
Submitted:
07 May 2024
Posted:
08 May 2024
You are already at the latest version
Abstract
Uropathogenic E. coli (UPEC) is the main etiologic agent of urinary tract infections (UTI) and possesses virulence and resistance determinants commonly harbored in mobilizable genetic elements such as plasmids or pathogenicity islands (PAI). UPEC belongs to the group of Ex-traintestinal pathogenic E. coli, however the existence of hybrid strains has been described; these possess both diarrheagenic (DEC) and extraintestinal (ExPEC) E. coli traits and are considered as "hypervirulent strains," which represents a significant health problem. This study aimed to de-termine the prevalence of UPEC PAI, ExPEC sequence types (ST), DEC genes, carbapenemase and extended-spectrum β-lactamase (ESBL) phenotypes, fluoroquinolones/aminoglycosides resistance genotypes and the presence of plasmids in 40 clinical isolates previously characterized as UPEC. A 72.5% of the isolates presented PAI, mainly PAI IV536 (53%). Sixty-five percent of the β-lactam-resistant isolates were positive for ESBL phenotype, and 100% of the car-bapenem-resistant isolates were carbapenemase producers. The most prevalent ESBL-associated gene was blaCTX-M-2 (60%), while among isolates resistant to fluoroquinolones and aminoglycosides the most frequent resistance gene was aac(6')Ib (93%). Plasmid bands were present in 57% of the isolates. Seventy percent belonged to the ST131 clonal group. The presence of molecular markers associated with DEC pathotypes was detected in 20 isolates; 60% were classified as hybrid pathotypes. These data suggest, in addition to an important pathogenic potential, the existence of hybrid pathotypes in clinical isolates of E. coli causing UTI in the Mexican population.
Keywords:
Urinary tract infection
; ERIC
; Plasmids
; ESBL
; Pathogenicity islands
; Hybrid pathotypes
1. Introduction
E. coli is an important human pathogen that can cause infectious processes of varied severity in different anatomical sites. Its remarkable genomic plasticity has allowed it to adapt to intestinal and extraintestinal environments. According to its genetic content and anatomical site of infection, it has been divided into two large groups: The diarrheagenic E. coli (DEC) group, which includes at least six different pathotypes, and the extraintestinal pathogenic E. coli (EXPEC) group, which includes Uropathogenic E. coli, the main etiological agent of urinary tract infections (UTI) [1,2,3].
UTIs represent a critical health problem in Mexico since they are the second cause of morbidity, with more than three million cases per year and a 70% of prevalence in women [4]. UPEC possesses a wide variety of virulence factors commonly harbored within large DNA segments called pathogenicity islands (PAI). PAIs are mobilizable genetic elements (MGE) that promote the horizontal transfer of important virulence determinants between isolates. To date, it is known the existence of at least thirteen PAIs in the prototype strain of UPEC CFT073 (PAI I-XIII CFT073), seven in UPEC 536 (PAI I-VII 536, and two in UPEC J96 (PAI I-II J96) [5,6,7,8,9]. The genetic charge of these PAIs has been described, showing genes associated with adherence such the pyelonephritis-associated pilus operon (papA-K), F1C pilus (sfa), toxins such as alpha-hemolysin (hlyA and its gene operon), the secreted autotransporter toxin (sat), vacuolating autotransporter toxin (vat), cytotoxic necrotizing factor 1 (cnf-1), and uropathogenic specific protein (usp), genes associated with biofilm and immunoevasion such as antigen 43 (agn43), and capsule (kpsM), genes related to iron uptake such as salmochelin (iroN), yersiniabactin (fyuA), enterobactin (iha), and aerobactin (iucD) [10]. These virulence characteristics have allowed UPEC not only to adapt to the environment provided by the urinary tract but also to favor the success of its uropathogenic mechanism.
Another essential feature is the multidrug resistance (MDR) present in most clinical isolates of E. coli recovered from UTI cases. In Mexico, more than 70% of reported clinical isolates of UPEC are classified as MDR and show resistance to antibiotics used to treat UTI. β-lactam antibiotics, aminoglycosides, and fluoroquinolones are among the most implemented antibiotics for primary treatment [4,11,12]. Resistance to these drugs can be mediated by enzymes such as extended-spectrum β-lactamases (ESBL), acetylases or by different mechanisms such as expulsion pumps, and in the case of fluoroquinolones, proteins that protect the target site [13,14]. According to the reports in Mexico, there is a high prevalence of isolates with ESBL producers’ genotypes and phenotypes [4,15,16,17]. These characteristics and virulence factors can be encoded in MGE, mainly plasmids, which favor their spread between strains [18,19,20].
On the other hand, the existence of heteropathogenic E. coli strains and hybrid pathotypes has been documented; these are important because clinical isolates can possess virulence characteristics of more than one E. coli pathotype, which has led to consider them as hypervirulent strains and can cause more severe infectious processes [21,22].
despite its importance, information focused on the virulence or resistance characteristics of UPEC and the prevalence of hybrid pathotypes causing UTI in Mexico is scarce. Therefore, in this work, we detected the presence of MGE involved with virulence (PAI) in UPEC, the presence of plasmid possibly associated with their antimicrobial resistance, as well as ESBL and carbapenemase production phenotypes in 40 clinical isolates of E. coli, previously identified as the etiological agent of UTI in Mexican population. The presence of molecular markers associated with DEC pathotypes, sequence types (ST), and their adherence patterns were also determined. Finally, phylogenetic analysis was also performed by detection of ERIC (Enterobacteriaceae Repetitive Intergenic Consensus) profiles.
2. Materials and Methods
2.1. Clinical Isolates
Urine samples from adult women with UTI from Sonora, Mexico were obtained. The samples (midstream mictions) were collected in sterile containers and kept at 4°C until they were processed in a time not exceeding 3-4 hours. The samples were inoculated on nutrient agar to CFU/mL count and McConkey agar to identify their colonial morphology. Biochemical identification was carried out by indole, methyl red, Vogues-Prosakuer, and Simmons citrate (IMViC) tests and included: motility, lysine deamination-decarboxylation, urease production, ornithine decarboxylation and sugar fermentation (glucose, lactose, and sucrose). Additionally, microorganisms biochemically identified as E. coli were molecularly confirmed by amplification of the ybbW gene encoding for a particular allantoin receptor of E. coli [23,24]. Information about the clinical characteristics of patients, results for urinalysis, urine cultures, and biochemical or molecular identification of analyzed isolates were previously reported [10].
2.2. Genomic DNA Extraction
A single colony of each isolate was inoculated in 3 mL of Müller-Hinton broth (MHB) and incubated for 24 h at 37°C with constant shaking (150 rpm). Genomic DNA extraction was performed by the alkaline lysis method, according to previously established protocols [25] with some modifications. Briefly, 500 µL of the pre-culture was centrifuged at 16,200 xg for 10 min, subsequently 100 µL of a lysozyme (from Sigma-Aldrich; L6876) solution at 20 mg/mL was added, the mixture was vortexed for one minute and then 5 µL of RNAse A (Sigma-Aldrich; R5500) was added, the mixture was incubated for 1 hour at 37°C. Subsequently, 50 µL of 20X SDS (Sodium Dodecyl Sulfate from Sigma-Aldrich; L3771-500G) was added and homogenized for 1 minute, 200 µL of 5M NaCl was added, vortexed for 30 seconds, the extracts were kept on ice for 5 minutes and subsequently centrifuged at 16,200 xg for 20 minutes and 4°C. The supernatant was transferred to a new sterile tube and 700 µL of cold 2-propanol (from Sigma-Aldrich; I9516) was added, the tube was manually shaken for 5 minutes and centrifuged again for 15 minutes at 16,200 xg and 4°C, the supernatant was discarded, and the DNA pellet was allowed to dry at room temperature for 1 hour and then resuspended in 200 µL of nuclease-free water. The extraction product was resolved by electrophoresis in an 0.7% agarose gel, stained with ethidium bromide solution (10 mg/mL) and visualized with a UVP photodocumenter (AnalytikJena, USA) to determine the integrity of the DNA.
2.3. Pathogenicity Islands (PAI) Detection
The presence of eight pathogenicity islands was detected by PCR using the previously established oligonucleotides and conditions reported by Sabate (Supplementary Material 1) [8]. PCR products were resolved on 2% agarose gels and stained with GelStar (Lonza, Morrinstown, NJ, USA). Each PCR reaction was performed using a GoTaq® Green Master Mix (PROMEGA, USA) following the manufacturer guidelines.
2.4. Plasmid DNA Extraction.
Plasmid DNA extraction was performed using previously reported methods [25]. The extraction product was resolved on 2% agarose gel electrophoresis, stained with ethidium bromide solution (10 mg/mL) and visualized with an UVP photodocumenter (AnalytikJena, USA). The band profiles were analyzed with gel analyzer software 19.1 (www.gelanalyzer.com) using a 1 kb molecular weight marker to normalize the bands and obtain their sizes. For larger plasmids, we used as a control the E. coli strain EDL933, which has a 92 kb plasmid, which allowed to normalize the band sizes.
2.5. Detection of Antibiotic Resistance Associated Genes
The presence of genes related to the main β-lactamase enzymes that confer resistance to broad-spectrum β-lactams was detected; likewise, the presence of three DNA fragments associated with resistance to fluoroquinolones was identified. The genes associated with resistance to β-lactams were: blaCTX-M-1 and 8, blaCTX-M-9, blaCTX-M-2, blaCTX-M-15, blaTEM, and blaSHV. While those associated with resistance to fluoroquinolones and aminoglycosides were qepA, aac(6')-Ib (Also related with cross-resistance to aminoglycosides) and qnrB. The oligonucleotide sequences and conditions were previously reported (Supplementary Material 1) [26,27,28,29,30]. PCR products were resolved in a 2% agarose gel and stained with GelStar (Lonza, Morrinstown, NJ, USA).
2.6. Extended Spectrum β-lactamase (ESBL) Producing Phenotype
Previously characterized cefotaxime-, ceftazidime-, ceftriaxone-, aztreonam-, or cefepime-resistant isolates [10] were selected to evaluate their ability to produce ESBL. Determination of the ESBL phenotype was performed by the double-disk diffusion method, using clavulanic acid as an inhibitor. For each suspected isolate, a bacterial suspension adjusted to 0.5 on the McFarland scale was prepared in Müeller-Hinton broth; Müeller-Hinton agar plates were inoculated with the adjusted suspensions with a sterile swab, allowed to dry for 5 minutes, and the antibiotics discs to be evaluated were placed. The discs were placed at 1.5 cm from the inhibitor center to center. The presence of an inhibition halo, indicative of synergy, between inhibitor and any of the β-lactams evaluated was considered positive for the phenotype. Additionally, negative results were confirmed following the guidelines established by CLSI [31] for the determination of ESBL phenotypes in Enterobacterales; for this purpose, Müeller-Hinton agar plates were inoculated with the isolates and discs of each β-lactam to be evaluated were placed in the presence and absence of 10 µg of clavulanic acid, a difference of 5 mm in inhibition halo diameter between antibiotics with clavulanate and without clavulanate indicated a positive ESBL phenotype.
2.7. Carbapenemase Producing Phenotype
For the detection of enzymes with carbapenemase activity, only isolates resistant to carbapenems were selected [10]. This detection was performed following the criteria established by CLSI [31] using the modified carbapenems inactivation method (mCIM); for this purpose, tubes of TSB broth were inoculated with the isolate in question, the suspension was homogenized with a vortex for 10-15 seconds; with sterile forceps, a single disc of carbapenem (10 µg) was immersed in the bacterial suspension and incubated at 37°C for 4 hours. Subsequently, a 0.5 McFarland’s adjusted suspension of E. coli strain ATCC 25922 (carbapenem sensitive) was prepared in a sterile saline solution. The adjusted inoculum was mass seeded with a sterile swab on a Müeller-Hinton agar plate and allowed to dry for 10 minutes. The carbapenem discs were extracted from the problem strain inoculum, drained, and plated onto the MH agar plate previously inoculated with E. coli ATCC 25922. The plate was incubated at 37°C for 18-24 h, and the results were interpreted according to CLSI, considering a difference of ≥5 mm between hydrolyzed antibiotic vs non-hydrolyzed antibiotic as a positive phenotype.
2.8. Bacterial Adherence Patterns to HeLa Cells
Adherence assays were performed in twenty clinical isolates according to previously reported protocols [10]. The cell lineage used was HeLa ATCC CCL-2 which is a Cervix Cancer cell line; cells were grown in culture plates with DMEM (Dulbecco’s Modified Eagle Medium) with 5% fetal bovine serum (D5F) (GIBCO, USA) and incubated at 37°C with 5% CO2 until sub-confluence of 70-90%. Once grown, the cells were washed with sterile phosphate buffer solution (PBS) and trypsinized for two minutes to detach the monolayer. Subsequently, ten milliliters of fresh D5F were added and centrifuged for seven minutes at 500x g to remove the trypsin and to obtain the cell pellet, the culture medium was decanted, and then 3 mL more was added to resuspend the cells. One hundred microliters of the suspension were taken for cell counting in a hemocytometer chamber. Once the counting was done, sterile coverslips were placed in each well of a six-well polystyrene plate with 2 mL of D5F, and a suspension of 50,000 HeLa cells was added. The plate was incubated for 24 hours at 37°C and 5% CO2 until 70-90% confluence.
A bacterial suspension adjusted to 0.5 on the McFarland scale (1X108 bacteria per milliliter) was prepared from a 24 h pre-culture in Luria-Bertani broth (LB) of each clinical isolate to be evaluated; the adjustment was made in D5F broth without antibiotic. Once prepared, 15 µL of the adjusted inoculum was added to each well of the plate with HeLa cells (MOI: 30:1 Bacteria:Hela). The plate was incubated for three hours at 37°C and 5% CO2. Then, the monolayer was washed with sterile PBS to remove unattached bacteria. This last procedure was performed three times. The cells were then fixed with methanol for ten minutes, allowed to dry at room temperature, and stained with Giemsa for fifteen minutes. The coverslips were removed and mounted on slides to count the adherent bacteria per HeLa cell. The adherence profile for each clinical isolate was also determined by lightfield microscopy with the 100x objective. E. coli EDL933 (EHEC), which is characterized by a localized adherence pattern, was used as a control strain,
2.9. Pathotypes Detection
The presence of genes related to specific DEC pathotypes was detected in twenty clinical isolates by PCR according to primer sequences and conditions previously reported [32,33,34,35] (Supplementary Material 1). Hybrid pathotypes were classified as those isolates that simultaneously presented any of the UPEC associated virulence genes or PAIs in combination with any of the DEC pathotypes associated genes (daaE, eaeA, bfpA, pCVD432). The daaE gene is associated with Diffuse Adherent E. coli (DAEC) and diffuse adherent pattern, eaeA and bfpA are genes related to Enteropathogenic E. coli (EPEC) and localized adherent pattern. In contrast, plasmid pCVD432 is associated with Enteroaggregative E. coli (EAEC) and aggregative adherent pattern.
2.10. Multiplex PCR for Dominant Sequence Types (STs) in EXPEC Isolates
The STs (ST131, ST69, and ST73 from clinical isolates were identified by multiplex PCR reactions according to previously reported oligonucleotides and conditions [36] (Supplementary Material 1).
2.11. Enterobacteriaceae Repetitive Intergenic Consensus (ERIC) PCR
Clonal relationship was detected by amplification of ERIC (Enterobacteriaceae Repetitive Intergenic Consensus) fragments by PCR (ERIC-PCR) using previously standardized oligonucleotides [37]. Each PCR reaction was performed using a mixture containing 2 µL of buffer, 0.5 µL of a mixture of dNTP (10 mM), 1.5 µL of MgCl2 (25 mM), 1 µL of each primer (20 µM), 0.1 µL of GoTaq® Flexi DNA polymerase (PROMEGA, USA), 1.5 µL of template DNA (50-75 ng) and nuclease-free water until a final volume of 15.5 µL of reaction was obtained. The mixture was submitted to one cycle at 95°C for five minutes, followed by 40 cycles at 95°C for 1 min, 50°C 1 minute, 72°C for 8 min and one more cycle of 72°C for 16 minutes. PCR products were resolved on 1.5% agarose gels and stained with GelStarTM (Lonza, USA). Band profiles were analyzed using GelAnalyzer 19.1 software (available on http://www.gelanalyzer.com/?i=1) to determine the sizes of each product, and subsequently, dendrograms were constructed by the unweighted paired-pairing by arithmetic mean (UPGMA) method and the DICE algorithm to detect similarities between ERIC patterns, dendogram was constructed with iTOL 6.8 [38].
2.11. Statistical Analysis
For the statistical analysis, Pearson's correlation test was implemented. Analysis was performed with JASP software version 0.16.1.
3. Results
3.1. Clinical Isolates of E. coli from UTI Cases in Mexico Harbor Pathogenicity Islands
The presence of PAI was observed in 29 (72.5%) of the forty analyzed clinical isolates. One isolate has six PAI (isolate 26). Three of these isolates presented five of the identified PAIs; three showed only one PAI, and the other thirty-three -five isolates showed between two and four PAIs.
The most prevalent pathogenicity island was PAI IV 536 (72.41%), followed by PAI II CFT073 (48.27%) and PAI I J96 (48.27%). The eight PAI investigated were found in the E. coli strains analyzed. (Figure 1).
The PAI profiles of each isolate are shown in Table 1. Statistical analysis was performed to detect a possible correlation between the co-occurrence of PAI in the clinical isolates, and a strong positive correlation was observed in the co-occurrence of PAI I 536-PAI I J96 (r = 0. 3126; p = 0.02), PAI I 536-PAI I CFT073 (r = 0.3725; p = 0.009), PAI III 536-PAI II CFT073 (r = 0.4871; p = 0.001), PAI IV 536-PAI II CFT073 (r = 0. 4881; p = 0.001), PAI I J96-PAI I CFT073 (r = 0.3698; p = 0.009), PAI I 536-PAI II J96 (r = 0.3059; p = 0.02) and PAI II J96-PAI I CFT073 (r = 0.5412; p = <0.001).
3.2. Clinical Isolates of E. coli Are ESBL Producers
The antibiotic resistance profiles of the analyzed clinical isolates were previously reported [10] and are shown in supplementary material 2. In total, 34 (85%) isolates were resistant to third or fourth generation cephalosporins or to the tested monobactam antibiotics; these 34 isolates were selected to detect the ESBL-producing phenotype. Twenty-two (65%) of the 34 selected clinical isolates exhibited ESBL-producing phenotype. The produced enzymes hydrolyzed one to five of the evaluated antibiotics. The most frequent substrate profiles are shown in Table 2. Interestingly, five of the isolates (ID: 5, 27, 31, 35, and 36), despite being resistant to most of the β-lactam antibiotics tested (they were resistant to cefoxitin, third generation cephalosporins and at least one of the two β-lactams with inhibitors tested, did not show an ESBL-producing phenotype.
As ExPECted a statistical significance was observed in co-occurrence of ESBL phenotypes and resistance to β-lactam antibiotics, mainly between ESBL producers and resistance to CTX/CFZ (r = 0.49, p = 0.005), CRO (r = 0.41, p = 0.017), ATM (r = 0.46, p = 0.009); CFZ and CRO (r = 0.71, p = <0.001), FEP (r = 0.49, p = 0.005); FEP and ATM (r = 0.68, p = <0.001) . On the other hand, when compared virulence with resistance characteristics in this study, a negative correlation between positive ESBL phenotype for CTX and prevalence of PAI I J96 (r = -0.40, p = 0.05) and PAI IV 536 (r = -0.28; p = 0.04) was observed.
3.3. Carbapenem Resistance is Mediated by Carbapenemases Production
Five of the analyzed clinical isolates were resistant to carbapenems (ID: 7, 27, 35, 39, and 40, Supplementary Material 2). These isolates were selected to detect the carbapenemase (CAR) production phenotype using the mCIM method previously described. Interestingly, all evaluated isolates were CAR positives, three for imipenem and two for meropenem. Two isolates (27 and 35) were classified previously as a non-producer ESBL phenotype with CX and AMC resistance, while two more isolates (39 and 40) were also ESBL producers, and the last clinical isolate (7) was resistant only to carbapenems (Table 2 and Supplementary material 2).
3.4. Clinical Isolates Harbored Antibiotic Resistance Genes
β-lactams
ESBL Producers, aminoglycosides, and fluoroquinolones-resistant isolates were selected. Thirty-two (80%) were resistant to amikacin or gentamicin, while only fifteen (38%) were fluoroquinolone-resistant (Ciprofloxacin, levofloxacin, or norfloxacin) (Supplementary material 2).
Among twenty-two isolates with ESBL-producing phenotype, we observed the enzyme-associated genes in twenty (91%) of them. The prevalence of each analyzed gene was as follows: blaCTX-M-2 (60%), blaCTX-M-1 and 8 (55%), blaTEM (30%), blaCTX-M-9 (20%), and blaCTX-M-15 (10%). No positive isolates for the blaSHV gene were observed.
On the other hand, despite showing resistance to β-lactams, twelve of the isolates did not exhibit a positive ESBL phenotype. However, they presented genes associated with these enzymes, mainly blaTEM (7 isolates), blaCTX-M-1 and 8 (4 isolates), blaCTX-M-2 (4 isolates), blaCTX-M-9 (2 isolates), and blaCTX-M-15 (1 isolate). Interestingly, within this group of isolates are the five isolates that also showed resistance to cephamycins (cefoxitin) and at least one of the antibiotics with β-lactamase inhibitors (clavulanic acid or sulbactam). The remaining six isolates did not show resistance to the β-lactam antibiotics tested and did not have a production phenotype, but blaCTX-M-1 and 8 (3 isolates), blaTEM (3 isolates), and blaCTX-M-2 (1 isolate) were detected, this is probably due to non-expressed genes or non-functional proteins.
The statistical analysis showed a significance in the correlation between the prevalence of blaCTX-M-2 and ESBL phenotype for CFZ (r = 0.49, p = 0.005), CRO (r = 0.41, p = 0.02). Likewise, a positive correlation was observed for blaCTX-M-15 and CRO (r = 0.4, p = 0.03) and FEP (r = 0.4, p = 0.04). The co-occurrence of blaCTX-M-9 and blaCTX-M-15 was also statistically significant (r = 0.045, p = 0.009).
Aminoglycosides or Fluoroquinolones
Thirty-two (80%) isolates were resistant to at least one of the tested aminoglycosides (AMK and GM, Supplementary Material 2), while fifteen (38%) were resistant to at least one of the evaluated fluoroquinolones (CIP, LVX, and NOR). These isolates were selected to determine the prevalence of genes associated with resistance to these two families of antimicrobials.
Fourteen (93%) of the fifteen isolates resistant to fluoroquinolones showed the presence of at least one of the determined genes, being the acetylase associated gene aac(6')-Ib (93%) the most prevalent, followed by qepA (66%), associated with efflux pumps, and qnrB (60%) related to protective proteins of the target site of antibiotic. On the other hand, the aac(6')-Ib gene was also found in high prevalence among aminoglycoside-resistant isolates (54.2%). A strong positive correlation was observed in the prevalence of aminoglycoside vs fluoroquinolone resistance and each resistance gene analyzed (r ≥ 0.99, p <0.001, Supplementary Material 3 A). Interestingly, also a strong positive correlation between ESBL producer phenotypes, resistance to aminoglycosides, fluoroquinolones, and the presence of previously mentioned genes was observed (r ≥ 0.99, p <0.001) (Supplementary Material 3 A).
Regarding virulence, no statistical significance was observed between the prevalence of PAI and the evaluated antibiotic-resistance genes.
3.5. Plasmids of Clinical Isolates are Correlated with Resistance Genes Prevalence
The presence of plasmid bands was determined in the 40 analyzed clinical isolates. Only twenty-three (57.5%) of the isolates possessed plasmids. The number of bands per isolate ranged from one to eleven, with a mean of four plasmid bands per isolate. The interval band sizes observed, and their prevalence were as follows: 1-5 kb (91.3%), 6-10 kb (39.1%), 11-20 kb (17.4%), 21-30 kb (9%), 31-50 kb (13%), and 101-120 kb (9%). No statistically significant was observed between the presence of plasmids vs the virulence and resistance evaluated features (p >0.05) (supplementary material 3 B). However, a strong positive correlation was found between the prevalence of 51-100 kb with resistance to FEP (r = 0.38, p = 0.01), and 101-120 kb with resistance to LVX (r = 0.33, p = 0.037), FEP (r = 0.69, p <0.001), ATM (r = 0.4, p = 0.01), IMP (r = 0.37, p = 0. 02), with the β-lactamase-producing phenotypes ESBL-FEP (r = 0.37, p = 0.02), ESBL-ATM (r = 0.54, p <0.001), and qepA gene prevalence (r = 0.4, p = 0.01) (supplementary material 3 C).
3.6. ST131 Is the most Prevalent Clonal Group
Within the 40 analyzed isolates, only 30 (75%) showed the presence of molecular markers associated with the determined STs. The 60% of the isolates were classified as belonging to the ST131 clonal group, followed by the ST69 group (13%), and ST73 (3%). No isolates belonging to the ST95 group were observed.
No significant association was shown between the presence of PAI, and the ST groups identified. However, a statistically significant association was observed in the higher prevalence of resistance to fluoroquinolones (p = 0.05) in the ST131 clonal group (Table 3). In this group we also showed a high prevalence of isolates with genes associated with antibiotic resistance, mainly blaCTX-M-1 and 8 (58.4%, p = 0.005), blaCTX-M-9 (17%, p = 0.02), qepA (33.3%, p = 0.001), aac(6')-Ib (46%, p = 0.002), and qnrB (33.3%, p = <0.001). On the other hand, a strong positive correlation was shown in the higher prevalence of the blaTEM gene (r = 0.46, p = 0.002) in the ST69 clonal group.
3.7. Clinical Isolates Show Mixed Adherence Patterns, Including some Associated with Diarrheagenic Pathotypes of E. coli
According to the profile of genes related to adherence of each strain [10], twenty isolates were randomly selected their adherence patterns were determined. The patterns observed were bricks (85%), localized (55%), aggregative (20%), and diffuse (15%) (Figure 2). Interestingly, only eight (40%) of the isolates exhibited single adherence pattern, six of them shown the Tandem bricks pattern, while twelve (60%) exhibited mixed adherence patterns associated with different pathotypes of E. coli and of these twelve, pathogenicity islands related to prototype strains of Uropathogenic E. coli were found in nine (77%) (table 4 and Supplementary Material 4).
These findings and the presence of virulence factors, characteristic of DEC, in the clinical isolates suggest the presence of UPEC-DEC hybrid pathotypes in the study bacterial population.
3.8. Clinical Isolates have Molecular Markers Associated with DEC Pathotypes
Considering the adherence results, molecular markers associated with DEC pathotypes were determined for the 20 clinical isolates.
In 80% of the selected isolates, the bfpA gene was detected, the plasmid-associated gene pCVD432 was found in 30%, the daaE gene in 20%, and the eaeA gene in 5%. Four isolates did not present any of the DEC genes (Table 4). On the other hand, nine isolates presented between 2 and 4 of the genes determined. The presence of the eaeA gene and the absence of bfpA is related to atypical EPEC, in this study, only one isolate harbored the eaeA gene, but this was also positive for bfpA. Interestingly, ten isolates harbored PAI in addition to one or more pathotypes associated genes. These results and virulence factors related to UPEC in analyzed clinical isolates (Supplementary Material 4) indicate the existence of hybrid clinical isolates of E. coli in the Mexican population with UTI, including triple and quadruple hybrid strains (Table 4).
3.9. ERIC Fingerprint Pattern in Clinical Isolates
All analyzed isolates showed between 1 and 12 bands, with a higher prevalence of isolates with 8 (38%) and 6 (20%) bands. The size interval of the bands was from 0.15 kb to 10 kb.
According with the results obtained in the bands analysis and the dendogram generated by applying UPGMA method and the DICE coefficient with a similarity value of 80%, the forty analyzed clinical isolates were distributed in ten different clusters (C-1-C-10) (Figure 3). Only two isolates (Ec-25 and Ec-26 presented the same ERIC profile; both isolates belonged to the ST69 group and did not presented genes associated with other pathotypes different from UPEC. Regarding their virulence characteristics, differences were observed in the profile and number of PAI of each isolate; however, the remaining characteristics associated with resistance were the same.
It was investigated if the presence of the different molecular markers of virulence or resistance and the phenotypes of resistance were associated with any of the different clusters obtained. A positive correlation with statistical significance was found between PAI III536 vs. clinical isolates belonging to cluster 6 (r = 0.44, p = 0.002), resistance to third generation cephalosporins (r = 0.27, p = 0.05), and the ESBL phenotype (r = 0.29, p = 0.04) vs. cluster 8. On the other hand, fluoroquinolone resistance (r = -0.28, p = 0.04), the prevalence of PAI ICFT073 (r = -0.36, p = 0.01), and PAI III536 (r = 0.31, p = 0.03) was negatively correlated with clusters 6, 8, and 10 respectively.
4. Discussion
Urinary tract infections are the second leading cause of morbidity in Mexico. E. coli is the main etiological agent of this disease and possesses a wide variety of genetic virulence and resistance determinants that have allowed it to become a successful pathogen. These genetic characteristics are commonly harbored in pathogenicity islands or plasmids that can be transferred, which promotes the spreading of these important characteristics.
In this work, the presence of molecular markers associated with pathogenicity islands was determined in 40 clinical isolates of E. coli obtained from Mexican women with UTI, previously, these isolates had been classified as highly virulent with important characteristics for the development of both lower and upper UTI, and a co-occurrence of genes that are commonly found harbored in pathogenicity islands was observed [10]. Therefore, in this work we determined the presence of these elements in the previously mentioned isolates, upon analysis we found that 72.5% of them presented PAI associated with UPEC, with a high prevalence of PAI IV of E. coli 536 (53%), followed by PAI II CFT073 (35%) and PAI I J96 (35%). It is important to mention that we only determined the presence of 8 pathogenicity islands, however there are others such as those of UPEC UMN026 (5 PAIs), UPEC UTI 89 (8 PAIs), five more of UPEC 536 and 11 more of UPEC CFT073 (Desvaux et.al. ,2020) that were not identified in this study and that could probably also explain this co-occurrence of virulence-associated genes that we previously reported.
The PAI IV of E. coli 536, also known as high pathogenicity island (HPI), is one of the most stable islands reported in E. coli and is thought to be fixed in the chromosome [8,40], probably being one of the first PAI to be acquired, which could explain its high prevalence in this and other previously reported studies, including in commensal strains, which has led to the assumption that rather than a pathogenicity island it could be a fitness island, however its content of virulence-associated genes makes it an important marker of pathogenicity [6,41]. On the other hand, PAI I of E. coli 536 was the less prevalent, it has been reported that this island together with PAI II 536 and PAI III 536 present higher instability. Interestingly, conditions such as the presence of antimicrobials such as nitrofurantoin, gentamicin, cotrimoxazole, ampicillin and fosfomycin, all implemented in the treatment of UTI in Mexico, induce the expression of integrases on this MGE and their cleavage [42], which could explain the lower prevalence of these PAI in our study.
The prevalence of clinical isolates with PAI found in this study is higher than previously reported in another work performed in Sonora and Puebla, which until now was the only report of these specific pathogenicity islands in our country; however, the prevalence of PAI of E. coli 536 had not been previously described [12]. The prevalence of PAI IV 536 and the other determined islands are similar to those reported by Sabaté in 2006 in clinical isolates of UPEC, however, Sabaté and other authors do not report the presence of E. coli J96 PAIs [6,8,41,43], which could indicate an association to the geographic area or patient group, since in this work and in the previously reported in Mexico both islands were observed in adult females [12], whereas, reports in other countries include male and pediatric population.
PAIs of E. coli 536 harbor genes related to upper UTI, including toxins such as alpha hemolysin and its gene operon (hlyA-D) (PAI I), the pyelonephritis-associated pilus operon (papA-F) (PAI II), S-type fimbriae (sfa) and its gene operon, iron scavengers such as salmochelin and its gene operon (PAI III), yersiniabactin-related genes (PAI IV) [44], and genes associated with capsule (kps) production (PAI V) [45]. This together with the high prevalence of PAI of E. coli CFT073 (isolate characterized as pyelonephritic), PAI of E. coli J96 and their co-occurrence confirm the great genomic plasticity of E. coli and the elevated pathogenic potential of the analyzed clinical isolates. Interestingly, despite its importance, this is the second article in our country that reports the presence of these genetic elements in clinical isolates of E. coli obtained from patients with UTI.
In addition to its virulence and its evident impact on the development of UTI, E. coli possesses a wide variety of antibiotic resistance determinants. In this regard, β-lactam antibiotics, fluoroquinolones, and aminoglycosides represent three of the categories that, depending on patient characteristics, are widely employed in UTI therapeutics. Recently, our working group reported a systematic review in which a high prevalence of resistance to these groups of antimicrobials was observed in Mexico, with fluoroquinolones (58%) having the highest resistance, followed by β-lactam antibiotics (second to fourth generation cephalosporins: 35-57%), and aminoglycosides (26%) [4].
Resistance to this group of drugs is harbored mainly into MGE such as plasmids, and in the case of β-lactams is mediated by genes encoding for extended spectrum β-lactamases (ESBL) [46]. In this sense, in the present study, of the 40 clinical isolates analyzed in 34 resistance to third and fourth generation cephalosporins or monobactams was observed, of these in 22 strains (65%) an ESBL producing phenotype was present and 20 isolates showed genes associated with β-lactamases, being more prevalent blaCTX-M-2 genes (60%), followed by blaCTX-M-1 and 8 (55%) and blaTEM (30%), these results are similar to those reported by other authors in our country with respect to the prevalence of blaCTX-M-1 and blaTEM [47,48,49] however the only report found in Mexico on the frequency of blaCTX-M-2 [50] was considerably lower than the reported in our study. Likewise, a high prevalence of the blaCTX-M-15 gene (33-96%) has been observed in other Mexican states, such as Mexico City and Jalisco (Alcántar-Curiel et al., 2015; Paniagua-Contreras et al., 2019; Rayo Morfín-Otero, Soraya Mendoza-Olazarán, Jesús Silva-Sánchez, Eduardo Rodríguez-Noriega, Jorge Laca-Díaz, Perla Tinoco-Carrillo, Luis Petersen, Perla López, Fernando Reyna-Flores, Dolores Alcantar -Curiel, Ulises Garza-Ramos, 2013; Reyna-Flores et al., 2013), associated with the ST131 clone in our study we observed a reduced prevalence of this gene compared to those studies (10%) even though 70% of our isolates were molecularly identified as belonging to the ST131 clonal group, which could indicate a probable association with the geographical area or patients group.
Five of the analyzed isolates showed resistance to third generation cephalosporins, monobactam aztreonam, and one strain was also resistant to cefepime. Interestingly, these isolates did not present an ESBL-producing phenotype, but their susceptibility profile showed resistance to cephamycins and combined antibiotics (amoxicillin-clavulanic acid and/or ampicillin-sulbactam). This resistance profile is commonly associated with pathogens AmpC β-lactamases producers, mainly those harbored in plasmids (pAmpC), which although in E. coli are infrequent, their existence has been reported in clinical isolates obtained from patients with UTI in other countries [54,55]. In Mexico, there are reports of genes associated with these pAmpC in clinical isolates of UPEC, being CMY (mainly CMY 1, 2, and 23) the most prevalent [53,56]. The potential existence of pAmpC in the clinical isolates analyzed is important due to the resulting complications in the treatment of the infectious process, since its presence is associated with therapeutic failures and due to its location in plasmids, its dispersion is more probable. Therefore, it would be interesting and epidemiologically important to determine molecular markers associated with plasmidic AmpC enzymes in the clinical isolates mentioned.
Regarding resistance to aminoglycosides and quinolones, this was observed in 80% and 38%, respectively, of the analyzed isolates. The occurrence of these phenotypes was coincident with the presence of genes associated with resistance to these groups of antimicrobials, with a high prevalence of the aac(6')-Ib gene (93%), which is related to cross-resistance to fluoroquinolones and aminoglycosides [57,58] followed by qepA (60%), and qnrB (30%). The aac(6')-Ib gene is also localized in integrons, mainly class 1 integrons which are present in plasmids [59]. Therefore, and considering the high prevalence of resistance to fluoroquinolones in our country, which exceeds 50% according to the reports available until 2021 [4] would be interesting to determine the presence of these elements in the analyzed strain.
A strong positive correlation was observed between ESBL production with each of the determined genes and the resistance phenotypes to aminoglycosides and fluoroquinolones; this could be because these genes and most of those associated with ESBL are commonly harbored in plasmids, and there are reports of their co-existence [60,61]. In addition, when analyzing the plasmid profiles, a strong positive correlation was found between each resistance gene and phenotypes determined with plasmid bands of variable sizes. This reinforces our hypothesis regarding the co-occurrence of these characteristics in the population analyzed; however, further studies are needed to confirm it since many of the observed bands could be plasmid isoforms.
In addition, it was observed that five of the isolates were carbapenemase producers, and due to their resistance profile and the number of antibiotic categories to which they are resistant, they were previously classified as multidrug-resistant (Supplementary Material 2). This result is alarming, considering that carbapenems are implemented as a last resource in the treatment of infectious processes and reflect the urgency of searching for therapeutic alternatives.
On the other hand, 60% of the analyzed isolates in this study belong to the ST131 group. This clonal group of E. coli is globally distributed and is important due to its significant genetic content associated with virulence and resistance. It has been reported that ST131 strains are predominantly non-susceptible to fluoroquinolones, in addition to presenting co-resistance to antimicrobials such as aminoglycosides, co-trimoxazole, and β-lactams [62,63]. In our study, isolates from the ST131 clonal group showed higher resistance to all evaluated antibiotic categories. However, only resistance to fluoroquinolones was statistically correlated.
In the same way, the genes associated with ESBL, except for blaTEM and all those related to fluoroquinolones and aminoglycoside resistance, were more prevalent in this clonal group. Our results agree with those previously reported in countries such as Turkey [64] concerning the association of blaCTX-M-1 and blaCTX-M-15 with the ST131 group. However, we observed a low prevalence of blaCTX-M-15 (10%) in the analyzed isolates, which differs from that reported in Turkey (38%), in India by Hussain (100%) [65], and in other states of Mexico by Reyna-Flores [52] (97%), these results suggest that the prevalence of blaCTX-M-15 is more influenced by the geographical area and the environment to which the pathogen is exposed than the clonal group to which they belong and that, considering the higher prevalence of blaCTX-M-2 (60%), as well as the prevalence of blaCTX-M-9 (55%) and blaTEM (30%) the ST131 clonal group presents a greater diversity of genes associated with β-lactam resistance than that reported in the previously mentioned works.
Finally, the existence of E. coli isolates that possess virulence determinants of more than one pathotype has been previously described in other countries [66,67]; these strains are considered hypervirulent and represent an important health risk. In this study we determined the presence of molecular markers associated with intestinal pathotypes of E. coli, the genes were selected according to the observed adherence profiles. There is no characteristic adherence pattern for UPEC, however in previous studies in Mexico, it has been reported that some clinical isolates of UPEC can adopt an adherence pattern that has been called "train wagon” [68] in HeLa cells, which is like what we observed and identified as tandem bricks pattern.
Of the 20 isolates analyzed, genetic determinants of intestinal pathotypes were found in 16 (80%), being mainly the bfpA gene, which is associated with the localized adherence pattern and enteropathogenic E. coli, this result together with the existence of UPEC virulence factors such as P-type pili (papC, papG-II), S-type pili (sfa). aerobactin (iucD, iutA), yersiniabactin (fyuA), capsular antigens (kpsM), and vacuolizing autotransporter toxin (vat) that were previously reported and that are considered as molecular markers of ExPEC-UPEC [69], in addition to the presence of pathogenicity islands associated with UPEC in these isolates, confirm the existence of hybrid pathotypes in the Mexican population with UTI, so it would be important to determine their implications in the clinical and diagnosis of the disease.
5. Conclusions
The clinical isolates analyzed showed virulence characteristics attributed to intestinal pathotypes and UPEC, which demonstrates the presence of hybrid pathotypes (UPEC/DEC) of E. coli causing urinary tract infections in our country. To our knowledge, this is the first article reporting the existence of UPEC/DEC hybrid pathotypes in Mexico. These isolates belong mainly to the ST131 clonal group and harbor a considerable number of genes associated with resistance to fluoroquinolones, aminoglycosides and β-lactams, which are antibiotics widely employed for the treatment of UTI. Furthermore, the high prevalence of ESBL-producing strains in our study and the presence of potential pAmpC- or carbapenemase-producing isolates is alarming because of their impact on antimicrobial therapy.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
M.G.B.-M.: conceptualization, methodology, formal analysis, investigation, and writing—original draft; P.M.P.: writing—review and editing and resources; B.O: writing-review and editing; Y.E.M.: writing—review and editing; M.M.A.H.: writing—review and editing; M.L.A.A.: writing—review and editing and resources; B.D.M: writing-review and editing; E.B.M.: writing—review and editing; E.B.V.: writing—review and editing; D.V.: writing—review and editing, resources, project administration, and supervision.
Funding
This research was funded by Departamento de Ciencias Químico-Biológicas de la Universidad de Sonora, Campus Caborca.
Informed Consent Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Institutional Ethics Committee of Universidad de Sonora (CEI-UNISON) (Registry number 07.2019. 12 March 2019). Informed consent was obtained from all subjects involved in the study.
Acknowledgments
The authors are pleased to acknowledge the Departamento de Ciencias Químico-Biológicas y Agropecuarias de la Universidad de Sonora, Campus Caborca. Manuel G. Ballesteros-Monrreal had a CONACYT Fellowship during the performance of this work (Scholarship No. 617232).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Croxen, M.A.; Law, R.J.; Scholz, R.; Keeney, K.M.; Wlodarska, M.; Finlay, B.B. Recent Advances in Understanding Enteric Pathogenic E. coli. Clin Microbiol Rev 2013, 26, 822–880. [Google Scholar] [CrossRef] [PubMed]
- Russo, T.A.; Johnson, J.R. Proposal for a New Inclusive Designation for Extraintestinal Pathogenic Isolates of E. coli: ExPEC. J Infect Dis 2000. [Google Scholar] [CrossRef]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary Tract Infections: Epidemiology, Mechanisms of Infection and Treatment Options. Nat Rev Microbiol 2015, 13, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros-Monrreal, M.G.; Mendez-Pfeiffer, P.; Barrios-Villa, E.; Arenas-Hernández, M.M.P.; Enciso-Martínez, Y.; Sepúlveda-Moreno, C.O.; Bolado-Martínez, E.; Valencia, D. Uropathogenic E. coli in Mexico, an Overview of Virulence and Resistance Determinants: Systematic Review and Meta-Analysis. Arch Med Res 2023. [Google Scholar] [CrossRef] [PubMed]
- Gyles, C.; Boerlin, P. Horizontally Transferred Genetic Elements and Their Role in Pathogenesis of Bacterial Disease. Veterinary Pathology 2014, 51, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Samei, A.; Haghi, F.; Zeighami, H. Distribution of Pathogenicity Island Markers in Commensal and Uropathogenic E. coli Isolates. Folia Microbiol (Praha) 2016, 61, 261–268. [Google Scholar] [CrossRef]
- Schmidt, H.; Hensel, M. Pathogenicity Islands in Bacterial Pathogenesis. Clin Microbiol Rev 2004, 17, 14–56. [Google Scholar] [CrossRef] [PubMed]
- Sabate, M.; Moreno, E.; Perez, T.; Andreu, A.; Prats, G. Pathogenicity Island Markers in Commensal and Uropathogenic E. coli Isolates. Clin Microbiol Infect 2006, 12, 880–886. [Google Scholar] [CrossRef]
- Desvaux, M.; Dalmasso, G.; Beyrouthy, R.; Barnich, N.; Delmas, J.; Bonnet, R. Pathogenicity Factors of Genomic Islands in Intestinal and Extraintestinal E. coli. Front Microbiol 2020, 11. [Google Scholar] [CrossRef]
- Ballesteros-Monrreal, M.G.; Arenas-Hernández, M.M.P.; Barrios-Villa, E.; Juarez, J.; Álvarez-Ainza, M.L.; Taboada, P.; De la Rosa-López, R.; Bolado-Martínez, E.; Valencia, D. Bacterial Morphotypes as Important Trait for Uropathogenic E. coli Diagnostic; a Virulence-Phenotype-Phylogeny Study. Microorganisms 2021, 9, 2381. [Google Scholar] [CrossRef]
- Amábile-Cuevas, C.F. Antibiotic Resistance in Mexico: A Brief Overview of the Current Status and Its Causes. J Infect Dev Ctries 2010, 4, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros-Monrreal, M.G.; Arenas-Hernández, M.M.P.; Enciso-Martínez, Y.; Martinez de la Peña, C.F.; Rocha-Gracia, R. del C.; Lozano-Zarain, P.; Navarro-Ocaña, A.; Martínez-Laguna, Y.; de la Rosa-López, R. Virulence and Resistance Determinants of Uropathogenic E. coli Strains Isolated from Pregnant and Non-Pregnant Women from Two States in Mexico. Infect Drug Resist 2020, 13, 295–310. [Google Scholar] [CrossRef]
- Galindo-Méndez, M. Reservoirs of CTX-M Extended Spectrum β -Lactamase-Producing Enterobacteriaceae in Oaxaca, Mexico. J. Microbiol. Exp 2019, 7, 43–47. [Google Scholar] [CrossRef]
- Gonçalves, L.F.; De Oliveira Martins, P.; De Melo, A.B.F.; Da Silva, R.C.R.M.; De Paulo Martins, V.; Pitondo-Silva, A.; De Campos, T.A. Multidrug Resistance Dissemination by Extended-Spectrum β-Lactamase-Producing E. coli Causing Community-Acquired Urinary Tract Infection in the Central-Western Region, Brazil. J Glob Antimicrob Resist 2016, 6, 1–4. [Google Scholar] [CrossRef]
- Miranda-Estrada, L.I.; Ruíz-Rosas, M.; Molina-López, J.; Parra-Rojas, I.; González-Villalobos, E.; Castro-Alarcón, N. Relación Entre Factores de Virulencia, Resistencia a Antibióticos y Los Grupos Filogenéticos de E. coli Uropatógena En Dos Localidades de México. Enferm Infecc Microbiol Clin 2017, 35, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Zavala-Cerna, M.G.; Segura-Cobos, M.; Gonzalez, R.; Zavala-Trujillo, I.G.; Navarro-Perez, S.F.; Rueda-Cruz, J.A.; Satoscoy-Tovar, F.A. The Clinical Significance of High Antimicrobial Resistance in Community-Acquired Urinary Tract Infections. Canadian Journal of Infectious Diseases and Medical Microbiology 2020, 2020, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lagunas-Rangel, F.A. Antimicrobial Susceptibility Profiles of Bacteria Causing Urinary Tract Infections in Mexico: Single-Centre Experience with 10 Years of Results. J Glob Antimicrob Resist 2018, 14, 90–94. [Google Scholar] [CrossRef]
- Basu, S.; Mukherjee, M. Incidence and Risk of Co-Transmission of Plasmid-Mediated Quinolone Resistance and Extended-Spectrum β-Lactamase Genes in Fluoroquinolone-Resistant Uropathogenic E. coli: A First Study from Kolkata, India. J Glob Antimicrob Resist 2018, 14, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Randall, L.P.; Cooles, S.W.; Osborn, M.K.; Piddock, L.J. V; Woodward, M.J. Antibiotic Resistance Genes, Integrons and Multiple Antibiotic Resistance in Thirty-Five Serotypes of Salmonella enterica Isolated from Humans and Animals in the UK. Journal of Antimicrobial Chemotherapy 2004, 53, 208–216. [Google Scholar] [CrossRef]
- Chávez-Jacobo, V.; Ramírez-Díaz, M.; Silva-Sánchez, J.; Cervantes, C. Resistencia Bacteriana a Quinolonas: Determinantes Codificados En Plásmidos. REB. Revista de educación bioquímica 2015. [Google Scholar]
- Bielaszewska, M.; Schiller, R.; Lammers, L.; Bauwens, A.; Fruth, A.; Middendorf, B.; Schmidt, M.A.; Tarr, P.I.; Dobrindt, U.; Karch, H.; et al. Heteropathogenic Virulence and Phylogeny Reveal Phased Pathogenic Metamorphosis in E. coli O2: H6. EMBO Mol Med 2014, 6, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Moreno, E.; Caporal-Hernandez, L.; Mendez-Pfeiffer, P.A.; Enciso-Martinez, Y.; De la Rosa López, R.; Valencia, D.; Arenas-Hernández, M.M.P.; Ballesteros-Monrreal, M.G.; Barrios-Villa, E. Characterization of Diarreaghenic E. coli Strains Isolated from Healthy Donors, Including a Triple Hybrid Strain. Antibiotics 2022, 11, 833. [Google Scholar] [CrossRef]
- Walker, D.I.; McQuillan, J.; Taiwo, M.; Parks, R.; Stenton, C.A.; Morgan, H.; Mowlem, M.C.; Lees, D.N. A Highly Specific E. coli QPCR and Its Comparison with Existing Methods for Environmental Waters. Water Res 2017, 126, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Carreón, E. Estudio Molecular de La Resistencia y Virulencia de Cepas de E. coli Productoras de β-Lactamasas de Espectro Extendido Aisladas de Vegetales Crudos., Benemerita Universidad Autonoma de Puebla, 2019.
- Sambrook, J. [Molecular Cloning A Laboratory Manual; 2012; Vol. 33; ISBN 9781936113415.
- Eftekhar, F.; Seyedpour, S.M. Prevalence of Qnr and Aac(6’)-Ib-Cr Genes in Clinical Isolates of Klebsiella Pneumoniae from Imam Hussein Hospital in Tehran. Iran J Med Sci 2015, 40, 515–521. [Google Scholar]
- Garza-González, E.; Bocanegra-Ibarias, P.; Bobadilla-del-Valle, M.; Ponce-de-León-Garduño, L.A.; Esteban-Kenel, V.; Silva-Sánchez, J.; Garza-Ramos, U.; Barrios-Camacho, H.; López-Jácome, L.E.; Colin-Castro, C.A.; et al. Drug Resistance Phenotypes and Genotypes in Mexico in Representative Gram-Negative Species: Results from the Infivar Network. PLoS One 2021, 16, e0248614. [Google Scholar] [CrossRef]
- Garza-Ramos, U.; Davila, G.; Gonzalez, V.; Alpuche-Aranda, C.; López-Collada, V.R.; Alcantar-Curiel, D.; Newton, O.; Silva-Sanchez, J. The BlaSHV-5 Gene Is Encoded in a Compound Transposon Duplicated in Tandem in Enterobacter cloacae. Clinical Microbiology and Infection 2009, 15, 878–880. [Google Scholar] [CrossRef] [PubMed]
- Robicsek, A.; Strahilevitz, J.; Sahm, D.F.; Jacoby, G.A.; Hooper, D.C. Qnr Prevalence in Ceftazidime-Resistant Enterobacteriaceae Isolates from the United States. Antimicrob Agents Chemother 2006, 50, 2872–2874. [Google Scholar] [CrossRef] [PubMed]
- Yamane, K.; Wachino, J.; Suzuki, S.; Arakawa, Y. Plasmid-Mediated QepA Gene among E. coli Clinical Isolates from Japan. Antimicrob Agents Chemother 2008, 52, 1564–1566. [Google Scholar] [CrossRef] [PubMed]
- CLSI supplement M100. Clinical and Laboratory Standards Institute; 2023.
- Mansan-Almeida, R.; Pereira, A.L.; Giugliano, L.G. Diffusely Adherent E. colistrains Isolated from Children and Adults Constitute Two Different Populations. BMC Microbiol 2013, 13, 22. [Google Scholar] [CrossRef]
- Rodriguez-Angeles, M.G. Principales Características y Diagnóstico de Los Grupos Patógenos de E. coli. Salud Publica Mex 2002, 44, 464–475. [Google Scholar] [CrossRef]
- Aranda, K.R.S.; Fabbricotti, S.H.; Fagundes-Neto, U.; Scaletsky, I.C.A. Single Multiplex Assay to Identify Simultaneously Enteropathogenic, Enteroaggregative, Enterotoxigenic, Enteroinvasive and Shiga Toxin-Producing E. coli Strains in Brazilian Children. FEMS Microbiol Lett 2007, 267, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Franke, J.; Franke, S.; Schmidt, H.; Schwarzkopf, A.; Wieler, L.H.; Baljer, G.; Beutin, L.; Karch, H. Nucleotide Sequence Analysis of Enteropathogenic E. coli (EPEC) Adherence Factor Probe and Development of PCR for Rapid Detection of EPEC Harboring Virulence Plasmids. J Clin Microbiol 1994, 32, 2460–2463. [Google Scholar] [CrossRef] [PubMed]
- Doumith, M.; Day, M.; Ciesielczuk, H.; Hope, R.; Underwood, A.; Reynolds, R.; Wain, J.; Livermore, D.M.; Woodford, N. Rapid Identification of Major E. coli Sequence Types Causing Urinary Tract and Bloodstream Infections. J Clin Microbiol 2015, 53, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Versalovic, J.; Koeuth, T.; Lupski, R. Distribution of Repetitive DNA Sequences in Eubacteria and Application to Finerpriting of Bacterial Genomes. Nucleic Acids Res 1991. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros-Monrreal, M.G.; Arenas-Hernández, M.M.P.; Barrios-Villa, E.; Juarez, J.; Álvarez-Ainza, M.L.; Taboada, P.; De la Rosa-López, R.; Bolado-Martínez, E.; Valencia, D. Bacterial Morphotypes as Important Trait for Uropathogenic E. coli Diagnostic; a Virulence-Phenotype-Phylogeny Study. Microorganisms 2021, 9, 2381. [Google Scholar] [CrossRef]
- Wilde, C.; Mazel, D.; Hochhut, B.; Middendorf, B.; Le Roux, F.; Carniel, E.; Dobrindt, U.; Hacker, J. Delineation of the Recombination Sites Necessary for Integration of Pathogenicity Islands II and III into the E. coli 536 Chromosome. Mol Microbiol 2008, 68, 139–151. [Google Scholar] [CrossRef]
- Navidinia, M.; Najar Peerayeh, S.; Fallah, F.; Bakhshi, B.; Adabian, S.; Alimehr, S.; Gholinejad, Z. Distribution of the Pathogenicity Islands Markers (PAIs) in Uropathogenic E. Coli Isolated from Children in Mofid Children Hospital. Arch Pediatr Infect Dis 2013, 1, 75–79. [Google Scholar] [CrossRef]
- Chittò, M.; Berger, M.; Klotz, L.; Dobrindt, U. Sub-Inhibitory Concentrations of SOS-Response Inducing Antibiotics Stimulate Integrase Expression and Excision of Pathogenicity Islands in Uropathogenic E. coli Strain 536. International Journal of Medical Microbiology 2020, 310, 151361. [Google Scholar] [CrossRef]
- Firoozeh, F.; Soleimani-Moorchekhorti, L.; Zibaei, M. Evaluation of Pathogenicity Islands in Uropathogenic E. coli Isolated from Patients with Urinary Catheters. The Journal of Infection in Developing Countries 2017, 11, 557–562. [Google Scholar] [CrossRef]
- Dobrindt, U.; Blum-Oehler, G.; Nagy, G.; Schneider, G.; Johann, A.; Gottschalk, G.; Hacker, J. Genetic Structure and Distribution of Four Pathogenicity Islands (PAI I536 to PAI IV536) of Uropathogenic E. coli Strain 536. Infect Immun 2002, 70, 6365–6372. [Google Scholar] [CrossRef]
- Schneider, G.; Dobrindt, U.; Brüggemann, H.; Nagy, G.; Janke, B.; Blum-Oehler, G.; Buchrieser, C.; Gottschalk, G.; Emödy, L.; Hacker, J. The Pathogenicity Island-Associated K15 Capsule Determinant Exhibits a Novel Genetic Structure and Correlates with Virulence in Uropathogenic E. coli Strain 536. Infect Immun 2004, 72, 5993–6001. [Google Scholar] [CrossRef]
- Dahmen, S.; Métayer, V.; Gay, E.; Madec, J.-Y.; Haenni, M. Characterization of Extended-Spectrum Beta-Lactamase (ESBL)-Carrying Plasmids and Clones of Enterobacteriaceae Causing Cattle Mastitis in France. Vet Microbiol 2013, 162, 793–799. [Google Scholar] [CrossRef]
- Paniagua-Contreras, G.L.; Monroy-Pérez, E.; Bautista, A.; Reyes, R.; Vicente, A.; Vaca-Paniagua, F.; Díaz, C.E.; Martínez, S.; Domínguez, P.; García, L.R.; et al. Multiple Antibiotic Resistances and Virulence Markers of Uropathogenic E. coli from Mexico. Pathog Glob Health 2018, 112, 415–420. [Google Scholar] [CrossRef]
- Galindo-Méndez, M. Caracterización Molecular y Patrón de Susceptibilidad Antimicrobiana de E. coli Productora de β;-Lactamasas de Espectro Extendido En Infección Del Tracto Urinario Adquirida En La Comunidad. Revista chilena de infectología 2018, 35, 29–35. [Google Scholar] [CrossRef]
- Alcántar-Curiel, M.D.; Alpuche-Aranda, C.M.; Varona-Bobadilla, H.J.; Gayosso-Vázquez, C.; Jarillo-Quijada, M.D.; Frías-Mendivil, M.; Sanjuan-Padrón, L.; Santos-Preciado, J.I. Risk Factors for Extended-Spectrumb-Lactamases-Producing E. coli urinary Tract Infections in a Tertiary Hospital. Salud Publica Mex 2015, 57, 412–418. [Google Scholar] [CrossRef]
- Gallegos-Miranda, V.; Garza-Ramos, U.; Bolado-Martínez, E.; Navarro-navarro, M.; Félix-Murray, K.R.; Carmen, M.; Sanchez-Martinez, G.; Dúran-Bedolla, J.; Silva-Sanchez, J.; Garza-Ramos, U.; et al. ESBL-Producing E. coli and Klebsiella Pneumoniae from Health-Care Institutions in Mexico. Journal of Chemotherapy 2020, 0, 1–5. [Google Scholar] [CrossRef]
- Rayo Morfín-Otero, Soraya Mendoza-Olazarán, Jesús Silva-Sánchez, Eduardo Rodríguez-Noriega, Jorge Laca-Díaz, Perla Tinoco-Carrillo, Luis Petersen, Perla López, Fernando Reyna-Flores, Dolores Alcantar -Curiel, Ulises Garza-Ramos. Characterization of Enterobacteriaceae Isolates Obtained from a Tertiary Care Hospital in Mexico, Which Produce Extended-Spectrum β-Lactamase. 2013, 19, 378–383. [CrossRef]
- Reyna-Flores, F.; Barrios, H.; Garza-Ramos, U.; Sánchez-Pérez, A.; Rojas-Moreno, T.; Uribe-Salas, F.J.; Fagundo-Sierra, R.; Silva-Sanchez, J. Molecular Epidemiology of E. coli O25b-ST131 Isolates Causing Community-Acquired UTIs in Mexico. Diagn Microbiol Infect Dis 2013, 76, 396–398. [Google Scholar] [CrossRef]
- Paniagua-Contreras, G.L.; Monroy-Pérez, E.; Díaz-Velásquez, C.E.; Uribe-García, A.; Labastida, A.; Peñaloza-Figueroa, F.; Domínguez-Trejo, P.; García, L.R.; Vaca-Paniagua, F.; Vaca, S. Whole-Genome Sequence Analysis of Multidrug-Resistant Uropathogenic Strains of E. coli from Mexico. Infect Drug Resist 2019, 12, 2363–2377. [Google Scholar] [CrossRef]
- Drinkovic, D.; Morris, A.J.; Dyet, K.; Bakker, S.; Heffernan, H. Plasmid-Mediated AmpC Beta-Lactamase-Producing E. coli Causing Urinary Tract Infection in the Auckland Community Likely to Be Resistant to Commonly Prescribed Antimicrobials. N Z Med J 2015, 128, 50–59. [Google Scholar]
- Ehsan, B.; Haque, A.; Qasim, M.; Ali, A.; Sarwar, Y. High Prevalence of Extensively Drug Resistant and Extended Spectrum Beta Lactamases (ESBLs) Producing Uropathogenic E. coli Isolated from Faisalabad, Pakistan. World J Microbiol Biotechnol 2023, 39, 132. [Google Scholar] [CrossRef]
- Ramírez-Castillo, F.Y.; Moreno-Flores, A.C.; Avelar-González, F.J.; Márquez-Díaz, F.; Harel, J.; Guerrero-Barrera, A.L. An Evaluation of Multidrug-Resistant E. coli Isolates in Urinary Tract Infections from Aguascalientes, Mexico: Cross-Sectional Study. Ann Clin Microbiol Antimicrob 2018, 17, 34. [Google Scholar] [CrossRef]
- Robicsek, A.; Strahilevitz, J.; Jacoby, G.A.; Macielag, M.; Abbanat, D.; Hye Park, C.; Bush, K.; Hooper, D.C. Fluoroquinolone-Modifying Enzyme: A New Adaptation of a Common Aminoglycoside Acetyltransferase. Nat Med 2006, 12, 83–88. [Google Scholar] [CrossRef]
- Frasson, I.; Cavallaro, A.; Bergo, C.; Richter, S.N.; Palù, G. Prevalence of Aac(6’)-Ib-Cr Plasmid-Mediated and Chromosome-Encoded Fluoroquinolone Resistance in Enterobacteriaceae in Italy. Gut Pathog 2011, 3, 12. [Google Scholar] [CrossRef]
- Raherison, S.; Jove, T.; Gaschet, M.; Pinault, E.; Tabesse, A.; Torres, C.; Ploy, M.-C. Expression of the Aac(6′)-Ib-Cr Gene in Class 1 Integrons. Antimicrob Agents Chemother 2017, 61. [Google Scholar] [CrossRef]
- Seni, J.; Falgenhauer, L.; Simeo, N.; Mirambo, M.M.; Imirzalioglu, C.; Matee, M.; Rweyemamu, M.; Chakraborty, T.; Mshana, S.E. Multiple ESBL-Producing E. coli Sequence Types Carrying Quinolone and Aminoglycoside Resistance Genes Circulating in Companion and Domestic Farm Animals in Mwanza, Tanzania, Harbor Commonly Occurring Plasmids. Front Microbiol 2016, 7. [Google Scholar] [CrossRef]
- Juraschek, K.; Malekzadah, J.; Malorny, B.; Käsbohrer, A.; Schwarz, S.; Meemken, D.; Hammerl, J.A. Characterization of QnrB-Carrying Plasmids from ESBL- and Non-ESBL-Producing E. coli. BMC Genomics 2022, 23, 365. [Google Scholar] [CrossRef]
- Riley, L.W. Pandemic Lineages of Extraintestinal Pathogenic E. coli. Clinical Microbiology and Infection 2014, 20, 380–390. [Google Scholar] [CrossRef]
- Moreira da Silva, R.C.R.; de Oliveira Martins Júnior, P.; Gonçalves, L.F.; de Paulo Martins, V.; de Melo, A.B.F.; Pitondo-Silva, A.; de Campos, T.A. Ciprofloxacin Resistance in Uropathogenic E. coli Isolates Causing Community-Acquired Urinary Infections in Brasília, Brazil. J Glob Antimicrob Resist 2017, 9, 61–67. [Google Scholar] [CrossRef]
- Demirci, M.; Ünlü, Ö.; İstanbullu Tosun, A. Detection of O25b-ST131 Clone, CTX-M-1 and CTX-M-15 Genes via Real-Time PCR in E. coli Strains in Patients with UTIs Obtained from a University Hospital in Istanbul. J Infect Public Health 2019, 12, 640–644. [Google Scholar] [CrossRef]
- Hussain, A.; Ewers, C.; Nandanwar, N.; Guenther, S.; Jadhav, S.; Wieler, L.H.; Ahmed, N. Multiresistant Uropathogenic E. coli from a Region in India Where Urinary Tract Infections Are Endemic: Genotypic and Phenotypic Characteristics of Sequence Type 131 Isolates of the CTX-M-15 Extended-Spectrum-β-Lactamase-Producing Lineage. Antimicrob Agents Chemother 2012, 56, 6358–6365. [Google Scholar] [CrossRef] [PubMed]
- Valiatti, T.B.; Santos, F.F.; Santos, A.C.M.; Nascimento, J.A.S.; Silva, R.M.; Carvalho, E.; Sinigaglia, R.; Gomes, T.A.T. Genetic and Virulence Characteristics of a Hybrid Atypical Enteropathogenic and Uropathogenic E. coli (AEPEC/UPEC) Strain. Front Cell Infect Microbiol 2020, 10. [Google Scholar] [CrossRef]
- Tanabe, R.H.S.; Dias, R.C.B.; Orsi, H.; de Lira, D.R.P.; Vieira, M.A.; dos Santos, L.F.; Ferreira, A.M.; Rall, V.L.M.; Mondelli, A.L.; Gomes, T.A.T.; et al. Characterization of Uropathogenic E. coli Reveals Hybrid Isolates of Uropathogenic and Diarrheagenic (UPEC/DEC) E. coli. Microorganisms 2022, 10, 645. [Google Scholar] [CrossRef]
- Pérez Ama, A. Estudio de Las Propiedades de Virulencia En Cepas de E. coli Uropatógena; Benemérita Universidad Autónoma de Puebla: Puebla, 2018. [Google Scholar]
- Santos, A.C. de M.; Santos, F.F.; Silva, R.M.; Gomes, T.A.T. Diversity of Hybrid- and Hetero-Pathogenic E. coli and Their Potential Implication in More Severe Diseases. Front Cell Infect Microbiol 2020, 10. [Google Scholar] [CrossRef]
Figure 1.
Prevalence of Pathogenicity Islands (PAI) in this Study.
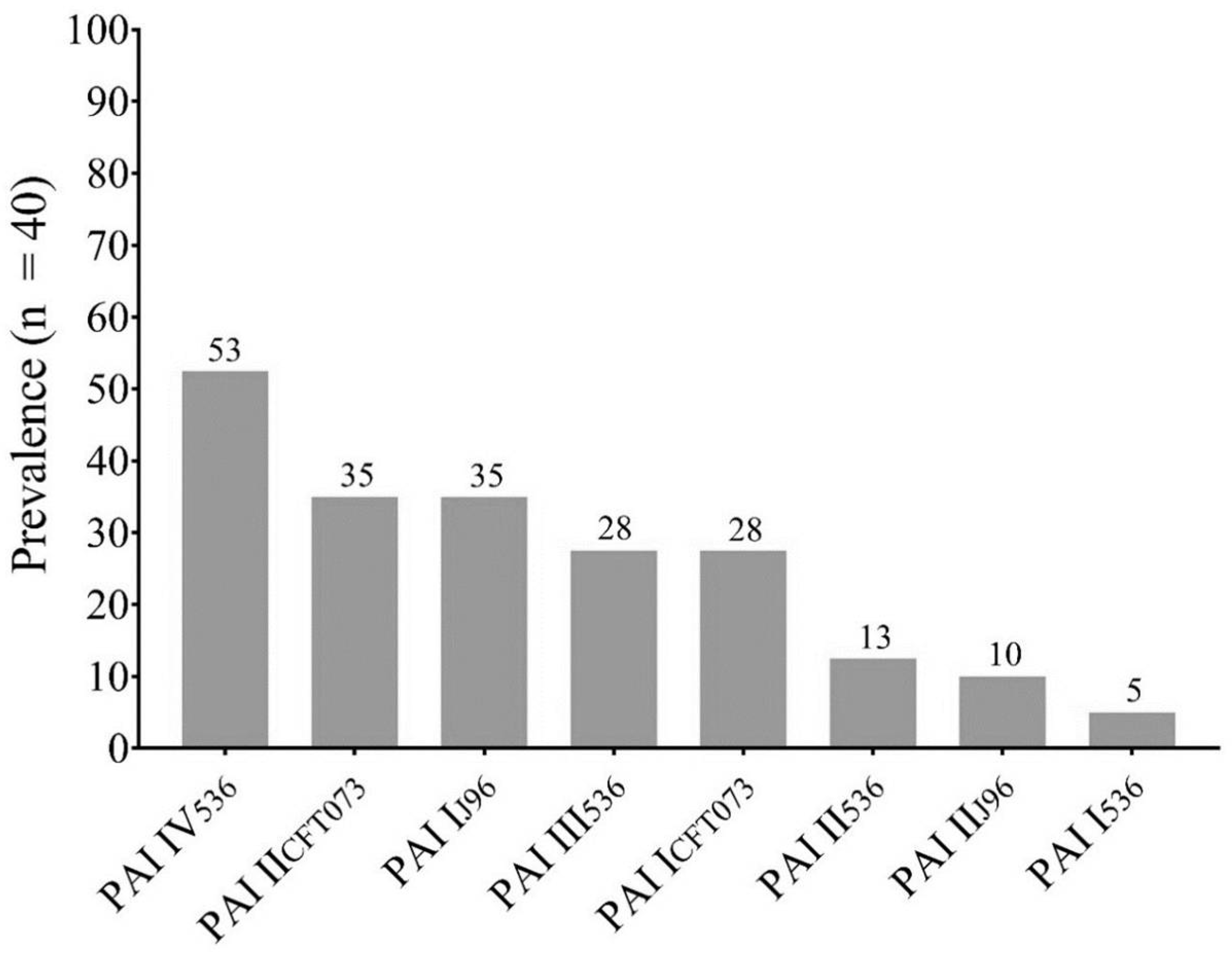
Figure 2.
Adherence patterns detected in the analyzed clinical isolates. A) Tandem bricks pattern; B) Aggregative pattern; C) Diffuse pattern; D) Localized pattern. Bright field microscopy 50X objective.
Figure 2.
Adherence patterns detected in the analyzed clinical isolates. A) Tandem bricks pattern; B) Aggregative pattern; C) Diffuse pattern; D) Localized pattern. Bright field microscopy 50X objective.
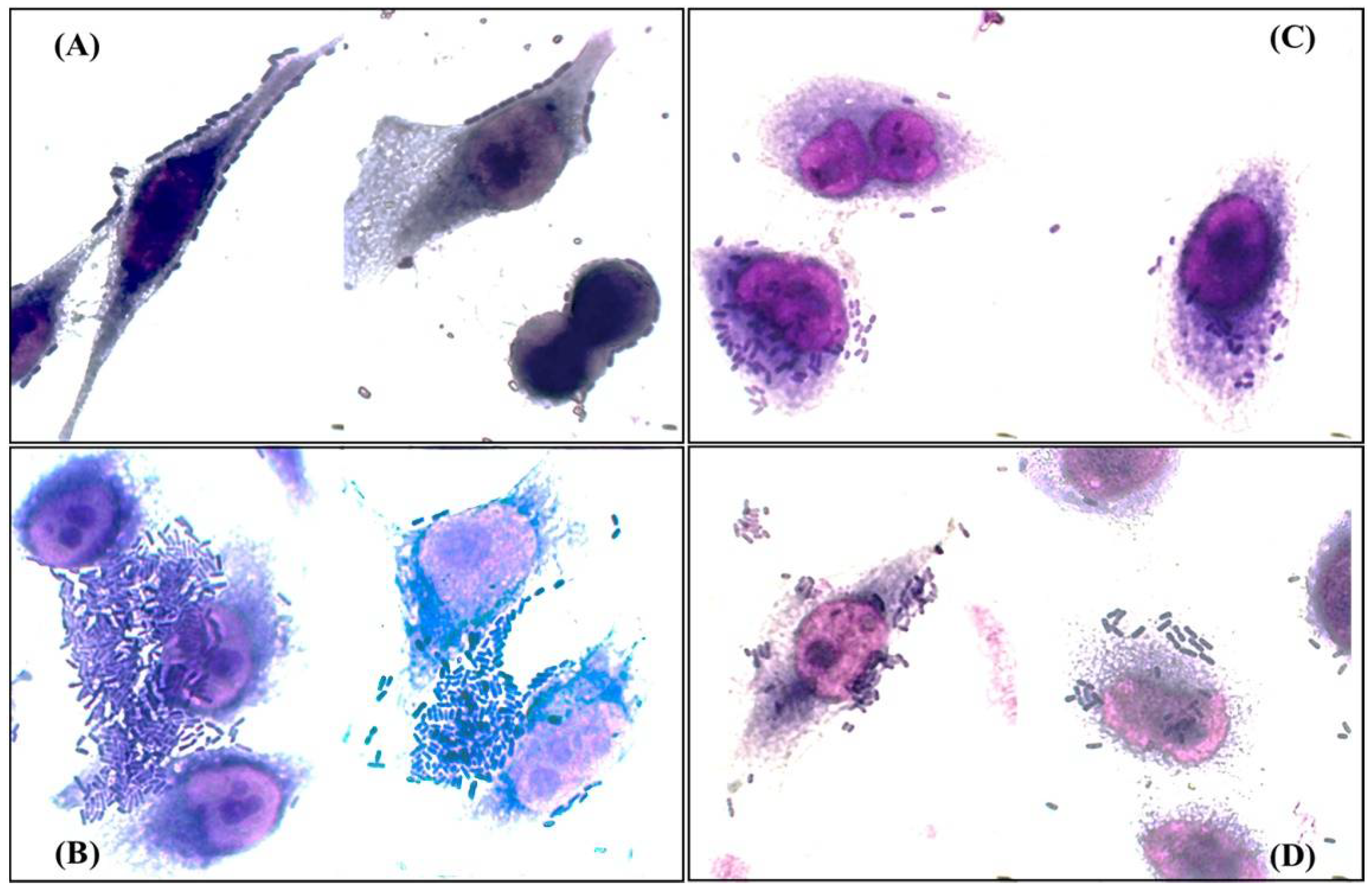
Figure 3.
Dendrogram derived from the UPGMA pairing and the DICE coefficient representing ERIC-PCR patterns, PAI profile, DEC pathotypes associated genes, STs, and resistance characteristics of clinical E. coli isolates analyzed. PAI: Pathogenicity island; DAEC: Diffuse adherent E. coli (presence of daaE gene); EPEC: Enteropathogenic E. coli (presence of bfpA and eaeA genes); EAEC: Enteroaggregative E. coli (presence of pCVD432 gene); ST: Sequence types; Amg: Aminoglycosides; FQ: Fluoroquinolones; Cx: Cefoxitin; 3rd CFS: Third generation cephalosporins; FEP: Cefepime; ATM: Aztreonam; Pen+Inh: Penicillins with ESBL inhibitor; CAR: Carbapenemase.
Figure 3.
Dendrogram derived from the UPGMA pairing and the DICE coefficient representing ERIC-PCR patterns, PAI profile, DEC pathotypes associated genes, STs, and resistance characteristics of clinical E. coli isolates analyzed. PAI: Pathogenicity island; DAEC: Diffuse adherent E. coli (presence of daaE gene); EPEC: Enteropathogenic E. coli (presence of bfpA and eaeA genes); EAEC: Enteroaggregative E. coli (presence of pCVD432 gene); ST: Sequence types; Amg: Aminoglycosides; FQ: Fluoroquinolones; Cx: Cefoxitin; 3rd CFS: Third generation cephalosporins; FEP: Cefepime; ATM: Aztreonam; Pen+Inh: Penicillins with ESBL inhibitor; CAR: Carbapenemase.
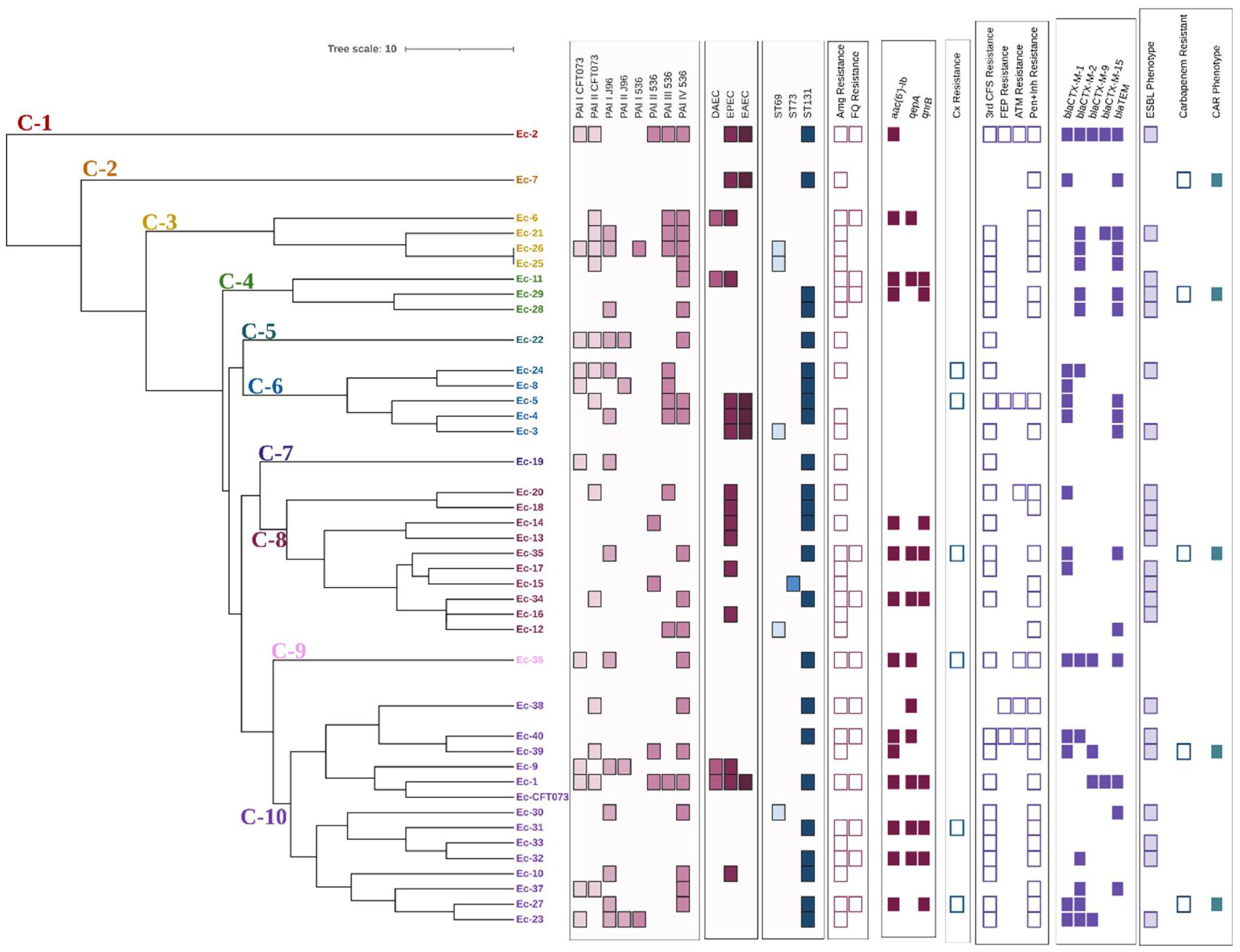

Table 1.
Pathogenicity Islands (PAI) Profile in each Clinical Isolate of E. coli from ITU.
| PAI Profile | Isolate ID |
|---|---|
| PAI I 536, PAI I CFT073 | 25 |
| PAI I 536, PAI I J96, PAI II J96, PAI I CFT073 | 23 |
| PAI I 536, PAI III 536, PAIV 536, PAI J96, PAI I CFT073, PAI II CF073 | 26 |
| PAI I J96, PAI I CFT073 | 19 |
| PAI I J96, PAII J96, PAI I CFT073 | 9 |
| PAI II 536 | 14,15 |
| PAI II 536, PAI III 536, PAI IV 536, PAI I CFT073, PAI II CFT073 | 1, 2 |
| PAI II 536, PAI IV 536, PAI II CFT073 | 39 |
| PAI III 536, PAI II CFT073 | 20 |
| PAI III 536, PAI II J96, PAI I CFT073 | 8 |
| PAI III 536, PAI IV 536 | 12 |
| PAI III 536, PAI IV 536, PAI I J96, PAI II CFT073 | 21 |
| PAI III 536, PAI IV 536, PAI II CFT073 | 5,6 |
| PAI III 536, PAI IV 536, PAI J96 | 4 |
| PAI III 536, PAI J96, PAI I CFT073, PAI II CFT073 | 24 |
| PAI IV 536, PAI II CFT073 | 34 |
| PAI IV 536 | 11 |
| PAI IV 536, PAI I CFT073, PAI II CFT073 | 37 |
| PAI IV 536, PAI I J96 | 10, 27, 28, 30, 35 |
| PAI IV 536, PAI I J96, PAI I CFT073 | 36 |
| PAI IV 536, PAI I J96, PAI II J96, PAI I CFT073, PAI II CFT073 | 22 |
| PAI IV 536, PAI II CFT073 | 25, 38 |
| Negatives | 3, 4,13,16-18,29,31-33,40 |
PAI: Pathogenicity islands.
Table 2.
ESBL Phenotype and ESBL Profiles respect to β-lactam Resistance Phenotype.
| ID | Antibiotic Resistance* | ESBL Phenotype | ESBL Profile | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CX | AMC | AMS | CFZ | CTX | CRO | FEP | ATM | CTX | CFZ | CRO | FEP | ATM | ||
| 1 | R | R | R | |||||||||||
| 5 | R | R | R | R | R | R | R | R | ||||||
| 10 | R | R | ||||||||||||
| 12 | R | R | R | |||||||||||
| 19 | R | R | ||||||||||||
| 22 | R | R | ||||||||||||
| 25 | R | R | R | |||||||||||
| 26 | R | R | R | |||||||||||
| 27 | R | R | R | R | R | R | ||||||||
| 31 | R | R | R | R | R | |||||||||
| 35 | R | R | R | R | R | |||||||||
| 36 | R | R | R | R | R | R | ||||||||
| 21 | R | R | R | R | R | + | + | CFZ, CRO | ||||||
| 3 | R | R | R | R | + | CRO | ||||||||
| 11 | R | R | R | + | CTX | |||||||||
| 13 | R | R | + | CTX | ||||||||||
| 14 | R | R | + | CTX | ||||||||||
| 15 | R | R | + | CTX | ||||||||||
| 16 | R | R | + | CTX | ||||||||||
| 17 | R | + | CTX | |||||||||||
| 18 | R | R | R | + | CTX | |||||||||
| 23 | R | R | R | + | CTX | |||||||||
| 20 | R | R | R | R | + | + | CTX, ATM | |||||||
| 40 | R | R | R | R | R | R | R | + | + | CTX, ATM | ||||
| 24 | R | R | R | R | R | + | + | CTX, CFZ | ||||||
| 30 | R | R | R | R | + | + | CTX, CFZ | |||||||
| 28 | R | R | R | R | R | + | + | + | CTX, CFZ, CRO | |||||
| 32 | R | R | R | R | R | + | + | + | CTX, CFZ, CRO | |||||
| 34 | R | R | R | R | R | + | + | + | CTX, CFZ, CRO | |||||
| 2 | R | R | R | R | R | R | R | + | + | + | + | + | CTX, CFZ, CRO,FEP, ATM | |
| 29 | R | R | R | R | R | + | + | + | + | + | CTX, CFZ, CRO,FEP, ATM | |||
| 39 | R | R | R | R | + | + | CTX, CRO | |||||||
| 33 | R | R | R | R | R | + | + | + | CTX, CRO, ATM | |||||
| 38 | R | R | R | R | + | + | FEP, ATM | |||||||
Predetermined resistance profile [10]. R: Resistant; Blank space: Negative ESBL Phenotype; + Positive ESBL Phenotype; CX: Cephamycin; AMC: Amoxicillin-Clavulanic Acid; AMS: Ampicillin-Sulbactam; CTX: Cefotaxime; CFZ: Ceftazidime; CRO: Ceftriaxone; FEP: Cefepime; ATM: Aztreonam. In underlined, CX and AMC or AMS resistant isolates without ESBL phenotype are shown. Only results for β-lactam Resistant isolates is shown.
Table 3.
Prevalence of Virulence and Resistance Determinants by Clonal Group.
| Feature | ST73 %(n = 1) |
ST69 %(n = 5) |
STNT %(n = 10) |
ST131 %(n= 24) |
p value |
|---|---|---|---|---|---|
| PAI III536 | 0 | 40 (2) | 20 (2) | 29.2 (7) | |
| PAI IV536 | 0 | 80 (4) | 50 (5) | 50 (12) | |
| PAI IICFT073 | 0 | 40 (2) | 40 (4) | 33.3 (8) | |
| PAI II536 | 100 (1) | 0 | 10 (1) | 13 (3) | |
| PAI IJ96 | 0 | 40 (2) | 20 (2) | 42 (10) | |
| PAI ICFT073 | 0 | 20 (1) | 20 (2) | 33.3 (8) | |
| PAI I536 | 0 | 20 (1) | 0 | 4.2 (1) | |
| PAI IIJ96 | 0 | 0 | 10 (1) | 13 (3) | |
| Aminoglycosides | 100 (1) | 80 (4) | 70 (7) | 83.3 (20) | |
| Fluoroquinolones | 0 | 0 | 30 (3) | 50 (12) * | 0.05 |
| Cotrimoxazole | 100 (1) | 60 (3) | 40 (4) | 46 (11) | |
| Cephamycins | 0 | 0 | 0 | 25 (6) | |
| 3st Cephalosporins | 100 (1) | 100 (5) | 70 (7) | 83.3 (20) | |
| 4th Cephalosporins | 0 | 0 | 0 | 17 (4) | |
| Monobactams | 0 | 0 | 20 (2) | 33.3 (8) | |
| Pen-Inh | 100 (1) | 100 (5) | 70 (7) | 79.2 (19) | |
| Carbapenems | 0 | 0 | 10 (1) | 17 (4) | |
| ESBL Phenotype | 100 (1) | 40 (2) | 70 (7) | 50 (12) | |
| CAR Phenotype | 0 | 0 | 10 (1) | 17 (4) | |
| blaCTX-M 1 y 8 | 0 | 0 | 40 (4) | 58.3 (14)* | 0.005 |
| blaCTX-M-9 | 0 | 0 | 10 (1) | 17 (4)* | 0.02 |
| blaTEM | 0 | 100 (5)* | 20 (2) | 38 (9) | |
| blaCTX-M-2 | 100 (1) | 60 (3) | 30 (3) | 46 (11) | |
| blaCTX-M-15 | 0 | 0 | 10 (1) | 8.3 (2)* | 0.05 |
| qepA | 0 | 0 | 20 (2) | 33.3 (8)* | 0.001 |
| aac(6')-Ib | 0 | 0 | 30 (3) | 46 (11)* | 0.002 |
| qnrB | 0 | 0 | 10 (1) | 33.3 (8)* | <0.001 |
PAI: Pathogenicity island; 3srdCephalosporins: Third generation cephalosporins; 4th Cephalosporins: Fourth generation cephalosporins; Pen-Inh: Penicillin’s combined with β-lactamases Inhibitors; ESBL: β-lactamase positive phenotype; CAR: Carbapenemase positive phenotype; bla: β-lactamase associated genes; qepA: efflux pump associated with fluoroquinolone resistance; aac(6’)-Ib: Acetylase associated gene; qnrB: Protective proteins associated with fluoroquinolone resistance; NT: Non typable- The p values were obtained by Fisher Exact Test.
Table 4.
Adherence Patterns, Pathotypes Associated Genes, PAI Profile and Potential Hybrid Pathotypes of Clinical Isolates.
Table 4.
Adherence Patterns, Pathotypes Associated Genes, PAI Profile and Potential Hybrid Pathotypes of Clinical Isolates.
| ID | Pattern | Profile | PAI Profile | Pathotype |
|---|---|---|---|---|
| 1 | Bs/Lo | pCVD432, eaeA, daaE, bfpA |
PAI III536, PAI IV536, PAI IICFT073, PAI II536, PAI ICFT073 |
UPEC/EPEC/DAEC/EAEC |
| 2 | Bs/Dif | pCVD432, bfpA | PAI III536, PAI IV536, PAI IICFT073, PAI II536, PAI ICFT073 |
UPEC/EAEC/EPEC |
| 3 | Bs/Lo/Ag | pCVD432, bfpA | - | UPEC/EAEC/EPEC |
| 4 | Bs/Lo | pCVD432, bfpA | PAI III536, PAI IV536, PAIIJ96 | UPEC/EAEC/EPEC |
| 5 | Ag | pCVD432, bfpA | PAI III536, PAI IV536, PAI IICFT073 | UPEC/EAEC/EPEC |
| 6 | Bs/Lo/Di | daaE, bfpA | PAI III536, PAI IV536, PAI IICFT073 | UPEC/DAEC/EPEC |
| 7 | Bs | pCVD432, bfpA | - | UPEC/EAEC/EPEC |
| 8 | Bs/Lo | - | PAI III536, PAI ICFT073, PAI IIJ96 | UPEC |
| 9 | Bs/Ag | daaE, bfpA | PAI IJ96, PAI ICFT073, PAI IIJ96 | UPEC/DAEC/EPEC |
| 10 | Bs/Lo | bfpA | PAI IV536, PAIIJ96 | UPEC/EPEC |
| 11 | Bs | daaE, bfpA | PAI IV536 | UPEC/DAEC/EPEC |
| 12 | Lo | - | PAI III536, PAI IV536 | UPEC |
| 13 | Ag/Lo | bfpA | - | UPEC/EPEC |
| 14 | Bs | bfpA | PAI II536 | UPEC/EPEC |
| 15 | Bs | - | PAI II536 | UPEC |
| 16 | Ag/Bs/Lo | bfpA | - | UPEC/EPEC |
| 17 | Bs | bfpA | - | UPEC/EPEC |
| 18 | Bs | bfpA | - | UPEC/EPEC |
| 19 | Bs/Ag/Lo | - | PAI IJ96, PAI ICFT073 | UPEC |
| 20 | Bs/Lo/Di | bfpA | PAI III536, PAI IICFT073 | UPEC/EPEC |
ID: Isolate; PAI: Pathogenicity islands; Bs: Tandem bricks pattern; Lo: Localized pattern; Di: Diffuse pattern; Ag: Aggregative pattern.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



