Submitted:
29 May 2024
Posted:
30 May 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
: Cytokine storm is usually described as one of the main reasons behind COVID-associated mor-tality. Cytokines are essential protein molecules engaged in immune responses; they play a critical role in protection against infections. However, they also contribute to inflammatory reactions and tissue damage, becoming a double-edged sword in the context of COVID-19. Recent studies have suggested various cytokines and chemokines that play a crucial role in the immune response to SARS-CoV-2 infection. One such cytokine is Interleukin 27 (IL-27), which has been found to be elevated in the blood plasma of patients with COVID-19. Within this study, we will explore the role of IL-27 in immune responses and address our own findings concerning this cytokine in COVID-19. It affects a wide variety of immune cells; regardless of the pathological process it is involved in, IL-27 is critical for upholding the necessary balance between tissue damage and cytotoxicity against in-fectious agents and/or tumors. In COVID-19, it is involved in multiple processes, including antiviral cytotoxicity via CD8+ cells, IgG subclass switching, and even the activation of Tregs.
Keywords:
cytokines
; IL-27
; COVID-19
; cellular immunity
; inflammation
1. Introduction
The COVID-19 pandemic has caused a global health crisis with millions of confirmed cases and deaths worldwide [1]. COVID-19 is caused by the SARS-CoV-2 virus, which enters the body through the respiratory system and attaches to ACE2 (angiotensin converting enzyme II), widely present on cells in different parts of the body: the respiratory system, intestines, blood vessels, heart tissue, thyroid gland, epididymis, pancreas, liver, and even placenta [2]. Upon entry, the virus initiates immune reactions, which can cause inflammatory responses to become hyperergic. This hyperinflammation has the potential to cause a cytokine storm and widespread inflammation throughout the body, resulting in tissue damage to various organs such as the respiratory tract, heart, kidneys, and nervous system [3]. Cytokine storm is usually described as one of the main reasons behind COVID-associated mortality [4].
The virus primarily affects the respiratory system; therefore, symptoms of infection vary in their severity from the common cold to pneumonia with severe acute respiratory syndrome [5]. However, the pathophysiology behind COVID-19 is rather complex; it involves multiple organ systems, including the immune system.
Cytokines are essential protein molecules engaged in immune responses; they play a critical role in protection against infections. However, they also contribute to inflammatory reactions and tissue damage, becoming a double-edged sword in the context of COVID-19. Recent studies have suggested various cytokines and chemokines that play a crucial role in the immune response to SARS-CoV-2 infection. One such cytokine is Interleukin 27 (IL-27), which has been found to be elevated in the blood plasma of patients with COVID-19.
Within this study, we will explore the role of IL-27 in immune responses and address our own findings concerning this cytokine in COVID-19.
2. Interleukin 27 Conformation and Its Biological Function in Immunity
IL-27 consists of two subunits, IL-27p28 and EBI3, that combine to form a heterodimer. IL-27p28 (also known as IL-30) was initially discovered as an orphan homologue of the IL-6/12 family in 2002 through a computational analysis of expressed sequence tags [6]. In contrast, EBI3 was first identified in 1996 as an erythropoietin receptor-like protein induced by Epstein-Barr virus infection in B cells [7]. Antigen-presenting cells typically express both subunits of IL-27, and the secretion of human p28 is reliant on the secretion of human EBI3. However, mouse p28 can be secreted without relying on other factors [8]. The receptor for IL-27 (IL-27R) is also a heterodimer; it is composed of IL-27Rα and glycoprotein 130 (gp130). IL-27Rα is unique to IL-27R in the IL-6/IL-12 family, while gp130 is also a subunit of the IL-6 receptor and IL-35 receptor [9]. The IL-27 receptor is present on various cell types, such as DCs, monocytes, macrophages, neutrophils, mast cells, eosinophils, T, B, and NK cells.
The IL-30/EBI3 combination, known as IL-27, has both regulatory and anti-inflammatory functions. At first, it was believed to be similar to IL-12, which promotes Th1 immunity by inducing T-bet and IL12R-beta-2 expression through STAT1 activation during Th1 differentiation [10]. However, it was later found that IL-27 also has a strong inhibitory effect during Th17 differentiation [11].
IL-27 uses a STAT1-dependent mechanism to stop activated T cells from developing ROR-gamma-t and IL-17 expression [12]. It also inhibits Th2 immunity by preventing Th2 differentiation and cytokine production or by directly suppressing ILC2s [13]. IL-27 is involved in Treg suppression; it suppresses TGF-beta-induced Foxp3+ regulatory T cell differentiation. Therefore, IL-27 has a wide range of functions that control T-cell immunity by targeting multiple pathways [14]. IL-27 can also function in collaboration with other cytokines, including IL-6, IL-10, IL-11, and interferons [15,16,17]. The immunomodulatory effects of IL-27 on a number of adaptive and innate immune cells are presented in Figure 1. On the spectrum between pro- and anti-inflammatory effects, it tends to lean toward anti-inflammation [18].
Interestingly, IL-27 has both pro-inflammatory and anti-inflammatory properties that are mediated by STAT1 and STAT3 activation, respectively. However, its pro-inflammatory effects rely on the induction of T-bet and IL-12Rβ2 expression. IL-27 has been found to have a dual function in preventing tissue damage caused by excessive inflammation. Its impact on NK cells and their ability to regulate tumor growth have been observed in certain mouse models, but not in all of them, suggesting that the effects of IL-27 on mouse NK cells may vary depending on the type of tumor [19].
In other words, IL-27 has a varying spectrum of functions, and it may be difficult to predict the overall effects of this cytokine as its functional activity relies on several factors, such as signaling pathways and target cells.
In oncology, IL-27 is often discussed as a potential agent capable of stimulating anti-tumor immunity [20]. Various preclinical murine models have provided evidence that IL-27 has strong antitumor effects against different types of tumors. These effects are achieved through several mechanisms and do not seem to cause any negative effects. These involve CD8+ T cells, natural killer cells, and macrophages, as well as antibody-dependent cell-mediated cytotoxicity and inhibition of cyclooxygenase-2 and prostaglandin E2 expression, as well as direct effects on tumor cells, including inhibition of cell proliferation, tumor cell apoptosis [21] and suppression of epithelial-mesenchymal transition.
However, anti-tumor immunity is rarely achieved through the activity of only IL-27. In the review by Kourko et al., IL-27 effects are investigated in combination with other cytokines (IL-30, IL-35). Interactions of these so-called ‘sister’ cytokines have the ability to hinder the development of tumors by either interacting with cancer cells directly or indirectly by activating various types of immune cells. For instance, IL-27 and other cytokines increase NK cell-mediated cytotoxic activity, promote the cytotoxic activity of CD8+ effector T cells, and stimulate the generation of CD8+ memory T cells. At the same time, IL-27 provides certain pro-tumor effects: for instance, it is known to be produced by tumor cells and inhibits chemotherapeutic effects [22]. When tumor cells undergo apoptosis, they are endocytosed by antigen-presenting cells, which stimulate regulatory T cell activation [23].
Overall, in the context of cancer, IL-27 is mostly discussed as a pro-inflammatory cytokine that stimulates the anti-tumor effects via activation of tumor-specific cytotoxicity. Unlike the stimulation of tumor immunity, IL-27 is often suggested as a therapeutic option in autoimmune diseases because it has anti-inflammatory effects.
In vivo studies of autoimmune encephalomyelitis showed that the introduction of IL-27 affected T helper 17 cells and suppressed autoinflammation [24]. In the review by Meka et al., IL-27 is described as one of the most important cytokines in autoimmune diseases. In rheumatoid arthritis, IL-27 is often found in the synovial tissue from affected joints. injections of IL-27 cause disease improvement when injected in a murine model of rheumatoid arthritis. Overall, a similar picture is seen for other autoimmune pathologies, i.e., systemic lupus erythematosus, colitis, psoriasis, and diabetes mellitus.
As mentioned above, IL-27 can play an ambiguous role in terms of inflammatory reactions. It is known to augment antigen-specific generation of cytotoxic lymphocytes [25], specifically via activation of T-bet (T-box transcription factor) and EOMES (Eomesodermin/Tbr2). In addition, IL-27 plays a key role in the generation of Th1 immune responses. IL-27 is produced by the antigen-presenting cells upon activation and promotes the rapid proliferation of CD4+ T cells that have previously encountered the antigen. It also works together with IL-12 to induce IFN-gamma production by CD4+T cells. However, IL-27 does not have the same effect on memory CD4+ T cells [27]. Lucas et al. showed that IL-27 is insufficient to stimulate full-scale IFN production by the innate cells, but it plays a contributing role to Th1 responses [28]. At the same time, IL-27 suppresses Th2 cell differentiation by blocking the expression of GATA-3, a transcription factor deeply involved in Th2 responses [29]. In the study conducted by Schneider R et al., it was demonstrated that IL-27 presence empowered CD8+ T cell proliferation and their development into Tc1 cells [29]; IL-27 promotes the expansion of self-renewing CD8+ T cells [30].
Overall, IL-27 interacts with cells via the IL-27 receptor and activates or inhibits their activity through signaling molecules STAT1/3.
3. Interleukin 27 in Infectious Pathology
Due to the abovementioned diversity of IL-27 effects, its role in infectious diseases is often a matter of debate. It launches pro-inflammatory and protective effects by stimulating Th1-mediated immune responses; at the same time, it elicits regulatory effects by causing the expression of co-inhibitory receptors, including PD-L1, LAG-3, TIM-3, CTLA-4, and TIGIT on T cells [31]. During infections, IL-27 levels are commonly increased and have been associated with the generation of IL-10, which could create a mechanism for controlling inflammation while still allowing the infectious agent to thrive [32].
In bacterial infections, especially those that are often followed by sepsis, IL-27 is usually elevated in the blood plasma [33]. The same tendency was noted in tuberculosis: IL-27 showed elevated concentrations in the plasma of patients with tuberculosis when compared to healthy donors [34]. In one of the studies, IL-27 was named ‘a wolf in sheep’s clothing’, due to the fact that in its absence, better control of mycobacterium growth was noted, but at the same time, its absence resulted in chronic hyperinflammation in later stages [35]. In another study on M. Tuberculosis, it was shown that IL-27 inhibits the anti-mycobacterial activity of macrophages [36]. In respiratory chlamydia infections, IL-27 is known to stimulate Th1-mediated inflammatory responses [37].
In viral infections, for instance, influenza, IL-27 plays an ambiguous role. In murine experiments conducted by Liu et al., IL-27 was found to limit immunopathology, neutrophil accumulation, and dampened Th1 or Th17 responses via IL-10-dependent and -independent pathways. And in the absence of IL-27 signals, a more severe disease course was observed [38]. The results indicate that IL-27 plays a crucial role in regulating immunopathology and that administering IL-27 at appropriate times could be an effective treatment for severe inflammation caused by infections.
At the same time, in parasitic infections, Th1 responses mediated by IL-27 signaling are directed towards the limitation of inflammation [39].
4. Interleukin 27 in COVID-19
As shown in other pathologies, IL-27 demonstrates a diversity of functions. We believe IL-27 to be a potential key player in the development of COVID-19 and protection against it. There are several studies concerning the involvement of IL-27 in COVID-associated immunity. IL-27 is a protein that is produced by immune cells when they are activated by pathogens or TLR ligands. It can stimulate the release of inflammatory cytokines from various types of cells, including mast cells, monocytes, and keratinocytes [40].
Initially, our investigation was aimed at analyzing the role of this cytokine in COVID-19 patients. Upon comparing acute-phase COVID-19 patients with healthy donors, we observed a statistically significant increase in IL-27 plasma levels in acute COVID-19 patients. Conversely, we also noted an opposite trend in convalescent patients, where concentrations of IL-27 were lower than those in healthy donors [41]. This tendency was seen independently of disease severity in the first-wave COVID patients in early 2020.
In a study performed by Zamani et al., IL-27 was proposed as a potential predictor for the severity and outcome of COVID-19 [42], along with IL-32 and neutrophil-to-lymphocyte ratio (NLR). IL-27 was identified as a factor indicating the necessity for hospitalization in patients who have a higher risk due to demographic factors such as being male, of black or Hispanic ethnicity, and having a median age over 63 years [43]. Another research used machine learning to predict the outcome of COVID-19 and identified a few cytokines, including IL-27, IL-9, IL-12p40, and MCP-3, as potential indicators for mild cases of the disease [49]. The 2023 publication authored by Valdés-López and Urcuqui-Inchima revealed that the identification of mRNA encoding IL-27 subunits in PBMCs and monocytes from individuals with COVID-19 was linked to the severity of the disease [45]. Moreover, mRNA expression levels of IL-27 signaling components in COVID-derived PBMCs showed positive correlations between the expression level of mRNA encoding the IL-27p28 subunit and IL-27 signaling components in COVID-derived PBMCs, including Gp130, JAK2, STAT1, STAT3, and SOCS3. In addition to our previous studies on the role of cytokines in COVID-19, we conducted another investigation to explore the association between Interleukin 27 (IL-27) concentrations and disease outcome. Our research showed that there was a significant increase in concentrations among both COVID-19 survivors and non-survivors when compared to healthy individuals. However, there was no statistical significance found when comparing the survivors and non-survivors [46]. And while it is possible that IL-27 may have predictive value, our analysis showed no sufficient sensitivity or specificity to support this claim. We used ROC analysis to assess the predictive value of IL-27 and found that it was not a reliable marker for prognosis.
In studies concerning other respiratory pathologies, IL-27 has shown a positive correlation with the severity of pulmonary inflammation [47]. In patients with community-associated pneumonia (CAP), serum levels of IL-27 were elevated in comparison with healthy donors [48].
However, as we delved further into the investigation of IL-27 and its involvement in COVID-19, we also analyzed the correlation between IL-27 and the SARS-CoV-2 genetic variant. In one of our studies, we performed multiplex cytokine profiling in plasma samples from patients infected with different SARS-CoV-2 variants (i.e., 2019-nCoV strain or Wuhan strain, B.1.1.7 or Alpha variant, B.1.617.2 or Delta variant, and B.1.1.529 or Omicron variant) [49]. Our results suggest that IL-27 plasma levels are not a reliable indicator of the severity of COVID-19 infection, as they do consistently vary with the viral variant. However, the four cytokines (IL-6, IL-10, IL-18, and IL-27) that were consistently elevated in COVID-19 patients may provide valuable insights into the equilibrium behind disease progression and host infection. Further research is needed to confirm these findings and determine their clinical significance.
When analyzing CD8+ T cell subpopulations in acute COVID-19 patients, convalescents, and healthy donors, we noted a negative correlation with the CCR6+CD8+ T cell frequencies within effector memory T cells and terminally differentiated effector memory T cells re-expressing CD45RA in patients with acute COVID-19 [50]. This tendency was not evident in COVID-19 convalescents or healthy controls. Moreover, serum IL-27 levels also showed a negative correlation with the absolute numbers of central memory and terminally differentiated effector memory cells (Tc17 cells) from patients with acute COVID-19 but not from convalescent or healthy donors. In the study by Schneider R et al., IL-27 presence was affecting CD8+ T cell proliferation and inducing their development into Tc1 cells [51]. In viral infections, it was highlighted that IL-27 promotes the expansion of self-renewing CD8+ T cells [52].
All findings concerning IL-27 in COVID-19 are presented in Table 1.
IL-27, among other cytokines, is deeply involved in immune-mediated processes associated with COVID-19. As its receptor is widely present on a variety of cells, its function is strongly interconnected with other cytokines and chemokines. We present our view on how IL-27 is integrated into immune processes in COVID-19 in Figure 2.
5. Conclusions
Interleukin 27 is a cytokine endowed with diverse functional activities. It can play both pro- and anti-inflammatory roles, depending on the signaling pathway and the target cells it affects. Overall, regardless of the pathological process it is involved in, IL-27 is critical for upholding the necessary balance between tissue damage and cytotoxicity against infectious agents and/or tumors.
In COVID-19, its role is also ambiguous. Being one of the few cytokines that show statistically significant changes in concentrations in the acute phase of the infection, IL-27 is directly linked to the markers of cell-mediated immunity. And even though its full role in the immune responses behind COVID-19 is yet to be explored, it is now clear that IL-27 may be one of the most interesting cytokines involved in this infection.
6. Future Directions
We are open to discussing this topic further and welcome any insights or perspectives that may help us better understand the role of IL-27 and other cytokines in COVID-19. By gaining a better understanding of the immune response to SARS-CoV-2, we hope to identify new therapeutic targets and develop more effective treatments for this devastating disease.
Author Contributions
Z.R.K. – Investigation, Formal Analysis, Visualization, Writing – original draft; N.A.A. –Investigaion, Methodology, Resources; A.S. – Conceptualization, Writing – review & editing; A.A.T – Conceptualization, Project administration, Visualization, Writing – review & editing.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable.
Acknowledgments
We thank the staff of the core facility centre, ‘Cytometry and biomarkers’ in Pasteur Institute, Russia, for helping with experimental parts of our work.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Filip, Roxana, Roxana Gheorghita Puscaselu, Liliana Anchidin-Norocel, Mihai Dimian, and Wesley K. Savage. 2022. “Global Challenges to Public Health Care Systems during the COVID-19 Pandemic: A Review of Pandemic Measures and Problems.” Journal of Personalized Medicine 12 (8): 1295. [CrossRef]
- Salamanna, Francesca, Melania Maglio, Maria Paola Landini, and Milena Fini. 2020. “Body Localization of ACE-2: On the Trail of the Keyhole of SARS-CoV-2.” Frontiers in Medicine 7 (December). [CrossRef]
- Boechat, José Laerte, Inês Chora, António Morais, and Luís Delgado. 2021. “The Immune Response to SARS-CoV-2 and COVID-19 Immunopathology – Current Perspectives.” Pulmonology 27 (5). [CrossRef]
- Lu, Ligong, Hui Zhang, Meixiao Zhan, Jun Jiang, Hua Yin, Danielle J Dauphars, Shiyou Li, Yong Li, and You-Wen He. 2020. “Preventing Mortality in COVID-19 Patients: Which Cytokine to Target in a Raging Storm?” Frontiers in Cell and Developmental Biology 8 (July). [CrossRef]
- Gibson, Peter G, Ling Qin, and Ser Hon Puah. 2020. “COVID -19 Acute Respiratory Distress Syndrome ( ARDS ): Clinical Features and Differences from Typical Pre- COVID -19 ARDS.” Medical Journal of Australia 213 (2). [CrossRef]
- Kastelein, Robert A., Christopher A. Hunter, and Daniel J. Cua. 2007. “Discovery and Biology of IL-23 and IL-27: Related but Functionally Distinct Regulators of Inflammation.” Annual Review of Immunology 25 (1): 221–42. [CrossRef]
- Devergne, O, M Hummel, H Koeppen, M M Le Beau, E C Nathanson, E Kieff, and M Birkenbach. 1996. “A Novel Interleukin-12 P40-Related Protein Induced by Latent Epstein-Barr Virus Infection in B Lymphocytes.” Journal of Virology 70 (2): 1143–53. [CrossRef]
- Pflanz, Stefan, Jackie C Timans, Jeanne Cheung, Rency Rosales, Holger Kanzler, Jonathan Gilbert, Linda Hibbert, et al. 2002. “IL-27, a Heterodimeric Cytokine Composed of EBI3 and P28 Protein, Induces Proliferation of Naive CD4+ T Cells.” Immunity 16 (6): 779–90. [CrossRef]
- Mei, Youwen, Zi Lv, Liling Xiong, Hanwen Zhang, Nanlin Yin, and Hongbo Qi. 2021. “The Dual Role of IL-27 in CD4+T Cells.” Molecular Immunology 138 (October): 172–80. [CrossRef]
- Min, Booki, Dongkyun Kim, and Matthias J Feige. 2021. “IL-30† (IL-27A): A Familiar Stranger in Immunity, Inflammation, and Cancer.” Experimental & Molecular Medicine 53 (5): 823–34. [CrossRef]
- Kishi, Yasuhiro, Takaaki Kondo, Sheng Xiao, Nir Yosef, Jellert Gaublomme, Chuan Wu, Chao Wang, et al. 2016. “Protein c Receptor (PROCR) Is a Negative Regulator of Th17 Pathogenicity.” Journal of Experimental Medicine 213 (11): 2489–2501. [CrossRef]
- Marques, Hanna Santos, Breno Bittencourt de Brito, Filipe Antônio França da Silva, Maria Luísa Cordeiro Santos, Júlio César Braga de Souza, Thiago Macêdo Lopes Correia, Luana Weber Lopes, et al. 2021. “Relationship between Th17 Immune Response and Cancer.” World Journal of Clinical Oncology 12 (10): 845–67. [CrossRef]
- Mchedlidze, T, M Kindermann, A T Neves, D Voehringer, M F Neurath, and S Wirtz. 2016. “IL-27 Suppresses Type 2 Immune Responses in Vivo via Direct Effects on Group 2 Innate Lymphoid Cells.” Mucosal Immunology 9 (6): 1384–94. [CrossRef]
- Do, Jeongsu, Dongkyun Kim, Sohee Kim, Alice Valentin-Torres, Nina Dvorina, Eunjung Jang, Vivekananthan Nagarajavel, et al. 2017. “Treg-Specific IL-27Rα Deletion Uncovers a Key Role for IL-27 in Treg Function to Control Autoimmunity.” Proceedings of the National Academy of Sciences of the United States of America 114 (38): 10190–95. [CrossRef]
- Pyle, Chloe J., Faith I. Uwadiae, David P. Swieboda, and James A. Harker. 2017. “Early IL-6 Signalling Promotes IL-27 Dependent Maturation of Regulatory T Cells in the Lungs and Resolution of Viral Immunopathology.” Edited by Carolina B. Lopez. PLOS Pathogens 13 (9): e1006640. [CrossRef]
- Kourko, Olena, Kyle Seaver, Natalya Odoardi, Sameh Basta, and Katrina Gee. 2019. “IL-27, IL-30, and IL-35: A Cytokine Triumvirate in Cancer.” Frontiers in Oncology 9 (October). [CrossRef]
- Morita, Yugo, Elysia A. Masters, Edward M. Schwarz, and Gowrishankar Muthukrishnan. 2021. “Interleukin-27 and Its Diverse Effects on Bacterial Infections.” Frontiers in Immunology 12 (May). [CrossRef]
- Shimizu, Jun, Fumio Kaneko, and Noboru Suzuki. 2013. “Skewed Helper T-Cell Responses to IL-12 Family Cytokines Produced by Antigen-Presenting Cells and the Genetic Background in Behcet’s Disease.” Genetics Research International 2013 (December): 1–11. [CrossRef]
- Zwirner, Norberto Walter, and Andrea Ziblat. 2017. “Regulation of NK Cell Activation and Effector Functions by the IL-12 Family of Cytokines: The Case of IL-27.” Frontiers in Immunology 8 (January). [CrossRef]
- Yoshimoto, Takayuki, Yukino Chiba, Jun-Ichi Furusawa, Mingli Xu, Ren Tsunoda, Kaname Higuchi, and Izuru Mizoguchi. 2015. “Potential Clinical Application of Interleukin-27 as an Antitumor Agent.” Cancer Science 106 (9): 1103–10. [CrossRef]
- Kourko, Olena, Kyle Seaver, Natalya Odoardi, Sameh Basta, and Katrina Gee. 2019. “IL-27, IL-30, and IL-35: A Cytokine Triumvirate in Cancer.” Frontiers in Oncology 9 (October). [CrossRef]
- Jia, Haiyan, Paula Dilger, Chris Bird, and Meenu Wadhwa. 2016. “IL-27 Promotes Proliferation of Human Leukemic Cell Lines through the MAPK/ERK Signaling Pathway and Suppresses Sensitivity to Chemotherapeutic Drugs.” Journal of Interferon & Cytokine Research 36 (5): 302–16. [CrossRef]
- Sekar, Divya, Christina Hahn, Bernhard Brüne, Edward Roberts, and Andreas Weigert. 2012. “Apoptotic Tumor Cells Induce IL-27 Release from Human DCS to Activate TReg Cells That Express CD69 and Attenuate Cytotoxicity.” European Journal of Immunology 42 (6): 1585–98. [CrossRef]
- Batten, Marcel, Ji Li, Sothy Yi, Noelyn M. Kljavin, Dimitry M. Danilenko, Sophie Lucas, James Lee, Frederic J. de Sauvage, and Nico Ghilardi. 2006. “Interleukin 27 Limits Autoimmune Encephalomyelitis by Suppressing the Development of Interleukin 17–Producing T Cells.” Nature Immunology 7 (9): 929–36. [CrossRef]
- Morishima, Naohiko, Itaru Mizoguchi, Masae Okumura, Yukino Chiba, Mingli Xu, Motomu Shimizu, Masanao Matsui, Junichiro Mizuguchi, and Takayuki Yoshimoto. 2010. “A Pivotal Role for Interleukin-27 in CD8+T Cell Functions and Generation of Cytotoxic T Lymphocytes.” Journal of Biomedicine and Biotechnology 2010 (January): 1–10. [CrossRef]
- Pflanz, Stefan, Jackie C Timans, Jeanne Cheung, Rency Rosales, Holger Kanzler, Jonathan Gilbert, Linda Hibbert, et al. 2002. “IL-27, a Heterodimeric Cytokine Composed of EBI3 and P28 Protein, Induces Proliferation of Naive CD4+ T Cells.” Immunity 16 (6): 779–90. [CrossRef]
- Lucas, Sophie, Nico Ghilardi, Ji Li, and Frédéric J. de Sauvage. 2003. “IL-27 Regulates IL-12 Responsiveness of Naïve CD4+ T Cells through Stat1-Dependent and -Independent Mechanisms.” Proceedings of the National Academy of Sciences of the United States of America 100 (25): 15047–52. [CrossRef]
- Yoshimoto, Tomohiro, Takayuki Yoshimoto, Koubun Yasuda, Junichiro Mizuguchi, and Kenji Nakanishi. 2007. “IL-27 Suppresses Th2 Cell Development and Th2 Cytokines Production from Polarized Th2 Cells: A Novel Therapeutic Way for Th2-Mediated Allergic Inflammation.” The Journal of Immunology 179 (7): 4415–23. [CrossRef]
- Schneider, Raphael, Teodora Yaneva, Diane Beauseigle, Lama El-Khoury, and Nathalie Arbour. 2010. “IL-27 Increases the Proliferation and Effector Functions of Human Naïve CD8+ T Lymphocytes and Promotes Their Development into Tc1 Cells.” European Journal of Immunology 41 (1): 47–59. [CrossRef]
- Harker, James A., Aleksandr Dolgoter, and Elina I. Zuniga. 2013. “Cell-Intrinsic IL-27 and Gp130 Cytokine Receptor Signaling Regulates Virus-Specific CD4+ T Cell Responses and Viral Control during Chronic Infection.” Immunity 39 (3): 548–59. [CrossRef]
- Meka, Rakeshchandra R., Shivaprasad H. Venkatesha, Steven Dudics, Bodhraj Acharya, and Kamal D. Moudgil. 2015. “IL-27-Induced Modulation of Autoimmunity and Its Therapeutic Potential.” Autoimmunity Reviews 14 (12): 1131–41. [CrossRef]
- Do, Jeong-su, Anabelle Visperas, Yibayiri Osee Sanogo, Jennifer J Bechtel, Nina Dvorina, Sohee Kim, Eunjung Jang, et al. 2016. “An IL-27/Lag3 Axis Enhances Foxp3+ Regulatory T Cell–Suppressive Function and Therapeutic Efficacy.” Mucosal Immunology 9 (1): 137–45. [CrossRef]
- Wong, Hector R., Christopher J. Lindsell, Patrick Lahni, Kimberly W. Hart, and Sebastien Gibot. 2013. “Interleukin 27 as a Sepsis Diagnostic Biomarker in Critically Ill Adults.” Shock 40 (5): 382–86. [CrossRef]
- Skouras, Vasileios S, Sophia F Magkouta, Ioannis Psallidas, Irene Tsilioni, Panagiotis Maragozidis, Konstantinos I Gourgoulianis, and Ioannis Kalomenidis. 2015. “Interleukin-27 Improves the Ability of Adenosine Deaminase to Rule out Tuberculous Pleural Effusion regardless of Pleural Tuberculosis Prevalence.” Infectious Diseases 47 (7): 477–83. [CrossRef]
- Gollnick, Hailey, Jamie Barber, Robert J Wilkinson, Sandra Newton, and Ankita Garg. 2023. “IL-27 Inhibits Anti- Mycobacterium Tuberculosis Innate Immune Activity of Primary Human Macrophages.” Tuberculosis 139 (March): 102326–26. [CrossRef]
- Zeng, Jiajia, Shuaini Yang, Yuqing Tuo, Xiaoyu Zha, Ruoyuan Sun, Tingsha Lu, Hong Zhang, Lu Tan, Sai Qiao, and Hong Bai. 2023. “IL-27 Signaling Promotes Th1 Response by Downregulating IL-10 Production in DCs during Chlamydial Respiratory Infection.” Microorganisms 11 (3): 604–4. [CrossRef]
- Liu, Francesca Diane M., Elisabeth E. Kenngott, Micha F. Schröter, Anja Kühl, Silke Jennrich, Ralf Watzlawick, Ute Hoffmann, et al. 2014. “Timed Action of IL-27 Protects from Immunopathology While Preserving Defense in Influenza.” PLoS Pathogens 10 (5): e1004110. [CrossRef]
- Montes de Oca, Marcela, Fabian de Labastida Rivera, Clay Winterford, Teija C. M. Frame, Susanna S. Ng, Fiona H. Amante, Chelsea L. Edwards, et al. 2020. “IL-27 Signalling Regulates Glycolysis in Th1 Cells to Limit Immunopathology during Infection.” Edited by Phillip Scott. PLOS Pathogens 16 (10): e1008994. [CrossRef]
- Ansari, Nasim Akhtar, Rajiv Kumar, Shalini Gautam, Susanne Nylén, Om Prakash Singh, Shyam Sundar, and David Sacks. 2011. “IL-27 and IL-21 Are Associated with T Cell IL-10 Responses in Human Visceral Leishmaniasis.” The Journal of Immunology 186 (7): 3977–85. [CrossRef]
- Arsentieva, N A, N E Liubimova, O K Batsunov, Z R Korobova, O V Stanevich, A A Lebedeva, E A Vorobyov, et al. 2021. “Plasma Cytokines in Patients with COVID-19 during Acute Phase of the Disease and Following Complete Recovery.” Медицинская иммунoлoгия 23 (2): 311–26. [CrossRef]
- Zamani, Batool, Maedeh Najafizadeh, Hossein Motedayyen, and Reza Arefnezhad. 2022. “Predicting Roles of IL-27 and IL-32 in Determining the Severity and Outcome of COVID-19.” International Journal of Immunopathology and Pharmacology 36 (January): 039463202211458-039463202211458. [CrossRef]
- Klingler, Jéromine, Gregory Lambert, Juan C Bandres, Rozita Emami-Gorizi, Arthur Nádas, Kasopefoluwa Y Oguntuyo, Fatima Amanat, et al. 2022. “Immune Profiles to Distinguish Hospitalized versus Ambulatory COVID-19 Cases in Older Patients” 25 (12): 105608–8. [CrossRef]
- Laatifi, Mariam, Samira Douzi, Hind Ezzine, Chadia El Asry, Abdellah Naya, Abdelaziz Bouklouze, Younes Zaid, and Mariam Naciri. 2023. “Explanatory Predictive Model for COVID-19 Severity Risk Employing Machine Learning, Shapley Addition, and LIME.” Scientific Reports 13 (1). [CrossRef]
- Juan Felipe Valdés-López, and Silvio Urcuqui-Inchima. 2023. “Antiviral Response and Immunopathogenesis of Interleukin 27 in COVID-19.” Archives of Virology 168 (7). [CrossRef]
- Valdés-López, J.F., Urcuqui-Inchima, S. Antiviral response and immunopathogenesis of Interleukin 27 in COVID-19. Arch Virol 168, 178 (2023). [CrossRef]
- Arsentieva, Natalia A, Natalia E Liubimova, Oleg K Batsunov, Zoia R Korobova, Raisa N Kuznetsova, Artem A Rubinstein, Oksana V Stanevich, et al. 2023. “Predictive Value of Specific Cytokines for Lethal COVID-19 Outcome.” Infekciâ I Immunitet 12 (5): 859–68. [CrossRef]
- Xu, Fang, Qiong Liu, Shihui Lin, Na Shen, Yibing Yin, and Ju Cao. 2013. “IL-27 Is Elevated in Acute Lung Injury and Mediates Inflammation.” Journal of Clinical Immunology 33 (7): 1257–68. [CrossRef]
- Xu, Zheng, Xin-Ming Wang, Peng Cao, Chen Zhang, Chun-Mei Feng, Ling Zheng, De-Xiang Xu, Lin Fu, and Hui Zhao. 2022. “Serum IL-27 Predicts the Severity and Prognosis in Patients with Community-Acquired Pneumonia: A Prospective Cohort Study.” International Journal of Medical Sciences 19 (1): 74–81. [CrossRef]
- Korobova, Zoia R., Natalia A. Arsentieva, Natalia E. Liubimova, Oleg K. Batsunov, Vladimir G. Dedkov, Anna S. Gladkikh, Alena A. Sharova, et al. 2022. “Cytokine Profiling in Different SARS-CoV-2 Genetic Variants.” International Journal of Molecular Sciences 23 (22): 14146. [CrossRef]
- Kudryavtsev, Igor V., Natalia A. Arsentieva, Zoia R. Korobova, Dmitry V. Isakov, Artem A. Rubinstein, Oleg K. Batsunov, Irina V. Khamitova, Raisa N. Kuznetsova, Tikhon V. Savin, Tatiana V. Akisheva, and et al. 2022. "Heterogenous CD8+ T Cell Maturation and ‘Polarization’ in Acute and Convalescent COVID-19 Patients" Viruses 14, no. 9: 1906. [CrossRef]
- Schneider, Raphael, Teodora Yaneva, Diane Beauseigle, Lama El-Khoury, and Nathalie Arbour. 2010. “IL-27 Increases the Proliferation and Effector Functions of Human Naïve CD8+ T Lymphocytes and Promotes Their Development into Tc1 Cells.” European Journal of Immunology 41 (1): 47–59. [CrossRef]
- Harker, James A., Aleksandr Dolgoter, and Elina I. Zuniga. 2013. “Cell-Intrinsic IL-27 and Gp130 Cytokine Receptor Signaling Regulates Virus-Specific CD4+ T Cell Responses and Viral Control during Chronic Infection.” Immunity 39 (3): 548–59. [CrossRef]
Figure 1.
Simplified role of Interleukin 27 in immune responses. Produced by dendritic cells, this cytokine serves multiple purposes as it affects IL-27 receptor on CD4+, CD8+, B and NK cells.
Figure 1.
Simplified role of Interleukin 27 in immune responses. Produced by dendritic cells, this cytokine serves multiple purposes as it affects IL-27 receptor on CD4+, CD8+, B and NK cells.
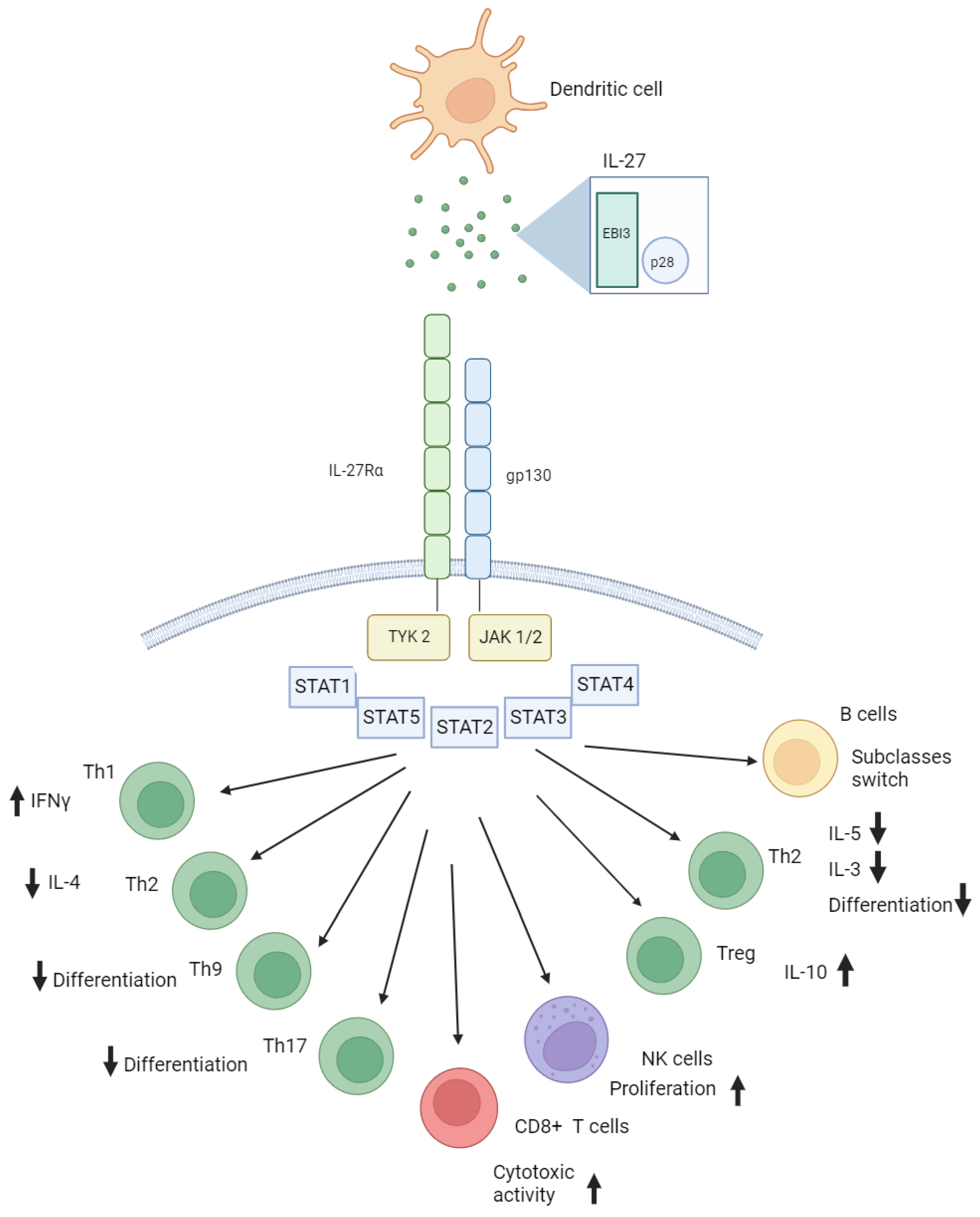
Figure 2.
Simplified representation of immune responses to SARS-CoV-2 and the outcomes associated with them. A – SARS-CoV-2 interacts with innate immune cells (e.g., macrophage) via TLR4 receptor. Subsequent signaling leads to production of cytokines and chemokines. B – Immune cell produce pro-inflammatory cytokines, IFNs and chemokines that attract other immune cells. C – IFN γ also stimulates production of IFN-dependent cytokines and chemokines, such as CXCL10/IP-10. In its presence, additionally to the influence of viral SARS-CoV-2 peptides, production of MDC/CCL22 by macrophages decreases, which leads to depletion in concentrations of this chemokine noted in our studies earlier. D - Upon cytokine production, Th0 differentiation branches out depending on cytokines involved, including Th1-mediated immune responses with further production of IFN γ and involvement of macrophages. E - Involvement of cytotoxic lymphocytes (CTLs) in immune responses is also mediated by IL-27. It leads to cytotoxicity towards infected cells. F – In germinal center, via activation of Th2 immune responses and IL-4 production, B cells differentiate into effector plasma cells or memory B cells. G - Plasma cells produce antigen-specific antibodies, whereas IL-27 is involved in subclasses switch. H – anti-inflammatory effects of IL-27 are affecting differentiation of T regulatory cells. IL-27 is also involved in switching Th2, Th17 immune responses towards anti-viral Th1 immunity (B-E).
Figure 2.
Simplified representation of immune responses to SARS-CoV-2 and the outcomes associated with them. A – SARS-CoV-2 interacts with innate immune cells (e.g., macrophage) via TLR4 receptor. Subsequent signaling leads to production of cytokines and chemokines. B – Immune cell produce pro-inflammatory cytokines, IFNs and chemokines that attract other immune cells. C – IFN γ also stimulates production of IFN-dependent cytokines and chemokines, such as CXCL10/IP-10. In its presence, additionally to the influence of viral SARS-CoV-2 peptides, production of MDC/CCL22 by macrophages decreases, which leads to depletion in concentrations of this chemokine noted in our studies earlier. D - Upon cytokine production, Th0 differentiation branches out depending on cytokines involved, including Th1-mediated immune responses with further production of IFN γ and involvement of macrophages. E - Involvement of cytotoxic lymphocytes (CTLs) in immune responses is also mediated by IL-27. It leads to cytotoxicity towards infected cells. F – In germinal center, via activation of Th2 immune responses and IL-4 production, B cells differentiate into effector plasma cells or memory B cells. G - Plasma cells produce antigen-specific antibodies, whereas IL-27 is involved in subclasses switch. H – anti-inflammatory effects of IL-27 are affecting differentiation of T regulatory cells. IL-27 is also involved in switching Th2, Th17 immune responses towards anti-viral Th1 immunity (B-E).
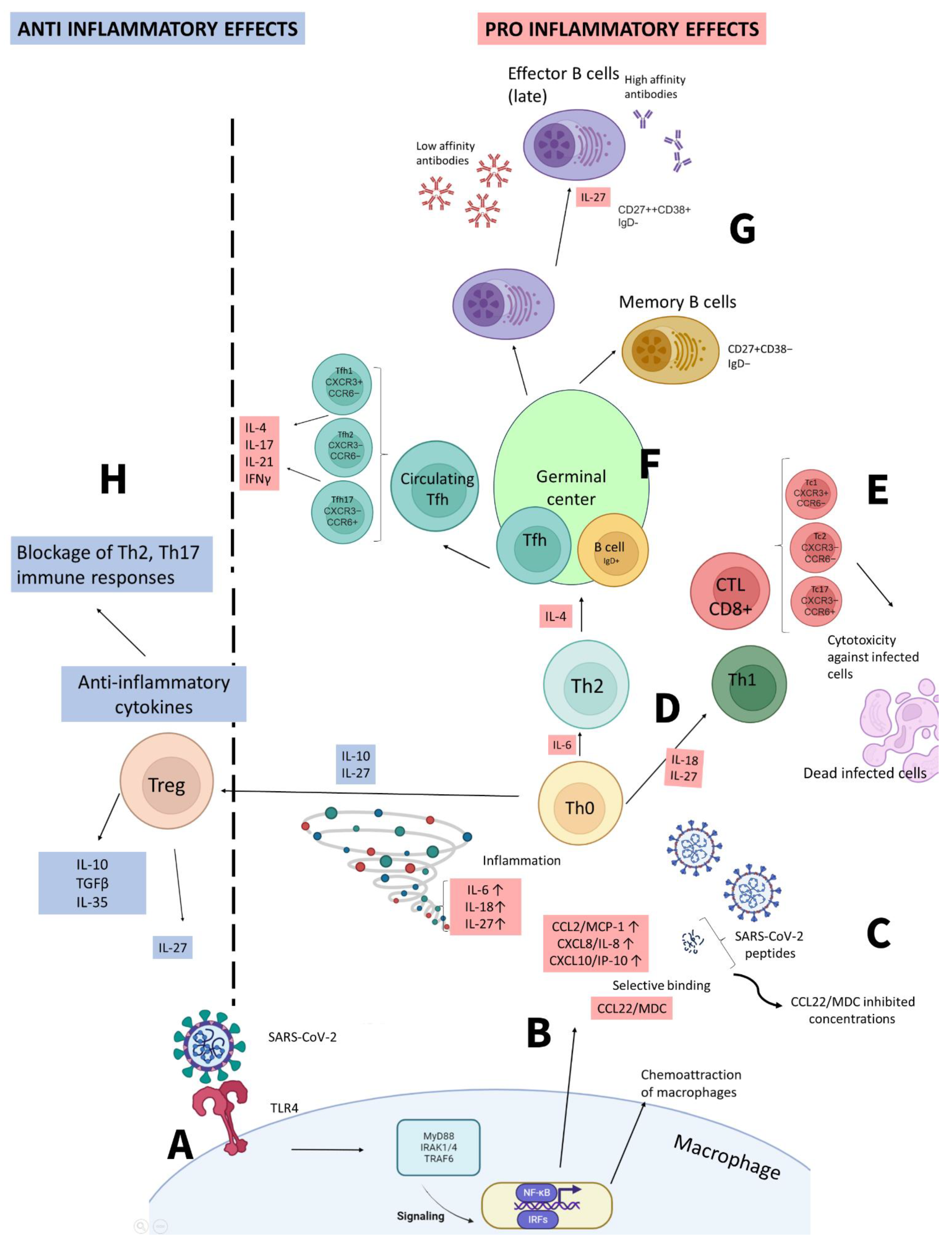

Table 1.
Research overview on the role of IL-27 in COVID-19.
| Year of publication | Authors | Findings | Reference |
|---|---|---|---|
| 2021 | Arsentieva N.A. et al. | Increase in IL-27 levels in acute COVID-19 patients vs. healthy donors (p<0.0001); Decrease of IL-27 concentrations in convalescents when compared to healthy donors (p=0.0015). | [41] |
| 2022 | Arsentieva N.A. et al. | IL-27 showed statistically significant increase in concentrations in COVID-19 acute patients when compared to healthy donors (p<0.001 for non-survivors and p<0.05 for survivors). | [46] |
| 2022 | Korobova Z.R. et al. | IL-27 was one of the four biological markers showing statistically significant changes in concentrations in blood plasma of patients infected with different variants of SARS-CoV-2 (p<0.001). | [49] |
| 2022 | Kudryavtsev I.V. et al. | The correlation between Tc17 cells of central memory and TEMRA cells and serum IL-27 levels was negative in patients with acute COVID-19, a tendency not observed in convalescent or healthy donors. | [50] |
| 2022 | Zamani B. et al. | IL-27, along with IL-32 and neutrophil-to-lymphocyte ratio (NLR) was highlighted as one of the markers of severe COVID-19 and lethal outcomes. | [42] |
| 2022 | Klingler J et al. | In patients with higher demographic risk factors (i.e., male, black/Hispanic descent and median age over 63 years old), IL-27 was suggested as one of the factors to prove the need for hospitalization | [43] |
| 2023 | Laatifi M. et al. | COVID-19 prognosis was based on machine learning and included several cytokines, and IL-27, along with IL-9, IL-12p40, and MCP-3, to play the role of markers for non-severe COVID-19 | [44] |
| 2023 | Valdés-López and Urcuqui-Inchima | IL-27 triggers a strong pro-inflammatory and antiviral reaction that relies on STAT1, without requiring IFN, in COVID-19-derived PBMCs and monocytes, which is linked to a severe clinical outcome of COVID-19. This effect is also seen in macrophages that have been stimulated by S protein. | [45] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



