
Submitted:
18 June 2024
Posted:
19 June 2024
You are already at the latest version
Abstract
Increased norepinephrine (NE) release from the sympathetic nerves during pregnancy in women or animal models, as occurs during stress, modifies the fetal environment and increases flow of NE to the fetus, programing the progeny and affecting its cardiovascular and metabolic function in adulthood. In the present work, gestating rats were exposed to stress during gestation and the female progeny was analyzed for heart and metabolic function. Pregnant Sprague–Dawley rats were exposed to cold stress (4 °C/3 h/day); rats’ female progeny were euthanized at 4, 20 and 60 days old and their hearts evaluated to determine the β-adrenergic receptor (βAR) mRNA, abundance and affinity to agonists and NE concentration. The in vivo response to glucose overload and the arterial pressure response to isoproterenol (ISO, 125 µg/kg weight/day/10 days) were monitored. Results: At 20 and 60 days old, stressed female progeny presented an increase in ventricular weight and no changes in the cardiac NE as compared with their age-matched controls. The β1AR mRNA in control rats decreased between 4 and 20 days of age, but stressed rats maintained 50% higher levels of mRNA as compared with their age-matched controls. This difference was also found in the protein abundance of β1AR. No differences were found at 60 days old. The β2AR mRNA levels were higher than in controls at both 4 and 20 days old, but not expressed in the amount of protein. An increase in the ratio of β1/β2 receptors was found at 20 days old but not at 60 days old. The displacement of 3H-dihydroalprenolol (DHA) from a membrane fraction with propranolol (β antagonist) showed decreased affinity (at 60 days old) but no changes in the βAR number. In vivo exposure to ISO induced a β-adrenergic overload and provoked death in 40% of stressed females by day 3 of ISO treatment. Regarding metabolic function, we found that stressed rats presented insulin resistance when adults. These data suggest permanent changes to the heart’s adrenergic response and metabolic function after rat progeny were stressed in the uterus.
Keywords:
gestational programming
; cardiovascular
; norepinephrine
; beta receptors
1. Introduction
During gestation, all organisms adapt to the local conditions existing in the uterus. In the placental environment, they are exposed to different stimuli that could be dangerous if they are chronic. If there are no changes in their genetic load; this adaptation to the surrounding environment will produce changes in a single generation, which is much faster than in genetic adaptation [1,2,3]. Increasing evidence supports the notion that these changes in the offspring could have transgenerational effects. In this regard, if chronic stress occurs during the gestation period, this stress modifies the placental environment to produce changes in the embryo, affecting the development of the organs, the physiology and the metabolism of the organism (fetal programming) [4,5]. Various studies in animals have shown the effects of fetal programming. In Wistar rats, maternal protein restriction during gestation and lactation resulted in increased levels of cardiac fibrosis in the adult offspring [6], lower concentrations of cardiac β1-adrenergic receptors and a reduced baseline heart rate [7]. Similar effects were found in young animals that were stressed during the gestation period. For example, in Sprague–Dawley rats, adult pups, which had suffered restraint stress during gestation, had a more significant increase in blood pressure and heart rate due to stress [8]. Sympathetic stress stimulates norepinephrine (NE) release from the sympathetic nerves. During pregnancy, it modifies the fetal environment, increasing the access of NE to the fetus through the placental NE transporter and probably affecting adult physiological functions [9]. The effects of chronic stress on people’s health and, in particular, on embryos are only now beginning to be analyzed, making it important to understand the effects of fetal programming. Regarding the stress response, the activation of the sympathetic nervous system (SNS) is observed in “fight or flight” situations, as would be the case for a cold situation without hypothermia [1,10,11]. Along with increased stress levels within the population, the frequency and mortality associated with cardiovascular diseases in humans have increased, with a direct association between stress and these diseases. Currently, cardiovascular diseases are considered one of the leading causes of death in the adult population, and their prevalence is expected to continue to increase [12].
Our laboratory works with an intermittent chronic cold stress protocol (4 °C/3 h/day) applied to the gestating rat throughout the gestation period. This stress model produces a selective sympathetic response and a decrease in the placental norepinephrine transporter (NET), which causes the overexposure to NE of the fetus during the gestation period [10,11,13]. This stress modifies the reproductive function of the adult female progeny with the induction of a polycystic ovarian phenotype (presence of ovarian cysts, hyperandrogenic condition [14], insulin resistance in the ovary [15] and modified reproduction of the progeny [16,17]). Interestingly, cold stress could have broad implications for the development of the offspring. Prenatal cold stress has been associated with an altered pattern of phosphorylation in ERK1/2 and the induction of BDNF expression in the hippocampus [18]. Additionally, prenatal cold stress increases the blood pressure in the offspring due to lower levels of dopamine D1 vascular receptor [19]. Based on these considerations regarding the effects of gestational stress in female progeny and the possible impact of a high noradrenergic state during gestation in cardiovascular disease on the progeny, we recently discussed the effect of gestational sympathetic stress in the cardiovascular system in male progeny [20]. We found a differential response in the males when subjected to a sympathetic overload with isoproterenol (ISO). The dose of ISO used in males (1mg/k,[21]) was much higher than that used in females (125 ug/k,[22]) and thus it could have led to gender differences in the response to the sympathetic overload. Thus, in the present work, we hypothesized that chronic gestational exposure to higher levels of NE, due to cold stress, would cause cardiovascular alterations in the female adult offspring. During this experimental protocol, both males and females were born; the results for the male progeny and some of the females born corresponded to already published reproductive studies [17,20]. In the present report, we describe the effects on the cardiovascular system of the remaining female progeny.
2. Materials and Methods
Primiparous female Sprague–Dawley rats weighing between 250 and 300 g, kept at 20 °C and with a 12:12 light–dark cycle, were used. Food and water were constantly and freely available. Mating procedure: Rats were checked for estrous cyclic activity to determine the proestrus day. On the night of the proestrus phase (when ovulation occurs), the rats were mated with males of proven fertility. The following morning, the rats were checked for the presence of a vaginal sperm plug. The presence of a vaginal plug was considered to indicate day 0 of pregnancy. Twenty pregnant rats were randomized into two groups of 8 control rats and 12 stressed rats. All the rats of the female progeny were euthanized at the age shown in Figure 1, depending on the experimental protocol used. The rats were euthanized by decapitation at the end of the experiments; the heart (ventricular tissue) and plasma of each rat were collected. Tissue specimens were weighed on a laboratory scale with a sensitivity level of up to 1 mg. Decapitation was performed according to the AVMA Guidelines for the Euthanasia of Animals (2020 Edition) [23]. This was performed by specialized personnel and approved by the Bioethics Committee of the Faculty of Chemistry and Pharmaceutical Sciences at the University of Chile (protocol number: CBE2016-13 to BP and CBE2017-05 to HL). All procedures presented in this paper complied with the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 8023, revised 1978) and national guidelines (CONICYT Guide for the Care and Use of Laboratory Animals). Figure 1 graphically shows the different experimental protocols used in the biological assays and the total number of rats.
The total number of rats used in the study is shown in each figure. An age-matched group of rats underwent the same procedure as stressed rats but, during pregnancy, the mothers were not exposed to gestational stress. An additional group of 60 days old was used to determinate the response to a Glucose tolerance test
2.1. Gestational Stress Induction
From day 1 of pregnancy, pregnant female rats from the experimental group were exposed daily to a temperature of 4 °C for 3 h a day throughout the gestation period. The control group was kept at a constant temperature of 20 °C throughout the gestation period. Once the rats were born, their sex was determined, and they were assigned to foster mothers up to 20 days old. The progeny were regulated to 12 rats each, with half of the pups being male and the other half being female (to maintain the average number of pups naturally born and ensure sufficient nursery). The male progeny were recently published [20] and studied at 20 and 60 days old. The female progeny were divided into two groups. One of them was included in another protocol to study fertility during a multigeneration study [17], and the remaining female progeny were used for the present work. The chronic cold stress protocol for pregnant rats was the same as that previously used for adult rats [9,16].
2.2. Adrenergic Overload Protocol
When female progeny reached 50 days old, we used two groups with 10 controls and 10 rats exposed to gestational stress during fetal growth. Both the control and stressed rats received a daily injection of isoproterenol (ISO, β-adrenergic agonist, Sigma Chemical Co., St. Louis, MO, USA) at a daily dose of 1 mg/kg of weight for 10 days. This dose produced heart hypertrophy after 10 days in control untreated rats [21]. ISO was dissolved in saline supplemented with ascorbic acid (Sigma Chemical Co., St Louis, MO, USA) at a concentration of 0.5 mM to prevent the oxidation of the ISO. Each day, a solution was freshly prepared, and the solution was sterilized by passage through a 0.22 µm filter. We also studied two groups (control and stressed) injected with saline as controls for the ISO group. These animals were euthanized at 60 days old after the 10th dose of ISO, before they resisted adrenergic overload with ISO exposure. For the in vivo recording of arterial pressure, we used the same protocol as previously described [20]. We implanted a microtransducer (TRM54P, from Millar, Inc., Houston, TX, USA) coupled to a PowerLab DAQ (ADInstruments, Sydney, Australia). Fifty-day-old stressed and control female rats were anesthetized with an i.m. dose of ketamine 60 mg/kg/xylazine 10 mg/kg solution under aseptic conditions. A transverse mid-lumbar incision of ~1.5 cm length was created in anesthetized rats, and the microdevice was implanted in the peritoneal cavity. A cannula connected to the abdominal aorta was sealed with instant glue. Rats were maintained in postoperative observation for 3 days before starting to record the pressure. The device was wirelessly charged through a platform that also received diastolic and systolic pressure signals. The signals were recorded through the program incorporated into the system.
2.3. Quantification of NE Levels
The quantification of the NE concentration in the heart and plasma was performed by homogenizing the tissues in 10 volumes of 0.2 M perchloric acid in ice. The NE levels were quantified using the Norepinephrine ELISA Kit—Research© (IMMUSMOL, Pessac, France).The minimal detectable value was 1.3 pg/ml, with a range of 0.2–32 ng/ml NE was extracted using a cis-diol-specific affinity gel, acylated and then derivatized enzymatically. The antigen was bound to the solid phase of the microtiter plate. The derivatized standards, controls and samples and the solid-phase-bound analyte competed for a fixed number of antiserum binding sites. The antibody bound to the solid phase was detected using an anti-rabbit IgG-peroxidase conjugate and 3,3′,5,5′-tetramethylbenzidine (TMB) as a substrate. The reaction was monitored at 450 nm. The results were expressed as the total amount of NE in ng per ovary. The sensitivity was 2 pg/ml, and the intra- and inter-assay variability were 8.4 and 8.0%, respectively. The cross-reactivity found was 0.14% for adrenaline and 1.8% for dopamine.
2.4. Determination of plasma glucose
Plasma glucose concentrations were determined in 60-day-old adult female rats using Accu-chek Active test strips. Blood was drawn by inserting a small cannula (PE-10, Intramedic, Clay Adams) into the tail vein. After an overnight fast, the glucose concentration was determined at times of 0, 30, 60, 90 and 120 minutes after the oral administration of 2 g of glucose/ kg weight of a 40% solution. At each time, sufficient blood was collected for immediate glucose determination and the subsequent determination of plasma insulin.
2.5. Plasma insulin determination by ELISA kit
Insulin levels were determined in the plasma of 60-day-old control and stressed adult female rats. This was performed using ELISA (80-INSRT-E01, E10, ALPCO). The sensitivity of this kit was 0.124 ng/ml; the kit presented 120% cross-reactivity with human insulin but less than 0.01% with rat C-peptide. The intra-assay variability was less than 5.8% and the inter-assay variability was less than 0.95%. The HOMA index was used to determine the presence or absence of insulin resistance; it was calculated using the following formula [24]:
The presence of insulin resistance was considered when the index was greater than 2.5.
2.6. Binding of [3H]dihydroalprenolol to Cardiac Membranes
2.6.1. Preparation of the Crude Membrane Fraction
One hundred milligrams of the basal area of the heart tissue was weighed and homogenized in an ultraturrax with 0.7 ml of 20 mM Tris–HCl/0.25 M sucrose buffer at pH 7.4. Then, it was centrifuged at 13,000 rpm for 20 min at 4 °C. The supernatant was discarded, and the precipitate was suspended in 0.7 ml of 20 mM Tris–HCl/0.25 M sucrose buffer at pH 7.4 and centrifuged again at 13,000 rpm for 20 min at 4 °C. The supernatant was removed, the precipitate was suspended in 0.7 ml of 20 mM Tris–HCl/10 mM MgCl2 buffer pH 7.4 without sucrose, and the protein concentration was measured by the Bradford method [25].
2.6.2. Assay for β-Adrenergic Receptors
Glass tubes were used, working in duplicate; they contained the membrane samples obtained after tissue homogenization (volume equivalent to 300 μg of protein) and 20 μl of 10 nM 3H-DHA in a total volume of 0.2 ml. The assay contained 0.5–40 nM 3Hdihydroalprenolol (92.1 Ci/mmol, NEN DuPont). Nonspecific binding was assessed: 20 μl of 10−3 M propranolol (only for the tubes corresponding to nonspecific binding) and 20 mM Tris–HCl/10 mM MgCl2 buffer pH 7.4 without sucrose (in a sufficient quantity to reach 200 μL as the final volume). The tubes were incubated for 30 min at 37 °C. Then, they were filtered with Whatman GF/C filters in a vacuum system and washed three times with 2 ml of 20 mM Tris–HCl/10 mM MgCl2 buffer pH 7.4 without sucrose with 10–5 M propranolol. Finally, the filters were placed in scintillation vials, 0.5 ml of distilled water and 4 ml of scintillation mix were added to each vial, each tube was vortexed, and the radioactivity was measured in a scintillation counter 24 h later [26, 27]. Statistical analyses were performed using GraphPad Prism 6 (GraphPad Software, San Diego, CA, USA).
2.7. Determination of Cardiac β1- and β2-Adrenergic Receptor mRNA
RNA was extracted by the method described by Chomczynski and Sacchi [35] from the heart (control and stress). For the first-strand cDNA synthesis, we used SUPERSCRIPT II (Invitrogen, Carlsbad, CA, USA; 200 units) in a 20 µL reaction volume with 1 µg of total RNA. The primers used were from the published work of [36], and a BLAST analysis was performed to determine the specificity of the sequences. The sequences of the primers for real-time PCR were as follows.
| Genes | Forward | Reverse |
| 18S | 5’-TCAAGAACGAAAGTCGGAGG-3’ | 5’-GGACATCTAAGGGCATCACA-3’ |
| β1AR | 5’-TCGTAGTGGGCAACGTGTTGGTGAT-3’ | 5’-GTCTACGAAGTCCAGAGCTCACAGAA-3’ |
| β2AR | 5’-CAGGCCTATGCTATCGCTTCCTCTAT-3’ | 5’-GGCTGAGGTTTTGGGCATGAAATC-3’ |
The PCR reaction mix contained 10 μl of Brilliant II SYBR Green QPCR Master Mix (Agilent Technologies, Inc., California, USA), 0.25 μM of each 18S primer, 0.1 μM of each βAR primer, 2 μl of cDNA and sterile water for a final volume of 20 μl. PCR reactions were performed using the IQ5 real-time thermocycler (BioRad) under the following conditions: 94°C for 20 s, 60°C for 20 s for βAR (60°C for GAPDH), 78°C for 20 s and a final extension of 72°C for 10 min. All samples for RT-qPCR analysis were run in triplicate and a mean value was used for the determination of the mRNA levels. The relative quantification of the βAR mRNA levels was performed using the ratio between the βAR and 18S mRNA levels.
2.8. Determination of Cardiac β1- and β2-Adrenergic Receptors by Western Blot
Protein extraction: Approximately 100 mg of cardiac tissue was weighed and homogenized in a glass using RIPA buffer (Tris–HCl pH 7.2 10 mM, Triton X-100 1%, NaCl 150 mM, SDS 0.1%), a process carried out on ice. The obtained product was centrifuged at 13,000 rpm for 20 min at 4 °C. The pellet was discarded, the supernatant was recovered, and the protein concentration was subsequently measured using the Bradford method [25].
Electrophoresis in polyacrylamide gels: Gels were loaded with 20 µg of protein. Electrophoresis was performed at a constant voltage of 120 V in 1X electrophoresis buffer (Tris base 0.3% (w/v), glycine 1.44% (w/v), SDS 0.1% (w/v); pH 8, 6) for 90 min. The proteins were transferred to a pore nitrocellulose membrane of 0.45 μm at a constant level of 400 mA for 90 min in transfer buffer (Tris base 0.5% (w/v), glycine 2.4% (w/v) and methanol 20% (v/v)). After the transfer was completed, the membrane was stained with 5% Ponceau Red to verify the correct transfer. Subsequently, the staining was eliminated via washing with tap water and three washes with TBST 0.1% (Tris base 0.224% (p/v), NaCl 0.8% (p/v), Tween 0.1% (v/v), adjusted to pH 7.6), each for 5 min. Nitrocellulose membranes were incubated with gentle shaking with 5% milk in 0.1% TBST at room temperature to block nonspecific binding sites. We performed three washes with 0.1% TBST, each for 5 min; subsequently, incubation was performed overnight at 4 °C with primary antibodies to the β1- or β2-adrenergic receptor using a dilution of 1:3000 or 1:2500, respectively. The antibodies used for beta-1 were a polyclonal anti-beta 1 adrenergic receptor antibody (ab3442, from ABCAM) and a recombinant anti-beta 2 adrenergic receptor antibody (ab182136, from ABCAM). Both antibodies were diluted in TBST 0.1 and 5% milk to avoid nonspecific binding. After incubation, the membrane was washed three times for 5 min with TBST 0.1% and incubated for 1 h, with shaking and at room temperature and with the secondary antibody (anti-rabbit polyclonal antibody from ABCAM), in both conditions using a dilution of 1:10,000 in TBST 0, 1%. The membrane was washed three times for 5 min each time. GAPDH, a highly conserved protein present in similar amounts, independent of the sample type, was used as a loading control. The primary antibody was incubated for 1 h using a dilution of 1:40,000 and subsequently incubated with a secondary anti-rabbit antibody incubated at 1:10,000 for 45 min. Using the GeneSys G-Box system (Syngene Headquarters, Frederick, MD, USA), chemiluminescence was developed using the EZ-ECL reagent. The intensity of the bands was evaluated using the ImageJ program.
2.9. Morphometric Analysis
The medial portion of the left ventricle was fixed in Bouin, dehydrated at 4 °C for 24 h and washed with 70% ethanol, and the tissue was embedded in paraffin. We used 5 µm slices stained with hematoxylin–eosin and Masson’s trichrome and examined them under a light microscope. We measured the area and perimeter of the cardiomyocytes according to Nakamura et al. [28]. Briefly, we analyzed four contiguous slices, and when an apparent nucleus appeared, the area and perimeter were measured with the ImageJ software, version 1.53t. All analyses were performed with at least 100 cells per heart.
2.10. Statistical Analysis
The data were expressed as the mean ± SEM. Statistical analyses were performed using GraphPad Prism 6 (GraphPad Software, San Diego, CA, USA). To test for significant differences between two groups, we used Student’s t-test and the Mann–Whitney test. To test for significant differences among more than two groups, we used one-way ANOVA, followed by the Newman–Keuls post hoc test to check for the differences between pairs of data. The number of animals for all experiments was calculated as the minimum number of animals according to the variability of the experimental procedures. The minimum number of animals was calculated according to Equation [29]:
where is the number of animals for each condition, S is the standard deviation, d is the difference needed to obtain statistical significance, Zα is the probability of type I error (significance) and Zβ is the probability of type II error (power). We proposed α = 0.05, i.e., the probability of finding a statistically significant difference was 0.05; β = 0.3 denoted the probability of having a difference between the populations; the intrapopulation variation was 0.2; and d, the smallest difference in the population, was 0.11. Thus, we obtained n = 4.5. Therefore, to obtain a statistically significant difference of p < 0.05, we needed to use four or five animals per study group.
3. Results
3.1. Animal and Heart Relative Weight
Stress did not modify the gestation days, pup numbers, or male and female pup proportions (data not shown but presented in [20]).
No changes in the weight of stressed or control rats during development were found (Figure 2, left graph, A), but a slight but significant increase in the weight of the heart was found at 20 and 60 days old (upper right, B). There were no changes in the serum levels and heart concentrations of NE (letters C and D).
3.2. Changes in β1 and β2 Adrenergic Receptor Protein Abundance
mRNA determination was performed for the β1- and β2-adrenergic receptors (Figure 3A). Control rats presented an age-dependent decrease in the β1 mRNA at 20 days old. Stressed rats maintained higher levels of the mRNA for the receptor during all periods. The β2AR mRNA in the hearts of stressed rats decreased as they aged to reach lower levels at 60 days old. Control rats presented a similar tendency but at lower levels. The β-adrenergic receptor abundance was studied by Western blot in the heart tissue (Figure 3B). The hearts of control rats presented increased levels of the β1AR protein with age, and a further increase was found in the hearts of stressed rats at 20 days old. The β2AR abundance decreased in the hearts of control and stressed rats at 20 days but the level was recovered at 60 days old (Figure 3B). No changes were observed in β2-adrenergic receptor abundance in stressed versus control rats (Figure 3B). Consequently, the cardiac level ratio of β1/β2 receptors increased in stressed rats (Figure 3C).
3.3. Concentration of β-Adrenergic Binding Sites
The β-adrenergic receptor affinity (IC50) and receptor numbers were measured in the heart tissue of the control and stressed female rats sacrificed at 60 days old. Competition curves were obtained to determine the possible effects of the gestational stress protocol on the affinity of these receptors using the radioligand technique in fractions of cardiac membranes from female rats sacrificed at 60 days old. Increasing concentrations (10−3–10−8 M) of propranolol, a nonselective β-adrenergic antagonist, were used, which allowed us to analyze the affinity of the total β-adrenergic receptor (Figure 4A–B). The IC50 (necessary dose of a drug to obtain 50% displacement) was used for each case (Figure 4C). A significant increase in the IC50 was observed in the hearts of stressed rats at 60 days old (Figure 4C). No changes in the β-adrenergic receptor number in each condition were observed (Figure 4D).
3.4. Effects of Isoproterenol on Ventricle Size and Cardiomyocyte Area in Gestationally Stressed Rats
We analyzed the ventricular size and cardiomyocyte area to analyze whether exposure to gestational stress also modified the morphological parameters of the heart ventricles and the cardiomyocyte size. As shown in Figure 5A, cardiomyocytes from the hearts of stressed rats presented a slight but significant increase in the perimeter and area of ventricles at 60 days old; however, after the ISO challenge for 10 days, the increases in ventricle size and area were greater in control rats (Figure 5A, B).
We wished to analyze the relative percentage of collagen in the heart under the different experimental conditions. In Figure 6 is show heart slices stained with Masson’s trichrome to visualize the collagen fiber in a blue-green color. There were no differences in the percentage of collagen under stress and under the isoproterenol challenge.
3.5. Cardiac Functional Parameters in Stressed Offspring Challenged with Isoproterenol Overload
Since we found differences in the affinity of β-adrenergic receptors and the ventricle size and cardiomyocyte area in stressed animals, we performed a functional analysis to verify the pathophysiological impact of exposure to an adrenergic loading protocol previously found to increase the size of cardiomyocytes [30]. In Figure 7-I, we show that 40% of the progeny of rats exposed in utero to sympathetic stress did not resist the administration of 125ug/kg of ISO and died by the fourth day of the daily dose of ISO, but none of the control rats treated with ISO did. To verify whether the effect was a consequence of the response to ISO, we implanted a microchip with a pressure transducer in the descending aorta to record the in vivo response of the cardiovascular system to ISO administration. In figure 7-II it is presented the systolic and diastolic pressure and pulse frequency for a control rat. When ISO was administered, there was a rapid increase in heart frequency with no change in the pressure response because there were no changes in systolic and diastolic pressure (Figure 7-III). Probably there was compensation in the pressure, most probably by vasodilation. However, in the rats exposed to gestational stress, irregularities in the pressure registration appeared, showing multiples extrasystole (Figure 7-IV, green circle), reaching a point at which 40% of the rats were not able to compensate (Figure 7-V). This was probably the reason that, in the rats under gestational stress, not only the morphological and biochemical processes of the heart were modified but also the capacity to respond to an adrenergic overload.
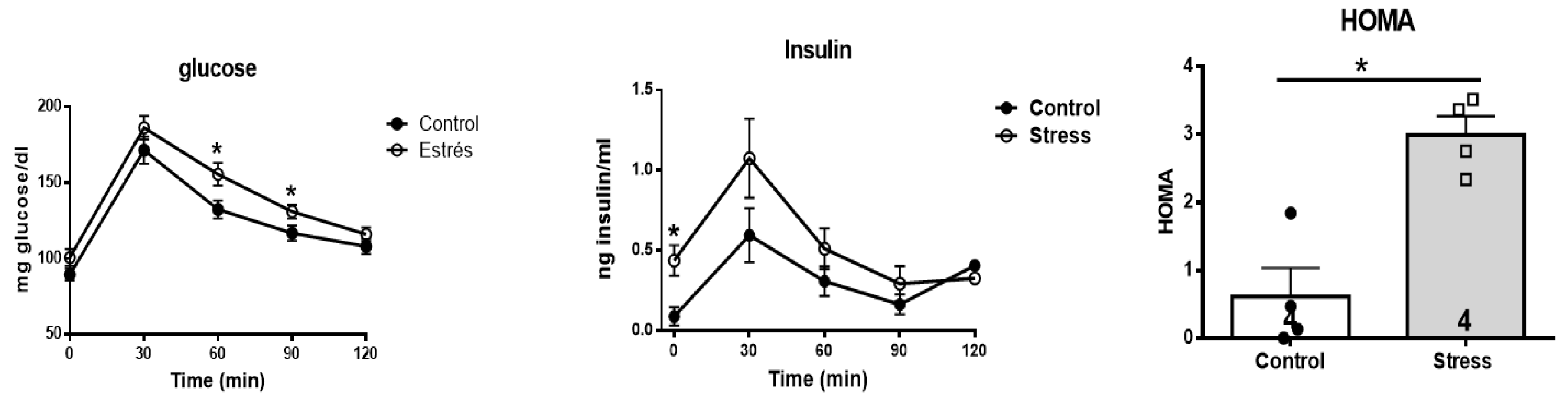
3.6. Effect of stress on the glycemic and insulin response to a glucose overload
To investigate the impact of prenatal stress on some of the metabolic markers in the progeny in adulthood, we analyzed the response to a glucose overload and the impact on blood glucose concentrations in control and stressed adult female offspring after the oral administration of 2 g of glucose/kg weight of a 40% glucose solution (Figure 8). It was observed that the curve corresponding to the stressed females was above the curve of the control females, and an increase in blood glucose concentrations was found at 60 and 90 minutes after the administration of glucose. In addition to glucose determination, blood insulin concentrations were determined in control and stressed adult female offspring. It was observed that, as in the previously measured glucose concentrations (Figure 8, middle), the curve corresponding to the stressed females was above the curve of the control females until 90 minutes after glucose administration. An increase in basal blood insulin concentrations was found prior to the administration of oral glucose (at time 0). With these data, we calculated the HOMA index (used to determine the presence or absence of insulin resistance, IR), considering IR when the index was greater than 2.5. We thus confirmed the presence of IR in the progeny of rats exposed to stress in the uterus.
4. Discussion
Most modern theories about stress recognize that although stress is not a disease, it may trigger the majority of diseases when an allostatic overload is generated. During stress, glucocorticoids and catecholamines play a key role in the regulation of physiological parameters and homeostasis. The cold stress protocol is classified as a sympathetic stress protocol, since only this system is activated, producing the consequent release of NE without a change in the concentration of ACTH or A [10]. This result indicates that there is no activation of the HHA axis [13]. During pregnancy, the placenta acts as an interface between the mother and the fetus, so any alteration in this connection can induce long-term changes in the health of the offspring. In this case, we previously found that applying this cold stress protocol in rats increases the plasma concentration of NE in the stressed mother, which leads to the lower expression of placental NETs [9]. A decrease in the functionality of the transporter leads the fetus (either male or female) to develop in an environment with high concentrations of catecholamines such as NE, which can lead to long-term effects on offspring in adulthood. In the present study, we choose groups of 20- and 60-day-old rats because they represent two stages of development, prepubescent and adult, respectively (albino rats reach sexual maturity by the fourth week of age, either in females or males [31]). Unlike adult rats, prepubescent rats still have an immature sympathetic nervous system in the peripheral organs, such as the ovary [32] and the vas deferens [33], where the sympathetic nervous system changes throughout postnatal development until they reach sexual maturity, and the nervous system is already functionally activated. We found important changes not only in the size of the heart but, age-dependent changes in the heart β-adrenergic receptors but most importantly, in the functional activity of the heart and in βAR sensitivity.
Effects of Gestational Stress in the Heart
The β1- and β2-adrenergic receptors are the subtypes predominantly expressed in the hearts of many mammalian species, including humans, and they are the principal regulators of cardiovascular function [34]. Under normal physiological conditions, the β1-adrenergic receptor, the predominant subtype in cardiomyocytes, comprises approximately 80% of the total β-adrenergic receptors, whereas the β2-adrenergic receptor comprises approximately 20%. Regarding β-adrenergic receptors, it has been observed in humans that heart failure produces alterations in the population of β-adrenergic receptors in the ventricular myocardium, causing a decrease in the concentration of β1-adrenergic receptors due to a downregulation mechanism, without presenting changes in the β2-adrenergic receptor population. The stoichiometry of the two β-adrenergic receptor subtypes changes from 77:23 in the healthy heart to approximately 60:40 under heart failure conditions, which is mainly caused by the selective downregulation of β1-adrenergic receptor expression [35]. The decrease in β1 mRNA at 20 days old, found in control rats, was not followed by similar changes in the mRNA from stressed rats. The β2AR mRNA decreased during all time periods studied, thus increasing the ratio of β1/β2. Jevjdovic et al. [36] showed that prenatal stress might induce region- and sex-specific β1- and β2-adrenergic receptor expression patterns within the left ventricle. Using an unpredictable stress paradigm during gestation, they found a decrease in the ratio of receptors, but they did not find differences in the female progeny as we did. This difference could arise from the fact that our stress paradigm reflected sympathetic stress, and the one used by Jevjdovic was a mixed stress paradigm that also produced changes in corticoids; thus, they could have revealed a mixed effect that could be opposed, as we have previously found with restriction and cold stress [37]. In addition [38], the effect of different stressors could differentially affect the expression of the β1 and β2 receptors; diet restriction increased the β2 receptor levels in females but not in males. Thus, maternal protein restriction programs cardiac sympathetic activity in a sex-specific manner, and this might explain the increased susceptibility to ischemia–reperfusion injury in males subject to fetal undernutrition. Similarly, Fernandez-Twinn et al. [7] found that low-protein offspring from stressed prenatal rats had reduced β-adrenergic responsiveness and attenuated adrenergic and insulin signaling, suggesting that intrauterine undernutrition also alters the heart failure risk. The multiple effects found by different stressors offer the possibility for multiple studies to understand how the maternal medium affects the progeny. Our studies may suggest that the changes in the β receptor ratio could be manifested in changes at the heart level and also in cardiovascular function.
On the other hand, regarding the cardiac consequences of this prenatal stress, in the present work, we found that the hearts of female rat progeny from stressed mothers presented slight changes in the weight of the heart (probably ventricular hypertrophy), although we found clear changes in adrenergic receptor abundance and the response to a challenge with ISO. These results agree with previous data from our laboratory in which we observed that the sisters of the male progeny presented a more significant and prolonged cardiovascular response to a stress situation than previously described in adult females [8]. In addition, they were more susceptible to β-adrenergic stimulation with isoproterenol [22] than males (125 µg/kg instead of 1 mg/kg in males), indicating that the existence of a higher concentration of receptors provides greater sensitivity to adrenergic stimulation.
The increase in the IC50 of propranolol found in rats exposed to sympathetic stress in utero indicates the lower affinity of drugs for β-adrenergic receptors. Although the decrease in the affinity of the ligand for its receptor could be the result of post-transductional changes in the receptors, the displacement results did not show a decrease in the number of receptors. The decrease in affinity could be explained by protein modification mechanisms, occurring via intrauterine stress. Fewer β1-adrenergic receptors and low affinity could result in conditions similar to those found in myocardial infarctions, in which there is a decrease in the concentration of β1-adrenergic receptors without changes in the concentration of β2-adrenergic receptors [39], and they could be associated with changes in the pathological responses of the heart to stress.
On functional grounds, the fact that gestational stress did not change the noradrenergic activity in the hearts of the female progeny, with no changes in the receptor protein, means that either higher sympathetic activity or a higher concentration of β1 or β2 agonists is needed to obtain normal function. In contrast, we observed unchanged NE and increased β1-adrenergic receptor abundance, probably as a compensatory mechanism. In addition, under the lower NE activity, the administration of ISO could act on other receptors, especially at the central level, to modify the sympathetic discharge from the PVN and produce an imbalance in systolic pressure and frequency (local effect in the pacemaker) [40]. This condition could generate a pressure response in which the heart cannot maintain a high frequency, which finally produces extrasystolic activity and thus noncontrolled heart function once the sympathetic nervous system is activated in the face of a stressful situation, as in this cold stress protocol, where the release of NE occurs, which acts on the β-adrenergic receptors. In this condition, gestational stress programs the β-adrenergic receptors during prenatal development, and the effects of this programming are seen in postnatal development, with the heart being more sensitive to neuronal overload, without the ability to regulate and thus being weaker to adapt to more stressful conditions.
Consequently, ISO administration caused death in 40% of rats from stressed mothers during the first 4 days of treatment, versus the 100% survival of control rats treated with ISO, which prematurely demonstrated the effect of maternal cold-stress-induced physiological changes in the heart, so that they were more susceptible to the adrenergic overload. This greater reactivity to treatment with ISO in the rats stressed in utero could be associated with the change in the affinity of the β1-adrenergic receptors. In this sense, it was observed that it was associated with the larger size of the left ventricle and the larger area of cardiomyocytes, as shown in our work.
During the stress reaction, the sympathetic adrenal medullary system and the hypothalamus–pituitary–adrenal cortex axis are activated, causing β-adrenergic receptor overstimulation and the remodeling of the β1/β2-adrenergic receptor ratio in the membrane fraction of the cardiac tissue. Cardiovascular disorders have also been related to altered β-adrenergic receptor signaling at or beyond the receptor level. A common feature of many of these conditions seems to be decreased β1-adrenergic receptor signaling coinciding with increased β2-adrenergic receptor signaling. These may play an adaptive role in the increased sympathetic drive to the heart, protecting the cardiac tissue from the cardiotoxic effects mediated by β1-adrenergic receptors without altering the cardiac output, since this would be sustained by the β2-adrenergic receptors, which would also have cardioprotective effects against myocyte apoptosis and remodeling. Our data found in stressed rats describe increased β1AR, meaning that, in this condition, there is no adaptive role but a disease state fits better. Thus, the selective maintenance of the β2-adrenergic receptor population might help to diminish the risk of the catecholaminergic overstimulation of the heart since this adrenoceptor subtype couples to Gs and Gi and thus constitutes a part of the “suppressive” action of glucocorticoids, which limits the stress reaction.
Moreover, the apoptotic and anti-apoptotic effects of catecholamines mediated by β1- and β2-adrenergic receptors, respectively, represent the “protective” role of glucocorticoids during the stress reaction [41]. Alterations in the density of cell surface receptors or the affinity of these receptors to different agonists, regardless of whether they are endogenous, as well as in the coupling of these receptors to intracellular effectors, are of crucial clinical importance, since these processes may alter the functioning of the cardiac tissue and its sensitivity to endogenous mediators or therapeutic drugs. In the human heart atria, increased coupling of the β1- and β2-adrenergic receptors to their intracellular effectors triggers the processes of arrhythmia. In this regard, our results showed the strong presence of arrhythmia, which could be associated with maladaptive adrenergic stimulation.
The insulin resistance due to gestational stress confirmed our results that overactivation by sympathetic stress during gestation influences insulin resistance, and it confirmed a finding previously described in the rat ovary, which was closely related to the formation of polycystic ovaries [15].
Finally, the cardiotoxic effects of catecholamines are mediated via prolonged adrenergic receptor stimulation. Thus, the increase in β1-adrenergic receptors could be associated with a mechanism to protect cardiac function from excessive adrenergic stimulation. Chronic β1-adrenergic receptor hyperstimulation by ISO results in their downregulation, and, consequently, there is a marked reduction in the inotropic cardiac reserve, as evidenced by frequent tachyarrhythmia leading to death.
5. Conclusions
The present study demonstrated that the exposure of the rat’s progeny to catecholamine during fetal development permanently modifies their cardiovascular function, making the rat more susceptible to changes in the catecholaminergic environment and thus decreasing the capacity to respond to changes in the external medium in adulthood. The identification of the mechanisms that induce such alterations in the β-AR, as well as the protective mechanisms against these alterations, may be of major importance in the clarification of the pathological processes related to stress due to noradrenergic exposure, as well as its therapy, to prevent cardiovascular risks derived from exposure to stress during gestation.
Author Contributions
B.P. performed most of the experimental work with rats and the biochemical analysis, discussed the theoretical phases and experimental design of the work and contributed to the draft and the final version of the paper. A.P. carried out the molecular determination and Western blot analysis. S.P. carried out all morphological work and isoproterenol administration. C.A. was responsible for the peripheral neurotransmitters. H.E.L. directed the work and drafted the paper. All authors have read and agreed to the published version of the manuscript.
Funding
FONDECYT 1130049, 1170291 (to H.E.L.) and Conicyt grant for doctoral thesis 21120077 (to B.P.).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board (protocol number: CBE2016-13 to BP and CBE2017-05 to HL).
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are available under written request.
Acknowledgments
This work was performed in partial fulfillment of the requirements of a Ph.D. degree in Pharmacology to B.P. This work was supported, in part, by FONDECYT 1130049, 1170291 (to H.E.L.) and a Conicyt grant for doctoral thesis 21120077 (to B.P.).
Conflicts of Interest
The authors declare that there are no conflicts of interest that could be perceived as prejudicing the impartiality of the research reported.
References
- Cottrell EC, Seckl JR. Prenatal stress, glucocorticoids and the programming of adult disease. Front Behav Neurosci. 2009;3:19. [CrossRef]
- Kajantie E. Fetal origins of stress-related adult disease. Ann N Y Acad Sci. 2006;1083:11-27. [CrossRef]
- Hales CN, Barker DJ. The thrifty phenotype hypothesis. Br Med Bull. 2001;60:5-20. [CrossRef]
- Godfrey KM, Barker DJ. Fetal programming and adult health. Public Health Nutr. 2001;4(2B):611-24. [CrossRef]
- Phillips DI, Jones A, Goulden PA. Birth weight, stress, and the metabolic syndrome in adult life. Ann N Y Acad Sci. 2006;1083:28-36. [CrossRef]
- Lim K, Zimanyi MA, Black MJ. Effect of maternal protein restriction in rats on cardiac fibrosis and capillarization in adulthood. Pediatr Res. 2006;60(1):83-7. [CrossRef]
- Fernandez-Twinn DS, Ekizoglou S, Wayman A, Petry CJ, Ozanne SE. Maternal low-protein diet programs cardiac beta-adrenergic response and signaling in 3-mo-old male offspring. Am J Physiol Regul Integr Comp Physiol. 2006;291(2):R429-36.
- Igosheva N, Klimova O, Anishchenko T, Glover V. Prenatal stress alters cardiovascular responses in adult rats. J Physiol. 2004;557(Pt 1):273-85. [CrossRef]
- Piquer B, Fonseca JL, Lara HE. Gestational stress, placental norepinephrine transporter and offspring fertility. Reproduction. 2017;153(2):147-55. [CrossRef]
- Goldstein DS, Kopin IJ. Adrenomedullary, adrenocortical, and sympathoneural responses to stressors: a meta-analysis. Endocr Regul. 2008;42(4):111-9.
- Pacak K, Palkovits M, Yadid G, Kvetnansky R, Kopin IJ, Goldstein DS. Heterogeneous neurochemical responses to different stressors: a test of Selye's doctrine of nonspecificity. Am J Physiol. 1998;275(4 Pt 2):R1247-55. [CrossRef]
- Writing Group M, Mozaffarian D, Benjamin EJ, Go AS, Arnett DK, Blaha MJ, et al. Heart Disease and Stroke Statistics-2016 Update: A Report From the American Heart Association. Circulation. 2016;133(4):e38-360.
- Benedict CR, Fillenz M, Stanford C. Noradrenaline release in rats during prolonged cold-stress and repeated swim-stress. Br J Pharmacol. 1979;66(4):521-4. [CrossRef]
- Dorfman M, Arancibia S, Fiedler JL, Lara HE. Chronic intermittent cold stress activates ovarian sympathetic nerves and modifies ovarian follicular development in the rat. Biol Reprod. 2003;68(6):2038-43. [CrossRef]
- Dorfman M, Ramirez VD, Stener-Victorin E, Lara HE. Chronic-intermittent cold stress in rats induces selective ovarian insulin resistance. Biol Reprod. 2009;80(2):264-71. [CrossRef]
- Barra R, Cruz G, Mayerhofer A, Paredes A, Lara HE. Maternal sympathetic stress impairs follicular development and puberty of the offspring. Reproduction. 2014;148(2):137-45. [CrossRef]
- Piquer B, Ruz F, Barra R, Lara HE. Gestational Sympathetic Stress Programs the Fertility of Offspring: A Rat Multi-Generation Study. Int J Environ Res Public Health. 2022;19(5). [CrossRef]
- Lian S, Wang D, Xu B, Guo W, Wang L, Li W, et al. Prenatal cold stress: Effect on maternal hippocampus and offspring behavior in rats. Behav Brain Res. 2018;346:1-10. [CrossRef]
- Sun D, Chen K, Wang J, Zhou L, Zeng C. In-utero cold stress causes elevation of blood pressure via impaired vascular dopamine D1 receptor in offspring. Clin Exp Hypertens. 2020;42(2):99-104. [CrossRef]
- Piquer B, Olmos D, Flores A, Barra R, Bahamondes G, Diaz-Araya G, et al. Exposure of the Gestating Mother to Sympathetic Stress Modifies the Cardiovascular Function of the Progeny in Male Rats. Int J Environ Res Public Health. 2023;20(5). [CrossRef]
- Ayala P, Montenegro J, Vivar R, Letelier A, Urroz PA, Copaja M, et al. Attenuation of endoplasmic reticulum stress using the chemical chaperone 4-phenylbutyric acid prevents cardiac fibrosis induced by isoproterenol. Exp Mol Pathol. 2012;92(1):97-104. [CrossRef]
- Luna SL, Neuman S, Aguilera J, Brown DI, Lara HE. In vivo beta-adrenergic blockade by propranolol prevents isoproterenol-induced polycystic ovary in adult rats. Horm Metab Res. 2012;44(9):676-81.
- AVMA. American Veterinary Medical Association Guidelines for the Euthanasia of Animals: 2020 Edition. 2020.
- Muniyappa R, Chen H, Muzumdar RH, Einstein FH, Yan X, Yue LQ, et al. Comparison between surrogate indexes of insulin sensitivity/resistance and hyperinsulinemic euglycemic clamp estimates in rats. Am J Physiol Endocrinol Metab. 2009;297(5):E1023-9. [CrossRef]
- Kruger NJ. The Bradford method for protein quantitation. Methods Mol Biol. 1994;32:9-15.
- Barria A, Leyton V, Ojeda SR, Lara HE. Ovarian steroidal response to gonadotropins and beta-adrenergic stimulation is enhanced in polycystic ovary syndrome: role of sympathetic innervation. Endocrinology. 1993;133(6):2696-703. [CrossRef]
- Lara HE, Porcile A, Espinoza J, Romero C, Luza SM, Fuhrer J, et al. Release of norepinephrine from human ovary: coupling to steroidogenic response. Endocrine. 2001;15(2):187-92. [CrossRef]
- Nakamura K, Fushimi K, Kouchi H, Mihara K, Miyazaki M, Ohe T, et al. Inhibitory effects of antioxidants on neonatal rat cardiac myocyte hypertrophy induced by tumor necrosis factor-alpha and angiotensin II. Circulation. 1998;98(8):794-9. [CrossRef]
- Zar J. Biostatistical Analysis. second edition ed. NJ: Prentice Hall; 1984 1984. 5 p.
- Benjamin IJ, Jalil JE, Tan LB, Cho K, Weber KT, Clark WA. Isoproterenol-induced myocardial fibrosis in relation to myocyte necrosis. Circ Res. 1989;65(3):657-70. [CrossRef]
- Sengupta P. The Laboratory Rat: Relating Its Age With Human's. Int J Prev Med. 2013;4(6):624-30.
- Ricu M, Paredes A, Greiner M, Ojeda SR, Lara HE. Functional development of the ovarian noradrenergic innervation. Endocrinology. 2008;149(1):50-6. [CrossRef]
- Burnstock G, Verkhratsky A. Vas deferens--a model used to establish sympathetic cotransmission. Trends Pharmacol Sci. 2010;31(3):131-9. [CrossRef]
- Wang J, Gareri C, Rockman HA. G-Protein-Coupled Receptors in Heart Disease. Circ Res. 2018;123(6):716-35. [CrossRef]
- Bristow MR, Altman NL. Heart Rate in Preserved Ejection Fraction Heart Failure. JACC Heart Fail. 2017;5(11):792-4. [CrossRef]
- Jevjdovic T, Dakic T, Kopanja S, Lakic I, Vujovic P, Jasnic N, et al. Sex-Related Effects of Prenatal Stress on Region-Specific Expression of Monoamine Oxidase A and beta Adrenergic Receptors in Rat Hearts. Arq Bras Cardiol. 2019;112(1):67-75.
- Galvez A, Paredes A, Fiedler JL, Venegas M, Lara HE. Effects of adrenalectomy on the stress-induced changes in ovarian sympathetic tone in the rat. Endocrine. 1999;10(2):131-5.
- Elmes MJ, Haase A, Gardner DS, Langley-Evans SC. Sex differences in sensitivity to beta-adrenergic agonist isoproterenol in the isolated adult rat heart following prenatal protein restriction. Br J Nutr. 2009;101(5):725-34.
- Bristow MR, Ginsburg R, Umans V, Fowler M, Minobe W, Rasmussen R, et al. Beta 1- and beta 2-adrenergic-receptor subpopulations in nonfailing and failing human ventricular myocardium: coupling of both receptor subtypes to muscle contraction and selective beta 1-receptor down-regulation in heart failure. Circ Res. 1986;59(3):297-309. [CrossRef]
- Cayupe B, Morgan C, Puentes G, Valladares L, Burgos H, Castillo A, et al. Hypertension in Prenatally Undernourished Young-Adult Rats Is Maintained by Tonic Reciprocal Paraventricular-Coerulear Excitatory Interactions. Molecules. 2021;26(12). [CrossRef]
- Sapolsky RM, Romero LM, Munck AU. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr Rev. 2000;21(1):55-89. [CrossRef]
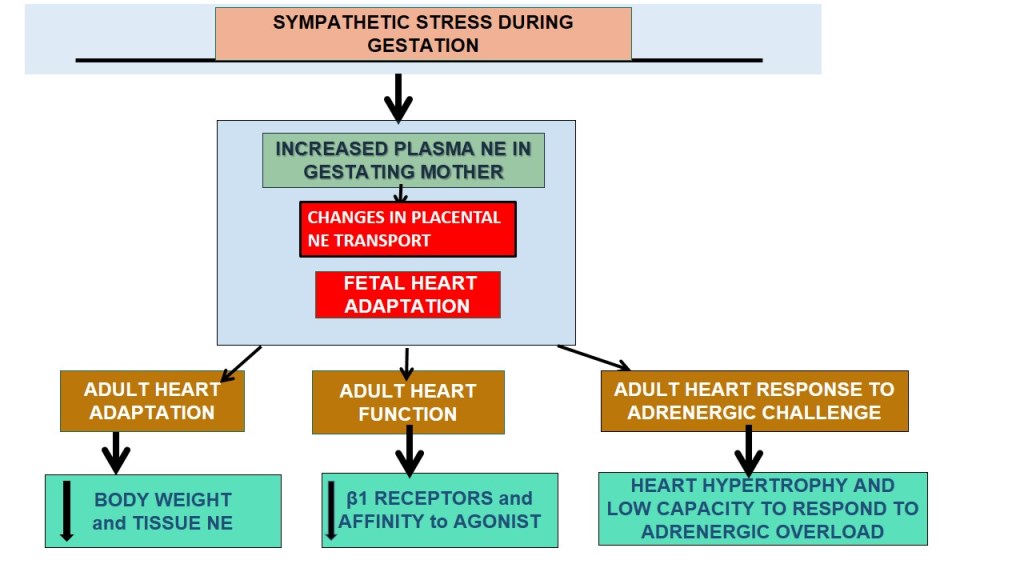
Figure 1.
Schematic representation of the experimental groups used in the study. Group A represented the effect of sympathetic gestational stress on sympathetic neurotransmitter levels in female progeny and its relationship with β-adrenergic receptor in the heart. In group B, we studied the in vivo effect of gestational stress on the development of cardiomyocytes in the adult heart and the response to an adrenergic overload with a β-agonist in vivo.
Figure 1.
Schematic representation of the experimental groups used in the study. Group A represented the effect of sympathetic gestational stress on sympathetic neurotransmitter levels in female progeny and its relationship with β-adrenergic receptor in the heart. In group B, we studied the in vivo effect of gestational stress on the development of cardiomyocytes in the adult heart and the response to an adrenergic overload with a β-agonist in vivo.
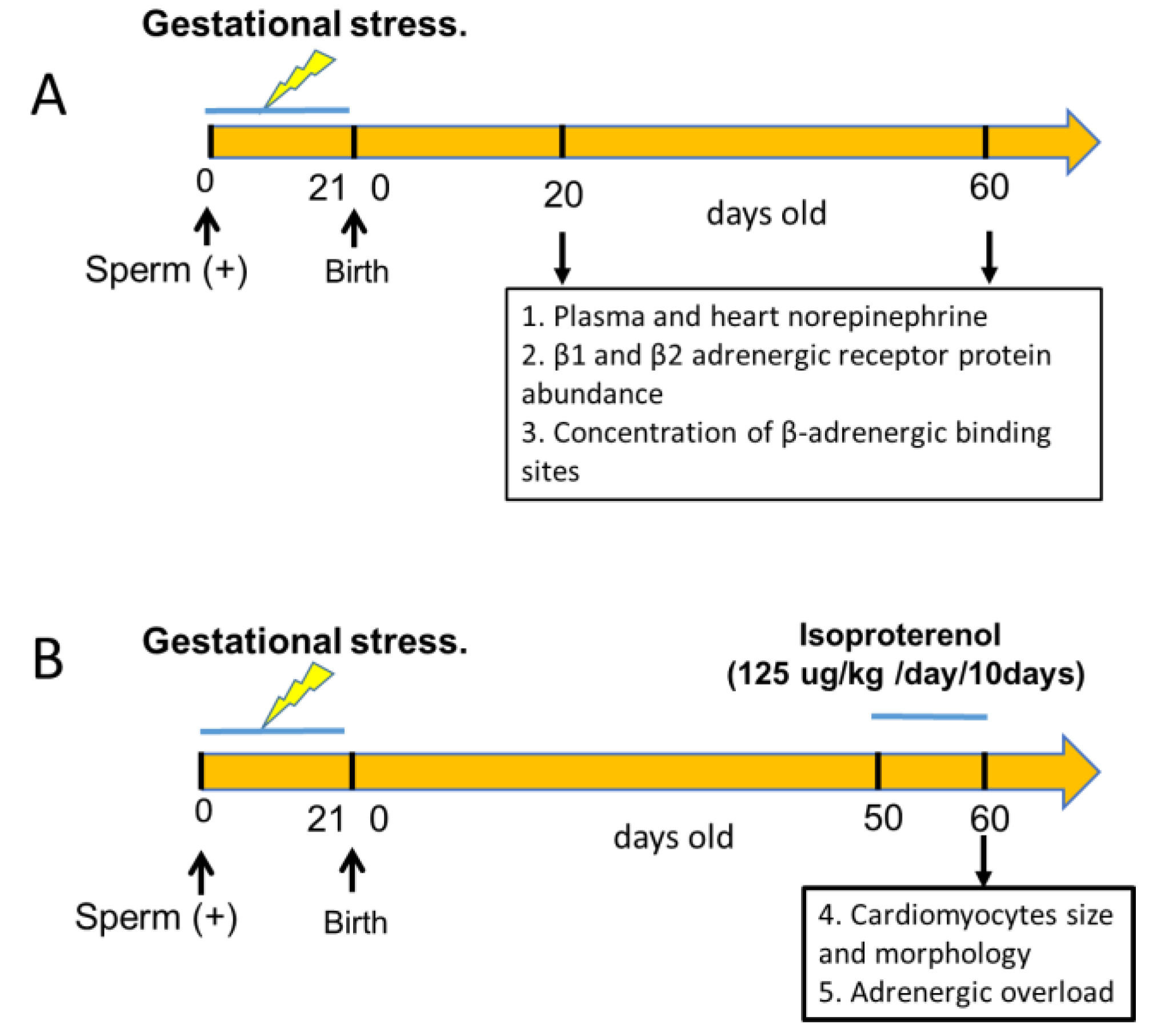
Figure 2.
Changes in the weight of the pups and heart during development and plasma and heart norepinephrine. Results are expressed as mean ± SEM of the number or rats shown in the bars. * = p <0.05.
Figure 2.
Changes in the weight of the pups and heart during development and plasma and heart norepinephrine. Results are expressed as mean ± SEM of the number or rats shown in the bars. * = p <0.05.
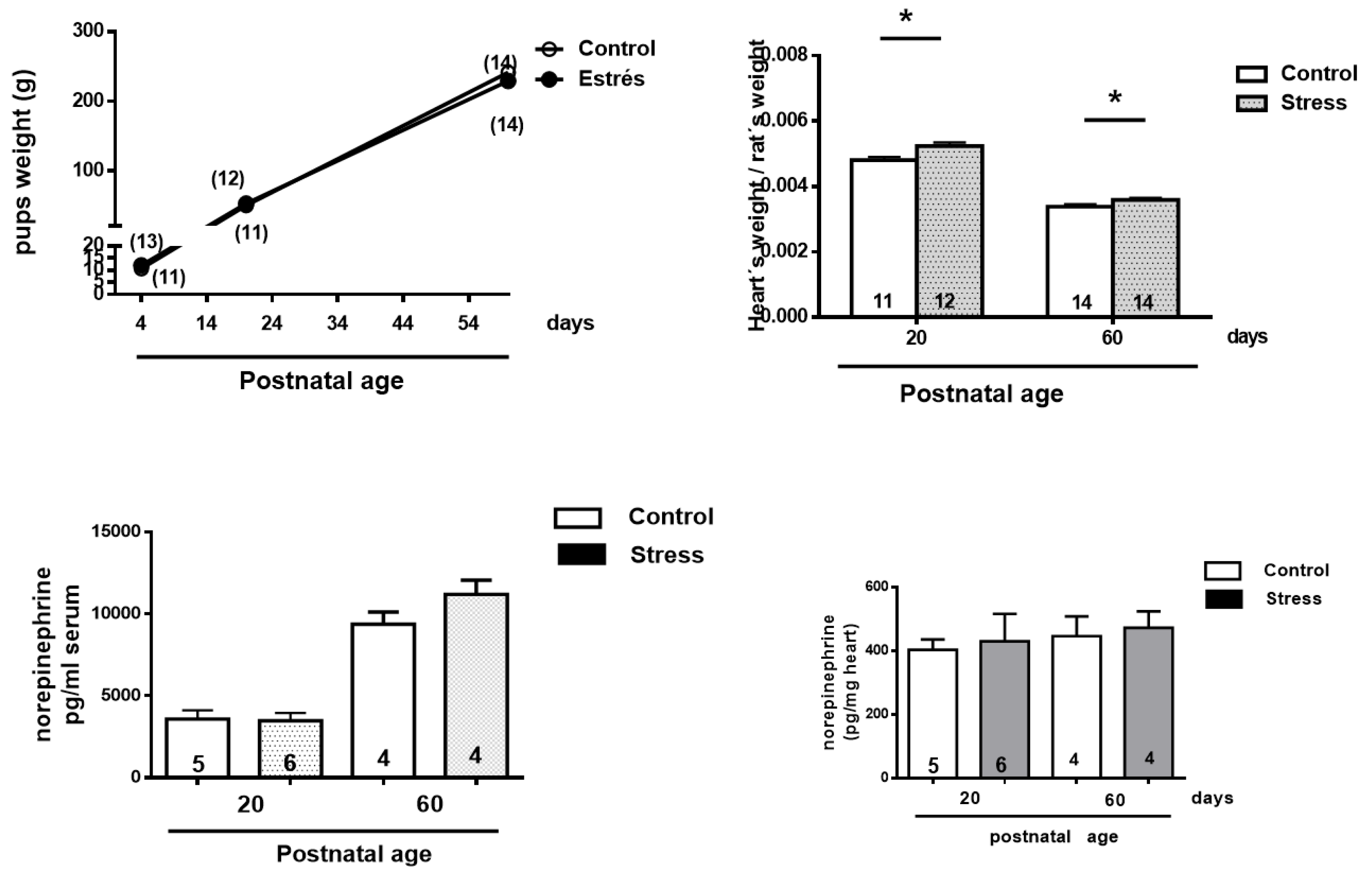
Figure 3.
mRNA determination (A) and Western blot analysis (B) of β1- and β2-adrenergic receptors (Figure 3). In Figure 2A (left side), we show the expression of β1AR; the lower-left side shows β2AR (A), normalized to 18S mRNA. The right side (B) shows the quantification of the signal normalized by GAPDH for 4 control and 5 stressed rats at 20 and 60 days old. β1AR represents β1 adrenergic receptor protein. We also show data for the β2-adrenergic receptor normalized by GAPDH for 4 rats in the control group and 5 rats in the stressed group at 20 and 60 days old. β2AR represents β2 adrenergic receptor protein. A representative blot is shown on the right side of 2(A,B) for all the conditions. The results are mean ± SEM of 4-5 rats in each condition. * p < 0.05 stress vs. control, ** p < 0.01 stress vs. control *** p < 0.001 stress vs. control.
Figure 3.
mRNA determination (A) and Western blot analysis (B) of β1- and β2-adrenergic receptors (Figure 3). In Figure 2A (left side), we show the expression of β1AR; the lower-left side shows β2AR (A), normalized to 18S mRNA. The right side (B) shows the quantification of the signal normalized by GAPDH for 4 control and 5 stressed rats at 20 and 60 days old. β1AR represents β1 adrenergic receptor protein. We also show data for the β2-adrenergic receptor normalized by GAPDH for 4 rats in the control group and 5 rats in the stressed group at 20 and 60 days old. β2AR represents β2 adrenergic receptor protein. A representative blot is shown on the right side of 2(A,B) for all the conditions. The results are mean ± SEM of 4-5 rats in each condition. * p < 0.05 stress vs. control, ** p < 0.01 stress vs. control *** p < 0.001 stress vs. control.
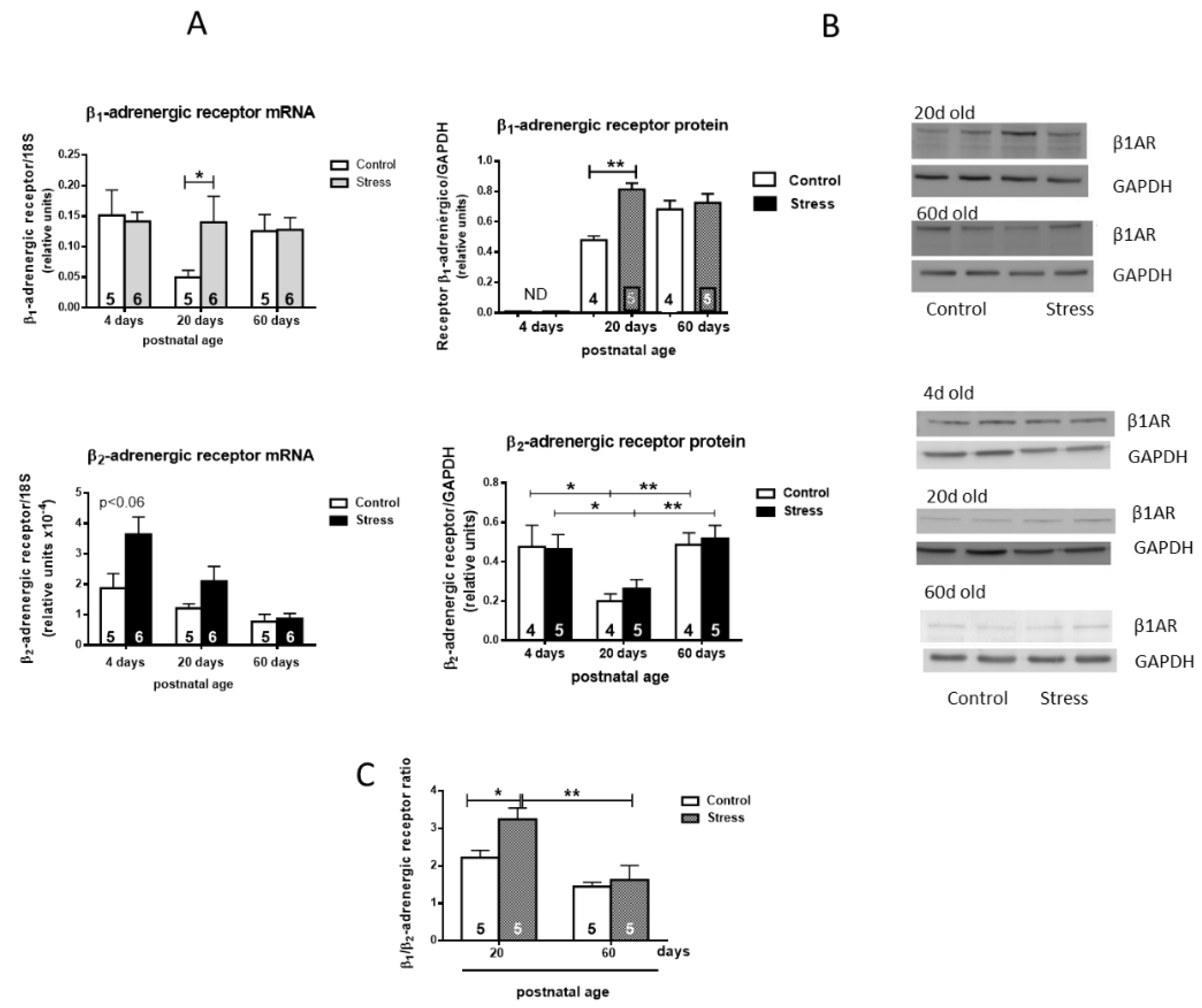
Figure 4.
Effect of gestational stress on the displacement of 3H-DHA and the affinity of β receptors from a membrane fraction of the heart homogenate from female progeny at 60 days old. In (A–B), the displacement of the binding of dihydroalprenolol by increasing doses of propranolol is shown. In C, we show the affinity of each of the β ligands; in D, the number of binding sites is shown. The results correspond to the mean ± SEM of 5 individual experiments for control and stressed rats. * p < 0.05 stress vs. control.
Figure 4.
Effect of gestational stress on the displacement of 3H-DHA and the affinity of β receptors from a membrane fraction of the heart homogenate from female progeny at 60 days old. In (A–B), the displacement of the binding of dihydroalprenolol by increasing doses of propranolol is shown. In C, we show the affinity of each of the β ligands; in D, the number of binding sites is shown. The results correspond to the mean ± SEM of 5 individual experiments for control and stressed rats. * p < 0.05 stress vs. control.
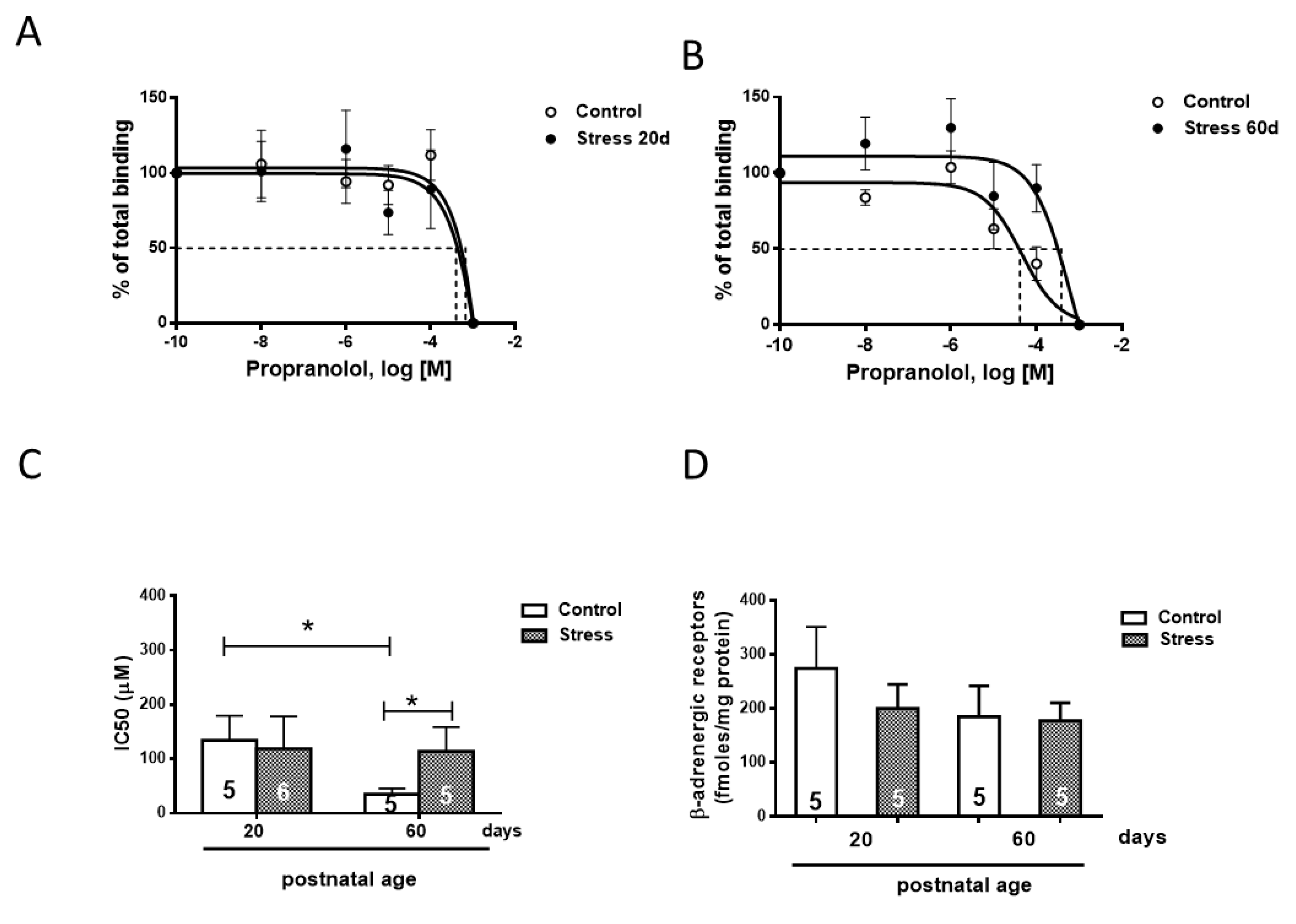
Figure 5.
Perimeter and area (A) and morphology (B) of cardiomyocytes of 60-day-old male rats after 10 days of isoproterenol or PBS treatment. Hearts were collected at 60 days old after 10 days of treatment with a daily dose of 125 ug/kg of isoproterenol. The results are the mean value ± SEM of 5 animals in each group. The bar indicates the magnification of the picture. A: ** p < 0.01 control-ISO vs. stress-ISO; C:*** p < 0.001 control-ISO vs. stress-ISO.
Figure 5.
Perimeter and area (A) and morphology (B) of cardiomyocytes of 60-day-old male rats after 10 days of isoproterenol or PBS treatment. Hearts were collected at 60 days old after 10 days of treatment with a daily dose of 125 ug/kg of isoproterenol. The results are the mean value ± SEM of 5 animals in each group. The bar indicates the magnification of the picture. A: ** p < 0.01 control-ISO vs. stress-ISO; C:*** p < 0.001 control-ISO vs. stress-ISO.
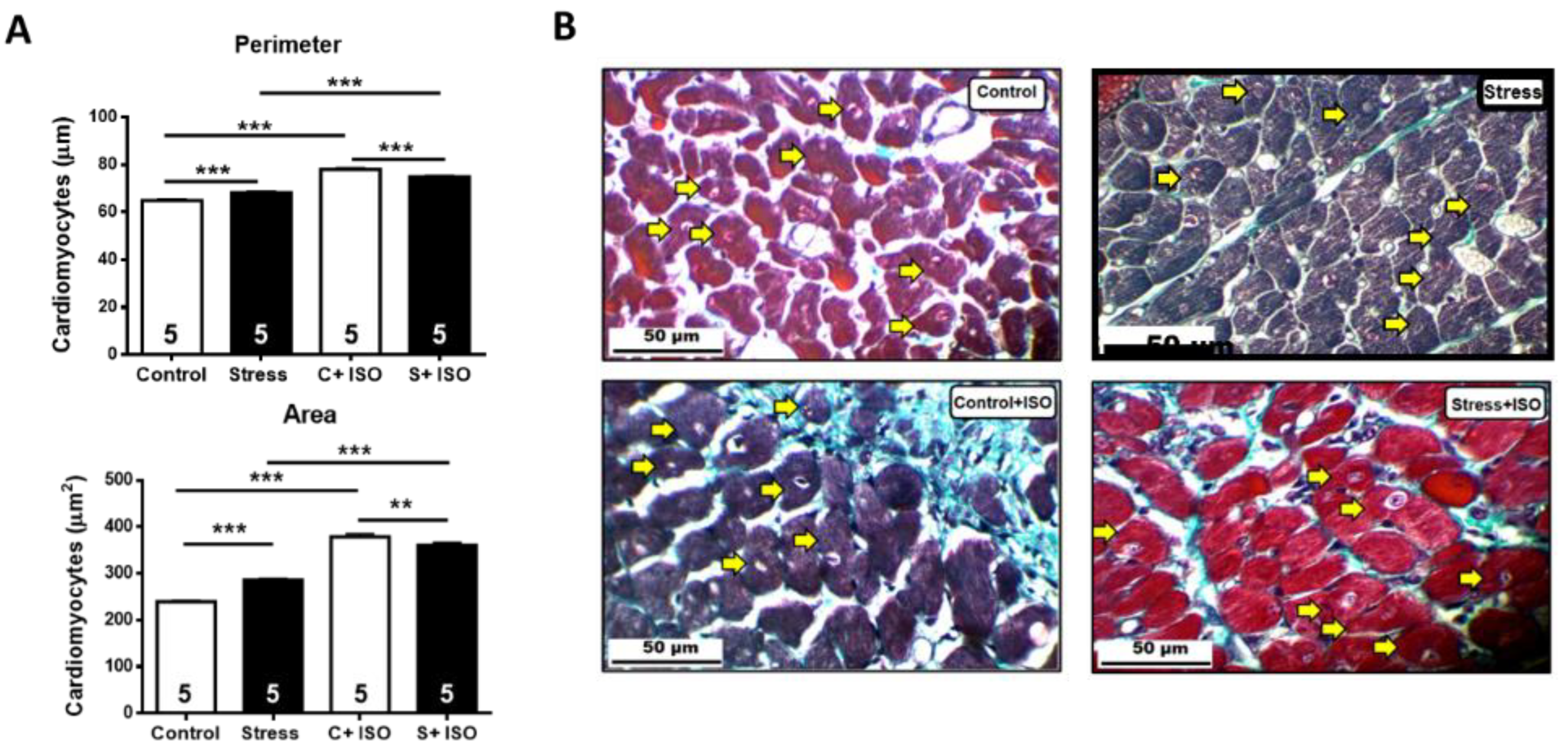
Figure 6.
Collagen determination in hearts of control and stressed rats with or without isoproterenol treatment. A. On the left is the quantification of collagen as a percentage of the total cells. B. Representative picture of a heart slice stained with Masson’s trichrome, in which the blue-green fibers are shown (yellow arrows). Results correspond to the mean value ± SEM of the number shown in each bar. C+ ISO: Control+ ISO; S+ ISO: Stress ISO.
Figure 6.
Collagen determination in hearts of control and stressed rats with or without isoproterenol treatment. A. On the left is the quantification of collagen as a percentage of the total cells. B. Representative picture of a heart slice stained with Masson’s trichrome, in which the blue-green fibers are shown (yellow arrows). Results correspond to the mean value ± SEM of the number shown in each bar. C+ ISO: Control+ ISO; S+ ISO: Stress ISO.
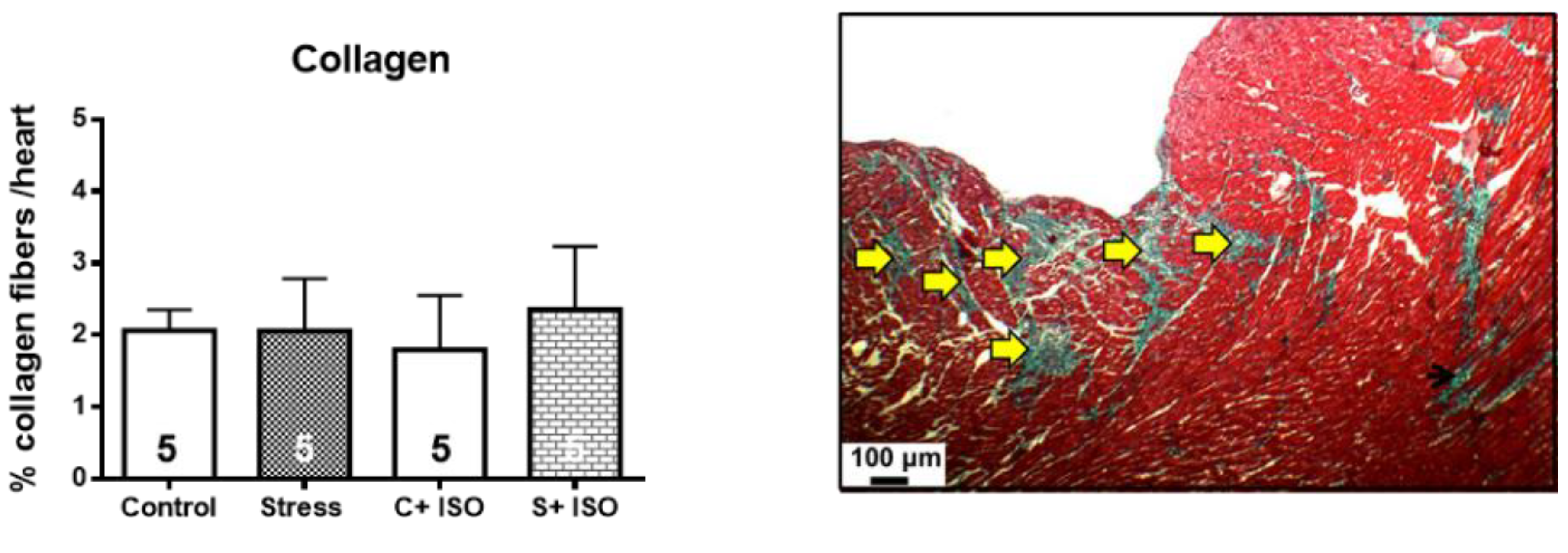
Figure 7.
I. Gestational stress increased the cardiac sensitivity of the female progeny to adrenergic overload. Rats were treated with daily doses of 125 ug/kg weight isoproterenol or saline for control or stressed rats. The graph reflects the percentage of survival of young adult rats after their corresponding treatments. The results correspond to the mean ± mean standard error. Each group consisted of 10 animals. II. The pressure and pulse frequency response recorded in a control rat. III. The admistration of isoproterenol (to a control rat) on the pulse frequency with no change in pressure. IV. The presence of multiples extrasystoles in the stressed rats and, V. the response to isoproterenol on the pressure with a collapse in the pressure after the administration of Isoproterenol (ISO).
Figure 7.
I. Gestational stress increased the cardiac sensitivity of the female progeny to adrenergic overload. Rats were treated with daily doses of 125 ug/kg weight isoproterenol or saline for control or stressed rats. The graph reflects the percentage of survival of young adult rats after their corresponding treatments. The results correspond to the mean ± mean standard error. Each group consisted of 10 animals. II. The pressure and pulse frequency response recorded in a control rat. III. The admistration of isoproterenol (to a control rat) on the pulse frequency with no change in pressure. IV. The presence of multiples extrasystoles in the stressed rats and, V. the response to isoproterenol on the pressure with a collapse in the pressure after the administration of Isoproterenol (ISO).
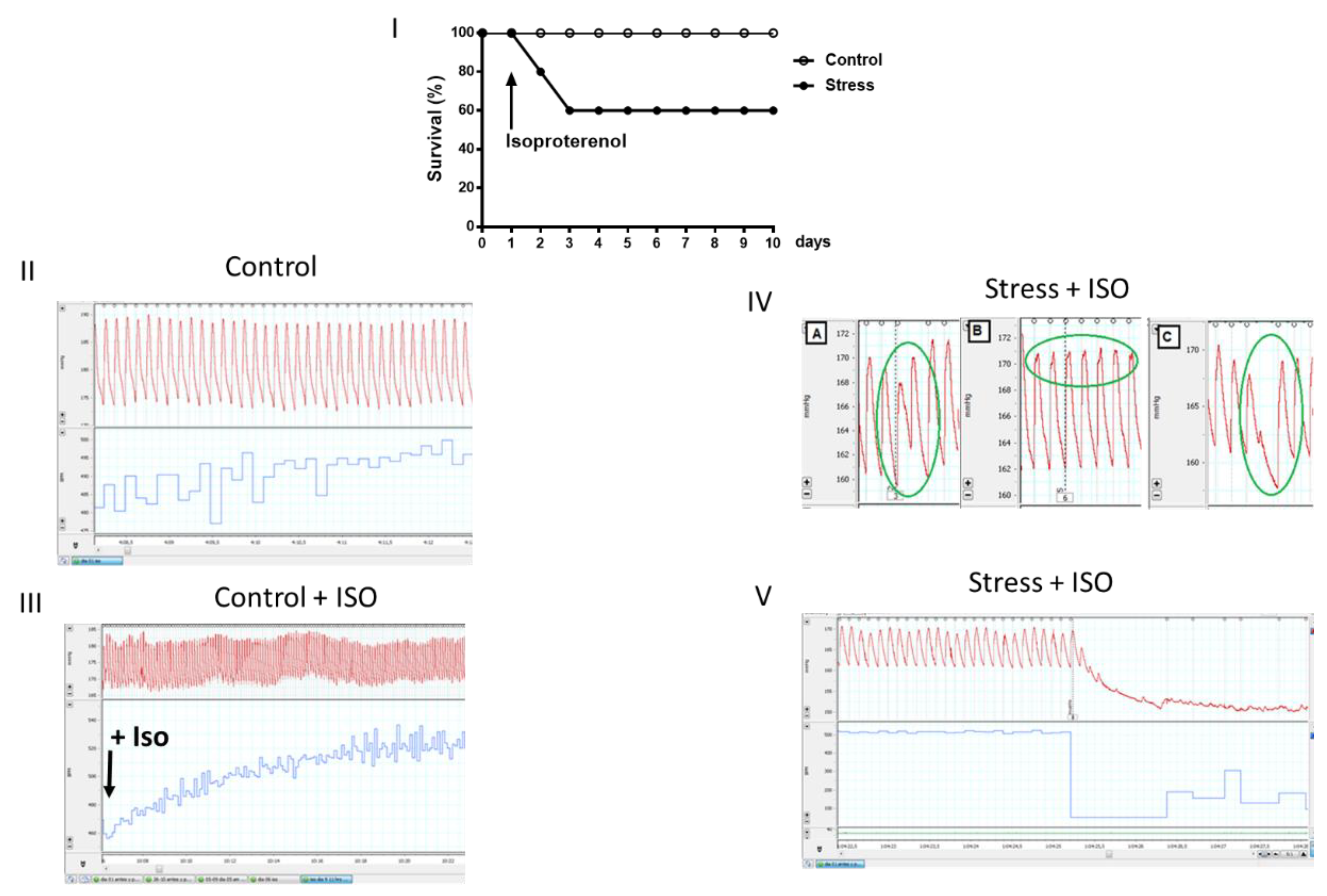
Figure 8.
On the left side is shown the blood glucose concentration after a glucose overload in control and stressed female offspring. Blood samples were obtained by the tail vein after the oral administration of 2 g of glucose/kg body weight, in which we measured the blood concentration at 0, 30, 60, 90 and 120 minutes in thirteen control female rats and twelve female rats. In the same samples, we determined the insulin concentration (Figure 7, middle). In the right side shows the HOMA index for the same rats. The results correspond to the average ± SEM. The number of animals (n=4 in each condition) is indicated in the graph. * p<0.05.
Figure 8.
On the left side is shown the blood glucose concentration after a glucose overload in control and stressed female offspring. Blood samples were obtained by the tail vein after the oral administration of 2 g of glucose/kg body weight, in which we measured the blood concentration at 0, 30, 60, 90 and 120 minutes in thirteen control female rats and twelve female rats. In the same samples, we determined the insulin concentration (Figure 7, middle). In the right side shows the HOMA index for the same rats. The results correspond to the average ± SEM. The number of animals (n=4 in each condition) is indicated in the graph. * p<0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



