Submitted:
11 May 2024
Posted:
14 May 2024
You are already at the latest version
Abstract
Reviewed are recent efforts leading to chemo-enzymatic syntheses of strongly fluorescent ribosides of nucleobase analogs, potentially applicable in analytical biochemistry and cell biology. Described are syntheses and properties of fluorescent purine, 8-azapurine and etheno-purine ribosides, obtained using various types of the purine nucleoside phosphorylase (PNP) as catalysts of ribosylation, and -ribose-1-phosphate (r1P) as a second substrate. In several instances, the ribosylation sites were different than the canonical purine N9. Some of the obtained ribosides show fluorescence yields close to 100%. Their main applications are assays of PNP, nucleoside hydrolases, and other enzyme activities both in vitro and, possibly, within the living cells, using fluorescence microscopy.
Keywords:
purine-nucleoside phosphorylase
; nucleoside analogs
; fluorescence
; chemo-enzymatic synthesis
1. Introduction: Fluorescent Nucleoside Analogs – An Outlook
Fluorescence spectroscopy is one of the most sensitive methods of bio-molecules detection and measurements of their interactions and activities, at both the ensemble and single-molecule levels [1,2,3]. For this reason, various fluorescence techniques are frequently used in biological, biochemical and biophysical research. Unfortunately, some of the most important biomolecules (nucleic acids, membranes, some proteins) are not fluorescent under natural conditions (neutral aqueous medium, moderated temperatures). Therefore, direct application of fluorescence methods to biological research in vivo or in vitro requires finding appropriate fluorescent markers, which can provide structural and dymamic information about molecules as well as their activities.
Among these, fluorescent nucleoside and nucleotide analalogs play key role [4]. They are applied, inter alia, to structural and dynamic research of nucleic acids, including ribozymes [5,6,7,8], their interaction with proteins [9], and detection of enzymatic activities of enzymes related to nucleic acid metabolism both in vitro and in vivo with aid of the fluorescence microscopy [10,11,12].
Natural nucleosides and nucleotides are virtually nonfluorescent in neutral aqueous media [13] with exception of some weakly fluorescent minor RNA subunits [14]. First fluorescent nucleoside analogs, among them the intensely fluorescent 2-aminopurine riboside, were described five decades ago [15], and some of them still are employed in biophysical research [16,17]. Since that, hundreds of fluorescent nucleobase and nucleoside analogs were developed for various applications, most of them by multi-step chemical procedures [4,6,10,18,19,20,21].
There are two broad classses of nucleoside analogs [5]: (i) nucleosides linked to strong aromatic or heteroaromatic fluorophores, like coumarin, pyrene or fluorene, typically via the C-8, N7, or ribose moiety, and (ii) nucleosides with the heteroaromatic ring (purine or pyrimidine) replaced by a fluorescent analog of nucleobase [5,21]. The most useful analogs are those which combine good spectral parameters with substrate or inhibitory activities towards the selected enzymatic or other biological systems, and this condition is usually fulfilled by isosteric or isomorphic (that is, mimicking the natural nucleosides in size, shape, and hydrogen bonding abilities) analogs. In this brief review we concentrate mainly on this kind of nucleoside analogs.
There is a growing number of such analogs known, and recent finding of Tor’s group enlarged the available “libraries” by the substituted thieno- and thiazolo-pyrimidine compounds, isosteric and, to the large extent, isofunctional with natural purine nucleosides [22]. All the thieno- and thiazolo-purine nucleoside analogs are strongly fluorescent in the visible region [22,23]. These are all C-nucleosides, what makes them resistant to some nucleolytic enzymes, what is important for biological applications.
Many nucleoside analogs, including elements of “RNA emissive alphabet” can be selectively incorporated into polynucleotides and ribozymes, either chemically, or using enzymes like T7 RNA polymerase and other polymerases [5,22].
Most of the fluorescent analogs are generated chemically, using sometimes quite complex multi-step procedures, but some can be obtained in much simpler, enzymatic way [24,25]. Purine-nucleoside phosphorylase (see below) offer possibilty of nucleoside (but not of the C-nucleoside) analogs synthesis from nucleobase and pentose analogs.
There are also examples of fluorescent pyrimidine nucleoside analogs [5,6,21,26,27], but nearly all of them belong to the first (“noncanonical”) group, with pyrimidine moiety liked to organic fluorophore, or significantly modified. They are nevertheless utilized in polynucleotide and nucleic acid reserch, e.g. to detection of apurinic sites in double-stranded polynucleotides or DNA fragments [28].
2. Purine Nucleoside Phosphorylase as a Synthetic Tool
Purine-nucleoside phosphorylase (PNP, E.C. 2.4.2.1) catalyzes a reversible phosphorolysis of natural purine nucleosides (or deoxynucleosides) to the respective nucleobases and α-(deoxy)ribose-1-phosphate (r1P, [29]). It is a key enzyme of the so-called purine-salvage pathway [30], and its physiological role is regulation of nucleoside concentration within the cells.
Genetic deficiency of PNP activity leads to severe impairment of immune system in man [29,31], and is a subject of experimental gene therapy [31]. Inhibitors of PNP reveal potent pharmacological activities towards bacterial, viral, and parasitic infections (including malaria), and also can be applied as antineoplastic and immunosuoppresive agents [29,32]. One of them, the transition-state inhibitor forodesine (known as immucilin H, [33]), with inhibition constant ~60 pM, has been recently approved in Japan for the treatment of relapsed/refractory peripheral T-cell lymphoma [34]. Even more potent is ulodesine [35], with Ki ~9 pM, actually under examination.
Various forms of PNP are employed as catalysts in enzymatic syntheses of nucleosides and their analogs (for reviews, see refs. [24,25,36,37,38,39]). For canonical purines and many analogs, the reaction equillibrium is shifted towards nucleoside synthesis [29], but there are also ribosides for which phosphorolysis is virtually irreversible, and these are ideal to serve as ribose-donors in the synthetic “one pot” reactions, while the purified r1P is rarely used because of its high price and relative instability [36]. The commonly known example is a commercially available 7-methylguanosine [40], and another good ribose donors involve 7-methyl-thioguanosine, 7-methyl-inosine and nicotinamide riboside [29]. Another way of the “one-pot” purine (deoxy)nucleoside syntheses is that proposed long ago by Krenitsky [41], with uridine or thymidide phosphorylase, acting on uridine or thymidine, serving as a source of r1P or its deoxy analog.
More than 100 natural forms of PNP are known [38]. Various forms of PNP, both natural and modified via the site-directed mutagenesis [39] differ quantitatively or qualitatively in substrate specificities, and may be used for synthetic purposes. There are two broad classes of PNP (i) mammalian (trimeric) forms which are specific towards 6-oxopurine nucleosides [29], but single genetic mutation in the active site, N243D, can alter this specificity [42], and (ii) bacterial forms, which are mostly hexameric and reveal broader substrate specificity, particularly in respect to heteroaromatic moieties of the substrates. Thermophilic PNP variants [43] offer opportunities for large-scale applications. Whole-cell catalysis is also recognized for its relative simplicity and enzyme stabilty [36,39]. Various advanced techniques, including those utilizing enzyme immobilisation on magnetic expoxide microspheres, led to effective syntheses on a gram or even kilogram scale [44]. Analysis of the indutrial potential of such techniques has been recenty presented [39].
PNP, particularly its bacterial forms, is characterized by broad substrate specificity, and this refers both to the heteroaromatic and sugar moieties, as well to various non-typical arrangements of these [38]. Early works pointed unexpected activities of mammalian PNP against N3- and N7- ribosides of canonical purines [45], as well as effective modifications of enzyme specificities via the site-directed mutagenesis of the active-site amino acids [42]. These observations lead to many applications in syntheses of chemically, biochemically, and pharmacologically important nucleoside analogs, like arabinosyl-purines, cordycepin (3-deoxyribosyl-adenine) and analogs, halogenated purine and benzimidazole ribosides and deoxyribosides, including cladribine (2-Chlorodeoxyadenosine), fludarabine (2-fluoro-arabinyl-adenine) arabino- and other nucleosides modified on sugar and/or heteroarometic moieties, reviewed recently [37]. All known PNP forms maintain strict regioselectivity in the ribosylation site (purine N9) and pentose configuration (β), rare exceptions to the former rule are discussed below. Attemtpts to further modify enzyme specificity in a controlled manner are under way [39,43,46].
Another enzymatic systems effecting in deoxynucleoside production are deoxyribosyl transferases (DNT’s) of bacterial origin [36,47]. Analogous synteses of nucleotides from nucleobases are possible using phosphoribosyl tranferases [48].
In the present paper, we summarize recent progress in obtaining new highly fluorescent nucleoside analogs and point their potential for analytical applications. This paper is an update of our previous, more extended review [49].
3. Chemo-Enzymatic Synthesis of Fluorescent Ribosides of Purines and 8-Azapurines
The only strongly fluorescent purine riboside is 2-aminopurine riboside, but this compound is apparently not ribosylated by PNP. Its deoxy analog can be synthesized using DNT as a catalyst [36]. Weak fluorescence is exhibited by 2,6-diaminopurine riboside and N6,N6-dimethyladenosine [49], and these analogs, and the corresponding (deoxy)ribosides, can be synthesized chemo-enzymatically using either PNP from E. coli or bacterial DNT’s as catalysts [36]. Moderately fluorescent 7-methylguanosine and 7-methylinosine undergo almost irreversible phosphorpolysis by both trimeric and hexameric PNP’s [29], so they cannot be synthesized this way. In contrast, the 8-azapurines show much better spectral properties and are substrates for many forms of PNP in the synthetic pathway [49,50].
8-azapurines are isosteric and isomorphic analogs of natural purine bases, substituting the latters in a variety of biochemical transformations [49,50,51,52,53]. Some of them are quite toxic, and to date no direct medical application of these compounds is repoted, but are widely employed in laboratories for various purposes, including mutagenicity tests [51,52]. Their chemical and physicochemical properties differ somewhat from those of the analogous purines, particularly in the acido-basic equillibria and prototropic tautomerism [52]. Nevertheless, they can be classified as probably the most isomorphic purine analogs, although their emissive properties are inferior to those of the “RNA alphabet”.
In contrast to the canonical nucleobases, some of 8-azapurines, as well as the corresponding nucleosides and nucleotides, reveal marked fluorescence in neutral aqueous medium (see refs [49,54], and Table 1). Their nucleotides can be selectively incorporated to mRNA molecules, including ribosymes, and have been applied to mechanistic research of ribozyme action [55,56].
Figure 1.
Purines and 8-azapurines.

3.1. 8-Azaadenosine, 8-Azainosine, 8-Azaxanthosine and Derivatives
Although 8-azaadenine is very weakly fluorescent, and only as a neutral species (pH<6.1), 8-azaadenosine exhibits moderate fluorescence in a broad pH range [50]. Incorporated into ribozyme molecules, it served as a reporter of acid-base equillibria important for the reaction mechanism [55]. As expected, wild-type PNP from E.coli accepts 8-azaadenine as a substrate, but the ribosylation goes about 50-fold slower than that for adenine [50]. Mutations of bacterial PNP sofar were not effective in improving the rate of ribosylation.
8-azainosine (synthesized chemically) was investigated by Seela and co-workers [53], who reported moderate fluorescence of the anionic form of the nucleoside (pKa ~8). 8-Azahypoxanthine reveals poor substrate properties towards PNP in the synthetic pathway, since in the neutral aqueous medium it exists as an anion [53].
8-azaxanthine (8-azaXan) and particularly its methyl derivatives are known to be strongly fluorescent in aqueous medium [57], but the chemo-enzymatic synthesis of 8-azaxanthosine from 8-azaxanthine is difficult, probably because this compound is even more acidic than 8-azaisoguanine (pKa ~4.8, [51]). It has been however reported that the inducible xanthosine phosphorylase from E. Coli, known as PNP-II [58], accepts 8-azaxanthine as a substrate in the ribosylation pathway, although the ribosylation site is cetrainly not N9, but either N7 or N8, as inferred from the spectral properties of the products, strongly fluorescent at 440 nm in the neutral aqueous medium, similarly to those of N7- or N8-methyl, but not to N9-methyl derivatives of 8-azaXan [57].
3.2. 8-Azaguanosine and Analogs
8-Azaguanine is known as a very toxic analog of guanine, which is applied in the mutagenicity tests [59]. It reveals quite intense fluorescence as a neutral species (<pH 7), ascribed to its minor N(8)H tautomer [50]. By contrast, 8-azaguanosine is very weakly fluorescent below pH 6, where it exists in the neutral form, but strongly as a monoanion (pKa~8). This compound is readily, and almost irreversibly, synthesized enzymatically from 8-azaguanine, using calf spleen PNP as a catalyst, and r1P as a ribose donor [50]. It has been applied to mechanistic research of ribozyme action, primarily as a probe of micro-acididity of the active guanine residue of glms ribozyme [56].
The mutated form of calf PNP, N243D, in analogous (slower) reaction gave a mixture of N9- and moderately fluorescent N-7 riboside [60]. By contrast, the bacterial (E.coli) enzyme lead (rather slowly) maily to N9- riboside, with some minor contribution of another strongly fluorescent product, tentatively identified as N8-riboside [60,61].
8-Azaguanine forms weak, but fluorescent complex with calf spleen PNP, which was a subject of spectroscopic investigations [62,63]. Fluorescence excitation spectrum of the complex is markedly shifted to the long wavelength relative to free 8-azaGua. Interestingly, the weakly fluorescent N9-phosphonomethoxy analog of 8-azaguanine binds to this enzyme quite tightely as a bi-substrate analog, also showing marked increase in fluorescence [62]. Somewhat surprisingly, these two complexes are spectrally quite different [63], the fact interpreted in terms of prototropic tautomerism of the bound 8-azaguanine, possibly involving the enol tautomer.
8-Aza-deoxyguanosine has been also synthesized, on a larger scale, using PNP as a catalyst [64], and its fluorescence is reported to be very similar to that of the riboside.
3.3. Other 8-Azapurine Analogs and Their Enzymatic Ribosylation
Although 2,6-diaminopurine is only moderately fluorescent, and its riboside even less [65], their 8-aza analog, 2,6-diamino-8-azapurine (DaaPu) reveals high fluorescence yield in neutral aqueous media [53]. Even stronger emission is observed in some ribosides of DaaPu (Table 1), which could be generated enzymatically using mammalian (calf) or bacterial (E. coli) forms of PNP [61]. The ribosylation site depends in this case qualitatively on catalyst used. Wild-type calf PNP ribosylates DaaPu at N7 and N8, its mutated form (D243N) at N7 and N9, and bacterial (E.coli) enzyme at N9 and N8 [61]. All three ribosides differ markedly by their spectroscopic parameters, particularly fluorescence maxima (spectral data summarized in Table 1). The N9-riboside of DaaPu is probably the most intensely fluorescent nucleoside analog known todate, with yield reaching 90% (Table 1).
The non-typical ribosides of DaaPu, after purification, revealed good and selective substrate properties towards human erythrocytic and bacterial (E.coli) forms of PNP, allowing quantitation of PNP activity in whole human blood, diluted 1000-fold, without necessity of removal of hemoglobin [66]. Mammalian and bacterial forms of PNP can be detected selectively, since N7-riboside of DaaPu is resistant to the E. coli PNP, and N8-riboside is apparently not phosphorolysed by human erythrocytic enzyme [66]. This assay is possible because of marked spectral differences between the above-mentioned ribosides and the nucleobase analog (DaaPu). This selectivity in PNP ribosylation is a subject of molecular modeling investigations [67], which can lead to further broadening of their synthetic possibilities.
The effect of non-typical ribosylation of DaaPu may be kinetic in nature, at least in the case of E.coli PNP, as evidenced by the bi-phasic nature of the reaction progress (unpublished data). Initially, probably kinetically produced N8-riboside is eventually converted to the final thermodynamic product, which is the “canonical” N9-riboside. This reaction progress is easily observed using UV or fluorescence spectroscopy (unpublished data).
It may be also interesting that the phosphonomethoxy analog of DaaPu, PME-DaaPu, prepared by Holy et al. [68], is also intensely fluorescent in near UV and as a bi-substrate analog strongly binds to the mammalian PNP in the absence of phoshate ions [69].
8-Aza-deoxy-isoguanosine has been obtained chemically by Seela and Jiang [70] and polymerized to study oligonucleoside associations. This deoxyriboside is strongly fluorescent, particularly in the anionic form [70]. Unlike many other nucleosides, 8-aza-isoguanosine ribosides are fluorescent also in polymers [70,71], and may serve as reporters of their structure and interactions. Chemo-enzymatic synthesis of 8-aza-isoguanosine is possible, but difficult because of marked acidity of the corresponding purine analog (pK ~5.5, [53]), so at pH>6 only small fraction of molecules is in neutral form. Preliminary experiments (unpublished) conducted in acetate buffer at pH 5.5, at 30℃, showed slow reaction to occur, leading to moderately fluorescent product, but the ribosylation site remains problematic.
3.4. 8-Aza-7-Deazapurine and 8-aza-9-Deazapurine Ribosides
The title compounds (known also as pyrazolo[3,4-d]pyrimidine and pyrazolo[4,3-d]pyrimidine ribosides, respectively) are isomorphic purine analogs, revealig strong biological and pharmaceutical activities, exemplified by allopurinol (4-oxo-pyrazolo[3,4-d]pyrimidine), an ihibitor of xanthine oxidase, widely applied in anti-gout therapy [72]. The corresponding ribosides and deoxyribosides can be synthesized enzymatically using bacterial (E. coli) PNP [64]. This reaction is much slower than that for natural purines, apparently because protonation of purine N7 is essential for the substrate activation by D204 residue [53]. According to Mikhailopulo, this role is in this case taken by Ser90 residue of the E. coli PNP, interacting with puirine N8 [73].
Some of the pyrazolo[3,4-d]pyrimidine ribosides are fluorescent, as indiciated by fluorescence of the commercial nucleobase analog 8-aza-7-deaza-isoguanine (4-amino-6-hydroxyprazolo[3,4-d]pyrimidine), which is a slow substrate of PNP from E.coli (unpublished). Fluorescence changes, observed during the reaction (unpublished) show that at least one of the products must be fluorescent.
The ribosides of 2-aminopurine analog, 8-aza-7-deaza-2-aminopurine, and its 2,6-diaminopurine analog (IUPAC name 4,6-diaminopyrazolo[3,4-d]pyrimidine) were prepared and modified chemically by Seela et al. [74] and enzymatically by Mikhailopulo’s group [64]. Both compounds exhibit measurable fluorescence, which was applied in the polynucleotide studies [74].
8-Aza-9-deazapurine ribosides, known as formycins [75], are C-nucleosides and as such are not phosphorolysed by PNP, but reveal good substrate properties towards other enzymes of purine metabolism and some of them are fluorescent in neutral aqueous media [76,77]. These compounds strongly inhibit hexameric PNP’s [29,78] and their fluorescence was utilized in investigations of enzyme-inhibitor complexes, especially to identify tautomeric forms of the bound ligand(s) [79].
3.5. Other Bicyclic Heteroaromatics
Benzimidazole fluorescence is known from many years, and this compound, and many derivatives, are readily (deoxy)ribosylated by PNP from E. coli [24]. Also halogenated derivatives of benzimidazole were reported to be substrates of PNP [24,80,81]. Another fluorescent isomer, indazole (benzopyrazole), is probably poor or not a substrate. Benzoxazole is apparently not a substrate for the PNP from E. coli [24], but enzymatic ribosylation of 2-aminobenzoxazole is possible, and apparently takes place on the amine group [73]. N1-deaza and N3-deaza purines can be ribosylated by bacterial forms of PNP, but their fluorescence is not known [73].
4. Chemo-Enzymatic Synthesis and Properties of the Tri-Cyclic Purine Analogs and Their Ribosides
Tri-cyclic purine analogs, although cannot be classified as isomorphic with the parent purines, typically combine good spectral characteristics with substrate activities towards many enzymes as well as biological activities [82,83,84]. The simplest way to obtain such analogs is chloroacetyldehyde treatment of amino-group containing purines, the reaction which can be performed in aqueous medium and relatively mild conditions [85]. Some of the “etheno” derivatives are good substrates for various forms of PNP in the synthetic pathway, and their ribosylation leads to strongly fluorescent ribosides, as shown below.
Much more difficult is synthesis of the “extended” nucleobase and nucleoside analogs, containig an aditional phenyl ring between the pyrimidine and imidazole moieties [83]. These “lin-benzo” and other “extended nucleosides” reveal strong fluorescence in the visible range and substrate activities towards some important enzymes of purine metabolism (e.g. adenosine and guanosine deaminase) [83]. Chemistry and biological activity of the tri-cyclic purine analogs have been summarized in recent reviews [85,86].
4.1. Adenosine Analogs
Strongly fluorescent 1,N6-ethenoadenosine (εAdo) is easily generated chemically from the parent adenosine by treatment with chloroacetaldehyde in aqueous medium at room temperature [82]. Although not susceptible to phosphorolysis by calf PNP, it can be readily phosphorolysed to 1,N6-ethenoadenine (εAde) by the bacterial (E. Coli) enzyme [87]. This latter reaction is revesible, but in the synthetic pathway, HPLC analysis revealed minor contribution of the adenine rybosylated at N7. By contrast, the ribosylation of etheno-adenine base by calf PNP as a catalyst and r1P as a ribose source provides quite effectively a non-typical, and moderately fluorescent N6-riboside while the mutated form of calf PNP (N242D) provided mixture of N6- and N9- ribosides [87]. The N6-riboside is only moderately fluorescent (Table 2), but can be useful for detection of PNP activity in biological or clinical material, including diluted whole blood, since it is rapidly phosphorolysed by the human erythrocytic PNP to strongly fluorescent 1,N6-ethenoadenine [66].
4.2. Guanosine Analogs
Two isomeric ethenoguanosine derivatives (Figure 2) are generated from the parent guanosine by treatment with chloroacetaldehyde in aqueous medium [88]. This reaction is slower than that of adenosine, so several different strategies were proposed to achieve this goal [89]. One of the etheno-guanine isomers (the linear 1,N2-ethenoguanine) is a good substrate for bacterial (E. coli), but not for calf PNP, providing poorly fluorescent nucleoside (Table 2). The second isomer, N2,3-ethenoguanine, is nonfluorescent and resistant to ribosylation by PNP [90].
Somewhat stronger fluorescence (yield 0.11, see Table2) was detected from N2,3-etheno-6-O-methylguanosine [90], which can be enzymatically generated from the base analog by the E. coli PNP, but in this case the ribosylation rate was ~300-fold slower than that for guanosine.
4.3. 2-Aminopurine Riboside and Isoguanosine Analogs
The reaction of chloroacetaldehyde with 2-aminopurine riboside is rapid and gives essentially one product, identified as 1,N2-etheno-2-aminopurine-N9-β-D-riboside [91]. This compound is only moderately fluorescent, but the analogous reaction with 2-aminopurine base yielded two products, identified as 1,N2-etheno-2-aminopurine (~95%) and N2,3-etheno-2-aminopurine(~5%), the latter showing fluorescence yield >70% [92]. The minor product was found to be good substrate of both calf PNP and E.coli PNP , giving fluorescent riboside products [92]. Much slower reaction was observed for the “linear” 1,N2-etheno-2-aminopurine with both enzymes. Somewhat unexpectedly, the ribosylation takes place on N2 nitrogen, rather then the “canonical” N9 [92]. The purified riboside of N2,3-etheno-2-aminopurine is rapidly phosphorolysed by calf and human erythrocytic PNP, and the reaction is easily followed fluorimetrically thanks to marked spectral difference between substrate and product [92]. Applications of these compounds may be somewhat uncomfortable because of their sensitivity to air oxidation. Another possible application of the “linear” etheno-2-aminopurine derivative may be in fluorimetric detection of xanthine oxidase activity (in preparation).
Isoguanine (2-oxo-adenine), which is an almost nonfluorescent product of adenine radiolysis [93], reacts with chloroacetaldehyde fairly rapidly, giving essentially one product, identified as 1,N6-etheno-isoguanine or 1,N6-etheno-2-oxo-adenine [90], moderately fluorescent in neutral aqueous medium (Table 2). This compound is readily ribosylated by the E. coli PNP, and more slowly by the calf enzyme [90], with r1P as a ribose donor. There are at least three products of such ribosylation, all of them fluorescent, with fluorescence quantum yield reaching 0.66 (see Table 2) for the compound identified as 1,N6-etheno-2-oxo-adenine-6-β-D-riboside [90], and this was the main product when the reaction was catalyzed by calf PNP (wild type).
Attemps to obtain highly fluorescent nucleobase analogs from 2,6-diaminopurine were not sucessful [94], but our preliminary data indicate that some of the related ribosides are in fact intensely fluorescent (unpublished).
4.4. Tri-Cyclic aza-Purine Analogs
Enzymatic phosphorolysis of the moderately fluorescent 2-aza analog of εAdo, prepared originally by Tsou et al. [95], has been reported [49], but the reverse reaction was not examined.
Some other tri-cyclic 2-aza-purine, 8-aza-purine and 2,8-diazapurine nucleoside analogs were synthesized chemically, with results summarized in ref. [96]. The reported fluorescence quantum yields are lower than those obtained for purine analogs, but the reported spectra are batochromically shifted relative to those of εAdo [96]. To our knowledge, no data on enzymatic hydrolysis of phosphorolysis of these analogs exist.

Table 2.
Fluorescence parameters of selected “etheno” substituted purine ribosides in water. Unless otherwise indicated, all data refer to neutral species of the fluorophores. Data from references [82,87,90,92]; nd = no data.
| Compound | excitation: λmax [nm] |
emission: λmax [nm] |
φ | τ [ns] |
|---|---|---|---|---|
| 1,N6-etheno-adenosine | 305 | 410 | 0.56 | 21 |
| 1,N6-etheno-6-β-D-ribosyl-adenine | 310 | 380 | 0.10 | nd |
| 1,N2-ethenoguanosine (anion) | 400 | <0.01 | nd | |
| N2,3-ethenoguanosine | 261 | 400 | 0.02 | nd |
| N2,3-etheno-O6-methylguanosine | 272 | 405 | 0.11 | nd |
| 1,N2-etheno-2-aminopurine | 248 | 473 | 0.18 | 6.9;10.3 |
| 1,N2-etheno-2-aminopurine-9-β-D- riboside |
295 | 463 | 0.14 | 3.8;8.5 |
| 1,N2-etheno-2-aminopurine-2-β-D- riboside |
338 | 406 | 0.73 | nd |
| N2,3-etheno-2-aminopurine | 315 | 357 | 0.29 | 2.15 |
| N2,3-etheno-2-aminopurine-2-β-D- riboside |
315 | 365 | 0.29 | nd |
| 1,N6-etheno-2-oxo-adenine | 295 | 415 | 0.17 | nd |
| 1,N6-etheno-2-oxo-adenosine | 295 | 415 | 0.34 | 6.1 |
| 1,N6-etheno-2-oxo-adenine-7-β-D- riboside |
294 | 360 | 0.036 | 0.8;5.2 |
| 1,N6-etheno-2-oxo-adenine-6-β-D- riboside |
303 | 425 | 0.66 | nd |
5. Conclusion and Perspectives
This brief review illustrates possibility of rapid and effective syntheses of many strongly fluorescent ribosides, which can be used in future biophysical and analytical research. The presented examples involve various forms of PNP used as catalysis, and illustrate possibilities of modifying enzyme specificity via site-directed mutagenesis not only in quantitative, but sometimes in qualitative sense. Variability in the ribosylation sites, exhibited by PNP in some analogs, is helpful for preparation of ribosides with better spectral characteristics, suitable, e. g., for fluorescence microscopy. This variabilty was not observed for canonical purines, with the only exception of recently reported synthyesis of xanthine-N7-β-D-riboside [97].
Efforts to modify specificity of PNP using various genetic modifications are published [98]. In the future, specific enzymes may be designed for specific substrates to be ribosylated at specific sites, and theoretical methods potentially leading to this goal are being developed [99]. Sensitive and specific fluorimetric assays for PNP, briefly presented above, can be regarded as “bonuses” of this kind of research.
It would be interesting to examine analogous possibilities of other enzymatic systems, for example phosphoribolsyl transferases, catalysing formation of nucleotides from nucleobases and 5-phosphoribose-1-pyrophosphate (PrPP), and extensively investigated because of their medical significance [100,101]. Various bacterial ezymatic systems, like deoxy-riboside transferases, phosphoribosyl transferases and/or nucleoside hydrolases (acting in non-aqueous media) may be also useful but their specificities towards fluorescent or fluorogenic analogs are mostly unknown.
Supplementary Materials
- not applicable
Author Contributions
Conceptualization, J.W. and A.S.-W..; methodology, J.W. and A.S.-W.; software, A.S.-W.; validation, J.W., A.S.-W.; formal analysis, J.W.; investigation, J.W. and A.S.-W.; resources, J.W.; data curation, J.W.; writing—original draft preparation, J.W.; writing—review and editing, J.W. and A.S.-W.; visualization, J.W. and A.S.-W.; supervision, J.W.; project administration, J.W.; funding acquisition, A.S.-W. All authors have read and agreed to the published version of the manuscript.
Funding
This workwas supported by the University of Warmia & Masury in Olsztyn, internal grant # 17.610.011-110.
Institutional Review Board Statement
- not applicable
Informed Consent Statement
- not applicable
Data Availability Statement
-
Acknowledgments
We thank Professor Beata Wielgus-Kutrowska from Warsaw University for valuable discussions and support.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Lakowicz, J. Principles of fluorescence spectroscopy, 3rd Edition. Springer, New York, 2006.
- Zalejski, J.; Sun, J.; Sharma, A. Unravelling the Mystery inside Cells by Using Single-Molecule Fluorescence Imaging. J. of Imaging 2023, 9, art#192. [Google Scholar] [CrossRef]
- Adhikari, S; Smit, R and Orrit, M. Future Paths in Cryogenic Single-Molecule Fluorescence Spectroscopy. J. Physical Chem. C, 2023, 128, 3-18. DOI:10.1021/acs.jpcc.3c06564; b) Wang, D.; Shalamberidze, A.; Arguello, A. E.; Purse, B. W.; Kleiner, R. E. Live-Cell RNA Imaging with Metabolically Incorporated Fluorescent Nucleosides. J. Am. Chem. Soc. 2022, 144, 14647–14656, DOI: 10.1021/jacs.2c04142.
- Wilhelmsson M, Tor Y. Fluorescent Analogues of Biomolecular Building Blocks: Design and Applications. John Wiley & Sons; New York, USA, 2016.
- Xu, W.; Ke, M.C.; Kool, E.T. Fluorescent nucleobases as tools for studying DNA and RNA. Nature Chem. 2017, 9, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Tanpure, A.A.; Pawar, G.; Srivatsan, S.G. Fluorescent Nucleoside Analogs: Probes for Investigating Nucleic Acid Structure and Function. Israel Journal of Chemistry 2013, 53, 366–378. [Google Scholar] [CrossRef]
- Hocek, M. Enzymatic Synthesis of Base-Functionalized Nucleic Acids for Sensing, Cross-linking, and Modulation of Protein–DNA Binding and Transcription. Accounts of Chemical Research 2019, 52, 1730–1737. [Google Scholar] [CrossRef]
- Michel, B.Y.; Dziuba, D.; Benhida, R.; Demchenko, A.P.; Burger, A. Probing of Nucleic Acid Structures, Dynamics, and Interactions With Environment-Sensitive Fluorescent Labels. Front. Chem. 2020, 8, 112. [Google Scholar] [CrossRef]
- a) Xu, Wang; Ke, Min Chan; Kool, E.T. Fluorescent nucleobases as tools for studying DNA and RNA. Nature Chem. 2017, 9, 1043–1055. https://doi.org/10.1038/nchem.2859 b) Wang, D.; Shalamberidze, A.; Arguello, A. E.; Purse, B. W.; Kleiner, R. E. Live-Cell RNA Imaging with Metabolically Incorporated Fluorescent Nucleosides. J. Am. Chem. Soc. 2022, 144, 14647– 14656, DOI: 10.1021/jacs.2c04142.
- Saito, Y.; Hudson, R.H.E. Base-modified fluorescent purine nucleosides and nucleotides for use in oligonucleotide probes. J. Photochem. Photobiol. C: Photochemistry Reviews 2018, 36, 48–73. [Google Scholar] [CrossRef]
- Singh, H.; K Tiwari, R. Tiwari, S.K. Pramanik, A. Das. Small molecules as fluorescent probes for monitoring intracellular enzymatic transformations. Chemical Revs. 2019, 119, 11718-11760. [CrossRef]
- Gilbault, G.G. in: Practical Fluorescence, editor: G. G. Gilbault, Marcel Dekker, New York, 1990, Chapter 12.
- Callis, P. R. Electronic States and Luminescence of Nucleic Acids Systems. Annu. Rev. Phys. Chem. 1983, 34, 329–357. [Google Scholar] [CrossRef]
- Leng, M.; Pochon, F.; Michelson, M. Photochemistry of polynucleotides.2. Mononucleotide and dinucleotide fluorescence at ordinary temperatures. Biochim Biophys Acta 1968, 169, 338–349. [Google Scholar] [CrossRef]
- Ward, D. C.; Reich, E.; Stryer, L. Fluorescence Studies of Nucleotides and Polynucleotides. 1. Formycin, 2-aminopurine riboside, 2,6-diaminopurine riboside and their derivatives. J. Biol. Chem. 1969, 244, 1228–1237. [Google Scholar] [CrossRef]
- Jones, A. C.; Neely, R. K. 2-aminopurine as a fluorescent probe of DNA conformation and the DNA–enzyme interface. Q. Rev. Biophys. 2015, 48, 244–279. [Google Scholar] [CrossRef]
- Kirk, S. R.; Luedtke, N. W.; Tor, Y. 2-Aminopurine as a Real-Time Probe of Enzymatic Cleavage and Inhibition of Hammerhead Ribozymes. Bioorg. Med. Chem. 2001, 9, 2295–2301. [Google Scholar] [CrossRef]
- P. Herdewijn, Ed. Modified nucleosides in biochemistry, biotechnology and medicine. Wiley-VCH, New York, 2008.
- Sinkeldam, R. W.; Greco, N. J.; Tor, Y. Fluorescent Analogs of Biomolecular Building Blocks: Design, Properties and Applications. Chem. Rev. 2010, 110, 2579–2619. [Google Scholar] [CrossRef]
- Okamoto, A.; Saito, Y.; Saito I. Design of base-discriminating fluorescent nucleosides. J. Photochem. Photobiol., C 2005, 6, 108–122. [CrossRef]
- Tor, Y. Isomorphic Fluorescent Nucleosides. Acc. Chem. Res. 2024, 57(9), 1325–1335. [Google Scholar] [CrossRef]
- Ludford, P.; Yang, Shengua; Bucardo, M.S.; Tor, Y. A New Variant of Emissive RNA Alphabets. Chemistry Europ. J. 2022, 28, e202104472. [CrossRef]
- Dziuba, D.; Didier, P.; Ciaco, S.; Barth, A.; Seidel, C.A.M.; Mély, Y. Fundamental photophysics of isomorphic and expanded fluorescent nucleoside analogues. Chemical Society Reviews 2021, 50, 7062–7107. [Google Scholar] [CrossRef] [PubMed]
- Mikhailopulo, A.I.; Miroshnikov, A.I. Biologically important nucleosides: Modern trends in biotechnology and application. Mendeleev Comm. 2011, 21, 57–68. [Google Scholar] [CrossRef]
- Cosgrove, S.C.; Miller, G.J. Advances in biocatalytic and chemoenzymatic synthesis of nucleoside analogues. Expert Opinion on Drug Discovery 2022, 17, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, P. A.; Sinkeldam, R. W.; Tor, Y. Visibly Emissive and Responsive Extended 6-Aza-Uridines. Org. Lett. 2014, 16, 5290– 5293. [CrossRef]
- Ardhapure, A.V.; Gayakhe, V.; Bhilare, S.; Kapdi, A.R.; Bag, S.S.; Sanghvi, Y.S.; Gunturu, K.C. Extended fluorescent uridine analogues: synthesis, photophysical properties and selective interaction with BSA protein. New J. Chem. 2020, 44, 14744–14754. [Google Scholar] [CrossRef]
- Greco, N. J.; Tor, Y. Simple Fluorescent Pyrimidine Analogs Detect the Presence of DNA Abasic Sites. J. Am. Chem. Soc. 2005, 127, 10784–10785. [Google Scholar] [CrossRef]
- Bzowska A, Kulikowska E. Shugar D. Purine nucleoside phosphorylases: Properties, functions, and clinical aspects. Pharmacol. Therap. 2000, 88, 349-425. [CrossRef]
- Rodwell, V.W. Metabolism of purine and pyrimidine nucleotides. In: Harper’s Illustrated Biochemistry, 30-th edition, McGraw & Hill, 2008, pp. 347-358.
- a) Grunebaum, E.; Cohen, A.; Roifman, C.M. Recent advances in understanding and managing adenosine deaminase and purine nucleoside phosphorylase deficiencies. Curr. Opin. Allergy Clin. Immunol. 2013, 13, 630–638. DOI: 10.1097/ACI.0000000000000006; b) Fox, T.A.; Booth, C. Gene therapy for primary immunodeficiencies. British Journ. Haematol. 2021, 193, 1044-1059. DOI: 10.1111/bjh.17269.
- Ardiani, A.; Johnson, A.J.; Ruan, H.M.; Sanchez-Bonilla, M.; Serve, K.; Black, M.E. Enzymes To Die For: Exploiting Nucleotide Metabolizing Enzymes for Cancer Gene Therapy. Current Gene Therap. 2012, 12, 77–91. [Google Scholar] [CrossRef]
- Schramm, V.L. Enzymatic Transition States and Drug Design. Chem. Revs. 2018, 118, 11194–11258. [Google Scholar] [CrossRef]
- a) Lewis, D.J.; Duvic, M. Forodesine in the treatment of cutaneous T-cell lymphoma. Expert Op Investig Drugs 2017, 26, 771-775. DOI: 10.1080/13543784.2017.1324569; b) Makita S.; Maeshima A.M.; Maruyama D.; Izutsu K.; Tobinai K. "Forodesine in the treatment of relapsed/refractory peripheral T-cell lymphoma: an evidence-based review". OncoTargets & Therapy 2018, 11, 2287–2293. doi:10.2147/OTT.S140756. PMC 5916385. PMID 29719411.
- Ho, M.C.; Shi, W.; Rinaldo-Matthis, A.; Tyler, P.C.; Evans, G.B.; Clinch, K.; Almo, S.C.; Schramm, V.L. Four generations of transition-state analogues for human purine nucleoside phosphorylase. Proc. Natl. Acad. Sci. USA 2010, 107, 4805–4812. [Google Scholar] [PubMed]
- Mikhailopulo, I.A. Biotechnology of Nucleic Acid Constituents – State of the Art and Perspectives. Curr. Org. Chem. 2007, 11, 317–335. [Google Scholar] [CrossRef]
- Kamel, S.; Yehia, H.; Neubauer, P.; Wagner, A. Enzymatic Synthesis of Nucleoside Analogues by Nucleoside Phosphorylases. In: Enzymatic and Chemical Synthesis of Nucleic Acid Derivatives; Fernandez-Lucas, J., Camarasa-Rius, M.J., Eds.; Wiley-VCH, New York, 2019. Chapter 1.
- Yehia, H.; Kamel, S.; Paulick, K.; Neubauer, P.; Wagner, A. Substrate Spectra of Nucleoside Phosphorylases and their Potential in the Production of Pharmaceutically Active Compounds. Curr. Pharm. Des. 2017, 23, 6913–6935. [Google Scholar] [CrossRef]
- Westarp, S.; Kaspar, F.; Neubauer, P.; Kurreck, A. Industrial potential of the enzymatic synthesis of nucleoside analogs: existing challenges and perspectives. Current Op. Biotech. 2022, 78, 102829. [Google Scholar] [CrossRef]
- Hennen, W.J.; Wong, C.-H. A new method for the enzymic synthesis of nucleosides using purine nucleoside phosphorylase. J. Org. Chem. 1989, 4692–4695. [Google Scholar] [CrossRef]
- Krenitsky, T.A.; Koszalka, G.W.; Tuttle, J.V. Purine nucleoside synthesis: An efficient method employing nucleoside phosphorylases. Biochemistry 1981, 20, 3615–3621. [Google Scholar] [CrossRef]
- Stoeckler, J.D.; Poirot, A.F.; Smith, R.M.; Parks, R.E., Jr.; Ealick, S.E.; Takabayashi, K.; Erion, M.D. Purine-nucleoside phosphorylase. 3. Reversal of purine base specificity by site-directed mutagenesis. Biochemistry1997, 36, 11749–11756. [CrossRef]
- Kamel, S.; Thiele, I.; Neubauer, P., Wagner, A. Thermophilic nucleoside phosphorylases: Their properties, characteristics and applications. Biochim. Biophys. Acta-Proteins Proteomics 2020, 1868, 140304. [CrossRef]
- Zhou, X.R.; Mikhailopulo, I.A.; Bournazou, M.N.C.; Neubauer, P. Immobilization of thermostable nucleoside phosphorylases on MagReSyn® epoxide microspheres and their application for the synthesis of 2,6-dihalogenated purine nucleosides. J. Mol. Catal. B 2015, 115, 119-127. [CrossRef]
- a) Bzowska, A.; Kulikowska, E.; Poopeiko, N.E.; Shugar, D. Kinetics of phosphorolysis of 3-(β-D-ribofuranosyl)adenine and 3-(β-D-ribofuranosyl)hypoxanthine, non-conventional substrates of purine-nucleoside phosphorylase. Eur. J. Biochem. 1996, 239, 229–234. DOI: 10.1111/j.1432-1033.1996.0229u.x; b) Bzowska, A.; Ananiev, A.V.; Ramzaeva, N.; Alksnis, E.; Maurins, J.A.; Kulikowska, E.; Shugar, D. Purine nucleoside phosphorylase: Inhibition by purine N(7)- and N(9)-acyclonucleosides; and substrate properties of 7-β-D-ribofuranosylguanine and 7-β-D-ribofuranosylhypoxanthine. Biochem. Pharmacol. 1994, 48, 937–947.
- Fernandez-Lucas, J. Multienzymatic synthesis of nucleic acid derivatives: a general perspective. Appl Microbiol Biotechnol 2015, 99, 4615–4627. [CrossRef]
- Del Arco, J.; Acosta, J.; Fernandez-Lucas, J. New trends in the biocatalytic production of nucleosidic active pharmaceutical ingredients using 2’-deoxyribosyl transferases. Biotechnol. Advances 2021, 51, 107701. [Google Scholar] [CrossRef] [PubMed]
- Del Arco, J.; Fernandez-Lucas, J. Purine and pyrimidine phosphoribosyl transferases: a versatile tool for enzymatic synthesis of nucleoside-5’-mono phosphates, Curr. Pharm. Des. 2017, 23, 6898 – 6912. [CrossRef]
- Wierzchowski J., Stachelska-Wierzchowska, A., Wielgus-Kutrowska, B. Bzowska, A. 1,N6-ethenoadenine and other fluorescent nucleobase analogues as substrates for purine-nucleoside phosphorylases: spectroscopic and kinetic studies. Curr. Pharmaceut. Design 2017, 23, 6972-6990. [CrossRef]
- Wierzchowski, J.; Wielgus-Kutrowska, B.; Shugar, D. Fluorescence emission properties of 8- azapurines and their nucleosides, and application to the kinetics of the reverse synthetic reaction of PNP. Biochim Biophys Acta 1996, 1290, 9–17. [Google Scholar] [CrossRef]
- Albert, A. Chemistry of 8-azapurines. Adv Heterocycl Chem 1986, 39, 117–178. [Google Scholar] [CrossRef]
- Giorgi, I.; Scartoni, V. 8-Azapurine nucleus: a versatile scaffold for different targets. Mini-Reviews Medicinal Chemistry 2009, 9 1367–1378. [CrossRef]
- Wierzchowski J., Antosiewicz J.M., Shugar D. 8-Azapurines as isosteric purine fluorescent probes for nucleic acid and enzymatic research. Molecular BioSystems 2014, 10, 2756-2774. [CrossRef]
- Seela F., Jawalekar A.M., Münster, I. Replacement of Canonical DNA Nucleobases by Benzotriazole and 1,2,3-Triazolo[4,5-d]pyrimidine: Synthesis, Fluorescence, and Ambiguous Base Pairing. Helvetica Chimica Acta 2005, 88, 751-765. [CrossRef]
- Cottrell, J.W, Scott, L.G., Fedor, M.J. The pH dependence of hairpin ribozyme catalysis reflects ionization of an active site adenine, J. Biol. Chem. 2011, 286, 17658–17664. [CrossRef]
- Viladoms, J., Scott, L.G., Fedor, M.J. An active-site guanine participates in glms ribozyme catalysis in its protonated state. J. Amer. Chem. Soc. 2011, 133, 18388–18396. [CrossRef]
- a) Wierzchowski, J.; Sepiol, J.; Sulikowski, D.; Kierdaszuk, B.; Shugar, D. Fluorescence emission properties of 8-azaxanthine and its N-alkyl derivatives: Excited-state proton transfer, and potential applications in enzymology. J. Potochem. Photobiol. A-Chemistry 2006, 179, 276-282. DOI10.1016/j.jphotochem.2005.08.027; b) Wierzchowski, J.; Smyk, B. Excited-State Proton Transfer in 8-Azapurines I: A Kinetic Analysis of 8-Azaxanthine Fluorescence. Molecules 2020, 25, #2740.
- Dandanell, G.; Szczepanowski, R.H.; Kierdaszuk, B.; Shugar, D.; Bochtler, M. Escherichia coli purine nucleoside phosphorylase II, the product of the xapA gene. J. Mol. Biol. 2005, 348, 113–125. [Google Scholar] [CrossRef]
- Albertini, R.J. Albertini, R.J. HPRT mutations in humans: biomarkers for mechanistic studies. Mutat. Res. 489, 1-16. Mutation Res., 2001, 489, 1–16. [CrossRef]
- Stachelska-Wierzchowska, A; Wierzchowski, J; Wielgus-Kutrowska B.; Mikleusevic, G. Enzymatic Synthesis of Highly Fluorescent 8-Azapurine Ribosides Using a Purine Nucleoside Phosphorylase Reverse Reaction: Variable Ribosylation Sites. Molecules 2013, 18, 12587–12598. [CrossRef]
- Stachelska-Wierzchowska, A.; Wierzchowski, J.; Bzowska, A.; Wielgus-Kutrowska, B. Site-selective ribosylation of fluorescent nucleobase analogs using purine-nucleoside phosphorylase as a catalyst: effects of point mutations. Molecules 2016, 21: 44. [CrossRef]
- Wierzchowski, J.; Bzowska, A.; Stępniak, K.; Shugar, D. Interactions of Calf Spleen Purine Nucleoside Phosphorylase with 8-Azaguanine, and a Bisubstrate Analogue Inhibitor: Implications for the Reaction Mechanism. Z. Naturforsch C. 2004, 59, 713–725. [Google Scholar] [CrossRef]
- Wierzchowski, J.; Stępniak, K.; Bzowska, A.; Shugar, D. Spectroscopic and kinetic studies of interactions of calf spleen purine nucleoside phosphorylase with 8-azaguanine and its 9-(2-phosphonylmethoxyethyl) derivative. Nucleosides, Nucleotides & Nucl Acids 2005, 24, 459-464. [CrossRef]
- Stepchenko, V.A.; Seela, F.; Esipov, R.S.; Miroshnikov, A.I.; Sokolov, Y.A.; Mikhailopulo, I.A. Enzymatic Synthesis of 2’-Deoxy-β-D-ribonucleosides of 8-Azapurines and 8-Aza-7-deazapurines. Synlett 2012, 10, 1541-45. [CrossRef]
- Wierzchowski, J.; Ogiela, M.; Iwańska, B.; Shugar, D. Selective fluorescent and fluorogenic substrates for purine-nucleoside phosphorylases from various sources, and direct fluorimetric determination of enzyme levels in human and animal blood. Analytica Chimica Acta 2002, 472, 63–74. [Google Scholar] [CrossRef]
- Stachelska-Wierzchowska, A.; Wierzchowski, J. Non-typical nucleoside analogs as fluorescent and fluorogenic indicators of purine-nucleoside phosphorylase activity in biological samples. Analytica Chimica Acta 2020, 1139, 119–128. [Google Scholar] [CrossRef]
- Pyrka, M.; Maciejczyk, M. Why Purine Nucleoside Phosphorylase Ribosylates 2,6-Diamino-8-azapurine in Noncanonical Positions? A Molecular Modeling Study. J. Chem. Inf. Model. 2020, 60, 1595–1606. [Google Scholar] [CrossRef]
- a) Holy, A. Phosphonomethoxyalkyl analogs of nucleotides. Curr. Pharm. Design 2003, 9, 2567-2592; DOI: 10.2174/1381612033453668 b) De Clercq, E.; Holy, A. Acyclic nucleoside phosphonates: A key class of antiviral drugs. Nature Revs Drug Disc. 2005, 4, 928-940. https://doi.org/10.1038/nrd1877.
- Wierzchowski, J.; Kulikowska, E.; Bzowska, A.; Holy, A.; Magnowska, L.; Shugar, D. Interactions of purine nucleoside phosphorylase with antiviral acyclic nucleoside phosphonate inhibitors – kinetics and emission studies. Nucleosides & Nucleotides 1999, 18, 875-876. [CrossRef]
- Jiang, D.W.; Seela, F. Oligonucleotide Duplexes and Multistrand Assemblies with 8-Aza-2′-deoxyisoguanosine: A Fluorescent isoGd Shape Mimic Expanding the Genetic Alphabet and Forming Ionophores. J. Amer. Chem. Soc. 2010, 132, 4016–4024. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Jiang, D.W.; Schäfer, A.H.; Seela, F. 8-Aza-2′-deoxyisoguanosine Forms Fluorescent Hydrogels whereas 8-Aza-2′-deoxyguanosine Assembles into Nucleoside Nanotubes. ChemPlusChem 2017, 82, 778-784. [CrossRef]
- Emmerson, B.T. Drug therapy - The management of gout. New England J. Med. 1996, 334, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Fateev, I.V.; Kharitonova, M.I.; Antonov, K.V.; Konstantinova, I.D.; Stepanenko, V.N.; Esipov, R.S.; Seela, F.; Temburnikar, K.W.; Seley-Radtke, K.L.; Stepchenko, V.A.; et al. Recognition of Artificial Nucleobases by E. coli Purine Nucleoside Phosphorylase versus its Ser90Ala Mutant in the Synthesis of Base-Modified Nucleosides. Chem. Eur. J. 2015, 21, 13401–13419. [CrossRef]
- Seela, F.; Becher, G. Synthesis, base pairing, and fluorescence properties of oligonucleotides containing 1H-pyrazolo[3,4-d]pyrimidin-6-amine (8-aza-7-deazapurin-2-amine) as an analogue of purin-2-amine. Helvetica Chim. Acta 2000, 83, 928-942. https://doi.org/10.1002/(SICI)1522-2675(20000510)83:5<928::AID-HLCA928>3.0.CO;2-5; b) Kondhare, D.; Leonard, P.; Seela, F. Isoguanine (2-Hydroxyadenine) and 2-Aminoadenine Nucleosides with an 8-Aza-7-deazapurine Skeleton: Synthesis, Functionalization with Fluorescent and Clickable Side Chains, and Impact of 7-Substituents on Physical Properties. J.Org. Chem. 2021, 86, 14461-14475. DOI10.1021/acs.joc.1c01283.
- De Clercq, E. C-Nucleosides To Be Revisited. J. Med. Chem. 2016, 59, 2301–2311. [Google Scholar] [CrossRef]
- Wierzchowski, J.; Shugar, D. Luminescence studies on formycin, its aglycone, and their N-methyl derivatives: tautomerism, sites of protonation and phototautomerism. Photochem. Photobiol. 1982, 35, 445–458. [Google Scholar] [CrossRef]
- Karlish, S.J.D. Use of formycin nucleotides, intrinsic protein fluorescence, and... FLUORESCEIN ISOTHIOCYANATE-labeled enzymes for measurement of conformational states of NA+,K+-ATPase. Methods Enzymol. 1988, 156, 271-277. [CrossRef]
- Kierdaszuk, B.; Modrak-Wojcik, A.; Wierzchowski, J.; Shugar, D. Formycin A and its N-methyl analogues, specific inhibitors of E-coli purine nucleoside phosphorylase (PNP): induced tautomeric shifts on binding to enzyme, and enzyme -> ligand fluorescence resonance energy transfer. Biochim. Biophys. Acta - Protein Struct. Mol. Enzymol. 2000, 1476, 109-128. [CrossRef]
- Wlodarczyk, J.; Galitonov, G.S.; Kierdaszuk, B. Identification of the tautomeric form of formycin A in its complex with Escherichia coli purine nucleoside phosphorylase based on the effect of enzyme-ligand binding on fluorescence and phosphorescence. Europ. Biophys. J. Biophys. Lett. 2004, 33, 377-385. [CrossRef]
- Konstantinova, I.D.; Selezneva, O.M.; Fateev, I.V.; Balashova, T.A.; Kotovskaya, S.K.; Baskakova, Z.M.; Charushin, V.N.; Baranovsky, A.V.; Miroshnikov, A.I.; Balzarini, J.; Mikhailopulo, I.A. Chemo-Enzymatic Synthesis and Biological Evaluation of 5,6-Disubstituted Benzimidazole Ribo- and 2′-Deoxyribonucleosides. Synthesis-Stuttgart 2013, 45, 272-280. [CrossRef]
- Kharitonova, M.I.; Fateev, I.V.; Kayushin, A.L.; Konstantinova, I.D.; Kotovskaya, S.K.; Andronova, V.L.; Galegov, G.A.; Charushin, V.N.; Miroshnikov, A.I. Chemoenzymatic Synthesis and Antiherpes Activity of 5-Substituted 4,6-Difluorobenzimidazoles Ribo- and 2′-Deoxyribonucleosides. Synthesis-Stuttgart 2016, 48, 394-406. [CrossRef]
- Leonard, N.J. Etheno-substituted nucleotides and coenzymes: Fluorescence and biological activity. Crit. Revs. Biochem. 1984, 15, 125–199. [Google Scholar] [CrossRef] [PubMed]
- Leonard, N.J. Adenylates: bound and unbound. Biopolymers 1985, 24, 9–28. [Google Scholar] [CrossRef] [PubMed]
- Secrist, J. A.; Barrio, J. R.; Leonard, N. J.; Weber, G. Fluorescent modifications of adenosine containing coenzymes. Biological activities and spectroscopic properties. Biochemistry 1972, 11, 3499– 3506. [CrossRef]
- Jahnz-Wechmann, Z.; Framski, G.R.; Januszczyk, P.A.; Boryski, J. Bioactive fused heterocycles: Nucleoside analogs with an additional ring. Eur. J. Med. Chem. 2015, 97, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Jahnz-Wechmann, Z.; Framski, G.R.; Januszczyk, P.A.; Boryski, J. Base-modified nucleosides: Etheno derivatives. Frontiers Chem. 2016, 4, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Stachelska-Wierzchowska, A.; Wierzchowski, J.; Bzowska, A.; Wielgus-Kutrowska, B. Tricyclic nitrogen base, 1,N6-ethenoadenine, and its ribosides, as substrates for purine-nucleoside phosphorylases: spectroscopic and kinetic studies. Nucleos. Nucleot. Nucleic Acids 2018, 37, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Kuśmierek, J.T.; Jensen, D.E.; Spengler, S.J.; Stolarski, R.; Singer, B. Synthesis and properties of N2,3-ethenoguanosine and N2,3-erthenoguanosine 5’diphosphate. J. Org. Chem. 1987, 52, 2374–2378. [Google Scholar] [CrossRef]
- Boryski, J. 1,N-2-ethenoguanosine – 3 methods of synthesis. Nucl. Nucleot. 1990, 9, 803–813. [Google Scholar] [CrossRef]
- Stachelska-Wierzchowska, A.; Wierzchowski, J.; Górka, M.; Bzowska, A.; Wielgus-Kutrowska, B. Tricyclic nucleobase analogs and their ribosides as substrates of purine-nucleoside phosphorylases. II. Guanine and isoguanine derivatives, Molecules 2019 24, 1493. [CrossRef]
- Virta, P.; Holmstrom, T.; Roslund, M.U.; Mattjus, P.; Kronberg, L.; Sjoholm, R. New nucleoside analogs from 2-amino-9-(β-d-ribofuranosyl)-purine. Org. Biomol. Chem. 2004, 2, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Stachelska-Wierzchowska, A.; Wierzchowski, J.; Górka, M.; Bzowska, A.; Stolarski, R.; Wielgus-Kutrowska, B. Tricyclic nucleobase analogs and their ribosides as substrates and inhibitors of purine-nucleoside phosphorylases III. Aminopurine derivatives, Molecules 2020, 25, 681. [CrossRef]
- Von Sonntag, C.; Schuchmann, H.P. Radical-mediated damege of DNA in the presence of oxygen. Methods. Enzymol. 1990, 186, 511–520. [Google Scholar] [CrossRef]
- Horejsí, K; Pohl, R and Holy, A. Tricyclic purine analogs derived from 2-amino-6-chloropurine and 2,6-diaminopurine and their methylated quaternary salts. Coll. Czechosl. Chem. Comm. 2006, 71, 77-90. [CrossRef]
- Tsou, K.C.; Yip, K.-F., Miller, E.E., Lo, K.W. Synthesis of l,N6-etheno-2-aza-adenosine (2-aza-ε-adenosine): a new cytotoxic fluorescent nucleoside. Nucleic Acid Res. 1974, 1, 518-538. [CrossRef]
- Budow, S.; Seela, F. 2-Azapurine Nucleosides: Synthesis, Properties, and Base Pairing of Oligonucleotides. Chemistry and Biodiversity 2010, 7, 2145–2190. [Google Scholar] [CrossRef]
- Westarp, S.; Brandt, F.; Neumair, L.; Betz, C.; Dagane, A.; Kemper, S.; Jacob, C.R.; Neubauer, P.; Kurreck, A.; Kaspar, F. Nucleoside phosphorylases make N7-xanthosine. Nature Comm. 2024, 15, 3625. [Google Scholar] [CrossRef] [PubMed]
- Liu, Gaofei; Wang, Jialing; Chu, Jianlin; Jiang, Tianyue; Qin, Song; Gao, Zhen; He, Bingfang. Engineering Substrate Promiscuity of Nucleoside Phosphorylase Via an Insertions-Deletions Strategy. JACS AU, 2024, 4, 454-464. [CrossRef]
- Lovelock SL, Crawshaw R, Basler S, Levy C, Baker D, Hilvert D, Green AP: The road to fully programmable protein catalysis. Nature 2022, 606, 49–58. [CrossRef] [PubMed]
- Nyhan, W.L. Lesch-Nyhan Disease and Related Disorders of Purine Metabolism. TZU CHI Medical J. 2007, 19, 105–108. [Google Scholar] [CrossRef]
- De Bruyn, C.H.M.M. Hypoxanthine-guanine phosphoribosyl transferase deficiency. Human Genet. 1976, 31, 127–150. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
Tri-cyclic analogs of adenosine and guasnosine.
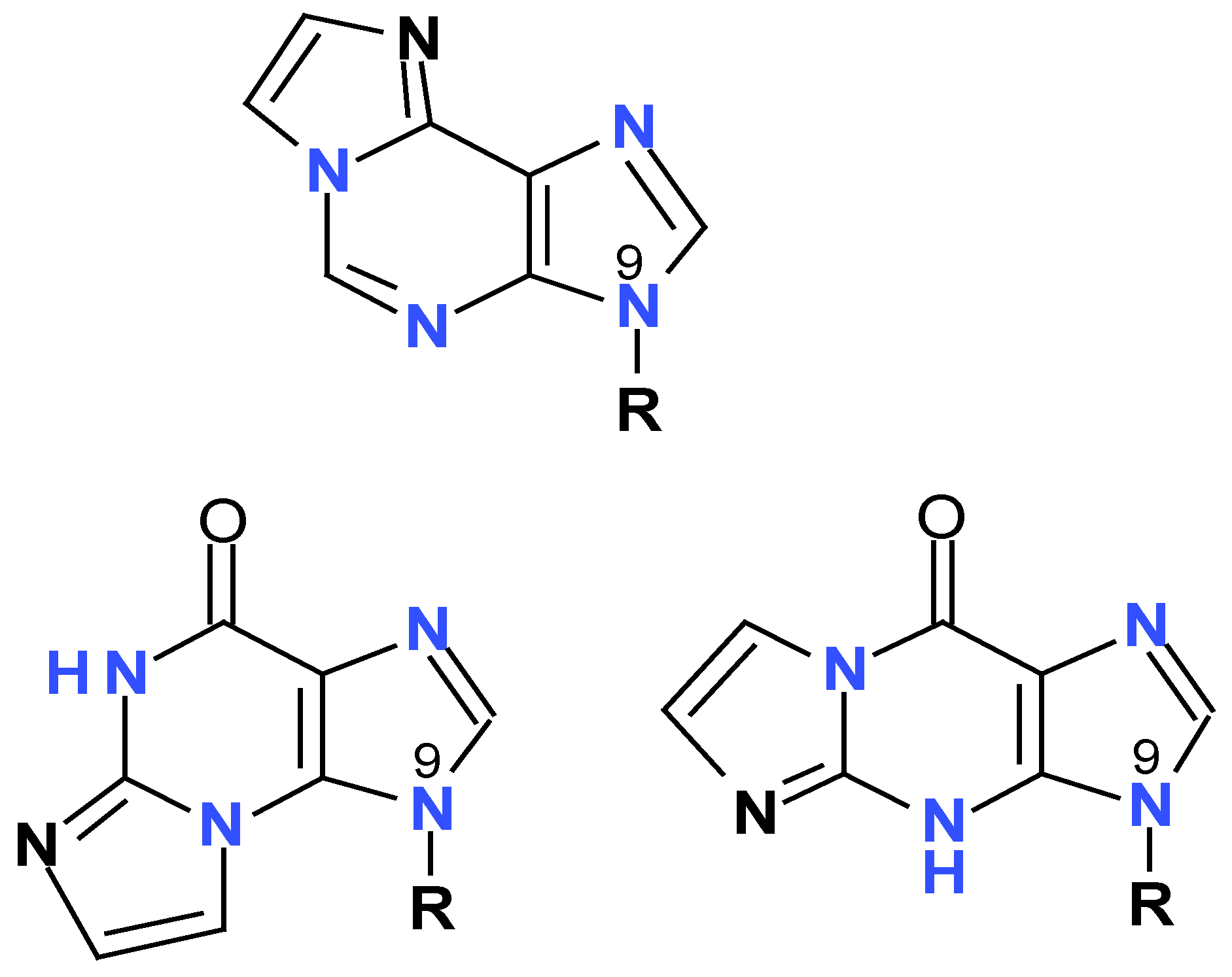
Table 1.
Fluorescence parameters of selected 8-azapurine ribosides, compared to other isomorphic purine and purine nucleoside analogs. Unless otherwise indicated, all data refer to neutral species of the fluorophores. Data from references [15,49,53].
| Compound | excitation: λmax [nm] |
emission: λmax [nm] |
φ | τ [ns] |
|---|---|---|---|---|
| 2-aminopurine riboside | 305 | 380 | 0.68 | ~8 |
| 8-azaxanthosine* | 290 | 440 | >0.1 | nd |
| 8-azaguanosine (neutral) | 260 | 347 | <0.01 | |
| 8-azaguanosine (anionic) | 278 | 362 | 0.55 | 5.6 |
| 8-azaguanine-N7-β-D-riboside (anionic) | 304 | 420 | 0.03 | nd |
| 8-azaguanine-N8-β-D-riboside | ~290 | nd | >0.1 | nd |
| 8-azaadenosine | 278 | 352 | 0.068 | 0.8 |
| 8-azainosine (anionic) | 275 | 357 | 0.018 | nd |
| 2,6-diamino-8-azapurine- N9-β-D-riboside |
285 | 365 | 0.9 | ~6 |
| 2,6-diamino-8-azapurine- N7-β-D-riboside |
314 | 420 | 0.063 | 1.5;0.45 |
| 2,6-diamino-8-azapurine- N8-β-D-riboside |
313 | 430 | 0.41 | 10.5 |
| 8-aza-7-deaza-isoguanine | 288 | 370 | nd | nd |
| 8-aza-isoguanosine (anionic) | 285 | 370 | ~0.1 | nd |
| 8-aza-7-deaza-isoguanosine* | 285 | 430? | nd | nd |
*The ribosylation site is uncertain.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



