Submitted:
04 June 2024
Posted:
06 June 2024
You are already at the latest version
Abstract
(1) Background: Hepatitis B virus (HBV) sequencing data are important for monitoring HBV evolution. We aimed to molecularly characterize HBV sequences from participants with HBV surface antigen (HBsAg+) positive serology and occult hepatitis B infection (OBI+). (2) Methods: We utilized archived plasma samples from a cohort of people living with (PLWH) in Botswana. HBV DNA was sequenced, genotyped and analyzed for mutations. We compared mutations from study sequences to those from previously generated HBV sequences in Botswana. The impact of OBI-associated mutations on protein function was assessed using the Protein Variation Effect Analyzer. (3) Results: Sequencing success was higher in HBsAg+ than OBI+ samples [86/128 (67.2%) vs 21/71 (29.2%)]. Overall, 93.5% (100/107) sequences were genotype A1, 2.8% (3/107) were D3 and 3.7% (4/107) were E. We identified 13 escape mutations in 18/90 (20%) sequences with HBsAg coverage, with K122R having the highest frequency. The mutational profile of current sequences differed from previous Botswana HBV sequences, suggesting possible mutational changes over-time. Mutations deemed to have an impact on protein function were tpQ6H, surfaceV194A and preCW28L. (4) Conclusions: We characterized HBV sequences from PLWH in Botswana. Escape mutations were prevalent and were not associated with OBI. Longitudinal HBV studies are needed to investigate HBV natural evolution.
Keywords:
Hepatitis B virus
; Genotypes
; Escape mutations
; people living with HIV
; Botswana
; Africa
1. Introduction
Hepatitis B virus (HBV) infection remains a significant global health concern particularly in Africa, which has the highest HBV prevalence globally at 5.8% [1]. In Botswana, the prevalence of hepatitis B surface antigen (HBsAg) varies by region, reaching levels as high as 22% in certain areas of the country [2]. Only three studies have reported occult hepatitis B infection (OBI) prevalence in Botswana, with a range from 6.6% to 33% in adults [2,3,4]. OBI is defined as the presence of replicative competent HBV deoxyribonucleic acid (DNA) in the blood and/or liver of individuals testing negative for HBsAg [5].
Understanding the genetic diversity of HBV within specific geographic locations is important for devising effective prevention and control strategies as HBV genotypes have clinical relevance. HBV genotypes are associated with vaccine efficacy [6], treatment response [7] , tendency of chronicity [8], HBsAg and hepatitis B e antigen (HBeAg) seroconversion [9]. Previous studies have identified HBV subgenotypes A1, D2, D3, and genotype E in Botswana [3,10,11,12,13]. Notably, the prevalence of subgenotype A1 varies among different demographic groups [3,10,11,14]. There is need for HBV surveillance and molecular characterization efforts especially in settings with high HIV prevalence and widespread antiretroviral treatment (ART) use, in which HBV drug resistance can be common [15]. Furthermore, HBV genomics are important in studying vaccine and treatment response, as well as transmission dynamics within the country.
OBI is not reported in national, regional, and global reports however it has clinical relevance. OBI was determined to be an independent risk factor for hepatocellular carcinoma (HCC) in one study [16]. Drug resistance associated mutations have been identified in participants with OBI [15]. Furthermore, infants born to mothers with positive HBsAg (HBsAg+) were diagnosed with OBI in one study [17]. One mechanism postulated to lead to the OBI phenotype is the presence of mutations that impair HBsAg detection. Some OBI associated mutations in different HBV open reading frames (ORFs), (pres1, pre2, surface, core, pre-core, X, and the polymerase domains) have been identified and studied in Botswana and South Africa [18,19]. The impact of these mutations on protein function was assessed using available online in silico tools as screening methods for potential candidates of functional in vitro studies. The Protein Variation Effect Analyzer (PROVEAN) was more accurate than other tools being studied [18,19].
In Botswana, the HBsAg positivity remains high, and the OBI prevalence is 3 to 4 times that of HBsAg positivity as prior studies have reported an adult OBI prevalence of 6.6% to 33% [2,3,4]. Therefore, the genetic diversity of HBV in both HBsAg+ and OBI participants needs to be further studied. We aimed to molecularly characterize HBV sequences from people living with HIV (PLWH) who tested positive for HBsAg and OBI, and to determine the impact of OBI-associated mutations on protein function in a cohort of PLWH in Botswana.
2. Materials and Methods
2.1. Study Population
Archived plasma samples from participants in the Botswana Combination Prevention Project (BCPP), that had previously tested positive for HBsAg (HBsAg+), and OBI (OBI+) were used [20]. Details of the BCPP study are described elsewhere [21,22]. Briefly, the BCPP study was a pair-matched cluster randomized study that enrolled 12,610 all consenting adults residing in a random sample of ~20% of households in 30 geographically dispersed villages throughout Botswana between the years 2013 and 2018. The main aim of the BCPP study was to assess if a combination of HIV prevention strategies would reduce HIV incidence at a community level compared to the standard of care. At baseline 3,596 BCPP study participants were PLWH and (83% of whom knew their HIV status) [21]. Our study was approved by the Human Research Development Committee (HRDC) at the Botswana Ministry of Health (HRDC number: 01028).
2.2. Laboratory Procedures
HBV screening has been described in our previous report [2]. Briefly, available plasma samples from 3304/3596 (91.9%) of PLWH in the BCPP cohort were screened for HBsAg and total core-antibodies (anti-HBc). HBsAg+ samples were further screened for HBeAg and immunoglobulin M core antibodies (anti-HBc IgM). HBV viral load was quantified for HBsAg+ samples with sufficient sample volume and performed in samples that tested negative for HBsAg to determine OBI prevalence using the Roche COBAS Ampliprep/Taqman Analyzer [2].
The QIAamp DNA Blood Mini kit (Qiagen, Hilden, Germany) was used to extract DNA from 200μL of HBsAg+ and OBI+ plasma samples according to the manufacturer’s protocol with a final elution volume of 30μL. HBV DNA was amplified using tiling primers adopted from Choga’s protocol [23] (Table S1). Briefly, two 10μM pools of tiling primers were prepared. A master mix for each primer pool with 5μL of DNA template was prepared, and HBV DNA was amplified using a protocol from our previous reports [15,24]. After amplification, these PCR products were combined, and library preparation followed suite [15,23,25]. The library was loaded into flow cells version R9.4.1 (Oxford Nanopore Technologies, Oxford, UK) and the GridION platform was used for sequencing.
2.3. Sequence Analyses
2.3.1. Genotypic and Mutational Analysis
Generated FASTQ files were uploaded into Genome Detective 1.132/1.133 for reference-based assembly of HBV [26] (last accessed 20 April 2023). The generated consensus sequences were downloaded and further imputed and assessed in AliView ver. 1.26 for viewing, trimming and alignment [27]. Geno2pheno (https://hbv.geno2pheno.org) (last accessed 11 December 2023), was used to assign HBV genotypes/subgenotypes. Genotypes were confirmed using phylogenetic analysis. BCPP generated sequences and reference HBV sequences from GenBank were used to construct a phylogenetic tree of the complete surface ORF using a Bayesian Markov chain Monte Carlo (MCMC) in the Bayesian Evolutionary Analysis by Sampling Trees (BEAST) v1.8.2 (BEAST Developers) program with a chain length of 100,000,000 and sampling every 10,000 generations. The analysis utilized an uncorrelated log-normal relaxed molecular clock, the Hasegawa, Kishino, and Yano (HKY) model, and the general time reversible model with gamma distributed rates of variation among sites and a proportion of invariable sites (GTR+G+I). Tracer v1.7 (BEAST Developers) was used to visualize results and confirm chain convergence. Tree Annotator v1.7.3 (BEAST Developers) was used to choose the maximum clade credibility tree after a 10% burn-in. The mutational profile of sequences generated form this study was compared to mutations in sequences generated in previous Botswana studies that were predominantly from PLWH (2009-2015) [3,10,11,28].
2.3.2. Impact of Occult-Associated Mutations on Protein Function
Sequences with a depth of >100 were used for this analysis. The Protein Variation Effect Analyzer (PROVEAN), available at http://provean.jcvi.org/index.php (accessed 24 April 2024) was used to determine the impact of occult-associated mutations on protein function Occult-associated mutations were defined as mutations identified only in OBI+ sequences and those that were overrepresented in OBI+ sequences versus HBsAg+ sequences.
3. Results
3.1. Participants Clinical Characteristics
Table 1 summarizes the clinical characteristics of participants whose plasma samples were used in the analysis. Most participants were female (66.4%) and had a median age of 43 (IQR: 36 – 49). Most participants had a low HBV viral load of <2000IU/ml (58.9%). Approximately 94.4% of participants were on antiretroviral therapy (ART) and were mostly on tenofovir disoproxil fumarate (TDF) containing regimen (61.6%). The TDF regimen also had emtricitabine (FTC) for all participants except for one who was on a dolutegravir/TDF regimen. The majority of participants had undetectable HIV-1 RNA (<40copies/mL) (88.8%). Median duration time on ART was 7 years (IQR: 4.7 – 9.9) Table 1.
3.2. Sequencing Success
Figure 1 shows the total number of sequences generated. Sequencing was attempted on 128 samples out of the 271 HBsAg+ samples and the success rate was 67.2% (86/128). Success rate for OBI+ was 29.2% (21/72). Samples that were successfully sequenced had HBV viral loads ranging from target not detected (TND) to >1.7 x 108 IU/mL.
In total, 27 out of the 30 BCPP study sites contributed HBV sequences to this analysis (Figure S2). Samples from participants residing in Mmadinare, Molapowabojang and Otse (BCPP study sites) were not successfully sequenced. Most sequences were generated from participants in the Central district with 50 sequences, followed by the North-West district with 26 sequences. Kgatleng district had the least number of sequences generated (n=5) (Figure 2).
3.3. Genotypic Analysis
Figure 3 shows the clustering of the study sequences with reference sequences. All previous HBV sequences from Botswana clustered together and BCPP sequences clustered randomly (Figure 3). Overall, 93.5% (100/107) sequences were genotype/subgenotype A1, 2.8% (3/107) were D3 and 3.7% (4/107) were E. Among the HBsAg+ sequences, 93.0% (80/86) sequences were genotype/subgenotype A1, 3.5% (3/80) were D3 and 3.5% (3/80) were E. For OBI sequences, 95.2% (20/21) sequences were genotype A1 and 4.8% (1/21) were E.
3.4. Mutational Analysis
3.4.1. Escape Mutations
A total of 13 escape mutations were detected in 20% (18/90) of sequences with surface gene coverage (Table 2). surfaceK122R had the highest frequency in 9/18 (50%) of participants with escape mutations. surfaceT114S, surfaceS114L, surfaceC139R, surfaceN146S and surfaceC147Y are associated with impaired virion secretion. Vaccine escape mutations (surfaceG130C, surfaceN131T, surfaceT121N) were identified in four participants. We also identified mutations that may impact HBsAg detection (surfaceT118M, surfaceC121R, surfaceK122R, surfaceG130C) (Table 2).
3.4.2. Comparison of Botswana reference HBV sequences and BCPP HBV sequences.
For all downstream analyses, only subgenotype A1 sequences were used as they constituted >93% of the sequences. Table 3 and Table 4 show previously generated Botswana sequences (REFERENCE-unique) and BCPP unique mutations with a prevalence of >20%. The full list of mutations is shown in Tables S3 and S4. BCPP sequences tended to have more unique mutations in all ORFs with a high prevalence (>20%) that were not observed in previous Botswana sequences. Some amino acid substitutions unique to BCPP sequences had a much higher prevalence, such as preCV17F (53.6%), xP33S (42.2%), and surfaceI195M (55.7%). Among mutations that were identified in both sets of sequences, the prevalence of pres2A7T and pres2A11T was higher in the BCPP sequences (19.9% and 10.3% vs 5.3% for both mutations in the reference sequences). However, pres2T38I was lower in the BCPP sequences (30.3% vs 41%). For the transcriptional transactivator protein (HBx), the prevalence of xG22S, xA21T and xS46P was higher in the BCPP sequences than in the reference sequences (43.8%, 28.1%, and 67.2% vs 8.3%, 8.3% and 41.7%). In the surface protein, surfaceN131T and surfaceV194A had a lower prevalence in the BCPP sequences compared to the reference sequences (2.2% and 7.7% vs 37.5% and 14.8%) while the opposite was observed for surfaceK122R (10.1% vs 2.3%).
In Table 4, we report mutations specific to the polymerase domains. It is noticeable that resistance associated mutations (rtM204V, rtL180M, rtV173L) were unique to the BCPP cohort. In the same RT region, some mutations had a noticeably higher prevalence in the reference sequences as compared to the BCPP sequences. These are rtV7A (40.0% vs 16.3%), rtL53I (35.0% vs 11.4%), rtH122N (35.0% vs 2.4%), rtN332S (39.3% vs 15.9%) and rtQ333K (44.4% vs 14.5%). In the terminal protein (TP) region, tpH182Q had a noticeably higher prevalence in the BCPP cohort than in the reference sequences (21.7% vs 5.1%). spY86H, spS125N and spS129N in the spacer domain occurred more frequently in the BCPP sequences than the reference sequences. In the RNase H domain, RNaseHY116F was observed at a much higher prevalence in the BCPP cohort compared to the reference sequences (30.0% vs 8.3%).
3.4.3. Impact of Occult-Associated Mutations on Protein Function
For this analysis, we focused on sequences that had a depth of >100 and we identified mutations that were in sequences isolated from OBI participants only (coreT142S, tpE88R, tpQ6H, rtM250L, preCW28L). Other mutations were overly represented in OBI+ sequences compared to HBsAg+ sequences. surfaceV194A and surfaceS55P appeared in 3/13 (23.1%) OBI participants each versus 1/53 (1.9%), HBsAg participant each, p-value 0.004. Using PROVEAN, three mutations were deemed deleterious, that is were deemed to affect protein function negatively: tpQ6H, surfaceV194A and preCW28L.

Table 5.
Impact of mutations identified only/overrepresented in participants with OBI.
| ORF | Mutation | PROVEAN Prediction |
|---|---|---|
| Core | coreT142S | Neutral |
| Terminal protein | tpE88R | Neutral |
| tpQ6H | Deleterious | |
| Surface | surfaceV194A | Neutral |
| surfaceS55P | Deleterious | |
| Reverse transcriptase | rtM250L | Neutral |
| Precore | preCW28L | Deleterious |
ORF; open reading frame, PROVEAN; Protein Variation Effect Analyzer.
4. Discussion
In this study, we identified HBV subgenotypes A1, D3, and E across a wide geographic area in Botswana, with subgenotype A1 representing more than 93% of all sequences. We also report the mutational profile of HBV in the Botswana population including mutations with deleterious impact on protein function in participants with OBI.
Our findings are consistent with prior HBV studies in Botswana which identified the same subgenotypes however with varying genotype prevalence [3,10,11]. We also report immune, vaccine, and diagnostic escape mutations in this population, some of which have been identified in other populations in Botswana [10,11,28]. surfaceK122R was the most prevalent escape mutation and was identified only among participants with HBsAg positive serology. This mutation is associated with decreased HBsAg expression and HBsAg detection failure [33,34,35], however it was not detected among the OBI samples. We did not perform quantitative HBsAg ELISA which could have revealed HBsAg levels in samples with these mutations compared to those without. We also identified known mutations (surfaceT118M, surfaceN146S), and uncharacterized mutations at positions associated with immune escape (surfaceC121R, surfaceQ129C, surfaceG130C,) in OBI participants although these mutations are not unique to individuals with OBI in other studies [30,41,45,46]. All these mutations were identified in the major hydrophilic region (MHR) (position 99 to 169) of the HBsAg with the majority of these being found in the “α” determinant of the MHR (position 124 to 147) which is a major cluster of antigenic epitopes [47]. We also identified vaccine escape mutations (VEMs) in 16.7% of participants with escape mutations, which is a cause for concern as these may counter-act vaccination efforts in the country. This was at position 131 of the surface region also in the “α” determinant of the MHR known for mutations that allow the virus to evade vaccine induced immune response [48]. We have identified a potential vaccine escape mutation, G130C which was reported as a novel mutation in 2017 [49]. Mutations at position 130 are reported to be vaccine escape mutations and have been isolated in vaccinated individuals [38,39].
There was a change in mutation patterns between sequences previously generated in Botswana (2009-2015) [3,10,11,28] and sequences we generated in the BCPP study (2013–2018). For example, the RT region of BCPP sequences harbors more drug resistance associated mutations than the reference sequences. Over 90% of PLWH among the BCPP participants were ART-experienced while in the previous studies, participants were mostly ART-naïve [3,10,11,28]. Due to the overlapping pattern of the HBV genome, some of the mutations observed in the RT region of the polymerase affected the surface region. Position surfaceI195 corresponds to the rtM204 [50], therefore its prevalence is higher in BCPP sequences compared to previous Botswana sequences. Furthermore, surfaceE164D is known to alter HBsAg antigenicity and tends to occur with surfaceI195M, as observed in our study and a previous study [51].
Most of the mutations identified in our study are uncharacterized. Other mutational variations of interest between BCPP and previously generated Botswana sequence are the xP33S in the X region. This mutation was only observed in the BCPP sequences and had not been previously identified in Botswana. It is a B-cell epitope mutation that has been shown to result in increased endoplasmic reticulum (ER) stress [52] and reduced protein stability in combination with other mutations [53]. There were some mutations that were common to both sequence datasets; however, they were more prevalent in the BCPP sequences. For example, the xG22S, which is reported to be an HCC-related HBx mutation [54]. BCPP sequences also had the xT36A, a functionally characterized HCC-associated mutation at >20% prevalence. This mutation is reported to enhance viral genome integration into the host cell resulting in insertion mutations and a 3’-terminal truncation of HBx [55,56]. While previously generated Botswana sequences and the BCPP sequences were generated from different parts of the country, we cannot rule out the possibility that more people are progressing to chronicity and HCC in the population.
To further elucidate on OBI-associated mutations, we used a freely available online tool, PROVEAN, previously shown to be more accurate in predicting the impact of functionally characterized HBV mutations on protein function than other prediction tools [18,19]. This analysis also allows for the selection of mutations that could be candidates for further in vitro studies. Three OBI associated mutations were deemed deleterious: tpQ6H, surfaceV194A and preCW28L. The tpQ6H mutation has not been characterized however it falls within the N-terminal helices of the TP. Mutations and deletions in this subdomain were shown to impact RNA packaging, DNA synthesis and protein priming [57]. preCW28L has not been characterized however a stop codon at this position has been identified in a participant with OBI [58] and in patients with HCC in a much older study [59]. Other surface mutations (surfaceS55P and surfaceV194A) were overrepresented in OBI+ sequences compared to HBsAg+ sequences, and other studies also show that these mutations are not unique to OBI+ sequences [30,60]. It is worth exploring the HBsAg levels in HBsAg+ and OBI+ samples with these mutations. surfaceS55P has not been functionally characterized, while surfaceV194A is a well know mutation associated with decreased extracellular HBsAg levels [61].
Our analysis focused on the differences between sequences from HBsAg+ and OBI+ infection, but it could be that the host factors have a key role to play on the OBI phenotype. Host factors such as immune response, human leukocyte antigen classA2 and interleukin-10 have been associated with OBI persistence [31,62,63]. Some studies attribute this phenotype to viral factors such as epigenetic control mechanisms [64,65] and mutations [18,19,66] that lead to reduced HBsAg expression and DNA replication..
A strength of the study is that we used samples from the BCPP study which recruited participants from different communities in Botswana. However, we analyzed sequences from only 27 out of the original 30 BCPP study sites due to insufficient volumes and the low sequencing success rate. Sequencing success was generally low, especially for OBI+ samples, however, this study still provides the largest number of HBV sequences in Botswana. We generated HBV sequences from participants with a target not detectable viral load result, a group usually excluded during sequencing. Escape mutations, surfaceT114S, surfaceN131T and surfaceK122R were identified in these participants which shows that there may be an underrepresentation of mutations in different reports that may not sequence participants with these viral load results. There is need however, to adopt a nucleic acid testing assay with a lower limit of detection than the one used in our current study. A limitation of the study is that participants were all PLWH predominantly on ART which limits generalizability to the general Botswana population. We make comparisons between previously generated Botswana sequences and the BCPP sequences, which comes with a limitation as these datasets are not from the same communities. Therefore, we cannot rule out the impact of host-genetic factors which could limit the conclusions in the differences we note. However, the mutational profiles of these sequences are fully documented in this study.
5. Conclusions
We have molecularly characterized HBV from PLWH in Botswana and identified that subgenotype A1 is predominant countrywide. We have also reported vaccine escape mutations which shows the importance of periodic monitoring of circulating HBV strains in the population. It is essential to generate HBV sequencing data to monitor the evolution of HBV and the emergence of mutations that could evade immunity and vaccines, potentially affecting HBV prevention and management strategies.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: HBV primers used for sequencing PCR; Figure S2: Number of sequences generated per BCPP communities in the different Botswana districts; Table S3: List of mutations in HBV ORFs; Table S4: List of mutations in HBV Pol domains.
Author Contributions
Conceptualization, Bonolo Phinius, Motswedi Anderson, Margaret Mokomane, Irene Gobe and Simani Gaseitsiwe; Data curation, Bonolo Phinius; Formal analysis, Bonolo Phinius and Wonderful Choga; Funding acquisition, Motswedi Anderson, Joseph Makhema, Roger Shapiro, Shahin Lockman, Rosemary Musonda, Sikhulile Moyo and Simani Gaseitsiwe; Investigation, Bonolo Phinius; Methodology, Bonolo Phinius, Tsholofelo Ratsoma, Basetsana Phakedi and Gorata Mpebe; Project administration, Bonolo Phinius; Resources, Joseph Makhema, Roger Shapiro, Shahin Lockman, Sikhulile Moyo and Simani Gaseitsiwe; Supervision, Motswedi Anderson, Margaret Mokomane, Irene Gobe and Simani Gaseitsiwe; Validation, Wonderful Choga and Simani Gaseitsiwe; Visualization, Wonderful Choga and Lynnette Bhebhe; Writing – original draft, Bonolo Phinius; Writing – review & editing, Bonolo Phinius, Wonderful Choga, Motswedi Anderson, Margaret Mokomane, Irene Gobe, Tsholofelo Ratsoma, Basetsana Phakedi, Gorata Mpebe, Lynnette Bhebhe, Tendani Gaolathe, Mosepele Mosepele, Joseph Makhema, Roger Shapiro, Shahin Lockman, Rosemary Musonda, Sikhulile Moyo and Simani Gaseitsiwe.
Funding
This work was supported by the Wellcome Trust (grant number 218770/Z/19/Z). W. T. C., S. M., and S. G. are partly supported through the Sub-Saharan African Network for TB/HIV Research Excellence (SANTHE 2.0) from the Bill & Melinda Gates Foundation (INV-033558). S. G. and B.B.P are supported by the Fogarty International Center at the US National Institutes of Health (D43 TW009610). B. B. P., R. M., and S. M. are also supported by Trials of Excellence in Southern Africa (TESAIII), which is part of the EDCTP2 program supported by the European Union (grant number CSA2020NoE-3104 TESAIII). S. L., R. S. and S.M. received support from the NIH (award numbers K24 AI131928, K24 AI131924 and K43 TW012350 respectively). S.G and W.T.C are supported partly by NIH (award number 1G11TW012503). B. B. P. and S. G. were supported by the National Institutes of Health (NIH) Common Fund, award number U41HG006941 (H3ABioNet). H3ABioNet is an initiative of the Human Health and Heredity in Africa Consortium (H3Africa) program of the African Academy of Science. BCPP was supported by the United States President’s Emergency Plan for AIDS Relief (PEPFAR) through the Centers for Disease Control and Prevention (CDC, cooperative agreements U01 GH000447 and U2G GH001911). The views and opinions in this manuscript represent are solely the authors and do not represent the official position of the funding agencies. The funders had no role in the design, conduct of the study and decision to publish the results.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by Human Research Development Committee (HRDC) at the Botswana Ministry of Health (HRDC number: 01028).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the original study. Only samples from participants who consented for future research were used.
Data Availability Statement
The datasets [generated/analyzed] for this study are available upon request.
Acknowledgments
The authors thank the Botswana Prevention Combination Project study participants, Dikgosi and other community leaders, the clinic staff, District Health Management Teams, and Community Health Facilities at study sites; the Ya Tsie Study Team at the Botswana Harvard Health Partnership, the Harvard T. H. Chan School of Public Health, the Centers for Disease Control and Prevention (CDC) Botswana, CDC Atlanta, and the Botswana Ministry of Health. The authors also thank those who served on the Ya Tsie Community Advisory Board, Laboratory Staff, and Management of Botswana Harvard HIV Reference Laboratory.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A
Table S1.
HBV primers used for sequencing PCR.
| Primer Name | Sequence | Positions |
|---|---|---|
| SC_1_LEFT | TTCCACCAAGCTCTGCAAGATC | 11 - 32 |
| SC_1_RIGHT | AGAGGAATATGATAAAACGCCGCA | 384-407 |
| SC_2_LEFT | CTCCAATCACTCACCAACCTCC | 325-346 |
| SC_2_RIGHT | AAAGCCCTACGAACCACTGAAC | 692-713 |
| SC_3_LEFT | AAATACCTATGGGAGTGGGCCT | 632-653 |
| SC_3_RIGHT | TTGTGTAAATGGAGCGGCAAAG | 1655-1676 |
| SC_4_LEFT | AGAAAACTTCCTGTTAACAGACCTATTG | 949-976 |
| SC_4_RIGHT | GGACGACAGAATTATCAGTCCCG | 1326-1348 |
| SC_5_LEFT | TCCATACTGCGGAACTCCTAGC | 1265-1286 |
| SC_5_RIGHT | TGTAAGACCTTGGGCAGGATTTG | 1632-1654 |
| SC_6_LEFT | CTTCTCATCTGCCGGTCCGTGT | 1559-1580 |
| SC_6_RIGHT | AGAAGTCAGAAGGCAAAAACGAGA | 1947-1970 |
| SC_7_LEFT | GGCTTTGGGGCATGGACATT | 1890-1909 |
| SC_7_RIGHT | ATCCACACTCCGAAAGAGACCA | 2256-2277 |
| SC_8_LEFT | GACAACTATTGTGGTTTCATATTTCT | 2193-2218 |
| SC_8_RIGHT | TTGTTGACACCTATTAATAATGTCCTCA | 2576-2594 |
| SC_9_LEFT | TGGGCTTTATTCCTCTACTGTCCC | 2492-2515 |
| SC_9_RIGHT | GGGAACAGAAAGATTCGTCCCC | 2889-2910 |
| SC_10_LEFT | TTGCGGGTCACCATATTCTTGG | 2816-2837 |
| SC_10_RIGHT | GGCCTGAGGATGACTGTCTCTT | 3189-3210 |
Primers adopted from Choga et al 2023 [23] (https://dx.doi.org/10.17504/protocols.io.5qpvo3xxzv4o/v1).
Figure S2.
Number of sequences generated per BCPP communities in the different Botswana districts.
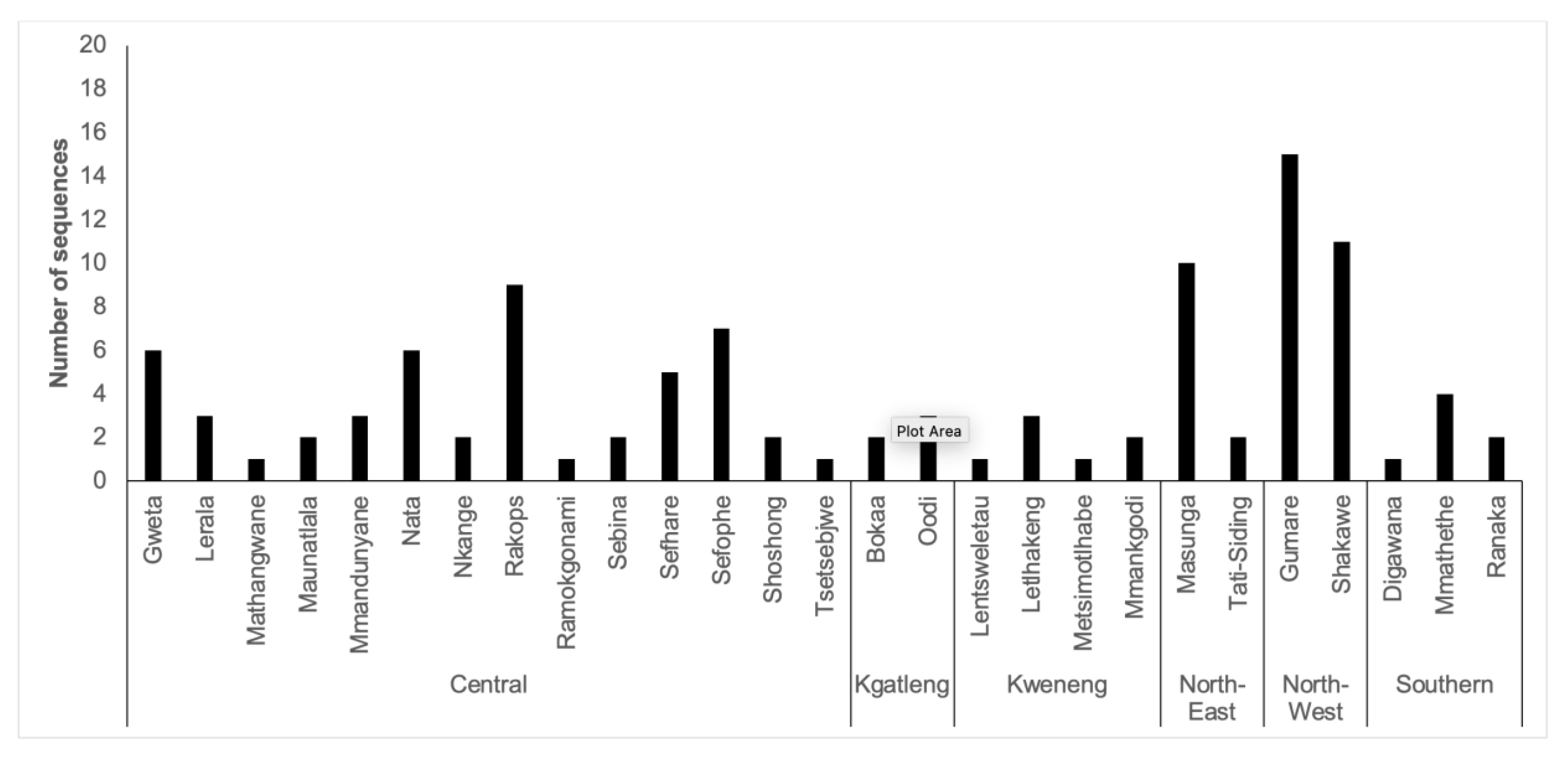
Table S3.
List of mutations in HBV ORFs
| REFERENCE SEQUENCES | BCPP SEQUENCES | COMMON MUTATIONS | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Mutation | Frequency | Prevalence | Mutation | Frequency | Prevalence | REF | BCPP | ||
| PreS1 | S5P | 3 | 8.1% | G35R | 12 | 12.1% | T94P | 44.4% | 50.5% |
| A6S | 6 | 16.2% | N37Y | 15 | 15.2% | ||||
| K10N | 3 | 8.1% | I48V | 21 | 21.2% | ||||
| F25L | 5 | 13.5% | A90V | 23 | 23.2% | ||||
| V74F | 2 | 5.4% | |||||||
| I84V | 5 | 13.5% | |||||||
| A86T | 2 | 5.4% | |||||||
| V88L | 3 | 8.1% | |||||||
| A90T | 6 | 16.2% | |||||||
| A90P | 9 | 24.3% | |||||||
| V91S | 5 | 13.5% | |||||||
| P92L | 4 | 11.1% | |||||||
| P92S | 5 | 13.9% | |||||||
| P93S | 4 | 11.1% | |||||||
| PreS2 | V17I | 2 | 5.3% | Y21C | 2 | 2.2% | A7T | 5.3% | 16.9% |
| Y21S | 2 | 5.1% | L32V | 8 | 9.0% | A11T | 5.3% | 10.3% | |
| F22L | 3 | 7.7% | N37T | 7 | 7.9% | T38I | 41.0% | 30.3% | |
| R48T | 5 | 12.8% | A39V | 9 | 10.1% | A53V | 12.8% | 4.5% | |
| H41P | 2 | 2.2% | L54P | 15.4% | 14.6% | ||||
| H41L | 3 | 3.4% | |||||||
| I45T | 18 | 20.2% | |||||||
| S46F | 8 | 9.0% | |||||||
| S47L | 2 | 2.2% | |||||||
| T49I | 6 | 6.7% | |||||||
| D51V | 6 | 6.7% | |||||||
| L54S | 21 | 23.6% | |||||||
| preC | C14Y | 2 | 7.14% | None | |||||
| W28L | 3 | 10.71% | |||||||
| V17F | 15 | 53.57% | |||||||
| Core | S35A | 2 | 8.3% | P45S | 8 | 9.8% | None | ||
| A41S | 2 | 8.3% | C61R | 2 | 2.4% | ||||
| C48G | 2 | 8.3% | G63R | 2 | 2.4% | ||||
| E64D | 3 | 12.5% | E64K | 3 | 3.7% | ||||
| L65V | 3 | 12.5% | M66T | 2 | 2.4% | ||||
| T67N | 4 | 16.7% | N74I | 3 | 3.7% | ||||
| N74S | 2 | 8.3% | S106F | 2 | 2.4% | ||||
| E77D | 2 | 8.3% | T142S | 2 | 2.7% | ||||
| A131P | 3 | 12.5% | V148A | 2 | 2.7% | ||||
| R151C | 4 | 16.7% | R181P | 3 | 3.7% | ||||
| D153A | 3 | 12.5% | |||||||
| S183P | 2 | 8.3% | |||||||
| X | E80A | 2 | 8.3% | L5V | 16 | 26.7% | S11P | 12.5% | 18.3% |
| I88S | 2 | 8.3% | Y6C | 9 | 15.0% | G22S | 8.3% | 43.8% | |
| K130M | 3 | 15.8% | S12A | 11 | 18.3% | A31T | 8.3% | 28.1% | |
| V131I | 4 | 21.1% | R26C | 13 | 20.3% | S46P | 41.7% | 67.2% | |
| P29S | 17 | 26.6% | |||||||
| A31S | 8 | 12.5% | |||||||
| G32R | 8 | 12.5% | |||||||
| P33S | 27 | 42.2% | |||||||
| T36G | 3 | 4.7% | |||||||
| T36A | 14 | 21.9% | |||||||
| S47A | 5 | 7.8% | |||||||
| D48V | 6 | 9.4% | |||||||
| C78R | 8 | 12.7% | |||||||
| A85S | 2 | 3.4% | |||||||
| L93S | 2 | 4.3% | |||||||
| V116L | 2 | 4.3% | |||||||
| S147P | 2 | 4.3% | |||||||
| Surface | M1R | 2 | 5.4% | N3S | 3 | 3.5% | T114S | 3.4% | 2.2% |
| S45P | 3 | 8.1% | G7R | 3 | 3.5% | K122R | 2.3% | 10.1% | |
| L49R | 2 | 5.4% | F8L | 2 | 2.3% | N131T | 14.8% | 2.2% | |
| L98V | 3 | 3.4% | R24K | 4 | 4.5% | V194A | 37.5% | 5.7% | |
| M103I | 3 | 3.4% | G44S | 2 | 2.3% | ||||
| P111A | 3 | 3.4% | S45T | 2 | 2.3% | ||||
| S117R | 2 | 2.3% | S45A | 4 | 4.5% | ||||
| Q129R | 4 | 4.5% | P46T | 3 | 3.4% | ||||
| G130N | 2 | 2.3% | S55P | 9 | 10.2% | ||||
| T140I | 2 | 2.3% | T57I | 5 | 5.6% | ||||
| A159V | 5 | 5.7% | V96A | 9 | 10.1% | ||||
| E164G | 4 | 4.5% | Q101K | 2 | 2.2% | ||||
| V168A | 2 | 2.3% | W156Q | 2 | 2.3% | ||||
| L173P | 3 | 3.4% | A159I | 2 | 2.3% | ||||
| V184A | 4 | 4.5% | E164D | 25 | 28.7% | ||||
| S204N | 8 | 9.1% | F179L | 2 | 2.3% | ||||
| V180A | 6 | 6.9% | |||||||
| W182R | 2 | 2.3% | |||||||
| F183S | 3 | 3.5% | |||||||
| V184L | 2 | 2.7% | |||||||
| G185W | 2 | 2.7% | |||||||
| I195M | 39 | 55.7% | |||||||
| W199R | 2 | 2.9% | |||||||
| Y200C | 2 | 2.9% | |||||||
| P203Q | 2 | 2.9% | |||||||
| Y206H | 2 | 2.9% | |||||||
| Y206R | 3 | 4.3% | |||||||
| I213M | 2 | 2.9% | |||||||
| P214L | 4 | 5.7% | |||||||
Table S4.
List of mutations in HBV polymerase domains.
| REFERENCE SEQUENCES | BCPP SEQUENCES | COMMON MUTATIONS | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Mutation | Frequency | Prevalence | Mutation | Frequency | Prevalence | REF | BCPP | ||
| TP | D16A | 3 | 7.7% | Q6H | 2 | 2.8% | V71I | 28.2% | 34.7% |
| T18L | 3 | 7.7% | A20V | 2 | 2.8% | Q87H | 53.8% | 47.2% | |
| V71N | 4 | 10.3% | A20G | 2 | 2.8% | H182Q | 5.1% | 21.7% | |
| E75D | 4 | 10.3% | L23P | 2 | 2.8% | ||||
| N120K | 2 | 5.1% | D32G | 2 | 2.8% | ||||
| S121G | 2 | 5.1% | A40I | 2 | 2.8% | ||||
| K161Q | 2 | 5.1% | G46Q | 2 | 2.8% | ||||
| L181I | 3 | 7.7% | W54L | 2 | 2.8% | ||||
| F61L | 2 | 2.8% | |||||||
| S80P | 2 | 2.8% | |||||||
| P82H | 2 | 2.8% | |||||||
| E88R | 2 | 2.8% | |||||||
| I91S | 13 | 18.1% | |||||||
| R93K | 2 | 2.8% | |||||||
| Q95E | 2 | 2.8% | |||||||
| M113I | 2 | 2.8% | |||||||
| D128N | 2 | 2.8% | |||||||
| Q138H | 15 | 20.8% | |||||||
| L153T | 2 | 3.8% | |||||||
| G157R | 2 | 3.8% | |||||||
| R162K | 2 | 3.8% | |||||||
| S167G | 2 | 4.3% | |||||||
| Spacer | V4A | 3 | 7.9% | S15P | 7 | 11.9% | P64A | 44.7% | 36.3% |
| S5I | 6 | 15.8% | S18P | 12 | 20.3% | I84T | 13.2% | 12.1% | |
| Q6K | 15 | 39.5% | V29I | 6 | 6.7% | Y86H | 13.2% | 26.4% | |
| A7T | 23 | 60.5% | R34Q | 11 | 12.2% | H93S | 44.7% | 45.1% | |
| R10W | 3 | 7.9% | Q36L | 15 | 16.7% | S125N | 5.6% | 22.9% | |
| Q19K | 5 | 13.2% | H47R | 19 | 20.9% | S129N | 5.3% | 12.3% | |
| P20S | 2 | 5.3% | R63W | 2 | 2.2% | ||||
| L24P | 5 | 13.2% | F74L | 2 | 2.2% | ||||
| V29T | 3 | 7.9% | S91A | 11 | 12.1% | ||||
| P46S | 3 | 7.9% | H93P | 2 | 2.2% | ||||
| P56S | 4 | 10.5% | K102E | 3 | 3.3% | ||||
| H83R | 5 | 13.2% | F126I | 4 | 8.2% | ||||
| G85D | 2 | 5.3% | P127S | 12 | 23.1% | ||||
| S87I | 2 | 5.3% | P128S | 5 | 9.6% | ||||
| S89N | 6 | 15.8% | S130F | 2 | 3.2% | ||||
| S89T | 9 | 23.7% | A131S | 9 | 12.7% | ||||
| S90K | 5 | 13.2% | Q136E | 2 | 2.5% | ||||
| S91F | 5 | 13.2% | Q136K | 5 | 6.2% | ||||
| S92L | 3 | 7.9% | G137R | 12 | 14.5% | ||||
| L108F | 3 | 8.1% | P138L | 2 | 2.4% | ||||
| V121M | 2 | 5.4% | S141P | 3 | 3.6% | ||||
| F123L | 2 | 5.6% | T150I | 2 | 2.2% | ||||
| S129G | 2 | 5.4% | T150S | 7 | 7.9% | ||||
| S135N | 2 | 5.4% | Q151E | 5 | 5.6% | ||||
| F140S | 4 | 10.5% | K155D | 6 | 6.7% | ||||
| L158I | 10 | 26.3% | S159T | 4 | 4.5% | ||||
| L161I | 2 | 5.3% | |||||||
| RT | E1D | 5 | 12.5% | V7T | 2 | 2.3% | V7A | 40.0% | 16.3% |
| H9Q | 3 | 7.5% | E11K | 3 | 3.5% | L53I | 35.0% | 11.4% | |
| S105T | 5 | 12.5% | R15K | 3 | 3.5% | I103V | 37.5% | 23.5% | |
| S106C | 3 | 7.5% | I16T | 2 | 2.3% | P109S | 20.0% | 14.1% | |
| R110G | 12 | 30.0% | A21T | 2 | 2.3% | H122N | 35.0% | 2.4% | |
| S119C | 3 | 7.5% | R51G | 2 | 2.3% | W153R | 7.5% | 19.0% | |
| Q139H | 12 | 30.0% | G52E | 2 | 2.3% | K266V | 27.5% | 14.5% | |
| R242K | 3 | 7.5% | L53N | 2 | 2.3% | K266I | 65.0% | 65.2% | |
| H271P | 2 | 5.0% | L53S | 2 | 2.3% | N332S | 39.3% | 15.9% | |
| H271S | 3 | 7.5% | T54Y | 2 | 2.3% | Q333K | 44.4% | 14.5% | |
| H271C | 13 | 32.5% | T54S | 4 | 4.5% | ||||
| T322I | 3 | 9.1% | V63A | 7 | 8.0% | ||||
| H122L | 2 | 2.4% | |||||||
| N124H | 19 | 22.4% | |||||||
| Y126H | 20 | 23.5% | |||||||
| L129M | 2 | 2.4% | |||||||
| N131D | 16 | 18.8% | |||||||
| Y151F | 2 | 2.4% | |||||||
| R167H | 2 | 2.4% | |||||||
| V173L | 23 | 27.4% | |||||||
| L180M | 41 | 47.1% | |||||||
| I187A | 2 | 2.3% | |||||||
| V190A | 2 | 2.3% | |||||||
| R192S | 2 | 2.8% | |||||||
| R192L | 3 | 4.2% | |||||||
| R193V | 2 | 2.8% | |||||||
| M204V | 37 | 53.6% | |||||||
| V207A | 2 | 2.9% | |||||||
| V214A | 2 | 2.9% | |||||||
| T222A | 2 | 2.9% | |||||||
| A223V | 2 | 2.9% | |||||||
| T225A | 2 | 2.9% | |||||||
| L229F | 2 | 2.9% | |||||||
| N238T | 9 | 13.0% | |||||||
| S246F | 3 | 4.3% | |||||||
| M250L | 4 | 5.8% | |||||||
| V253I | 10 | 14.5% | |||||||
| S256G | 6 | 8.7% | |||||||
| T259S | 8 | 11.6% | |||||||
| Q262R | 2 | 2.9% | |||||||
| I282V | 8 | 11.6% | |||||||
| C287Y | 2 | 2.9% | |||||||
| R289E | 2 | 2.9% | |||||||
| R289K | 2 | 2.9% | |||||||
| I290F | 2 | 2.9% | |||||||
| Y312H | 2 | 2.9% | |||||||
| M336V | 2 | 2.9% | |||||||
| Y339C | 2 | 2.9% | |||||||
| Y339L | 2 | 2.9% | |||||||
| RNaseH | V138D | 3 | 12.50% | R1Q | 2 | 9.5% | S2P | 41.7% | 61.9% |
| C5M | 2 | 9.5% | Y116F | 8.3% | 30.0% | ||||
| T37A | 2 | 9.1% | R151K | 8.3% | 7.8% | ||||
| A38T | 2 | 9.1% | |||||||
| K53N | 2 | 4.1% | |||||||
| L54I | 2 | 3.7% | |||||||
| G75S | 3 | 4.7% | |||||||
| T77A | 10 | 15.6% | |||||||
| G84R | 2 | 3.1% | |||||||
| A93V | 2 | 2.9% | |||||||
| P100L | 2 | 2.9% | |||||||
| L107V | 4 | 5.7% | |||||||
| Y108S | 3 | 4.3% | |||||||
| R113S | 2 | 2.9% | |||||||
| L114P | 2 | 2.9% | |||||||
| P115L | 5 | 7.1% | |||||||
| R117H | 2 | 2.9% | |||||||
| R117C | 2 | 2.9% | |||||||
| R117Q | 6 | 8.6% | |||||||
| T119S | 8 | 11.4% | |||||||
| V128D | 18 | 25.7% | |||||||
| F142S | 3 | 4.3% | |||||||
| V148A | 24 | 36.9% | |||||||
References
- WHO. Global hepatitis report 2024: action for access in low- and middle-income countries; World Health Organization: Geneva, 2024.
- Phinius, B.B.; Anderson, M.; Gobe, I.; Mokomane, M.; Choga, W.T.; Mutenga, S.R.; Mpebe, G.; Pretorius-Holme, M.; Musonda, R.; Gaolathe, T.; et al. High Prevalence of Hepatitis B Virus Infection Among People With HIV in Rural and Periurban Communities in Botswana. Open Forum Infect Dis 2023, 10, ofac707. [Google Scholar] [CrossRef]
- Mbangiwa, T.; Kasvosve, I.; Anderson, M.; Thami, P.K.; Choga, W.T.; Needleman, A.; Phinius, B.B.; Moyo, S.; Leteane, M.; Leidner, J.; et al. Chronic and Occult Hepatitis B Virus Infection in Pregnant Women in Botswana. Genes (Basel) 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Ryan, K.; Anderson, M.; Gyurova, I.; Ambroggio, L.; Moyo, S.; Sebunya, T.; Makhema, J.; Marlink, R.; Essex, M.; Musonda, R.; et al. High Rates of Occult Hepatitis B Virus Infection in HIV-Positive Individuals Initiating Antiretroviral Therapy in Botswana. Open Forum Infect Dis 2017, 4, ofx195. [Google Scholar] [CrossRef] [PubMed]
- Raimondo, G.; Pollicino, T.; Cacciola, I.; Squadrito, G. Occult hepatitis B virus infection. J Hepatol 2007, 46, 160–170. [Google Scholar] [CrossRef]
- Kato, M.; Hamada-Tsutsumi, S.; Okuse, C.; Sakai, A.; Matsumoto, N.; Sato, M.; Sato, T.; Arito, M.; Omoteyama, K.; Suematsu, N.; et al. Effects of vaccine-acquired polyclonal anti-HBs antibodies on the prevention of HBV infection of non-vaccine genotypes. J Gastroenterol 2017, 52, 1051–1063. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Liao, Y.; Chen, J.; Cai, B.; Su, Z.; Ying, B.; Lu, X.; Tao, C.; Wang, L. Epidemiology study of HBV genotypes and antiviral drug resistance in multi-ethnic regions from Western China. Scientific Reports 2015, 5, 17413. [Google Scholar] [CrossRef] [PubMed]
- Kao, J.H.; Chen, P.J.; Lai, M.Y.; Chen, D.S. Hepatitis B genotypes correlate with clinical outcomes in patients with chronic hepatitis B. Gastroenterology 2000, 118, 554–559. [Google Scholar] [CrossRef]
- Kao, J.H.; Liu, C.J.; Chen, D.S. Hepatitis B viral genotypes and lamivudine resistance. J Hepatol 2002, 36, 303–304. [Google Scholar] [CrossRef]
- Anderson, M.; Choga, W.T.; Moyo, S.; Bell, T.G.; Mbangiwa, T.; Phinius, B.B.; Bhebhe, L.; Sebunya, T.K.; Lockman, S.; Marlink, R.; et al. Molecular Characterization of Near Full-Length Genomes of Hepatitis B Virus Isolated from Predominantly HIV Infected Individuals in Botswana. Genes (Basel) 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Choga, W.T.; Anderson, M.; Zumbika, E.; Moyo, S.; Mbangiwa, T.; Phinius, B.B.; Melamu, P.; Kayembe, M.K.; Kasvosve, I.; Sebunya, T.K.; et al. Molecular characterization of hepatitis B virus in blood donors in Botswana. Virus Genes 2018. [Google Scholar] [CrossRef] [PubMed]
- Baruti, K.; Lentz, K.; Anderson, M.; Ajibola, G.; Phinius, B.B.; Choga, W.T.; Mbangiwa, T.; Powis, K.M.; Sebunya, T.; Blackard, J.T.; et al. Hepatitis B virus prevalence and vaccine antibody titers in children HIV exposed but uninfected in Botswana. PLoS One 2020, 15, e0237252. [Google Scholar] [CrossRef] [PubMed]
- Shaver, Z.M.; Anderson, M.; Bhebhe, L.; Baruti, K.; Choga, W.T.; Ngidi, J.; Mbangiwa, T.; Tau, M.; Setlhare, D.R.; Melamu, P.; et al. Decreased hepatitis B virus vaccine response among HIV-positive infants compared with HIV-negative infants in Botswana. AIDS 2022, 36, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Phinius, B.B.; Anderson, M.; Gobe, I.; Mokomane, M.; Choga, W.T.; Mokomane, M.; Choga, W.T.; Mpebe, G.; Makhema, J.; Shapiro, R.; et al. High Prevalence of Hepatitis B Virus Drug Resistance Mutations to Lamivudine Among People with HIV/HBV Coinfection in Rural and Peri-Urban Communities in Botswana. Preprints 2024. [Google Scholar] [CrossRef]
- Phinius, B.B.; Anderson, M.; Gobe, I.; Mokomane, M.; Choga, W.T.; Phakedi, B.; Ratsoma, T.; Mpebe, G.; Makhema, J.; Shapiro, R.; et al. High Prevalence of Hepatitis B Virus Drug Resistance Mutations to Lamivudine among People with HIV/HBV Coinfection in Rural and Peri-Urban Communities in Botswana. Viruses 2024, 16, 592. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Hollinger, F.B. Occult hepatitis B virus infection and hepatocellular carcinoma: a systematic review. J Viral Hepat 2014, 21, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Eilard, A.; Andersson, M.; Ringlander, J.; Wejstal, R.; Norkrans, G.; Lindh, M. Vertically acquired occult hepatitis B virus infection may become overt after several years. J Infect 2019, 78, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.; Choga, W.T.; Moyo, S.; Bell, T.G.; Mbangiwa, T.; Phinius, B.B.; Bhebhe, L.; Sebunya, T.K.; Makhema, J.; Marlink, R.; et al. In Silico Analysis of Hepatitis B Virus Occult Associated Mutations in Botswana Using a Novel Algorithm. Genes (Basel) 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Olagbenro, M.; Anderson, M.; Gaseitsiwe, S.; Powell, E.A.; Gededzha, M.P.; Selabe, S.G.; Blackard, J.T. In silico analysis of mutations associated with occult hepatitis B virus (HBV) infection in South Africa. Arch Virol 2021, 166, 3075–3084. [Google Scholar] [CrossRef] [PubMed]
- Phinius, B.B.; Anderson, M.; Bhebhe, L.; Baruti, K.; Manowe, G.; Choga, W.T.; Mupfumi, L.; Mbangiwa, T.; Mudanga, M.; Moyo, S.; et al. Increased Prevalence of Liver Fibrosis and HIV Viremia among Patients with HIV, HBV, and Tuberculosis in Botswana. Pathogens 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Gaolathe, T.; Wirth, K.E.; Holme, M.P.; Makhema, J.; Moyo, S.; Chakalisa, U.; Yankinda, E.K.; Lei, Q.; Mmalane, M.; Novitsky, V.; et al. Botswana’s progress toward achieving the 2020 UNAIDS 90-90-90 antiretroviral therapy and virological suppression goals: a population-based survey. Lancet HIV 2016, 3, e221–230. [Google Scholar] [CrossRef] [PubMed]
- Makhema, J.; Wirth, K.E.; Pretorius Holme, M.; Gaolathe, T.; Mmalane, M.; Kadima, E.; Chakalisa, U.; Bennett, K.; Leidner, J.; Manyake, K.; et al. Universal Testing, Expanded Treatment, and Incidence of HIV Infection in Botswana. N Engl J Med 2019, 381, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Choga, W. Complete Hepatitis B Virus Sequencing using an ONT-Based Next-Generation Sequencing Protocol; 2023.
- Phinius, B.B.; Anderson, M.; Mokomane, M.; Gobe, I.; Choga, W.T.; Ratsoma, T.; Phakedi, B.; Mpebe, G.; Ditshwanelo, D.; Musonda, R.; et al. Atypical Hepatitis B Virus Serology Profile-Hepatitis B Surface Antigen-Positive/Hepatitis B Core Antibody-Negative-In Hepatitis B Virus/HIV Coinfected Individuals in Botswana. Viruses 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- ONT. PCR Tiling of Sars Cov 2 Virus With Rapid Barcoding SQK rbk110 PCTR - 9125 - v110 - Revh - 24mar2021 Minion. Available online: https://community.nanoporetech.com/docs/prepare/library_prep_protocols/pcr-tiling-of-sars-cov-2-virus-with-rapid-barcoding-and-midnight/v/mrt_9127_v110_revw_14jul2021 (accessed on 28 March 2023).
- Vilsker, M.; Moosa, Y.; Nooij, S.; Fonseca, V.; Ghysens, Y.; Dumon, K.; Pauwels, R.; Alcantara, L.C.; Vanden Eynden, E.; Vandamme, A.M.; et al. Genome Detective: an automated system for virus identification from high-throughput sequencing data. Bioinformatics 2019, 35, 871–873. [Google Scholar] [CrossRef] [PubMed]
- Larsson, A. AliView: a fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.; Gaseitsiwe, S.; Moyo, S.; Wessels, M.J.; Mohammed, T.; Sebunya, T.K.; Powell, E.A.; Makhema, J.; Blackard, J.T.; Marlink, R.; et al. Molecular characterisation of hepatitis B virus in HIV-1 subtype C infected patients in Botswana. BMC Infect Dis 2015, 15, 335. [Google Scholar] [CrossRef] [PubMed]
- Kwei, K.; Tang, X.; Lok, A.S.; Sureau, C.; Garcia, T.; Li, J.; Wands, J.; Tong, S. Impaired virion secretion by hepatitis B virus immune escape mutants and its rescue by wild-type envelope proteins or a second-site mutation. J Virol 2013, 87, 2352–2357. [Google Scholar] [CrossRef] [PubMed]
- Konopleva, M.V.; Belenikin, M.S.; Shanko, A.V.; Bazhenov, A.I.; Kiryanov, S.A.; Tupoleva, T.A.; Sokolova, M.V.; Pronin, A.V.; Semenenko, T.A.; Suslov, A.P. Detection of S-HBsAg Mutations in Patients with Hematologic Malignancies. Diagnostics (Basel) 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.L.; Li, X.; Li, J.; Zhang, Z.H. Genetic variation of occult hepatitis B virus infection. World J Gastroenterol 2016, 22, 3531–3546. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Xu, Y.; Zhang, Z.; Meng, Z.; Qin, L.; Lu, M.; Yang, D. The amino Acid residues at positions 120 to 123 are crucial for the antigenicity of hepatitis B surface antigen. J Clin Microbiol 2007, 45, 2971–2978. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.G.; Ueda, K. A meta-analysis on genetic variability of RT/HBsAg overlapping region of hepatitis B virus (HBV) isolates of Bangladesh. Infect Agent Cancer 2019, 14, 33. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.M.; Welge, J.A.; Rouster, S.D.; Shata, M.T.; Sherman, K.E.; Blackard, J.T. Mutations associated with occult hepatitis B virus infection result in decreased surface antigen expression in vitro. J Viral Hepat 2012, 19, 716–723. [Google Scholar] [CrossRef]
- Yong-Lin, Y.; Qiang, F.; Ming-Shun, Z.; Jie, C.; Gui-Ming, M.; Zu-Hu, H.; Xu-Bing, C. Hepatitis B surface antigen variants in voluntary blood donors in Nanjing, China. Virol J 2012, 9, 82. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Li, F.; Guo, H.; Yang, S.; Zhang, Y.; Zhu, H.; Wang, J.; Mao, R.; Qin, Y.; Xu, J.; et al. Amino acid substitutions Q129N and T131N/M133T in hepatitis B surface antigen (HBsAg) interfere with the immunogenicity of the corresponding HBsAg or viral replication ability. Virus Research 2018, 257, 33–39. [Google Scholar] [CrossRef]
- Huang, C.H.; Yuan, Q.; Chen, P.J.; Zhang, Y.L.; Chen, C.R.; Zheng, Q.B.; Yeh, S.H.; Yu, H.; Xue, Y.; Chen, Y.X.; et al. Influence of mutations in hepatitis B virus surface protein on viral antigenicity and phenotype in occult HBV strains from blood donors. J Hepatol 2012, 57, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Wang, Y. Comprehensive analysis of the prevalence of hepatitis B virus escape mutations in the major hydrophilic region of surface antigen. J Med Virol 2012, 84, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.N.; Oon, C.J. Changes in the antigenicity of a hepatitis B virus mutant stemming from lamivudine therapy. Antimicrob Agents Chemother 2000, 44, 1765. [Google Scholar] [CrossRef] [PubMed]
- Khodadad, N.; Seyedian, S.S.; Moattari, A.; Biparva Haghighi, S.; Pirmoradi, R.; Abbasi, S.; Makvandi, M. In silico functional and structural characterization of hepatitis B virus PreS/S-gene in Iranian patients infected with chronic hepatitis B virus genotype D. Heliyon 2020, 6, e04332. [Google Scholar] [CrossRef] [PubMed]
- Colagrossi, L.; Hermans, L.E.; Salpini, R.; Di Carlo, D.; Pas, S.D.; Alvarez, M.; Ben-Ari, Z.; Boland, G.; Bruzzone, B.; Coppola, N.; et al. Immune-escape mutations and stop-codons in HBsAg develop in a large proportion of patients with chronic HBV infection exposed to anti-HBV drugs in Europe. BMC Infect Dis 2018, 18, 251. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, F.; Slanina, H.; Roderfeld, M.; Roeb, E.; Trebicka, J.; Ziebuhr, J.; Gerlich, W.H.; Schüttler, C.G.; Schlevogt, B.; Glebe, D. A Novel Insertion in the Hepatitis B Virus Surface Protein Leading to Hyperglycosylation Causes Diagnostic and Immune Escape. Viruses 2023, 15, 838. [Google Scholar] [CrossRef] [PubMed]
- Mokaya, J.; McNaughton, A.L.; Hadley, M.J.; Beloukas, A.; Geretti, A.M.; Goedhals, D.; Matthews, P.C. A systematic review of hepatitis B virus (HBV) drug and vaccine escape mutations in Africa: A call for urgent action. PLoS Negl Trop Dis 2018, 12, e0006629. [Google Scholar] [CrossRef] [PubMed]
- Bi, X.; Tong, S. Impact of immune escape mutations and N-linked glycosylation on the secretion of hepatitis B virus virions and subviral particles: Role of the small envelope protein. Virology 2018, 518, 358–368. [Google Scholar] [CrossRef] [PubMed]
- Adesina, O.A.; Akanbi, O.A.; Opaleye, O.O.; Japhet, M.O.; Wang, B.; Oluyege, A.O.; Klink, P.; Bock, C.T. Detection of Q129H Immune Escape Mutation in Apparently Healthy Hepatitis B Virus Carriers in Southwestern Nigeria. Viruses 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Deshkar, S.T.; Patil, N.B.; Lad, A.A.; Papal, N.S.; S. , S.S. Monitoring of hepatitis B virus surface antigen escape mutations and concomitant nucleostide analog resistance mutations in patients with chronic hepatitis B. International Journal of Research in Medical Sciences 2022, 10, 2261–2267. [Google Scholar] [CrossRef]
- Lazarevic, I. Clinical implications of hepatitis B virus mutations: recent advances. World J Gastroenterol 2014, 20, 7653–7664. [Google Scholar] [CrossRef] [PubMed]
- Cooreman, M.P.; Leroux-Roels, G.; Paulij, W.P. Vaccine- and hepatitis B immune globulin-induced escape mutations of hepatitis B virus surface antigen. J Biomed Sci 2001, 8, 237–247. [Google Scholar] [CrossRef]
- Gencay, M.; Hübner, K.; Gohl, P.; Seffner, A.; Weizenegger, M.; Neofytos, D.; Batrla, R.; Woeste, A.; Kim, H.S.; Westergaard, G.; et al. Ultra-deep sequencing reveals high prevalence and broad structural diversity of hepatitis B surface antigen mutations in a global population. PLoS One 2017, 12, e0172101. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.H.; Wu, C.C.; Chen, X.W.; Li, X.; Li, J.; Lu, M.J. Genetic variation of hepatitis B virus and its significance for pathogenesis. World J Gastroenterol 2016, 22, 126–144. [Google Scholar] [CrossRef] [PubMed]
- Enriquez-Navarro, K.; Maldonado-Rodriguez, A.; Rojas-Montes, O.; Torres-Ibarra, R.; Bucio-Ortiz, L.; De la Cruz, M.A.; Torres-Flores, J.; Xoconostle-Cazares, B.; Lira, R. Identification of mutations in the S gene of hepatitis B virus in HIV positive Mexican patients with occult hepatitis B virus infection. Annals of Hepatology 2020, 19, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Mondal, R.K.; Khatun, M.; Banerjee, P.; Ghosh, A.; Sarkar, S.; Santra, A.; Das, K.; Chowdhury, A.; Banerjee, S.; Datta, S. Synergistic impact of mutations in Hepatitis B Virus genome contribute to its occult phenotype in chronic Hepatitis C Virus carriers. Scientific Reports 2017, 7, 9653. [Google Scholar] [CrossRef]
- Cortese, M.F.; González, C.; Gregori, J.; Casillas, R.; Carioti, L.; Guerrero-Murillo, M.; Riveiro-Barciela, M.; Godoy, C.; Sopena, S.; Yll, M.; et al. Sophisticated viral quasispecies with a genotype-related pattern of mutations in the hepatitis B X gene of HBeAg-ve chronically infected patients. Sci Rep 2021, 11, 4215. [Google Scholar] [CrossRef] [PubMed]
- Ave, A.R.; Putra, A.E.; Miro, S. Gene X Two Triple Mutations Predominance on Chronic Hepatitis B Virus in Padang, West Sumatra, Indonesia. The Indonesian Journal of Gastroenterology, Hepatology and Digestive Endoscopy 2022, 23, 106–111. [Google Scholar] [CrossRef]
- Zhang, X.; Ding, H.G. Key role of hepatitis B virus mutation in chronic hepatitis B development to hepatocellular carcinoma. World J Hepatol 2015, 7, 1282–1286. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Liu, S.; Zhao, Y.; Guo, Z.; Xu, J. X protein mutations in hepatitis B virus DNA predict postoperative survival in hepatocellular carcinoma. Tumour Biol 2014, 35, 10325–10331. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.N.; Flanagan, J.M.; Hu, J. Mapping of Functional Subdomains in the Terminal Protein Domain of Hepatitis B Virus Polymerase. J Virol 2017, 91. [Google Scholar] [CrossRef]
- Taherkhani, R.; Farshadpour, F. Prevalence, genotype distribution and mutations of hepatitis B virus and the associated risk factors among pregnant women residing in the northern shores of Persian Gulf, Iran. PLoS One 2022, 17, e0265063. [Google Scholar] [CrossRef]
- Park, Y.M.; Kim, B.S.; Tabor, E. Precore codon 28 stop mutation in hepatitis B virus from patients with hepatocellular carcinoma. Korean J Intern Med 1997, 12, 201–207. [Google Scholar] [CrossRef]
- Hudu, S.A.; Harmal, N.S.; Saeed, M.I.; Alshrari, A.S.; Malik, Y.A.; Niazlin, M.T.; Hassan, R.; Sekawi, Z. Naturally occurring hepatitis B virus surface antigen mutant variants in Malaysian blood donors and vaccinees. Eur J Clin Microbiol Infect Dis 2015, 34, 1349–1359. [Google Scholar] [CrossRef] [PubMed]
- Salpini, R.; Battisti, A.; Piermatteo, L.; Carioti, L.; Anastasiou, O.E.; Gill, U.S.; Di Carlo, D.; Colagrossi, L.; Duca, L.; Bertoli, A.; et al. Key mutations in the C-terminus of the HBV surface glycoprotein correlate with lower HBsAg levels in vivo, hinder HBsAg secretion in vitro and reduce HBsAg structural stability in the setting of HBeAg-negative chronic HBV genotype-D infection. Emerg Microbes Infect 2020, 9, 928–939. [Google Scholar] [CrossRef]
- Askari, A.; Hassanshahi, G.H.; Ghalebi, S.R.; Jafarzadeh, A.; Mohit, M.; Hajghani, M.; Kazemi Arababadi, M. Intensity of HLA-A2 Expression Significantly Decreased in Occult Hepatitis B Infection. Jundishapur J Microbiol 2014, 7, e10298. [Google Scholar] [CrossRef]
- Arababadi, M.K.; Pourfathollah, A.A.; Jafarzadeh, A.; Hassanshahi, G. Serum Levels of IL-10 and IL-17A in Occult HBV-Infected South-East Iranian Patients. Hepat Mon 2010, 10, 31–35. [Google Scholar]
- Vivekanandan, P.; Kannangai, R.; Ray, S.C.; Thomas, D.L.; Torbenson, M. Comprehensive genetic and epigenetic analysis of occult hepatitis B from liver tissue samples. Clin Infect Dis 2008, 46, 1227–1236. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Li, Y.; Mu, S.; Zhang, J.; Yan, Z. Evidence that methylation of hepatitis B virus covalently closed circular DNA in liver tissues of patients with chronic hepatitis B modulates HBV replication. J Med Virol 2009, 81, 1177–1183. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, M.; Huang, J.; Xu, R.; Liao, Q.; Shan, Z.; Zheng, Y.; Rong, X.; Tang, X.; Li, T.; et al. Novel hepatitis B virus surface antigen mutations associated with occult genotype B hepatitis B virus infection affect HBsAg detection. J Viral Hepat 2020, 27, 915–921. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Consort diagram of successfully sequenced samples.
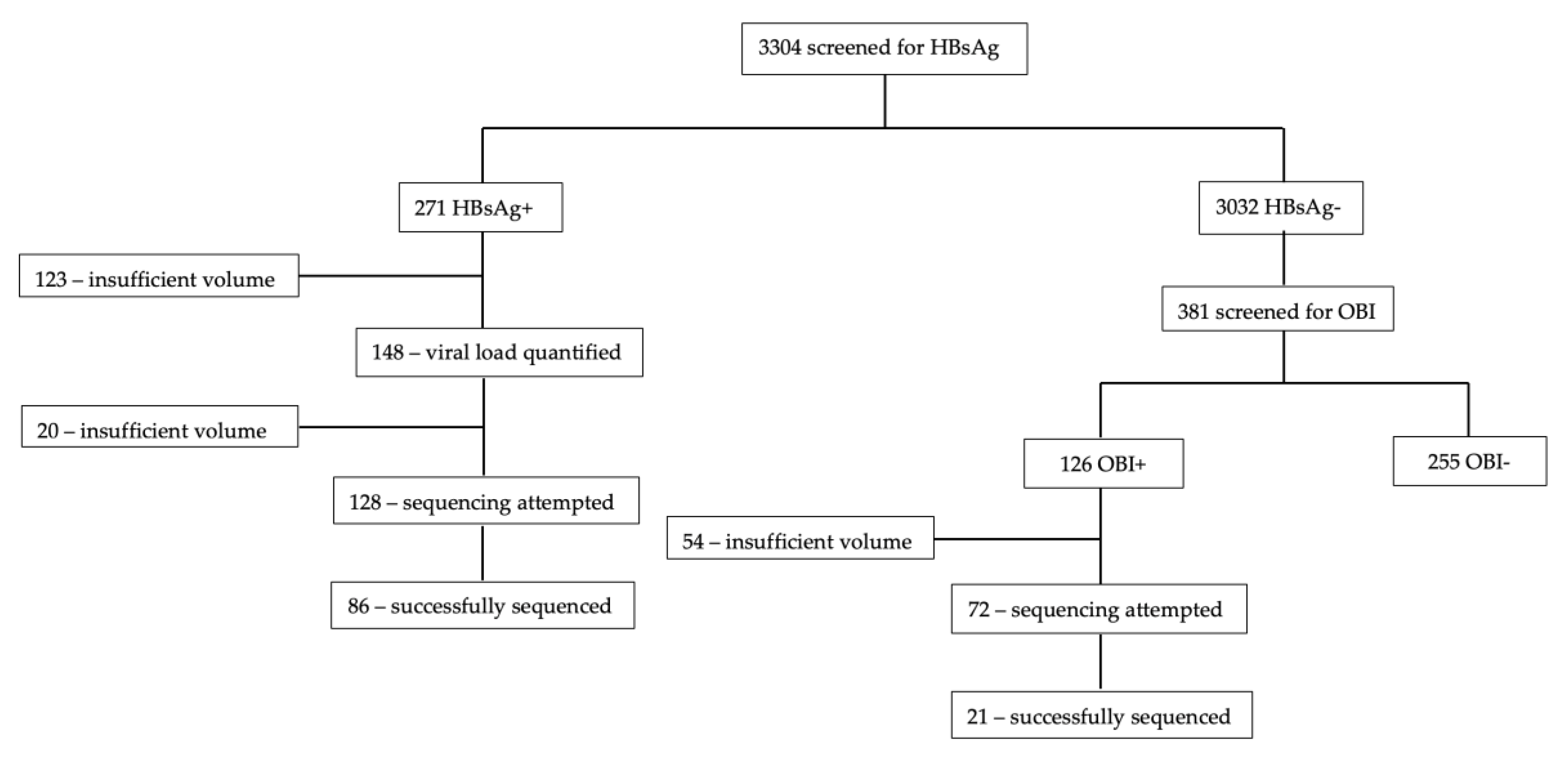
Figure 2.
Generated sequences in the different districts of Botswana.
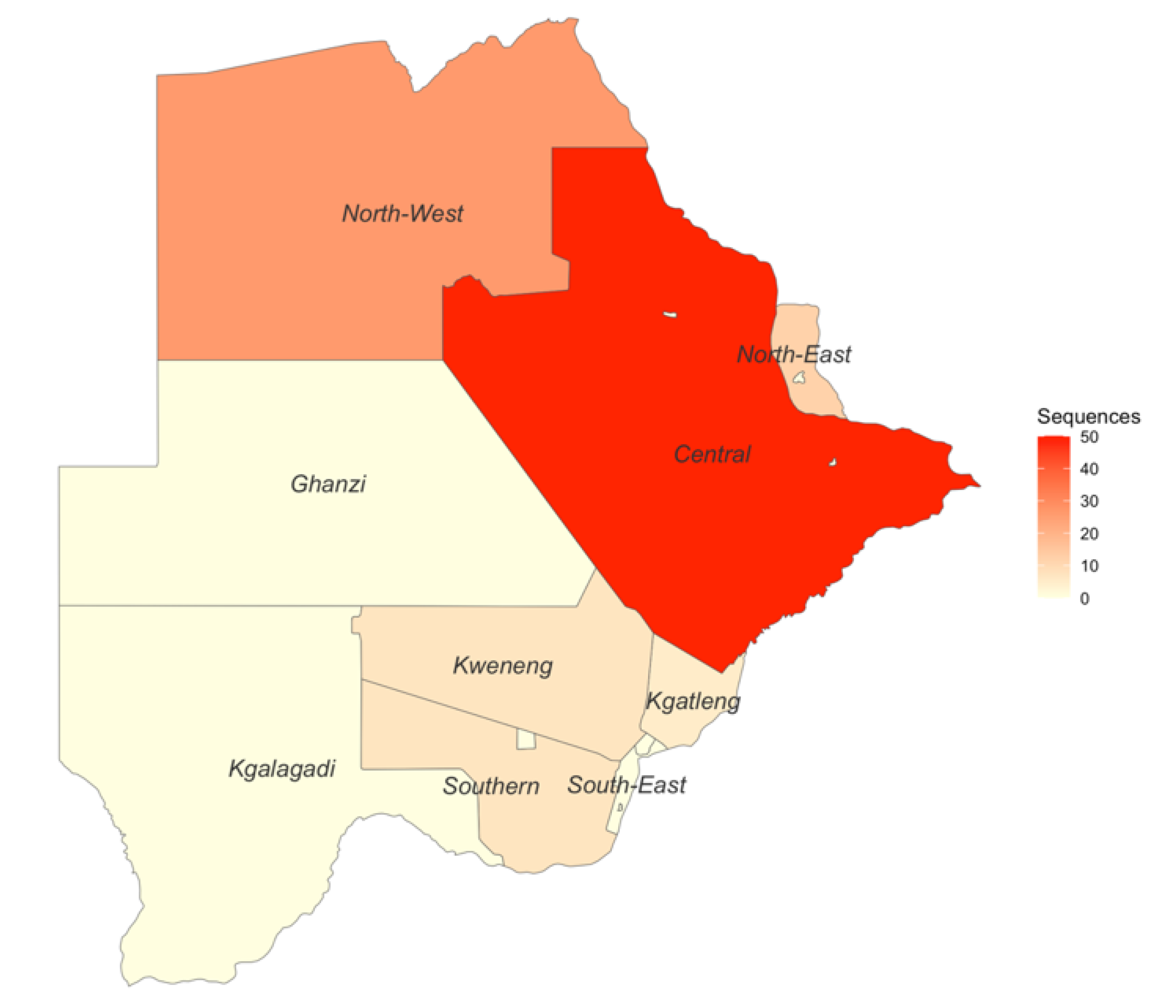
Figure 3.
Bayesian phylogenetic tree of sequences generated in the BCPP cohort and reference sequences. Study sequences are denoted by BBP followed by a number while references sequences start with accession number followed by country. Sequences in blue are genotype A1, those in red are genotype D3 and those in black are E.
Figure 3.
Bayesian phylogenetic tree of sequences generated in the BCPP cohort and reference sequences. Study sequences are denoted by BBP followed by a number while references sequences start with accession number followed by country. Sequences in blue are genotype A1, those in red are genotype D3 and those in black are E.
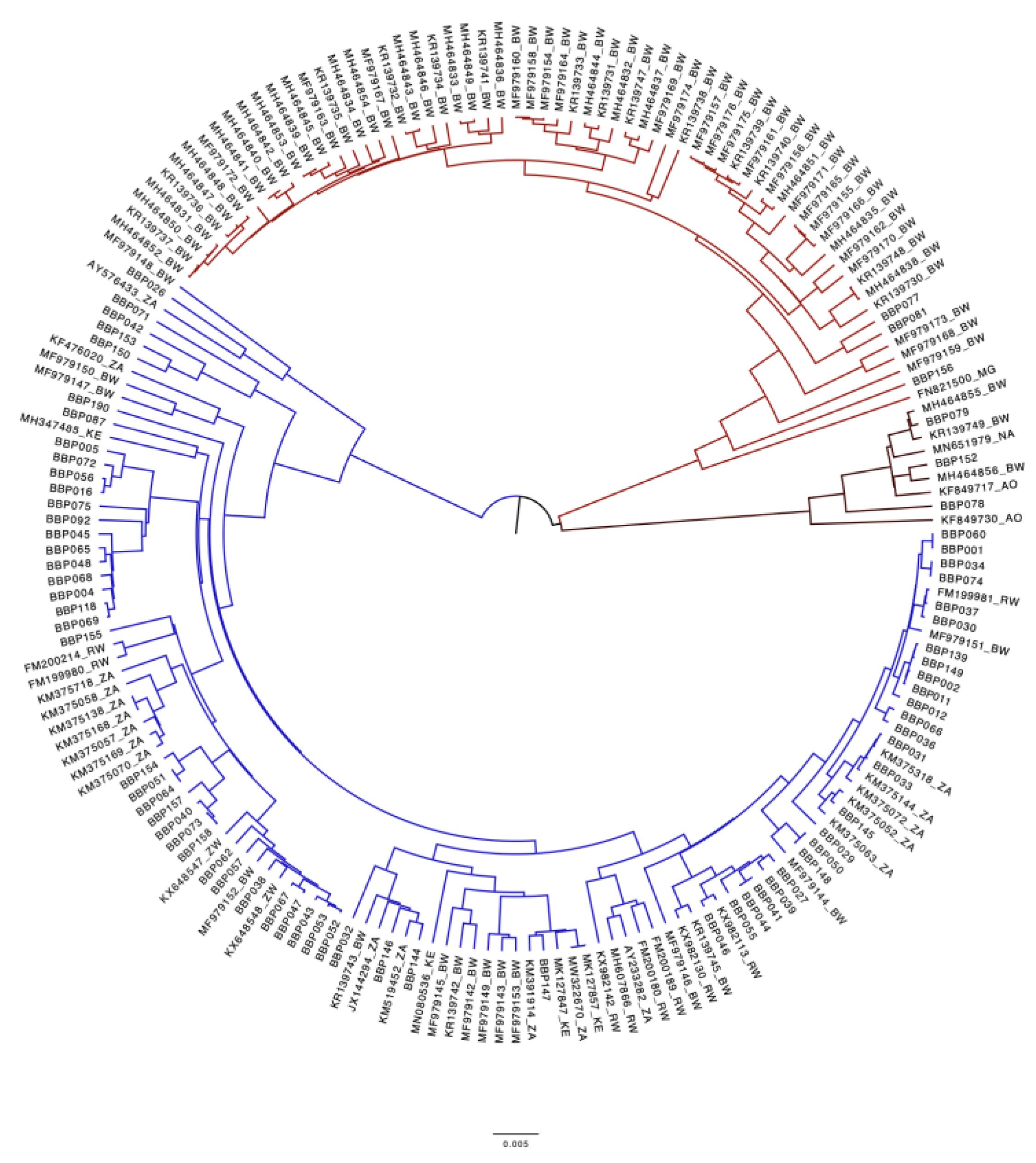
Table 1.
Participants clinical and socio-economic characteristics at enrollment.
| Characteristics | Number (%) n=107 |
|---|---|
| Sex Female |
71 (66.4) |
| Age, years, median (IQR) | 43 (36 – 49) |
| HBV type HBsAg+ OBI+ |
86 (80.4) 21 (19.6) |
| Total anti-HBc, n=104 Positive |
87 (83.7) |
| Anti-HBc IgM status*, n=81 Positive |
5 (6.2) |
| HBeAg status*, n=82 Positive |
15 (18.3) |
| HBV viral load Target not detected <2000 ≥2000 |
21 (19.6) 63 (58.9) 23 (21.5) |
| HIV viral load Undetectable Detectable |
95 (88.8) 12 (11.2) |
| ART status Naïve On ART |
6 (5.6) 101 (94.4) |
| ART regimen None 3TC/TDF-containing regimen 3TC-containing regimen TDF-containing regimen# Unknown |
1 (1.0) 27 (26.7) 45 (44.6) 28 (27.7) |
| Duration on ART, years, n=76, median (IQR) | 7.0 (4.7 – 9.9) |
HBV; hepatitis B virus, IQR; interquartile range, HBsAg; hepatitis B surface antigen, OBI; occult hepatitis infection, HBeAg; hepatitis B e antigen, anti-HBc: hepatitis B core antibody, anti-HBc IgM; hepatitis B core antigen immunoglobulin M antibodies, HIV; human immunodeficiency virus, ART; antiretroviral therapy, 3TC; lamivudine, TDF; tenofovir disoproxil fumarate. *Only in HBsAg positive participants. #The TDF containing regimen also had emtricitabine (FTC) for all participants except one who was on a dolutegravir/TDF regimen.
Table 2.
Escape mutations in the surface region.
| Mutation | Frequency | Genotype | HBV type | Reported impact | Reference |
|---|---|---|---|---|---|
| T114S | 2 | A1 | HBsAg | Other substitutions at position 114 (R) reported to impair virion secretion | [29] |
| S114L | 1 | E | HBsAg | Other substitutions at position 114 (R) reported to impair virion secretion | [29] |
| T118M | 1 | A1 | OBI | Impair antigenicity, detection escape | [30] |
| C121R | 1 | A1 | OBI | Other substitutions at position 121 (S) reported to reduce antigenicity and impair HBsAg detection | [31,32] |
| K122R | 9 | A1 | HBsAg | Decreased HBsAg expression, detection failure | [33,34,35] |
| Q129C | 1 | A1 | OBI | Other Q129 (N) mutations lead to impaired antigenicity and immunogenicity, Q129R leads to impaired virion/S protein secretion, Q129H leads to decreased virion secretion | [29,36,37] |
| G130C | 1 | A1 | OBI | Other G130 mutations to lead to diagnostic escape, vaccine/ immunoglobulin therapy escape, altered antigenicity | [38,39] |
| N131T | 2 | A1 | HBsAg | Vaccine escape | [40] |
| T131N | 1 | D3 | HBsAg | Vaccine escape, diagnostic escape, hepatitis B immunoglobulin resistance | [41,42,43] |
| C137I | 1 | A1 | HBsAg | Other C137 mutations are reported to decrease antigenicity | [31] |
| C139R | 1 | A1 | HBsAg | Impair virion/S protein secretion | [37] |
| N146S | 1 | E | OBI | Impair virion secretion | [29,44] |
| C147Y | 1 | A1 | HBsAg | Impair virion secretion | [29] |
HBsAg; hepatitis B surface antigen, OBI; occult hepatitis infection.
Table 3.
Mutations in HBV ORFs.
| REFERENCE SEQUENCES | BCPP SEQUENCES | COMMON MUTATIONS | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Mutation | Frequency | Prevalence | Mutation | Frequency | Prevalence | Mutation | REFERENCE | BCPP | |
| PreS1 | A90P | 9/37 | 24.3% | I48V | 21/99 | 21.2% | T94P | 44.4% | 50.5% |
| A90V | 23/99 | 23.2% | |||||||
| PreS2 | None | I45T | 18/89 | 20.2% | A7T | 5.3% | 16.9% | ||
| L54S | 21/89 | 23.6% | A11T | 5.3% | 10.3% | ||||
| T38I | 41.0% | 30.3% | |||||||
| A53V | 12.8% | 4.5% | |||||||
| preC | None | V17F | 15/28 | 53.6% | None | ||||
| Core | None | None | None | ||||||
| X | V131I | 4/19 | 21.1% | R26C | 13/64 | 20.3% | S11P | 12.5% | 18.3% |
| P29S | 17/64 | 26.6% | G22S | 8.3% | 43.8% | ||||
| P33S | 27/64 | 42.2% | A31T | 8.3% | 28.1% | ||||
| T36A | 14/64 | 21.9% | S46P | 41.7% | 67.2% | ||||
| Surface | E164D | 25/87 | 28.7% | K122R | 2.3% | 10.1% | |||
| I195M | 39/70 | 55.7% | N131T | 14.8% | 2.2% | ||||
| V194A | 37.5% | 5.7% | |||||||
BCPP; Botswana Combination Prevention Project.
Table 4.
Mutations in the polymerase domains.
| BW REFRENCE SEQUENCES | BCPP REFERENCES | COMMON MUTATIONS | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mutation | Frequency | Prevalence | Mutation | Frequency | Prevalence | REFERENCE | BCPP | |||
| TP | Q138H | 15/72 | 20.8% | V71I | 28.2% | 34.7% | ||||
| Q87H | 53.8% | 47.2% | ||||||||
| H182Q | 5.1% | 21.7% | ||||||||
| Spacer | Q6K | 15/38 | 39.5% | S18P | 12/59 | 20.3% | P64A | 44.7% | 36.3% | |
| A7T | 23/38 | 60.5% | H47R | 19/91 | 20.9% | I84T | 13.2% | 12.1% | ||
| S89T | 9/38 | 23.7% | P127S | 12/52 | 23.1% | Y86H | 13.2% | 26.4% | ||
| L158I | 10/38 | 26.3% | H93S | 44.7% | 45.1% | |||||
| S125N | 5.6% | 22.9% | ||||||||
| S129N | 5.3% | 12.3% | ||||||||
| RT | R110G | 12/40 | 30.0% | N124H | 19/84 | 22.6% | V7A | 40.0% | 16.3% | |
| Q139H | 12/40 | 30.0% | Y126H | 20/84 | 23.8% | L53I | 35.0% | 11.4% | ||
| H271C | 13/40 | 32.5% | V173L | 23/84 | 27.4% | I103V | 37.5% | 23.5% | ||
| L180M | 41/87 | 47.1% | P109S | 20.0% | 14.1% | |||||
| M204V | 37/69 | 53.6% | H122N | 35.0% | 2.4% | |||||
| W153R | 7.5% | 19.0% | ||||||||
| K266V | 27.5% | 14.5% | ||||||||
| K266I | 65.0% | 65.2% | ||||||||
| N332S | 39.3% | 15.9% | ||||||||
| Q333K | 44.4% | 14.5% | ||||||||
| Rnase H | V128D | 18/70 | 25.7% | S2P | 41.7% | 61.9% | ||||
| V148A | 24/70 | 34.3% | Y116F | 8.3% | 30.0% | |||||
| R151K | 8.3% | 7.8% | ||||||||
BCPP; Botswana Combination Prevention Project, TP; terminal protein, RT; reverse transcriptase.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



