Submitted:
10 May 2024
Posted:
12 May 2024
You are already at the latest version
Abstract
The flowering period of rice significantly impacts variety adaptability and yield formation. Properly shortening the reproductive period of rice varieties can expand their ecological range without significant yield reduction. Targeted genome editing, like CRISPR/Cas9, is an ideal tool to fine-tune rice growth stages and boost yield synergistically. In this study, we developed a CRISPR/Cas9-mediated multiplex genome editing vector containing five genes related to three traits: Hd2, Ghd7, and DTH8 (flowering stage genes), along with the recessive rice blast resistance gene Pi21 and the aromatic gene BADH2. This vector was introduced into the high-quality Geng rice variety in Zhejiang province, Jiahe212 (JH212), resulting in 34 T0 plants with various effective mutations. Among the 17 mutant T1 lines, several displayed diverse flowering dates, but most exhibited undesirable agronomic traits. Notably, three homozygous mutant lines (JH-C15, JH-C18, and JH-C31) showed slightly earlier flowering dates without significant differences in yield-related traits compared to JH212. Through special Hyg and Cas marker selection of T2 plants, we identified 7, 6, and 2 fragrant glutinous plants devoid of transgenic components, respectively. These single plants will serve as sib lines of JH212 and potential resources for breeding applications, including maintenance lines for Xian-Geng interspecific three-line hybrid rice. In summary, our research offers practical insights and potential resources for targeted improvement of Geng rice varieties, and would also promote the Geng rice varieties expending from the southward to the northward.
Keywords:
Rice
; Flowering period
; Multiplex Genome Editing
; Breed improvement
1. Introduction
Rice growth and yield are influenced by light-temperature interaction, affecting flowering, fruiting, and yield formation. The flowering period, a complex trait governed by multiple genes [1], plays a crucial role in rice adaptation and yield optimization [2] by maximizing light and temperature resources [3]. Modifying the flowering period expands the cultivation area for high-quality rice varieties, addressing production challenges. Rice blast, caused by Magnaporthe oryzae, poses a significant threat to rice yield stability [4]. Developing disease-resistant rice varieties is pivotal for effective pathogen control.
Traditional rice breeding is time-consuming due to limited heritable variation and extensive phenotypic screening. Genetic engineering techniques, particularly CRISPR/Cas9, which is a cutting-edge gene editing tool and widely applied in crop genetic research for revolutionize trait improvement by precise DNA-level editing, accelerating breeding with enhanced efficiency and targeted outcomes. The approach offers rapid enhancements in flowering date, grain quality, stress resistance, and crop yield, minimizing time costs [5,6,7,8,9]. Noteworthy studies include editing the DTH8 gene to alter tasselling periods of Geng rice 99-25 and obtaining DTH8 mutant material with an earlier tasselling period [10]. In addition, this technology has been used to enhance resistant starch content through SS3a and SS3b editing, achieving a significant increase of 4.7%-5.0% in single knockouts, and the double-knocked out mutant exhibiting significant differences, with the resistant starch increasing 9.5%-9.7%, creating new nutritious rice germplasm [11]. Wang Kejian's team at the China National Rice Research Institute employed CRISPR/Cas9 gene editing technology to knock out four reproduction-related genes, PAIR1, REC8, OSD1, and MTL, in the hybrid rice Chunyou 84, achieving fusion-free reproduction of hybrid rice from 0 to 1, which is of great theoretical and practical significance [12]. Zhou Wenjia used CRISPR/Cas9 to edit the flowering period gene Hd2 and the aroma gene Badh2 of Sui-Geng 14, to shorten the flowering period [13]. Zhou employed CRISPR/Cas9 to edit broad-spectrum rice blast resistance genes Bsr-d1, Pi21, and ERF922. Single- or triple-mutants displayed high resistance to rice blast, with Pi21 or erf922 single mutants exhibiting enhanced resistance to leaf blight [14] Plant gene editing technology offers avenues for creating high-quality crop varieties, demonstrating the feasibility of directly editing genes in superior crop varieties for improved commercial crops. Cite a case, rice flowering is controlled by multiple genes, with allele variants influencing regional adaptability under varying day lengths. Hd2, a core gene in rice photoperiodic flowering regulation system, functions as a flowering repressor under long daylight conditions [15]. Its expression level correlates significantly with flowering time, regulating rice flowering by enhancing photoreceptivity [16] Ghd7, a major QTL, controls grain number per spike, plant height, and flowering period in rice [17,18]. In the photoperiodic pathway under prolonged sunlight, Ghd7 represses Ehd1 expression, inhibiting flowering through the Ghd7-Ehd1-Hd3a/RFT1 pathway, and upregulates OsCOL10 and OsMFT1 as downstream flowering repressors [19,20]. Similarly, DTH8, a member of the rice HAP family (containing the HAP3D subunit), affects flowering, plant height, and grain number per spike [21] under extended sunlight, repressing flowering through protein interactions with Hd1, Ghd7, DTH7, and HAPL1 [22].
Previous studies have demonstrated the Ehd1-dependent photoperiodic flowering pathway, three flowering repressor genes, Hd2, Ghd7, and DTH8 negatively regulated during the flowering period in rice varieties. Delaying rice flowering under prolonged sunlight conditions produced increased plant height and number of grains per spike, leading to higher yields. Therefore, it is important to shorten the rice flowering period by editing Hd2, Ghd7, and DTH8, to genetically improve the flowering period of Geng rice variety. Pi21, a recessive rice blast resistance gene [23], contributes to persistent rice blast resistance in Japanese rice varieties. The recessive version of the Pi21 gene has a deletion of the first and second proline domains corresponding to 21 bp and 48 bp, respectively, associated with its resistance to rice blast [24]. Pi21 negatively regulates disease resistance, and its loss-of-function shows resistance to rice blast, and the use of this resistance is expected to overcome the problem of rice varieties prone to lose rice blast resistance.
Jiahe212 (JH212) is a traditional Geng rice variety renowned for its superior quality and serves as the maintainer line for Jiahe212A (JH212A), a high-quality hybrid line widely used in southern China. However, hybrid combinations involving JH212A often exhibit prolonged growth periods and low heat tolerance, hindering their widespread adoption. To address this, shortening the flowering period can mitigate heat stress during sowing, allowing for more flexible planting arrangements and facilitating the expansion of cultivation areas, especially northward. In this study, we employed the CRISPR/Cas9 system to conduct multi-gene editing targeting key genes in rice flowering regulation (Hd2, Ghd7, and DTH8), the rice blast resistance gene Pi21, and the flavor gene Badh2 in JH212. The objective was to develop homozygous and stable mutant lines that enhance disease resistance and expedite the flowering period while preserving rice quality. These modifications aim to enhance breeding flexibility and promote wider adoption of high-quality Xian/Geng interspecific hybrid rice varieties.
2. Results
2.1. Target Design and Vector Construction
The CRISPR-GE website (http://skl.scau.edu.cn/) was utilized for gRNA target site design. Primer sequences were selected based on high scores and proximity to the ATG start site. Target sites were designed on the first CDS1 of Hd2, DTH8, and Pi21, and on the second CDS2 of Ghd7 and Badh2 for vector construction. The target site sequences included NGG (Figure 1). All five target sites were positioned close to the ATG start codon to facilitate the loss of function of the target gene. Finally, the rice genome was analyzed using BLAST through NCBI to confirm the specificity of the target sites.
2.2. Target Design and Off-Target Analysis
A set of candidate target sequences was generated by inputting these gene sequences into the target design tool on the CRISPR-GE webpage. Following the criteria for efficient target site selection, we opted for sequences with a GC content between 45% and 70%, off-target scores less than 0.6, and sgRNA sequences with fewer than 8 base pairs of complementarity. These selections were made to minimize off-target effects and enhance gene editing efficiency. All five targets chosen for this experiment exhibited low off-target indices (Table 1), indicating the feasibility of gene editing at these sites. However, it is important to remain vigilant about potential off-target risks during subsequent mutant screening.
2.3. Vector Construction and Genetic Transformation
In the laboratory of Kejian Wang at the China Rice Research Institute, the primary objective was to construct CRISPR/Cas9 expression vectors targeting five genes simultaneously using traditional homotetrameric enzyme technology [12]. Initially, the five target sequences were assembled into the intermediate vector SK gRNA. Subsequently, SK-gRNA-Hd2, SK-gRNA-Ghd7, SK-gRNA-DTH8, and SK-gRNA-Pi21 were combined into a single intermediate vector named SK-gRNA-Hd2-Ghd7-DTH8-Pi21. Separately, SK-gRNA-Badh2 was assembled into another intermediate vector, which was then ligated with the intermediate vector containing four sgRNAs to form an intermediate vector encompassing five genes. Finally, the module with the five sgRNAs in tandem was integrated into the expression vector pCB2 using digestion-ligation methods. The resultant tandem module of five sgRNAs was further integrated into the expression vector pC1300-Cas9 via enzymatic cleavage and ligation. The constructed expression vector, designated CRISPR/Cas9-JH212, was subsequently sent to Wuhan Boyuan Company for genetic transformation of the Geng rice variety 'Jiahe 212' (Figure 2).
2.4. Acquisition of T0 Transgenic Positive Plants
The expression vector CRISPR/Cas9-JH212, carrying five targets, was employed to transform the recipient material JH212 via Agrobacterium-mediated methods, resulting in the generation of T0 transgenic lines. Detection of the vector region in the transgenic seedlings was conducted using vector-specific primers Cas9-F/Cas9-R to screen for transgene-positive lines. The analysis revealed that among 34 T0 transgenic seedlings, 30 were positive, corresponding to a positive rate of 88.23% (Figure 3).
2.5. Mutation Analysis of Transgenic Seedlings in T0 Generation
To analyze the mutation of Hd2, Ghd7, DTH8, Pi21, and Badh2 genes in JH212, the upstream and downstream regions of the five target sequences were amplified using target detection primers. Subsequently, the PCR products were sequenced and analyzed. The sequencing results revealed mutations at the following frequencies: 26 positive plants were mutated at the Hd2 target site position (mutation frequency of 86.66%), 25 positive seedlings were mutated at the Ghd7 target site (mutation frequency of 83.33%), 25 positive seedlings were mutated at the DTH8 target site (mutation frequency of 83.33%), and 26 positive seedlings were mutated at the Pi21 target site (mutation frequency of 86.66%) (Table 2). The mutation genotypes included homozygous homozygous mutations, double allelic mutations, and various mutation types, such as base insertions and deletions.
2.6. Analysis of Mutation Sites in T1 Mutants
Based on the PCR sequencing results of the T1 plants, Hd2 exhibited six types of homozygous mutations, including base insertions and deletions, notably insertions of T, A, C, and G between the third and fourth bases of PAM, and deletions of 4 bp and 46 bp. Ghd7 displayed a total of eight types of homozygous mutations, including base insertions and deletions, notably insertions of T, A, and deletions of 3 bp, 4 bp, 5 bp, 7 bp, 28 bp, and 45 bp. DTH8 showed four types of homozygous mutations, involving deletions of 1 bp, 4 bp and 6 bp. Pi21 exhibited nine types of homozygous mutations, including insertions, deletions, and other base mutations, particularly insertions of single bases T, A, and G, as well as deletions of various bases (Figure 4).
The green letters indicate the PAM sequence; Red letters and hyphen indicate base insertions and missing bases, respectively. a, The 6 mutation types of Hd2; b, The four mutation types of DTH8; c, The 8 mutation types of Ghd7; d, The 9 mutation types of Pi21.
2.7. Analysis of Mutation Sites in Homozygous T1 Mutants
After genotyping and analyzing the T1 generation mutant plants, 17 different genotypes were identified (Table 3), the flowering time of which were 24.25 days to 38.3 days earlier than that of JH212 (Table 4). Following phenotypic observations and agronomic trait investigations, three T2 lines were selected for the subsequent experiment: four-gene, three-gene, and two-gene homozygous and mutant lines. Subsequently, DNA sequences upstream and downstream o f the target sites of the mutant plants were amplified using target detection primers (Table 4). Sequencing of the PCR products revealed that the mutations at the target sites of the T1 generation mutants were consistent with those observed in the T0 generation. This indicated that the genotypes of these three homozygous and mutant lines could be stably inherited. The three homozygous and mutant lines were established by self-pollinating the aforementioned three homozygous and mutant lines in the T2 generation and were named JH-C15, JH-C18, and JH-C31 (Table 4).
2.8. T1 Generation and T2 Generations of Cas9-Free Plants Were Obtained.
To prevent ongoing gene editing by the Cas9 vector in transgenic plants, we implemented a strategy to remove the Cas9 vector starting from the T0 generation. This involved self-pollinating the transgenic plants across three consecutive generations. At the outset of each transgenic seedling generation, once we confirmed gene editing in the transgenic lines, we conducted PCR amplification using two pairs of specific primers: Hyg-F/Hyg-R and Cas9-F/Cas9-R. If the genome of a transgenic plant amplified with both primer pairs did not yield the target fragment, we inferred that the target gene in that plant was mutated and the Cas9 vector was removed. These plants were then transplanted into the greenhouse for further cultivation. After two successive generations of self-pollination, we assessed the removal of the vector from the CRISPR/Cas9 knockout lines. While attempting to screen for plants without transgenic components in the T1 generation, we did not achieve homozygous mutant lines without transgene mutations due to the high temperature encountered during the T0 generation. Our focus was primarily on counting the removal of the Cas9 vector in the CRISPR/Cas9 knockout lines in the selected homozygotes lines in T2 generation, namely, JH-C15, JH-C18 and JH-C31. The results indicated that out of 15 independent lines, we successfully obtained seven transgenic lines without the Cas9 vectors (Figure 5a,b). The three selected homozygotes lines showed significantly earlier flowering and maturity, significantly reduced plant height, but satisfactory performance in yield traits (Figure 5c–h).
2.9. qRT-PCR Detection of Gene Editing in Homozygous Mutant Lines
The expression levels of flowering and flowering genes Hd3a, Hd2, Ghd7, DTH8, and Pi21 were assessed using qRT-PCR, and in the three homozygous and mutant lines JH-C15, JH-C18, and JH-C31 mentioned previously. The expression of Hd3a was significantly higher in the JH-C15, JH-C18, and JH-C31 homozygous mutant lines compared to the wild-type. Conversely, Hd2 was significantly lower in all three homozygous and mutant lines compared to the wild type. Ghd7 expression was significantly lower in the JH-C18 four-gene homozygous and mutant line compared with the wild type. DTH8 expression was significantly reduced in the JH-C15 three-gene homozygous and mutant line and the JH-C18 four-gene homozygous and mutant line compared to the wild-type. Additionally, the expression of Pi21 was significantly lower in all three homozygous mutant lines compared to the wild type. These findings suggest that the transcript levels of Hd2, Ghd7, DTH8, and Pi21, may have been affected and partially degraded in vivo following target editing by the Cas9 system, resulting in a significant downregulation of their expression in the mutant lines JH-C15, JH-C18, and JH-C31 (Figure 6a–c).
2.9. Phenotypic Analysis of T2 Generation Knockout Mutants
In this study, we investigated the phenotypes of the CRISPR/Cas9 knockout lines, focusing on flowering and yield-related traits in three pure mutant lines mentioned earlier. JH-C15 showed wider grain (Figure 7a–e); JH-C18 exhibited lager grain length/width without significant influence on grain weight (Figure 7a–e); JH-C31 displayed wider grains without significant influence on grain weight (Figure 7a–e). All the three lines exhibited significantly reduced plant height compared to JH212 (Figure 8f). The number of effective panicles has a significant impact on the yield of a single plant, such as JH-C15, the main reason for the decrease in its yield is the significant decrease in the number of effective panicles (Figure 7g,h). The yield of JH-C18 and JH-C31 showed no significant decrease ((Figure 7g,h), indicating their potential utilization value.
Among the identified flowering repressors, Hd2 encodes a PRR protein homologous to Arabidopsis TOC1, Ghd7 is a CO-like protein containing a CCT structural domain that controls plant height, flowering period, and spike number in rice, while DTH8 serves as a pleiotropic regulator of plant height and flowering period. Ehd1 promotes the expression of Hd3a and RFT1 under long sunlight conditions, with Hd2, Ghd7, and DTH8 acting as inhibitors of Ehd1. In the JH-C15 triple mutant strain, we observed a base T insertion at the Hd2 target site and a base G deletion in the DTH8 target site, resulting in premature protein translation termination and loss of gene function (Table S3). Consequently, under long sunlight conditions, the JH-C15 mutant exhibited significantly earlier flowering compared to the wild-type, indicating successful knockout of the DTH8 in JH-C15 and JH-C18 strains. Similarly, in the JH-C18 four-mutant line, a 46-bp deletion was observed at the Hd2 target site, along with a base T insertion at the Ghd7 target site and a four-base deletion at the DTH8 gene sequence target site (Table S3). This resulted in significantly earlier flowering in the JH-C18 mutant compared to the wild-type, confirming successful knockout of the Hd2, Ghd7, and DTH8 genes in JH-C18. In the JH-C31 double mutant strain, a 4-bp deletion in the Hd2 gene led to earlier flowering of the spike compared to the wild-type, indicating successful knockout of the Hd2 gene (Figure 8f, Table S3).Among the three families obtained, JH-C31 had fewer mutated genes and showed better performance in yield traits, effective tillering, yield per plant, grain length and width ratio relative to the other two families, while JH-C15 in turn showed lower yield, especially decreased panicle number per plant than JH-C18 and JH-C31. This result may be attributed to the different number of genes mutated in these three lines as well as the different ways in which the genes were mutated and the different magnitude of the effect of these three spike stage genes. Hence, it resulted in showing different agronomic traits.
When testing rice blast resistance, we inoculated JH212 and three mutant lines at the three-leaf, one-center stage with rice blast. Surprisingly, there was no notable difference in disease resistance between the edited lines and JH212. Additionally, the selected physiological minor seed used in this study exhibited high resistance both in JH212 and the edited strains, resulting in a phenotype lacking disease resistance following rice blast inoculation.
3. Discussion
Rice yield and adaptability are influenced by factors like spiking duration, disease resistance of specific physiological minor varieties, and cropping schedules. While extending the spiking period is commonly seen as a method to boost yields, there is growing interest in short-duration high-quality rice varieties due to their flexible sowing nature, which allows for expansion without significant yield loss. The spiking period, a critical physiological phase determining rice variety adaptability [25], has garnered attention from breeders. Improving the flowering period of existing Geng varieties can expand high-quality rice cultivation and address flowering period-related challenges in popularized varieties. Maximizing rice yield relies on cultivating varieties with optimal flowering periods for the planting region. Gene editing offers a rapid and stable breeding pathway favored by experts [26], boasting high efficiency, stability, and speed compared to traditional breeding methods like hybridization and backcrossing. CRISPR/Cas9 gene editing technology has rapidly advanced in recent years, extensively applied in creating germplasm resources [27], genetic enhancement [28], fertility improvement [29], disease resistance [14], and quality improvement of crops [29]. Its adoption in rice research has become instrumental in unraveling gene functions and molecular mechanisms [26].
In this study, we employed CRISPR/Cas9 gene editing technology to modify Hd2, Ghd7, and DTH8 in the rice flowering pathway and rice blast resistance gene Pi21. We successfully created new allelic variants in the JH212 background, establishing novel breeding materials for the flowering variation. In the T0 generation, 88.23% efficiency was achieved, with 30 out of 34 transgenic seedlings showing positive results. Sequenced analyses revealed high mutation frequencies: Hd2 (86.6%), Ghd7 (83.3%), DTH8 (83.3%), and Pi21 (86.6%), predominantly involving base insertions and deletions. Our results align with previous studies: Li et al. achieved 77.8% simultaneous editing of Hd2, Ghd7, and DTH8 across seven cultivars [30]; Yang et al. obtained an 86.7% mutation rate for Pi21; Xu et al. reported mutation frequencies of 75% for Pita, 85% for Pi21, and 65% for ERF922 [31]. These data fully proved the effectiveness of our new-constructed vector.
Important agronomic traits in rice are also jointly regulated by multiple genes or gene families. Using the CRISPR-Cas9 multiple knockout system, multiple genes can be targeted simultaneously to obtain plants with variable traits and to investigate the interactions among genes [32], offering new materials for crop breeding. In this study, we found that the probability of single gene mutation was very limited, and many of the mutations occurred at the same time, primarily as polygenic mutations. Notably, we identified a total of eight homozygous mutation types for Ghd7, six homozygous mutation types for Hd2, nine homozygous mutation types for Pi21, and four homozygous mutation types for DTH8. These results are basically consistent with previous researches. Yang et al. utilized CRISPR/Cas9 to simultaneously knockdown eight genes highly expressed in the glutenin gene family and obtained seven mutants with varying combinations of mutations without affecting the content of storage substances and rice appearance, while the rice protein content was down-regulated to varying degrees [33]. Shen et al successfully developed co-knockout vectors for eight genes associated with agronomic traits in rice using CRISPR-Cas9, and obtained homozygous six-mutant, seven-mutant, and eight-mutant strains [34]. This approach provides a strategy for the rapid introduction of genetic diversity in the development of crop breeding. In constructing knockout lines, we have obtained materials to further study the dosage effect of alleles on phenotypes and genetic interactions between non-alleles, but also expanded the possibilities for the use of CRISPR/Cas9 gene editing technology in targeted editing of the genomes of rice varieties, to obtain polymerization of favorable alleles. In future work, we can better utilize these haplotypes to enhance the diversity of flowering times. Moreover, by integrating several genes with flowering-promoting impacts, it is possible to breed varieties with very early flowering times.
In the 17 homozygous mutant lines obtained, the four-gene homozygous mutant line JH-C18, the three-gene homozygous mutant line JH-C15, and the two-gene homozygous mutant line JH-C31 were chosen for the following experiments by observing phenotypes, and investigation of agronomic traits. In analyzing the expression of flowering-related genes in the gene-edited homozygous lines, the expression of Hd3a in the homozygous mutant lines JH-C15, JH-C18, and JH-C31 was significantly higher than that of JH212, and the transcript levels of Hd2, Ghd7, DTH8, and Pi21, might be impacted after being target-edited by the Cas9 system, partially degrading the RNAs in vivo, producing a significant down-regulation of their gene expression in JH-C15, JH-C18, and JH-C31. Previous studies reported that in the Ehd1-dependent photoperiodic flowering pathway, Hd2, Ghd7, and DTH8 are the primary genes negatively regulated by flowering in rice varieties, delaying rice flowering under long sunlight conditions. Li et al. designed specific targets for three flowering genes, including Hd2, Ghd7, and DTH8, showing that the knockout progeny could reduce the flowering period to varying degrees [30]. Zhou et al. employed gene editing technology to target the flavor gene Badh2 and the flowering period gene Hd2 to obtain improved rice varieties with improved flavor and early maturity [35]. Multi-target editing of Pi21 and the flavor suppressor gene OsBadh2 resulted in highly significant reductions in expression relative to the wild-type, and improved rice blast resistance and accumulation of 2-AP. In this study, the knockdown of Hd2, Ghd7, and DTH8 resulted in significant advancement in the flowering period of mutant lines compared to the wild-type, indicating that the CRISPR/Cas9-mediated gene editing at the flowering period can achieve targeted fertility improvement, facilitate the introduction of high-quality rice resources, and accelerate local breeding programs. The analysis of genetic and molecular mechanisms of flowering regulation in rice and the development of flowering regulatory pathways will offer breeders more targeted and effective options for selecting the most suitable ecotypes for local growing areas and provide optimal forms and combinations of flowering gene alleles for future molecular design breeding. The use of CRISPR/Cas9 can regulate gene expression, creating new alleles that do not exist in nature and offering broad application prospects in future breeding of new breakthrough and diversified rice varieties.
Recently, the CRISPR/Cas9 gene editing system has become increasingly advanced. Compared to previous generations of gene editing technology, the CRISPR/Cas9 system offers advantages such as easier construction, lower cost, and simpler operation. However, CRISPR/Cas9 is not without flaws, and its high off-target rate has been a subject of criticism. In this study, the Badh2 gene target experienced off-target effects. To address this issue, Fu et al. developed a series of progressively shorter guide RNAs (gRNAs) for the EGFP reporter gene, including 15, 17, 19, or 20 complementary nucleotides. gRNAs with 17 or 18 complementary nucleotides could function at the intended target sites, reducing the off-target effects of paired Cas9 endonucleases without compromising the efficiency of targeted genome editing [37] Research has shown that gRNAs can direct Cas9 for on-target editing via base pairing with "seed sequences" located 8-12 nucleotides upstream of the protospacer adjacent motif (PAM). Therefore, appropriately lowering the length of the gRNA by 1-3 nucleotides can effectively reduce the off-target probability without impacting its targeting ability [38]. In addition, we selected physiological minor seeds with significant resistance against rice blast in varieties such as ZH11 and ZH8015 to inoculate JH212 with rice blast in this study [31,36]. The final plants did not get the relevant phenotypes, leading to the failure of the experiment. The reason for this failure may be due to the fact that JH212 is a variety with good blast resistance, or the fact that the selected physiological seed is not suitable for the inoculation, or the genetic effect of mutation type were not strong enough. In conclusion, despite certain limitations, CRISPR/Cas9 has attracted increasing attention from researchers due to its unique ability to precisely modify genes. This has prompted in-depth studies to overcome its challenges and examine its full potential. The three homozygous mutant lines obtained in this study, namely, JH-C15, JH-C18 and JH-C31, especially the last two, could serve as potential resources for expanding the promotion and application areas of JH212.
4. Materials and Methods
4.1. Experimental Materials and Planting
In this study, JH212 served as the recipient material for genetic transformation. Wild-type and transgenic plants from generations T0 and T2 were cultivated at the transgenic experimental base of the China Rice Research Institute. T1 plants were grown in Hainan during winter, receiving conventional water and fertilization practices. The expression vectors pC1300-Cas9 and SK-gRNA intermediate vectors were supplied by the laboratory of Wang Kejian at the China Rice Research Institute.
4.2. gRNA Target Site Design
The CRISPR-P website (http://cbi.hzau.edu.cn/cgibin/CRISPR) was utilized for designing gRNA target sites. Primer sequences with high scores and proximity to the ATG start site were selected. Gene sequences for Hd2, Ghd7, DTH8, Pi21, and Badh2 were inputted into the CRISPR-P website to facilitate target site design. The CRISPR/Cas9 system was used to recognize approximately 20 bases upstream of the PAM to identify suitable target sites. After identifying the target sites, we added the GGCA sticky end to the front and the CCCA sticky end to the back of the target site sequence. The specific primer sequences are detailed in Table 1. These five target sites were positioned close to the ATG start codon to facilitate functional loss of the target genes. Lastly, the specificity of the target sites was confirmed by BLAST analysis of the rice genome using NCBI.
4.3. Construction of CRISPR/Cas9 Expression Vector
Five genes were targeted for knockout using the CRISPR/Cas9 system. The successfully constructed expression vectors were introduced into Agrobacterium through homotetramer ligation. Primers Hd2-g-F/R, Ghd7-g-F/R, DTH8-g-F/R, Pi21-g-F/R, and Badh2-g-F/R (Table S1) were synthesized based on the target site sequences. These primers were mixed in equal amounts, denatured, and annealed to form fragments with sticky ends. The sgRNA fragments for Hd2, Ghd7, DTH8, and Pi21 were digested with Kpn I/Xho I, Sal I/Xba I, Nhe I/BamH I, and Kpn I/Bgl II, respectively, and then assembled into an intermediate vector. Subsequently, the two sgRNA fragments were combined into an intermediate vector containing all five sgRNAs. Finally, the intermediate vector containing the five sgRNAs was digested with KpnⅠ/BglⅡ and ligated into the final expression vector pC1300-Cas9 (KpnⅠ/BamHⅠ double digestion). The constructed CRISPR/Cas9-2 expression vector was used for the genetic transformation of the Geng rice variety JH212 by Wuhan biorun biosciences co., ltd.
4.4. Acquisition of T0 Generation Positive Strains and Identification of Target Sites
The vector CRISPR/Cas9-2 was transferred to JH212 rice callus using the Agrobacterium-mediated method, and hygromycin was applied for screening. At the full tillering stage, leaf DNA from the gene-edited rice plants was extracted using the CTAB method. Positive gene-edited plants were identified by PCR using primers Hyg-F/Hyg-R specific for the thaumatin resistance gene in the vector. Next, primers flanking the target sequences of genes Hd2, Ghd7, DTH8, Pi21, and Badh2 were designed to amplify target bands from the T0-positive plants. The PCR products were then sequenced by Hangzhou Qingke Biotech Co., using the genome sequences of corresponding wild-type materials as references (Table S1). Sequencing results were compared with the wild-type sequences. A single peak in the sequencing results indicated a homozygous mutation, while a double peak indicated a heterozygous mutation, which required further isolation and verification.
4.5. Screening for GM-Free Components in T1 and T2 Generations
Upon analyzing mutations at the editing site of transgenic plants in the T0 generation, it was observed that only a small proportion had homozygous mutations, with most showing heterozygous mutations. To obtain homozygous mutant plants without the Cas9 carrier, we employed self-cross segregation to remove the carrier from the transgenic plants. Leaf samples from T1 and T2 generation mutant plants were collected, and genomic DNA was extracted using the CTAB method. Genomic sequences of the mutant plants were amplified using vector-specific primers Hyg-F/Hyg-R and Cas9-F/Cas9-R. The amplified products were analyzed using 1% agarose gel electrophoresis. If both pairs of primers failed to amplify a band, the plant was identified as a positive mutant plant lacking the exogenous transgene components.
4.6. qRT-PCR Analysis of Transgenic Plants
Total RNA was extracted using the RNAprep homozygous Plant Kit, and first strand cDNA was synthesized with the ReverTra Aceq PCR RT Master Mix with gDNA Remover (ToYoBo, Japan), following the manufacturer’s instructions. The expression levels of Hd2, Ghd7, DTH8, and Pi21 were analyzed in both wild-type and mutants using real-time fluorescence quantitative PCR (qRT-PCR) (Table S2). The qRT-PCR protocol involved pre-denaturation at 95 °C for 30 seconds, followed by denaturation at 95 °C for 5 seconds, annealing at 58 °C for 30 seconds, and extension at 72 °C for 15 seconds, repeated for 35 cycles. The primers for quantitative PCR are shown in Table 2. OsActin (accession number LOC_Os03g08020) served as the internal reference gene. Relative gene expression was calculated using the 2 -∆∆Ct method.
4.7. Mutant Phenotype Analysis
Phenotypic investigations were conducted on transgenic plants in the T2 generation, focusing on plant height and flowering period. For the flowering period, the measurement was taken 2 cm from the leaf sheath of the main spike of each plant, while plant height was measured as the distance from the top of the tallest spike to the soil surface after full irrigation. Data was recorded daily. To verify the effect of gene editing on the flowering period, a t-test was employed to compare wild-type rice materials and different mutants. A significance threshold (α) of 10-6 was set for the t-test. Specifically, if P ≤ 10-6 in the t-test comparing the days to flowering of the mutant with that of the control, it indicated that gene editing had significantly altered the flowering period.
5. Conclusions
In this study, we edited a specially designed Cas9 multi gene editing vector and transformed three flowering stage genes, Hd2, Ghd7, and DTH8, into the high quality Geng rice variety, JH212. This approach enabled us to produce progeny with different genotypes, producing more significant results regarding the relationship between genotypes and the flowering stage. Finally, we obtained three homozygous mutant lines, namely, JH-C15, JH-C18 and JH-C31, which exhibited significantly earlier flowering without significantly production loss. Our research would provide materials and theoretical basis for the ecological adaptability improvement of Geng rice, and also promote the Geng rice varieties expending from the southward to the northward.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Primers used in this study; Table S2: Quantitative primers used in this study; Table S3, Detailed mutation information of the three selected homozygotes lines.
Author Contributions
S.W., L.S., X. Z., and D.H. designed and conceived the study. D.H., X.C., R.Z., C.H., and D.L. performed the experiments. D.H.,. D.L. and L.S. analyzed all experiment data. Z.P. and Y.L. completed the field trial work. D.H. wrote the manuscript. S. W., L. S., X. Z., L. C., and S.C. reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by a fund of Guangxi Science and Technology Major Project grant number (GuiKe AA23062015), Zhejiang Provincial Key Special Projects, grant number (2021C02063-1), China Agriculture Research System, grant number (CARS-01-03), the Start-up Project for Studying 2023 High-level Talents of Hechi University (Grant No. 2023GCC020 and 2023GCC021), the National Natural Science Foundation of China, grant number (Grant No. 32272191), Chinese Academy of Agricultural Sciences Innovation Project, grant number CAAS-ASTIP-2013-CNRRI.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to thank Pro. Kejian Wang, Dr. Yong Huang and their laboratory pro providing the CRISPR/Cas9- mediated multiplex genome editing system and technical guidance.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhou, S.; Zhu, S.; Cui, S.; Hou, H.; Wu, H.; Hao, B.; Cai, L.; Xu, Z.; Liu, L.; Jiang, L.; et al. Transcriptional and Post-Transcriptional Regulation of Flowering Date in Rice. New Phytologist 2021, 230, 943–956. [Google Scholar] [CrossRef] [PubMed]
- Yuqi, S.U.N.; Qingyun, B.U.; Yunqing, C.; Xiaojie, T.; Zhenyu, W.; Yuekun, R.E.N.; Jiaqi, T.; Xiufeng, L.I. Research progress on the Flowering Date of rice in Northeast China. tryzw 2018, 7, 177–183. [Google Scholar] [CrossRef]
- Error: DOI Not Found Available online:. https://doi.org/10.11689/j.issn.2095-2961.2018.02.010 (accessed on 8 May 2024).
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 Fungal Pathogens in Molecular Plant Pathology. Mol Plant Pathol 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Qu, R.; Zhang, P.; Liu, Q.; Wang, Y.; Guo, W.; Du, Z.; Li, X.; Yang, L.; Yan, S.; Gu, X. Genome-Edited ATP BINDING CASSETTE B1 Transporter SD8 Knockouts Show Optimized Rice Architecture without Yield Penalty. Plant Commun 2022, 3, 100347. [Google Scholar] [CrossRef]
- Chen, W.; Chen, L.; Zhang, X.; Yang, N.; Guo, J.; Wang, M.; Ji, S.; Zhao, X.; Yin, P.; Cai, L.; et al. Convergent Selection of a WD40 Protein That Enhances Grain Yield in Maize and Rice. Science 2022, 375, eabg7985. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, S.; Huang, Y.; Ma, X.; Tan, L.; Liu, F.; Lv, Q.; Zhu, Z.; Hu, M.; Fu, Y.; et al. OsMADS17 Simultaneously Increases Grain Number and Grain Weight in Rice. Nat Commun 2023, 14, 3098. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Yang, Y.; Qin, R.; Li, H.; Qiu, C.; Li, L.; Wei, P.; Yang, J. Rapid Improvement of Grain Weight via Highly Efficient CRISPR/Cas9-Mediated Multiplex Genome Editing in Rice. J Genet Genomics 2016, 43, 529–532. [Google Scholar] [CrossRef]
- Huang, Y.; Dong, H.; Mou, C.; Wang, P.; Hao, Q.; Zhang, M.; Wu, H.; Zhang, F.; Ma, T.; Miao, R.; et al. Ribonuclease H-like Gene SMALL GRAIN2 Regulates Grain Size in Rice through Brassinosteroid Signaling Pathway. J Integr Plant Biol 2022, 64, 1883–1900. [Google Scholar] [CrossRef]
- ZHANG, H.; LIU, X.; XUAN, N.; ZHANG, H.; GAO, R.; ZHAO, Q.; YAO, F. Editing DTH8 Gene Using CRISPR /Cas9 Technology to Improve Heading Date of Rice 99-25. Acta Agriculturae Boreali-Sinica 2020, 35, 58–66. [Google Scholar]
- Huang, L.; Xiao, Y.; Zhao, W.; Rao, Y.; Shen, H.; Gu, Z.; Fan, X.; Li, Q.; Zhang, C.; Liu, Q. Creating High-Resistant Starch Rice by Simultaneous Editing of SS3a and SS3b. Plant Biotechnol J 2024, 22, 787–789. [Google Scholar] [CrossRef]
- Wang, C.; Liu, Q.; Shen, Y.; Hua, Y.; Wang, J.; Lin, J.; Wu, M.; Sun, T.; Cheng, Z.; Mercier, R.; et al. Clonal Seeds from Hybrid Rice by Simultaneous Genome Engineering of Meiosis and Fertilization Genes. Nat Biotechnol 2019, 37, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Wenjia, Z.; Xiaojie, T.; Yuekun, R.E.N.; Xiangjin, W.E.I.; Yang G. a., O.; Lihong, X.I.E.; Huazhao, L.I.U.; Qingyun, B.U.; Xiufeng, L.I. Breeding of Early-maturatity and Fragrant Rice via CRISPR/Cas9 Mediated Genome Editing. tryzw 2017, 6, 146–152. [Google Scholar] [CrossRef]
- Zhou, Y.; Xu, S.; Jiang, N.; Zhao, X.; Bai, Z.; Liu, J.; Yao, W.; Tang, Q.; Xiao, G.; Lv, C.; et al. Engineering of Rice Varieties with Enhanced Resistances to Both Blast and Bacterial Blight Diseases via CRISPR/Cas9. Plant Biotechnol J 2022, 20, 876–885. [Google Scholar] [CrossRef]
- Sun, K.; Huang, M.; Zong, W.; Xiao, D.; Lei, C.; Luo, Y.; Song, Y.; Li, S.; Hao, Y.; Luo, W.; et al. Hd1, Ghd7, and DTH8 Synergistically Determine the Rice Flowering Date and Yield-Related Agronomic Traits. J Genet Genomics 2022, 49, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Qu, X.; Zhou, Y.; Song, G.; Abiri, N.; Xiao, Y.; Liang, F.; Jiang, D.; Hu, Z.; Yang, D. OsPRR37 Confers an Expanded Regulation of the Diurnal Rhythms of the Transcriptome and Photoperiodic Flowering Pathways in Rice. Plant Cell Environ 2018, 41, 630–645. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Xing, Y.; Weng, X.; Zhao, Y.; Tang, W.; Wang, L.; Zhou, H.; Yu, S.; Xu, C.; Li, X.; et al. Natural Variation in Ghd7 Is an Important Regulator of Flowering Date and Yield Potential in Rice. Nat Genet 2008, 40, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Chen, S.; Wu, M.; Zheng, T.; Zhou, L.; Li, C.; Zhang, H.; Wang, J.; Xu, X.; Chai, J.; et al. Early Flowering 7 Interacts with DTH8, and Regulates Flowering Time in Rice. Plant Cell Rep 2019, 38, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Zheng, X.-M.; Chen, D.; Zhang, Y.; Ma, W.; Zhang, H.; Sun, L.; Yang, Z.; Zhao, C.; Zhan, X.; et al. OsCOL16, Encoding a CONSTANS-like Protein, Represses Flowering by up-Regulating Ghd7 Expression in Rice. Plant Sci 2017, 260, 60–69. [Google Scholar] [CrossRef]
- Tan, J.; Jin, M.; Wang, J.; Wu, F.; Sheng, P.; Cheng, Z.; Wang, J.; Zheng, X.; Chen, L.; Wang, M.; et al. OsCOL10, a CONSTANS-Like Gene, Functions as a Flowering Time Repressor Downstream of Ghd7 in Rice. Plant Cell Physiol 2016, 57, 798–812. [Google Scholar] [CrossRef]
- Zong, W.; Ren, D.; Huang, M.; Sun, K.; Feng, J.; Zhao, J.; Xiao, D.; Xie, W.; Liu, S.; Zhang, H.; et al. Strong Photoperiod Sensitivity Is Controlled by Cooperation and Competition among Hd1, Ghd7 and DTH8 in Rice Flowering. New Phytol 2021, 229, 1635–1649. [Google Scholar] [CrossRef]
- Du, A.; Tian, W.; Wei, M.; Yan, W.; He, H.; Zhou, D.; Huang, X.; Li, S.; Ouyang, X. The DTH8-Hd1 Module Mediates Day-Length-Dependent Regulation of Rice Flowering. Mol Plant 2017, 10, 948–961. [Google Scholar] [CrossRef]
- Fukuoka, S.; Okuno, K. QTL Analysis and Mapping of Pi21, a Recessive Gene for Field Resistance to Rice Blast in Japanese Upland Rice. Theor Appl Genet 2001, 103, 185–190. [Google Scholar] [CrossRef]
- Tao, H.; Shi, X.; He, F.; Wang, D.; Xiao, N.; Fang, H.; Wang, R.; Zhang, F.; Wang, M.; Li, A.; et al. Engineering Broad-Spectrum Disease-Resistant Rice by Editing Multiple Susceptibility Genes. J Integr Plant Biol 2021, 63, 1639–1648. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Liu, X.; Tang, W. Research Progress of Photoperiod Regulation in Rice Flowering. Chinese Journal OF Rice Science 2021, 35, 207. [Google Scholar] [CrossRef]
- Lorenzo, C.D.; Debray, K.; Herwegh, D.; Develtere, W.; Impens, L.; Schaumont, D.; Vandeputte, W.; Aesaert, S.; Coussens, G.; De Boe, Y.; et al. BREEDIT: A Multiplex Genome Editing Strategy to Improve Complex Quantitative Traits in Maize. Plant Cell 2023, 35, 218–238. [Google Scholar] [CrossRef] [PubMed]
- Romero, F.M.; Gatica-Arias, A. CRISPR/Cas9: Development and Application in Rice Breeding. Rice Science 2019, 26, 265–281. [Google Scholar] [CrossRef]
- Wang, F.; Wang, C.; Liu, P.; Lei, C.; Hao, W.; Gao, Y.; Liu, Y.-G.; Zhao, K. Enhanced Rice Blast Resistance by CRISPR/Cas9-Targeted Mutagenesis of the ERF Transcription Factor Gene OsERF922. PLOS ONE 2016, 11, e0154027. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, X.; Wu, C.; He, Y.; Ma, Y.; Hou, H.; Guo, X.; Du, W.; Zhao, Y.; Xia, L. Engineering Herbicide-Resistant Rice Plants through CRISPR/Cas9-Mediated Homologous Recombination of Acetolactate Synthase. Mol Plant 2016, 9, 628–631. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhou, W.; Ren, Y.; Tian, X.; Lv, T.; Wang, Z.; Fang, J.; Chu, C.; Yang, J.; Bu, Q. High-Efficiency Breeding of Early-Maturing Rice Cultivars via CRISPR/Cas9-Mediated Genome Editing. J Genet Genomics 2017, 44, 175–178. [Google Scholar] [CrossRef]
- Xu, P.; Wang, H.; Tu, R .; Liu, Q.; Wu, W.; Fu, X.; Cao, L.; SHEN, X. Orientation Improvement of Blast Resistance in Rice via CRISPR/Cas9 System. Chinese Journal of Rice Science 2019, 33, 313–322. [Google Scholar] [CrossRef]
- LIANG, M.; ZHANG, H.; CHEN, J.; DAI, D.; DU, C.; WANG, H.; MA, L. Developing Fragrant Early indica TGMS Line with Blast Resistance by Using CRISPR/Cas9 Technology. Chinese Journal of Rice Science 2022, 36, 248–258. [Google Scholar] [CrossRef]
- Yang, Y.; Shen, Z.; Li, Y.; Xu, C.; Xia, H.; Zhuang, H.; Sun, S.; Guo, M.; Yan, C. Rapid Improvement of Rice Eating and Cooking Quality through Gene Editing toward Glutelin as Target. J Integr Plant Biol 2022, 64, 1860–1865. [Google Scholar] [CrossRef]
- Shen, L.; Hua, Y.; Fu, Y.; Li, J.; Liu, Q.; Jiao, X.; Xin, G.; Wang, J.; Wang, X.; Yan, C.; et al. Rapid Generation of Genetic Diversity by Multiplex CRISPR/Cas9 Genome Editing in Rice. Sci China Life Sci 2017, 60, 506–515. [Google Scholar] [CrossRef]
- Zhou, W.; Tian, X.; Ren, Y.; Wei, X.; Gao, Y.; Xie, L.; Liu, H.; Bu, Q.; Li, X. Breeding of Early-maturatity and Fragrant Rice via CRISPR/Cas9 Mediated Genome Editing[J]. Soils and Crops, 2017; 6, 146–152. [Google Scholar] [CrossRef]
- Liu, Q.; Ning, Y.; Zhang, Y.; Yu, N.; Zhao, C.; Zhan, X.; Wu, W.; Chen, D.; Wei, X.; Wang, G.-L.; et al. OsCUL3a Negatively Regulates Cell Death and Immunity by Degrading OsNPR1 in Rice. Plant Cell 2017, 29, 345–359. [Google Scholar] [CrossRef]
- Fu, Y.; Sander, J.D.; Reyon, D.; Cascio, V.M.; Joung, J.K. Improving CRISPR-Cas Nuclease Specificity Using Truncated Guide RNAs. Nat Biotechnol 2014, 32, 279–284. [Google Scholar] [CrossRef]
- Pattanayak, V.; Lin, S.; Guilinger, J.P.; Ma, E.; Doudna, J.A.; Liu, D.R. High-Throughput Profiling of off-Target DNA Cleavage Reveals RNA-Programmed Cas9 Nuclease Specificity. Nat Biotechnol 2013, 31, 839–843. [Google Scholar] [CrossRef]
Figure 1.
Target site location data. a-e correspond to Hd2, Ghd7, DTH8, Pi21, and Badh2, respectively. The letters in the red box indicate the PAM sequence.
Figure 1.
Target site location data. a-e correspond to Hd2, Ghd7, DTH8, Pi21, and Badh2, respectively. The letters in the red box indicate the PAM sequence.
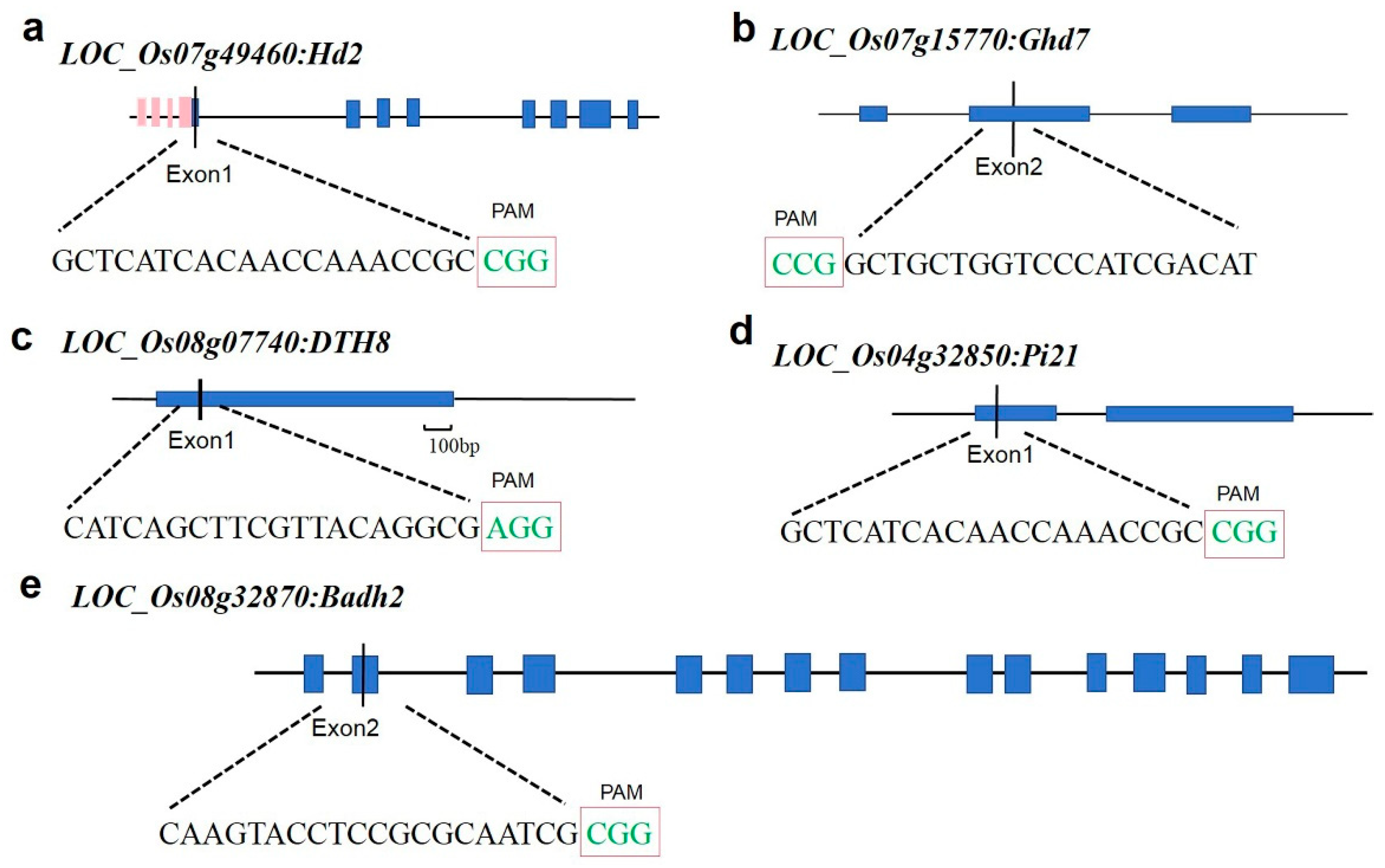
Figure 2.
Expression vector construction procedure.
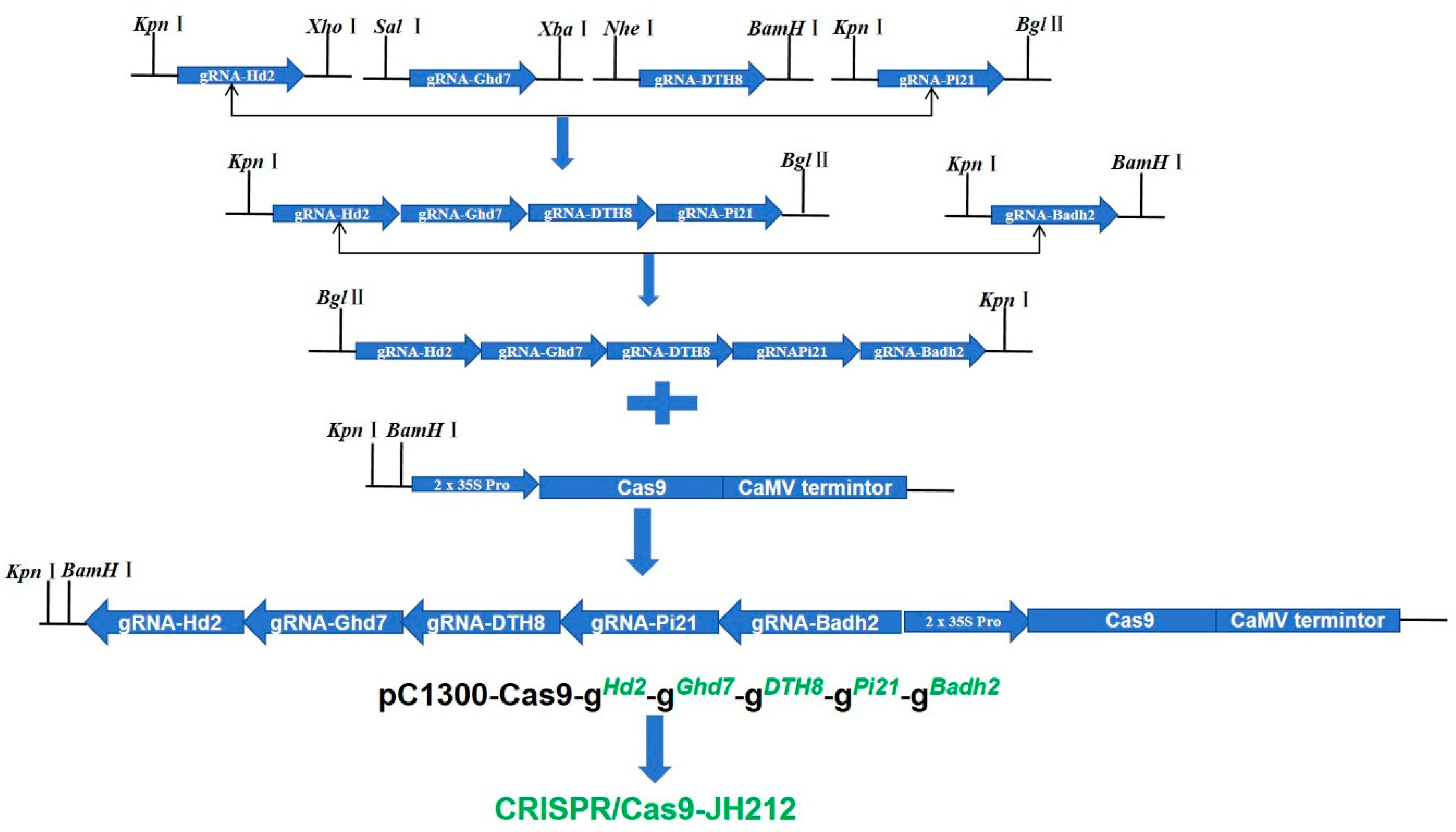
Figure 3.
PCR screening T0 generation positive strains. M, 2K DNA marker; 1-34 is the test line number.
Figure 3.
PCR screening T0 generation positive strains. M, 2K DNA marker; 1-34 is the test line number.

Figure 4.
Mutant genotype of the positive T1 plants tested.
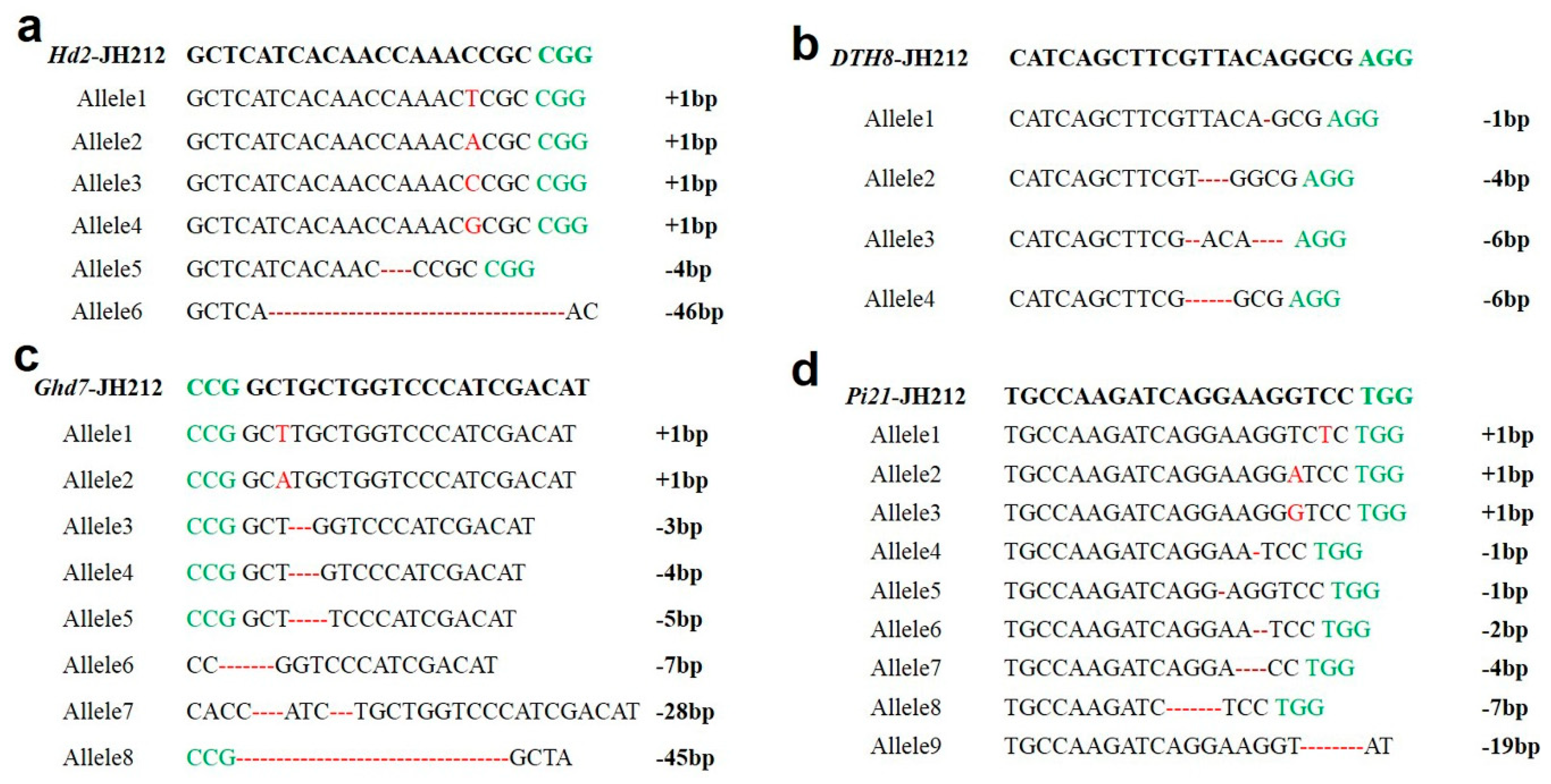
Figure 5.
PCR screening of T2 generation mutant lines without transgenic components and the phenotype of the three selected homozygotes lines. a and b, PCR screening of the Cas9 vector region and Hygromycin region in the single plants sampled from the three selected homozygotes lines. M, 2K DNA marker; 1-3, are negative control, distilled water, positive control, 4-16 are strains of JH-C15, 17- 30 are strains of JH-C18, a31-a45 and 31-45 are strains of JH-C31. c, e and g, the phenotype of JH-C15, JH-C18 and JH-C31 at flowering stage; d, f and h, the phenotype of JH-C15, JH-C18 and JH-C31 at harvest stage.
Figure 5.
PCR screening of T2 generation mutant lines without transgenic components and the phenotype of the three selected homozygotes lines. a and b, PCR screening of the Cas9 vector region and Hygromycin region in the single plants sampled from the three selected homozygotes lines. M, 2K DNA marker; 1-3, are negative control, distilled water, positive control, 4-16 are strains of JH-C15, 17- 30 are strains of JH-C18, a31-a45 and 31-45 are strains of JH-C31. c, e and g, the phenotype of JH-C15, JH-C18 and JH-C31 at flowering stage; d, f and h, the phenotype of JH-C15, JH-C18 and JH-C31 at harvest stage.

Figure 6.
qPCR analysis of flowering related genes in the selected homozygotes lines.
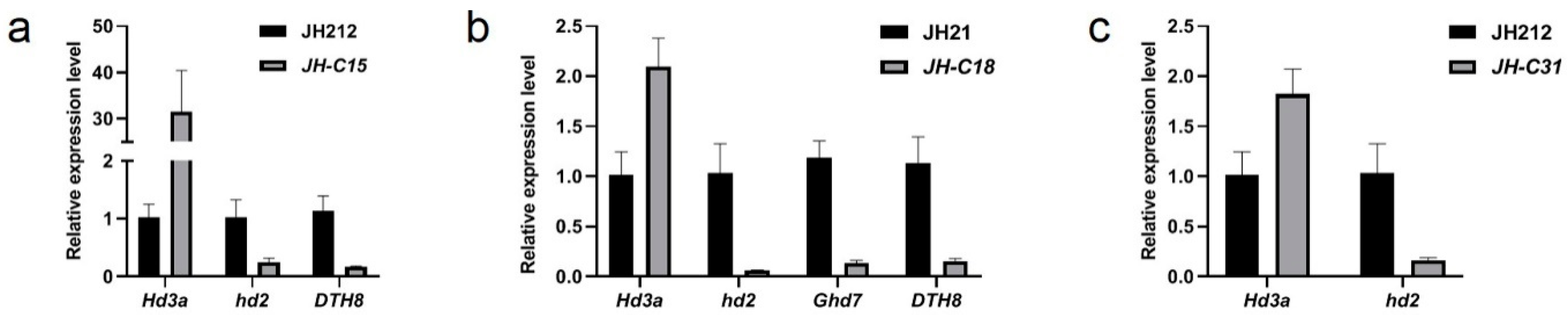
Figure 8.
Analysis of grain shape and agronomic traits of the selected homozygotes lines.
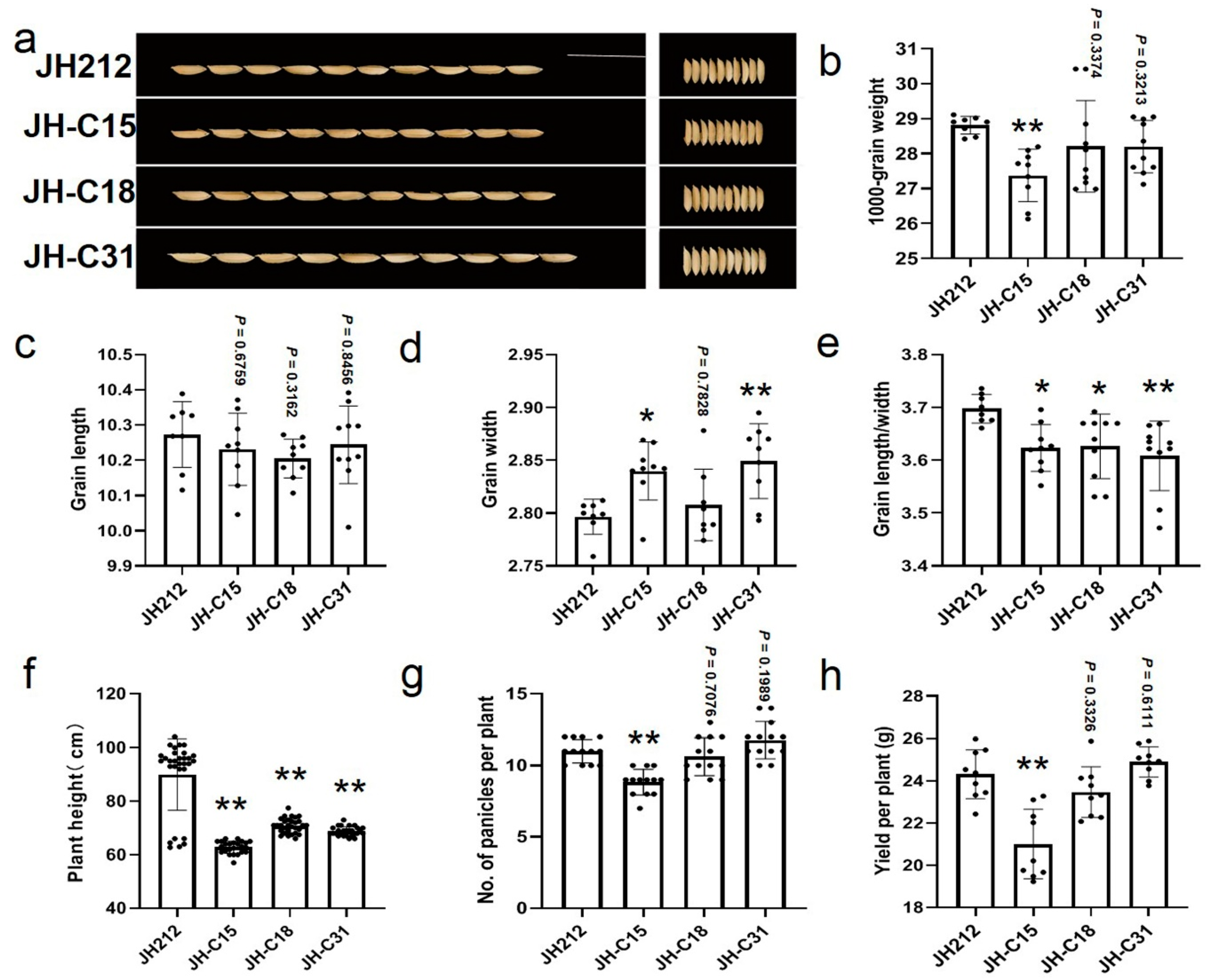

Table 1.
Off-target frequency assessment.
| Target | Position | Strand | GC(%) | Region | Potential off-target sites (Max score) |
Pairing with sg RNA (>= 8 nt) |
|---|---|---|---|---|---|---|
| Hd2 | 738 - 757 | + | 55 | CDS1 | 0.074 | None |
| Ghd7 | 366 - 385 | + | 60 | CDS2 | 0.25 | None |
| DTH8 | 986 - 1005 | + | 55 | CDS1 | 0.238 | None |
| Pi21 | 503 - 522 | + | 55 | CDS1 | 0.133 | None |
| Badh2 | 499 - 518 | + | 60 | CDS2 | 0.083 | None |
Table 2.
Mutation frequency of positive plants in T0 generation.
| Gene | No. of plants | Number of mutant strains | Mutation rate |
|---|---|---|---|
| Hd2 | 30 | 26/30 | 86.66% |
| Ghd7 | 30 | 25/30 | 83.33% |
| DTH8 | 30 | 25/30 | 83.33% |
| Pi21 | 30 | 26/30 | 86.66% |
Table 3.
Genotype combination analysis of T1 homozygous lines.
| Line No.\Gene | Hd2 | Ghd7 | DTH8 | Pi21 | Badh2 | Flowering after sowing (Days) |
|---|---|---|---|---|---|---|
| JH212 | WT | WT | WT | WT | WT | 88.8±3.5 |
| JH-C15 | Allele1: +1bp | WT | Allele1: -4bp | Allele3: +1bp | WT | 57.38±2.75** |
| JH-C17 | Allele1: +1bp | Allele1: +1bp | WT | Allele6: -2bp | WT | 53.22±2.75** |
| JH-C18 | Allele6: -46bp | Allele1: +1bp | Allele1: -1bp | Allele1: +1bp | WT | 61.4±2.25** |
| JH-C19 | Allele2: +1bp | Allele7: -28bp | WT | Allele2: +1bp | WT | 56.55±3.25** |
| JH-C22 | Allele2: +1bp | Allele1: +1bp | WT | Allele9: -19bp | WT | 56.55±4.25** |
| JH-C23 | Allele3: +1bp | Allele5: -5bp | WT | Allele2: +1bp | WT | 50.5±2.75** |
| JH-C24 | Allele3: +1bp | Allele4: -4bp | WT | Allele9: -19bp | WT | 55.5±3.25** |
| JH-C26 | Allele1: +1bp | Allele5: +1bp | WT | Allele6: -2bp | WT | 50.5±2.25** |
| JH-C27-5 | Allele2: +1bp | WT | WT | Allele7: -4bp | WT | 51.55±4.25** |
| JH-C27-2 | WT | Allele7: -28bp | WT | Allele7: -4bp | WT | 52.55±3.25** |
| JH-C28-5 | Allele4: +1bp | Allele2: +1bp | WT | Allele8: -7bp | WT | 60.55±2.75** |
| JH-C31 | Allele5: -5bp | WT | WT | Allele4: -1bp | WT | 59.07±2.75** |
| JH-C32 | Allele2: +1bp | WT | WT | Allele4: -1bp | WT | 60.55±3.25** |
| JH-C34 | Allele5: -5bp | Allele5: -5bp | WT | Allele5: -1bp | WT | 63.55±2.75** |
| JH-C37 | Allele3: +1bp | Allele3: -3bp | WT | Allele2: +1bp | WT | 59.55±3.25** |
| JH-C38 | Allele2: +1bp | Allele8: -45bp | WT | Allele9: -19bp | WT | 55.5±3.75** |
| JH-C40 | Allele4: +1bp | Allele3: -3bp | WT | Allele4: -1bp | WT | 60.55±2.75** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



