
Submitted:
22 May 2024
Posted:
23 May 2024
You are already at the latest version
Abstract
(1) Background: Double-strand breaks (DSBs) in a single nucleus are usually measured using the sperm chromatin structure assay (SCSA), sperm chromatin dispersion (SCD) test, and comet assay (CA). We developed mono-dimensional single-cell pulsed-field gel electrophoresis (1D-SCPFGE) and angle-modulated two-dimensional single-cell pulsed-field gel electrophoresis (2D-SCPFGE) to observe DNA fragmentation in separated motile sperm. (2) Methods: We set up comparative standards, calibration curves, the required sensitivity, and eligibility criteria for test sperm to validate their measurement principles. (3) Results: We revealed that the conventional methods overlooked the interference of nucleoproteins in their measurements. In-gel proteolysis improves the measurement accuracies of 1D- and 2D-SCPFGE. We recommend using naked DNA for comparative standards and test specimens. Moreover, we have observed several dysfunctions that may induce DNA damage in the separated motile sperm. Overall, our discussion highlights the need to revisit the conventional univariable analyses based on the SCSA, SCD test, and CA.
(4) Conclusions: Human infertility is a complex syndrome, and the aim of quality control in intracytoplasmic sperm injection is to identify the underlying dysfunctions remaining in the separated motile sperm that render them ineligible for injection. Well-designed, multivariable analyses with special consideration to confounding factors are necessary in future cohort studies.
Keywords:
DNA fragmentation
; single-cell pulsed-field gel electrophoresis
; human sperm
; sperm chromatin structure assay
; sperm chromatin dispersion test
; comet assay
; quantitative analysis
Overview
The three major DNA-related contributors to human male infertility are de novo mutations, a decline in DNA repair capacity, and DNA fragmentation. Each of us is born with a few novel genetic changes, known as de novo mutations, that occur either during gamete formation or postzygotically [1]. Pathogenic de novo mutations that cause severe congenital anomalies are mostly of paternal origin [2], and they play a prominent role in severe male infertility [3]. Apoptosis regulates spermatogenesis in the testis during the spermatogonium, spermatocyte, and spermatid phases [4]. More than half of differentiating spermatogenic cells undergo apoptosis before maturing into sperm and are selectively and rapidly eliminated through phagocytosis by Sertoli cells [5]. Mature sperm lack DNA repair mechanisms and depend on maternal factors for repair after fertilization [6]. Owing to the absence of homologous templates, zygotic repair of paternal double-strand breaks (DSBs) depends on non-homologous repair mechanisms that are considered to be error-prone. Moreover, apoptosis degenerates the sperm awaiting ejaculation in the epididymis [4,5]. DNA fragmentation is the most characteristic feature of the apoptotic process; upon activation, Ca/Mg-dependent endonucleases cleave DNA strands into nucleosomal subunits of ~185 base pairs in length [7]. Although intracytoplasmic sperm injection (ICSI) is a major fertilization procedure in assisted reproductive technology (ART), many cohort studies have revealed DNA fragmentation in human sperm nuclei as a major risk factor not only for early post-implantation embryo development [8] but also for sperm-derived congenital anomalies in ART [9].
For the past two decades, numerous researchers have discussed the significance of nonspecific single-nuclear DNA damage based on sperm chromatin structure assay (SCSA) [10,11], sperm chromatin dispersion (SCD) test [12,13], and comet assay (CA) [14,15]. We have developed mono-dimensional single-cell pulsed-field gel electrophoresis (1D-SCPFGE) to observe single-nucleus DNA fragmentation [16,17,18]. Even in normozoospermic semen, that method revealed that almost all of the immotile sperm in the ejaculate already exhibit end-stage DNA fragmentation [18,19,20]. Importantly, we discovered that the apparent density of motile sperm with fibrous DNA was 1.12–1.17 g/mL, whereas that of the immotile sperm exhibiting end-stage DNA fragmentation was higher than 1.17 g/mL [18,19,20]. The motile and immotile sperm were easily separated from each other using single-layer density-gradient centrifugation [18,19,20]. These two cell types differed in terms of not only the sequentiality of DNA [18,19,20] but also the integrity of the plasma and mitochondrial membranes [20], the status of the acrosome [20], production of endogenous reactive oxygen species (ROS) in the midpiece [20], and antigenicity for anti-sperm antibodies [19]. These differences corresponded to those between cells that had not yet undergone apoptosis and those that had. The difference in apparent density was due to apoptotic volume decrease [21].
As mature sperm lack DNA repair mechanisms [6], the critical threshold of DSBs in the nucleus is very low [22,23]. The incidence of sperm-derived congenital anomalies is not proportional to the number of DSBs, as a number exceeding the threshold results in fertilization failure or pregnancy loss. In conventional genome sequencing, DNA is extracted from a population of cells. Hence, genetic variations unique to individual cells are lost to the population average, and de novo mutations in a single cell are concealed in the bulk signal. Highly advanced technologies, e.g., high-throughput, single-nuclear, whole-genome amplification [24,25], are major candidates for detecting de novo mutations. However, even after the motile and immotile sperm have been separated, some of the motile sperm contain nonspecific DNA damage [16,17,18,19,20].
In the present review, we first introduce analytical methods to observe the early symptoms of DNA fragmentation in the motile sperm fraction, including the establishment of comparative standards, the calibration curve, the required sensitivity, and the eligibility criteria.
Human infertility is a complex syndrome influenced by various dysfunctions besides DNA damage in the separated motile sperm fraction. Therefore, well-designed multivariable analyses with special consideration to confounding factors are necessary to conduct cohort studies. Thereafter, we discuss the design of cohort studies in which the pathology and epidemiology of DNA fragmentation are evaluated.
1. First Step of Qualitative-Method Validation—Establishment of Comparative Standards
We, first, purified the sperm with fibrous DNA and those with granular segments from human semen, some analytical methods require to distinguish these two extremes at the first step qualitative validation. The detailed procedures for the separation of the two types of sperm were described in our previous reports [18,19,20]. Briefly, the diluted semen was fractionated by means of the sedimentation equilibrium in isotonic Optiprep (final apparent density of 1.17 g/mL, hereafter referred to as OP; Axis Shield, San Jose, CA, USA) and differential velocity sedimentation in an isotonic, 90% Percoll (GE Healthcare, Chicago, IL, USA) density gradient.
The sperm from the sediment of the OP/intermediate layer of the Percoll solution were almost immotile and auto-agglutinated, and their apparent density was estimated to exceed 1.17 g/mL. The fraction was termed “denatured sperm” (DS). The sperm recovered from the interface layer of OP/sediment of the Percoll solution was progressively motile, and their apparent density was estimated to be in the range of 1.12–1.17 g/mL. Finally, the motile sperm fraction was prepared using the swim-up method and termed “live sperm” (LS).
2. 1D-SCPFGE
Traditional pulsed-field gel electrophoresis (PFGE) [26,27] is widely used for bacterial typing. Typically, cells are embedded in an agarose plug, proteolyzed, and further digested with restriction endonucleases [28]. Thereafter, DNA segments are separated in macro-gel; and to analyze the banding patterns. We developed micro-PFGE to observe single-nuclear DNA sequentiality without downsizing. The DNA morphology can be directly observed under a microscope using 1D-SCPFGE as described in previous reports [18,19,20].
The in-gel digested LS exserted the tips of DNA fibers outward (Figure 1A). After 1D-SCPFGE, most of the LS exhibited a bundle of elongated long-chain fibers without any segment (Figure 1B). The remainder of the LS exhibited a few to several dozen fibrous segments beyond the anterior end of the elongated fibers. In contrast, in the DS, granular segments were circumferentially dispersed (Figure 1C), and upon 1D-SCPFGE, the segments resembled comet tails (Figure 1D). Thus, according to the 1D-SCPFGE features, we define sperm with a bundle of elongated long-chain fibers without any fibrous segments as intact and those with at least one fibrous fragment as damaged.
As described in Section 4, we also developed a novel angle-modulated two-dimensional single-cell pulsed-field gel electrophoresis (2D-SCPFGE) [29], It revealed some technical issues in 1D-SCPFGE, a fraction of long-chain fibers remained at the electrophoretic origin and long segments were tangled in the bundle of elongated fibers even after the first run. Therefore, the definitions of intact and damaged obtained via 1D-SCPFGE are incomplete. Thus, 1D-SCPFGE can be used to observe mid- or end-stage DNA fragmentation, but not the early stages.
3. Conventional Analytical Methods for DNA Fragmentation do Not Pass the First Step of Qualitative Validation
Comparative studies between LS and DS have revealed various technical problems in the measurement principles of the SCSA [10,11], SCD test [12,13], and CA [14,15]. They do not pass the first step of qualitative validation.
3.1. Lack of Proteolysis Produces False Negative Results in the CA
The CA is used to estimate DNA damage based on the amount of granular segments discharged from the origin, the so-called “comet tail” [14,15]. 1D-SCPFGE visualized the progression of the fragmentation; at first, a few large fibrous segments appear, and cleavage proceeds until all DNA fibers are degraded into granular segments [16,17,18,19,20]. Our observations raised doubts over whether the comet tail was derived from sperm with end-stage fragmentation [16,17,18]. Human sperm nucleoproteins include protamines [30,31], histones [32], condensins [33], and cohesins [33]. The DNA-nucleoprotein complex is fixed to the nuclear membrane with the nuclear matrix [34,35]. The CA extracted nucleoproteins, such as protamines [36], with a high-salt solution [14,15]. In-gel proteolysis of DS dramatically increases the number of granular segments extended via electrophoresis. In a neutral CA, unextracted nucleoproteins remain fixed to the DNA, and only a fraction of the granular segments unfixed were extended as the comet tail [18]. Naked chromosomal DNA fibers in LS are very susceptible to degradation via alkaline hydrolysis; a mere 10 mmol/L NaOH cleaves such fibers into granular segments [18]. Alkaline CA treated DNA with 300 mmol/L NaOH, the binding capacities of the unextracted nucleoproteins tolerate for this radical treatment, they still fixed to DNA, preventing migration of the newly generated granular segments [18]. Therefore, a lack of proteolysis produces false-negative results in both neutral and alkaline CAs. Furthermore, a CA without a pulsed-field current is insufficient to extend the fibrous DNA [14,15].
The enzymatic properties of bovine pancreatic trypsin (EC.3.4.21.4) [37], with respect to substrate specificity and pH dependency, are optimal for the digestion of sperm embedded in agarose. Protamines are arginine-rich, basic proteins in which the guanidyl residue is tightly coupled with phosphoric acid in DNA through electrostatic bonding, and neighboring protamines are cross-linked through disulfide bonds [30,31,36]. Trypsin specifically cleaves the carboxyl end of lysine and arginine residues [37]. In-gel digestion of the embedded cells needs to be initiated after the agarose is completely solidified to avoid free diffusion of the DNA fibers. As the activity of trypsin is strictly dependent on pH, trypsin can be kept inactive at a pH of 4.7 until the gel solidifies, and reactivated by immersing the gel into the cell-lytic reagents (pH = 8). As commercial preparations of trypsin are usually contaminated with pancreatic deoxyribonucleases (DNases), it is not suitable for DNA studies as is. To remove autolyzed trypsin and DNases, twice-crystallized bovine pancreatic trypsin must be further purified via affinity chromatography by using lima bean trypsin inhibitor–conjugated Sephacryl [38]. The purified trypsin should be stored in a solution with a pH < 2.0 to avoid autolysis. Trypsin undoubtedly has a high performance but lacks versatility.
Proteinase K [39] has a chymotrypsin-like broad substrate specificity for aliphatic and aromatic residues of amino acids and acts in a wide pH range; ready-to-use preparations are supplied commercially for the extraction of DNA from somatic cells. However, proteinase K is not very effective against protamines. Competitive dissociation of the protamine-DNA complexes with sodium dodecyl sulfate (SDS) and subsequent digestion of other nucleoproteins with proteinase K allows migration of DNA fibers via 1D-SCPFGE [29].
3.2. Red Fluorescence in the SCSA is Derived from Nucleoproteins but Not from Single-Stranded DNA
The SCSA employs a simple bisection principle wherein the intercalation of monomeric acridine orange (AO) into double-stranded DNA or the adsorption of oligomeric AO to single-stranded DNA produces green or red fluorescence, respectively [10,11]. After AO staining, the nucleus and cytoplasm of lymphocytes yield green and red fluorescence, respectively, and in-gel tryptic digestion of the sperm vanished the red fluorescence [40]. These phenomena raise doubts regarding whether the red fluorescence was truly derived from single-stranded DNA.
The SCSA is routinely performed using flow cytometry [10,11]. The fluorescent profiles of LS and those that underwent radical treatment (96˚C for 20 min) to destroy the DNA structure yielded similar cytograms. DNA and nucleoproteins are packed rigidly in the sperm head, and the combination of the two colors leads to color variation [40]. Prolonged exposure under a fluorescence microscope causes photobleaching of fluorescent dyes and the concurrent photo-breakage of DNA. Under such conditions, sperm nuclei do not undergo simple color quenching but rather a progressive, time-dependent discoloration from red to green, which contradicts the principle of the SCSA [40]. The red fluorescence is derived from AO adsorbing to nucleoproteins rather than single-stranded DNA. Discoloration during prolonged exposure is due to the higher tolerance of intercalated AO than that of AO adsorbed to proteins. The SCSA is normally used to examine unseparated semen, and a DNA fragmentation index (DFI) is calculated from the distribution of signals on the cytogram [10,11]. With such indirect signal analysis, one must carefully interpret the origin of the signals on the cytogram. One would have to account for signals from non-sperm debris to calculate an accurate DFI.
Several researchers have referred to the interaction of monomeric and oligomeric AO with non-DNA materials [41,42,43,44,45]. AO can permeate organelle membranes, and proton pump-driven intra-lysosomal acidity facilitates the accumulation of permeated AO, the oligomeric aggregates of which exhibit a red shift. When the lysosomal pH rises owing to membrane damage, oligomeric AO dissociates to form monomers, which shifts the emission from orange to green. Thus, AO acts as an indicator of lysosomal localization and proton pump damage. The principle of SCSA misunderstood the chemical properties of AO; it adsorbs to various intercellular materials besides DNA [40].
3.3. The Halo in the SCD Test Comprises Unextracted Nucleoproteins Adhered to DNA Fibers
After in-gel high-salt extraction of nucleoproteins in human sperm, crystal violet (CV) staining is used to visualize DNA fibers that are radiated outward. In the SCD test, the extent of DNA damage is defined as inversely proportional to the area of the violet halo [12,13]. As the violet halo disappears upon in-gel tryptic digestion, CV staining visualized a real picture of unextracted nucleoproteins adhered to DNA fibers [18]. SCD test could not clearly distinguish the halo between LS and DS [18]. The SCD test overlooked the interference of nucleoproteins in their measurements.
3.4. Tightly Packed DNA-Nucleoprotein Complex Physically Blocks Permeation of Certain Dyes and Terminal Deoxynucleotidyl Transferase
After reduction of inter- and intramolecular disulfide cross-linking of protamine with dithiothreitol (DTT), guanidine sulfate competitively dissociates the DNA-nucleoprotein complex, which results in sperm head swelling [20,46]. The dye exclusion assay with trypan blue, which is widely used to observe the plasma membrane integrity of somatic cells, resulted in no staining of almost all the membrane-excluded LS (Figure 2A), whereas the swollen head was stained deep blue (Figure 2B). A similar phenomenon has been observed with bromophenol blue and ponceau 4R [20]. The tightly packed DNA-nucleoprotein complex physically blocks permeation of certain organic dyes.
The terminal deoxynucleotidyl transferase (TdT)-mediated dUTP nick-end labeling (TUNEL) assay [47] catalyzes the addition of fluorescent-dUTP at the 3′-OH end of damaged DNA segments. The assay detects the nicks in the non-swollen DS (Figure 2C) and the swelling increased the intensity of the fluorescence (Figure 2D). The tightly packed nucleus physically blocks permeation of TdT, which has a molecular weight of 32,360 [48]. The signals observed in Figure 2C may be derived from the DNA segments at the surface, the swollen LS gave no fluorescent signal (data not shown). Pre-swelling of the sperm head is recommended for TUNEL assay; however, its sensitivity may be insufficient to detect the early stage of fragmentation.
4. Angle-Modulated 2D-SCPPFGE
2D-SCPFGE sleaves the tangled mass of DNA fibers during the first short run, and the subsequent change in the electrophoretic direction causes them to elongate and align. Variations in the rotation angle and the current-application time provides diverse alignment profiles (Figure 3) [29]. The main aim of this method is to detect small numbers of fibrous DNA segments that appear in the early-stage fragmentation. Figure 3A–C summarize the progress of fragmentation. There found no segments in the inner angle of the fan-like shape formed by the elongated DNA fibers (Figure 3A). A few fibrous segments appeared in the inner angle of the fan (Figure 3B). Along with fragmentation proceeded, various sizes of fibrous fragments are assembled in the inner angle of the fan (Figure 3C). Figure 3D shows the electrophoretic condition to align a set of single-nuclear DNA fibers in parallel without overlapping, they followed S-shaped curves according to the switching intervals of the current.
2D-SCPFGE revealed technical issues with 1D-SCPFGE. The lack of change in direction renders 1D-SCPFGE incapable of drawing out the long fibrous segments from the bundle of fibers [29]. Theoretically, the longest possible segments would arise from chromosome 1 being cleaved in half-and- half, and although 2D-SCPFGE with angle modulation has an improved sensitivity compared with 1D-SCPFGE, whether it can draw out such long segments remains unclear. An experiment should be performed in which a few to several dozen cleavages are made in a nucleus to determine the maximum size of detectable segments. A set of size markers will also be needed to calibrate the segments observed in the micrographs.
LS and DS, naturally occurring extremes, are suitable for qualitative-method validation. The calibration standard required to determine the limit of sensitivity must have a small number of cleavages and should be prepared via artificial cleavage of LS. To date, we have examined the cleavage of DNA fibers by means of heat denaturation [16,40], alkaline hydrolysis [18], hydroxy radicals produced via the Fenton reaction [49], and restriction endonuclease digestion (EcoRI) [29]. In each case, 1D-SCPFGE confirmed that the DNA was cleaved in a dose-dependent manner, the dose-cleavage curves were, however, too steep to simulate early-stage fragmentation.
A future subject to be explored in 2D-SCPFGE is determining from which chromosomes the fibers are derived. Fluorescence in situ hybridization (FISH) enables mapping of a specific DNA sequence in metaphase chromosomes and interphase nuclei. FISH applied to naked DNA fibers, the so-called “DNA fiber-FISH,” yields the highest mapping resolution [50]. The extended long DNA fibers are very fragile in an aqueous solution, and the oscillation of the fibers due to micro-Brownian motion readily cleaves the DNA strands (personal observation). After the fibers are aligned by means of 2D-SCPFGE, a technical issue that remains to be solved is the chemical fixation of the extended long DNA fibers on the solid phase. 2D-SCPFGE is a pioneering approach and is the most sensitive method at present to image large DNA. Therefore, no precedents exist for the preparation of standards with the length of chromosomal-level DNA fibers. Until calibration standards are established, the quantitative performance of 2D-SCPFGE will remain undefined. We tentatively propose 2D-SCPFGE as a reasonably accurate method to determine whether DNA is sequentially intact, defined as the absence of long DNA segments in the region of interest.
5. How to Develop and Validate Analytical Methods for DNA Fragmentation
We learned six lessons from the conventional analytical methods. First, with regards to comparative standards, chemical probes often interact with various intercellular materials besides DNA; therefore, multi-step validations using qualitative and quantitative standards, and subsequent cross-validations among different instruments, are essential to verify the measurement principles. Second, as the critical threshold of DSBs in a sperm is extremely low, analytical methods need to have the sensitivity to detect a low level of cleavage in sperm to predict sperm-derived congenital anomalies. Third, the calibration carves are essential to ensure the quantitative performance. Dose-dependent artificial DNA cleavage is suitable for making a set of the calibration standards, as aforementioned, those mimic the early-stage are not established. Fourth, the eligibility criteria for test samples should be rigorously defined and should be limited to the separated motile-sperm fraction. 1D-SCPFGE originally suggested that almost all immotile sperm are in end-stage fragmentation [19,20]. However, once the cells are fixed and stained or embedded and lysed, determining the type of sperm from which the DNA is derived is difficult. Fifth, when analytical technologies normally used on somatic cells are imported for use on sperm, the unique cellular features of sperm should be considered. For instance, the tightly packed DNA-nucleoprotein complex physically blocks permeation of TdT (Figure 2), and enzymatic analyses, including TUNEL assays, are impeded owing to steric hindrance. Sixth, the CA, SCSA, and SCD test overlooked the interference of nucleoproteins on their measurement principles. Elimination of the nucleoproteins by means of in-gel proteolysis improves the accuracy of measurements in 1D- and 2D-SCPFGE (Figure 1 and Figure 2). We recommend using the naked DNA mass as the comparative standard and the test specimen.
6. Separated Motile Sperm Include Various Impairments Besides DNA Fragmentation
For many years, sperm concentration, sperm motility, and the morphology of the head have been assessed as key parameters of “semen quality”, a term that is used as a surrogate for male fecundity. In ICSI, intra-operative sperm selection usually depends on gross morphology and motility. However, as cited above, 1D-SCPFGE revealed that even after separation of the motile sperm, some exhibit fibrous DNA segments [16,17]. Therefore, we developed several pre-operative methods to evaluate sperm quality, such as 2D-SCPFGE for the detection of the early symptoms of DNA fragmentation [29], sperm-specific dye- and lectin-exclusion assays for assessment of the plasma and acrosomal membranes [20], the dye-retention assay for assessment of the mitochondrial organelle membrane, assessments of endogenous ROS in the mitochondria [20], and visualization of vacuoles in the sperm head [46,51]. These inspections highlight the importance of multifaceted approach to discuss DNA fragmentation.
6.1. Sperm-Specific Two-Step Dye-Exclusion Assay to Observe Damage of Plasma Membrane
All the organelles in somatic cells are grouped into one compartment and enveloped with the plasma membrane. Mammalian sperm are well known to differ greatly from somatic cells in terms of their membrane organization. It is compartmentalized into at least four regions: the acrosomal cap at the anterior end of the head, the posterior region of the head, the midpiece, and the principal and terminal pieces of the tail. The acrosomal cap is composed of the plasma membrane and outer and inner acrosomal membranes [52,53]. In contrast, the posterior region of the head is enveloped solely with the plasma membrane, and the lack of cytoplasm places the nucleus just below the membrane. Thus, damage to the plasma membrane in this region may directly affect DNA integrity.
Ascorbic acid (AA) is a well-known antioxidant that acts as a pro-oxidant in the presence of transitional metals via the Fenton reaction [54]. We previously observed that AA cleaved DNA double strands in membrane-excluded human sperm [49], and that EDTA inhibits this action. DS with damaged plasma membranes have granular DNA segments [20]. Therefore, we hypothesize that damage to the plasma membrane will cause AA in the seminal plasma [55] to come into contact with DNA-transition metal complexes in the nucleus, causing DNA fragmentation.
We developed a sperm-specific two-step dye-exclusion assay with a high affinity for sperm-specific protamines (Figure 4) [20]. Reactive red 195 (RR195) and reactive blue 222 (RB222) both contain 5–6 sulfate residues per molecule, which strongly bind with guanidyl residues in protamines. The sperm were incubated with isotonic RR195, and the reaction mixture was adhered to a plane glass slide, treated with methanol to exclude the plasma membrane, and treated with RB222 for counter-staining. Heads that stained red indicated that RR195 permeated through a damaged plasma membrane. Those that were stained blue indicated that the counter-dye permeated after membrane exclusion, suggesting that the plasma membranes had been intact. As shown in Figure 4, in contrast to LS, almost all of the DS had already undergone plasma-membrane impairment.
6.2. Observation of the Plasma and Acrosomal Membranes via Two-Step Concanavalin A-Labeling
The acrosomal region is covered with three layers: the plasma, outer acrosomal membrane, and inner acrosomal membrane [52]. During the acrosome reaction, the plasma and outer acrosomal membranes fuse [53]; consequently, the DNA-nucleoprotein complex just below the acrosome is protected solely by the inner acrosomal and nuclear membranes. Whether they block extracellular materials in the same way as the plasma membrane is unclear. Furthermore, no methods have been developed to detect damage to the inner acrosomal membrane.
When Cy3-conjugated concanavalin A (Cy3-con A; Molecular Probes, Eugene, OR, USA) permeates through damaged plasma and outer-acrosomal membranes, it binds with high-mannose glycoproteins on the inner-acrosomal membrane. In one of our previous studies [20], sperm were incubated with isotonic Cy3-con A in the presence of methyl a-D-mannopyranoside, an antagonist of concanavalin A [56]. The reaction mixture was treated with methanol to exclude the plasma and outer-acrosomal membranes and subsequently incubated with Alexa 488-conjugated concanavalin A (Alexa488-con A; Molecular Probes) in the same manner as with Cy3-con A. Thereafter, the same field of view was observed under a fluorescent microscope, first with a green filter and then with a red filter. These images were digitally merged. Red fluorescence on the merged photograph indicated that the plasma and outer-acrosomal membranes had already been damaged, facilitating the permeation of Cy3-con A. Green fluorescence indicated that Alexa488-con A had bound to the inner acrosomal membrane that was exposed after methanol treatment. As shown in Figure 5, in contrast to LS, almost all DS had damaged plasma and outer-acrosomal membranes.
6.3. Dye-Retention Assay for Endogenous ROS Generated in the Mitochondria
Live mitochondria generate endogenous ROS as metabolites of oxidative phosphorylation in the tricarboxylic acid cycle. CellROX Orange (Thermo Fisher Scientific, Waltham, MA, USA) permeates the mitochondria and produces orange fluorescence upon oxidation by ROS (Figure 6A) [57]. MitoTracker FM (Thermo Fisher Scientific), which permeates and is retained by the mitochondria regardless of the membrane potential of the organelle [58], produces green fluorescence in the midpiece (Figure 6B). It is employed in a dye-retention assay to observe the integrity of the mitochondrial membrane. The LS exhibit dual fluorescence derived from cellROX and MitoTracker FM, but both of them found in DS [20]. These assays are the referential index of oxidative phosphorylation taking place in mitochondria with an intact organelle membrane.
6.4. Observation of Vacuoles in the Head of Sperm
When we previously investigated ATP-activated channels in human sperm [46,51], we observed that a well-known, highly potent P2Y purinergic-receptor antagonist, reactive blue 2 (RB2) [59], strongly binds with protamine. It faintly stains the sperm head, which is observed as a translucent bluish body, and the vacuoles appear as toneless spots. Staining with RB2, which has three sulfate residues, leads to the swelling and re-formation of the head into an oval shape and the disappearance of the vacuoles regardless of their original features [46], just as guanidine sulfate does (Figure 2B) [20]. Comparison of RB2 staining with or without DTT is the referential index of local failure of inter- and intra-disulfide cross-linkage during spermiogenesis (Figure 7).
7. Multivariable Analyses of DNA Damage and Confounding Factors in Cohort Studies
As non-homologous zygotic repair of paternal DSBs is prone to error, de novo mutations in the zygote are mostly of paternal origin [2]. Single-strand breaks (SSBs) are one of the most frequent DNA lesions, and they are produced daily in the nucleus during DNA repair. Base excision repair [60] is a primary response pathway for the repair of deleterious DNA lesions, including non-bulky DNA adducts, apurinic/ apyrimidinic (AP) sites [61,62], and SSBs. If unrepaired or incorrectly repaired, these lesions threaten genetic integrity through their potential conversion to lethal DSBs during DNA replication. Single-nuclear DSBs and SSBs (AP sites) should preferably be analyzed together, although, considering their frequencies, the latter may pose a greater risk to DNA integrity.
Early apoptotic events or oxidative stress induce phospholipid flip-flop in the plasma membrane [63]. Endogenous ROS in the mitochondria [20] and exogenous ROS due to plasma membrane damage at the posterior region of the sperm head [49] should be taken into account as potential confounding factors. Researchers have reported that protamine deficiency is responsible for vacuole formation, and secondarily, for DNA fragmentation [64,65]. We have also observed that local failure of disulfide cross-linkage may play a critical role in determining sperm-head morphology as well as vacuole formation [46]. The underlying mechanism may be either that insufficient cross-linkage destabilizes all of the DNA or that it damages only DNA at the inner surface of the vacuole. Detection of the latter, however, is challenging at present, owing to the difficulty of probes with high molecular weight to permeate through the tightly packed nucleus. The age-dependent decline in embryonic repair of paternal DNA [60] is another potentially confounding factor.
In general, laboratory mice are generated via repeated inbreeding and natural delivery, and they remain highly fecund over many generations. In livestock farming, the seed bull is selected through progeny testing, and the selected bull will have the highest fecundity. The experiments using inbreeding mice or seed bull fit to human couples who pregnant spontaneously, in contrast, often un-fit to the infertile patients. Mice used as pathogenic animal models are classically generated through selective pedigree breeding; however, an animal model of male infertility cannot be sustainably propagated in such a way. Male infertility is not an issue in livestock farming. In fact, therapy for male infertility is in high demand among monogamous humans.
Recent progress in gene manipulation has led to the creation of various knockout mice with particular gene variants. The diverse symptoms in human infertility are caused by interactions between congenital and acquired reproductive dysfunctions in both the man and the woman. Determining the genetic origins of the varying symptoms of infertile men may be difficult with current techniques. Thus, well-designed cohort studies with special consideration to confounding factors may accelerate our understanding of the etiology of male infertility.
8. Vision for the Future
As far as in the scope of the present review, the eligibility criteria for injectable sperm in clinical ICSI became more complex: a motile sperm with normal head, midpiece, and tail morphologies, intact acrosomal and mitochondrial membranes, physiological oxidative phosphorylation in the mitochondria, and an absence of vacuoles and DNA damages such as DSBs and SSBs (AP sites). The fundamental framework for quality control of ICSI is to clarify which underlying dysfunctions in the motile sperm render them ineligible for injection. According to our preliminary observations, the infertile men who exhibited normal “semen quality under phase contrast microscopy” but suffered from repetitive ICSI failures exhibited higher frequencies of the early-stage fragmentation, as well as vacuoles and damage to the plasma membrane than the men in couples who became pregnant within a few ICSI treatment cycles. We are busy developing of a modified 2D-SCPGFE and the corresponding calibration standards for the simultaneous observation of early-stage DSBs and SSBs (AP sites). When the development is complete, we plan to conduct multivariable analyses, step-by-step and multifaceted approaches are necessary to find answers through the cohort studies.
Another issue in ICSI quality control is intra-operative, non-invasive testing. The RR195-exclusion assay without counterstaining serves as an example of such a test; motile sperm that stain red should be discarded. Our preliminary experiments suggest that various acidic dyes, including those that contain sulfate residues, have high affinity to protamines [20]. We hope to discover a dye that is excluded completely by the intact plasma membrane, which will require detailed exposure experiments according to reproductive toxicity guidelines prior to clinical use.
Supplementary Materials
This review includes no Supplementary Materials.
Author Contributions
Conceptualization, S.K. and Y.O.; methodology, S.K. and Y.O.; validation, S.K. and Y.O.; formal analysis, S.K. and Y.O.; investigation, S.K. and Y.O.; resources, S.K. and Y.O.; data curation, S.K. and Y.O.; writing—original draft preparation, S.K. and Y.O.; writing—review and editing, S.K. and Y.O.; visualization, S.K. and Y.O.; supervision, S.K. and Y.O.; project administration, S.K. and Y.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The datasets generated and analyzed in the present study are available from the corresponding author upon reasonable request.
Conflict of Interest
The authors declare the absence of any conflicting interests. This review cited our published data, “Institutional Review Board Statement” and “Informed Consent Statement” are stated in our published articles.
References
- Acuna-Hidalgo, R.; Veltman, JA.; Hoischen, A. New insights into the generation and role of de novo mutations in health and disease. Genome Biol. 2020, 17, 241. [Google Scholar] [CrossRef] [PubMed]
- Middelkamp, S.; Van Tol, HTA. ; Spierings, DCJ.; Boymans, S.; Victor Guryev, V. Sperm DNA damage causes genomic instability in early embryonic development. Sci. Adv. 2020, 6, eaaz7602. [Google Scholar] [CrossRef] [PubMed]
- Oud, MS.; Smits, RM.; Smith, HE.; Mastrorosa, FK.; Holt, GS. A de novo paradigm for male infertility nature com. 2022, 13, 154.
- Ricci, G.; Perticarari, S.; Fragonas, E.; Giolo, E.; Canova, S. Apoptosis in human sperm: its correlation with semen quality and the presence of leukocytes. Human Reprod. 2002, 17, 2665–2672. [Google Scholar] [CrossRef]
- Nakanishi, Y.; Shiratsuchi, A. Phagocytic Removal of Apoptotic Spermatogenic Cells by Sertoli Cells: Mechanisms and Consequences. Biol. Pharma. Bull. 2004, 27, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, F. ; fmarchetti@lbl. gov, Essers, J.; R. Kanaar, R.; Wyrobek, AJ. Disruption of maternal DNA repair increases sperm-derived chromosomal aberrations PNAS, 2007, 104, 17725–17729. [Google Scholar]
- Arends, MJ.; Morris, RG.; Wyllie. AH. Apoptosis. The role of the endonuclease. Am. J. Pathol. 1990, 136, 593–608. [Google Scholar]
- Borini, A.; Tarozzi, N.; Bizzaro, D.; Bonu, MA.; Fava, L. Sperm DNA fragmentation: paternal effect on early post-implantation embryo development in ART. Human Reprod. 2006, 21, 2876–2881. [Google Scholar] [CrossRef] [PubMed]
- Wennerholm, UB.; Bergh, C.; Hamberger, L.; Lundin, K.; Nilsson, L.; M. Wikland, M.; Källén, B. Incidence of congenital malformations in children born after ICSI. Human Reproduction. 2000, 15, 944–948. [Google Scholar] [CrossRef]
- Evenson, DP. The sperm chromatin structure assay (SCSA®) and other sperm DNA fragmentation tests for evaluation of sperm nuclear DNA integrity as related to fertility. Anim. Reprod. Sci. 2016, 169, 56–75. [Google Scholar] [CrossRef]
- Van Brakel, J.; Dinkelman-Smit. M.; de Muinck Keizer-Schrama, SMPF.; Hazebroek, FWJ.; G R Dohle, GR. Sperm DNA damage measured by sperm chromatin structure assay in men with a history of undescended testes. Andrology 2017, 5, 838–843. [Google Scholar] [CrossRef]
- Fernández, JL.; Muriel, L.; Rivero, MT.; Goyanes, V.; Vazquez, R. The sperm chromatin dispersion test: A simple method for the determination of sperm DNA fragmentation. J. Androl. 2003, 24, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Fernández, JL.; Johnston, S.; Gosálvez, J. Sperm chromatin dispersion (SCD) assay, A clinician's guide to sperm DNA and chromatin damage. (ed. Zini, A., & Agarwal, A.), Springer nature, Barling, 2018,137-152.
- Tice, RR.; Agurell, E.; Anderson, D.; Burlinson, B.; Hartmann, A.; H Kobayashi, H.; Miyamae, Y.; Rojas, E.; Ryu, JC.; Y F Sasaki, YF. Single cell gel/comet assay: guidelines for in vitro and in vivo genetic toxicology testing. Environ. Mol. Mutagen. 2000, 35, 206–221. [Google Scholar] [CrossRef]
- Olive, PL.; Banáth, JP. The comet assay: A method to measure DNA damage in individual cells. nature Protoc. 2006, 1, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, S.; Yoshida, J.; Ishikawa, H.; Takamatsu, K. Single-cell pulsed-field gel electrophoresis to detect the early stage of DNA fragmentation in human sperm nuclei. PLoS One 2012, 7, e42257. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, S.; Yoshida, J.; Ishikawa, H.; Takamatsu, K. Single-nuclear DNA instability analyses by means of single-cell pulsed-field gel electrophoresis - Technical problems of the comet assay and their solutions for quantitative measurements. J. Mol. Biomark. Diagn. 2013, S5, 005. [Google Scholar] [CrossRef]
- Kaneko, S.; Takamatsu, K. Revalidation of the sperm chromatin dispersion test and the comet assay using intercomparative studies between purified human sperm without and with end-stage DNA fragmentation. J. Med. Diagn. Meth. 2023, 12. 406.
- Kaneko, S.; Takamatsu, K. Re-Evaluation of Significance of Anti-Sperm Antibodies in Clinical Immune Infertility-Antigenicity of Human Sperm Diminishes during DNA Fragmentation. J. Med. Diagn. Meth. 2023, 12, 439. [Google Scholar]
- Kaneko, S.; Okada, Y.; Takamatsu, K. Sperm Specific Two-Step Dye Exclusion Assays to Evaluate Integrity of Plasma and Organelle Membranes-New Approach for Quality Assurance of the Sperm for Intra-Cytoplasmic Sperm Injection. J. Med. Diagn. Meth. 2024, 12, 450. [Google Scholar]
- Bortner, CD.; Cidlowski, JA. Apoptotic volume decrease and the incredible shrinking cell. Cell Death Differ. 2002, 9, 1307–10. [Google Scholar] [CrossRef] [PubMed]
- van Gent, DC.; Hoeijmakers, JH.; Kanaar, R. Chromosomal stability and the DNA double-stranded break connection. Nat. Rev. Genet. 2001, 2, 196–206. [Google Scholar] [CrossRef]
- Ceccaldi, R.; Rondinelli, B.; D’Andrea, AD. Repair pathway choices and consequences at the double-strand break. Trends Cell Biol. 2016, 26, 52–64. [Google Scholar] [CrossRef]
- Kalisky, T.; Quake, ST. Single-cell genomics Nat. Methods 2011, 8, 311–314. [Google Scholar]
- Wang, J.; Fan, HC.; Behr, B.; Quake. SR. Genome-wide single-cell analysis of recombination activity and de novo mutation rates in human sperm. Cell 2012, 150, 402–12. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, DC.; Cantor, CR. Separation of yeast chromosome-sized DNAs by pulsed field gradient gel electrophoresis Cell 1984, 37, 67–75.
- Herschleb, J.; Ananiev, G.; Schwartz, DC. Pulsed-field gel electrophoresis Nat. Protoc. 2007, 2, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, DW. Restriction endonuclease digestion of DNA in agarose plugs. CSH Protoc. 2006, pdb.prot4031.
- Kaneko, S.; Takamatsu, K. Angle modulated two-dimensional single cell pulsed-field gel electrophoresis for detecting early symptoms of DNA fragmentation in human sperm nuclei Scientific Reports 2024, 14, 840.
- Balhorn, R. The protamine family of sperm nuclear proteins. Genome Biol. 2007, 8, 227. [Google Scholar] [CrossRef]
- Torres-Flores, U.; Hernández-Hernández, A. The interplay between replacement and retention of histones in the sperm genome. Front Genet. 2020, 11, 780. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y. Sperm chromatin condensation: epigenetic mechanisms to compact the genome and spatiotemporal regulation from inside and outside the nucleus. Genes & Genetic Systems 2022, 97, 41–53. [Google Scholar]
- Hagstrom, KA.; Meyer, BJ. Condensin and cohesin: More than chromosome compactor and glue. Nat. Rev. Genet. 2003, 4, 520–534. [Google Scholar] [CrossRef]
- Nickerson, J. Experimental observations of a nuclear matrix. J. Cell Sci. 2001, 114, 463–474. [Google Scholar] [CrossRef]
- Shaman, JA.; Yamauchi, Y.; Ward, WS. Function of the sperm nuclear matrix. Arch. Androl. 2007, 53, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Ada Soler-Ventura, A.; Castillo, J.; de la Iglesia, A.; Jodar, M.; Barrachina, F.; Ballesca, JL.; Oliva, R. Mammalian sperm protamine extraction and analysis, a step-by-step detailed protocol and brief review of protamine alterations. Protein & Peptide Letters. 2018, 25, 424–433. [Google Scholar]
- Rawlings, ND.; Barrett, AJ. Families of serine peptidases. Meth. Enzymol. 1994, 244, 19–61. [Google Scholar]
- Jameson, GW.; Elmore, DT. Affinity chromatography of bovine trypsin. A rapid separation of bovine α- and β-trypsin. Biochem. J. 1974, 141, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Ebeling, W.; Hennrich, N.; Klockow, M.; Metz, H.; Orth, HD.; Lang, H. Proteinase K from Tritirachium album Limber. Eur. J. Biochem. 1974, 47, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, S.; Takamatsu, K. Re-evaluation of sperm chromatin structure assay (SCSA). J. Med. Diagn. Meth. 2023, 12, 399. [Google Scholar]
- Moriyama, Y.; Takano, T.; Ohkuma, S. Acridine orange as a fluorescent probe for lysosomal proton pump. J. Biochem. 1982, 92, 1333–1336. [Google Scholar] [CrossRef] [PubMed]
- Traganos, F.; Darzynkiewicz, Z. Lysosomal proton pump activity: supravital cell staining with acridine orange differentiates leukocyte subpopulations. Methods Cell Biol. 1994, 41, 185–194. [Google Scholar]
- Han, J.; Burgess, K. Fluorescent indicators for intracellular pH. Chem. Rev. 2010, 110, 2709–2728. [Google Scholar] [CrossRef]
- Darzynkiewicz, Z.; Kapuscinski, J. Acridine orange: A versatile probe of nucleic acids and other cell constituents. Flow cytometry and sorting. Second edition, Wiley, New York, US, 1990, 291-314.
- Uchimoto, T.; Nohara, H.; Kamehara, R.; Iwamura, M.; Watanabe, N. Mechanism of apoptosis induced by a lysosomotropic agent, L-Leucyl-L-Leucine methyl ester. Apoptosis 1999, 4, 357–362. [Google Scholar] [CrossRef]
- Kaneko, S.; Okada,Y. ; Yokota, S.; Takamatsu, K. (2023) Reactive Blue Dye: Highlights of Vacuoles in Human Sperm. J. Med. Diagn. Meth. 2023, 12, 400. [Google Scholar]
- Crowley, LC.; Brooke, J.; Marfell, BJ.; Waterhouse, NJ. Detection of DNA Fragmentation in Apoptotic Cells by TUNEL. Cold Spring Harb. Protoc. 2016. [CrossRef]
- Chang, LMS.; Bollum, F. Deoxynucleotide-polymerizing Enzymes of Calf Thymus Gland. J. Biol. Chem. 1971, 909-916.
- Kaneko, S.; Yoshida, J.; Takamatsu, K. Direct visualization of ascorbic acid inducing double-stranded breaks in single nuclear DNA using single-cell pulsed field gel electrophoresis. Indian, J. App. Res. 2015, 5, 248–252. [Google Scholar]
- Anton, K.; Raap, AK.; Florijn, RJ.; Blonden, LAJ. ; Wiegant, J. Fiber FISH as a DNA Mapping Tool. Methods 1996, 9, 67–73. [Google Scholar]
- Kaneko, S.; Yoshida, J.; Takamatsu, K. Low density regions of DNA in human sperm appear as vacuoles after translucent staining with reactive blue 2. J. Med. Diagn. Meth. 2013, 2, 145. [Google Scholar] [CrossRef]
- Patel, KS. Human Sperm Morphology. Atlas of Assisted Reproductive Technologies (ed. Ghumman, S.), Springer nature, Berlin, Germany, 2023, 179-188.
- Yanagimachi, R. Mammalian sperm acrosome reaction: Where does it begin before fertilization? Biol. Reprod. 2011, 85, 4–5. [Google Scholar] [CrossRef]
- Shen, J.; Griffiths, PT.; Campbell, SJ.; Utinger, B.; Kalberer, M. Ascorbate oxidation by iron, copper and reactive oxygen species: review, model development, and derivation of key rate constants Scientific Reports 2021, 11, 7417.
- Colagar, AH.; Marzony, ET. Ascorbic Acid in Human Seminal Plasma: Determination and Its Relationship to Sperm Quality. J. Clin. Biochem. Nutr. 2009, 45, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Clegg, RM.; Loontiens, FG.; Van Landschoot, A.; Jovin, TM. Binding kinetics of methyl alpha-D-mannopyranoside to concanavalin A: temperature-jump relaxation study with 4-methylumbelliferyl alpha-D-mannopyranoside as a fluorescence indicator ligand. Biochemistry 1981, 20, 4687–4692. [Google Scholar] [CrossRef]
- Celeghini, EC.; Alves, MB.; De Arruda, RP.; de Rezende, GM.; Florez Rodriguez, SA. Efficiency of CellROX deep red® and CellROX orange® fluorescent probes in identifying reactive oxygen species in sperm samples from high and low fertility bulls. Anim. Biotechnol. 2021, 32, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Kholmukhamedov, A.; Schwartz, JM.; Lemasters, JJ. Mitotracker probes and mitochondrial membrane potential. Shock 2013, 39, 543. [Google Scholar] [CrossRef] [PubMed]
- Claes, P.; van Kolen, K.; Roymans, D.; Blero, D.; Vissenberg, K. Reactive blue 2 inhibition of cyclic AMP-dependent differentiation of rat C6 glioma cells by purinergic receptor independent inactivation of phosphatidylinositol 3-kinase. Biochem. Pharmacol. 2004, 67, 1489–1498. [Google Scholar] [CrossRef] [PubMed]
- Atamna, H.; Cheung, I.; Ames, BN. A method for detecting abasic sites in living cells: age-dependent changes in base excision repair. PNAS. 2000, 97, 686–691. [Google Scholar] [CrossRef]
- Talpaert-Borlè, M. Formation, detection and repair of AP sites. Mutation Res. 1987, 181, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Boiteux, S.; Guillet, M. Abasic sites in DNA: repair and biological consequences in Saccharomyces cerevisiae. DNA Repair (Amst) 2004, 3, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bratton, DL.; Fadok, VA.; Richter, DA.; Kailey, JM.; Guthrie, LA. Appearance of Phosphatidylserine on Apoptotic Cells Requires Calcium-mediated Nonspecific Flip-Flop and Is Enhanced by Loss of the Aminophospholipid Translocase. J. Biol. Chem. 1997, 272, 26159–26165. [Google Scholar] [CrossRef] [PubMed]
- García-Peiró, A.; Martínez-Heredia, J.; Oliver-Bonet, M.; Abad, C.; Amengual, MJ.; Navarro, J. Protamine 1 to protamine 2 ratio correlates with dynamic aspects of DNA fragmentation in human sperm. Fertil. Steril. 2011, 95, 105–109. [Google Scholar] [CrossRef]
- Ni, K.; Spiess, AN.; Schuppe. HC.; Steger, K. The impact of sperm protamine deficiency and sperm DNA damage on human male fertility: A systematic review and meta-analysis. Andrology 2016, 4, 789–799. [Google Scholar] [CrossRef]
Figure 1.
Mono-dimensional single-cell pulsed-field gel electrophoresis. A: LS digested in the gel with trypsin, B: Electrophoretogram of LS, C: DS digested in the gel with trypsin, D: Electrophoretogram of DS. The scale bars represent 20 mm.
Figure 1.
Mono-dimensional single-cell pulsed-field gel electrophoresis. A: LS digested in the gel with trypsin, B: Electrophoretogram of LS, C: DS digested in the gel with trypsin, D: Electrophoretogram of DS. The scale bars represent 20 mm.
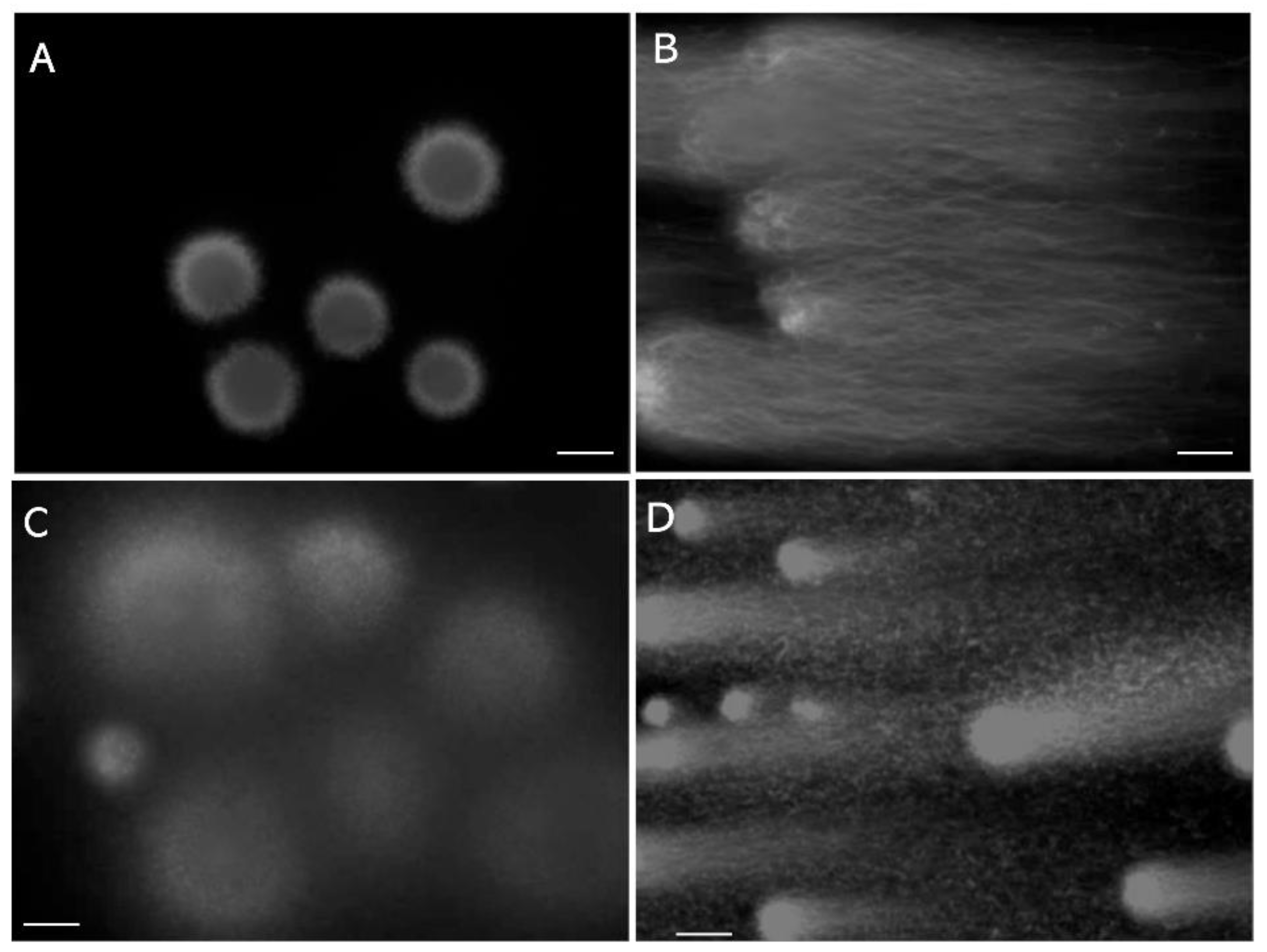
Figure 2.
Tightly packed DNA-nucleoprotein complex physically blocks permeation of certain organic dyes. A: Membrane-excluded LS stained with 0.05% trypan blue. B: Membrane-excluded LS swollen with 0.1 mol/L guanidine sulfate and 5.0 mmol/L DTT, stained in the same manner as those in A. C: TUNEL assay for non-swollen DS, performed using the TUNEL Assay Apoptosis Detection Kit (Cosmo-Bio, Tokyo, Japan). D: TUNEL assay for swollen DS, performed using the same Kit. The scale bars represent 10 µm.
Figure 2.
Tightly packed DNA-nucleoprotein complex physically blocks permeation of certain organic dyes. A: Membrane-excluded LS stained with 0.05% trypan blue. B: Membrane-excluded LS swollen with 0.1 mol/L guanidine sulfate and 5.0 mmol/L DTT, stained in the same manner as those in A. C: TUNEL assay for non-swollen DS, performed using the TUNEL Assay Apoptosis Detection Kit (Cosmo-Bio, Tokyo, Japan). D: TUNEL assay for swollen DS, performed using the same Kit. The scale bars represent 10 µm.
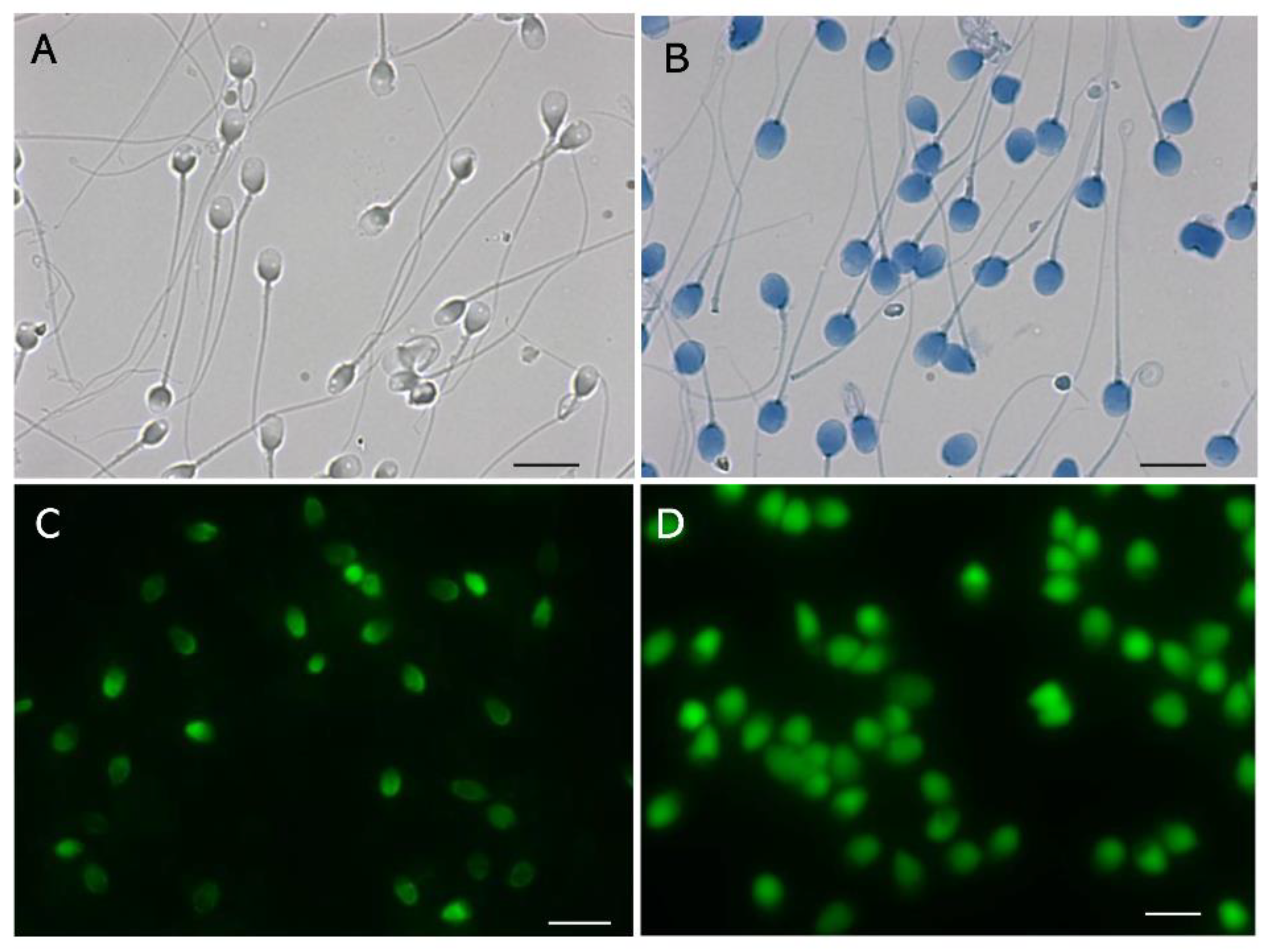
Figure 3.
Angle-modulated two-dimensional single-cell pulsed-field gel electrophoresis. LS were electrophoresed by means of 2D-SCPFGE; for details of image enhancement, please see reference [29]. A: After the first run, the glass slide is turned 150 degrees (2D-SCPFGE-0-150, first run: 5.0 min; second run: 3.5 min). DNA fibers are elongated in a fan-like shape, and no DNA is observed in the inner angle of the fan. B: The arrows indicate a few fibrous segments separated in the inner angle of the fan. C: Segments of various sizes are assembled in the inner angle of the fan. D: 2D-SCPFGE-(-75)-0 (3.0/10 min) aligned a set of DNA fibers in parallel without the need for downsizing. The scale bars represent 50 µm.
Figure 3.
Angle-modulated two-dimensional single-cell pulsed-field gel electrophoresis. LS were electrophoresed by means of 2D-SCPFGE; for details of image enhancement, please see reference [29]. A: After the first run, the glass slide is turned 150 degrees (2D-SCPFGE-0-150, first run: 5.0 min; second run: 3.5 min). DNA fibers are elongated in a fan-like shape, and no DNA is observed in the inner angle of the fan. B: The arrows indicate a few fibrous segments separated in the inner angle of the fan. C: Segments of various sizes are assembled in the inner angle of the fan. D: 2D-SCPFGE-(-75)-0 (3.0/10 min) aligned a set of DNA fibers in parallel without the need for downsizing. The scale bars represent 50 µm.
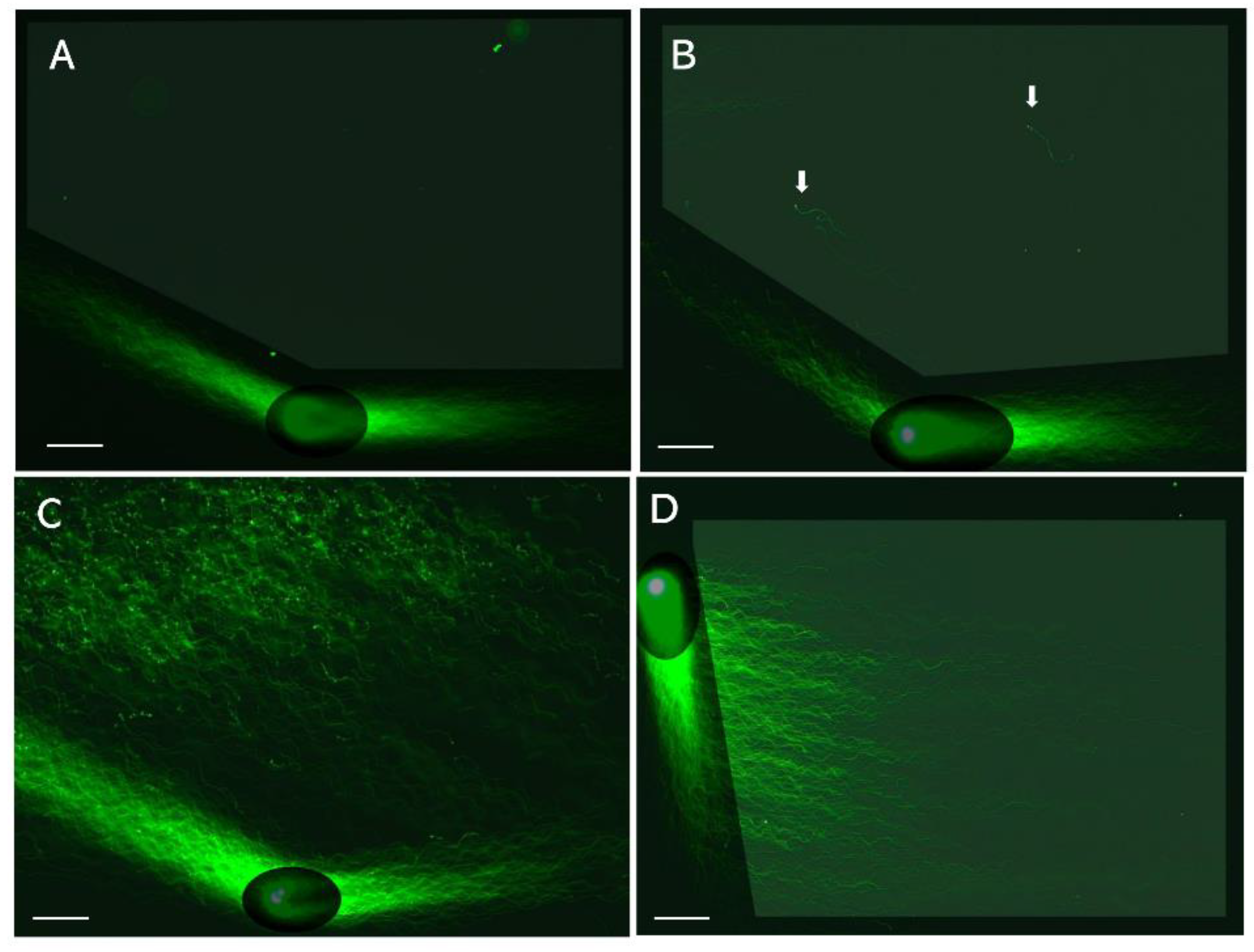
Figure 4.
Sperm-specific two-step dye-exclusion assay for living sperm and denatured sperm. A: LS, B: DS. Scale bars represent 10 µm.
Figure 4.
Sperm-specific two-step dye-exclusion assay for living sperm and denatured sperm. A: LS, B: DS. Scale bars represent 10 µm.
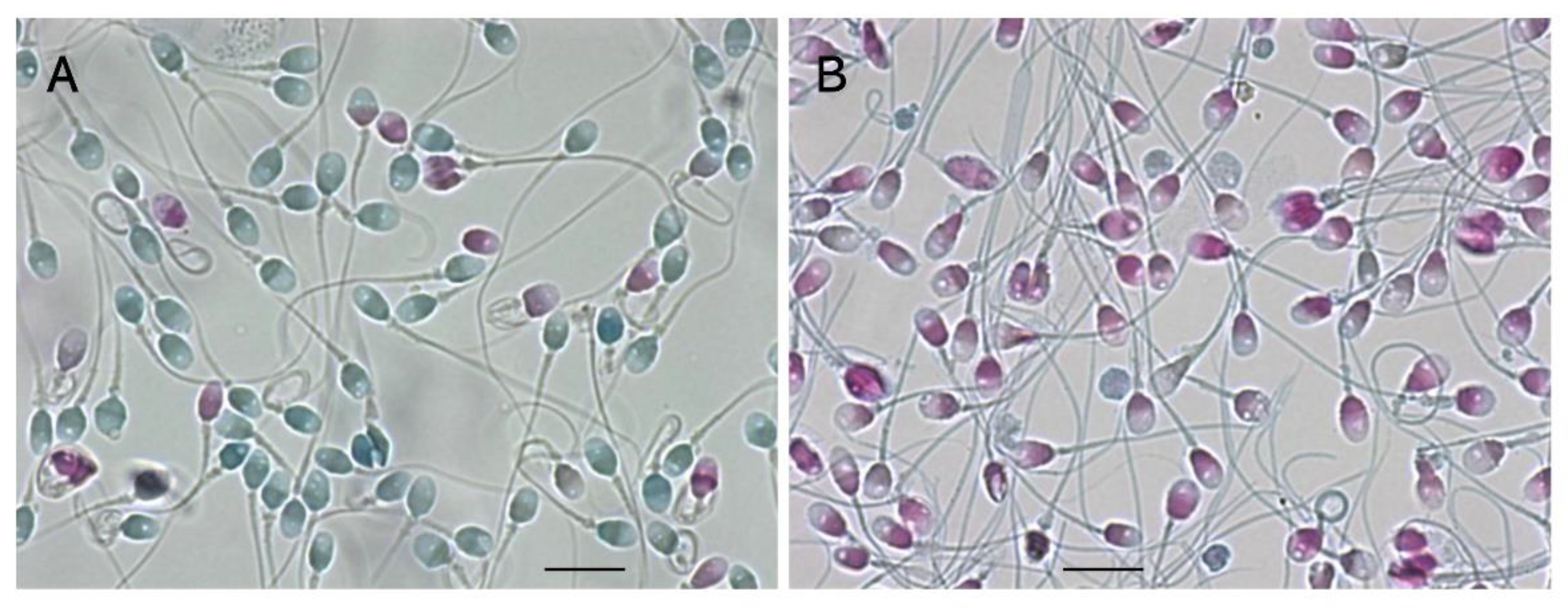
Figure 5.
Sperm-specific two-step concanavalin-A labeling for living and denatured sperm. A: LS, B: DS. The scale bars represent 10 µm.
Figure 5.
Sperm-specific two-step concanavalin-A labeling for living and denatured sperm. A: LS, B: DS. The scale bars represent 10 µm.
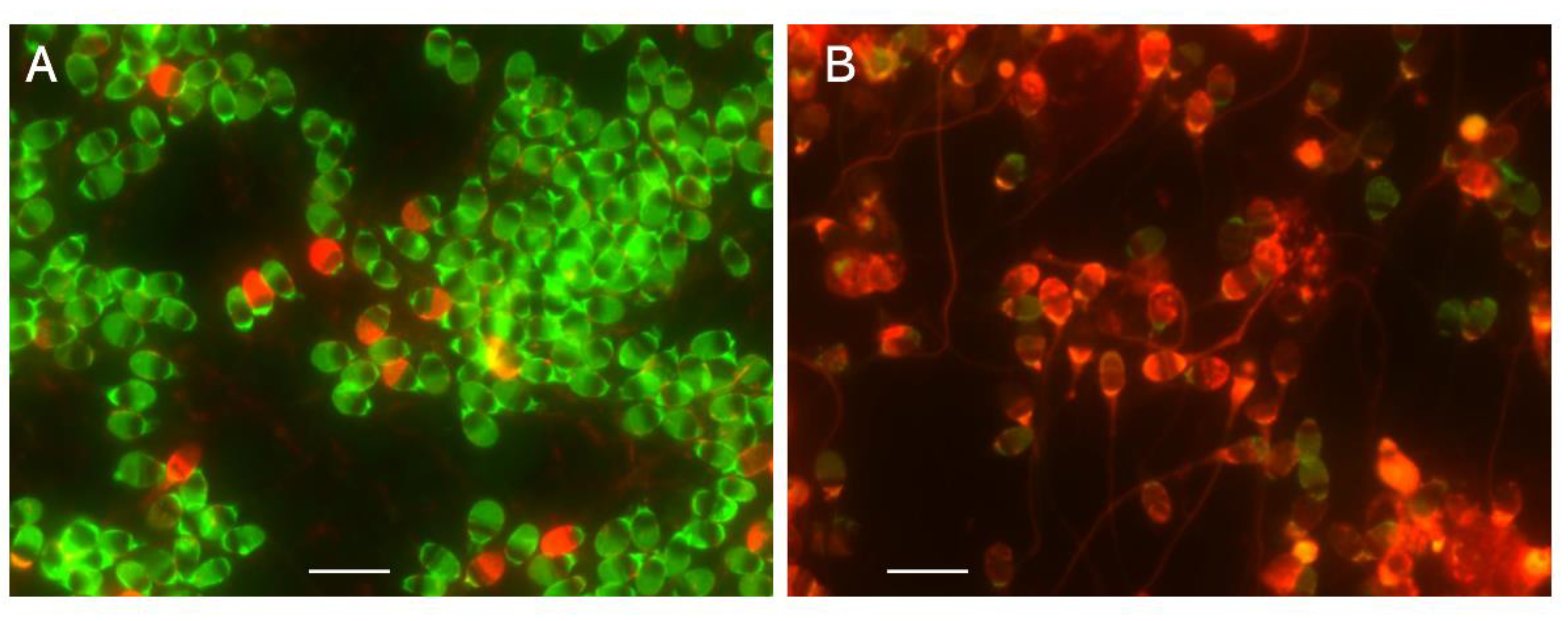
Figure 6.
Dye-retention assays for mitochondria of living sperm. A: Orange fluorescence was due to oxidative phosphorylation in the mitochondria of LS. The black-and-white photograph was taken with a highly sensitive charge-coupled device camera. B: Green fluorescence was derived from the retention of MitoTracker FM in the mitochondria of LS. The scale bars represent 10 µm.
Figure 6.
Dye-retention assays for mitochondria of living sperm. A: Orange fluorescence was due to oxidative phosphorylation in the mitochondria of LS. The black-and-white photograph was taken with a highly sensitive charge-coupled device camera. B: Green fluorescence was derived from the retention of MitoTracker FM in the mitochondria of LS. The scale bars represent 10 µm.
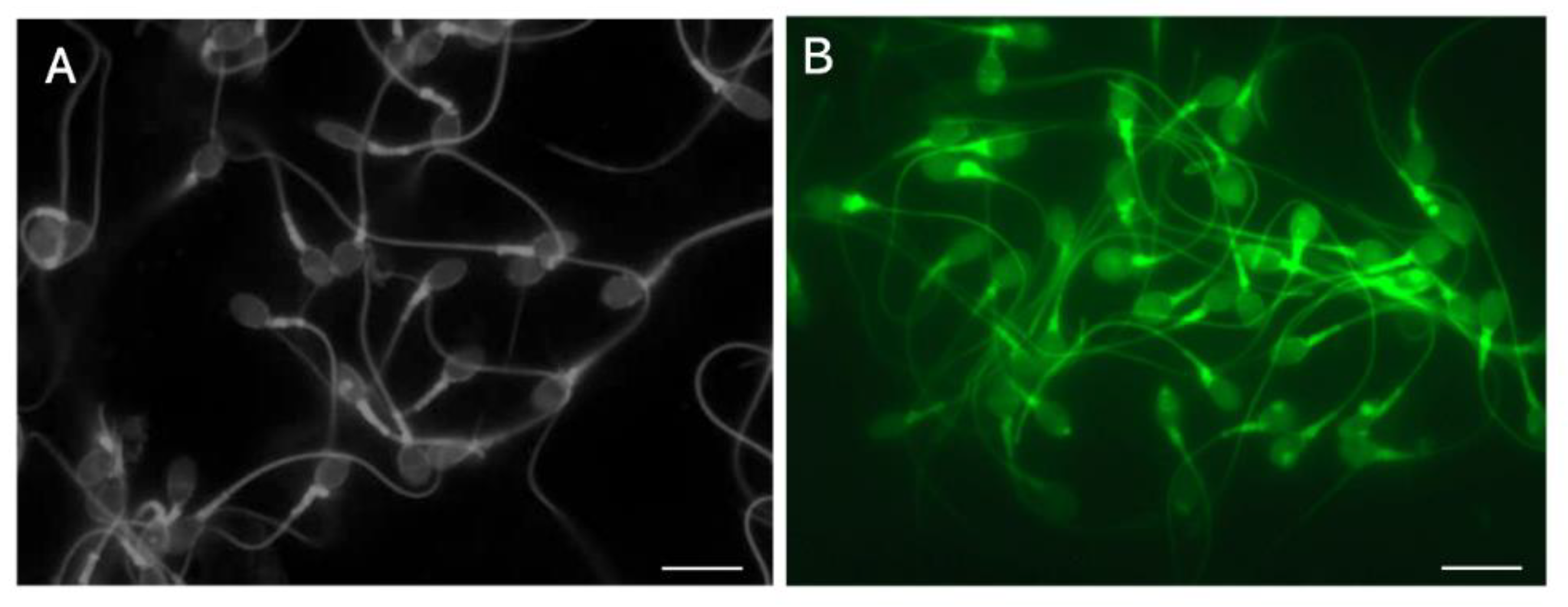
Figure 7.
Observation of the vacuole in human sperm with reactive blue 2 and swelling of the sperm head via reduction of disulfide bonds. LS were stained with 0.02% RB2 (0.1 mol/L Na2CO3–NaHCO3, pH = 10.0) in the absence (A) or presence (B) of 5.0 mmoL/L DTT. A: The vacuoles were visualized as toneless spots, B: DTT swelled the head and resulted in the disappearance of the vacuoles. The scale bars represent 10 µm.
Figure 7.
Observation of the vacuole in human sperm with reactive blue 2 and swelling of the sperm head via reduction of disulfide bonds. LS were stained with 0.02% RB2 (0.1 mol/L Na2CO3–NaHCO3, pH = 10.0) in the absence (A) or presence (B) of 5.0 mmoL/L DTT. A: The vacuoles were visualized as toneless spots, B: DTT swelled the head and resulted in the disappearance of the vacuoles. The scale bars represent 10 µm.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



