. Mangrove swamp assemblage (crustacean antennae excluded).
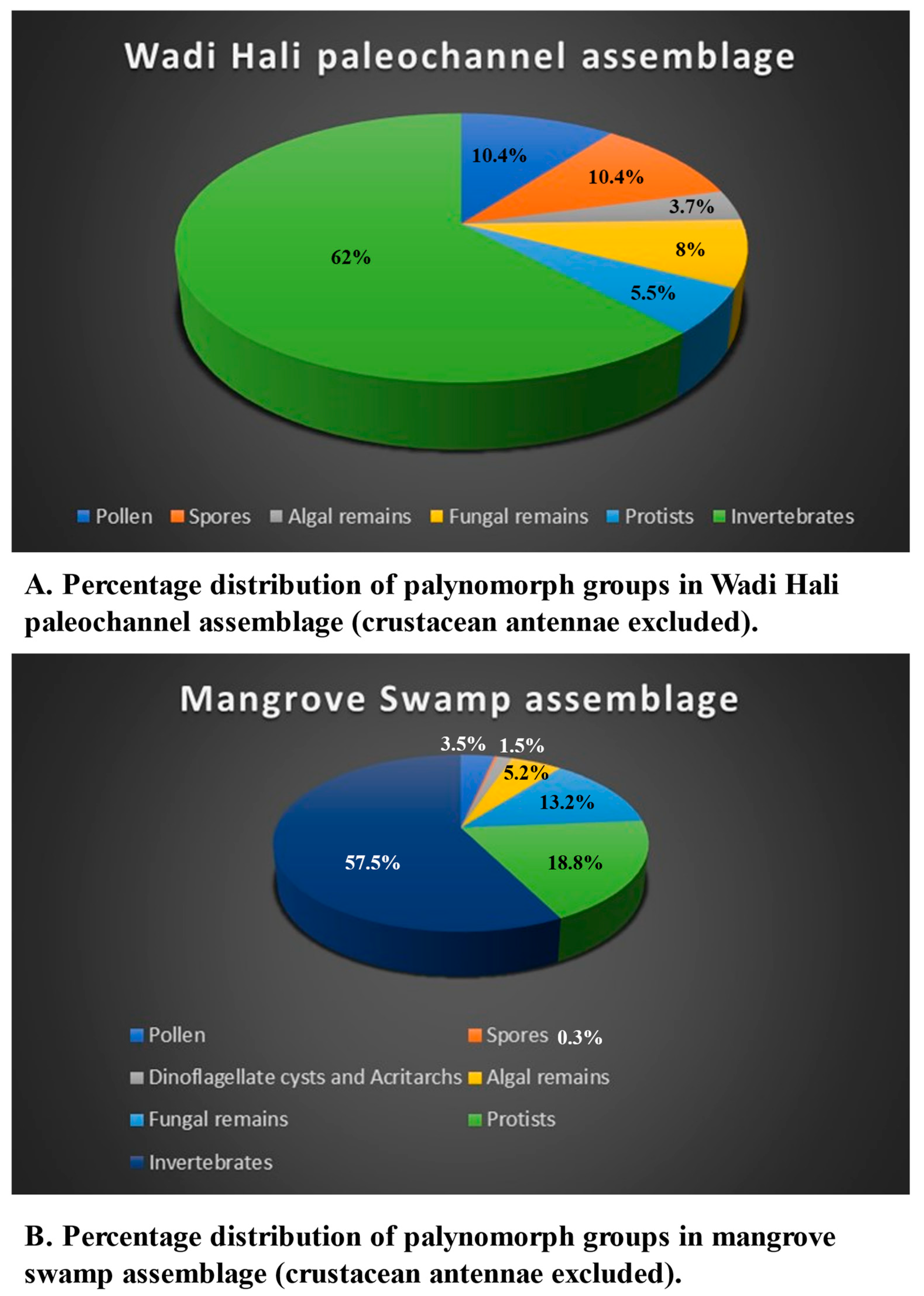
Figure 7.
Percentage distribution of palynomorph groups. A. Algal Mat assemblage, B. Sabia Island coral reef assemblage.
Figure 7.
Percentage distribution of palynomorph groups. A. Algal Mat assemblage, B. Sabia Island coral reef assemblage.
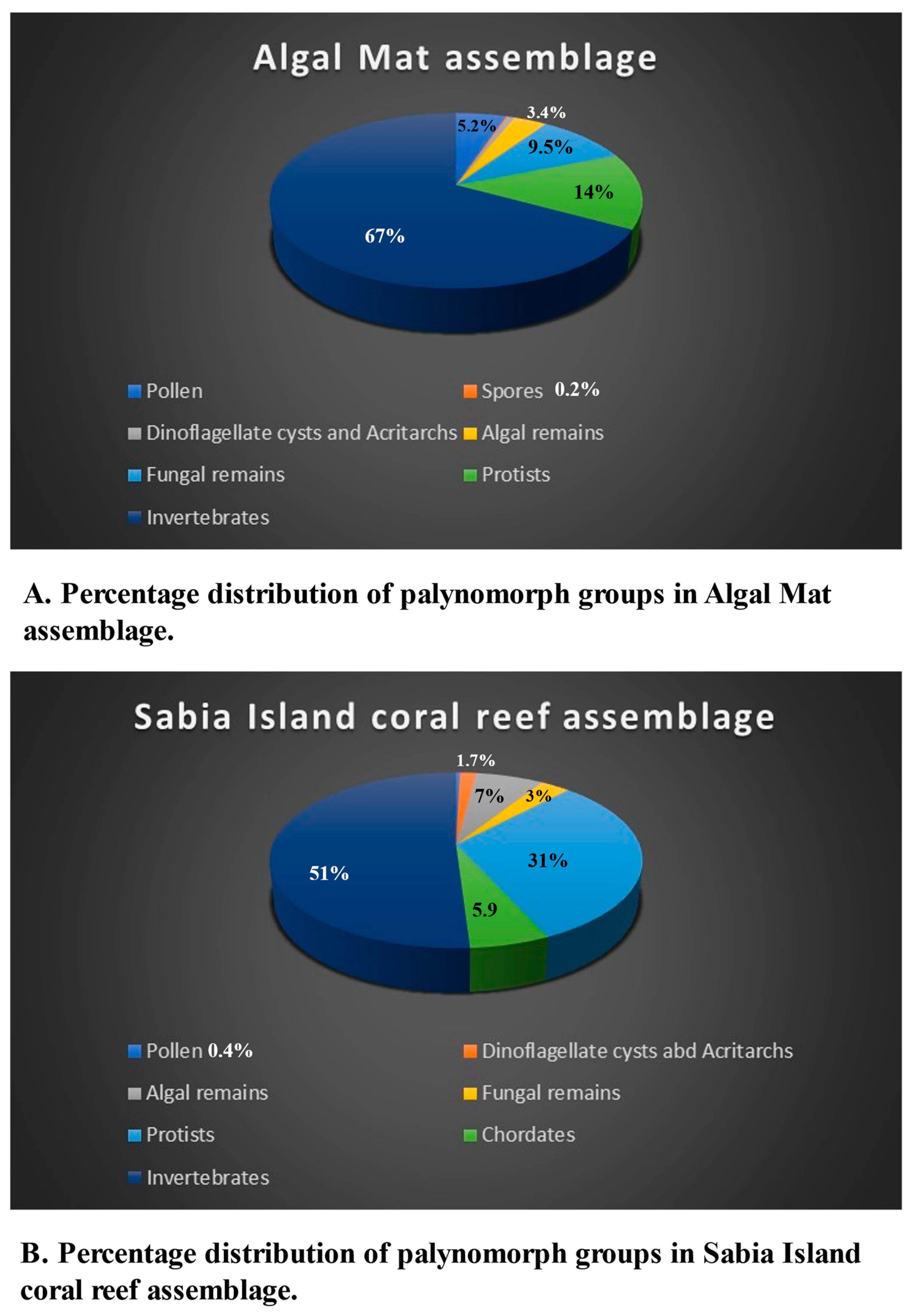
Figure 8.
Percentage representation of palynomorph groups in various environments. A. Wadi Hali paleochannel, B. Mangrove Swamp, C. Algal Mat, D. Sabia Island coral reef.
Figure 8.
Percentage representation of palynomorph groups in various environments. A. Wadi Hali paleochannel, B. Mangrove Swamp, C. Algal Mat, D. Sabia Island coral reef.
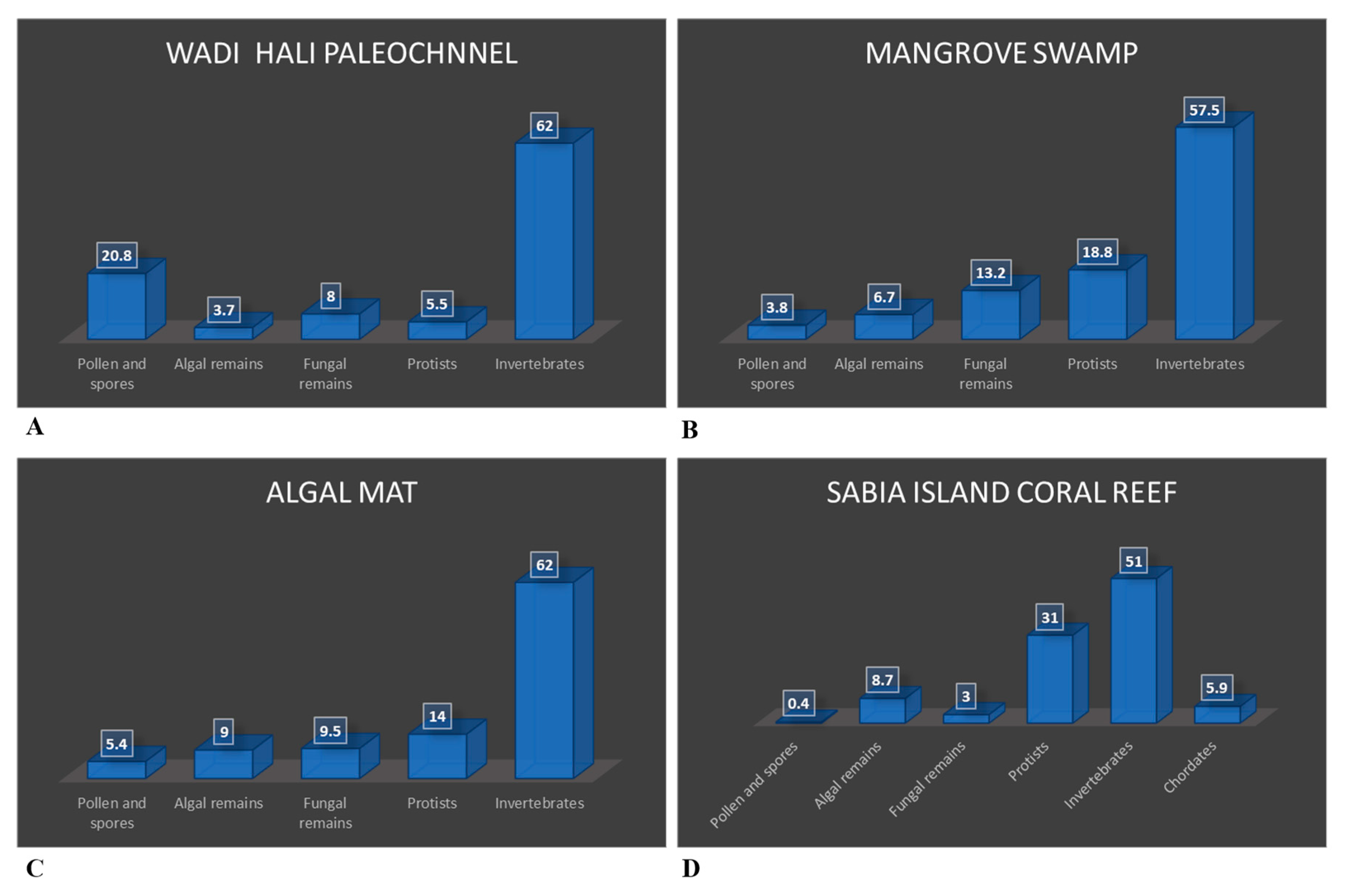
Figure 9.
Percentage distribution of palynomorph groups in the intertidal environments. Data based on seven samples [16].
Figure 9.
Percentage distribution of palynomorph groups in the intertidal environments. Data based on seven samples [16].
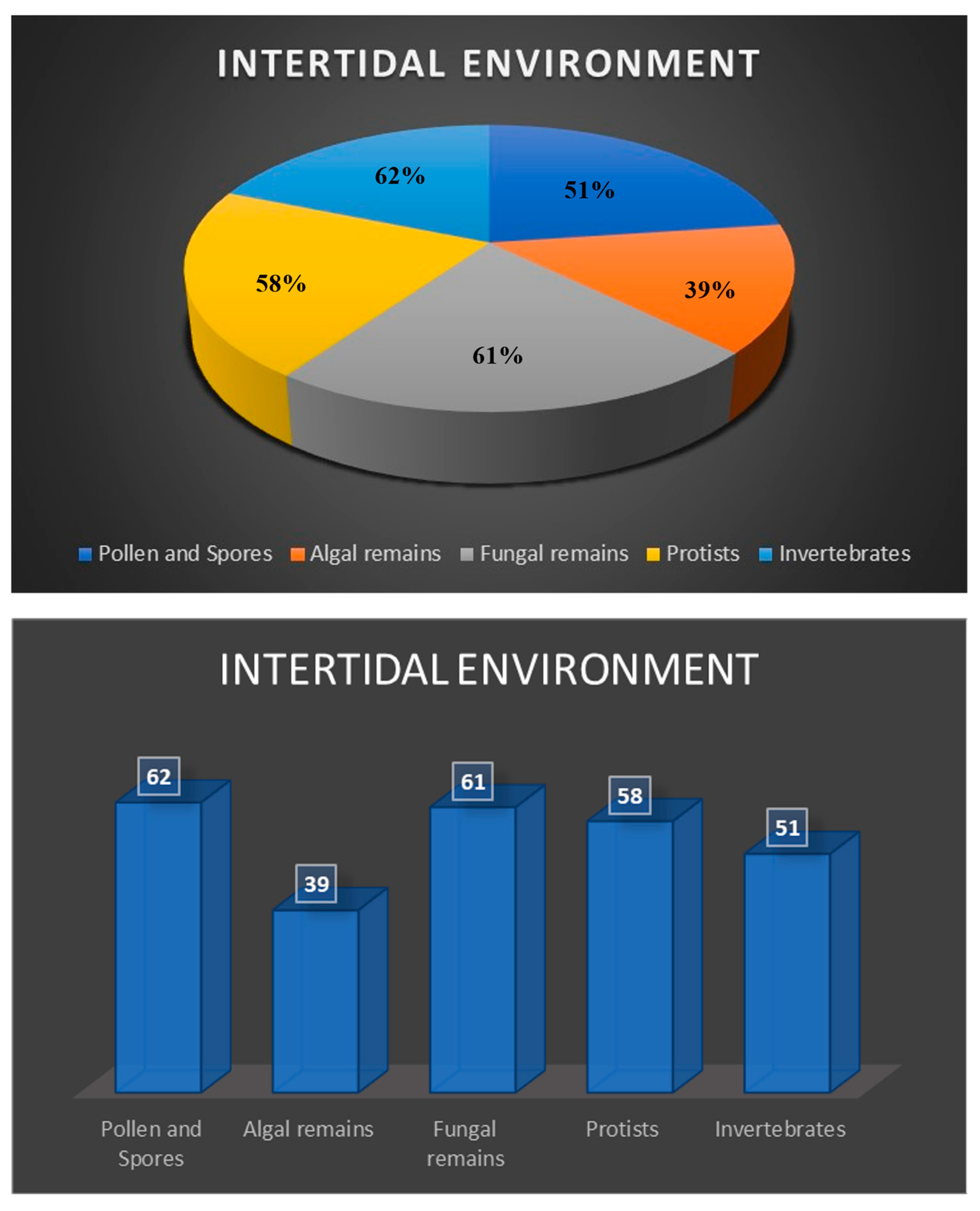

| Angiosperm Pollen | Mangrove Swamp | Palaeochannel (Middle Holocene) | Algal Mat | Sabia Is. | Intertidal | ||||||||||||||||||||
| M1 | M2 | M3 | M4 | M5 | M6 | M7 | WH1 | WH2 | WH3 | WH4 | WH5 | WH6 | WH7 | AM1 | AM2 | SI1 | SI2 | L1 | L2 | L3 | L4 | L5 | L6 | L7 | |
| Acacia sp. | 3 | 1 | 1 | 2 | 1 | 1 | |||||||||||||||||||
| Artemesia sp. | 1 | 1 | 1 | 1 | |||||||||||||||||||||
| Avicecennia marina | 2 | 1 | 1 | 1 | |||||||||||||||||||||
| cf Casurina sp. | 1 | ||||||||||||||||||||||||
| Calligonum sp. | 1 | ||||||||||||||||||||||||
| Chenopodiaceae/Amaranthaceae | 2 | 6 | |||||||||||||||||||||||
| Cyperaceae | 1 | 3 | 1 | 1 | 1 | ||||||||||||||||||||
| Euphorbia sp. | 1 | 1 | 1 | ||||||||||||||||||||||
| Neocouperipollis sp. | 1 | 2 | 2 | ||||||||||||||||||||||
| Palmaepollenites sp. | 2 | 4 | 1 | 1 | 2 | 3 | |||||||||||||||||||
| Palmidites sp. | 2 | 2 | 1 | 1 | 4 | 1 | 1 | 2 | 1 | ||||||||||||||||
| Potamogeton sp. | 1 | 1 | 2 | ||||||||||||||||||||||
| Proxapertites sp. | 1 | ||||||||||||||||||||||||
| Rannunculaceae | 1 | ||||||||||||||||||||||||
| Rhizophora mucronata | 1 | 1 | 1 | 1 | |||||||||||||||||||||
| Spinizonocolpites sp. (Nypa palm) | 3 | 3 | 2 | 4 | 1 | 2 | 2 | 5 | 1 | 2 | |||||||||||||||
| Trichotomosulcites sp. | 1 | 1 | |||||||||||||||||||||||
| Tricolpate psilate | 1 | ||||||||||||||||||||||||
| Tricolpate scabrate | 1 | 1 | 1 | ||||||||||||||||||||||
| Verrualetes sp. | 1 | ||||||||||||||||||||||||
| Zea mays (cultivated) | 1 | ||||||||||||||||||||||||
| Gymnosperm Pollen | |||||||||||||||||||||||||
| Inaperturate pollen | 3 | 5 | 2 | 3 | 3 | 2 | 6 | ||||||||||||||||||
| Juniperous pollen | 3 | 1 | 1 | ||||||||||||||||||||||
| Pinus sp. | 2 | ||||||||||||||||||||||||
| Podocarpus sp. | 1 | 1 | |||||||||||||||||||||||
| Spores | |||||||||||||||||||||||||
| Hypnaceae spore (Moss) | 1 | 1 | |||||||||||||||||||||||
| Laevigatosporites sp. | 1 | 1 | 2 | 1 | |||||||||||||||||||||
| Lycopodiumsporites sp. | 1 | 1 | 3 | 1 | |||||||||||||||||||||
| Polypodiisporites sp. | 6 | 1 | 2 | ||||||||||||||||||||||
| Triplanosporites sp. | 1 | ||||||||||||||||||||||||
| Verrucosiporites sp. | 2 | 1 | 2 | 1 | |||||||||||||||||||||
| Dinoflagellate Cysts | Mangrove Swamp | Palaeochannel (Middle Holocene) | Algal Mat | Sabia Is. | Intertidal | ||||||||||||||||||||
| M1 | M2 | M3 | M4 | M5 | M6 | M7 | WH1 | WH2 | WH3 | WH4 | WH5 | WH6 | WH7 | AM1 | AM2 | SI1 | SI2 | L1 | L2 | L3 | L4 | L5 | L6 | L7 | |
| cf. Brigantedinium simplex | 1 | ||||||||||||||||||||||||
| ?Operculodinium sp. | 1 | ||||||||||||||||||||||||
| Peridinium sp. | 1 | ||||||||||||||||||||||||
| Spiny round brown cyst | 1 | ||||||||||||||||||||||||
| Dinoflagellate cyst type A | 1 | 1 | |||||||||||||||||||||||
| Dinoflagellate cyst type B | 1 | ||||||||||||||||||||||||
| Dinoflagellate cyst type C | 1 | ||||||||||||||||||||||||
| Dinoflagellate cyst type D | 2 | 5 | 1 | 1 | 2 | 2 | 1 | ||||||||||||||||||
| Dinoflagellate cyst type E | 1 | ||||||||||||||||||||||||
| Dinoflagellate cyst type F | 1 | ||||||||||||||||||||||||
| Dinoflagellate cyst type G | 1 | ||||||||||||||||||||||||
| Dinoflagellate cyst type H | 1 | ||||||||||||||||||||||||
| Dinoflagellate cyst type I | 1 | 1 | |||||||||||||||||||||||
| Dinoflagellate cyst type J | 1 | ||||||||||||||||||||||||
| Dinoflagellate cyst type K | 1 | ||||||||||||||||||||||||
| Dinoflagellate cyst type L | 1 | ||||||||||||||||||||||||
| Dinoflagellate cyst type M | 1 | ||||||||||||||||||||||||
| Dinoflagellate cyst type N | 1 | ||||||||||||||||||||||||
| Acritarch (indeterminate) | 3 | 2 | |||||||||||||||||||||||
| Micrhystridium sp. | 1 | 1 | |||||||||||||||||||||||
| Algal remains | |||||||||||||||||||||||||
| Algal bodies (unidentified) | 4 | 4 | 7 | 7 | 10 | 19 | 11 | 1 | 2 | 1 | 1 | 1 | |||||||||||||
| Algal cells type A | 6 | 7 | 6 | 13 | 1 | 3 | 1 | ||||||||||||||||||
| Algal cells type B | 1 | 1 | 1 | ||||||||||||||||||||||
| Algal cyst (unidentified) | 1 | 1 | 2 | 3 | |||||||||||||||||||||
| Botryococcus sp. | 1 | ||||||||||||||||||||||||
| Colonial alga/Cyanobacteria | 1 | 2 | |||||||||||||||||||||||
| Concentricystis sp. | 2 | 1 | 1 | 3 | |||||||||||||||||||||
| Marine diatom (broken pieces) | 7 | 24 | |||||||||||||||||||||||
| Gloeotrichia sp. | 1 | 1 | |||||||||||||||||||||||
| Lecaniella sp. | 1 | 1 | 1 | 1 | |||||||||||||||||||||
| Rivularia sp. | 1 | ||||||||||||||||||||||||
| Zygospore Spirogyra | 1 | 1 | 1 | ||||||||||||||||||||||
| Zygospore Debraya glyptosperma | 1 | ||||||||||||||||||||||||
| Fungal Taxa | Intertidal | Palaeochannel | Mangrove Swamp | Algal Mat | Coral Reef |
| Alleppeysporonites elsikii | F | ||||
| Alleppeysporonites sp. | F | ||||
| Alternaria sp. 1 | F | ||||
| Alternaria sp. 2 | F | ||||
| Alternaria sp. cf. A. bellus | R | ||||
| Asterinites tellezii | F | ||||
| Callimothallus pertusus | F | ||||
| Cercospora sp. | R | R | |||
| Dicellaesporites bisariae | F | ||||
| Dicellaesporites foveolatus | F | ||||
| Dicellaesporites plicatus | F | ||||
| Dicellaesporites verrucatus | F | ||||
| cf. Endochaetophora sp. | R | ||||
| Exesisporites chandrae | C | ||||
| Exesisporites sp. | R | ||||
| Hypoxylonites sp. 1 | R | ||||
| Hypoxylonites sp. 2 | R | ||||
| Hypoxynonites sp. 3 | R | ||||
| Hypoxynonites sp. 4 | R | F | |||
| Inapertisporites kharei | F | ||||
| Inapertisporites foveolatus | F | ||||
| Inapertisporites kedvesii | F | ||||
| Inapertisporites microverrucatus | C | F | |||
| Inapertisporites sp. 1 | R | ||||
| ?Inapertisporites sp. | R | ||||
| Inaperturate elongated fungal spores | C | ||||
| Laevitubulus laxus | F | R | |||
| Laevitubulus sp. | R | R | F | ||
| Monoporisporites jansoniusii | F | ||||
| Monoporisporites pattersonii | F | ||||
| Monoporisporites valdiyae | F | ||||
| Monoporisporites rotundus | F | ||||
| Monoporisporites sp. 1 | R | ||||
| Monoporisporites sp. 2 | R | ||||
| Palaeomycites robustus | F | R | |||
| Palaeosclerotium sp. | R | ||||
| Papulosporonites multicellatus | F | ||||
| Pluricellaesporites mishrae | F | ||||
| Polyadosporites sp. | R | ||||
| Scolecosporites sp. | R | ||||
| Spinospororonites sp. | R | ||||
| Spirotremesporonites sp. | R | ||||
| Staphlosporites neyveliensis | R | ||||
| Tetraploa sp. 1 | F | ||||
| Tetraploa sp. 2 | R | ||||
| Trichothyrites amorphus | R | ||||
| Trichothyrites setifer | F | ||||
| Trichothyrites sp. | R | ||||
| Cluster of fungal hyphae | C | F | |||
| Fungal Remain Type 1 | R | ||||
| Fungal Remain Type 2 | R | ||||
| Fungal Remain Type 3 | R | ||||
| Fungal Remain Type 1 | R | ||||
| Sporangiun with spores | R |
| Microforaminifera | Mangrove Swamp | Palaeochannel (Middle Holocene) | Algal Mat | Sabia Is. | Intertidal | ||||||||||||||||||||
| M1 | M2 | M3 | M4 | M5 | M6 | M7 | WH1 | WH2 | WH3 | WH4 | WH5 | WH6 | WH7 | AM1 | AM2 | SI1 | SI2 | L1 | L2 | L3 | L4 | L5 | L6 | L7 | |
| Microforaminiferal test linings type 1 (Kumar 2020) | 4 | 2 | 2 | 2 | 1 | 7 | 1 | ||||||||||||||||||
| Microforaminiferal test linings type 2 (Kumar 2020) | 7 | 1 | |||||||||||||||||||||||
| Microforaminiferal test linings type 3 (Kumar 2020) | 4 | 1 | 5 | 14 | 1 | ||||||||||||||||||||
| Microforaminiferal test linings type 4 (Kumar 2020) | 1 | 3 | 6 | 4 | 1 | ||||||||||||||||||||
| Microforaminiferal test linings type 5 (Kumar 2020) | 1 | ||||||||||||||||||||||||
| Microforaminiferal test linings type 6 (Kumar 2020) | 2 | 14 | 1 | 1 | |||||||||||||||||||||
| Microforaminiferal test linings type 7 (Kumar 2020) | 3 | 2 | 2 | 1 | 1 | 3 | 9 | 5 | 1 | ||||||||||||||||
| Microforaminiferal test linings type 8 (Kumar 2021) | 1 | 10 | 1 | ||||||||||||||||||||||
| Microforaminiferal test linings type 9 (Kumar 2021) | 2 | 3 | 2 | 1 | 2 | ||||||||||||||||||||
| Microforaminiferal test linings type 10 (Kumar 2021) | 2 | 5 | |||||||||||||||||||||||
| Microforaminiferal test linings type 11 (Kumar 2021) | 1 | ||||||||||||||||||||||||
| Microforaminiferal test linings type 12 (Kumar 2021) | 44 | ||||||||||||||||||||||||
| Microforaminiferal test linings type 13 (Kumar 2021) | 1 | 3 | |||||||||||||||||||||||
| Microforaminiferal test linings type 14 (Kumar 2021) | 5 | ||||||||||||||||||||||||
| Microforaminiferal test linings type 15 (Kumar 2021) | 3 | 2 | |||||||||||||||||||||||
| Microforaminiferal test linings type 16 (Kumar 2021) | 11 | ||||||||||||||||||||||||
| Thecamoebians | |||||||||||||||||||||||||
| Arcella sp. | 1 | ||||||||||||||||||||||||
| Centropyxis aculeata | 3 | 1 | 1 | 1 | 1 | 4 | 1 | 2 | 2 | ||||||||||||||||
| Centropyxis sp. | 3 | 2 | |||||||||||||||||||||||
| Cyclopyxis sp. | 2 | ||||||||||||||||||||||||
| Difflugia sp. | 1 | 1 | |||||||||||||||||||||||
| Euglepha sp. | 1 | 1 | |||||||||||||||||||||||
| Geopyxella sylvicola | 1 | 1 | 1 | ||||||||||||||||||||||
| Thecamoebian type A (Kumar 2021) | 3 | ||||||||||||||||||||||||
| Thecamoebian type B (Kumar 2021) | 1 | 2 | |||||||||||||||||||||||
| Thecamoebian type F (Kumar 2021) | 2 | ||||||||||||||||||||||||
| ?Thecamoebians | 2 | ||||||||||||||||||||||||
| Tintinnomorphs | |||||||||||||||||||||||||
| Tintinnomorph type 9 (Kumar 2020) | 1 | ||||||||||||||||||||||||
| Tintinnomorph type 11 (Kumar 2020) | 1 | 1 | |||||||||||||||||||||||
| Tintinnomorph type 12 (Kumar 2020) | 1 | ||||||||||||||||||||||||
| Tintinnomorph type 13 (Kumar 2020) | 1 | ||||||||||||||||||||||||
| Tintinnomorph type 14 (Kumar 2020) | 1 | ||||||||||||||||||||||||
| Tintinnomorph type 16 (Kumar 2020) | 1 | 1 | |||||||||||||||||||||||
| Form taxa | |||||||||||||||||||||||||
| Katora arabica (Kumar 2023) | 1 | 1 | 4 | 2 | 18 | 10 | 14 | 1 | 4 | 7 | 12 | 30 | 1 | 7 | 5 | 2 | 3 | 1 | |||||||
| Katora oblonga (Kumar 2023) | 2 | 3 | 1 | 1 | 10 | 2 | 13 | 3 | 22 | 6 | 4 | 3 | 1 | ||||||||||||
| Katora twinmorpha (Kumar 2023) | 1 | 2 | 7 | 1 | |||||||||||||||||||||
| Katora elongata (Kumar 2023) | 1 | 5 | 1 | 5 | 6 | 1 | 2 | 1 | |||||||||||||||||
| Mangrovia hallii (Kumar 2023) | 3 | 3 | 1 | 2 | |||||||||||||||||||||
| Mangrovia redseaensis (Kumar 2023) | 2 | 4 | 7 | 38 | 29 | 7 | 21 | ||||||||||||||||||
| Sample Number | Total Number of Palynomorphs | Number of Foraminiferal Palynomorphs | Percentage of Foraminiferal Palynomorphs | |
| Intertidal | ||||
| L1 | 61 | 0 | 0 | |
| L2 | 55 | 0 | 0 | |
| L3 | 74 | 5 | 6.75 | |
| L4 | 19 | 2 | 10.52 | |
| L5 | 52 | 0 | 0 | |
| L6 | 66 | 1 | 1.51 | |
| L7 | 11 | 0 | 0 | |
| Total | 338 | 8 | Average = 2.68 | |
| Mangrove | ||||
| M1 | 99 | 10 | 10.1 | |
| M2 | 84 | 2 | 2.38 | |
| M3 | 67 | 3 | 4.47 | |
| M4 | 26 | 1 | 3.84 | |
| M5 | 107 | 28 | 26.16 | |
| M6 | 132 | 6 | 4.54 | |
| M7 | 57 | 5 | 8.77 | |
| Total | 572 | 55 | Average = 8.60 | |
| Algal Mat | ||||
| AM 1 | 60 | 0 | 0 | |
| AM 2 | 58 | 0 | 0 | |
| Total | 118 | 0 | 0 | |
| Coral Reef (Sabia Island) | ||||
| SI 1 | 66 | 14 | 21.21 | |
| SI 2 | 141 | 59 | 41.84 | |
| Total | 207 | 73 | Average = 31.52 | |
| Invertebrates | Mangrove Swamp | Palaeochannel (Middle Holocene) | Algal Mat | Sabia Is. | Intertidal | ||||||||||||||||||||
| M1 | M2 | M3 | M4 | M5 | M6 | M7 | WH1 | WH2 | WH3 | WH4 | WH5 | WH6 | WH7 | AM1 | AM2 | SI1 | SI2 | L1 | L2 | L3 | L4 | L5 | L6 | L7 | |
| Crustacean remains (unidentified) | 1 | 3 | 1 | 3 | 7 | ||||||||||||||||||||
| Ostracod jaw | 2 | 1 | 1 | ||||||||||||||||||||||
| cf. Leptocythere carapace (ostracod) | 3 | 2 | 7 | 1 | |||||||||||||||||||||
| Scolecodont | 2 | 3 | 5 | 4 | 2 | ||||||||||||||||||||
| Crustaceant wing | 1 | ||||||||||||||||||||||||
| Crustacean exoskeleton | 65 | 45 | 42 | 49 | 35 | 51 | 96 | 6 | 5 | 13 | 10 | 9 | 13 | 9 | 44 | 63 | 6 | 33 | 1 | 1 | 3 | 1 | |||
| Copepod fragment | 1 | 1 | |||||||||||||||||||||||
| Crustacean antennae | 5 | 29 | >500* | 33 | 8 | 12 | 11 | >500* | >500* | >500* | 119 | 165 | 132 | 141 | 52 | 57 | 83 | 88 | 2 | ||||||
| Crustacean hair | 9 | 7 | 8 | 12 | 8 | 13 | 5 | 2 | 3 | 5 | 12 | 5 | 4 | 3 | 4 | 6 | 1 | 18 | |||||||
| Crustacean eggs | 16 | 10 | 21 | 4 | 9 | 30 | 55 | 9 | 26 | 71 | 53 | ||||||||||||||
| Turbellarian worm | 3 | 2 | 1 | 1 | 2 | 1 | 1 | 1 | 2 | 2 | |||||||||||||||
| Turbellarian egg | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||
| Annelid palynomorph | 4 | 7 | 1 | 1 | 3 | ||||||||||||||||||||
| Annelid jaw | 1 | 1 | |||||||||||||||||||||||
| Worms | 1 | 1 | 5 | 8 | 27 | 5 | 2 | 1 | |||||||||||||||||
| Annelid remains (unidentified) | 77 | 18 | 3 | 9 | 11 | 17 | 7 | 20 | 1 | 1 | |||||||||||||||
| Mineral microssils | |||||||||||||||||||||||||
| Phytoliths | Few | Few | Few | ||||||||||||||||||||||
| Sponge spicules | 1 | ||||||||||||||||||||||||
| Ascidian spicule type 1 | 4 | 6 | |||||||||||||||||||||||
| Ascidian spicule type 2 | 2 | 15 | |||||||||||||||||||||||
| Ascidian spicule type 3 | 2 | 3 | |||||||||||||||||||||||
| Ascidian spicule type 4 | 3 | ||||||||||||||||||||||||
| Ascidian spicule type 5 | 1 | 4 | |||||||||||||||||||||||
| Ascidian spicule type 6 | 1 | ||||||||||||||||||||||||
| TAXA | STUDIED ENVIRONMENTS OF SOUTHERN RED SEA COAST OF SAUDI ARABIA | ||||||||||||||||||||||||
| Intertidal | Wadi Hali Paleochannel | Mangrove Swamp | Algal Mat | Coral Reef | |||||||||||||||||||||
| L1 | L2 | L3 | L4 | L5 | L6 | L7 | WH1 | WH2 | WH3 | WH4 | WH5 | WH6 | WH7 | M1 | M2 | M3 | M4 | M5 | M6 | M7 | AM1 | AM2 | SI1 | SI2 | |
| Pollen and Spores | |||||||||||||||||||||||||
| Angiospermous Pollen | F | F | C | R | R | C | R | R | R | R | R | F | F | F | C | F | R | R | R | R | R | R | |||
| Gymnospermous Pollen | R | R | R | R | R | F | |||||||||||||||||||
| Spores | F | R | F | R | R | R | R | R | R | R | |||||||||||||||
| Dinoflagellate cysts and Algal remains | |||||||||||||||||||||||||
| Acritarchs | R | R | R | ||||||||||||||||||||||
| Algal Remains (Indet.) | C | C | F | F | C | F | R | R | R | R | F | F | F | C | C | F | F | F | C | ||||||
| Botryococcus sp. | R | R | R | ||||||||||||||||||||||
| Concentricystis sp. | R | R | R | R | R | ||||||||||||||||||||
| Cyanobacteria | R | R | R | ||||||||||||||||||||||
| Dinoflagellate cysts | R | R | R | R | R | R | R | R | R | R | R | R | R | ||||||||||||
| Gloeotrichia filament | R | ||||||||||||||||||||||||
| Lecaniella sp. | R | R | R | R | |||||||||||||||||||||
| Marine diatom (broken pieces) | F | C | |||||||||||||||||||||||
| Oospore of Oedogonium | R | R | |||||||||||||||||||||||
| Tasmanites type | R | R | |||||||||||||||||||||||
| Tetraporina sp. | R | ||||||||||||||||||||||||
| Fungal spores, hyphae and fruit bodies | |||||||||||||||||||||||||
| Basidiosporites sp. | R | R | |||||||||||||||||||||||
| Cladosporites sp. | R | R | |||||||||||||||||||||||
| Dictyosporites sp. | C | C | C | C | |||||||||||||||||||||
| Dicellaesporites sp. | R | R | R | ||||||||||||||||||||||
| ? Frasnacritetrus sp. | R | R | |||||||||||||||||||||||
| Glomus sp. | R | C | R | F | R | C | C | F | C | C | R | F | C | R | |||||||||||
| Inapertisporites sp. | R | R | R | R | R | R | R | C | R | R | |||||||||||||||
| Lacrimasporonites sp. | R | ||||||||||||||||||||||||
| Monosporisporites sp. | R | R | C | R | R | R | |||||||||||||||||||
| Multicellites sp. | R | ||||||||||||||||||||||||
| Pluricellaesporites sp. | R | ||||||||||||||||||||||||
| Spores +Hyphae | C | R | C | F | F | R | R | R | C | R | |||||||||||||||
| Fruit bodies | A | F | VC | F | F | R | R | F | F | R | R | F | F | R | R | ||||||||||
| Indeterminate Forms | F | C | VC | F | C | C | R | ||||||||||||||||||
| Protists, Invertebrates and Mineral microfossils | |||||||||||||||||||||||||
| Ascidian spicules | C | VC | |||||||||||||||||||||||
| Crustacean Eggs | R | VC | VC | C | C | C | R | R | R | R | |||||||||||||||
| cf. Leptocythere (Ostracode) | R | R | F | R | |||||||||||||||||||||
| Tintinnomorphs | R | R | |||||||||||||||||||||||
| Scolecodonts | R | F | F | R | R | R | F | R | |||||||||||||||||
| Microforaminifera | R | R | F | R | R | R | F | F | F | R | R | F | C | VC | A | ||||||||||
| Crustacean Palynomorphs | R | F | C | C | C | A | F | A | A | A | A | A | A | A | A | A | A | A | A | A | A | A | A | A | A |
| Annelid Palynomorphs | C | VC | VC | R | R | C | R | A | R | C | R | F | C | C | F | C | R | F | |||||||
| Sponge spicules | R | ||||||||||||||||||||||||
| Thecamoebians | R | R | R | R | R | R | R | R | R | C | C | R | R | R | R | ||||||||||
| Thecamoebians (Indet.) | R | F | R | F | C | C | F | C | R | R | |||||||||||||||
| Turbellarian worm | R | R | R | R | R | R | R | ||||||||||||||||||
| Other Remains | |||||||||||||||||||||||||
| Crustacean Antennae ? | A | A | C | A | A | VC | VC | A | A | A | A | A | A | A | A | A | A | A | A | A | A | A | A | ||
| Phytoliths | F | F | F | ||||||||||||||||||||||
| Charcoal | F | F | F | F | F | F | F | F | F | F | F | F | F | F | F | F | F | F | F | F | F | F | F | F | F |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



