
Submitted:
31 May 2024
Posted:
03 June 2024
You are already at the latest version
Abstract
The extensive metabolic diversity of microalgae, coupled with their rapid growth rates and cost-effective production, position these organisms as highly promising resources for a wide range of biotechnological applications. These characteristics allow microalgae to address crucial needs in both the agricultural, medical, and industrial sectors. Microalgae are proving to be val-uable in various fields, including the remediation of diverse wastewater types, the production of biofuels and biofertilizers, and the extraction of various products from their biomass. For decades, the microalga Chlamydomonas has been widely used as a fundamental research model organism in various areas such as photosynthesis, respiration, sulfur and phosphorus metabolism, nitrogen metabolism, and flagella synthesis, among others. However, in recent years, the potential of Chlamydomonas as a biotechnological tool for bioremediation, biofertilization, biomass, and bio-products production has been increasingly recognized. Bioremediation of wastewater using Chlamydomonas presents significant potential for sustainable reduction of contaminants and fa-cilitates resource recovery and valorization of microalgal biomass, offering important economic benefits. Chlamydomonas has also established itself as a platform for the production of a wide va-riety of biotechnologically interesting products, such as different types of biofuels, and high-value-added products. The aim of this review is to achieve a comprehensive understanding of the potential of Chlamydomonas in these aspects, and to explore their interrelationship, which would offer significant environmental and biotechnological advantages.
Keywords:
Microalga
; Chlamydomonas
; bioremediation
; wastewater
; high-value-added products
1. Introduction. Why Microalgae and Why Chlamydomonas?
Microalgae represent a broad array of single-celled, photosynthetic organisms that serve as key contributors to primary production across our planet [1]. Microalgae can adopt photoautotrophic, heterotrophic, or mixotrophic modes of life, displaying a spectrum of cell sizes, shapes, and structures. Responsible for a significant portion of the global carbon capture, microalgae play a crucial role in supporting ecosystems [2]. Microalgae share a common evolutionary origin that can be traced back to a primary endosymbiotic event involving a cyanobacterium, which eventually evolved into the plastid [3]. This process has resulted in the emergence of a wide range of colorful and metabolically diverse algal groups, such as diatoms and dinoflagellates [4]. Microalgae are employed in activities such as wastewater treatment [5], biofuel generation [6], animal feed production [7], and the extraction of high-value-added products [8], among other applications. Additionally, microalgae show great potential as organisms for enhancing biological carbon sequestration aimed at mitigating global warming [9]. Consequently, microalgae hold significant ecological and economic potential.
Chlamydomonas is a microalga that is commonly found in freshwater and saltwater habitats, as well as in soil and snow. Taxonomically, the genus Chlamydomonas comprises more than 500 species [10]. Over time, it has evolved into a highly influential model organism, thanks to its numerous interesting characteristics [11]. Among the Chlamydomonas species, Chlamydomonas reinhardtii is the most commonly used due to its interesting characteristics. Among these features, C. reinhardtii has two flagella, grows well in axenic cultures, exhibits a relatively rapid doubling time of approximately 8-12 hours, and its nuclear, chloroplast, and mitochondrial genomes are sequenced. Additionally, C. reinhardtii exhibited an exceptional ability to adapt and thrive under nearly all experimental conditions tested in heterotrophic, phototrophic, and mixotrophic cultivations [12]. Moreover, the Chlamydomonas Sourcebook [13] provides a thorough overview of essential research areas, historical background, physiology, and methodologies related to Chlamydomonas. Additionally, the Chlamydomonas Resource Centre offers a wide range of resources, including biochemical assays, protocols, plasmids, and a diverse collection of mapped mutant strains. Furthermore, enhancing the yield of many biotechnological processes involving Chlamydomonas can be achieved through synergistic interactions with other microorganisms, predominantly bacteria [14].
However, there are still numerous challenges hindering the efficient utilization of Chlamydomonas biotechnologically in bioremediation and bio-product production. Consequently, substantial efforts are being directed towards gaining a deeper understanding of the biological mechanisms relevant to its applications. To the best of our knowledge, there has never been a single comprehensive review covering all these aspects of Chlamydomonas. Therefore, here we summarize and categorize these reports with the aim of highlighting the potential of Chlamydomonas to fulfill these tasks.
2. Wastewater and Advantages of Using Microalgae for Its Bioremediation
Wastewater comprises a diverse mixture of organic and inorganic compounds, as well as synthetic substances that reflect societal lifestyles and technology. Carbohydrates, fats, sugars, and amino acids are among the primary contaminants found in wastewater. Indeed, amino acids constitute three-quarters of the organic carbon in some wastewater [15]. Inorganic constituents found in wastewater include a variety of substances such as calcium, sodium, magnesium, potassium, sulfur, arsenic, bicarbonate, heavy metals, nitrates, chlorides, phosphates, and non-metallic salts [16]. Persistent organic pollutants include chlorinated and aromatic compounds, such as polychlorinated biphenyls, polycyclic aromatic hydrocarbons, and organochlorine pesticides [17]. The composition of wastewater varies depending on its source. Municipal wastewater is generated from households, commercial establishments, and institutions. It typically contains organic matter, nutrients, pathogens, various chemicals from soaps, and detergents [18]. Agricultural wastewater originates from farming activities and can contain organic matter, pesticides, herbicides, and fertilizers [19]. Industrial wastewater may include a diverse array of industry-specific pollutants, including heavy metals, organic chemicals, and oils [20]. Each type of wastewater has its own unique characteristics and requires specific treatment approaches to address its particular contaminants.
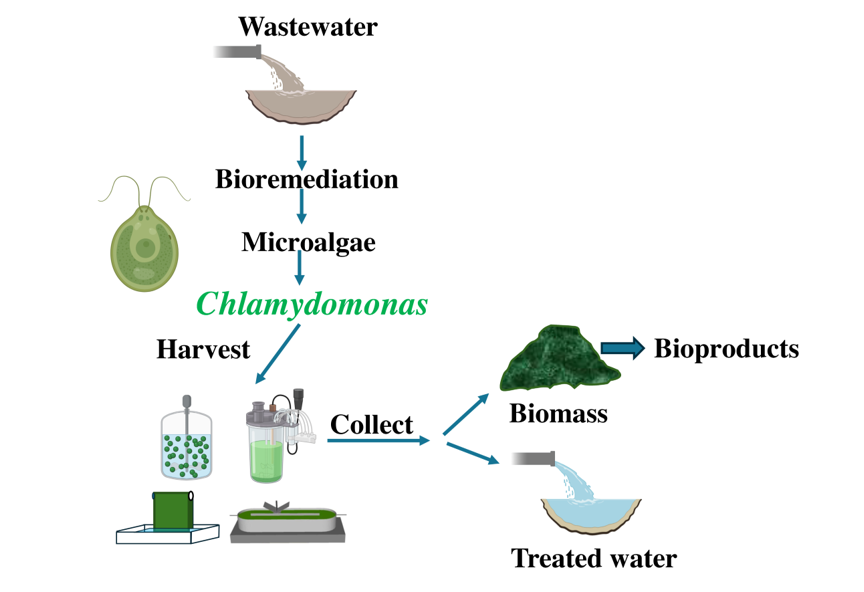
As anthropogenic activities increase, resulting in more complex wastewater compositions, it becomes crucial to develop wastewater treatment procedures that are easy to implement, efficient, and environmentally friendly. Traditional methods for treating wastewater include physical, mechanical, chemical, and biological approaches (Figure 1). Physical methods entail processes such as sedimentation, screening, and skimming, while mechanical methods include filtration techniques like ceramic membrane and sand filter technology [21]. Chemical methods involve processes such as neutralization, adsorption, precipitation, disinfection, ion exchange [22]. However, purely physical-chemical methods have proven ineffective in treating wastewater with complex compositions. Biological methods for wastewater treatment involve the use of microorganisms that consume pollutants in the wastewater as food [23]. However, biological wastewater treatment also has various drawbacks, including high energy consumption, expenses associated with aeration, and challenges in sludge management. Therefore, the integration of physical-chemical and biological methods is an effective approach for sustainable wastewater treatment [24].
Phycoremediation (where 'phyco' means algae in Greek) is a sustainable and environmentally friendly approach that utilizes various types of algae, including cyanobacteria, microalgae, and macroalgae, to remove or extract pollutants from wastewater (Figure 1). Among the benefits of phycoremediation are the removal of nutrients and xenobiotic substances, the reduction of excess nutrients from effluent with high organic material, CO2 mitigation, the treatment of effluents with heavy metal ions, and the monitoring of potentially toxic substances using algae as biosensors [25]. Microalgae have the ability to absorb and break down contaminants through processes such as biosorption, bioaccumulation, and biotransformation [26]. Phycoremediation not only helps in the removal of pollutants but also results in the production of algal biomass, which can be utilized for various valuable products such as food, feed, fertilizers, pharmaceuticals, and biofuels [27]. A wide range of non-pathogenic algae are utilized for wastewater treatment, such as Chlorella sp., Spirulina sp., Scenedesmus sp., Nostoc sp., and Oscillatoria sp., [28]. In this review, we will focus on those studies that use Chlamydomonas in phycoremediation.
3. Microalgae Cultivation Methods
Microalgae cultivation methods are categorized into suspended systems (including open reactors and closed photoreactors) and attached systems (such as biofilm reactors and encapsulated microalgae) (Figure 1).
3.1. Open Reactors
Open reactors include lakes and natural ponds, as well as specially designed high-rate algal ponds (HRAPs) that are tanks or lagoons featuring a paddle wheel that circulates wastewater. HRAPs can be an economical and sustainable method for treating wastewater, as microalgae efficiently absorb nutrients such as phosphorus and nitrogen, as well as help remove organic and inorganic contaminants [29]. Outdoor HRAPs are heavily influenced by various uncontrollable environmental factors, such as seasonal changes and weather conditions [30]. An inconvenience of this cultivation method is that open pond cultivation of mutant microalgal strains poses environmental and industrial risks. The release of mutants could damage local biodiversity and increase the probability of algal blooms.
3.2. Close Photoreactors
Closed photobioreactors (PBR) are enclosed systems utilized for the cultivation of microalgae and other phototrophic microorganisms. They provide excellent control over culture conditions with minimal risk of contamination. Different types of PBRs include flat panel, tubular, and stirred tank designs [31]. Many studies have employed PBRs with Chlamydomonas, primarily for basic research purposes. Next, we will focus on studies that have yielded practical or biotechnologically promising applications. The maximum biomass productivities were investigated in C. reinhardtii using two different PBRs: a torus-plane and a cylindrical reactor. The research highlighted that optimizing the design of Chlamydomonas PBR involves managing three key parameters: the specific illuminated area, the illuminated working volume fraction, and the mean value of the incident hemispherical photon flux density [32]. Additionally, optimization of a pilot-scale photobioreactor (120 L) for Chlamydomonas found that the gas diffuser design significantly impacted biomass production [33]. A 110 L PBR made of polypropylene and transparent Plexiglas, consisting of 64 tubes arranged in an 8x8 square pitch cell connected by U-bends, with a total length of 133 meters, has been specially designed for C. reinhardtii [34]. Recent studies have investigated the potential of utilizing the phototactic response of C. reinhardtii to reduce the economic cost of mixing in PBR. By exploiting its phototactic mechanism, C. reinhardtii can be stimulated to swim in opposite directions, thus providing mixing and ensuring access to nutrients without mechanical agitation, thereby enhancing PBR energy efficiency [35]. A multi-scale PBR, Antares I, with incident light modulations ranging from 10 to 300 µmol photons m−2 s−1, has been tested with C. reinhardtii and has shown improvement in biomass production compared to traditional PBRs. With Antares I the estimated doubling time for Chlamydomonas culture is half of that reported for culture using similar media and light conditions [36]. These findings demonstrate the diverse applications and research efforts related to the optimization of PBR for Chlamydomonas cultivation.
3.3. Biofilm Reactors
The cultivation of microalgae in biofilm reactors involves immobilizing the microalgae on a surface that acts as a support, forming a continuous layer. This method offers advantages such as higher concentration per unit volume of medium, reduction or absence of cells in the effluent, and ease of harvesting [37]. The extraction and dewatering of algae cells from biofilms are simplified by the ease of separating attached cells from their growth medium. In the context of technological applications, regulating the adhesion properties of Chlamydomonas could significantly enhance the efficiency of biofilm reactors by controlling surface colonization and biofilm formation. So far, the basic principles governing the colonization of surfaces by motile, photosynthetic microorganisms remain largely unexplored. Research has shown that the surface adhesion of C. reinhardtii is flagella-mediated and largely substrate-independent, enabling it to adhere to any type of surface [38]. However, it has been shown that the biofilm adhesion of C. reinhardtii is controlled by the type of light, being activated in blue light and deactivated under red light [39]. Interestingly, Chlamydomonas has the ability to secrete substances such as sulphated polysaccharides that act as antibiofilm agents for certain bacteria, preventing these bacteria from attaching to the biofilm [40]. This property can be highly beneficial in controlling the occurrence of bacterial contaminations.
3.4. Encapsulation
The encapsulation of microalgae is a process in which the microalgae are coated with a protective layer to enhance their stability, protect them from adverse conditions, and facilitate their application. This process offers various biotechnological advantages, such as protecting the formation of bioactive compounds, promoting release control, improving solubility, and enhancing bioavailability [41]. Various materials, including alginate, carrageenan, chitosan, and polyvinyl, have been used for the immobilization of microalgae [42]. In the case of Chlamydomonas, alginate has been the most successful and currently the most commonly used material for encapsulation. Some of its properties include low cost, biocompatibility, transparency, permeability, and mechanical defense, which reduces the risk of contamination [43]. The pore size of alginate beads in C. reinhardtii is critical, with the highest efficiency for contaminant removal obtained in gel beads with a pore size of 3.5 mm [44]. Silica hydrogels have been utilized to entrapped C. reinhardtii cells, offering advantages over alginate, such as greater stability against microbial attacks, and higher transparency [45]. One drawback of alginate encapsulation is its high porosity, which can lead to the release of large molecules. However, it has been observed with C. reinhardtii that the combination of alginate and silica to create hybrid beads can provide superior properties that overcome these limitations [46]. Single-cell encapsulation using metal-phenolic networks is an innovative technique aimed at protecting cells from stressors. This method involves coating individual cells with metal-phenolic networks to create a mechanical barrier. It was first employed with C. reinhardtii, finding that this type of encapsulation served as a mechanical barrier, delaying the proliferation of the coated cells and effectively promoting flocculation [47].
4. Chlamydomonas Phycoremediation
Microalgae, particularly Chlamydomonas, exhibit a remarkable capacity and diversity in bioremediating various molecules. Next, we will present the main mechanisms for bioremediation. Biosorption is a passive mechanism whereby microalgae serve as a biological sorbent to capture and accumulate pollutants. Microalgae utilize their cell wall and various chemical groups to attract and retain contaminants [48]. Microalgae can remove pollutants through bioaccumulation. The main differences between biosorption and bioaccumulation processes lie in their mechanisms. Biosorption is a passive process where microorganisms utilize their cellular structure to capture pollutants on the binding sites of the cell wall. On the other hand, bioaccumulation is an active process that involves the accumulation of pollutants in the biomass of microalgae, either by accumulation or uptake into intracellular spaces [49]. Bioaccumulation requires cellular growth and is typically slower than biosorption. Biotransformation involves the breakdown of pollutants, either inside or outside the cells, facilitated by enzymes [50]. While there aren't significant concerns with biosorption and bioaccumulation, biotransformation presents more challenges due to the possibility of its products being potentially more toxic than the original compounds.
Some studies have cultivated Chlamydomonas in PBRs for the decontamination of wastewater. In this regard, Chlamydomonas debaryana using dairy wastewater reduced nitrogen, phosphorus, organic carbon, and chemical oxygen demand by more than 85% [51]. C. debaryana and C. reinhardtii were able to effectively treat swine wastewater [52]. With C. reinhardtii, 55.8 mg of nitrogen and 17.4 mg of phosphorus per liter per day were effectively removed from industrial wastewater [53]. Wastewater collected from a paper industry was treated using C. reinhardtii, resulting in significant reductions of nitrate (86%), phosphate (88%), and chemical oxygen demand (93%) [54]. Using C. mexicana, a high removal efficiency of nitrogen (62%), phosphorus (28%), and inorganic carbon (29%) was achieved in piggery wastewater [55].
Numerous studies have reported the use of HRAP in wastewater treatment, primarily focusing on genera such as Scenedesmus and Chlorella [56]. However, very few records exist of applying HRAP with Chlamydomonas. In a pilot-scale HRAP experimental wastewater treatment, Chlamydomonas sp. was found to be one of the dominant genera. The study reported a reduction of the biochemical oxygen demand by 90%, chemical oxygen demand by 65%, total nitrogen by 46% and total phosphorus by 20% [57]. A study on the bioremediation of piggery wastewater using HRAP revealed that Chlamydomonas sp. was the dominant species, with average chemical oxygen demand and total nitrogen removal efficiencies of 76% and 88%, respectively [58]. In another study employing HRAP with Chlamydomonas sp. for treating municipal wastewater, average reductions of volatile suspended solids, total nitrogen, and biochemical oxygen demand were 63%, 76%, and 98%, respectively [59].
Chlamydomonas sp. JSC4 has been successfully employed in a biofilm reactor for the removal of phosphorus, nitrogen, and copper from swine wastewater [60]. In a biofilm reactor, Chlamydomonas pulvinata TCF-48 g has demonstrated significant polyphosphate accumulation and a high phosphorus removal rate of 70%, making it valuable for phosphate recovery applications [61]. The encapsulation of C. reinhardtii in alginate beads has been successfully carried out to remove various types of contaminants such as phosphorus, nitrogen, lead, mercury and cadmium [62] or even phenol [63].
C. reinhardtii has shown a significant capability for biosorption, effectively removing copper, boron, and manganese [64], arsenic [65], nickel [66], zinc, cadmium [67] and uranium [68]. In C. reinhardtii gene manipulation has been conducted to enhance the expression of the metal tolerance proteins metallothioneins [69], resulting in increased tolerance to cadmium [70], chromium [71], copper [72], mercury [73], and lead [74]. Biosorption in C. reinhardtii as a defense mechanism against silver nanoparticles involves an increase in phytochelatin and exopolysaccharides content, along with a decrease in glutathione levels [75]. C. reinhardtii has been shown to bioaccumulate several compounds as Prometryne (herbicide) [76], o-nitrophenol [77], and C. mexicana carbamazepine (antiepileptic agents) [78].
Some of the pollutants removed by biotransformation by C. reinhardtii include organophosphorus pesticide such as trichlorfon [79], polycyclic aromatic hydrocarbons such as benz(a)anthracene [80], polystyrene [81] and microplastics as bisphenol A [82]. The pharmaceuticals products that can be biotransformed by microalgae have been reviewed in [83]. Among these, Chlamydomonas has demonstrated high efficiency with the following compounds: Chlamydomonas sp. with 7-amino-cephalosporanic acid [84], C. mexicana with enrofloxacin [85], and C. reinhardtii with carbamazepine, ciprofloxacin, erythromycin, estrone, norfloxacin, ofloxacin, paracetamol, progesterone, roxithromycin, salicylic acid, sulfadiazine, sulfadimethoxine, sulfametoxydiazine, sulfamethazine, triclocarban, triclosan and trimethoprim [86], sulfadiazine [87] and ibuprofen [88]. C. reinhardtii has been found to biotransform antibiotics like azithromycin, erythromycin, and sulphapyridine [89]. C. reinhardtii was shown to be able to biotransform the hormones β-estradiol and 17α-ethinylestradiol [90] as well as the non-steroidal anti-inflammatory drug diclofenac [91].
Chlamydomonas can metabolize xenobiotics through a wide range of enzymatic processes, including CYP450 oxidation reactions, hydrolysis, glutamate conjugation, and methylation [92]. Chlamydomonas moewusii excretes laccases capable of breaking down and detoxifying phenolic pollutants [93]. The toxicity responses of different pollutants, such as benzophenone-3, bisphenol A, oxytetracycline, and atrazine, in C. reinhardtii showed a similar pattern: an increase in chlorophyll autofluorescence and a decrease in growth rate and vitality [94]. The biotransformation of five bisphenol derivatives (AF, B, F, S, and Z) by C. mexicana shows that all the biotransformed products were less toxic than the parent compounds [95]. Chlamydomonas has also been used in efforts to degrade commonly used plastic components such as Polyethylene terephthalate (PET). In Ideonella sakaiensis, a novel plastic degradation enzyme called PETase has been identified [96]. The I. sakaiensis PETase has been expressed through genetic recombination in the C. reinhardtii nucleus and chloroplast genomes, showing a significant ability to break down PET [97]. Under specific adverse conditions such as NaCl stress, EDTA exposure or acidic pH, C. reinhardtii can form multicellular aggregates called palmelloids. These are small clonal structures that result from cells failing to separate after division [98]. The defense mechanisms of C. reinhardtii under perchlorate stress were investigated, revealing palmelloid formation when exposed to 100 and 200 mM perchlorate [99]. These researchers highlight the metabolic versatility of Chlamydomonas in dealing with xenobiotic compounds, demonstrating its ability to transform and process a variety of chemicals through different mechanisms.
Microalgae have been actively employed in initiatives focused on reducing CO2 emissions due to their ability to absorb CO2 via photosynthesis. C. reinhardtii exhibits a superior ability to fix CO2 compared to other photosynthetic organisms [100]. Bio-fixation refers to the process by which certain organisms, such as microalgae, utilize CO2 from the air or other sources like flue gas streams to create biomass. The production of 1 gram of microalgae biomass leads to the sequestration of 1.8 grams of CO2 [101]. In Chlamydomonas the expression of a single H+-pump increase its tolerance to high concentrations of CO2, such as those found in industrial flue gas [102]. These findings illustrate the potential of C. reinhardtii to mitigate CO2 emissions from industrial sources.
5. Chlamydomonas Bioproduct Generation
5.1. Biomass
One of the main products derived from the cultivation of microalgae is their biomass, as it is used as raw material for obtaining other derived bioproducts. It would be highly beneficial economically to use the biomass resulting from the bioremediation process for bioproduct purification. However, the utilization of biomass derived from wastewater treatment encounters several inherent challenges. These challenges include the scalability of biomass production, the presence of xenobiotics and heavy metals, as well as the contamination with bacteria, fungi, and viruses, all of which limit their extensive application [103]. Although numerous efforts are being made to address this issue, the production of the main bioproducts obtained from microalgae still does not use wastewater as a cultivation source. Next, we will present studies that utilize Chlamydomonas to obtain certain bioproducts, some of which use biomass derived from wastewater remediation.
The composition of biomass is influenced by the strains of microalgae and the culture conditions [104]. One straightforward method to increase biomass productivity involves altering the culture medium conditions or adjusting the supply of certain macroelements. For example, in microalgae, some researchers have evaluated the effect of various carbon sources [105], pH variations [106], the photoperiods [107], as well as trace elements composition for biomass production [108]. Consequently, various approaches have been explored to optimize microalgae biomass enriched in specific biomolecules (Figure 2). The highest biomass concentration of Chlamydomonas obtained so far has been heterotrophically with acetate, reaching 23 g/l [109], far behind compared to other green algae that are able to consume glucose as a substrate, like Chlorella sp. and Scenedesmus sp., for which biomass reached 271 g/L and 286 g/L respectively [110].
5.2. Biochar
Biochar is a carbonaceous material produced through the pyrolysis of biomass (Figure 2), which can be obtained from microalgae, agricultural residues, wood, or organic waste [111]. Biochar is characterized by its high porosity and specific surface area, making it useful for improving soil quality and carbon sequestration. It is used in agriculture as a soil amendment to enhance soil structure, retain nutrients and water, and promote beneficial microbial activity. Additionally, biochar is considered a strategy for mitigating climate change, as burying it in the soil can store carbon stably for long periods [112]. C. reinhardtii biomass has been successfully used to prepare biochar [113]. The highest biochar yield was 93.9%, achieved by dry torrefaction at 200°C of Chlamydomonas sp. JSC4 [114]. Biochar prepared from Chlamydomonas sp. has been shown to have a high capacity for removing contaminants [115].
5.3. Biofertilizers
Microalgae are used as a biofertilizers and biostimulants by promoting crop growth and increasing soil nutrient contents, thereby reducing the usage of chemical fertilizers [116]. In contrast, Chlamydomonas species have received little attention and are not fully utilized in agriculture, despite being among the most abundant microalgae species in natural soil ecosystems. In this regard, a study on the effects of Chlamydomonas applanata M9V as a biofertilizer on wheat found that it performed even better than a certain amount of chemical fertilizer [117]. Acid-hydrolyzed dry biomass of C. reinhardtii improved the phosphorus, nitrogen, and carotenoid contents of Solanum lycopersicum [118]. The application of live Chlamydomonas cells significantly increased leaf size, shoot length, fresh weight, number of flowers, and pigment content of Medicago truncatula [119]. Lyophilized powders derived from C. reinhardtii have been found to positively affect the growth of maize plants by producing bioactive compounds that act as biostimulants, enhancing plant growth, crop performance, yields, and quality [120]. Biomass extracts of Chlamydomonas sp. exhibited auxin-like activity that increased the number of roots in cucumber [121]. Chlamydomonas sajao can improve soil physical properties, such as aggregation and stability, thereby contributing to enhanced soil structure and nutrient retention [122]. These results suggest that Chlamydomonas can be an effective alternative to chemical fertilizers for promoting crop growth and yield.
5.4. Bioplastic
Bioplastics are biodegradable materials derived from renewable biomass sources, offering a sustainable alternative to traditional plastics [123]. Various molecules can be used as building blocks for bioplastics, including polyhydroxybutyrate (PHB), starch, TAG, lactic acid, or polybutylene succinate. PHB can be naturally synthesized by certain bacteria, such as Azotobacter or Pseudomonas. PHB production involves three key enzymes: β-ketothiolase, acetoacetyl-CoA reductase, and PHB synthase, encoded by phbA, phbB, and phbC, respectively. Research has focused on engineering Chlamydomonas strains to enhance PHB de novo biosynthesis, as Chlamydomonas naturally cannot synthesize PHB. With this aim, the phbB and phbC genes from Ralstonia eutropha have been inserted into the C. reinhardtii genome, leading to the observation of PHB granules in the cytoplasm [124]. While cytosolic accumulation of PHB in Chlamydomonas often results in impaired cell growth and low yield, peroxisomes have emerged as a promising alternative. A complete PHB biosynthesis pathway has been successfully reconstructed by expressing the three PHB synthesis genes and targeting the proteins to the peroxisomes. Within the peroxisomes of these strains, PHB reached 21.6 mg/g, which represents a 3600-fold increase over cytosolic PHB production [125]. Another strategy is to use TAG as the building block for bioplastic. TAG synthesized by C. reinhardtii has been directly crosslinked with glycerol or ammonium persulfate and molded into plastic beads that are capable of withstanding compressive stress up to 1.7 megapascals [126]. Cell-plastics are a type of bioplastic that directly utilizes raw cells and the hydrolyzed cell broth. Unlike conventional bioplastics, cell-plastics do not require exhaustive processes for extracting and refining the biomolecules that serve as the building blocks. Recently, Chlamydomonas cells have arisen as the constituent blocks of this new type of bioplastic, as their cell size and protein-rich, cellulose-free cell wall were demonstrated to be ideal components for its fabrication [127].
5.5. Biofuels
Biofuels are fuels derived from renewable biological sources such as plants or plant-derived materials. First-generation biofuels are produced from food crops. Second-generation biofuels are derived from non-food sources such as waste, and third-generation biofuels are produced from sources that do not compete with arable land, such as microalgae [128]. Microalgae have regained attention as alternative resources for environmentally friendly production of biofuels, including biodiesel, bioethanol, biogas, and biohydrogen. These biofuels can be produced through thermochemical and biochemical conversions, photosynthesis-mediated microbial fuel production, and transesterification [129].
5.5.1. Biodiesel
Triacylglycerols (TAG) are crucial lipids in microalgae for biofuel production. Oleaginous microalgae, rich in TAG, can be converted into biodiesel through transesterification, a process that transforms TAG into fatty acid methyl esters, the key components of biodiesel [130]. Utilizing Chlamydomonas sp. JSC4, a direct transesterification process was employed, resulting in nearly 100% biodiesel production in a single step [131]. Given that biodiesel production is closely linked to the quantity of lipids and TAGs, various strategies have been explored to enhance their production in Chlamydomonas. Some studies have focused on elucidating the functions of key genes involved in lipid and TAG production. The down-regulation of the phosphoenolpyruvate carboxylase gene in C. reinhardtii resulted in a 74.4% increase in lipid content [132]. The overexpression of acetyl-CoA synthetase resulted in a 2.4-fold increase in the accumulation of TAG [133]. In C. reinhardtii, the mutation of ACX2, which encodes a member of the acyl-CoA oxidase responsible for the first step of peroxisomal fatty acid beta-oxidation, resulted in an accumulation of 20% more lipid [134]. A mutant of C. reinhardtii deficient in phospholipase showed an increase in TAG content of up to 190% [135]. The overexpression of the ferredoxin gene PETF in C. reinhardtii resulted in higher lipid content [136]. The Target of Rapamycin (TOR) plays a crucial role in regulating cell growth. It has been shown that mutants of C. reinhardtii lacking TOR experience an increase in TAG production [137]. The strategy of heterologously overexpressing genes in Chlamydomonas has been successful in increasing TAG content. In this sense, the heterologous expression of the Dunaliella tertiolecta fatty acyl-ACP thioesterase in C. reinhardtii leads to increased lipid production [138]. By expressing the diacylglycerol acyltransferase from Saccharomyces cerevisiae into C. reinhardtii, the fatty acids and TAG content increased by 22% and 32%, respectively [139]. The heterologous expression of Lobosphaera incisa glycerol-3-phosphate acyltransferase in C. reinhardtii enhances TAG production [140]. The synthesis of starch and lipids competes for carbon skeletons; thus, inhibiting starch synthesis is another strategy followed to increase TAG production. In this sense, silencing ADP-glucose pyrophosphorylase in C. reinhardtii resulted in a tenfold increase in TAG content [141]. Genetically modifying Chlamydomonas sp. JSC4 in the gene that encodes the starch debranching enzyme promotes carbohydrate degradation and redirects carbon resources into lipids, resulting in a 1.46-fold increase in lipid [142].
A commonly employed approach to accumulate TAG in Chlamydomonas is to induce stress conditions, particularly nutrient limitation or starvation [143]. C. reinhardtii exhibits a notable increase in TAG accumulation under low nitrogen concentration [144]. Under nitrogen deprivation, C. reinhardtii starch mutants exhibit almost a 10-fold increase in TAG [145]. Under nitrogen limitations, increasing the expression of S-adenosylmethionine synthetase in C. reinhardtii enhances cell viability and TAG production [146]. Phosphorus stress also triggers TAG production in Chlamydomonas [147]. Additionally, a higher TAG content is generated under conditions of low sulfur concentration [148]. The lipid content in C. mexicana was observed to rise as the concentration of NaCl was increased to 25 mM [149]. The lipid content of the C. reinhardtii starchless mutant BAF-J5 increased by 76% following a temperature shift to 32°C [150].
Increasing TAG levels by inducing stress conditions often comes at the expense of inhibited microalgal growth. Under these conditions, there is an inverse relationship between TAG yield and microalgal growth. To mitigate this, it has been reported that overexpressing the transcription factor MYB1 in C. reinhardtii, which mediates lipid accumulation, results in nearly 60% more TAG without negatively impacting cell growth [151]. In another strategy, a cultivation approach involving two stages has been proposed, wherein C. reinhardtii experiences nutrient stress only after an initial period of optimal growth, allowing for high TAG accumulation [152]. The development of effective methods for cultivating Chlamydomonas is essential in biodiesel production. In this regard, in C. reinhardtii a multi-parametric kinetic model developed using computational tools has been proven, resulting in significant increases in lipids (74%) [153].
5.5.2. Bioethanol
Bioethanol is a biofuel that can be obtained through the fermentation of various types of biomass containing high amounts of sugars. For bioethanol production, the high carbohydrate content present in both the cellulose and hemicellulose cell walls, as well as the starch-based cytoplasm, is broken down into monomeric sugars during enzymatic hydrolysis prior to fermentation. However, the cell wall of Chlamydomonas is not made of cellulose like in plants, but of five dense glycoprotein-rich layers [154]. Therefore, efforts have been focused on utilizing starch-rich Chlamydomonas for the production of bioethanol. The biomass of C. reinhardtii UTEX 90 was converted into glucose through two hydrolytic steps using α-amylase and amyloglucosidase, with nearly all the starch successfully transformed into glucose without damaging the cell wall, reducing the costs of bioethanol purification [155]. Pretreating C. reinhardtii UTEX 90 biomass with sulfuric acid (1-5%) at temperatures ranging from 100 to 120°C significantly increases the glucose release for the production of bioethanol [156]. The psychrophilic alga Chlamydomonas sp. KNM0029C was studied for its potential to produce bioethanol, showing promising results (Kim et al., 2020). The supraoptimal temperature treatment method, which involved cultivating C. reinhardtii at 39°C despite its optimal temperature being 25°C, was successfully applied and resulted in nearly a threefold enhancement of starch content [157]. In C. reinhardtii, optimizing the pH, temperature, initial concentrations of acetate and ammonium, along with the use of depigmented and defatted biomass, led to a bioethanol yield ranging from 90% to 94% (Banerjee et al., 2021). The hormones have also been described to have a very important role in starch accumulation; in this sense, in Chlamydomonas 100 µM of Indole-3-acetic acid produces an accumulation of up to nine times more starch [158]. Chlamydomonas sp. QWY37 has been effectively utilized for bioethanol production from swine wastewater, achieving a maximum bioethanol yield of 61 g/L [159].
5.5.3. Biogas
Biogas is a renewable energy source primarily composed of CH4, derived from the microbial anaerobic digestion of biomass obtained from various sources (Figure 2). The production of biogas involves multiple stages, including hydrolysis, acidogenesis, acetogenesis, and methanogenesis, which are facilitated by a microbial consortium that plays a crucial role in influencing both the composition and yield of the biogas [160]. This process eliminates the need to extract specific macromolecules, such as lipids, proteins, or carbohydrates, and can be carried out using wet biomass [161]. The fermentation of C. reinhardtii biomass produces approximately 587 ml of biogas per gram of volatile solids [162]. However, microalgae biomass is not ideal for biogas generation due to its high protein content, which results in an unfavorable low carbon-to-nitrogen ratio. This imbalance arises because the ammonia released during protein degradation inhibits the methanogenesis process [163]. C. reinhardtii biomass has been studied for its potential in overcoming this limitation. In this regard, the anaerobic digestion of C. reinhardtii biomass obtained in low nitrogen media has shown remarkable efficiency in biogas production due to its high carbon-to-nitrogen ratio [164].
The high resistance of microalgae biomass to microbial decomposition due to their rigid cell walls is a significant challenge in biogas production. However, since the main components of the C. reinhardtii cell wall are glycoproteins rather than cellulose, C. reinhardtii has been shown to produce larger quantities of biogas compared to species with more complex cell walls (such as Chlorella sp. and Scenedesmus sp.) [165]. The findings revealed that the C. reinhardtii cell wall was not an obstacle but instead became advantageous by enabling the gradual degradation of intracellular content [166]. One way to valorize the microalgal biomass produced during wastewater treatment is to utilize it as a source for biogas production, thereby reducing the economic costs of treatment [167]. In this regard, Chlamydomonas sp. Ck has demonstrated high efficiency in decontaminating piggery wastewater while simultaneously producing a high biogas yield [168]. For all the reasons mentioned, anaerobic digestion of C. reinhardtii biomass can be considered a cost-effective alternative for biogas production compared to other methods.
5.5.4. Hydrogen
The production of the preceding bioproducts shares the common step of first obtaining biomass, and then extracting these compounds from it. Next, we will present some products that Chlamydomonas releases into the culture medium and therefore can be purified without needing to be extracted from the biomass, thereby reducing the economic cost of their production (Figure 2). A prominent example of this is hydrogen, that has emerged as one of the most promising energy carriers for future energy demands. Hydrogen presents the opportunity to cultivate living organisms such as bacteria, cyanobacteria, and microalgae capable of releasing H2 into the media [169]. Hydrogen is generated through enzymes known as hydrogenases [170]. Chlamydomonas has two hydrogenases that have been extensively studied with the aim of increasing their production efficiency [171]. The hydrogenases catalyze the reduction of protons into H2 either using energy from light (biophotolysis) or by oxidizing organic compounds such as starch (dark fermentation). One of the primary biotechnological challenges of using Chlamydomonas as a factory to produce H2 is the rapid inactivation of its hydrogenases by oxygen, particularly considering that oxygen is generated during photosynthesis. Therefore, the initial evidence indicating that Chlamydomonas was capable of producing H2 was observed with Chlamydomonas moewusii under anaerobic condition [172], and subsequently with C. reinhardtii, also anaerobically [173]. The first successful strategy demonstrating significant and consistent H2 production under aerobic conditions involved using sulfur-starved C. reinhardtii [174]. The reason for this is that the absence of sulfur blocks protein synthesis, thereby halting photosynthesis and oxygen production. Alternative strategies for H2 production under non-stress conditions are also possible, particularly in media containing acetate, which is compatible with Chlamydomonas growth [175,176]. However, the rates of H2 production under non-stress conditions are lower compared to those under stressful condition [177].
In Chlamydomonas, numerous genetically engineered strains have been developed to enhance H2 production. One of the most successful approaches has been to improve the intrinsic oxygen tolerance of hydrogenase through mutagenesis [178]. A production of 1200 mL of H2 per liter has been reported after 6 days using the Photosystem I (PSI) cyclic electron transport mutant pgr5, which is defective in thylakoid proton gradient regulation [179]. Another strategy is diverting electron flow to the hydrogenase [180], and degrading or inhibiting the function of Photosystem II (PSII) to prevent oxygen production [181]. However, strategies that do not degrade PSII appear to be advantageous, as the long-term loss of PSII inhibits cell growth. In this sense, a PSI-hydrogenase chimera was created by inserting the HydA sequence into the PsaC (stromal subunit of PSI). This redirects photosynthetic electron flow towards proton reduction [182]. A disadvantage in the use of Chlamydomonas is that the hydrogen production rate is influenced by the size of microalgae cells. The hydrogen production rate of Chlorella is higher than that of Chlamydomonas due to its relatively smaller size [183].
5.6. High-Value Bioproducts
The term "high-value bioproducts" refers to a wide range of products derived from various sources, which economically have a higher value compared to low- to medium-value products. C. reinhardtii is a promising organism for the production of high-value bio-products [184]. Glycolate, a high-value cosmetic ingredient can be overproduced in Chlamydomonas. When Chlamydomonas is in an environment with low CO2 (0.04%), Rubisco oxygenates ribulose-1,5-bisphosphate instead of carboxylating it, consequently producing glycolate. In Chlamydomonas, glycolate is toxic, prompting an active system to excrete it. To facilitate the recovery of potentially lost carbon, the genes for photorespiratory metabolism are induced. Photorespiration detoxifies and recycles glycolate, generating glycerate and releasing CO2. In Chlamydomonas, glycolate dehydrogenase (GDH) is involved in photorespiration by oxidizing glycolate to glyoxylate. It has been observed that Chlamydomonas GDH mutants over-accumulate glycolate in the media [185]. Chlamydomonas has a CO2-concentrating mechanism (CCM) to prevent the rubisco oxygenation reaction and, consequently, glycolate excretion [186]. CIA5 is the primary transcription factor that induces the CCM, and its mutation has been shown to increase the amount of excreted glycolate [187]. By incorporating 6-Ethoxy-2-benzothiazolesulfonamide (EZA), a CCM inhibitor, glycolate production can be maximized without compromising cell viability. Under these conditions, glycolate accumulates in the medium, reaching a concentration of up to 41 mM [188]. In photorespiration, hydroxypyruvate is converted to glycerate by hydroxypyruvate reductase (HPR). In C. reinhardtii, the mutation of hpr1 results in increased excretion of glycolate into the medium [189].
Bioisoprenoids are natural compounds synthesized by plants, animals, and microorganisms through the isoprenoid biosynthetic pathway. These compounds are structurally and functionally diverse, with a wide range of applications, including their use as perfumes, cosmetics, pigments, medicines, and chemical signals. Bioisoprene production has gained attention due to its sustainability and efficiency compared to petrochemical sources [190]. It has been demonstrated that C. reinhardtii can be genetically modified to produce significant amounts of bioisoprene by overexpressing four different plant isoprene synthase genes (IspS), with the strain expressing the Ipomoea batatas IspS gene showing the highest isoprene levels [191].
Hydroxyalkanoyloxyalkanoates (HAA) are a type of lipidic surfactants that can be produced by certain bacteria that show great potential for a wide range of applications. They are synthesized by the condensation of hydroxyalkanoic acids, which are produced by the metabolism of fatty acids. The chloroplast genome of C. reinhardtii was engineered by inserting the gene encoding the acyltransferase of P. aeruginosa, a key enzyme in HAA synthesis, resulting in high concentrations of HAA not only in the intracellular fraction but also in the extracellular [192].
There is strong interest in developing bio-based hydrocarbons and their unsaturated analogs, the alkenes, as potential substitutes for hydrocarbons derived from petroleum. The alkene 7-heptadecene has high demand for various biotechnological processes. While the biological function of alkenes in microalgae remains completely unknown, it has been shown that in C. reinhardtii, the enzyme fatty acid photodecarboxylase is responsible for synthesizing 7-heptadecene [193]. This discovery opens the possibility of overproducing this alkene in C. reinhardtii. ε-Polylysine is a biodegradable polymer composed of 25-30 lysine monomers that has a variety of applications, including antimicrobial activity and anticancer agent [194]. It has been reported that ε-polylysine is produced from Chlamydomonas sp. supplemented with lysine, aspartate, and tricarboxylic acids, achieving a maximum production of 2.24 g/L [195].
Bio-polyamides, also known as nylons, are sustainable polymers derived from renewable resource. Bio-polyamides have excellent material properties, leading to a high demand for polyamide plastics with diverse applications across various industries [196]. Cadaverine and putrescine are polyamines commonly used as precursors and building block for the synthesis of bio-polyamides. By the heterologous expression of two E. coli lysine decarboxylases in C. reinhardtii, it was possible to significantly enhance the synthesis of cadaverine [197]. The mutation of essential genes in the C. reinhardtii polyamine biosynthesis pathway identified ornithine decarboxylase 1 (ODC1) as a crucial regulator that controls the accumulation of putrescine. Subsequently, the authors overexpressed different ODCs, resulting in a significant increase in cellular putrescine levels, reaching a maximum yield of 200 mg/L [198]. This achievement marks the first instance of microalgal bio-production of putrescine.
C. reinhardtii, along with Chlorella vulgaris, Dunaliella bardawil, Arthrospira platensis, Auxenochlorella protothecoides, and Euglena gracilis are among the very few microalgae recognized by the Food and Drug Administration as Generally Recognized as Safe (GRAS) organisms (GRAS Notice No. 773). This acknowledgment allows their use as a nutritional component in food, presenting new opportunities for the utilization of C. reinhardtii. Clinical studies on the human consumption of C. reinhardtii whole cells have demonstrated positive effects on gastrointestinal health and microbiota, showing that the intake of C. reinhardtii cells promotes microbiota eubiosis, reducing imbalances and improving the overall health of the intestine [199]. The development of alternative plant-based products to substitute meat has led to the exploration of heme-containing proteins for their ability to provide a meat-like color and flavor. One such compound that can provide these qualities is protoporphyrin IX (PPIX) a crucial intermediate in the heme biosynthetic pathway. In this regard, engineered C. reinhardtii strains have been shown to overexpress PPIX [200].
Antioxidants are widely recognized for their beneficial impact on health and their crucial role in protecting cells from the harmful effects of free radicals. Chlamydomonas agloeformis has garnered attention due to its exceptionally high antioxidant capacities that surpass those of higher plants [201]. Carotenoids are a diverse group of lipid-soluble pigments produced by plants and microorganisms, known for their benefits as vitamin precursors and antioxidants. Astaxanthin, a ketocarotenoid, is recognized as one of the most powerful natural antioxidants among carotenoids [202]. Astaxanthin is currently primarily produced industrially from the microalgae Haematococcus pluvialis, with the crucial enzyme involved in its biosynthesis being β-carotene ketolase (BKT)[203]. The synthetic redesign and overexpression of C. reinhardtii BKT has been shown to achieve Astaxanthin productivities of up to 4.3 mg/L/day, which is comparable to the results obtained with H. pluvialis [204]. This production does not impair the growth or biomass productivity of C. reinhardtii, presenting a promising alternative to natural astaxanthin-producing algal strains. Furthermore, the accumulation of astaxanthin has led to enhanced high light tolerance and increased biomass productivity [205]. Blocking the expression of ATG1 and ATG8, genes involved in autophagy in C. reinhardtii, leads to a 2.3 times increase in carotenoid biosynthesis, indicating that autophagy does play a role in regulating carotenoid levels [206].
Chlamydomonas has been shown to be able to synthesize vitamins C, A, E, B1, B7, B9, and ergosterol, the precursor of vitamin D2 [207]. However, for most of these vitamins, the mechanisms regulating their synthesis to achieve overproduction have not been studied in detail. In C. reinhardtii, oxidative stress leads to a substantial increase in vitamin C levels [208]. Omega-3 fatty acids play critical roles as nutrients and are extensively utilized in medicine. A comparison of C. reinhardtii with Chlorella and Spirulina revealed that C. reinhardtii contains superior amounts of omega-3 fatty acids, both in quality and quantity [209]. Sulphated polysaccharides (SPs) are polymer chains containing one or more monosaccharide units that have been modified with sulfate groups. C. reinhardtii is capable of synthesizing SPs, which have been associated with several beneficial properties, including potent antioxidant and anticancer effects [210], antineurodegenerative [211] and antibiotics [212].
More than 40 therapeutic proteins, such as antibodies, enzymes, viral proteins, hormones, among others, have been successfully expressed in C. reinhardtii [213]. ICAM-1, a protein belonging to the immunoglobulin superfamily, was targeted for secretion into the extracellular media and was found to be fully active, suggesting that C. reinhardtii can produce mammalian proteins that are correctly folded and functional. Additionally, it achieved a concentration of up to 46.6 mg/L, marking the highest reported concentration of any recombinant protein in C. reinhardtii to date [110]. The production of full-length spike protein, a crucial component for the infectivity of SARS-CoV-2, has been successfully achieved in C. reinhardtii as a secreted protein [214]. This achievement is crucial as it offers a simpler and more economical platform for producing recombinant spike proteins in microalgae.
6. Conclusions and Outlook
Throughout this review, various studies conducted with Chlamydomonas on bioremediation and the production of bioproducts have been presented. As observed, the studies in this regard are diverse and cover different fields. We believe that a key area to develop in the future, due to its significant economic and environmental impact, would be to combine these two aspects simultaneously. In other words, the biomass obtained from bioremediation should be used for the production of a specific bioproduct of interest. As indicated in this review, some attempts have been made in this direction, and although its development has significant challenges, we believe this would be a very promising strategy to pursue in the future.
Author Contributions
AL original idea, conceptualization, and preparation of the first draft; AL, C B-P and MJ T-P wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by Gobierno de España, Ministerio de Ciencia e Innovacion (Grant PID2020-118398GB-I00), Junta de Andalucía (Grant ProyExcel_00483), the “Plan Propio” from University of Cordoba, and a grant awarded by the Torres-Gutierrez foundation.
Data Availability Statement
All data required to evaluate the conclusions of this paper are included in the main text.
Acknowledgments
This paper is dedicated to Emilio Fernandez Reyes, who has recently retired after almost 40 years of studying Chlamydomonas reinhardtii as a reference organism. He was the driving force that promoted our research on Chlamydomonas, the pillar that allowed its advancement, and our great teacher whom we will never be able to repay for all the learnings received. We also thank Maribel Macias for her constant technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rani, S.; Gunjyal, N.; Ojha, C.S.P.; Singh, R.P. Review of Challenges for Algae-Based Wastewater Treatment: Strain Selection, Wastewater Characteristics, Abiotic, and Biotic Factors. J. Hazardous, Toxic, Radioact. Waste 2021, 25, 03120004. [Google Scholar] [CrossRef]
- Falkowski, P.G. The Role of Phytoplankton Photosynthesis in Global Biogeochemical Cycles. Photosynth. Res. 1994, 39, 235–258. [Google Scholar] [CrossRef]
- Ochoa De Alda, J.A.G.; Esteban, R.; Diago, M.L.; Houmard, J. The Plastid Ancestor Originated among One of the Major Cyanobacterial Lineages. Nat. Commun. 2014, 5, 4937. [Google Scholar] [CrossRef] [PubMed]
- Sibbald, S.J.; Archibald, J.M. Genomic Insights into Plastid Evolution. Genome Biol. Evol. 2020, 12, 978–990. [Google Scholar] [CrossRef] [PubMed]
- de Cassia Soares Brandão, B.; Oliveira, C.Y.B.; dos Santos, E.P.; de Abreu, J.L.; Oliveira, D.W.S.; da Silva, S.M.B.C.; Gálvez, A.O. Microalgae-Based Domestic Wastewater Treatment: A Review of Biological Aspects, Bioremediation Potential, and Biomass Production with Biotechnological High-Value. Environ. Monit. Assess. 2023, 195, 1384. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Ye, X.; Bi, H.; Shen, Z. Microalgae Biofuels: Illuminating the Path to a Sustainable Future amidst Challenges and Opportunities. Biotechnol. Biofuels Bioprod. 2024, 17, 10. [Google Scholar] [CrossRef]
- Gao, S.; Chen, W.; Cao, S.; Sun, P.; Gao, X. Microalgae as Fishmeal Alternatives in Aquaculture: Current Status, Existing Problems, and Possible Solutions. Environ. Sci. Pollut. Res. 2024, s11356. [Google Scholar] [CrossRef]
- Gupta, A.; Kang, K.; Pathania, R.; Saxton, L.; Saucedo, B.; Malik, A.; Torres-Tiji, Y.; Diaz, C.J.; Dutra Molino, J.V.; Mayfield, S.P. Harnessing Genetic Engineering to Drive Economic Bioproduct Production in Algae. Front. Bioeng. Biotechnol. 2024, 12, 1350722. [Google Scholar] [CrossRef] [PubMed]
- Udaypal; Goswami, R. K.; Mehariya, S.; Verma, P. Advances in Microalgae-Based Carbon Sequestration: Current Status and Future Perspectives. Environ. Res. 2024, 249, 118397. [Google Scholar] [CrossRef]
- Sasso, S.; Stibor, H.; Mittag, M.; Grossman, A.R. From Molecular Manipulation of Domesticated Chlamydomonas Reinhardtii to Survival in Nature. Elife 2018, 7, e39233. [Google Scholar] [CrossRef]
- Salomé, P.A.; Merchant, S.S. A Series of Fortunate Events: Introducing Chlamydomonas as a Reference Organism. Plant Cell 2019, 31, 1682–1707. [Google Scholar] [CrossRef] [PubMed]
- Oz Yasar, C.; Fletcher, L.; Camargo-Valero, M.A. Effect of Macronutrients (Carbon, Nitrogen, and Phosphorus) on the Growth of Chlamydomonas Reinhardtii and Nutrient Recovery under Different Trophic Conditions. Environ. Sci. Pollut. Res. 2023, 30, 111369–111381. [Google Scholar] [CrossRef] [PubMed]
- Goodenough, U. The Chlamydomonas Sourcebook. Volume 1: Introduction to Chlamydomonas and Its Laboratory Use. Elsevier Acad. Press. [CrossRef]
- Calatrava, V.; Tejada-Jimenez, M.; Sanz-Luque, E.; Fernandez, E.; Galvan, A.; Llamas, A. Chlamydomonas Reinhardtii, a Reference Organism to Study Algal–Microbial Interactions: Why Can’t They Be Friends? Plants 2023, 12, 788. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, Q.; Zhou, R.; Feng, J.; Zhang, K.; Li, X.; Ma, X.; Dietrich, A.M. Occurrence of Free Amino Acids in the Source Waters of Zhejiang Province, China, and Their Removal and Transformation in Drinking Water Systems. Water (Switzerland) 2020, 12, 73. [Google Scholar] [CrossRef]
- Ahamed, A.; Ge, L.; Zhao, K.; Veksha, A.; Bobacka, J.; Lisak, G. Environmental Footprint of Voltammetric Sensors Based on Screen-Printed Electrodes: An Assessment towards “Green” Sensor Manufacturing. Chemosphere 2021, 278, 130462. [Google Scholar] [CrossRef] [PubMed]
- Gaur, N.; Dutta, D.; Singh, A.; Dubey, R.; Kamboj, D.V. Recent Advances in the Elimination of Persistent Organic Pollutants by Photocatalysis. Front. Environ. Sci. 2022, 10, 872514. [Google Scholar] [CrossRef]
- Wagner, T. V.; Rempe, F.; Hoek, M.; Schuman, E.; Langenhoff, A. Key Constructed Wetland Design Features for Maximized Micropollutant Removal from Treated Municipal Wastewater: A Literature Study Based on 16 Indicator Micropollutants. Water Res. 2023, 244, 120534. [Google Scholar] [CrossRef] [PubMed]
- Morillas-España, A.; Lafarga, T.; Sánchez-Zurano, A.; Acién-Fernández, F.G.; González-López, C. Microalgae Based Wastewater Treatment Coupled to the Production of High Value Agricultural Products: Current Needs and Challenges. Chemosphere 2022, 291, 132968. [Google Scholar] [CrossRef]
- Elangovan, B.; Detchanamurthy, S.; Senthil Kumar, P.; Rajarathinam, R.; Deepa, V.S. Biotreatment of Industrial Wastewater Using Microalgae: A Tool for a Sustainable Bioeconomy. Mol. Biotechnol. 2023. [Google Scholar] [CrossRef]
- Talukdar, A.; Kundu, P.; Bhattacharya, S.; Dutta, N. Microplastic Contamination in Wastewater: Sources, Distribution, Detection and Remediation through Physical and Chemical-Biological Methods. Sci. Total Environ. 2024, 916, 170254. [Google Scholar] [CrossRef]
- Krishnan, R.Y.; Manikandan, S.; Subbaiya, R.; Biruntha, M.; Govarthanan, M.; Karmegam, N. Removal of Emerging Micropollutants Originating from Pharmaceuticals and Personal Care Products (PPCPs) in Water and Wastewater by Advanced Oxidation Processes: A Review. Environ. Technol. Innov. 2021, 23, 101757. [Google Scholar] [CrossRef]
- Singh, A.; Pal, D.B.; Mohammad, A.; Alhazmi, A.; Haque, S.; Yoon, T.; Srivastava, N.; Gupta, V.K. Biological Remediation Technologies for Dyes and Heavy Metals in Wastewater Treatment: New Insight. Bioresour. Technol. 2022, 343, 126154. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.A. Wastewater Treatment and Reuse for Sustainable Water Resources Management: A Systematic Literature Review. Sustainability 2023, 15, 10940. [Google Scholar] [CrossRef]
- Yadav, G.; Shanmugam, S.; Sivaramakrishnan, R.; Kumar, D.; Mathimani, T.; Brindhadevi, K.; Pugazhendhi, A.; Rajendran, K. Mechanism and Challenges behind Algae as a Wastewater Treatment Choice for Bioenergy Production and Beyond. Fuel 2021, 285, 119093. [Google Scholar] [CrossRef]
- Bhatia, S.K.; Mehariya, S.; Bhatia, R.K.; Kumar, M.; Pugazhendhi, A.; Awasthi, M.K.; Atabani, A.E.; Kumar, G.; Kim, W.; Seo, S.O.; et al. Wastewater Based Microalgal Biorefinery for Bioenergy Production: Progress and Challenges. Sci. Total Environ. 2021, 751, 141599. [Google Scholar] [CrossRef]
- Razaviarani, V.; Arab, G.; Lerdwanawattana, N.; Gadia, Y. Algal Biomass Dual Roles in Phycoremediation of Wastewater and Production of Bioenergy and Value-Added Products. Int. J. Environ. Sci. Technol. 2023, 20, 8199–8216. [Google Scholar] [CrossRef]
- Dayana Priyadharshini, S.; Suresh Babu, P.; Manikandan, S.; Subbaiya, R.; Govarthanan, M.; Karmegam, N. Phycoremediation of Wastewater for Pollutant Removal: A Green Approach to Environmental Protection and Long-Term Remediation. Environ. Pollut. 2021, 290, 117989. [Google Scholar] [CrossRef]
- Ibrahim, F.G.G.; Alonso Gómez, V.; Muñoz Torre, R.; de Godos Crespo, I. Scale-down of High-Rate Algae Ponds Systems for Urban Wastewater Reuse. J. Water Process Eng. 2023, 56, 104342. [Google Scholar] [CrossRef]
- Jebali, A.; Acién, F.G.; Rodriguez Barradas, E.; Olguín, E.J.; Sayadi, S.; Molina Grima, E. Pilot-Scale Outdoor Production of Scenedesmus Sp. in Raceways Using Flue Gases and Centrate from Anaerobic Digestion as the Sole Culture Medium. Bioresour. Technol. 2018, 262, 1–8. [Google Scholar] [CrossRef]
- Villalba, M.R.; Cervera, R.; Sánchez, J. Green Solutions for Urban Sustainability: Photobioreactors for Algae Cultivation on Façades and Artificial Trees. Buildings 2023, 13, 1541. [Google Scholar] [CrossRef]
- Takache, H.; Christophe, G.; Cornet, J.F.; Pruvost, J. Experimental and Theoretical Assessment of Maximum Productivities for the Microalgae Chlamydomonas Reinhardtii in Two Different Geometries of Photobioreactors. Biotechnol. Prog. 2010, 26, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Solsvik, J.; Wagner, J.L.; Zhang, D.; Hellgardt, K.; Park, C.W. CFD and Kinetic-Based Modeling to Optimize the Sparger Design of a Large-Scale Photobioreactor for Scaling up of Biofuel Production. Biotechnol. Bioeng. 2019, 116, 2200–2211. [Google Scholar] [CrossRef] [PubMed]
- Giannelli, L.; Torzillo, G. Hydrogen Production with the Microalga Chlamydomonas Reinhardtii Grown in a Compact Tubular Photobioreactor Immersed in a Scattering Light Nanoparticle Suspension. Int. J. Hydrogen Energy 2012, 37, 16951–16961. [Google Scholar] [CrossRef]
- Martinez-Carvajal, G.D.; Taidi, B.; Jarrahi, M. Towards a Low Energy, Stirless Photobioreactor Using Photosynthetic Motile Microalgae. Algal Res. 2024, 77, 103350. [Google Scholar] [CrossRef]
- Rodríguez-Bolaños, M.; Vargas-Romero, G.; Jaguer-García, G.; Aguilar-Gonzalez, Z.I.; Lagos-Romero, V.; Miranda-Astudillo, H. V. Antares I: A Modular Photobioreactor Suitable for Photosynthesis and Bioenergetics Research. Appl. Biochem. Biotechnol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Moreno Osorio, J.H.; Pollio, A.; Frunzo, L.; Lens, P.N.L.; Esposito, G. A Review of Microalgal Biofilm Technologies: Definition, Applications, Settings and Analysis. Front. Chem. Eng. 2021, 3, 737710. [Google Scholar] [CrossRef]
- Kreis, C.T.; Grangier, A.; Bäumchen, O. In Vivo Adhesion Force Measurements of Chlamydomonas on Model Substrates. Soft Matter 2019, 15, 3027–3035. [Google Scholar] [CrossRef] [PubMed]
- Catalan, R.E.; Fragkopoulos, A.A.; von Trott, N.; Kelterborn, S.; Baidukova, O.; Hegemann, P.; Bäumchen, O. Light-Regulated Adsorption and Desorption of Chlamydomonas Cells at Surfaces. Soft Matter 2023, 19, 306. [Google Scholar] [CrossRef] [PubMed]
- Vishwakarma, J.; Waghela, B.; Falcao, B.; Vavilala, S.L. Algal Polysaccharide’s Potential to Combat Respiratory Infections Caused by Klebsiella Pneumoniae and Serratia Marcescens Biofilms. Appl. Biochem. Biotechnol. 2022, 194, 671–693. [Google Scholar] [CrossRef]
- Vieira, M. V.; Pastrana, L.M.; Fuciños, P. Microalgae Encapsulation Systems for Food, Pharmaceutical and Cosmetics Applications. Mar. Drugs 2020, 18, 644. [Google Scholar] [CrossRef]
- Han, M.; Zhang, C.; Ho, S.H. Immobilized Microalgal System: An Achievable Idea for Upgrading Current Microalgal Wastewater Treatment. Environ. Sci. Ecotechnology 2023, 14, 100227. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Xie, P.; Ren, N.; Ho, S.H. Cytoprotective Alginate Microcapsule Serves as a Shield for Microalgal Encapsulation Defensing Sulfamethoxazole Threats and Safeguarding Nutrient Recovery. J. Hazard. Mater. 2024, 465, 133454. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Jeong, D.; Im, S.J.; Jang, A. Optimization of Alginate Bead Size Immobilized with Chlorella Vulgaris and Chlamydomonas Reinhardtii for Nutrient Removal. Bioresour. Technol. 2020, 302, 122891. [Google Scholar] [CrossRef] [PubMed]
- Homburg, S.V.; Kruse, O.; Patel, A. V. Growth and Photosynthetic Activity of Chlamydomonas Reinhardtii Entrapped in Lens-Shaped Silica Hydrogels. J. Biotechnol. 2019, 302, 58–66. [Google Scholar] [CrossRef]
- Zhang, B.B.; Wang, L.; Charles, V.; Rooke, J.C.; Su, B.L. Robust and Biocompatible Hybrid Matrix with Controllable Permeability for Microalgae Encapsulation. ACS Appl. Mater. Interfaces 2016, 8, 8939–8946. [Google Scholar] [CrossRef] [PubMed]
- Mandsberg, N.K.; Liao, W.; Yamanouchi, Y.A.; Boisen, A.; Ejima, H. Encapsulation of Chlamydomonas Reinhardtii into a Metal-Phenolic Network. Algal Res. 2022, 61, 102569. [Google Scholar] [CrossRef]
- Leong, Y.K.; Chang, J.S. Bioremediation of Heavy Metals Using Microalgae: Recent Advances and Mechanisms. Bioresour. Technol. 2020, 303, 122886. [Google Scholar] [CrossRef]
- Hoyos, B.S.; Hernandez-Tenorio, F.; Miranda, A.M.; Villanueva-Mejía, D.F.; Sáez, A.A. Systematic Analysis of Genes Related to Selenium Bioaccumulation in Microalgae: A Review. Biology (Basel). 2023, 12, 703. [Google Scholar] [CrossRef]
- Touliabah, H.E.S.; El-Sheekh, M.M.; Ismail, M.M.; El-Kassas, H. A Review of Microalgae-and Cyanobacteria-Based Biodegradation of Organic Pollutants. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Arora, N.; Patel, A.; Sartaj, K.; Pruthi, P.A.; Pruthi, V. Bioremediation of Domestic and Industrial Wastewaters Integrated with Enhanced Biodiesel Production Using Novel Oleaginous Microalgae. Env. Sci Pollut Res 2016, 23, 20997–21007. [Google Scholar] [CrossRef]
- Hasan, R. Bioremediation of Swine Wastewater and Biofuel Potential by Using Chlorella Vulgaris, Chlamydomonas Reinhardtii, and Chlamydomonas Debaryana. J. Pet. Environ. Biotechnol. 2014, 05, 3–7. [Google Scholar] [CrossRef]
- Kong, Q.X.; Li, L.; Martinez, B.; Chen, P.; Ruan, R. Culture of Microalgae Chlamydomonas Reinhardtii in Wastewater for Biomass Feedstock Production. Appl. Biochem. Biotechnol. 2010, 160, 9–18. [Google Scholar] [CrossRef]
- Sasi, P.; Viswanathan, A.; Mechery, J.; Thomas, D.; Jacob, J.; Paulose, S. Phycoremediation of Paper and Pulp Mill Effluent Using Planktochlorella Nurekis and Chlamydomonas Reinhardtii-A Comparative Study. J. Environ. Treat. Tech. 2020, 809–817. [Google Scholar]
- Abou-Shanab, R.A.I.; Ji, M.-K.; Kim, H.-C.; Paeng, K.-J.; Jeon, B.-H. Microalgal Species Growing on Piggery Wastewater as a Valuable Candidate for Nutrient Removal and Biodiesel Production. J. Environ. Manage. 2013, 115, 257–264. [Google Scholar] [CrossRef]
- Leong, Y.K.; Huang, C.Y.; Chang, J.S. Pollution Prevention and Waste Phycoremediation by Algal-Based Wastewater Treatment Technologies: The Applications of High-Rate Algal Ponds (HRAPs) and Algal Turf Scrubber (ATS). J. Environ. Manage. 2021, 296, 113193. [Google Scholar] [CrossRef] [PubMed]
- Grönlund, E.; Hanaeus, J.; Johansson, E.; Falk, S. Performance of an Experimental Wastewater Treatment High-Rate Algal Pond in Subarctic Climate. Water Environ. Res. a Res. Publ. Water Environ. Fed. 2010, 82, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Godos, I. de; Blanco, S.; García-Encina, P.A.; Becares, E.; Muñoz, R. Long-Term Operation of High Rate Algal Ponds for the Bioremediation of Piggery Wastewaters at High Loading Rates. Bioresour. Technol. 2009, 100, 4332–4339. [Google Scholar] [CrossRef] [PubMed]
- Bohutskyi, P.; Phan, D.; Spierling, R.E.; Kopachevsky, A.M.; Bouwer, E.J.; Lundquist, T.J.; Betenbaugh, M.J. Production of Lipid-Containing Algal-Bacterial Polyculture in Wastewater and Biomethanation of Lipid Extracted Residues: Enhancing Methane Yield through Hydrothermal Pretreatment and Relieving Solvent Toxicity through Co-Digestion. Sci. Total Environ. 2019, 653, 1377–1394. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Yu, T.; Xie, Y.; Chen, J.; Ho, S.H.; Wang, Y.; Huang, F. Attached Culture of Chlamydomonas Sp. JSC4 for Biofilm Production and TN/TP/Cu(II) Removal. Biochem. Eng. J. 2019, 141, 1–9. [Google Scholar] [CrossRef]
- Schaedig, E.; Cantrell, M.; Urban, C.; Zhao, X.; Greene, D.; Dancer, J.; Gross, M.; Sebesta, J.; Chou, K.J.; Grabowy, J.; et al. Isolation of Phosphorus-Hyperaccumulating Microalgae from Revolving Algal Biofilm (RAB) Wastewater Treatment Systems. Front. Microbiol. 2023, 14, 1219318. [Google Scholar] [CrossRef] [PubMed]
- de-Bashan, L.E.; Bashan, Y. Immobilized Microalgae for Removing Pollutants: Review of Practical Aspects. Bioresour. Technol. 2010, 101, 1611–1627. [Google Scholar] [CrossRef] [PubMed]
- Nazos, T.T.; Ghanotakis, D.F. Biodegradation of Phenol by Alginate Immobilized Chlamydomonas Reinhardtii Cells. Arch. Microbiol. 2021, 203, 5805–5816. [Google Scholar] [CrossRef]
- Saavedra, R.; Muñoz, R.; Taboada, M.E.; Vega, M.; Bolado, S. Comparative Uptake Study of Arsenic, Boron, Copper, Manganese and Zinc from Water by Different Green Microalgae. Bioresour. Technol. 2018, 263, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.; Han, B.; Kong, F.; You, T.; Bi, R.; Zeng, X.; Wang, S.; Jia, Y. Enhancement of Arsenic Uptake and Accumulation in Green Microalga Chlamydomonas Reinhardtii through Heterologous Expression of the Phosphate Transporter DsPht1. J. Hazard. Mater. 2023, 459, 132130. [Google Scholar] [CrossRef]
- Nam, S.H.; Kwak, J. Il; An, Y.J. Assessing Applicability of the Paper-Disc Method Used in Combination with Flow Cytometry to Evaluate Algal Toxicity. Environ. Pollut. 2018, 234, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Ibuot, A.; Webster, R.E.; Williams, L.E.; Pittman, J.K. Increased Metal Tolerance and Bioaccumulation of Zinc and Cadmium in Chlamydomonas Reinhardtii Expressing a AtHMA4 C-Terminal Domain Protein. Biotechnol. Bioeng. 2020, 117, 2996–3005. [Google Scholar] [CrossRef] [PubMed]
- Baselga-Cervera, B.; García-Balboa, C.; Díaz-Alejo, H.M.; Costas, E.; López-Rodas, V. Rapid Colonization of Uranium Mining-Impacted Waters, the Biodiversity of Successful Lineages of Phytoplankton Extremophiles. Microb. Ecol. 2020, 79, 576–587. [Google Scholar] [CrossRef] [PubMed]
- Balzano, S.; Sardo, A.; Blasio, M.; Chahine, T.B.; Dell’Anno, F.; Sansone, C.; Brunet, C. Microalgal Metallothioneins and Phytochelatins and Their Potential Use in Bioremediation. Front. Microbiol. 2020, 11, 517. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Tang, Y.; Yu, F.; Peng, Z.; Yao, S.; Deng, X.; Long, H.; Wang, X.; Huang, K. Translatomics and Physiological Analyses of the Detoxification Mechanism of Green Alga Chlamydomonas Reinhardtii to Cadmium Toxicity. J. Hazard. Mater. 2023, 448, 130990. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zhang, B.; Li, Z.; Deng, P.; Deng, X.; Long, H.; Wang, X.; Huang, K. Overexpression of the Sulfate Transporter-Encoding SULTR2 Increases Chromium Accumulation in Chlamydomonas Reinhardtii. Biotechnol. Bioeng. 2023, 120, 1334–1345. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, P.; Fu, H.; Chen, J.; Ye, M.; Zhai, S.; Hu, F.; Zhang, C.; Ge, Y.; Fortin, C. A Comparative Study of the Accumulation and Detoxification of Copper and Zinc in Chlamydomonas Reinhardtii: The Role of Extracellular Polymeric Substances. Sci. Total Environ. 2023, 871, 161995. [Google Scholar] [CrossRef] [PubMed]
- Millet, R.T.; Santos, J.P.; Slaveykova, V.I. Exploring the Subcellular Distribution of Mercury in Green Alga Chlamydomonas Reinhardtii and Diatom Cyclotella Meneghiniana : A Comparative Study. Aquat. Toxicol. 2024, 267, 106836. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Jiang, Z.; Yu, H.; Li, Z.; Zhang, C.; Ge, Y. Assessment of Joint Toxicity of Arsenate and Lead by Multiple Endpoints in Chlamydomonas Reinhardtii. Bull. Environ. Contam. Toxicol. 2023, 111, 30. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhao, Z.; Yan, Z.; Zhou, G.; Zhang, W.; Wang, Y.; Li, X. Defense Pathways of Chlamydomonas Reinhardtii under Silver Nanoparticle Stress: Extracellular Biosorption, Internalization and Antioxidant Genes. Chemosphere 2022, 291, 132764. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.P.; Luo, K.; Zhang, S.; Zheng, Q.; Yang, H. Bioaccumulation and Catabolism of Prometryne in Green Algae. Chemosphere 2012, 87, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Cao, J.; Ma, X.; Ping, J.; Zhang, C.; Ke, T.; Zhang, Y.; Tao, Y.; Chen, L. The Simultaneous Removal of the Combined Pollutants of Hexavalent Chromium and O-Nitrophenol by Chlamydomonas Reinhardtii. Ecotoxicol. Environ. Saf. 2020, 198, 110648. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.Q.; Kurade, M.B.; Abou-Shanab, R.A.I.; Ji, M.K.; Choi, J.; Kim, J.O.; Jeon, B.H. Biodegradation of Carbamazepine Using Freshwater Microalgae Chlamydomonas Mexicana and Scenedesmus Obliquus and the Determination of Its Metabolic Fate. Bioresour. Technol. 2016, 205, 183–190. [Google Scholar] [CrossRef]
- Wan, L.; Wu, Y.; Ding, H.; Zhang, W. Toxicity, Biodegradation, and Metabolic Fate of Organophosphorus Pesticide Trichlorfon on the Freshwater Algae Chlamydomonas Reinhardtii. J. Agric. Food Chem. 2020, 68, 1645–1653. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Deng, J.; Cui, L.; Chang, P.; Dai, X.; Yang, C.; Li, N.; Ren, Z.; Zhang, X. The Potential Assessment of Green Alga Chlamydomonas Reinhardtii CC-503 in the Biodegradation of Benz(a)Anthracene and the Related Mechanism Analysis. Chemosphere 2020, 249, 126097. [Google Scholar] [CrossRef]
- Li, S.; Wang, P.; Zhang, C.; Zhou, X.; Yin, Z.; Hu, T.; Hu, D.; Liu, C.; Zhu, L. Influence of Polystyrene Microplastics on the Growth, Photosynthetic Efficiency and Aggregation of Freshwater Microalgae Chlamydomonas Reinhardtii. Sci. Total Environ. 2020, 714, 136767. [Google Scholar] [CrossRef]
- Carbó, M.; Chaturvedi, P.; Álvarez, A.; Pineda-Cevallos, D.; Ghatak, A.; González, P.R.; Cañal, M.J.; Weckwerth, W.; Valledor, L. Ferroptosis Is the Key Cellular Process Mediating Bisphenol A Responses in Chlamydomonas and a Promising Target for Enhancing Microalgae-Based Bioremediation. J. Hazard. Mater. 2023, 448, 130997. [Google Scholar] [CrossRef]
- Hena, S.; Gutierrez, L.; Croué, J.P. Removal of Pharmaceutical and Personal Care Products (PPCPs) from Wastewater Using Microalgae: A Review. J. Hazard. Mater. 2021, 403, 124041. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.Q.; Zheng, H.S.; Li, S.; Du, J.S.; Feng, X.C.; Yin, R.L.; Wu, Q.L.; Ren, N.Q.; Chang, J.S. Removal of Cephalosporin Antibiotics 7-ACA from Wastewater during the Cultivation of Lipid-Accumulating Microalgae. Bioresour. Technol. 2016, 221, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.Q.; Kurade, M.B.; Jeon, B.H. Ecotoxicological Effects of Enrofloxacin and Its Removal by Monoculture of Microalgal Species and Their Consortium. Environ. Pollut. 2017, 226, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.-J.; Ying, G.-G.; Liu, S.; Zhou, L.-J.; Chen, Z.-F.; Peng, F.-Q. Simultaneous Removal of Inorganic and Organic Compounds in Wastewater by Freshwater Green Microalgae. Environ. Sci. Process. Impacts 2014, 16, 2018–2027. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Dong, S.; Huang, F.; Lin, L.; Hu, Z.; Zheng, Y. Toxicological Effects of Microplastics and Sulfadiazine on the Microalgae Chlamydomonas Reinhardtii. Front. Microbiol. 2022, 13, 865768. [Google Scholar] [CrossRef] [PubMed]
- Seoane, M.; Conde-Pérez, K.; Esperanza, M.; Cid, Á.; Rioboo, C. Unravelling Joint Cytotoxicity of Ibuprofen and Oxytetracycline on Chlamydomonas Reinhardtii Using a Programmed Cell Death-Related Biomarkers Panel. Aquat. Toxicol. 2023, 257, 106455. [Google Scholar] [CrossRef] [PubMed]
- Hom-Diaz, A.; Jaén-Gil, A.; Rodríguez-Mozaz, S.; Barceló, D.; Vicent, T.; Blánquez, P. Insights into Removal of Antibiotics by Selected Microalgae (Chlamydomonas Reinhardtii, Chlorella Sorokiniana, Dunaliella Tertiolecta and Pseudokirchneriella Subcapitata). Algal Res. 2022, 61, 102560. [Google Scholar] [CrossRef]
- Hom-Diaz, A.; Llorca, M.; Rodríguez-Mozaz, S.; Vicent, T.; Barceló, D.; Blánquez, P. Microalgae Cultivation on Wastewater Digestate: β-Estradiol and 17α-Ethynylestradiol Degradation and Transformation Products Identification. J. Environ. Manage. 2015, 155, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Liakh, I.; Harshkova, D.; Hrouzek, P.; Bišová, K.; Aksmann, A.; Wielgomas, B. Green Alga Chlamydomonas Reinhardtii Can Effectively Remove Diclofenac from the Water Environment – A New Perspective on Biotransformation. J. Hazard. Mater. 2023, 455, 131570. [Google Scholar] [CrossRef] [PubMed]
- Stravs, M.A.; Pomati, F.; Hollender, J. Exploring Micropollutant Biotransformation in Three Freshwater Phytoplankton Species. Environ. Sci. Process. Impacts 2017, 19, 822–832. [Google Scholar] [CrossRef] [PubMed]
- Otto, B.; Beuchel, C.; Liers, C.; Reisser, W.; Harms, H.; Schlosser, D. Laccase-like Enzyme Activities from Chlorophycean Green Algae with Potential for Bioconversion of Phenolic Pollutants. FEMS Microbiol. Lett. 2015, 362, fnv072. [Google Scholar] [CrossRef]
- Míguez, L.; Esperanza, M.; Seoane, M.; Cid, Á. Assessment of Cytotoxicity Biomarkers on the Microalga Chlamydomonas Reinhardtii Exposed to Emerging and Priority Pollutants. Ecotoxicol. Environ. Saf. 2021, 208, 111646. [Google Scholar] [CrossRef]
- Yadav, N.; Ahn, H.J.; Kurade, M.B.; Ahn, Y.; Park, Y.K.; Khan, M.A.; Salama, E.S.; Li, X.; Jeon, B.H. Fate of Five Bisphenol Derivatives in Chlamydomonas Mexicana: Toxicity, Removal, Biotransformation and Microalgal Metabolism. J. Hazard. Mater. 2023, 454, 131504. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Hiraga, K.; Takehana, T.; Taniguchi, I.; Yamaji, H.; Maeda, Y.; Toyohara, K.; Miyamoto, K.; Kimura, Y.; Oda, K. A Bacterium That Degrades and Assimilates Poly(Ethylene Terephthalate). Science (80-. ). 2016, 351, 6278. [Google Scholar] [CrossRef] [PubMed]
- Di Rocco, G.; Taunt, H.N.; Berto, M.; Jackson, H.O.; Piccinini, D.; Carletti, A.; Scurani, G.; Braidi, N.; Purton, S. A PETase Enzyme Synthesised in the Chloroplast of the Microalga Chlamydomonas Reinhardtii Is Active against Post-Consumer Plastics. Sci. Rep. 2023, 13, 10028. [Google Scholar] [CrossRef] [PubMed]
- de Carpentier, F.; Maes, A.; Marchand, C.H.; Chung, C.; Durand, C.; Crozet, P.; Lemaire, S.D.; Danon, A. How Abiotic Stress-Induced Socialization Leads to the Formation of Massive Aggregates in Chlamydomonas. Plant Physiol. 2022, 190, 1927–1940. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Y.; Chen, Z.; Gu, P.; Li, X.; Wang, G. Exploring Cell Aggregation as a Defense Strategy against Perchlorate Stress in Chlamydomonas Reinhardtii through Multi-Omics Analysis. Sci. Total Environ. 2023, 905, 167045. [Google Scholar] [CrossRef]
- Nakanishi, A.; Sakihama, Y.; Ozawa, N. Improvement of Growth of Chlamydomonas Reinhardtii in CO2 – Stepwisely Aerating Condition. J. Appl. Biotechnol. Reports 2021, 8, 37–40. [Google Scholar] [CrossRef]
- Hariz, H.B.; Takriff, M.S.; Mohd Yasin, N.H.; Ba-Abbad, M.M.; Mohd Hakimi, N.I.N. Potential of the Microalgae-Based Integrated Wastewater Treatment and CO2 Fixation System to Treat Palm Oil Mill Effluent (POME) by Indigenous Microalgae; Scenedesmus Sp. and Chlorella Sp. J. Water Process Eng. 2019, 32, 100907. [Google Scholar] [CrossRef]
- Choi, H. Il; Hwang, S.W.; Kim, J.; Park, B.; Jin, E.S.; Choi, I.G.; Sim, S.J. Augmented CO2 Tolerance by Expressing a Single H+-Pump Enables Microalgal Valorization of Industrial Flue Gas. Nat. Commun. 2021, 12, 6049. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.; Tahir, F.; Ashfaq, H.; Akbar, I.; Razzaque, N.; Haider, M.N.; Xu, J.; Zhu, H.; Wang, N.; Shahid, A. Decontamination of Industrial Wastewater Using Microalgae Integrated with Biotransformation of the Biomass to Green Products. Energy Nexus 2022, 6, 100089. [Google Scholar] [CrossRef]
- Ahmad, A.; W. Hassan, S.; Banat, F. An Overview of Microalgae Biomass as a Sustainable Aquaculture Feed Ingredient: Food Security and Circular Economy. Bioengineered 2022, 13, 9521–9547. [Google Scholar] [CrossRef]
- Ma, X.; Mi, Y.; Zhao, C.; Wei, Q. A Comprehensive Review on Carbon Source Effect of Microalgae Lipid Accumulation for Biofuel Production. Sci. Total Environ. 2022, 806. [Google Scholar] [CrossRef]
- Cheng, C.L.; Lo, Y.C.; Huang, K. Lou; Nagarajan, D.; Chen, C.Y.; Lee, D.J.; Chang, J.S. Effect of PH on Biomass Production and Carbohydrate Accumulation of Chlorella Vulgaris JSC-6 under Autotrophic, Mixotrophic, and Photoheterotrophic Cultivation. Bioresour. Technol. 2022, 351, 127021. [Google Scholar] [CrossRef]
- Beigbeder, J.B.; Lavoie, J.M. Effect of Photoperiods and CO2 Concentrations on the Cultivation of Carbohydrate-Rich P. Kessleri Microalgae for the Sustainable Production of Bioethanol. J. CO2 Util. 2022, 58, 101934. [Google Scholar] [CrossRef]
- Salman, J.M.; Grmasha, R.A.; Stenger-Kovács, C.; Lengyel, E.; Al-sareji, O.J.; AL-Cheban, A.M.A.A.; Meiczinger, M. Influence of Magnesium Concentrations on the Biomass and Biochemical Variations in the Freshwater Algae, Chlorella Vulgaris. Heliyon 2023, 9, e13072. [Google Scholar] [CrossRef] [PubMed]
- Fields, F.J.; Ostrand, J.T.; Mayfield, S.P. Fed-Batch Mixotrophic Cultivation of Chlamydomonas Reinhardtii for High-Density Cultures. Algal Res. 2018, 33, 109–117. [Google Scholar] [CrossRef]
- Torres-Tiji, Y.; Fields, F.J.; Yang, Y.; Heredia, V.; Horn, S.J.; Keremane, S.R.; Jin, M.M.; Mayfield, S.P. Optimized Production of a Bioactive Human Recombinant Protein from the Microalgae Chlamydomonas Reinhardtii Grown at High Density in a Fed-Batch Bioreactor. Algal Res. 2022, 66, 102786. [Google Scholar] [CrossRef]
- Leong, Y.K.; Chang, J.S. Microalgae-Based Biochar Production and Applications: A Comprehensive Review. Bioresour. Technol. 2023, 389, 129782. [Google Scholar] [CrossRef]
- Fan, X.; Du, C.; Zhou, L.; Fang, Y.; Zhang, G.; Zou, H.; Yu, G.; Wu, H. Biochar from Phytoremediation Plant Residues: A Review of Its Characteristics and Potential Applications. Environ. Sci. Pollut. Res. 2024, 31, 16188–16205. [Google Scholar] [CrossRef]
- Torri, C.; Samorì, C.; Adamiano, A.; Fabbri, D.; Faraloni, C.; Torzillo, G. Preliminary Investigation on the Production of Fuels and Bio-Char from Chlamydomonas Reinhardtii Biomass Residue after Bio-Hydrogen Production. Bioresour. Technol. 2011, 102, 8707–8713. [Google Scholar] [CrossRef]
- Gan, Y.Y.; Ong, H.C.; Show, P.L.; Ling, T.C.; Chen, W.H.; Yu, K.L.; Abdullah, R. Torrefaction of Microalgal Biochar as Potential Coal Fuel and Application as Bio-Adsorbent. Energy Convers. Manag. 2018, 165, 152–162. [Google Scholar] [CrossRef]
- Zheng, H.; Guo, W.; Li, S.; Chen, Y.; Wu, Q.; Feng, X.; Yin, R.; Ho, S.H.; Ren, N.; Chang, J.S. Adsorption of P-Nitrophenols (PNP) on Microalgal Biochar: Analysis of High Adsorption Capacity and Mechanism. Bioresour. Technol. 2017, 244, 1456–1464. [Google Scholar] [CrossRef] [PubMed]
- Miranda, A.M.; Hernandez-Tenorio, F.; Villalta, F.; Vargas, G.J.; Sáez, A.A. Advances in the Development of Biofertilizers and Biostimulants from Microalgae. Biology (Basel). 2024, 13. [Google Scholar] [CrossRef]
- Sido, M.Y.; Tian, Y.; Wang, X.; Wang, X. Application of Microalgae Chlamydomonas Applanata M9V and Chlorella Vulgaris S3 for Wheat Growth Promotion and as Urea Alternatives. Front. Microbiol. 2022, 13, 1035791. [Google Scholar] [CrossRef] [PubMed]
- Mutale-joan, C.; Redouane, B.; Najib, E.; Yassine, K.; Lyamlouli, K.; Laila, S.; Zeroual, Y.; Hicham, E.A. Screening of Microalgae Liquid Extracts for Their Bio Stimulant Properties on Plant Growth, Nutrient Uptake and Metabolite Profile of Solanum Lycopersicum L. Sci. Rep. 2020, 10, 2820. [Google Scholar] [CrossRef] [PubMed]
- Gitau, M.M.; Farkas, A.; Balla, B.; Ördög, V.; Futó, Z.; Maróti, G. Strain-Specific Biostimulant Effects of Chlorella and Chlamydomonas Green Microalgae on Medicago Truncatula. Plants 2021, 10, 1060. [Google Scholar] [CrossRef]
- Martini, F.; Beghini, G.; Zanin, L.; Varanini, Z.; Zamboni, A.; Ballottari, M. The Potential Use of Chlamydomonas Reinhardtii and Chlorella Sorokiniana as Biostimulants on Maize Plants. Algal Res. 2021, 60, 102515. [Google Scholar] [CrossRef] [PubMed]
- Stirk, W.A.; Ördög, V.; Staden, J. Van; Jäger, K. Cytokinin-and Auxin-like Activity in Cyanophyta and Microalgae. J. Appl. Phycol. 2002, 14, 215–221. [Google Scholar] [CrossRef]
- Metting, B. Population Dynamics of Chlamydomonas Sajao and Its Influence on Soil Aggregate Stabilization in the Field. Appl. Environ. Microbiol. 1986, 51, 1161–1164. [Google Scholar] [CrossRef] [PubMed]
- Arora, Y.; Sharma, S.; Sharma, V. Microalgae in Bioplastic Production: A Comprehensive Review. Arab. J. Sci. Eng. 2023, 48, 7225–7241. [Google Scholar] [CrossRef] [PubMed]
- Chaogang, W.; Zhangli, H.; Anping, L.; Baohui, J. Biosynthesis of Poly-3-Hydroxybutyrate (PHB) in the Transgenic Green Alga Chlamydomonas Reinhardtii. J. Phycol. 2010, 46, 396–402. [Google Scholar] [CrossRef]
- Hao, Z.; Songlin, M.; Xiaotan, D.; Ru, C.; Han, L.; Zhanyou, C.; Song, X.; Yonghua, L.-B.; Fantao, K. Harnessing Algal Peroxisomes for Efficient Poly Hydroxybutyrate Production. ACS Sustain. Chem. Eng 2024, 12, 3312–3321. [Google Scholar] [CrossRef]
- Kato, N. Production of Crude Bioplastic-Beads with Microalgae: Proof-of-Concept. Bioresour. Technol. Reports 2019, 5, 326–330. [Google Scholar] [CrossRef]
- Nakanishi, A.; Nemoto, S.; Yamamoto, N.; Iritani, K.; Watanabe, M. Identification of Cell-Attachment Factors Derived from Green Algal Cells Disrupted by Sonication in Fabrication of Cell Plastics. Bioengineering 2023, 10, 893. [Google Scholar] [CrossRef] [PubMed]
- Chaos-Hernández, D.; Reynel-Ávila, H.E.; Bonilla-Petriciolet, A.; Villalobos-Delgado, F.J. Extraction Methods of Algae Oils for the Production of Third Generation Biofuels – A Review. Chemosphere 2023, 341, 139856. [Google Scholar] [CrossRef]
- Daneshvar, E.; Sik Ok, Y.; Tavakoli, S.; Sarkar, B.; Shaheen, S.M.; Hong, H.; Luo, Y.; Rinklebe, J.; Song, H.; Bhatnagar, A. Insights into Upstream Processing of Microalgae: A Review. Bioresour. Technol. 2021, 329, 124870. [Google Scholar] [CrossRef] [PubMed]
- Sivaramakrishnan, R.; Suresh, S.; Kanwal, S.; Ramadoss, G.; Ramprakash, B.; Incharoensakdi, A. Microalgal Biorefinery Concepts’ Developments for Biofuel and Bioproducts: Current Perspective and Bottlenecks. Int. J. Mol. Sci. 2022, 23, 2623. [Google Scholar] [CrossRef]
- Chen, C.L.; Huang, C.C.; Ho, K.C.; Hsiao, P.X.; Wu, M.S.; Chang, J.S. Biodiesel Production from Wet Microalgae Feedstock Using Sequential Wet Extraction/Transesterification and Direct Transesterification Processes. Bioresour. Technol. 2015, 194, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Kao, P.H.; Ng, I.S. CRISPRi Mediated Phosphoenolpyruvate Carboxylase Regulation to Enhance the Production of Lipid in Chlamydomonas Reinhardtii. Bioresour. Technol. 2017, 245, 1527–1537. [Google Scholar] [CrossRef] [PubMed]
- Rengel, R.; Smith, R.T.; Haslam, R.P.; Sayanova, O.; Vila, M.; León, R. Overexpression of Acetyl-CoA Synthetase (ACS) Enhances the Biosynthesis of Neutral Lipids and Starch in the Green Microalga Chlamydomonas Reinhardtii. Algal Res. 2018, 31, 183–193. [Google Scholar] [CrossRef]
- Kong, F.; Liang, Y.; Légeret, B.; Beyly-Adriano, A.; Blangy, S.; Haslam, R.P.; Napier, J.A.; Beisson, F.; Peltier, G.; Li-Beisson, Y. Chlamydomonas Carries out Fatty Acid β-Oxidation in Ancestral Peroxisomes Using a Bona Fide Acyl-CoA Oxidase. Plant J. 2017, 90, 358–371. [Google Scholar] [CrossRef]
- Shin, Y.S.; Jeong, J.; Nguyen, T.H.T.; Kim, J.Y.H.; Jin, E.S.; Sim, S.J. Targeted Knockout of Phospholipase A2 to Increase Lipid Productivity in Chlamydomonas Reinhardtii for Biodiesel Production. Bioresour. Technol. 2019, 271, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.-F.; Lin, J.-Y.; Pan, K.-Y.; Huang, C.-K.; Chu, Y.-K. Overexpressing Ferredoxins in Chlamydomonas Reinhardtii Increase Starch and Oil Yields and Enhance Electric Power Production in a Photo Microbial Fuel Cell. Int. J. Mol. Sci. 2015, 16, 19308–19325. [Google Scholar] [CrossRef]
- Pancha, I.; Chokshi, K.; Tanaka, K.; Imamura, S. Microalgal Target of Rapamycin (TOR): A Central Regulatory Hub for Growth, Stress Response and Biomass Production. Plant Cell Physiol. 2020, 61, 675–684. [Google Scholar] [CrossRef]
- Tan, K.W.M.; Lee, Y.K. Expression of the Heterologous Dunaliella Tertiolecta Fatty Acyl-ACP Thioesterase Leads to Increased Lipid Production in Chlamydomonas Reinhardtii. J. Biotechnol. 2017, 247, 60–67. [Google Scholar] [CrossRef]
- Zhu, Z.; Yuan, G.; Fan, X.; Fan, Y.; Yang, M.; Yin, Y.; Liu, J.; Liu, Y.; Cao, X.; Tian, J.; et al. The Synchronous TAG Production with the Growth by the Expression of Chloroplast Transit Peptide-Fused ScPDAT in Chlamydomonas Reinhardtii. Biotechnol. Biofuels 2018, 11, 156. [Google Scholar] [CrossRef] [PubMed]
- Iskandarov, U.; Sitnik, S.; Shtaida, N.; Didi-Cohen, S.; Leu, S.; Khozin-Goldberg, I.; Cohen, Z.; Boussiba, S. Cloning and Characterization of a GPAT-like Gene from the Microalga Lobosphaera Incisa (Trebouxiophyceae): Overexpression in Chlamydomonas Reinhardtii Enhances TAG Production. J. Appl. Phycol. 2016, 28, 907–919. [Google Scholar] [CrossRef]
- Li, Y.; Han, D.; Hu, G.; Sommerfeld, M.; Hu, Q. Inhibition of Starch Synthesis Results in Overproduction of Lipids in Chlamydomonas Reinhardtii. Biotechnol. Bioeng. 2010, 107, 258–268. [Google Scholar] [CrossRef]
- Kato, Y.; Oyama, T.; Inokuma, K.; Vavricka, C.J.; Matsuda, M.; Hidese, R.; Satoh, K.; Oono, Y.; Chang, J.S.; Hasunuma, T.; et al. Enhancing Carbohydrate Repartitioning into Lipid and Carotenoid by Disruption of Microalgae Starch Debranching Enzyme. Commun. Biol. 2021, 4, 450. [Google Scholar] [CrossRef]
- Khoo, K.S.; Ahmad, I.; Chew, K.W.; Iwamoto, K.; Bhatnagar, A.; Show, P.L. Enhanced Microalgal Lipid Production for Biofuel Using Different Strategies Including Genetic Modification of Microalgae: A Review. Prog. Energy Combust. Sci. 2023, 96, 101071. [Google Scholar] [CrossRef]
- Zheng, S.; Zou, S.; Wang, H.; Feng, T.; Sun, S.; Chen, H.; Wang, Q. Reducing Culture Medium Nitrogen Supply Coupled with Replenishing Carbon Nutrient Simultaneously Enhances the Biomass and Lipid Production of Chlamydomonas Reinhardtii. Front. Microbiol. 2022, 13, 1019806. [Google Scholar] [CrossRef] [PubMed]
- Goodenough, U.; Blaby, I.; Casero, D.; Gallaher, S.D.; Goodson, C.; Johnson, S.; Lee, J.H.; Merchant, S.S.; Pellegrini, M.; Roth, R.; et al. The Path to Triacylglyceride Obesity in the Sta6 Strain of Chlamydomonas Reinhardtii. Eukaryot. Cell 2014, 13, 591–613. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Ahn, J.W.; Park, E.J.; Choi, J.L. Overexpression of S-Adenosylmethionine Synthetase in Recombinant Chlamydomonas for Enhanced Lipid Production. J. Microbiol. Biotechnol. 2023, 33, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Maltsev, Y.; Kulikovskiy, M.; Maltseva, S. Nitrogen and Phosphorus Stress as a Tool to Induce Lipid Production in Microalgae. Microb. Cell Fact. 2023, 22. [Google Scholar] [CrossRef]
- Gonzalez, D.I.; Ynalvez, R.A. Comparison of the Effects of Nitrogen-, Sulfur- and Combined Nitrogen- and Sulfur-Deprivations on Cell Growth, Lipid Bodies and Gene Expressions in Chlamydomonas Reinhardtii Cc5373-Sta6. BMC Biotechnol. 2023, 23, 35. [Google Scholar] [CrossRef]
- Salama, E.S.; Kim, H.C.; Abou-Shanab, R.A.I.; Ji, M.K.; Oh, Y.K.; Kim, S.H.; Jeon, B.H. Biomass, Lipid Content, and Fatty Acid Composition of Freshwater Chlamydomonas Mexicana and Scenedesmus Obliquus Grown under Salt Stress. Bioprocess Biosyst. Eng. 2013, 36, 827–833. [Google Scholar] [CrossRef] [PubMed]
- James, G.O.; Hocart, C.H.; Hillier, W.; Price, G.D.; Djordjevic, M.A. Temperature Modulation of Fatty Acid Profiles for Biofuel Production in Nitrogen Deprived Chlamydomonas Reinhardtii. Bioresour. Technol. 2013, 127, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Ge, Y.; Liu, K.; Yamaoka, Y.; Zhang, D.; Chi, Z.; Akkaya, M.; Kong, F. Overexpression of a MYB1 Transcription Factor Enhances Triacylglycerol and Starch Accumulation and Biomass Production in the Green Microalga Chlamydomonas Reinhardtii. J. Agric. Food Chem. 2023, 71, 17833–17841. [Google Scholar] [CrossRef]
- Singh, P.; Kumari, S.; Guldhe, A.; Misra, R.; Rawat, I.; Bux, F. Trends and Novel Strategies for Enhancing Lipid Accumulation and Quality in Microalgae. Renew. Sustain. Energy Rev. 2016, 55, 1–16. [Google Scholar] [CrossRef]
- Figueroa-Torres, G.M.; Pittman, J.K.; Theodoropoulos, C. Optimisation of Microalgal Cultivation via Nutrient-Enhanced Strategies: The Biorefinery Paradigm. Biotechnol. Biofuels 2021, 14, 64. [Google Scholar] [CrossRef] [PubMed]
- Poulhazan, A.; Arnold, A.A.; Mentink-Vigier, F.; Muszyński, A.; Azadi, P.; Halim, A.; Vakhrushev, S.Y.; Joshi, H.J.; Wang, T.; Warschawski, D.E.; et al. Molecular-Level Architecture of Chlamydomonas Reinhardtii’s Glycoprotein-Rich Cell Wall. Nat. Commun. 2024, 15, 986. [Google Scholar] [CrossRef]
- Choi, S.P.; Nguyen, M.T.; Sim, S.J. Enzymatic Pretreatment of Chlamydomonas Reinhardtii Biomass for Ethanol Production. Bioresour. Technol. 2010, 101, 5330–5336. [Google Scholar] [CrossRef]
- Nguyen, M.T.; Choi, S.P.; Lee, J.; Lee, J.H.; Sim, S.J. Hydrothermal Acid Pretreatment of Chlamydomonas Reinhardtii Biomass for Ethanol Production. J. Microbiol. Biotechnol. 2009, 19, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, I.N.; Zachleder, V.; Vítová, M.; Barbosa, M.J.; Bišová, K. Starch Production in Chlamydomonas Reinhardtii through Supraoptimal Temperature in a Pilot-Scale Photobioreactor. Cells 2021, 10, 1084. [Google Scholar] [CrossRef] [PubMed]
- De Marco, M.A.; Curatti, L.; Martínez-Noël, G.M.A. High Auxin Disrupts Expression of Cell-Cycle Genes, Arrests Cell Division and Promotes Accumulation of Starch in Chlamydomonas Reinhardtii. Algal Res. 2024, 78, 103419. [Google Scholar] [CrossRef]
- Qu, W.; Loke Show, P.; Hasunuma, T.; Ho, S.H. Optimizing Real Swine Wastewater Treatment Efficiency and Carbohydrate Productivity of Newly Microalga Chlamydomonas Sp. QWY37 Used for Cell-Displayed Bioethanol Production. Bioresour. Technol. 2020, 305, 123072. [Google Scholar] [CrossRef]
- Kunatsa, T.; Xia, X. A Review on Anaerobic Digestion with Focus on the Role of Biomass Co-Digestion, Modelling and Optimisation on Biogas Production and Enhancement. Bioresour. Technol. 2022, 344, 126311. [Google Scholar] [CrossRef] [PubMed]
- Jha, P.; Ghosh, S.; Panja, A.; Kumar, V.; Singh, A.K.; Prasad, R. Microalgae and Biogas: A Boon to Energy Sector. Environ. Sci. Pollut. Res. 2023. [Google Scholar] [CrossRef] [PubMed]
- Mussgnug, J.H.; Klassen, V.; Schlüter, A.; Kruse, O. Microalgae as Substrates for Fermentative Biogas Production in a Combined Biorefinery Concept. J. Biotechnol. 2010, 150, 51–56. [Google Scholar] [CrossRef]
- Veerabadhran, M.; Gnanasekaran, D.; Wei, J.; Yang, F. Anaerobic Digestion of Microalgal Biomass for Bioenergy Production, Removal of Nutrients and Microcystin: Current Status. J. Appl. Microbiol. 2021, 131, 1639–1651. [Google Scholar] [CrossRef] [PubMed]
- Klassen, V.; Blifernez-Klassen, O.; Wibberg, D.; Winkler, A.; Kalinowski, J.; Posten, C.; Kruse, O. Highly Efficient Methane Generation from Untreated Microalgae Biomass. Biotechnol. Biofuels 2017, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Zabed, H.M.; Akter, S.; Yun, J.; Zhang, G.; Zhang, Y.; Qi, X. Biogas from Microalgae: Technologies, Challenges and Opportunities. Renew. Sustain. Energy Rev. 2020, 117, 109503. [Google Scholar] [CrossRef]
- Fernández-Rodríguez, M.J.; de la Lama-Calvente, D.; Jiménez-Rodríguez, A.; Borja, R.; Rincón-Llorente, B. Influence of the Cell Wall of Chlamydomonas Reinhardtii on Anaerobic Digestion Yield and on Its Anaerobic Co-Digestion with a Carbon-Rich Substrate. Process Saf. Environ. Prot. 2019, 128, 167–175. [Google Scholar] [CrossRef]
- Barros, R.; Raposo, S.; Morais, E.G.; Rodrigues, B.; Afonso, V.; Gonçalves, P.; Marques, J.; Cerqueira, P.R.; Varela, J.; Teixeira, M.R.; et al. Biogas Production from Microalgal Biomass Produced in the Tertiary Treatment of Urban Wastewater: Assessment of Seasonal Variations. Energies 2022, 15, 5713. [Google Scholar] [CrossRef]
- Klassen, V.; Blifernez-Klassen, O.; Bax, J.; Kruse, O. Wastewater-Borne Microalga Chlamydomonas Sp.: A Robust Chassis for Efficient Biomass and Biomethane Production Applying Low-N Cultivation Strategy. Bioresour. Technol. 2020, 315, 123825. [Google Scholar] [CrossRef] [PubMed]
- Nirmala, N.; Praveen, G.; AmitKumar, S.; SundarRajan, P.S.; Baskaran, A.; Priyadharsini, P.; SanjayKumar, S.P.; Dawn, S.S.; Pavithra, K.G.; Arun, J.; et al. A Review on Biological Biohydrogen Production: Outlook on Genetic Strain Enhancements, Reactor Model and Techno-Economics Analysis. Sci. Total Environ. 2023, 896, 165143. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhou, Q.; Yu, D. The Future of Hydrogen Energy: Bio-Hydrogen Production Technology. Int. J. Hydrogen Energy 2022, 47, 33677–33698. [Google Scholar] [CrossRef]
- King, S.J.; Jerkovic, A.; Brown, L.J.; Petroll, K.; Willows, R.D. Synthetic Biology for Improved Hydrogen Production in Chlamydomonas Reinhardtii. Microb. Biotechnol. 2022, 15, 1946–1965. [Google Scholar] [CrossRef]
- Frenkel, A.W. Hydrogen Evolution by the Flagellate Green Alga, Chlamydomonas Moewusii. Arch Biochem Biophys 1951, 38, 219–230. [Google Scholar] [CrossRef]
- Ben-Amotz, A.; Erbes, D.L.; Riederer-Henderson, M.A.; Peavey, D.G.; Gibbs, M. H2 Metabolism in Photosynthetic Organisms: I. Dark H2 Evolution and Uptake by Algae and Mosses. Plant Physiol 1975, 56, 72–77. [Google Scholar] [CrossRef]
- Melis, A.; Zhang, L.; Forestier, M.; Ghirardi, M.L.; Seibert, M. Sustained Photobiological Hydrogen Gas Production upon Reversible Inactivation of Oxygen Evolution in the Green Alga Chlamydomonas Reinhardtii. Plant Physiol. 2000, 122, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Jurado-Oller, J.L.; Dubini, A.; Galván, A.; Fernández, E.; González-Ballester, D. Low Oxygen Levels Contribute to Improve Photohydrogen Production in Mixotrophic Non-Stressed Chlamydomonas Cultures. Biotechnol. Biofuels 2015, 8, 149. [Google Scholar] [CrossRef] [PubMed]
- Elman, T.; Yacoby, I. A Two-Phase Protocol for Ambient Hydrogen Production Using Chlamydomonas Reinhardtii. STAR Protoc. 2022, 3, 101640. [Google Scholar] [CrossRef] [PubMed]
- Fakhimi, N.; Gonzalez-Ballester, D.; Fernández, E.; Galván, A.; Dubini, A. Algae-Bacteria Consortia as a Strategy to Enhance H2 Production. Cells 2020, 9, 1353. [Google Scholar] [CrossRef]
- Elman, T.; Schweitzer, S.; Shahar, N.; Swartz, J.; Yacoby, I. Engineered Clostridial [FeFe]-Hydrogenase Shows Improved O2 Tolerance in Chlamydomonas Reinhardtii. Int. J. Hydrogen Energy 2020, 45, 30201–30210. [Google Scholar] [CrossRef]
- Nagy, V.; Podmaniczki, A.; Vidal-Meireles, A.; Kuntam, S.; Herman, É.; Kovács, L.; Tóth, D.; Scoma, A.; Tóth, S.Z. Thin Cell Layer Cultures of Chlamydomonas Reinhardtii L159I-N230Y, Pgrl1 and Pgr5 Mutants Perform Enhanced Hydrogen Production at Sunlight Intensity. Bioresour. Technol. 2021, 333, 125217. [Google Scholar] [CrossRef]
- Milrad, Y.; Schweitzer, S.; Feldman, Y.; Yacoby, I. Green Algal Hydrogenase Activity Is Outcompeted by Carbon Fixation before Inactivation by Oxygen Takes Place. Plant Physiol. 2018, 177, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Kosourov, S.; Nagy, V.; Shevela, D.; Jokel, M.; Messinger, J.; Allahverdiyeva, Y. Water Oxidation by Photosystem II Is the Primary Source of Electrons for Sustained H2 Photoproduction in Nutrient-Replete Green Algae. PNAS 2020, 117, 29629–29636. [Google Scholar] [CrossRef]
- Kanygin, A.; Milrad, Y.; Thummala, C.; Reifschneider, K.; Baker, P.; Marco, P.; Yacoby, I.; Redding, K.E. Rewiring Photosynthesis: A Photosystem I-Hydrogenase Chimera That Makes H2: In Vivo. Energy Environ. Sci. 2020, 13, 2903–2914. [Google Scholar] [CrossRef]
- Lakatos, G.; Balogh, D.; Farkas, A.; Ördög, V.; Nagy, P. .; Bíró, T.; Maróti, G. Factors Influencing Algal Photobiohydrogen Production in Algal-Bacterial Co-Cultures. Algal Res 2017, 28, 161–171. [Google Scholar] [CrossRef]
- Masi, A.; Leonelli, F.; Scognamiglio, V.; Gasperuzzo, G.; Antonacci, A. ; Terzidis, chael A. Chlamydomonas Reinhardtii: A Factory of Nutraceutical and Food Supplements for Human Health. Molecules 2023, 28, 1185. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y. Disruption of the Glycolate Dehydrogenase Gene in the High-CO2-Requiring Mutant HCR89 of Chlamydomonas Reinhardtii. Can. J. Bot. 2005, 87, 820–833. [Google Scholar] [CrossRef]
- Wang, Y.; Stessman, D.J.; Spalding, M.H. The CO2 Concentrating Mechanism and Photosynthetic Carbon Assimilation in Limiting CO2: How Chlamydomonas Works against the Gradient. Plant J. 2015, 82, 429–448. [Google Scholar] [CrossRef]
- Yun, E.J.; Zhang, G.-C.; Atkinson, C.; Lane, S.; Liu, J.-J.; Ort, D.R.; Jin, Y.-S. Glycolate Production by a Chlamydomonas Reinhardtii Mutant Lacking Carbon-Concentrating Mechanism. J. Biotechnol. 2021, 335, 39–46. [Google Scholar] [CrossRef]
- Taubert, A.; Jakob, T.; Wilhelm, C. Glycolate from Microalgae: An Efficient Carbon Source for Biotechnological Applications. Plant Biotechnol. J. 2019, 17, 1538–1546. [Google Scholar] [CrossRef] [PubMed]
- Zabaleta, E.; Wang, Y.; Zhao, L.; Shi, M. Identification and Characterization of Genes Encoding the Hydroxypyruvate Reductases in Chlamydomonas Reveal Their Distinct Roles in Photorespiration. Front. Plant Sci. 2021, 12, 690296. [Google Scholar] [CrossRef] [PubMed]
- Lauersen, K.J. Eukaryotic Microalgae as Hosts for Light-Driven Heterologous Isoprenoid Production. Planta 2019, 249, 155–180. [Google Scholar] [CrossRef]
- Yahya, R.Z.; Wellman, G.B.; Overmans, S.; Lauersen, K.J. Engineered Production of Isoprene from the Model Green Microalga Chlamydomonas Reinhardtii. Metab. Eng. Commun. 2023, 16, e00221. [Google Scholar] [CrossRef] [PubMed]
- Miró-Vinyals, B.; Artigues, M.; Wostrikoff, K.; Monte, E.; Broto-Puig, F.; Leivar, P.; Planas, A. Chloroplast Engineering of the Green Microalgae Chlamydomonas Reinhardtii for the Production of HAA, the Lipid Moiety of Rhamnolipid Biosurfactants. N. Biotechnol. 2023, 76, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Moulin, S.L.Y.; Beyly-Adriano, A.; Cuiné, S.; Blangy, S.; Légeret, B.; Floriani, M.; Burlacot, A.; Sorigué, D.; Samire, P.P.; Li-Beisson, Y.; et al. Fatty Acid Photodecarboxylase Is an Ancient Photoenzyme That Forms Hydrocarbons in the Thylakoids of Algae. Plant Physiol. 2021, 186, 1455–1472. [Google Scholar] [CrossRef] [PubMed]
- Salma-Ancane, K.; Sceglovs, A.; Tracuma, E.; Wychowaniec, J.K.; Aunina, K.; Ramata-Stunda, A.; Nikolajeva, V.; Loca, D. Effect of Crosslinking Strategy on the Biological, Antibacterial and Physicochemical Performance of Hyaluronic Acid and ɛ-Polylysine Based Hydrogels. Int. J. Biol. Macromol. 2022, 208, 995–1008. [Google Scholar] [CrossRef] [PubMed]
- Sivaramakrishnan, R.; Suresh, S.; Incharoensakdi, A. Chlamydomonas Sp. as Dynamic Biorefinery Feedstock for the Production of Methyl Ester and ɛ-Polylysine. Bioresour. Technol. 2019, 272, 281–287. [Google Scholar] [CrossRef]
- Lee, J.A.; Kim, J.Y.; Ahn, J.H.; Ahn, Y.-J.; Lee, S.Y. Current Advancements in the Bio-Based Production of Polyamides. Trends Chem. 2023, 5, 873–891. [Google Scholar] [CrossRef]
- Freudenberg, R.A.; Baier, T.; Einhaus, A.; Wobbe, L.; Kruse, O. High Cell Density Cultivation Enables Efficient and Sustainable Recombinant Polyamine Production in the Microalga Chlamydomonas Reinhardtii. Bioresour. Technol. 2021, 323, 124542. [Google Scholar] [CrossRef] [PubMed]
- Freudenberg, R.A.; Wittemeier, L.; Einhaus, A.; Baier, T.; Kruse, O. Advanced Pathway Engineering for Phototrophic Putrescine Production. Plant Biotechnol. J. 2022, 20, 1968–1982. [Google Scholar] [CrossRef] [PubMed]
- Fields, F.J.; Lejzerowicz, F.; Schroeder, D.; Ngoi, S.M.; Tran, M.; McDonald, D.; Jiang, L.; Chang, J.T.; Knight, R.; Mayfield, S. Effects of the Microalgae Chlamydomonas on Gastrointestinal Health. J. Funct. Foods 2020, 65, 103738. [Google Scholar] [CrossRef]
- Murbach, T.S.; Glávits, R.; Moghadam Maragheh, N.; Endres, J.R.; Hirka, G.; Goodman, R.E.; Lu, G.; Vértesi, A.; Béres, E.; Pasics Szakonyiné, I. Evaluation of the Genotoxic Potential of Protoporphyrin IX and the Safety of a Protoporphyrin IX-Rich Algal Biomass. J. Appl. Toxicol. 2022, 42, 1253–1275. [Google Scholar] [CrossRef]
- Grande, T.; Vornoli, A.; Lubrano, V.; Vizzarri, F.; Raffaelli, A.; Gabriele, M.; Novoa, J.; Sandoval, C.; Longo, V.; Echeverria, M.C.; et al. Chlamydomonas Agloeformis from the Ecuadorian Highlands: Nutrients and Bioactive Compounds Profiling and In Vitro Antioxidant Activity. Foods 2023, 12, 3147. [Google Scholar] [CrossRef] [PubMed]
- Bjørklund, G.; Gasmi, A.; Lenchyk, L.; Shanaida, M.; Zafar, S.; Mujawdiya, P.K.; Lysiuk, R.; Antonyak, H.; Noor, S.; Akram, M.; et al. The Role of Astaxanthin as a Nutraceutical in Health and Age-Related Conditions. Molecules 2022, 27, 7167. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.K.; Tambat, V.S.; Chen, C.W.; Chauhan, A.S.; Kumar, P.; Vadrale, A.P.; Huang, C.Y.; Dong, C. Di; Singhania, R.R. Recent Advancements in Astaxanthin Production from Microalgae: A Review. Bioresour. Technol. 2022, 364, 128030. [Google Scholar] [CrossRef]
- Perozeni, F.; Cazzaniga, S.; Baier, T.; Zanoni, F.; Zoccatelli, G.; Lauersen, K.J.; Wobbe, L.; Ballottari, M. Turning a Green Alga Red: Engineering Astaxanthin Biosynthesis by Intragenic Pseudogene Revival in Chlamydomonas Reinhardtii. Plant Biotechnol. J. 2020, 18, 2053–2067. [Google Scholar] [CrossRef]
- Cazzaniga, S.; Perozeni, F.; Baier, T.; Ballottari, M. Engineering Astaxanthin Accumulation Reduces Photoinhibition and Increases Biomass Productivity under High Light in Chlamydomonas Reinhardtii. Biotechnol. Biofuels Bioprod. 2022, 15, 17. [Google Scholar] [CrossRef] [PubMed]
- Tran, Q.G.; Cho, K.; Kim, U.; Yun, J.H.; Cho, D. hyun; Heo, J.; Park, S. Bin; Kim, J.W.; Lee, Y.J.; Ramanan, R.; et al. Enhancement of Β-Carotene Production by Regulating the Autophagy-Carotenoid Biosynthesis Seesaw in Chlamydomonas Reinhardtii. Bioresour. Technol. 2019, 292, 121937. [Google Scholar] [CrossRef] [PubMed]
- Del Mondo, A.; Smerilli, A.; Sané, E.; Sansone, C.; Brunet, C. Challenging Microalgal Vitamins for Human Health. Microb. Cell Fact. 2020, 19, 201. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Meireles, A.; Neupert, J.; Zsigmond, L.; Rosado-Souza, L.; Kovács, L.; Nagy, V.; Galambos, A.; Fernie, A.R.; Bock, R.; Tóth, S.Z. Regulation of Ascorbate Biosynthesis in Green Algae Has Evolved to Enable Rapid Stress-Induced Response via the VTC2 Gene Encoding GDP-l-Galactose Phosphorylase. New Phytol. 2017, 214, 668–681. [Google Scholar] [CrossRef] [PubMed]
- Darwish, R.; Gedi, M.A.; Akepach, P.; Assaye, H.; Zaky, A.S.; Gray, D.A. Chlamydomonas Reinhardtii Is a Potential Food Supplement with the Capacity to Outperform Chlorella and Spirulina. Appl. Sci. 2020, 10, 6736. [Google Scholar] [CrossRef]
- Kamble, P.; Cheriyamundath, S.; Lopus, M.; Sirisha, V.L. Chemical Characteristics, Antioxidant and Anticancer Potential of Sulfated Polysaccharides from Chlamydomonas Reinhardtii. J. Appl. Phycol. 2018, 30, 1641–1653. [Google Scholar] [CrossRef]
- Choudhary, S.; Save, S.N.; Vavilala, S.L. Unravelling the Inhibitory Activity of Chlamydomonas Reinhardtii Sulfated Polysaccharides against α-Synuclein Fibrillation. Sci. Rep. 2018, 8, 5692. [Google Scholar] [CrossRef] [PubMed]
- Vishwakarma, J.; Vavilala, S.L. Evaluating the Antibacterial and Antibiofilm Potential of Sulphated Polysaccharides Extracted from Green Algae Chlamydomonas Reinhardtii. J. Appl. Microbiol. 2019, 127, 1004–1017. [Google Scholar] [CrossRef] [PubMed]
- Arias, C.A.D.; Oliveira, C.F.M. de; Molino, J.V.D.; Ferreira-Camargo, L.S.; Matsudo, M.C.; Carvalho, J.C.M. de Production of Recombinant Biopharmaceuticals in Chlamydomonas Reinhardtii. Int. J. Plant Biol. 2023, 14, 39–52. [Google Scholar] [CrossRef]
- Kiefer, A.M.; Niemeyer, J.; Probst, A.; Erkel, G.; Schroda, M. Production and Secretion of Functional SARS-CoV-2 Spike Protein in Chlamydomonas Reinhardtii. Front. Plant Sci. 2022, 13, 988870. [Google Scholar] [CrossRef]
Figure 1.
The Chlamydomonas-based phycoremediation process for wastewater treatment. The diagram depicts the three main sources of wastewater. Traditional treatments (physical, mechanical, and chemical) are shown to be ineffective for treating complex wastewater compositions. These methods consume high energy and fail to address certain contaminants. Microalgae and macroalgae play a crucial role in phycoremediation, offering several benefits as indicated. The utilization of Chlamydomonas in phycoremediation, detailing the main cultivation methods, the employed mechanisms, and the various compounds that can be bioremediated, is illustrated.
Figure 1.
The Chlamydomonas-based phycoremediation process for wastewater treatment. The diagram depicts the three main sources of wastewater. Traditional treatments (physical, mechanical, and chemical) are shown to be ineffective for treating complex wastewater compositions. These methods consume high energy and fail to address certain contaminants. Microalgae and macroalgae play a crucial role in phycoremediation, offering several benefits as indicated. The utilization of Chlamydomonas in phycoremediation, detailing the main cultivation methods, the employed mechanisms, and the various compounds that can be bioremediated, is illustrated.
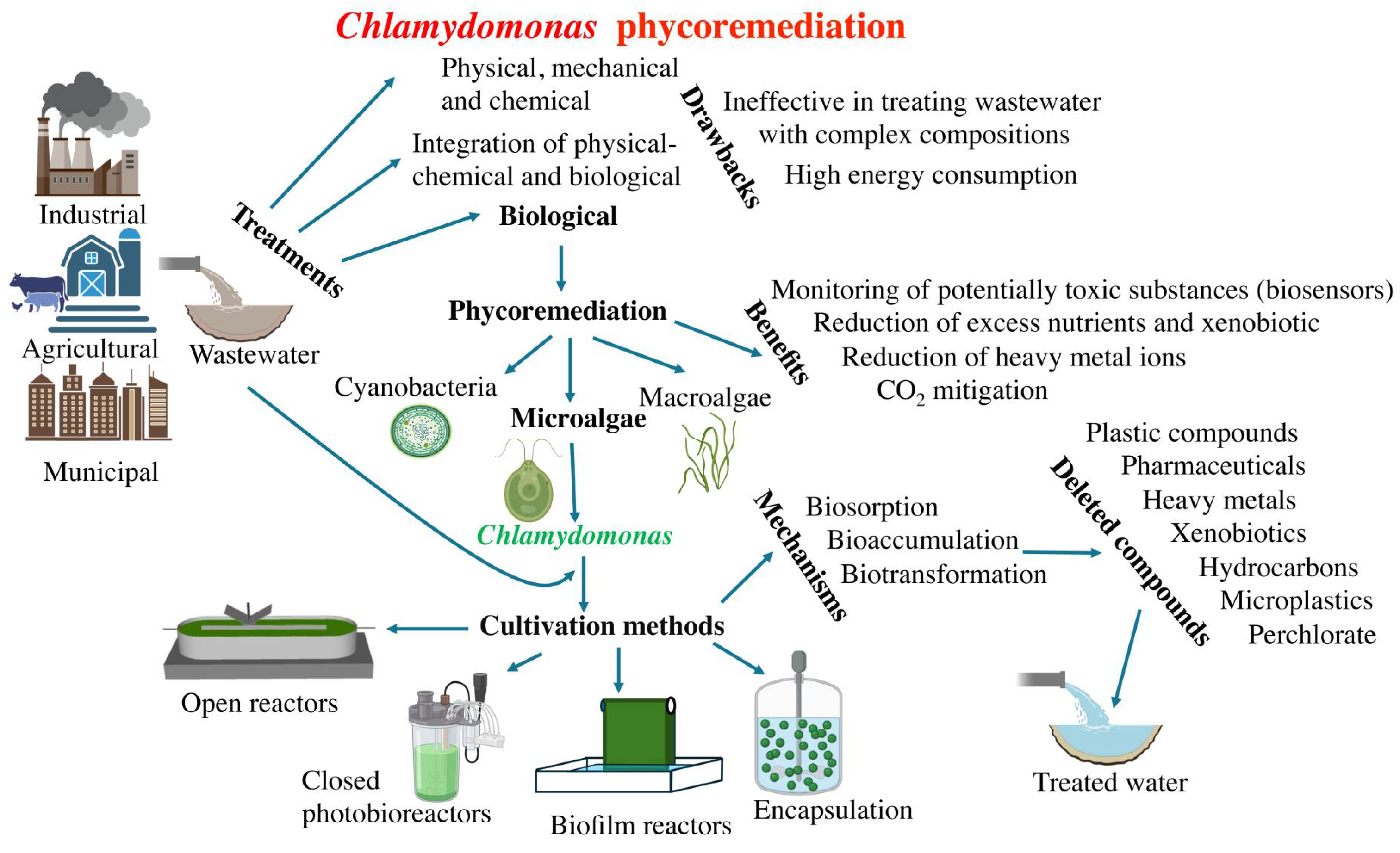
Figure 2.
The process of Chlamydomonas bioproduct generation. Chlamydomonas cells can be cultivated in various conditions. Cultivation optimization is a crucial factor and involves processes such as strain selection, nutrient availability, temperature control, light intensity, salinity, and pH levels, which impact the algae’s growth and productivity. After cultivation, the collected biomass and medium can then be processed as indicated to obtain the specified bioproducts.
Figure 2.
The process of Chlamydomonas bioproduct generation. Chlamydomonas cells can be cultivated in various conditions. Cultivation optimization is a crucial factor and involves processes such as strain selection, nutrient availability, temperature control, light intensity, salinity, and pH levels, which impact the algae’s growth and productivity. After cultivation, the collected biomass and medium can then be processed as indicated to obtain the specified bioproducts.
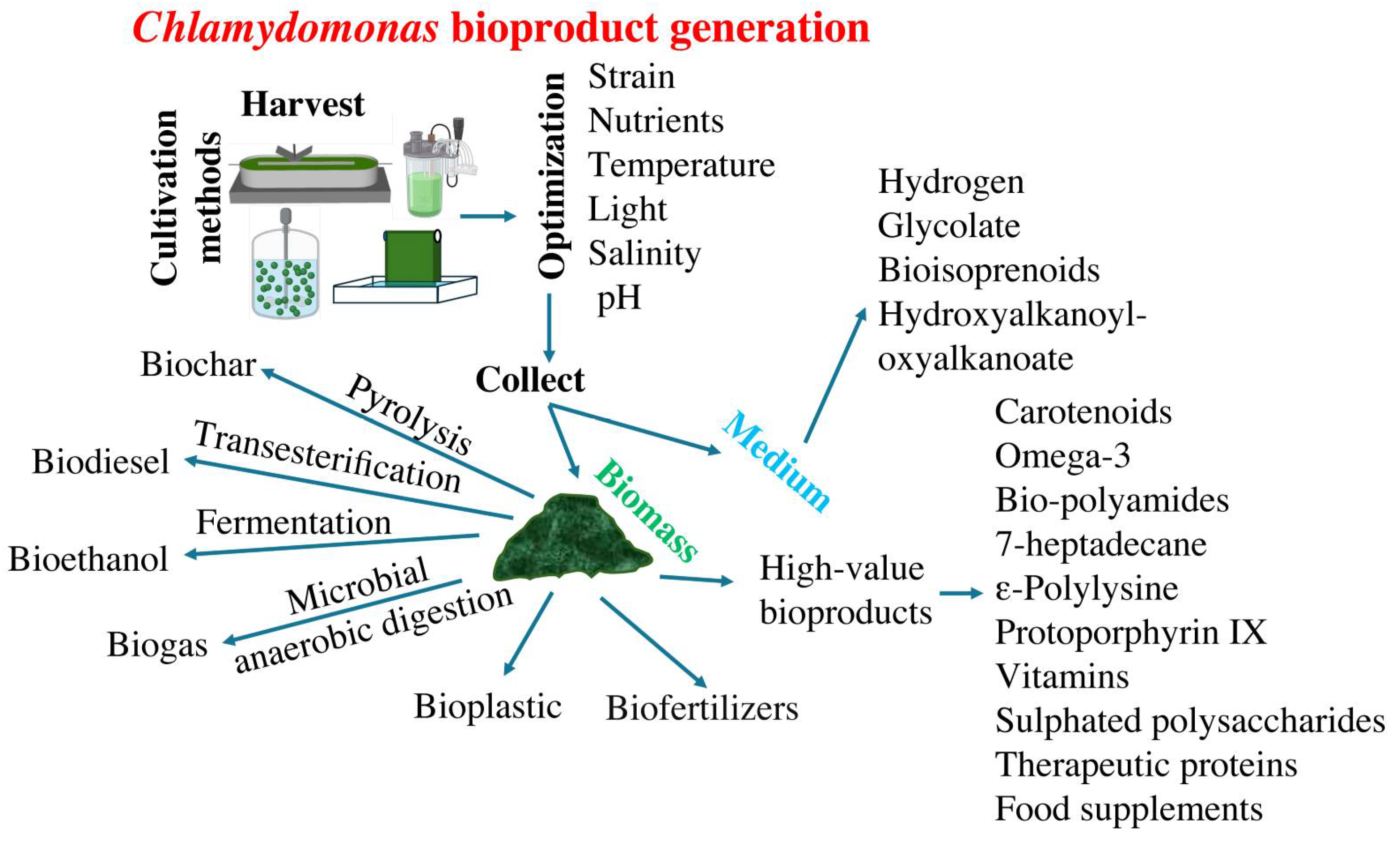
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



