Submitted:
06 June 2024
Posted:
07 June 2024
You are already at the latest version
Abstract
The taxonomy of the Fabeae (Vicieae) has long been problematic, but an analysis by Schaefer et al in 2012 gave an exceptionally clear view of the tribe and noted the possibility that some nomenclatural adjustments may be required at some future date. These authors suggested several options, expressing some preferences. However, there has been a recent change to formally accepted names implementing one of these possibilities, but without any additional relevant information. This change seems unjustified and unhelpful. We therefore present an argument for the retention, or re-instatement, of the genera Pisum, Vavilovia and Lens until such time as new data supports this requirement and there is no nomenclatural solution that is both accurate and convenient.
Keywords:
Leguminosae
; systematics
; phylogenetics
; Pisum sativum
; Lathyrus oleraceus
; nomenclature
1. Introduction
Legumes (Leguminoseae, or Fabaceae) are a large and diverse family of angiosperms [1,2] recently expanded and reorganized to include five subfamilies [2]. Most agriculturally important legumes are within the Papilionoideae, and within this subfamily an informally named monophyletic group of genera, the Inverted Repeat Lacking Clade (IRLC) [3,4], includes the majority of temperate legume crop and pasture species.
The IRLC, which includes the tribes Cicereae, Wisterieae, Trifolieae, and Fabeae (Vicieae), is defined as papilionoid legume species lacking one of the two inverted repeats of the plastid genome commonly found in land plants and freshwater green algae (the Viridiplantae). As with the other group of land plants where this type of plastid genome is found (twice in the Pinophyta), this structural rearrangement affects the evolutionary rate of plastid genes usually located within the inverted repeat [5]. Analysis of the phylogenetics of Medicago (Trifolieae) presented evidence for a gain of an IR structure in M. minima [6]. Taken together these observations suggest that phylogenies within the ILRC that depend on plastid genome divergence need to be treated with caution.
The Fabeae (Vicieae) sensu lato includes the genera Vicia, Lens, Lathyrus, Pisum and Vavilovia [7] and a comprehensive phylogenetic analysis of this group was presented by Schaefer et al. [8] and concluded that these genera were not monophyletic, noting that Pisum and Vavilovia were nested within Lathyrus, and that Lens was nested within Vicia and furthermore that a group of Vicia species, belonging to sections Ervum and Ervilla, were early-diverging lineages of the to the entire tribe. Schaefer et al. discussed the taxonomic implications of this phylogeny and suggested two broad possibilities; the first was to generate a multiplicity of new genera and the second was to amalgamate all into a single genus, Vicia. One alternative was to merge Pisum and Vavilovia with Lathyrus, and Lens with Vicia, creating three genera in the tribe, the basal genus comprised of Vicia sections Ervum and Ervilla, Vicia (excluding sections Ervum and Ervilla but including Lens) and Lathyrus broadened to include Pisum and Vavilovia.
This three-genus scheme has been implemented in part [9]; here, we discuss the basis and implications of this taxonomic restructuring and argue for delaying any change in the taxonomy this tribe until more robust information is available.
2. The Fabeae
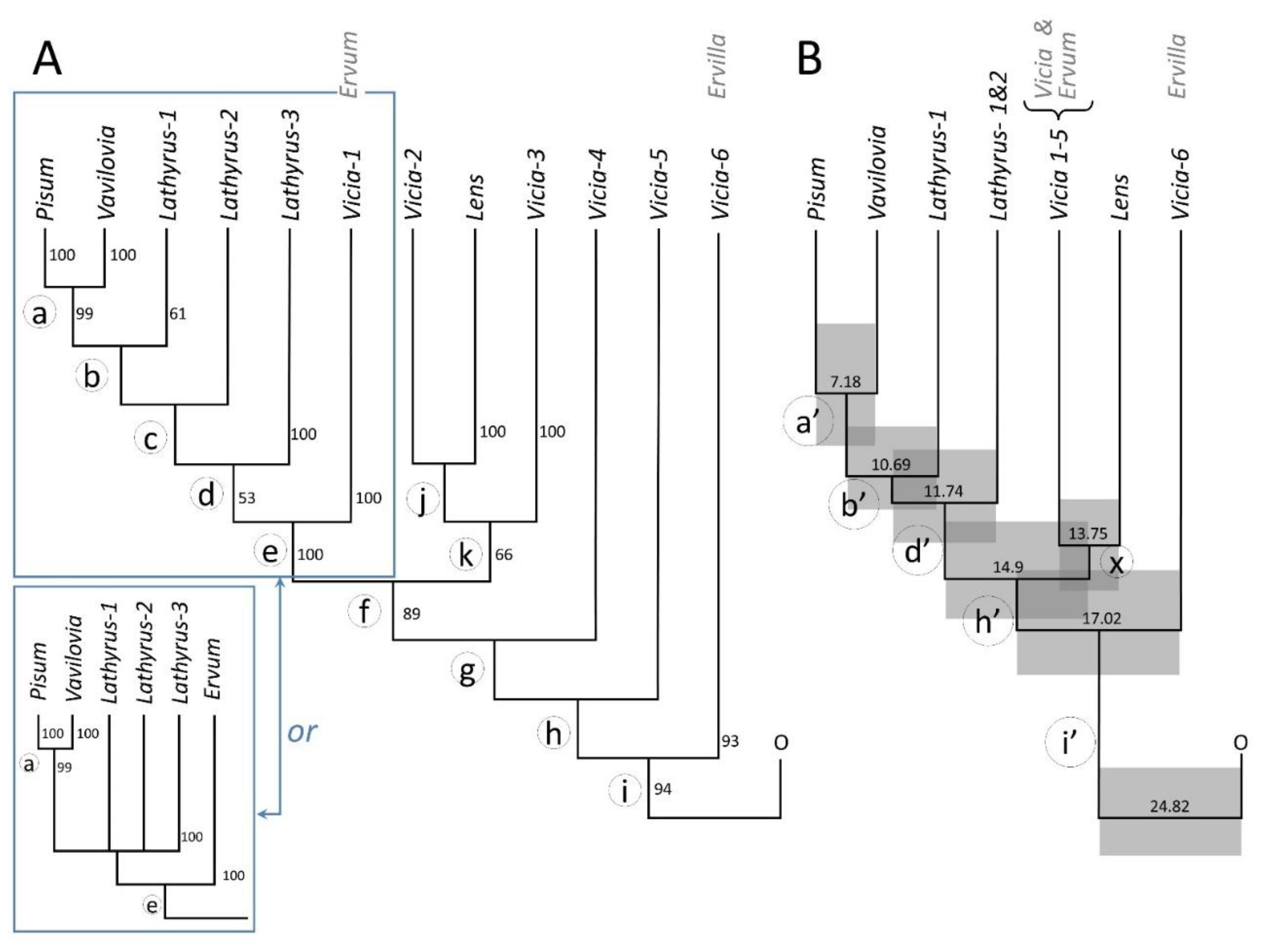
The general features of the phylogenetic tree presented by Schaefer et al. [8] are summarised in Figure 1; the data and interpretations of Schaefer et al. [8] remain the best available description of relationships within the Fabeae.
2.1. The Consensus Maximum Likelihood (ML) Phylogeny
The basal bifurcation in the consensus ML phylogeny separates Vicia-6 (V. cypria, V. lunata, V. sylvatica, V. ervilia, V. articulata, V. monanthos and V. hirsuta) from all other species with a high degree of certainty (93% bootstrap support). It is this bifurcation and the paraphyly of Vicia that argues for the entire tribe being considered to be the monophyletic genus, Vicia. While this interpretation is well supported, it adds no information regarding the internal relationships of the Fabeae as a whole, which is provided by the recognition of additional genera within the Fabeae.
The reinstatement of the genus, Ervilla, corresponding to this group of species, branching at node ‘i’, has the potential to enable the fragmentation of the tribe into more than one genus. These species have not been renamed; for example, the name Vicia ervilia (L.) Willd. is accepted in The World Checklist of Vascular Plants (WCVP) [9]. In addition, the remaining Vicia species are paraphyletic and the other genera are nested within Vicia.
The creation of another genus, Ervum, branching at node ‘e’, has also been proposed [8]. This genus would comprise Vicia pubescens, V. tenuissima and V. tetrasperma (Vicia1 of Figure 1). However, this retains a paraphyletic Vicia, including Lens. The genus Ervum is not recognized by WCVP [9], but Lens culinaris is no longer accepted and has been replaced by Vicia lens (L.) Coss. & Germ. Strangely Lens culinaris subsp. odemensis (Ladiz.) and Lens culinaris subsp. tomentosus (Ladiz.) remain [10].
The Ervum proposal creates a monophyletic group at node ‘d’ in the consensus ML phylogeny and this group includes Pisum and Vavilovia nested within three Lathyrus lineages. However, this group has only 53% bootstrap support. Pisum and Vavilovia are distinct each with 100% bootstrap support, even though the monospecific genus Vavilovia formosa was formerly Pisum formosum. The combined Pisum plus Vavilovia is monophyletic with 99% bootstrap support, and thus distinct from (or within) Lathyrus.
An important issue to resolve is the status of the nodes labelled ‘b’, ‘c’ and ‘d’ in the ML phylogeny. These do not have strong bootstrap support, raising the possibility that the three groups of Lathyrus species may be a monophyletic clade, rather that a paraphyletic grade as in this figure. Supposing that Lathyrus and Pisum plus Vavilovia are two monophyletic clades derived at node ‘d’ (condensing the nodes ‘b’, ‘c’, with less than 50% support, and ‘d’ which musters 53% support), then, in that case, there is no requirement to re-assign any generic status. On the other hand, were there two monophyletic lineages Lathyrus, as implied by the chronogram of Figure 1B,then, in that case, the choice would be whether to elevate one subgroup of Lathyrus to a new genus, or to combine Lathyrus with Pisum plus Vavilovia. Furthermore, it should be noted that the diversity within Pisum and Vavilovia is not fully represented in this tree, both with respect to the number of Pisum subspecies represented [11,12,13] and with respect to the molecular data available. Indeed, whereas Lathyrus accessions were described both by ITS and plastid DNA sequences, most Pisum accessions where only characterized by ITS sequences. Fifteen out of 18 Pisum accessions had either one or no chloroplast sequence, which may have significant effects on the robustness of the phylogenetic relationships inferred, especially if sampling of chloroplast data for Lathyrus is higher.
The majority of Lathyrus species are represented by Lathyrus-1 in Figure 1 and form a monophyletic group, but with 61% bootstrap support; in some trees, some of these species are placed differently; presumably in association with Lathyrus neurolobus. The third group of Lathyrus (L. articulates, L. clymenum and L. ochrus) is itself distinct and monophyletic with 100% bootstrap support in the ML tree. The arrangement of these three Lathyrus groups and the Pisum plus Vavilovia group is uncertain, with at best 53% bootstrap support for node ‘d’ and less than 50% support for nodes ‘b’ and ‘c’, suggesting that these dichotomies may collapse when adding more data and/or taxa for further phylogenetic inferences (Figure 1).
2.2. The Chronogram
There are many useful analyses in Schaefer et al. [8], one of which is a chronogram that attempted to date divergences within the Fabeae. The overall branching structure that relates to the present discussion is given in Figure 1B. There are several striking features of this structure. The first is that the proposed Ervilla group is basally branched, but the age estimate of this divergence overlaps slightly with the divergence of Vicia plus Lens from Lathyrus, Pisum and Vavilovia. All Vicia, including the proposed Ervum, are a monophyletic lineage but distinct from Lens in this analysis [8]. This is consistent with retaining Lens as a separate genus.
In the chronogram, the two minor Lathyrus lineages are distinct from the remaining (and majority of) Lathyrus species, but the age estimate for this divergence overlaps substantially with the estimated age of the divergence of Pisum plus Vavilovia from all Lathyrus. The estimate of the Pisum / Vavilovia divergence time overlaps to some degree with their divergence from the majority of Lathyrus, presumably reflected in the 99% bootstrap support for this clade in the ML tree.
3. Discussion
The phylogenetic analysis of the Fabeae presented by Schaefer et al. [8] is a thorough analysis of this tribe, and remains the best information we have on species and generic relationships in the Fabeae. These authors suggested several possible nomenclatural changes within the tribe that might better reflect the new understanding of species relationships which they uncovered, but did not present a firm and comprehensive set of new species and generic names. No new data have yet become available to lend weight to any of the possible new systems of species and generic names, yet a new system has been partially implemented by WCVP [9] and other authors [14,15,16]. This seems premature and precipitate.
Kenicer and Parsons [10] argue “A major change is that Pisum and Vavilovia are now included in Lathyrus. If an alternative approach had been taken with Pisum and Vavilovia, maintaining them as separate genera, then Lathyrus would have to lose L. clymenum, L. neurolobus and L. ochrus. On balance the inclusion of Pisum and Vavilovia in Lathyrus entails fewer name changes and more accurately reflects the morphology of the group, meaning the inwardly hairy style is a defining character of the genus”. Whether this in fact entails fewer name changes depends on the number of species within Pisum plus Vavilovia. In fact, this change demands at least four name changes (Pisum sativum, P. elatius and P. fulvum plus Vavilovia formosa) compared to renaming four Lathyrus species. This numerical point of view is not the only consideration; a search in Web of Science [16] with the term Pisum sativum returned 41,916 results, for this species, while a search with the term Lathyrus returned 4,834 results, for this genus, reflecting the focus of interest in one of the species to be renamed.
We recognize that taxonomic names should reflect our understanding of species relationships; nomenclature should above all be useful for distinguishing Organismal Taxonomical Units under study. When there are substantial changes to our understanding, a new nomenclatural system is appropriate, and this should recognize due priority as well the value of nomenclatural stability. Pisum has been used as a name since antiquity. However, we see no good evidence that Pisum, Vavilovia (and Lens) need to be renamed. This may become desirable given new knowledge, which is not yet available. If ou when this situation changes, we think it would be wise to implement changes that will minimally disrupt the scientific literature. If the creation of a small number of new genera enables the preservation of Vicia, Lens, Lathyrus, Pisum and Vavilovia, then that would seem a sensible change, as indeed Schaefer et al. [8] proposed for Ervum and Ervilla. Similarly, Kenicer and Parsons [14] presented a diagram of the relationships within the Fabeae with eight taxonomic groups combined in a way that creates four genera.
If future phylogenetic analysis does confirm that there is a monophyletic clade where Pisum and Vavilovia are indeed embedded within Lathyrus, then the creation of a new genus from within Lathyrus might enable the retention of Pisum and Vavilovia; if so this would seem a desirable change.
Author Contributions
T.H.N.E. prepared the first draft of the manuscript. P.S., N.M., J.B., and Y.B.K. contributed to the taxonomic interpretations as presented and edited the text. C.J.C, C.D. and N.C. edited the manuscript and contributed to the discussion.
Funding
T.H.N.E. and N.C. acknowledge long-term investment of the UK Research Infrastructure Biotechnology and Biological Sciences Research Council (UKRI-BBSRC) through National Bioscience Research Infrastructure grant (BBS/E/JI/23NB0001) and the UK Department for Environment, Food, and Rural Affairs (Defra) through the Provision and Maintenance of the Pea Genebank to Facilitate R&D Need grant (C5515). CD acknowledge funding support from UKRI BBSRC (BBS/E/J/000PR799). P.S. acknowledges funding from Grant Agency of Palacky University, PrF-2024-001.
Data Availability Statement
N/A
Acknowledgments
We thank Julie Hofer and Tom Warkentin for useful discussions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lewis, G.P.; Schrire, B.; Mackinder, B.; Lock, M. Legumes of the World. Kew Publishing, Kew, UK. 2005.
- Azani, N.; Babineau, M.; Bailey, C.D.; Banks, H.; Barbosa, A.R.; Pinto, R.B.; Boatwright, J.S.; Borges, L.M.; Brown, G.K.; Bruneau, A.; Candido, E.; Cardoso, D.; Chung, K.; Clark, R.P.; Conceição, A.d.S.; Crisp, M.; Cubas, P.; Delgado-Salinas, A.; Dexter, K.G.; Doyle, J.J.; Duminil, J.; Egan, A.N.; de la Estrella, M.; Falcão, M.J.; Filatov, D.A.; Fortuna-Perez, A.P.; Fortunato, R.H.; Gagnon, E.; Gasson, P.; Rando, J.G.; de Azevedo Tozzi, A.M.G.; Gunn, B.; Harris, D.; Haston, E.; Hawkins, J.A.; Herendeen, P.S.; Hughes, C.E.; Iganci, J.R.V.; Javadi, F.; Kanu, S.A.; Kazempour-Osaloo, S.; Kite, G.C.; Klitgaard, B.B.; Kochanovski, F.J.; Koenen, E.J.M.; Kovar, L.; Lavin, M.; le Roux, M.; Lewis, G.P.; de Lima, H.C.; López-Roberts, M.C.; Mackinder, B.; Maia, V.H.; Malécot, V.; Mansano, V.F.; Marazzi, B.; Mattapha, S.; Miller, J.T.; Mitsuyuki, C.; Moura, T.; Murphy, D.J.; Nageswara-Rao, M.; Nevado, B.; Neves, D.; Ojeda, D.I.; Pennington, R.T.; Prado, D.E.; Prenner, G.; de Queiroz, L.P.; Ramos, G.; Filardi, F.L.R.; Ribeiro, P.G.; de Lourdes Rico-Arce, M.; Sanderson, M.J.; Santos-Silva, J.; São-Mateus, W.M.B.; Silva, M.J.S.; Simon, M.F.; Sinou, C.; Snak, C.; de Souza, É.R.; Sprent, J.; Steele, K.P.; Steier, J.E.; Steeves, R.; Stirton, C.H.; Tagane, S.; Torke, B.M.; Toyama, H.; da Cruz, D.T.; Vatanparast, M.; Wieringa, J.J.; Wink, M.; Wojciechowski, M.F.; Yahara, T.; Yi, T. and Zimmerman, E. A new subfamily classification of the Leguminosae based on a taxonomically comprehensive phylogeny: The Legume Phylogeny Working Group (LPWG). Taxon. 2017, 66, 44-77. [CrossRef]
- Wojciechowski, M.F.; Sanderson, M.J.; Hu, J.M., Evidence on the monophyly of Astragalus (Fabaceae) and its major subgroups based on nuclear ribosomal DNA ITS and chloroplast DNA trnL intron data. Syst. Bot. 1999, 24, 409–437. [CrossRef]
- Duan, L.; Li, S-J.; Su, C.; Sirichamorn, Y.; Han, L-N.; Ye, W.; Lôc, P.K.; Wen, J.; Compton, J.A.; Schrire, B.; Nie, Z-L.; Chen, H.K. Phylogenomic framework of the IRLC legumes (Leguminosae subfamily Papilionoideae) and intercontinental biogeography of tribe Wisterieae. Molecular Phylogenetics and Evolution. 2021, 163, 107235. [CrossRef]
- Zhu, A.; Guo, W.; Gupta, S.; Fan, W.; Mower, J.P. (2016), Evolutionary dynamics of the plastid inverted repeat: the effects of expansion, contraction, and loss on substitution rates. New Phytol, 2016, 209, 1747-1756. [CrossRef]
- Choi, I-S.; Jansen, R.; Ruhlman, T. Lost and Found: Return of the Inverted Repeat in the Legume Clade Defined by Its Absence, Genome Biology and Evolution. 2019, 11, 1321–1333. [CrossRef]
- Kupicha, F.K., Vicieae. In "Advances in Legume Systematics". Eds. R.M. Polhill and P.M. Raven. pp. 377-381. Royal Botanic Gardens, Kew. (1981).
- Schaefer, H.; Hechenleitner, P.; Santos-Guerra, A.; Menezes de Sequeira, M.; Pennington, R.T.; Kenicer, G. and Carine, M.A. Systematics, biogeography, and character evolution of the legume tribe Fabeae with special focus on the middle-Atlantic island lineages. BMC Evol. Biol. 2012, 12, 250. [CrossRef]
- Royal Botanic Gardens, Kew. The World Checklist of Vascular Plants (WCVP): Fabaceae. (2023) Checklist dataset https://doi.org/10.15468/mvhaj3 accessed via GBIF.org on 09/05/2024.
- Ferguson, M.E.; Maxted, N.; van Slageren, M.V. and Robertson, L.D. () A re-assessment of the taxonomy of Lens Mill. (Leguminosae, Papilionoideae, Vicieae). Bot. J. Linn. Soc. 2000, 133: 41-59. [CrossRef]
- Hellwig, T., Abbo, S., Sherman, A., Coyne, C., Saranga, Y., Lev-Yadun, S., Main, D., Zheng, P. and Ophir, R. Limited divergent adaptation despite a substantial environmental cline in wild pea, Mol. Ecol. 2020, 29, 4322–4336.. [CrossRef]
- Timo Hellwig, Shahal Abbo, Amir Sherman, Ron Ophir, Prospects for the natural distribution of crop wild-relatives with limited adaptability: The case of the wild pea Pisum fulvum, Plant Science. 2021, 310, 110957. [CrossRef]
- Hellwig, T., Abbo, S. and Ophir R. Phylogeny and disparate selection signatures suggest two genetically independent domestication events in pea (Pisum L.) Plant J. 2022, 110, 419–439. [CrossRef]
- Kenicer G. and Parsons R. Lathyrus the complete guide Royal Horticultural Society, Peterborough (2021).
- Rix, M., Nesbitt, M. & King, C. 1063. Lathyrus oleraceus Lam. Curtis's Botanical Magazine, 2023, 40, 197–205. Available from: . [CrossRef]
- Kosterin, O. E. Abyssinian pea (Lathyrus schaeferi Kosterin nom. Nov. pro Pisum abyssinicum A. Br.) is a problematic taxon. Vavilovskii Zhurnal Genetiki i Selektsii Vavilov J. Genet. Breed. 2017, 21, 158–169. [CrossRef]
- https://www.webofscience.com/wos/alldb/basic-search accessed 19/05/20024.
Figure 1.
(A) Summary of the consensus Maximum Likelihood phylogeny of the Fabeae from Additional file 12, Figure S10 of Schaefer et al. [7]. Bootstrap values are to the right and, for reference, nodes are indicated by a letter. Lathyrus-1 = most Lathyrus; Lathyrus-2 = L. neurolobus; Lathyrus-3 = L. articulates, L. clymenum and L. ochrus; Vicia-1 = Vicia pubescens, V. tenuissima and V. tetrasperma; Vicia-2 = most Vicia; Vicia-3 = V. malosana, V. claessensii, V. paucifolia, V. iranica and V. subvillosa; Vicia-4 = V. crocea; Vicia- 5 = V. gigantea, V. nigricans and V. menziesii; Vicia-6 = V. cypria, V. lunata, V. sylvatica, V. ervilia, V. articulata, V. monanthos and V. hirsuta. (B) Summary Chronogram of the Fabeae for relevant nodes, from Additional file 11, Figure S9 of Schaefer et al. [7]. Numbers are age estimates in Myr. The grey boxes indicate the 95% confidence intervals of the age estimates. In part B, nodes indicated by letters with a prime refer to the equivalent node, with the same letter, as shown in part A and as a letter when there is no corresponding bifurcation in the two source data files. Names in grey were suggested [8] new generic names.
Figure 1.
(A) Summary of the consensus Maximum Likelihood phylogeny of the Fabeae from Additional file 12, Figure S10 of Schaefer et al. [7]. Bootstrap values are to the right and, for reference, nodes are indicated by a letter. Lathyrus-1 = most Lathyrus; Lathyrus-2 = L. neurolobus; Lathyrus-3 = L. articulates, L. clymenum and L. ochrus; Vicia-1 = Vicia pubescens, V. tenuissima and V. tetrasperma; Vicia-2 = most Vicia; Vicia-3 = V. malosana, V. claessensii, V. paucifolia, V. iranica and V. subvillosa; Vicia-4 = V. crocea; Vicia- 5 = V. gigantea, V. nigricans and V. menziesii; Vicia-6 = V. cypria, V. lunata, V. sylvatica, V. ervilia, V. articulata, V. monanthos and V. hirsuta. (B) Summary Chronogram of the Fabeae for relevant nodes, from Additional file 11, Figure S9 of Schaefer et al. [7]. Numbers are age estimates in Myr. The grey boxes indicate the 95% confidence intervals of the age estimates. In part B, nodes indicated by letters with a prime refer to the equivalent node, with the same letter, as shown in part A and as a letter when there is no corresponding bifurcation in the two source data files. Names in grey were suggested [8] new generic names.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



