
Submitted:
31 May 2024
Posted:
06 June 2024
You are already at the latest version
Abstract
The oral microbiome is a diverse and complex ecosystem essential for maintaining oral and systemic health. Our study is the first to define the oral microbial community in Egyptian young adults and investigate the effects of natural antimicrobials on the oral microbiome. SuperMint (SM) is a proprietary blend of peppermint, Japanese mint, bergamot mint, and spearmint essential oils encapsulated in a tiny soft beadlet. This work aimed to evaluate the effects of SM beadlets on the oral microbiome. This study recruited twenty healthy participants. A baseline investigation of the oral microbiome of the selected participants was performed by collecting saliva and swab samples before treatment. Treatment included chewing four SM beadlets twice a day for 7 days, and then post-administration saliva and swab samples were collected at the end of treatment. The oral microbiome samples were analyzed by high-throughput amplicon sequencing of 16S rRNA gene fragments, and the community composition was determined. The results showed that the abundance of some microbial genera and families decreased after using SM, including Prevotella, Streptococcus, Neisseria, and Haemophilus. However, some genera showed inconsistent patterns. We also found that the subject's gender and SM usage were significantly associated with diverse microbial composition. The results suggest that SM treatment decreased the abundance of several bacteria associated with halitosis and periodontal diseases, such as Actinomyces and Streptococcus. Furthermore, Corynebacterium species increased while Streptococcus decreased after SM usage. More research is needed to fully understand the antimicrobial effects of mint oils and their potential applications in maintaining good oral health.
Keywords:
Baseline study
; Oral microbiome
; SuperMint beadlets
; Mentha piperita EO
; Mentha canadensis EO
; Mentha citrata EO
; Mentha spicata EO
1. Introduction
The human body is colonized by a vast number of microbial populations known as the human microbiome [1,2]. Trillions of bacteria, viruses, fungi, and other microorganisms (about 100 trillion cells) colonize various locations in the body, such as the oral cavity, nasal cavity, gut, reproductive tract, and skin [1,3]. The oral microbiome is a diverse and complex ecosystem consisting of over 1,000 species of bacteria, archaea, viruses, fungi, and protozoa. These microorganisms are essential to maintaining good oral health by helping with nutrient digestion and absorption, preventing harmful microorganisms from taking hold, and boosting the immune system [4]. Several factors can affect the oral microbiome, including diet, antibiotic use, stress, oral hygiene practices, genetics, diseases like diabetes, and the environment [5]. A diet high in sugars can result in tooth decay and gum disease. On the other hand, a fiber and probiotics-rich diet can encourage the growth of beneficial bacteria that enhance oral health [6]. Poor oral hygiene practices like infrequent brushing and flossing can lead to the accumulation of harmful bacteria and plaque, which can cause dental diseases. Moreover, imbalances in the oral microbiome community can have systemic effects on the body [7]. A strong correlation was reported between periodontal pathogens and increased risk of respiratory infections, cardiovascular disease, diabetes, and some cancers [8,9,10]. Understanding the dynamic nature of the oral microbiome is essential to developing successful preventive and therapeutic strategies to maintain good oral and systemic health [5].
Several methods are used to study the oral microbiome, including 16S rRNA gene sequencing, metagenomics, culturing bacteria from oral samples, microscopy, and functional assays [1]. One of the most common methods is 16S rRNA gene sequencing, which involves analyzing the DNA of bacterial species in oral samples [11]. This method can identify different bacterial groups by targeting a conserved region of the bacterial genome and provide information on their abundance and diversity. The method involves extracting DNA from oral samples and amplifying the 16S rRNA gene using polymerase chain reaction (PCR). The amplified DNA is then sequenced and analyzed using bioinformatics tools to identify different bacterial groups [11,12]. Another method is metagenomics, which involves comprehensive sequencing of all the DNA in a sample, including bacterial, viral, and fungal DNA. The method involves extracting DNA from oral samples, sequencing the DNA using high-throughput sequencing technologies, and analyzing the data using bioinformatics tools to identify different microbial groups and determine their functions [13].
SuperMint (SM) is a proprietary blend of peppermint (Mentha piperita L.), Japanese mint (M. arvensis L.), bergamot mint (M. citrata Ehrh.), and spearmint (M. spicata L.) EOs. The SM blend is encapsulated in a tiny soft beadlet for oral use. Peppermint essential oil (EO) is known for its ability to inhibit the growth of microorganisms, making it an effective natural alternative to synthetic antimicrobials [14,15,16,17,18,19,20,21]. Studies have shown that peppermint EO can be effective against antibiotic-resistant microorganisms, which is particularly important given the increasing prevalence of antibiotic resistance [22,23,24]. The active ingredient in mint EO, menthol, has strong antimicrobial properties against various microorganisms [14,16,17,25,26]. The antimicrobial effects of mint EO have been observed in a variety of applications. For example, mint oil has been used in oral hygiene products (toothpaste, mouthwash, and dental floss) due to its ability to kill halitosis and gum disease-causing bacteria [27]. It has also been used in food preservation, as it can help to inhibit the growth of spoilage-causing microorganisms and extend the shelf life of food products [28]. Other potential applications of mint oil include its use in wound healing and skin care products. Mint oil has been found to have anti-inflammatory [29] and wound-healing properties [30,31]. Additionally, mint oil has been shown to have insecticidal properties [32,33], making it a potential natural alternative to synthetic insecticides.
Similar to peppermint oil, there is a plethora of research on the antimicrobial effects of Japanese mint [34,35], bergamot mint [36], and spearmint [16,18,19,20,37] EOs. Peppermint and spearmint EOs have been reported to have cleansing activity that may help protect the teeth [14,15,16,17,18,19,20]. While more research is needed to fully understand the antimicrobial effects of mint oils and their potential applications, the existing evidence suggests that they have excellent potential as a natural and effective way to combat harmful microorganisms. To the best of our knowledge, this work is the first study on the oral microbiome in Egypt. A major part of the novelty of this study is identifying the oral microbiome before and after the consumption of natural antimicrobials. In this pilot study, our objective was to evaluate the effects of SM beadlets on the oral microbiome profile in Egyptian youth using the 16S rRNA gene sequencing technique.
2. Materials and Methods
2.1. Essential Oil Chemistry
SM is a proprietary blend (dōTERRA®, Pleasant Grove, Utah). The volatile constituents were analyzed by a Shimadzu GC–MS-QP2010 Ultra (Shimadzu Scientific Instruments, Columbia, MD, USA) with electron impact (EI) mode with 70 eV, using 40–400 m/z range scans with a scan rate of 3.0 scan/sec as previously described [38]. Compound identification was performed by comparing mass spectral fragmentation patterns (over 80% similarity match) and retention indices (RI) based on a series of homologous C8-C20 n-alkanes with those reported in databases [NIST database and our in-house library] using the Lab Solutions GCMS post-run analysis software version 4.45 (Shimadzu Scientific Instruments, Columbia, MD, USA). The Gas Chromatography–Flame Ionization Detection (GC–FID) analysis was performed using a Shimadzu GC 2010 equipped with a flame ionization detector (Shimadzu Scientific Instruments, Columbia, MD, USA), as previously described [38], with a ZB-5 capillary column (Phenomenex, Torrance, CA, USA).
2.2. Ethical Review and Informed Consent
The Institutional Ethics Committee of the Faculty of Pharmacy, Suez Canal University, approved the study (IRB 2022207R1) in accordance with the 1964 Helsinki Declaration and its later amendments and comparable ethical standards. Written informed consent was obtained from all participants.
2.3. Study Population
For this investigation, a total of 20 healthy participants (out of 25 volunteers), including 10 males and 10 females, were recruited from the Faculty of Science (Suez University) and Faculty of Pharmacy (Suez Canal University) from Jan 2023 to Feb 2023. All participants read and signed the medical and nutritional history questionnaire along with an informed consent form. The inclusion criteria were age 18-35 years, good general health (free of systemic diseases), good oral health (free of oral pathologies), available for the duration of the study, and signed the informed consent form [39,40,41]. The exclusion criteria were age under 18 years old or above 35 years old, heart diseases or blood pressure alteration that requires medication, renal, hepatic, or gastrointestinal disease that requires medication, diabetes, sexually-transmitted diseases (STD), HIV, HCV infection, antibiotic use for the last 3 months prior to study, genetic disorders that could interfere with the evaluation of the study objectives, chronic obstructive pulmonary disease and asthma, any neoplastic lesion or cancer or paraneoplastic syndrome, current radiotherapy or chemotherapy, pregnant or lactating women, smoking or vaping, more than 8 missing teeth, accounted for by third molar extractions, teeth extracted for orthodontic purposes, teeth extracted because of trauma, or congenitally missing teeth, orthodontic appliances, tumors or significant pathology of the soft or hard tissue of the oral cavity (such as LPO, erythroplakia, leukoplakia, and candidiasis), chronic dry mouth, as assessed through questioning by an experienced clinician, clinically meaningful halitosis as determined by organoleptic assessment by and experienced clinician, diagnosis of periodontitis, untreated carious lesions or oral abscesses, and use of alcohol-containing mouthwash [39,40,41]. All data remain anonymous, and no names were associated with any data resulting from this study.
2.4. Treatment
A baseline investigation of the oral microbiome of the selected participants was performed by collecting saliva and swab samples before treatment. Four SM beadlets were chewed twice a day for 7 days. Post-administration saliva and swab samples were collected at the end of treatment.
2.5. Sampling
The sampling was performed at various locations in the oral cavity, including saliva, tooth surface, tongue, subgingival plaque around teeth, and buccal/palatal mucosa. For saliva collection, the volunteers were given 50 ml sterile collection tubes and were asked to allow 5 ml of saliva to flow naturally into the tube. An equal amount of sterile PBS was added to saliva and then centrifuged at 1,500 rpm for 10 min. The supernatant was then transferred to a new tube and centrifuged at 14,000 rpm for 20 minutes to obtain the bacterial pellet. All specimens were processed within 1 hour from collection. For sampling the soft tissue sites, swabs were used to collect samples from the tongue and buccal/palatal mucosa. Finally, toothpicks were used for the hard tissue sites to collect supra- and subgingival plaque around the teeth.
2.6. Oral DNA Extraction
Genomic DNA of the oral microbiome was isolated from the combination of saliva (bacterial pellet) and soft and hard tissue samples using the Quick—DNA Fungal/Bacterial Miniprep Kit (Zymo Research, Irvine, CA, USA). Toothpicks and swabs were rinsed in the recommended volume of BashingBead™ Buffer with 100 µL DNA shield (Zymo Research, Irvine, CA, USA). Then, pelleted cells (saliva) were resuspended in the same buffer. Proteinase K was added, and the cells were incubated at 65°C for 1 h to increase cell lysis. After that, the DNA extraction was carried out according to the manufacturer’s protocol. Extracted DNA was visualized by gel electrophoresis; the DNA concentration and purity were quantified using a Nanodrop One spectrophotometer (Thermo Scientific, Waltham, MA, USA) and then stored at −20 °C.
2.7. Amplicon library preparation and sequencing
Genomic DNA extracted from each person (20–50 ng/μL) was used as a template for the amplification of V3 and V4 regions of 16S rRNA. Briefly, amplicons were generated using Platinum® PCR SuperMix (Thermo Scientific, USA) with indexed primers. Forward Primer with Illumina adaptor (underlined) 5′TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGCCTACGGGNGGCWGCAG and Reverse Primer GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGACTACH VGGGTATCTAATCC [42]. Thermal cycling consisted of denaturation at 95 °C (3 min), followed by 30 cycles of denaturation at 95 °C (30 s), primer annealing at 60 °C (30 s), and primer extension at 72 °C (45 s), followed by extension at 72 °C for 5 min. Amplicons were analyzed by gel electrophoresis to verify size (~400 bp, including adaptor sequences). Library preparation and sequencing were performed using the Illumina MiSeq platform (Illumina) and accomplished at IGA Technology Services (Udine, Italy).
2.8. Bioinformatic Analyses and Ecological Statistics
Paired-end Illumina raw sequences were imported into Quantitative Insights into Microbial Ecology 2 (QIIME2 version 2023.2) [43]. Positional Quality plots were then used to decide the best positions for trimming and truncation. This was done using the QIIME2 command “qiime demux summarize.” Then, DADA2 [44] was used through QIIME2 by the qiime dada2 plugin for filtering out noisy sequences, correcting errors in marginal sequences, removing chimeric sequences, removing singletons, joining denoised paired-end reads, and then dereplicating these sequences, producing unique amplicon sequence variants (ASVs) feature table. The following parameters were used for the DADA2 pipeline: forward read sequences were truncated at 280, while reverse read sequences were truncated at 220. The first 24 bases were trimmed at 5′ end of each sequence for both forward and reverse sequences [45]. Taxonomy assignment of ASVs was performed based on trained RDP’s naive Bayesian classifier at 99% sequence similarity against the Greengenes 13_8 [46]. Further taxonomic classification of unrecognized features in our data set was performed using NCBI-BLAST. Quality filtered reads from all time points were submitted to the NCBI Sequence Read Archive (SRA) under BioProject PRJNA1090251. Alpha diversity rarefaction values for each sample were generated in QIIME2 using Faith’s Phylogenetic Diversity metric using the “qiime diversity alpha rarefaction” command [47]. The overall taxonomic diversity before and after treatment was estimated using different alpha diversity metrics based on species richness that estimate the number of OTUs and Shannon diversity index. Beta diversity analysis was carried out by QIIME2 to estimate both unweighted and weighted UniFrac, Bray–Curtis, distance matrix [47]. Alpha diversity (within sample variation) and beta diversity (between-sample variation) analysis were performed by running the QIIME2 scripts “qiime diversity core-metrics-phylogenetic” command. Taxonomic assignment tables generated by QIIME2 were imported into R to visualize the Shannon alpha diversity index and Weighted Unifrac beta diversity. The Vegan R package was used to calculate Shannon's index, which was visualized using ggplot2 and patchwork packages. Weighted Unifrac distance was also calculated and visualized in R using the Phyloseq package.
3. Results
3.1. Chemical Composition
SM is a proprietary blend from dōTERRA. Ingredients of the SM beadlets include Mentha piperita EO, M. canadensis EO, M. citrata EO, M. spicata EO, medium chain triglycerides, Guar gum, agar, glycerin, sodium alginate, and xylitol. The essential oil blend had a refreshing, minty scent. The volatile contents of the beadlets were analyzed by GC-MS and GC-FID. The major volatile components were menthol (46.19%), menthone (14.19%), linalool (4.40%), linalyl acetate (4.15%), menthyl acetate (3.87%), isomenthone (3.51%), carvone (3.31%), limonene (2.95%), 1,8-cineole (2.84%), neomenthol (2.37%), and β-caryophyllene (1.99%).
3.2. Demographic Features of the Study Population
Out of 25 initial volunteers and following the inclusion and exclusion criteria, twenty healthy participants (10 males and 10 females) with a mean age corresponding to 22.9 years (range 22–33) were enrolled in the study. The demographic features of the enrolled participants are summarized in Table 1. Overall, the mean number of teeth was 30.55 (range 30–32). Twenty participants used manual toothbrushes, while only four used dental floss.
3.3. Microbiome Diversity and Composition
The oral microbiome samples were assessed by high-throughput amplicon sequencing of 16S rRNA gene fragments. Six different specimens were collected from each subject including saliva, three soft tissues sampled by sterile swabs (tongue, buccal/palatal mucosa), and two hard tissues sampled by sterile curettes (supra- and sub-gingival plaque). Genomic DNA extracted from each person (20–50 ng/μL) was used as a template for the amplification of V3 and V4 region of 16S rRNA using indexed primers. Following DNA amplification, samples were analyzed using the Illumina MiSeq platform by IGA Technology Services (Udine, Italy) for the community composition.
3.3.1. Sequencing Data Profiles
After the quality trimming of Illumina MiSeq raw sequencing data of the 40 oral samples, 1,482,526 read counts were maintained. The average read counts per sample were 37,490, while the minimum and maximum counts per sample were 11,536 and 63,725, respectively. The total number of ASV features that remained was 3026. The plateau was reached at 11,500 within the rarefaction curves. This sequencing depth largely captured the oral microbiomes; the max depth was 63000 (Figure S1).
3.3.2. Taxonomical Classification
Overall, the community compositions before and after treatment by high-throughput sequencing of 16S rRNA are shown in Figure (S2). The oral microbiome contains 60 families, 82 microbial genera, and 209 species. Firmicutes, Bacteroidetes, and Proteobacteria were the most abundant phyla in all study groups, followed by Fusobacteria and Actinobacteria. The phylum Firmicutes represent 20-50% of the total microbiome, followed by Bacteroidetes, which represent 19-47%, then Proteobacteria, Fusobacteria, and Actinobacteria, as shown in Figure 1.
It was observed that Firmicutes and Bacteroidetes made up to 80% of the community in some samples before SM usage, and the abundance of Bacteroidetes decreased after treatment in the two groups. Firmicutes increased or remained constant after treatment in females, whereas Firmicutes decreased in males or remained constant (Figure 2).
Overall, Archaea made up only 0.003%, while Gram-positive and Gram-negative bacteria represented 43.4% and 56.5% of total sequences, respectively. The well-represented families in the microbiomes included Veillonellaceae, Prevotellaceae, Streptococcaceae, Pasteurellaceae, Neisseriaceae, Fusobacteriaceae, Actinomycetaceae, and Paraprevotellaceae. The relative abundance of the 8 most abundant families (except Veillonellaceae and Prevotellaceae) varied significantly among participants. Notably, the highly abundant families Prevotellaceae, Streptococcaceae, Pasteurellaceae, Actinomycetaceae, and Neisseriaceae were decreased after treatment. One notable finding was the decrease or disappearance of Spirochaetaceae, a well-known cause of bad breath, after SM treatment as shown in Figure 3. Moreover, the family Odoribacteraceae was represented by the infamous genus Odoribacter in one of the male samples. Interestingly, it decreased after SM use (Figure 3). Cyanobacteria, Xanthomonadaceae, and Bifidobacteriaceae were detected in females only.
Figure 4 summarizes the relative abundance and distribution of the microbial genera detected in the oral cavity. Comparing the prevalence of the most abundant bacterial genera in males and females before and after mint usage (37 genera representing more than 1% of total bacteria) showed a clear difference in microbial distribution among males and females before and after treatment (Figure 4).
In particular, Prevotella was the most abundant genus in the oral cavity (6 to 40% in males and 5 to 28% in females). They were followed by Veillonella and Streptococcus, which ranged from 4 to 33% and 3 to 26%, respectively. Fusobacterium, Neisseria, and Haemophilus were highly prevalent genera, ranging from 2 to 15% of the total bacteria detected. As expected, almost all the highly abundant genera decreased in about 70% of the samples in both males and females after treatment, including Porphyromonas, Prevotella, Streptococcus, Neisseria, and Haemophilus. Moreover, moderately abundant genera were decreased by SM as Actinomyces, Porphyromonas Lautropia, Actinobacillus, Aggregatibacter, Treponema, Leptotrichia, Campylobacter, and Capnocytophaga as shown in Figure 5. The patterns were not consistent in some genera, for, Veillonella was decreased in 80% of males although increased in 50%and decreased in the other females. It was also observed that Fusobacterium increased in 60% of male samples while decreasing in 90% of females.
At the species level, Prevotella melaninogenica was the most prevalent (10.5% of the total detected species), followed by Veillonella dispar, representing 7.5% of the total detected species. Haemophilus Parainfluenzae was the most abundant species belonging to the Haemophilus genus, with a relative abundance of 5%. Neisseria subflava was abundant, representing about 3.5% of the total species detected. However, Treponema, which causes bad breath, was decreased from 0.06 to 3.28% (Figure 5).
3.3.3. Alpha Diversity
The analysis of microbiome alpha-diversity values in the twenty study participants, performed by the measurement of the Shannon index, evidenced, as expected, an appreciable inter-individual difference between participants, with alpha values ranging from 3.5 to 5.5. Moreover, statistically significant differences were observed (p = 0.036), suggesting that the microbiome differed among participants. The female's microbiome was more diverse than males. On the contrary, Shannon indices showed no statistically significant difference between microbiomes associated with females and males before and after treatment (Figure 6).
3.3.4. Beta Diversity
The dissimilarities between oral microbial communities were detected by measuring beta diversity by calculating the Bray–Curtis, Jaccard indices and weighted UniFrac test. Beta diversity analyses showed the separation of samples by gender identity and SM usage, although the patterns were not consistent (Figure 7).
4. Discussion
The use of molecular 16S rRNA gene NGS techniques has enabled a more comprehensive understanding of the bacterial component of the oral microbiome compared to culture-based methods. These techniques have revealed differences between the male and female oral microbiome (HOM) and the effect of treatment on oral microbiome. In this study, we have analyzed the oral microbiome of individuals in a normal, healthy state in Ismailia and Suez, Egypt, by profiling the oral microbiota using 16S rRNA next-generation sequencing.
The present study aimed to examine the changes in the oral microbiome of healthy adult participants before and after treatment with mint, using NGS as the analysis method. To achieve this goal, we systematically analyzed the oral microbiomes of 20 healthy participants (10 males and 10 females) using NGS for human oral microbiome characterization, both on site-specific oral samples (hard and soft tissues) and on saliva specimens. Our results showed that there were 3026 operational taxonomic units (OTUs) shared with the oral microbiota in each of the saliva, hard, and soft tissues of the 20 individuals. We found five phyla in healthy individuals, including Proteobacteria, Firmicutes, Actinobacteria, Fusobacteria, and Bacteroidetes. These five phyla accounted for 75-98 % of the entire oral microbiome. Thus, in agreement with other studies, the five phyla account for 80–95% of the entire oral microbiome [48].
In the study, it was discovered that healthy individuals had over 82 microbial genera in their HOM. However, there were statistically significant differences between the enrolled participants. Interestingly, the study found no significant differences related to gender, which is consistent with previous research that also found no gender-related differences in oral microbiome diversity [48]. Additionally, no significant differences were observed when using hygiene devices (such as floss) or mint. However, the study did find that the female oral microbiome composition was more variable than that of the males after treatment, suggesting a diverse alpha-diversity value in the male versus female group. At the genus level, the study found a total of 37 distinct genera present at a 1% abundance.
The study found that there were several abundant genera present in all participants, including Prevotella, Streptococcus, Selenomonas, Veillonella, Fusobacterium, Neisseria, Haemophilus, Actinomyces, and Porphyromonas, which were previously characterized. Additionally, Corynebacterium, Campylobacter, Capnocytophaga, Treponema, Aggregatibacter, Actinobacillus, Leptotrichia, Tannerella, and TM7 were present in lower abundance. This suggests the existence of a core microbiome in the oral environment, similar to what has been proposed in other studies [48,49]. Furthermore, most oral taxa found in unrelated healthy individuals were similar, indicating the presence of a healthy microbiome [50,51,52]. Prevotella melaninogenica was found to be the most abundant species, and its abundance was higher in participants aged 25 and 32 compared to those aged 21, in line with previous studies on human microbiomes within different age groups [53]. Overall, the study identified over 200 bacterial species with high accuracy (>99%), including Prevotella melaninogenica, Haemophilus influenzae, Veillonella dispar, Neisseria subflava, and Rothia mucilaginosa.
Our study found that the subject's gender and use of SM were the primary factors influencing differences among participants. Interestingly, the data revealed that the subject was significantly associated with a diverse microbial composition. We also sought to explore the impact of microorganisms that make up the human oral microbiome in healthy conditions. These findings suggest that the microbiome of the healthy subject (HOM) has common recognizable characteristics, whatever their gender, and that SM treatment did not change the microbiome diversity while affecting their abundance. It is worth noting that menthol, the major EO constituent in this study, has been shown to possess several biological properties, including antimicrobial, anticancer, and anti-inflammatory activities [18,19,20,21,25,26,29].
According to our results, SM treatment decreased the abundance of Streptococcus, Veillonella, Actinomyces, and Leptotrichia. These genera were previously reported as abundant in severe early childhood caries subjects [54]. Additionally, several studies on halitosis and periodontal disease-causing bacteria have identified Actinomyces spp., Bacteroides spp., Dialister spp., Eubacterium spp., Fusobacterium spp., Leptotrichia spp., Peptostreptococcus spp., Porphyromonas spp., Prevotella spp., Selenomonas spp., Solobacterium spp., Tannerella forsythia, and Veillonella spp. as bacteria that are most related to halitosis and periodontal diseases [55,56,57]. Notably, all of these genera were affected and decreased after using mint. Moreover, Streptococcus spp., the most common pathogen involved in dental caries pathogenesis, also decreased prevalence after mint usage. Our findings align with previous research indicating that menthol is effective against various bacteria, such as S. aureus, S. mutans, S. faecalis, S. pyogenes, and L. acidophilus [58,59]. Furthermore, Leptotrichia spp., known to ferment carbohydrates and produce lactic acid that may be involved with tooth decay, decreased after treatment. Rothia dentocariosa and R. aeria were present in the oral cavity before mint usage, comprising 0.2% to 5% of the content, but decreased after mint usage to range from 0.1% to 2.7%, approximately half of the initial content. While the genus Gemella increased after using mint, it was observed that Gemella species have clear ecological preferences in the oral cavity of healthy humans that maintain health in a state of equilibrium [60]. Our findings suggest that Gemella sp. increased while Porphyromonas gingivalis decreased in alignment with previous studies. It was reported previously that G. hemolysis inhibited the growth of P. gingivalis, a species closely related to periodontal disease and halitosis, in vitro [61]. Interestingly, Odoribacter, known for producing high amounts of volatile organic compounds (VOCs) that cause unpleasant odors, decreased after using SM.
Recently, oral corynebacteria have gained significant attention due to their crucial role in the oral microbiome's biogeography [62]. According to further metagenomic sequencing analyses, Corynebacterium spp., particularly C. matruchotii, are among the most common species found in adults, suggesting that Corynebacterium sp. is part of the core oral commensals [63]. It is worth noting that Corynebacterium species are present in the conjunctiva of healthy adults and are non-pathogenic. Our results suggest new interactions between oral commensals, as the synergism between Corynebacterium and Streptococcus sanguinis reveals that Corynebacteria increased while Streptococcus decreased.
5. Conclusions
The current study successfully evaluated the effects of SM beadlets on the human oral microbiome. The results showed a significantly decreased abundance of Prevotella, Streptococcus, Neisseria, and Haemophilus microbial genera and families after treatment. Although some genera showed inconsistent patterns, it was observed that SM treatment effectively reduced the abundance of several bacteria associated with halitosis and periodontal disease, such as Actinomyces and Streptococcus. Moreover, Corynebacterium species increased after treatment. These findings strongly suggest that SM beadlets have the potential to improve oral health by reducing harmful bacteria that cause dental diseases. However, further research is needed to fully understand the antimicrobial effects of mint oils and their potential applications in maintaining good oral health.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Alpha-rarefaction curve; Figure S2: Taxonomic composition of oral microbiome.
Author Contributions
Conceptualization, S.A., N.D., P.S., and L.E.; methodology, S.A. and L.E.; software, S.A. and M.E.; validation, S.A. and M.E.; formal analysis, S.A. and M.E.; investigation, S.A. and L.E.; resources, N.D., P.S., and A.H.; data curation, S.A. and L.E.; writing—original draft preparation, S.A. and N.D.; writing—review and editing, S.A., N.D., P.S., M.E., A.H., and L.E.; visualization, M.E.; supervision, N.D., P.S., and A.H.; project administration, N.D., P.S., and A.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data generated from this study are available within this article and the supplementary material.
Acknowledgments
We thank Scott Johnson for his valuable feedback and Alex DaBell for providing the SM beadlets.
Conflicts of Interest
The authors declare no conflicts of interest. dōTERRA International had no role in the design of the clinical study, the collection, analysis, or interpretation of data, the writing of the manuscript, or the decision to publish the results.
References
- Ursell, L.K.; Metcalf, J.L.; Parfrey, L.W.; Knight, R. Defining the Human Microbiome. Nutr Rev 2012, 70, S38–S44. [Google Scholar] [CrossRef]
- Lloyd-Price, J.; Abu-Ali, G.; Huttenhower, C. The Healthy Human Microbiome. Genome Med 2016, 8, 51. [Google Scholar] [CrossRef]
- Grice, E.A.; Segre, J.A. The Human Microbiome: Our Second Genome. Annu Rev Genomics Hum Genet 2012, 13, 151–170. [Google Scholar] [CrossRef]
- Deo, P.N.; Deshmukh, R. Oral Microbiome: Unveiling the Fundamentals. J Oral Maxillofac Pathol 2019, 23, 122–128. [Google Scholar] [CrossRef]
- Sedghi, L.; DiMassa, V.; Harrington, A.; Lynch, S.V.; Kapila, Y.L. The Oral Microbiome: Role of Key Organisms and Complex Networks in Oral Health and Disease. Periodontol 2000 2021, 87, 107–131. [Google Scholar] [CrossRef] [PubMed]
- Santonocito, S.; Giudice, A.; Polizzi, A.; Troiano, G.; Merlo, E.M.; Sclafani, R.; Grosso, G.; Isola, G. A Cross-Talk between Diet and the Oral Microbiome: Balance of Nutrition on Inflammation and Immune System’s Response during Periodontitis. Nutrients 2022, 14, 2426. [Google Scholar] [CrossRef]
- Maier, T. Oral Microbiome in Health and Disease: Maintaining a Healthy, Balanced Ecosystem and Reversing Dysbiosis. Microorganisms 2023, 11, 1453. [Google Scholar] [CrossRef] [PubMed]
- Bui, F.Q.; Almeida-da-Silva, C.L.C.; Huynh, B.; Trinh, A.; Liu, J.; Woodward, J.; Asadi, H.; Ojcius, D.M. Association between Periodontal Pathogens and Systemic Disease. Biomed J 2019, 42, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Matayoshi, S.; Tojo, F.; Suehiro, Y.; Okuda, M.; Takagi, M.; Ochiai, M.; Kadono, M.; Mikasa, Y.; Okawa, R.; Nomura, R.; et al. Effects of Mouthwash on Periodontal Pathogens and Glycemic Control in Patients with Type 2 Diabetes Mellitus. Sci Rep 2024, 14, 2777. [Google Scholar] [CrossRef]
- Joshipura, K.J.; Muñoz-Torres, F.J.; Morou-Bermudez, E.; Patel, R.P. Over-the-Counter Mouthwash Use and Risk of Pre-Diabetes/Diabetes. Nitric Oxide 2017, 71, 14–20. [Google Scholar] [CrossRef]
- Wang, S.; Song, F.; Gu, H.; Wei, X.; Zhang, K.; Zhou, Y.; Luo, H. Comparative Evaluation of the Salivary and Buccal Mucosal Microbiota by 16S RRNA Sequencing for Forensic Investigations. Front Microbiol 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Wade, W.G.; Prosdocimi, E.M. Profiling of Oral Bacterial Communities. J Dent Res 2020, 99, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.L.; Morton, J.T.; Dinis, M.; Alvarez, R.; Tran, N.C.; Knight, R.; Edlund, A. Deep Metagenomics Examines the Oral Microbiome during Dental Caries, Revealing Novel Taxa and Co-Occurrences with Host Molecules. Genome Res 2021, 31, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Işcan, G.; Kirimer, N.; Kürkcüoğlu, M.; Başer, K.H.C.; Demirci, F. Antimicrobial Screening of Mentha Piperita Essential Oils. J. Agric. Food Chem. 2002, 50, 3943–3946. [Google Scholar] [CrossRef] [PubMed]
- Inouye, S.; Yamaguchi, H.; Takizawa, T. Screening of the Antibacterial Effects of a Variety of Essential Oils on Respiratory Tract Pathogens, Using a Modified Dilution Assay Method. Journal of Infection and Chemotherapy 2001, 7, 251–254. [Google Scholar] [CrossRef] [PubMed]
- Soković, M.; Glamočlija, J.; Marin, P.D.; Brkić, D.; Griensven, L.J.L.D. van Antibacterial Effects of the Essential Oils of Commonly Consumed Medicinal Herbs Using an In Vitro Model. Molecules 2010, 15, 7532–7546. [Google Scholar] [CrossRef] [PubMed]
- Inouye, S.; Takizawa, T.; Yamaguchi, H. Antibacterial Activity of Essential Oils and Their Major Constituents against Respiratory Tract Pathogens by Gaseous Contact. Journal of Antimicrobial Chemotherapy 2001, 47, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Mandava, K.; Batchu, U.R.; Kakulavaram, S.; Repally, S.; Chennuri, I.; Bedarakota, S.; Sunkara, N. Design and Study of Anticaries Effect of Different Medicinal Plants against S.Mutans Glucosyltransferase. BMC Complement Altern Med 2019, 19, 197. [Google Scholar] [CrossRef]
- Fitsiou, E.; Mitropoulou, G.; Spyridopoulou, K.; Tiptiri-Kourpeti, A.; Vamvakias, M.; Bardouki, H.; Panayiotidis, M.; Galanis, A.; Kourkoutas, Y.; Chlichlia, K.; et al. Phytochemical Profile and Evaluation of the Biological Activities of Essential Oils Derived from the Greek Aromatic Plant Species Ocimum Basilicum, Mentha Spicata, Pimpinella Anisum and Fortunella Margarita. Molecules 2016, 21, 1069. [Google Scholar] [CrossRef]
- Imai, H.; Osawa, K.; Yasuda, H.; Hamashima, H.; Arai, T.; Sasatsu, M. Inhibition by the Essential Oils of Peppermint and Spearmint of the Growth of Pathogenic Bacteria. Microbios 2001, 106 Suppl 1, 31–39. [Google Scholar]
- Mimica-Dukić, N.; Bozin, B.; Soković, M.; Mihajlović, B.; Matavulj, M. Antimicrobial and Antioxidant Activities of Three Mentha Species Essential Oils. Planta Med 2003, 69, 413–419. [Google Scholar] [CrossRef]
- Boren, K.; Crown, A.; Carlson, R. Multidrug and Pan-Antibiotic Resistance—The Role of Antimicrobial and Synergistic Essential Oils: A Review. Nat Prod Commun 2020, 15, 1934578X2096259. [Google Scholar] [CrossRef]
- Muntean, D.; Licker, M.; Alexa, E.; Popescu, I.; Jianu, C.; Buda, V.; Dehelean, C.A.; Ghiulai, R.; Horhat, F.; Horhat, D.; et al. <p>Evaluation of Essential Oil Obtained from <em>Mentha×piperita</Em> L. against Multidrug-Resistant Strains</P>. Infect Drug Resist. [CrossRef]
- Jeyakumar, E.; Lawrence, R.; Pal, T. Comparative Evaluation in the Efficacy of Peppermint (Mentha Piperita) Oil with Standards Antibiotics against Selected Bacterial Pathogens. Asian Pac J Trop Biomed 2011, 1, S253–S257. [Google Scholar] [CrossRef]
- Kotan, R.; Kordali, S.; Cakir, A. Screening of Antibacterial Activities of Twenty-One Oxygenated Monoterpenes. Zeitschrift für Naturforschung C 2007, 62, 507–513. [Google Scholar] [CrossRef]
- Iraji, A.; Yazdanpanah, S.; Alizadeh, F.; Mirzamohammadi, S.; Ghasemi, Y.; Pakshir, K.; Yang, Y.; Zomorodian, K. Screening the Antifungal Activities of Monoterpenes and Their Isomers against Candida Species. J Appl Microbiol 2020, 129, 1541–1551. [Google Scholar] [CrossRef] [PubMed]
- Haghgoo, R.; Abbasi, F. Evaluation of the Use of a Peppermint Mouth Rinse for Halitosis by Girls Studying in Tehran High Schools. J Int Soc Prev Community Dent 2013, 3, 29. [Google Scholar] [CrossRef]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential Oils in Food Preservation: Mode of Action, Synergies, and Interactions with Food Matrix Components. Front Microbiol 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Park, S.J.; Yang, W.M. Inhalation of Essential Oil from Mentha Piperita Ameliorates PM10-Exposed Asthma by Targeting IL-6/JAK2/STAT3 Pathway Based on a Network Pharmacological Analysis. Pharmaceuticals 2020, 14, 2. [Google Scholar] [CrossRef] [PubMed]
- Ghodrati, M.; Farahpour, M.R.; Hamishehkar, H. Encapsulation of Peppermint Essential Oil in Nanostructured Lipid Carriers: In-Vitro Antibacterial Activity and Accelerative Effect on Infected Wound Healing. Colloids Surf A Physicochem Eng Asp 2019, 564, 161–169. [Google Scholar] [CrossRef]
- Modarresi, M.; Farahpour, M.-R.; Baradaran, B. Topical Application of Mentha Piperita Essential Oil Accelerates Wound Healing in Infected Mice Model. Inflammopharmacology 2019, 27, 531–537. [Google Scholar] [CrossRef]
- Kumar, S.; Wahab, N.; Warikoo, R. Bioefficacy of Mentha Piperita Essential Oil against Dengue Fever Mosquito Aedes Aegypti L. Asian Pac J Trop Biomed 2011, 1, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Asadollahi, A.; Khoobdel, M.; Zahraei-Ramazani, A.; Azarmi, S.; Mosawi, S.H. Effectiveness of Plant-Based Repellents against Different Anopheles Species: A Systematic Review. Malar J 2019, 18, 436. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, A.K.; Malik, A. Antimicrobial Action of Essential Oil Vapours and Negative Air Ions against Pseudomonas Fluorescens. Int J Food Microbiol 2010, 143, 205–210. [Google Scholar] [CrossRef]
- Scartazzini, L.; Tosati, J.V.; Cortez, D.H.C.; Rossi, M.J.; Flôres, S.H.; Hubinger, M.D.; Di Luccio, M.; Monteiro, A.R. Gelatin Edible Coatings with Mint Essential Oil (Mentha Arvensis): Film Characterization and Antifungal Properties. J Food Sci Technol 2019, 56, 4045–4056. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.K.; Goswami, P.; Verma, R.S.; Padalia, R.C.; Chauhan, A.; Singh, V.R.; Darokar, M.P. Chemical Composition and Antimicrobial Activity of Bergamot-Mint (Mentha Citrata Ehrh.) Essential Oils Isolated from the Herbage and Aqueous Distillate Using Different Methods. Ind Crops Prod 2016, 91, 152–160. [Google Scholar] [CrossRef]
- Snoussi, M.; Noumi, E.; Trabelsi, N.; Flamini, G.; Papetti, A.; De Feo, V. Mentha Spicata Essential Oil: Chemical Composition, Antioxidant and Antibacterial Activities against Planktonic and Biofilm Cultures of Vibrio Spp. Strains. Molecules 2015, 20, 14402–14424. [Google Scholar] [CrossRef] [PubMed]
- Decarlo, A.; Johnson, S.; Ouédraogo, A.; Dosoky, N.S.; Setzer, W.N. Chemical Composition of the Oleogum Resin Essential Oils of Boswellia Dalzielii from Burkina Faso. Plants 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Brzychczy-Sroka, B.; Talaga-Ćwiertnia, K.; Sroka-Oleksiak, A.; Gurgul, A.; Zarzecka-Francica, E.; Ostrowski, W.; Kąkol, J.; Drożdż, K.; Brzychczy-Włoch, M.; Zarzecka, J. Standardization of the Protocol for Oral Cavity Examination and Collecting of the Biological Samples for Microbiome Research Using the Next-Generation Sequencing (NGS): Own Experience with the COVID-19 Patients. Sci Rep 2024, 14, 3717. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, A.J.S.; Parmar, V.; Blaser, M.J. Assessing Saliva Microbiome Collection and Processing Methods. NPJ Biofilms Microbiomes 2021, 7, 81. [Google Scholar] [CrossRef] [PubMed]
- Caselli, E.; Fabbri, C.; D’Accolti, M.; Soffritti, I.; Bassi, C.; Mazzacane, S.; Franchi, M. Defining the Oral Microbiome by Whole-Genome Sequencing and Resistome Analysis: The Complexity of the Healthy Picture. BMC Microbiol 2020, 20, 120. [Google Scholar] [CrossRef]
- Illumina 16S Metagenomic Sequencing Library Preparation: Preparing 16S Ribosomal RNA Gene Amplicons for the Illumina MiSeq System. Part No. 15044223 Rev B. Illumina, San Diego, CA, USA. Available online: https://www.illumina.com/content/dam/illumina-support/documents/documentation/chemistry_documentation/16s/16s-metagenomic-library-prep-guide-15044223-b.pdf (accessed on 5 December 2020).
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat Biotechnol 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Mohsen, A.; Park, J.; Chen, Y.-A.; Kawashima, H.; Mizuguchi, K. Impact of Quality Trimming on the Efficiency of Reads Joining and Diversity Analysis of Illumina Paired-End Reads in the Context of QIIME1 and QIIME2 Microbiome Analysis Frameworks. BMC Bioinformatics 2019, 20, 581. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a Chimera-Checked 16S RRNA Gene Database and Workbench Compatible with ARB. Appl Environ Microbiol 2006, 72, 5069–5072. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-High-Throughput Microbial Community Analysis on the Illumina HiSeq and MiSeq Platforms. ISME J 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Jiang, W. Application of High-Throughput Sequencing in Understanding Human Oral Microbiome Related with Health and Disease. Front Microbiol 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Shade, A.; Handelsman, J. Beyond the Venn Diagram: The Hunt for a Core Microbiome. Environ Microbiol 2012, 14, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Zaura, E.; Keijser, B.J.; Huse, S.M.; Crielaard, W. Defining the Healthy “Core Microbiome” of Oral Microbial Communities. BMC Microbiol 2009, 9, 259. [Google Scholar] [CrossRef]
- Avila, M.; Ojcius, D.M.; Yilmaz, Ö. The Oral Microbiota: Living with a Permanent Guest. DNA Cell Biol 2009, 28, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Aas, J.A.; Paster, B.J.; Stokes, L.N.; Olsen, I.; Dewhirst, F.E. Defining the Normal Bacterial Flora of the Oral Cavity. J Clin Microbiol 2005, 43, 5721–5732. [Google Scholar] [CrossRef]
- Crielaard, W.; Zaura, E.; Schuller, A.A.; Huse, S.M.; Montijn, R.C.; Keijser, B.J. Exploring the Oral Microbiota of Children at Various Developmental Stages of Their Dentition in the Relation to Their Oral Health. BMC Med Genomics 2011, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Ling, Z.; Kong, J.; Jia, P.; Wei, C.; Wang, Y.; Pan, Z.; Huang, W.; Li, L.; Chen, H.; Xiang, C. Analysis of Oral Microbiota in Children with Dental Caries by PCR-DGGE and Barcoded Pyrosequencing. Microb Ecol 2010, 60, 677–690. [Google Scholar] [CrossRef] [PubMed]
- Musić, L.; Par, M.; Peručić, J.; Badovinac, A.; Plančak, D.; Puhar, I. Relationship Between Halitosis and Periodontitis: A Pilot Study. Acta Stomatol Croat 2021, 55, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-H.; Hong, J.-Y. Oral Microbiome as a Co-Mediator of Halitosis and Periodontitis: A Narrative Review. Frontiers in Oral Health 2023, 4. [Google Scholar] [CrossRef] [PubMed]
- Persson, S.; Edlund, M.; Claesson, R.; Carlsson, J. The Formation of Hydrogen Sulfide and Methyl Mercaptan by Oral Bacteria. Oral Microbiol Immunol 1990, 5, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Al-Bayati, F.A. Isolation and Identification of Antimicrobial Compound from Mentha Longifolia L. Leaves Grown Wild in Iraq. Ann Clin Microbiol Antimicrob 2009, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Yang, C.; Zhang, N.; Peng, Y.; Ma, Y.; Gu, K.; Liu, X.; Liu, X.; Liu, X.; Liu, Y.; et al. Menthone Exerts Its Antimicrobial Activity Against Methicillin Resistant Staphylococcus Aureus by Affecting Cell Membrane Properties and Lipid Profile. Drug Des Devel Ther 2023, Volume 17, 219–236. [Google Scholar] [CrossRef]
- Torres-Morales, J.; Mark Welch, J.L.; Dewhirst, F.E.; Borisy, G.G. Site-Specialization of Human Oral Gemella Species. J Oral Microbiol 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, T.; Oge, S.; Nakata, S.; Ueno, Y.; Ukita, H.; Kousaka, R.; Miura, Y.; Yoshinari, N.; Yoshida, A. Gemella Haemolysans Inhibits the Growth of the Periodontal Pathogen Porphyromonas Gingivalis. Sci Rep 2021, 11, 11742. [Google Scholar] [CrossRef]
- Mark Welch, J.L.; Rossetti, B.J.; Rieken, C.W.; Dewhirst, F.E.; Borisy, G.G. Biogeography of a Human Oral Microbiome at the Micron Scale. Proc Natl Acad Sci U S A 2016, 113, E791–800. [Google Scholar] [CrossRef]
- Esberg, A.; Barone, A.; Eriksson, L.; Lif Holgerson, P.; Teneberg, S.; Johansson, I. Corynebacterium Matruchotii Demography and Adhesion Determinants in the Oral Cavity of Healthy Individuals. Microorganisms 2020, 8. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Relative abundance and distribution of the highest microbial phyla detected in the oral cavity. A) Percentage distribution of detected phyla in males. B) Percentage distribution of detected phyla in females. Participant IDs with B and A are before and after treatment, respectively.
Figure 1.
Relative abundance and distribution of the highest microbial phyla detected in the oral cavity. A) Percentage distribution of detected phyla in males. B) Percentage distribution of detected phyla in females. Participant IDs with B and A are before and after treatment, respectively.
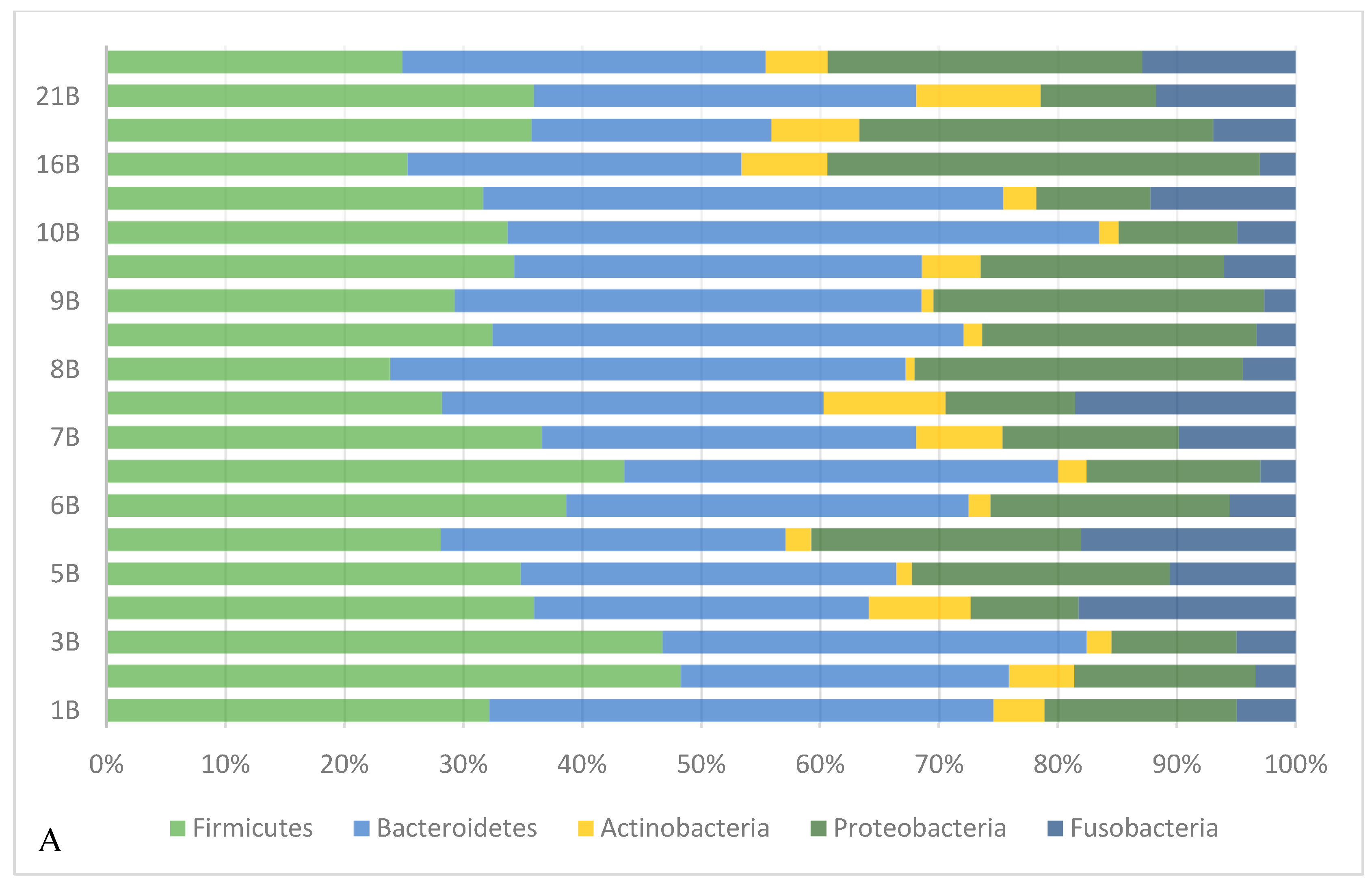
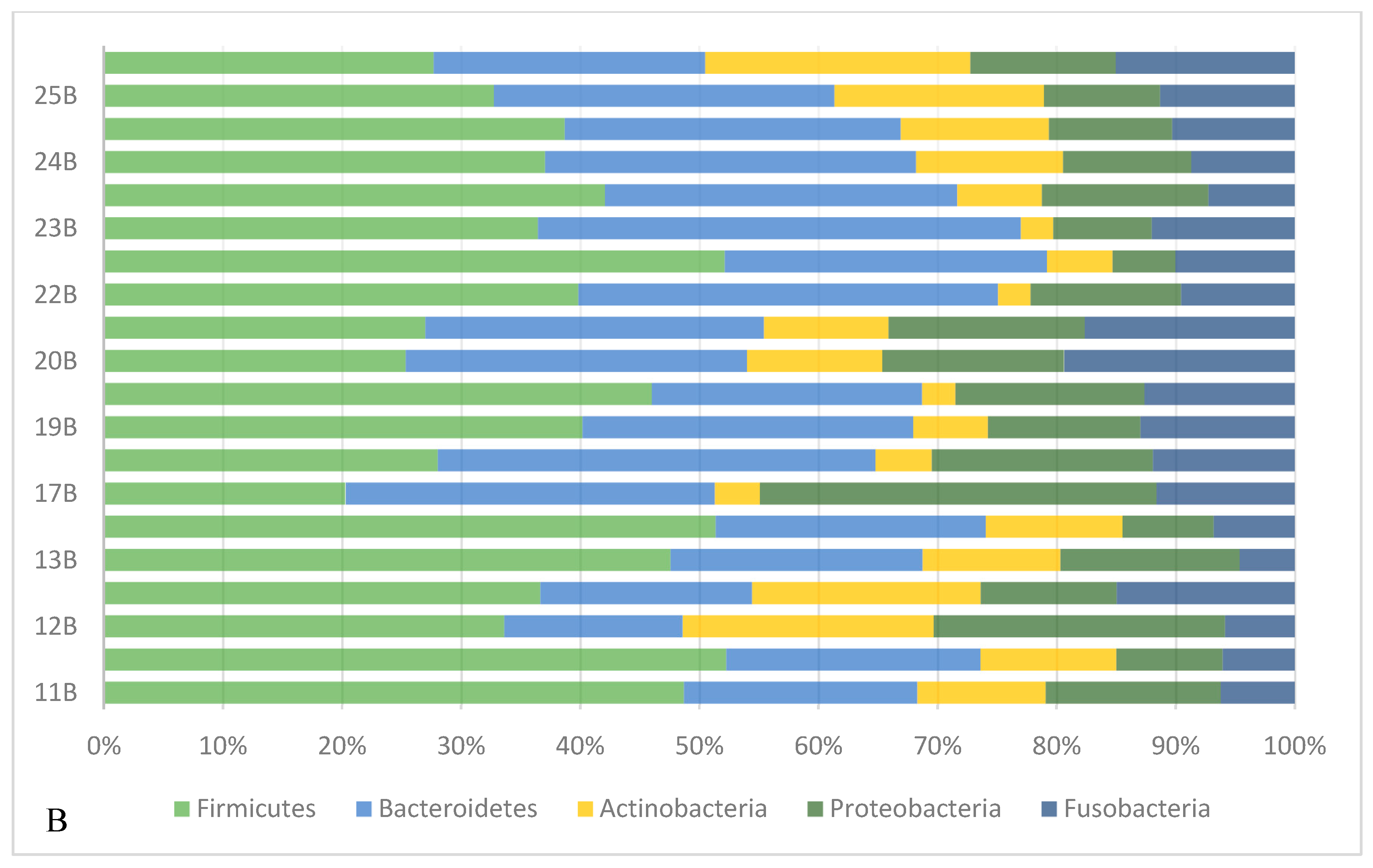
Figure 2.
Relative abundance of the two taxa Firmicutes and Bacteroidetes detected in the oral cavity before and after mint usage. A) Male. B) Female. Participant IDs with B and A are before and after treatment, respectively.
Figure 2.
Relative abundance of the two taxa Firmicutes and Bacteroidetes detected in the oral cavity before and after mint usage. A) Male. B) Female. Participant IDs with B and A are before and after treatment, respectively.
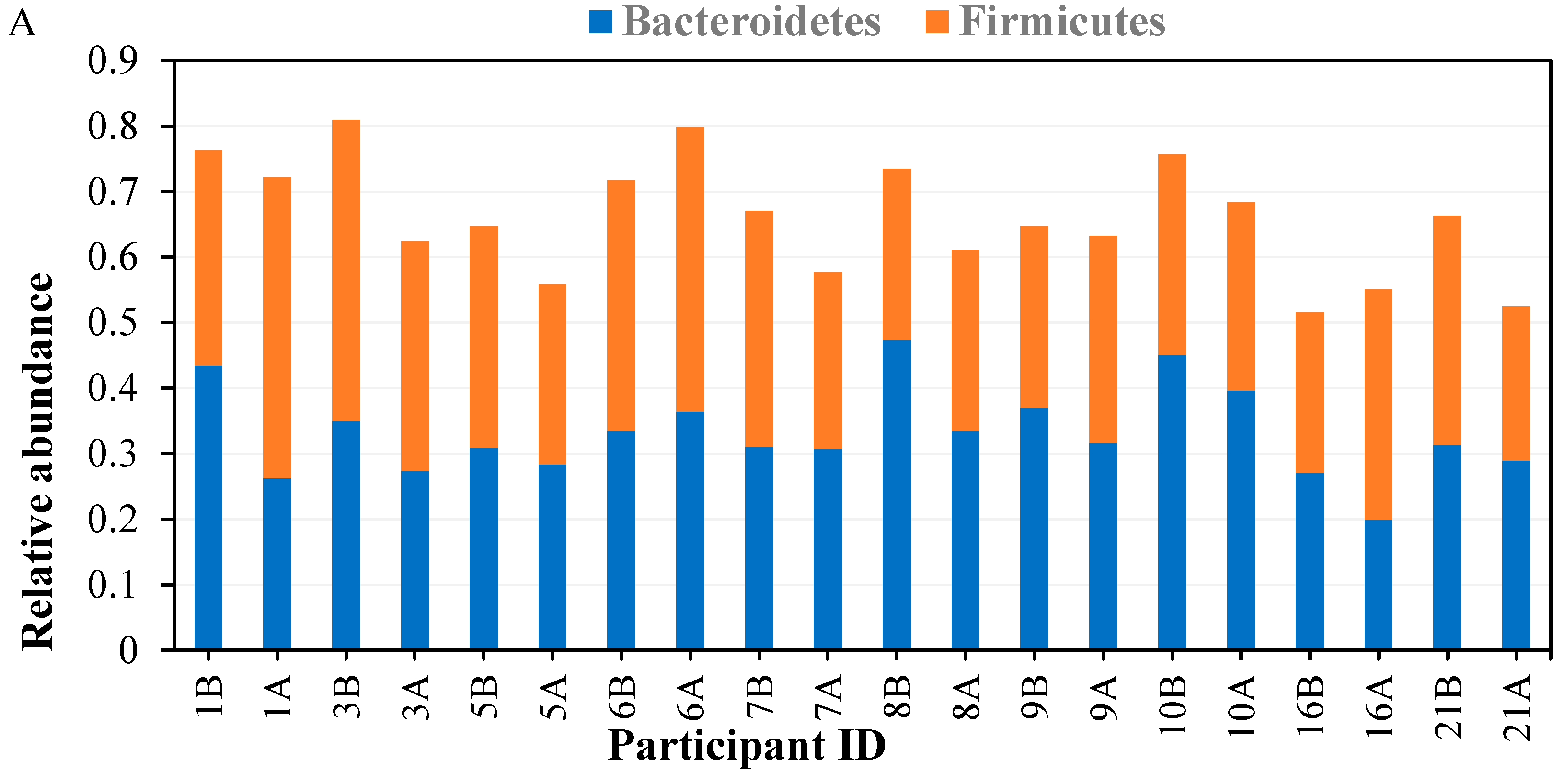
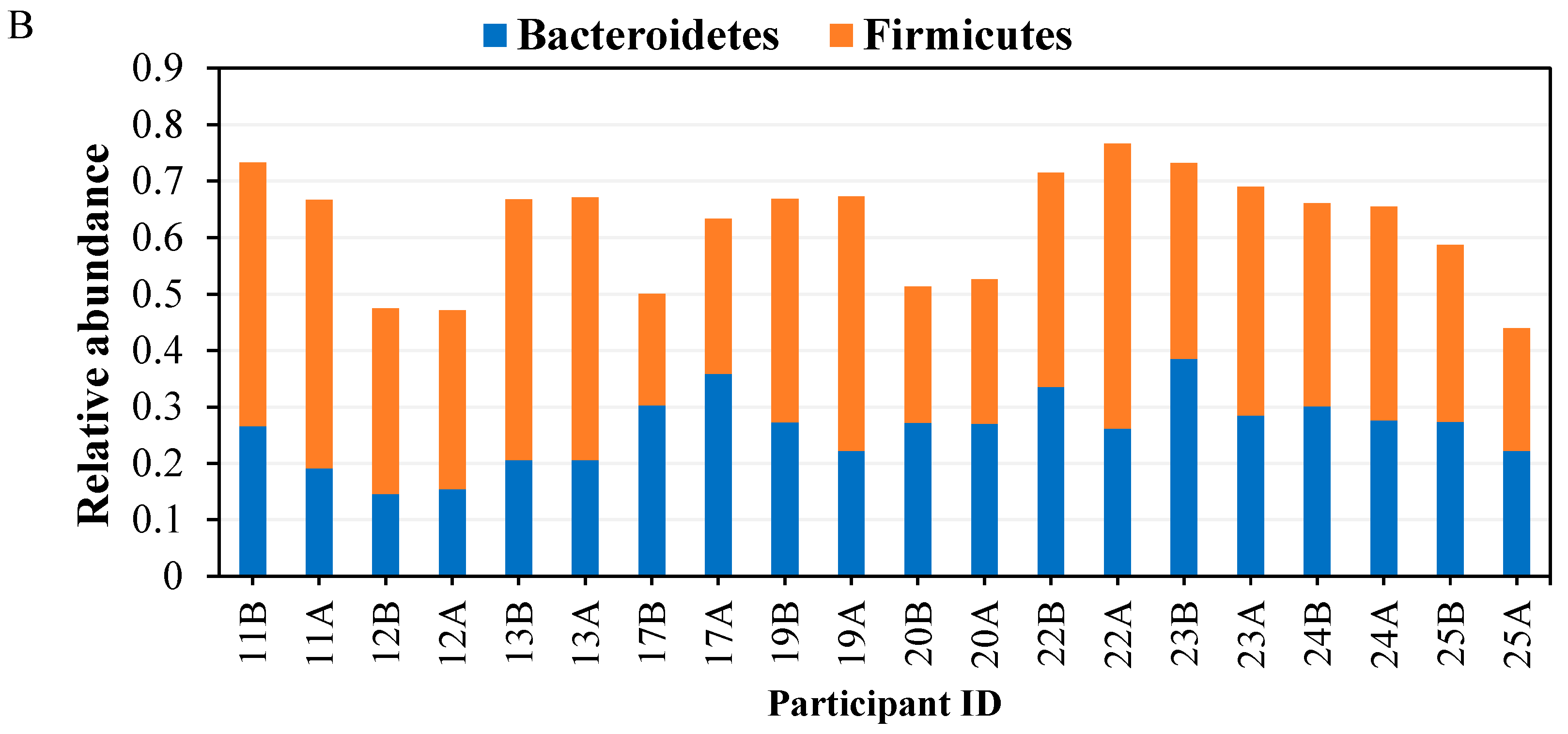
Figure 3.
Barplot representation of families detected by 16S rRNA analysis in the clustered before and after treatment.
Figure 3.
Barplot representation of families detected by 16S rRNA analysis in the clustered before and after treatment.
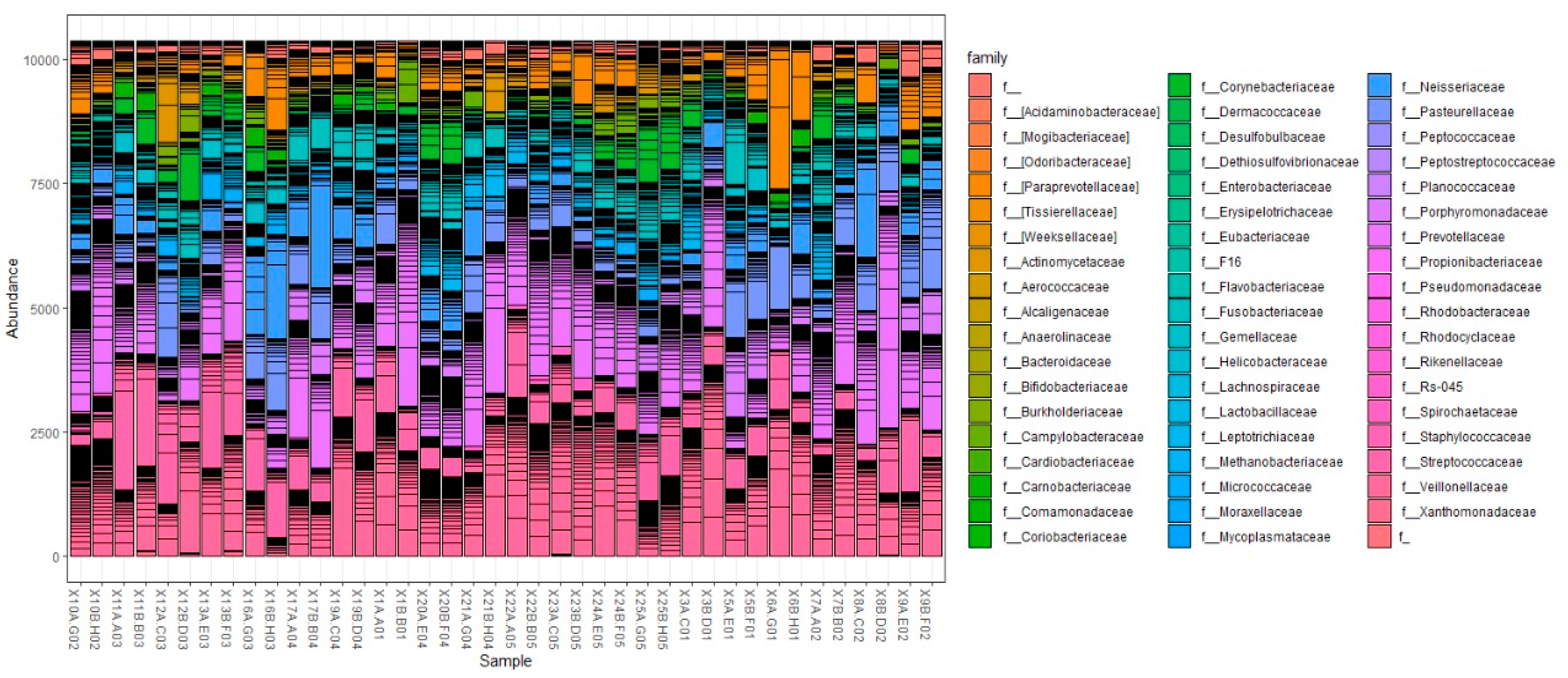
Figure 4.
Heatmap representation of genera (with relative abundance of more than 1%) detected by 16S rRNA community analysis in each sample from each enrolled subject before and after treatment.
Figure 4.
Heatmap representation of genera (with relative abundance of more than 1%) detected by 16S rRNA community analysis in each sample from each enrolled subject before and after treatment.
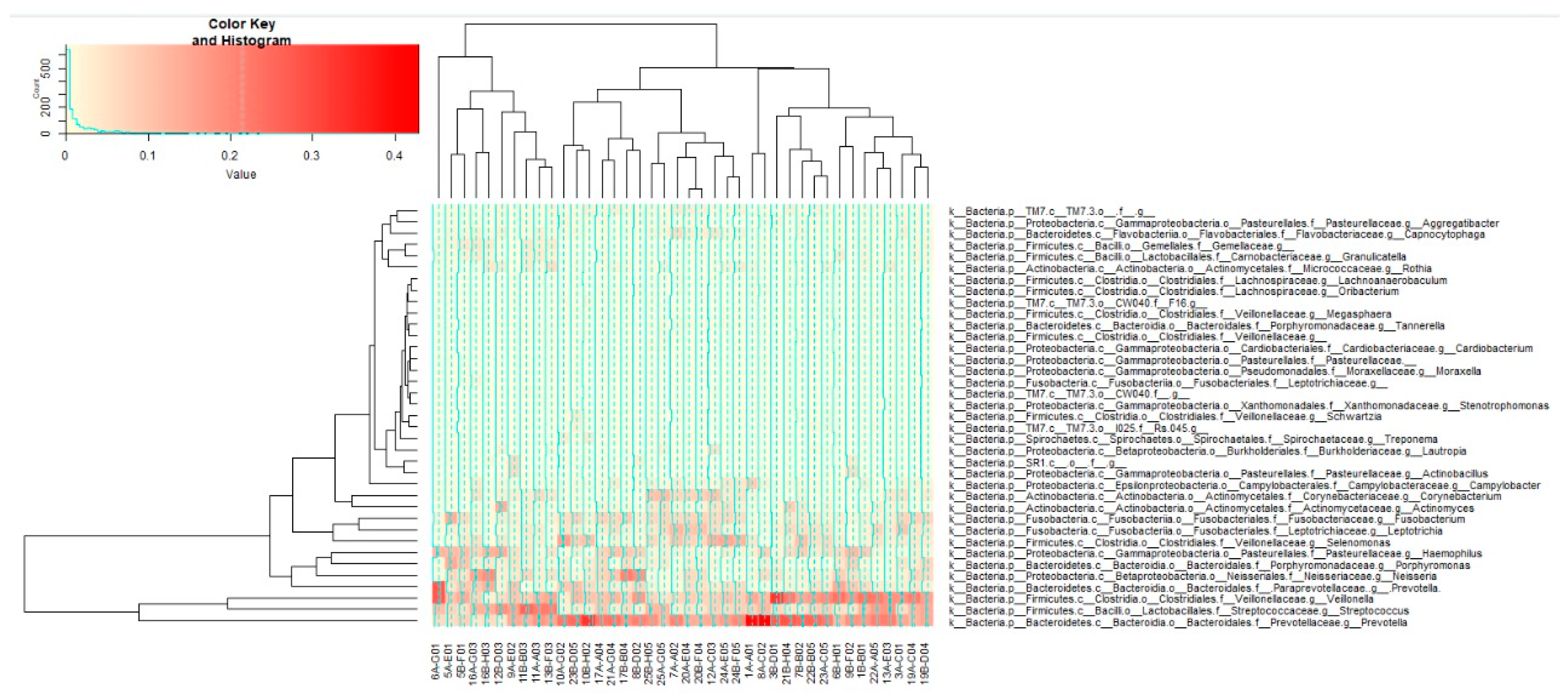
Figure 5.
Relative abundance of taxa responsible for carries, halitosis, and periodontal disease across samples. (A) at genus level in males before and after treatment for highly abundant genera, (B) at genus level in males before and after treatment for moderately abundant genera, (C) at genus level in females before and after treatment for highly abundant genera, and (D) at genus level in males before and after treatment for moderately abundant genera.
Figure 5.
Relative abundance of taxa responsible for carries, halitosis, and periodontal disease across samples. (A) at genus level in males before and after treatment for highly abundant genera, (B) at genus level in males before and after treatment for moderately abundant genera, (C) at genus level in females before and after treatment for highly abundant genera, and (D) at genus level in males before and after treatment for moderately abundant genera.
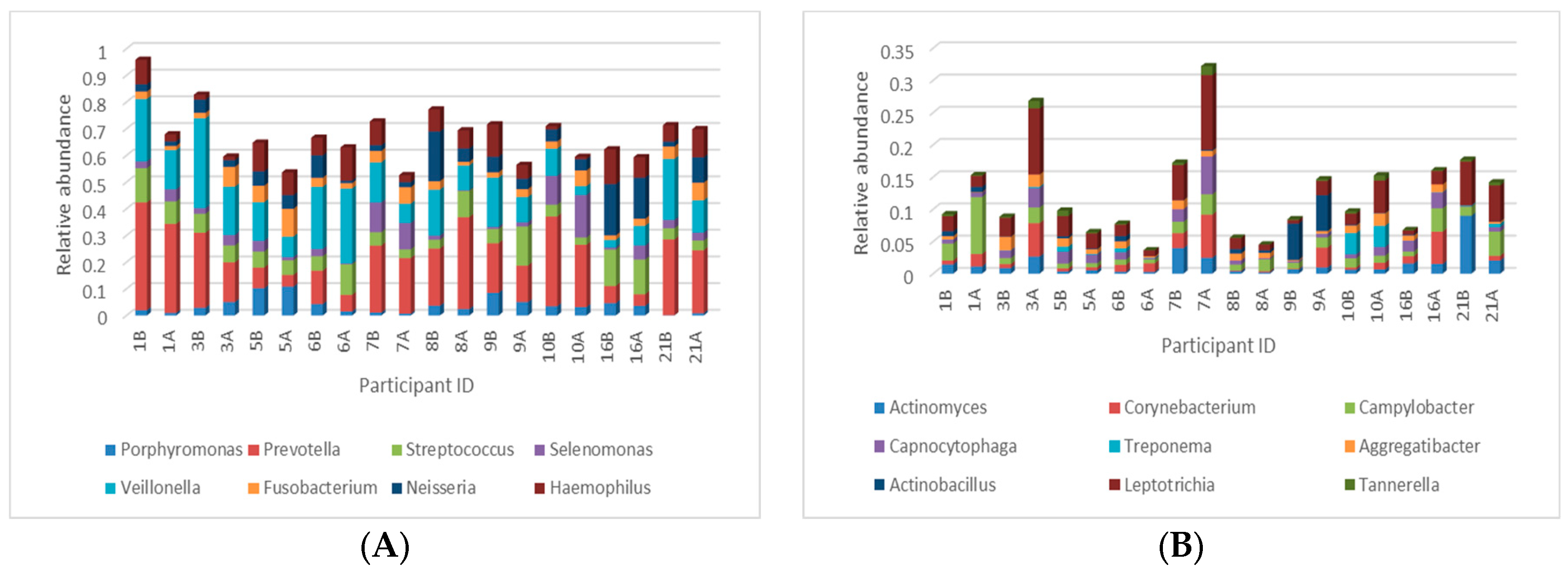
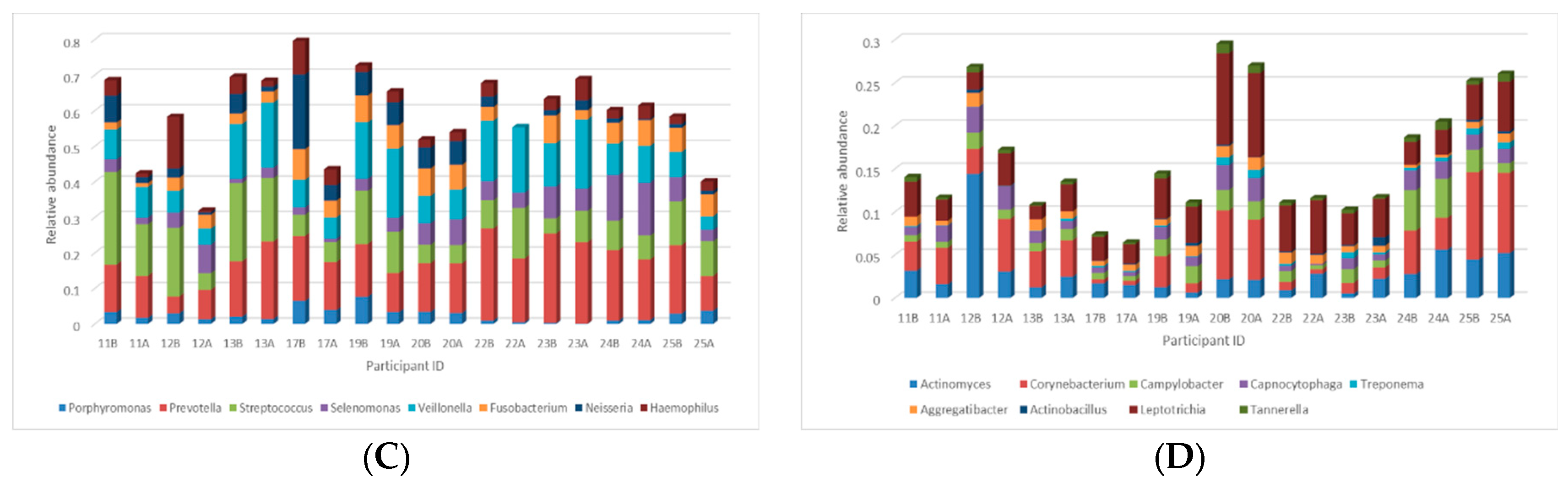
Figure 6.
Alpha diversity indices of oral microbial communities across samples. Alpha diversity, as calculated by the Shannon alpha diversity, differed by (A) gender, (B) before and after using mint, and (C) across samples. The only significant differences were between different participants. .
Figure 6.
Alpha diversity indices of oral microbial communities across samples. Alpha diversity, as calculated by the Shannon alpha diversity, differed by (A) gender, (B) before and after using mint, and (C) across samples. The only significant differences were between different participants. .
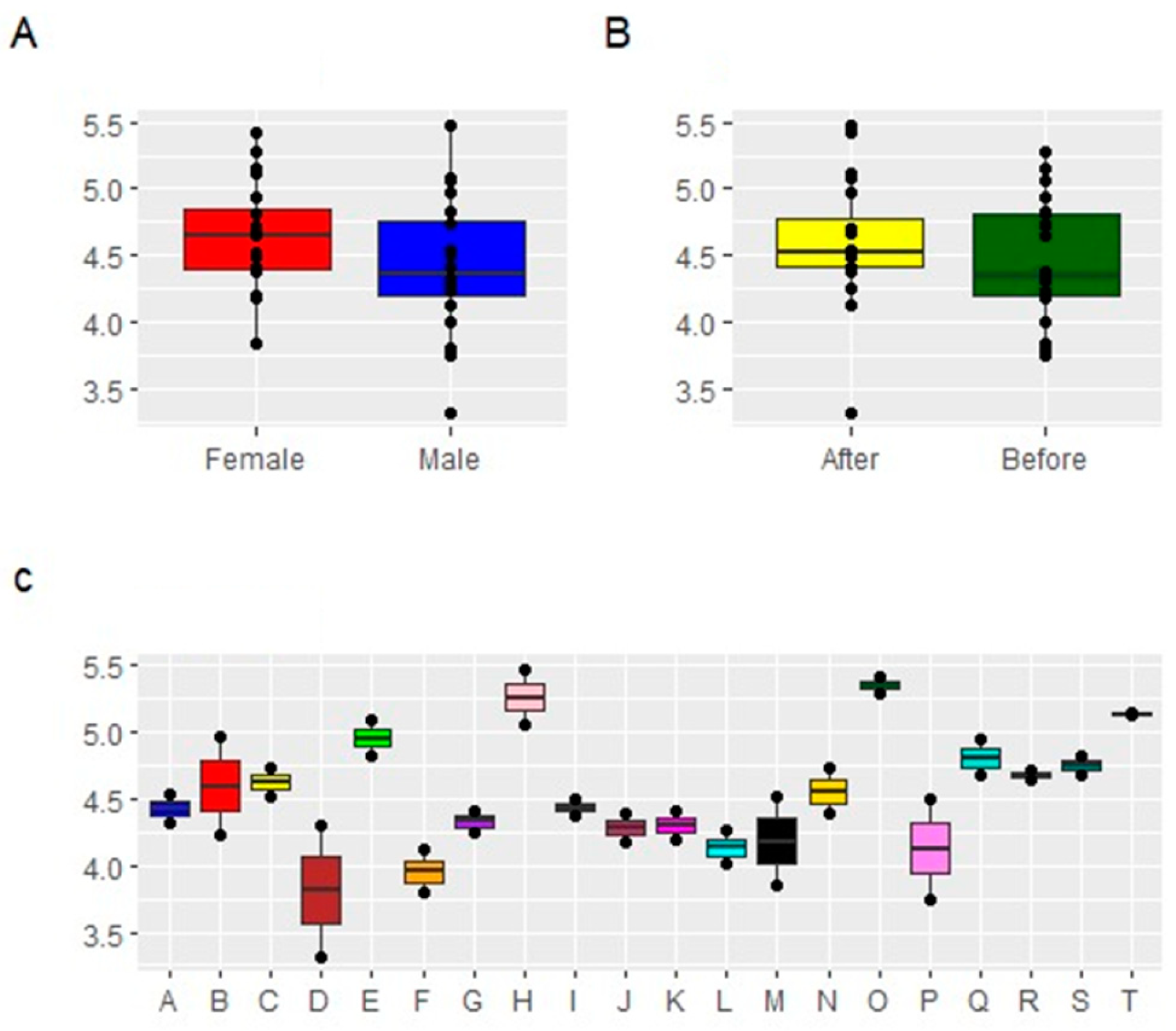
Figure 7.
Beta diversity of oral microbiomes calculated with Weighted UniFrac. (A) Gender, (B) Gender and before and after treatment. Beta diversity analysis showed samples primarily separated by gender.
Figure 7.
Beta diversity of oral microbiomes calculated with Weighted UniFrac. (A) Gender, (B) Gender and before and after treatment. Beta diversity analysis showed samples primarily separated by gender.
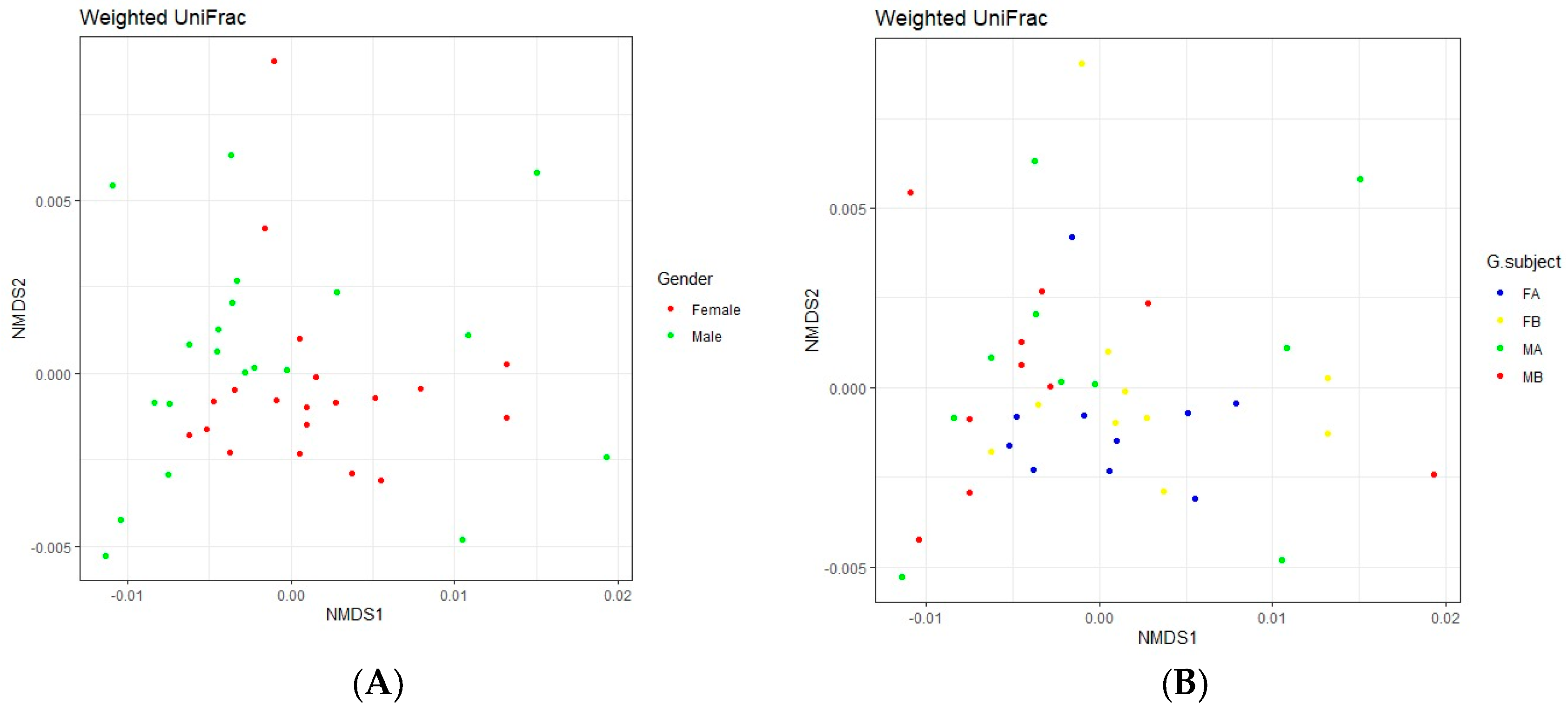

Table 1.
Demographic features of the study population.
|
Participant ID |
Gender | Age | Teeth n. |
Oral-Hygiene Devices at home |
City of residency |
| 1 | Male | 23 | 32 | None | Ismailia |
| 3 | Male | 23 | 32 | Floss | Cairo |
| 5 | Male | 24 | 32 | None | Cairo |
| 6 | Male | 23 | 30 | None | Ismailia |
| 7 | Male | 23 | 32 | None | Sinai |
| 8 | Male | 22 | 32 | None | Ismailia |
| 9 | Male | 22 | 31 | None | Ismailia |
| 10 | Male | 23 | 31 | None | Ismailia |
| 11 | Female | 21 | 32 | None | Eltour |
| 12 | Female | 21 | 32 | None | Suez |
| 13 | Female | 21 | 32 | None | Suez |
| 16 | Male | 21 | 32 | None | Suez |
| 17 | Female | 21 | 32 | None | Suez |
| 19 | Female | 21 | 31 | None | Suez |
| 20 | Female | 21 | 32 | None | Suez |
| 21 | Male | 23 | 32 | Floss | Ismailia |
| 22 | Female | 25 | 32 | None | Ismailia |
| 23 | Female | 22 | 32 | None | Ismailia |
| 24 | Female | 25 | 31 | Floss | Suez |
| 25 | Female | 33 | 32 | Floss | Ismailia |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



