Submitted:
07 May 2024
Posted:
07 May 2024
You are already at the latest version
Abstract
At the end of 2023 the Whole Mouse Brain Atlas was announced, revealing that there are about 5,300 molecularly defined neuronal types in the mouse brain. We ask whether brain models exist that contemplate how this is possible. The conventional columnar model, implicitly used by the authors of the Atlas, is incapable of doing so with only 20 brain columns (5 brain vesicles with 4 columns each). We argue that definition of some 1,250 distinct progenitor microzones, each producing over time minimally 4-5 neuronal types, may be sufficient. This is nearly achieved presently by the prosomeric model amplified by secondary dorsoventral and anteroposterior microzonation of progenitor areas, plus clonal variation of cell types produced on average by each of them.
Keywords:
neuronal cell types
; prosomeric brain model
; longitudinal zones
; neuromeres
; progenitor microzonal regionalization
; clonal properties
; tangential migration
; causal explanation
Introduction
The BRAIN Initiative Cell Census Network (BICCN) released the Whole Mouse Brain Atlas publication package in Nature on December 13, 2023 (https://www.nature.com/collections/fgihbeccbd). This single-cell transcriptomics, epigenomic, and spatial transcriptomic composite effort updates the number of different neuronal cell types present in the mouse brain to a staggering 5,300 total, revealing their molecular diversity in concert with their relative positions.
The issues we raise here are: ¿Can we explain how different cell types are produced and positioned? This question relates to ¿Do we have morphologic models allowing visualization of this level of variety in terms of relative position and neuronal typologic specification? The answers are, surprisingly: Perhaps; and: Nearly yes.
The BICCN publications implicitly use the conventional columnar brain model of Herrick (1910; Figure 1a-d), possibly in the modified version of Swanson (2004, 2012; Figure 1e), or that used by Dong (2008) in the Allen Institute’s Adult Mouse Brain Atlas [mouse.brain-map.org]. This model contemplates the telencephalon, diencephalon, midbrain, hindbrain and spinal cord as main partitions (5 rostrocaudal vesicles; Figure 1a). In this model, Herrick’s smallest units are represented by 4 functional entities (somatomotor, visceromotor, viscerosensory, and somatosensory columns defined in the brainstem and spinal cord; Sm, Vm, Vs, Ss; Figure 1a,d). This author extrapolated them into the forebrain (i.e., diencephalic Eth, Dth, Vth, Hth; telencephalic Hi, Pir, Str, Se; uniform-colored codes in Figure 1a-c). Note the forebrain columns possibly harbor other functions, though the diencephalon has been studied functionally as a continuation of the brainstem. This makes on the whole 5 vesicles x 4 columns = 20 columnar units that should produce the recently discovered 5,300 neuronal types (average of 265 cell types per column).
The columnar model never postulated any smaller generative parts than the cited columns, and it thus cannot explain today how specific neuronal types are generated by each column (in fact, columns were assumed to have a homogeneous cellular structure throughout; Kuhlenbeck 1973). Relatively recently, the brain was still assumed to contain only a few hundred neuronal types, perhaps close to 1,000 (Masland 2004). Nevertheless, neuroanatomical studies had identified many discrete neuronal populations within the columns, amply mapped in articles, books and atlases as nuclear, cortical or reticular cytoarchitectonic formations. These entities have remained devoid of causal explanation within the columnar model, representing what can be caricaturized as a ‘potato-sack’ view of order-less columnar ‘substructure’ (e.g., Krieg’s 1932 tridimensional image of the rat hypothalamic column; Figure 2; consider also modern notions about thalamic nuclei, another column). This sort of ‘unexplained hardcore neuroanatomic knowledge’ (see Figure 2) is so old now (over 100 years) that many think it is normal that we ignore why such columnar substructure exists.
We learn now that a ‘micropotato’ substructure of thousands of neuronal types exists, likewise without explanations of why each of them emerged and is where it is. They just are mapped by a computer as credible sacks of microscopic neuronal ‘potatoes’ with reproducible boundaries. These ‘micropotatoes’ are more real than the old bigger ones, since they are underpinned by more potent molecular technology, but seem more difficult to behold than the classic nuclei (we need computers to represent them). It recalls a similar reductive change that occurred when rocks, minerals and chemicals were reduced to atoms and subatomic particles. That scientific step was no doubt positive in the long run, and the present one may be so likewise for neuroscience. A neuronal theory of the brain is with us now for more than 100 years, pioneered by descriptive efforts by Ramón y Cajal and contemporary colleagues with the Golgi technique (has anyone counted how many neurons they described?). Unfortunately, this premature theory also lacked causal content and did not convey there would be thousands of neuronal types. We now see this matter apparently depends on developmental combinatorial usage of the genome during neuronal type specification. We thus should take seriously the issue of understanding causally (reductively) neuronal and glial cell type multiplicity in the brain, quite apart from the obvious consequences regarding substantial enrichment of our functional schemata.
During the last century there occurred also an important increase in our knowledge of brain connections and functions. Unfortunately, brain connections are likewise largely unexplained. They are so far largely unpredictable, excepting perhaps those of motoneurons. Helped by experimental hodological studies, lesion studies, and clinical cases, we have learned to schematize some of the basic functional wiring of the brain, a much-valued resource in clinical work (e.g., Figure 3). However, the apparent function of a brain pathway does not tell how it came to exist ontogenetically or evolutionarily, and may obscure less salient parallel functions (see Puelles 2024). Hodological schemata refer only to a few cell types. We cannot visualize 5,300 brain cell types in our functional diagrams, so that much analysis remains to be done in this context. This leads again, we pose, to knowledge without understanding, not a desirable position in science.
A significant conclusion to draw from the drawn morpho-physiological scenario is that the old-fashioned columnar brain model is irreversibly obsolete. This is due to its excessively simplistic concept of brain generative units (the postulated columns are conceived as homogeneous masses of postmitotic neurons, not even mentioning progenitors). Moreover, this model remains unable to formulate a molecular developmental causal basis for nuclear or cortical columnar substructure after 40 years of molecular neurodevelopmental advances. If we ever want to understand the why and where of 5,300 cell types in the brain, we need much more discriminative molecular and causal evo-devo-friendly brain models.
Alternative Models
Alternative possibilities already exist. They were recently sketched in “Towards a New Neuromorphology” (Nieuwenhuys and Puelles 2016), and are flowering likewise in much other recent literature. Developmental brain models have been growing in a less simplistic direction, potentiated by a variety of molecular procedures. We now can subdivide the brain more finely than in the classic 5 vesicles, or classify brain progenitors much more discriminatively than in the classical columns. These alternative approaches are already on the verge of being precise and complete enough (in their coverage of all brain territories) to account for thousands of cell types and specification of corresponding fixed relative positions. The possibility to understand the brain’s exquisite cellular structure is thus steadily increasing, a scenario that can only benefit functional and pathological analyses. Parallel technical advances in modern molecular hodology, physiology and neuropharmacology increasingly allow molecular properties of the neuronal cell membrane and related operative molecules to be translated into biological functions. This is why we answer the first question above as ‘perhaps’, and the second question as ‘nearly so’, due to the present partial incompleteness of such clarifying studies.
Longitudinal Zones of His
The alternative research program that is allowing us to surpass the columnar stalemate deals with mechanisms of developmental brain regionalization. This trend possibly started with the work of His in the last quarter of the 19th century (His 1893). This pioneering embryologist discovered longitudinal neurogenetic differences in the embryonic brain wall that led to differentiation of distinct floor, basal, alar and roof longitudinal zones (Figures 1f,g and 4a-f). This came jointly with the subsequently corroborated notion of a longitudinal general alar-basal boundary separating alar sensory neuronal classes from basal motor ones. These concepts were inherited by the neuromeric models mentioned next and were validated by accruing molecular results (Figures 1f,g and 4a-f). In the molecular era (eighties onwards) we learned that the zones of His result from dorsoventral patterning (DV) triggered by means of sonic hedgehog protein (SHH) diffusing out of the subjacent notochord into the brain primordium. The notochord ends rostrally under the mamillary body (Nch; M; Figure 1f,g). The four longitudinal zones of His run through the whole length of the neural primordium, coinciding at forebrain levels with a Nkx2.2-positive alar-basal boundary band (Figure 4b; this approximates the classic limiting sulcus of His, but the band can be seen already at neural plate stages; Sánchez-Arrones et al. 2009). These zones end rostrally at the singular acroterminal hypothalamic region, marked selectively by Dlk1 expression (Figure 4f,g; green-shaded in Figure 1g; this notion was introduced by Puelles et al. 2012). This rostral hypothalamic locus is contacted by the endodermal prechordal plate at the onset of gastrulation and forebrain neural induction, an interaction with a role in AP patterning (Acampora et al. 2001; García.Calero et al. 2008).
His’s epichordal dorsoventral zones (Figure 4a-g) reflect DV patterning as a causal antecedent of differently specified alar and basal progenitors leading secondarily to Herrick’s (1910) brainstem and spinal neuronal columns (Figure 1a); the latter result from secondary microzonal subdivisions of the basal and alar plates -see Figure 5e,f -, plus some motoneuronal tangential migrations (Ju et al. 2004 and Puelles et al. 2019). In contrast, the molecularly validated zones of His contradict Herrick’s and Swanson’s columnar hypotheses for the forebrain, due to their arbitrary forebrain axis ending in the telencephalon, a course objectively not followed by the observable molecularly-defined longitudinal zones (Figure 4a-f).
The Synthetic Neuromeric Models, Leading to the Prosomeric Model
A parallel second step forward resulted from 19th and 20th century descriptions of neuromeres, that is, series of transverse vesicular bulges of the embryonic neural tube wall (Figures 4b and 5a,b; Orr 1887, McClure 1890; Locy 1895, von Kupffer 1906; Bergquist and Källén 1954; Vaage 1969). We learned subsequently that neuromeres are specified differentially with partially unique molecular profiles leading to distinct adult fates. They also share some genetic determinants causing metamery, that is, serial repetition of given features among neighboring units. Some neuromeres clearly substitute for badly described columns in the forebrain (Figures 1f,g and 4b), and all of them establish an AP subdivision of His’s longitudinal zones (or Herrick’s columns) in the brainstem and spinal cord, while retaining the capacity to form modular plurineuromeric neuronal complexes corresponding to classic columns. This pattern allows in principle qualitatively different events happening in each neuromere -e.g., specification of different cell types-, without impeding the functional sensorial or motor unity stressed in the columnar system. The neuromeric pattern thus provides for significant modular cellular and functional diversity and, accordingly, reduces partially the ‘potato-sack’ problem (Figures 1f,g; 4b,c; 5a-c). Moreover, neuromeres can be explained as a result from AP patterning of the entire neural tube (review in Puelles 2018). Large tagmata (forebrain, hindbrain, and spinal cord), intermediate proneuromeres (hypothalamus, diencephalon, midbrain, etc.) and final individual neuromeres may be contemplated as molecularly diverse AP constituents (Figure 1g); note tagmata, proneuromeres and neuromeres can be still distinguished in the adult (Figure 5a-c).
It was long thought that neuromeres are transient early phenomena lacking both discrete derivatives in the adult brain and specific functions, but transgenic technology in the molecular era has shown that their molecular boundaries and derivatives persist even in adults (Figure 5c,d), though they become otherwise cryptic (invisible by inspection) as the brain wall thickens. Von Kupffer (1906), Bergquist and Källén (1954; Figure 1f) and Vaage (1969) listed a number of constant overt (bulging) neuromeres among vertebrates. Later some non-bulging or cryptic neuromeres showing only a molecular delimitation were added within the ‘prosomeric model’ (Figure 1g). This was the first molecularly-based neuromeric model, studied in the mouse (Figures 1g and 5b; Puelles and Rubenstein 1993, 2003; Rubenstein et al. 1994; Puelles and Rubenstein 2015; Tomás-Roca et al. 2016; Puelles 2013, 2018). Additional neuromere fate-mapping was done in the chick, as well as consistent differential gene mappings on chick and mouse embryos, leading to the conclusion that the four dorsoventral zones of His are subdivided into roughly 50 neuromeric segments (7 in the forebrain down to the isthmus, 12-13 in the hindbrain, and over 30 spinal cord units). This already represents a synthesis of 4 His zones x 50 neuromeres =200 different nominal generative units, theoretically responsible only for an average of 26,5 cell types each. These are surely still too many cell types to be explained as coming from a single generative unit. Neuromeric models divided into His primary DV zones thus strictly are also insufficient to solve the present ‘micro-potato-sack’ problem, but are no doubt much closer than the columnar models by an order of magnitude (average of 26,5 versus 265 cell types per column).
Dorsoventral Microzones
Further explanatory possibilities emerged as experimental molecular studies reviewed in Puelles (2013) and Nieuwenhuys (2017) indicated that each neuromere does not subdivide dorsoventrally merely into the 4 primary His zones (though these are real; Figure 4), but into 13-14 molecularly singular DV microzones per neuromere, each boundary resulting from antagonism between different pairs of ventral and dorsal genes (Figure 5e, f). Each microzone is capable of producing one or more different neuronal cell types (Figure 5g). We have: 1-2 floor plate microzones, 5 basal microzones, 6-7 alar microzones, 1-2 roof plate microzones. This pattern was first demonstrated in spinal and hindbrain neuromeric units (Figures 5e, f; 6a; Puelles 2013), but appears as well in the two hypothalamic neuromeres, in a particularly expanded manner if the numerous telencephalic subpallial and pallial subdomains are regarded as evaginated alar hypothalamic derivatives (Figures 5b; 6b,c; Flames et al. 2007; Puelles et al. 2012a, Diaz and Puelles 2022). Analogous alar DV microzonal patterning data were reported for the midbrain, pretectum and prethalamus (Puelles et al. 2012a,b; 2021; see Figure 6a). If such DV microzonal generative substructure would be found to apply to all or most neuromeres (unclear yet, but certainly possible), this would allow 13 microzones x 50 neuromeres = 650 distinct generative units instead of the 200 ones counted above only with 4 His zones. In this case a theoretical average of 8,15 cell types per DV microzone is reached (we are now already near the needed level of regionalization).
Anteroposterior Microzones
We will now look in a different direction. At least three forebrain neuromeres have been separately shown to display in their alar plates additional AP microzonal molecular regionalization, involving an AP tripartition (cases of the pretectum, Ferrán et al. 2008, 2009 -Figure 6d-f-, the prethalamus, see Puelles et al. 2021, and the midbrain m1 mesomere, see Puelles et al. 2012b; Puelles and Hidalgo-Sánchez 2023). Additionally, the thalamus shows at least two AP alar partitions, and a third one is possible (Puelles and Martínez 2013; see legend for Figure 6f). Note the alar cerebellum in r1 divides into hemispheric, parafloccular and floccular AP portions. In contrast, the smaller m2 mesomere only shows 1 AP alar domain (Puelles and Hidalgo-Sánchez 2023). These results accordingly cannot be generalized as yet, pending further studies. In any case, the m2 case may be exceptional due to its small size (Figure 4b). We think that advanced tripartite AP patterning within neuromeric alar fields may be yet found to exist in most neuromeres, allowing more distinct cell types to differentiate (there are also theoretic patterning reasons implying that neuromeres should have three AP parts). We thus may tentatively calculate potential 6 alar DV units x 3 alar AP units x 50 neuromeres = 900 DV/AP alar microzones, which, added to basal units, reach a new approximate total of 1,250 microzones. Note some brain alar areas, as the cerebral cortex (in hp1), show disproportionate tangential growth and seem to differentiate into many more than the 3 AP and 6 DV alar microzones of the average neuromere, mounting to some 200 alar cortical areas recently evaluated in the human isocortex (Nieuwenhuys et al. 2015), without counting other cortical areas, the complex subpallium that produces multiple types of interneurons (Flames et al. 2007), or the amygdala complex (Garcia-Calero et al. 2020). This potential result of progressive DV and AP microzonal patterning within neuromeric alar domains conceivably extends theoretical causal explanation to 1,250 or more microzones beyond columns and neuromeres. This very rough calculation implies now only production of 4,4 cell types on average per microzone. We already know several cases of microzones that produce sequentially a few different cell types over time (Figure 5g).
Areal Stratification and Clonal Typological Variation
A final important variable in this anti- ‘potato-sack’ brain regionalization theory is thus provided by the fourth dimension, time. The initial generative potency of microzonal progenitors often changes qualitatively and quantitatively over the local neurohistogenetic period, producing variously stratified or salt-and-pepper-mixed different cell types derived over time from a single microzonal multiclonal progenitor domain (e.g., the diverse retinal cell types, the 6-7 glutamatergic cell types produced sequentially at isocortical areas, or the 6-7 clonal cell types in the cerebellum apart of granule cells; Mathis et al. 1997 -see Figure 6g). The avian superior colliculus homolog -the optic tectum - forms 14 stratified and morphologically distinct neuronal types. Using the last present calculation, we only need on average the generation of 4-5 typologically diversified cell populations at each of the 1,250 microzones (some units may have less, others we know certainly have more) to reach the expected range of 5,300 neuronal types.
Recapitulation and Tangential Migrations
Looking back at our rationale as it applies in the case of the adult hypothalamus, Figure 7 displays first the ‘potato-sack-like’ unclassified and unexplained structure offered by columnar authors (Figure 7a,b; Krieg 1932; Dong 2008). Next we see AP (neuromeric) and DV (alar and basal microzonal) subdivisions postulated by a neuromeric author (Figures 6b,c; 7c; Puelles reference atlas for the P56 Allen Developing Mouse Brain Atlas in 2011; inset c’ is a detail of the alar paraventricular nucleus, showing internal tripartition according to c, consistent with adult Otp gene expression). Figure 7d takes as example the retromamillary basal area within the hp1 neuromere, where we already see two different populations labeled for two gene markers. Their behavior leads to a mixed (fuchsia and green) migration stream of retromamillary neurons, mapped with Foxa1-green- and Nr4a2 -fuchsia-, that exit hp1, enter the hp2 neuromere while eschewing invasion of the neighboring mamillary body, reach the ventral tuberal microzone and form there the compact ventral premamillary nucleus (PMV; Figure 7d,e; López-González et al. 2021). The higher magnification detail in Figure 7e illustrates multiple cell types in this migration (encircled green, white, yellow, fuchsia and red-fluorescent cells, indicating various combinations of the two gene markers used, together with the fluorescent migration tag). This multiplicity of PMV cell types was corroborated by single-cell transcriptomic studies discussed in the paper. This highlights surprising partial aspects of the causal cellular origin within a particular hypothalamic classic ‘potato’, the PMV nucleus, which was already known but not explained by Krieg (1932; Pv in Figure 2). The well-known subthalamic nucleus results also from a multitypological ventrodorsal migration which originates separately from the retromamillary area, though columnar tradition unaccountably does not recognize this nucleus as being hypothalamic (e.g., absent in Figure 7a from Krieg 1932; see López-González et al. 2021). A single microzone thus can give rise to multiple cell types in different parallel tangential migrations (unless the retromamillary area is actually a pair of microzones; the adjacent mamillary area also displays multiple subnuclei; Figure 7a). The prosomeric model allows further subdivisions.
As occurs in this case, different local neuron types of a microzone may stay aggregated together at a specific stratum or nucleus, or may rather mix in various ways with other neighboring cell types within the microzone or a larger enclosing primary alar or basal DV domain (when not moving between these; see Figure 6h). This microzonal-specific behavior pertaining to the precise positioning of individual neuronal types produced probably involves subtle neuronal adhesive and guidance properties we still ignore. Its analysis probably will explain in the future different sorts of cytoarchitectural nuclear or cortical configurations.
A different common phenomenon is that aligned similar neuromeric units with subtle molecular differences may compose a modular motor or sensory column (Marin and Puelles 1994; Cambronero and Puelles 2000; Marin et al. 2008; Tomás-Roca et al. 2016; García-Guillén et al. 2021). Some cell types may singly or collectively migrate out of their generative microzonal unit and neuromere, and incorporate functionally into neighboring microzones, columns, or neuromeres, or translocate actively into more distant neural domains. Tangential migrations were once thought to be rare, but now we realize that they occur often, in many parts of the brain, and always in a highly reproducible pattern, meaning they are under molecular control.
The logic for understanding brain cell type diversity at many brain positions thus necessarily involves distinguishing local intrinsic neurons from tangentially migrated neurons with more or less distant origins.
Further Considerations
If the reader belongs to those desiring the brain to be simple, he/she is in for deception. If 5,300 cell types were to wander around with unconstrained liberty, we would have a truly chaotic ‘potato-sack’ problem. That is not the expected scenario, though, since partial evidence at hand at the Whole Mouse Brain Atlas already indicates that many cell types respect specific boundaries (as in the case of the PMV nucleus); we only need to understand molecularly how these boundaries are first fixed and then how they are respected. We need more precise molecular identification of operative boundaries and chaos-restrictive guidance effects within and between the say 1,250 microzones in the brain. This seems a dire perspective, but we have merely 50 neuromeres (most of them in the repetitive spinal cord; only about 20 in the brain). Alternatively, we have just 3 tagmata: forebrain, hindbrain, and spinal cord. If the reader wants simplicity, generalities noted within a single tagma might be explored. While solid books probably can be written about the functions of any of these large brain domains, even disregarding their intrinsic neuromeric and microzonal phenomena (as anatomists and physiologists are used to do so far), there might persist a fogginess of conclusions due to lack of sufficient attention to the complex microzonal participation in those functions. If the reader rather prefers to be as close to the truth as possible, the complete microzonal scenario must be explored, similarly as was done with the atomic and subatomic world at the frontier of chemistry and physics (per aspera ad astra). The neuromeres and proneuromeres are just intermediate-level structural brain concepts which possibly explain aspects of axonal guidance, cell migration, and synaptogenesis (see Puelles 2024 on functionality of neuromeres). Just be wary of columns as sole background concepts, or of the thought that because we know the cell types, we understand the necessary underpinning regionalization.
The new neuromorphology presented in Nieuwenhuys and Puelles (2016) suggests in essence replacing unexplained knowledge (e.g., Figures 7a,b) by means of morphological, molecular and causal pigeon-holing of the conventional ‘potato-sack-like’ data sets. This can bring us finally to understand as predictable order the cellular adhesive, connective and functional complexity that emerges within the central nervous system (Figures 7d,e). This feasible approach should illuminate our perspective on large-scale functional assemblies of neurons and related upper-level psychological phenomena. Eventually we will glimpse the ethereal ‘butterflies of the soul’ (a poetic Cajalian concept).
Conclusions
It may thus be postulated that 1) progressive developmental phenomena of early DV and AP patterning leading to primary longitudinal zones of His and the crisscrossing series of transverse neuromeres, 2) amplified by subsequent DV and AP microzonation of neuromeric fields (hierarchy of microzones), 3) plus temporally patterned clonal neuronal phenotypic variation and differential migration, stratification and/or aggregation of the cell types produced over time may well be collectively able to account for the present 5,300 distinct neurons of the BICCN consortium and their typical positions.
References
- Acampora D, Boyl PP, Martinez-Barrera JP, Annino A, Signore M, Simaone A. 2001. Otx genes in evolution: are they involved in instructing the vertebrate brain morphology? J Anat 199: 53-62.
- Amat, J.A.; Martínez-De-La-Torre, M.; Trujillo, C.M.; Fernández, B.; Puelles, L. Neurogenetic Heterochrony in Chick, Lizard, and Rat Mapped with Wholemount Acetylcholinesterase and the Prosomeric Model. Brain, Behav. Evol. 2022, 97, 48–82. [Google Scholar] [CrossRef] [PubMed]
- Bergquist, H.; Källěn, B. Notes on the early histogenesis and morphogenesis of the central nervous system in vertebrates. J. Comp. Neurol. 1954, 100, 627–659. [Google Scholar] [CrossRef] [PubMed]
- Bulfone, A.; Puelles, L.; Porteus, M.; Frohman, M.; Martin, G.; Rubenstein, J. Spatially restricted expression of Dlx-1, Dlx-2 (Tes-1), Gbx-2, and Wnt- 3 in the embryonic day 12.5 mouse forebrain defines potential transverse and longitudinal segmental boundaries. J. Neurosci. 1993, 13, 3155–3172. [Google Scholar] [CrossRef] [PubMed]
- Cambronero, F.; Puelles, L. Rostrocaudal nuclear relationships in the avian medulla oblongata: A fate map with quail chick chimeras. J. Comp. Neurol. 2000, 427, 522–545. [Google Scholar] [CrossRef] [PubMed]
- Diaz, C.; de la Torre, M.M.; Rubenstein, J.L.R.; Puelles, L. Dorsoventral Arrangement of Lateral Hypothalamus Populations in the Mouse Hypothalamus: a Prosomeric Genoarchitectonic Analysis. Mol. Neurobiol. 2022, 60, 687–731. [Google Scholar] [CrossRef] [PubMed]
- Dong, HW. 2008. The Allen Reference Atlas: A Digital Color Brain Atlas of C57BL/6J Male Mouse, John Wiley & Sons.
- Ferran, J.L.; Sánchez-Arrones, L.; Bardet, S.M.; Sandoval, J.E.; Martínez-De-La-Torre, M.; Puelles, L. Early pretectal gene expression pattern shows a conserved anteroposterior tripartition in mouse and chicken. Brain Res. Bull. 2008, 75, 295–298. [Google Scholar] [CrossRef] [PubMed]
- Ferran, J.; de Oliveira, E.D.; Merchán, P.; Sandoval, J.; Sánchez-Arrones, L.; Martínez-De-La-Torre, M.; Puelles, L. Genoarchitectonic profile of developing nuclear groups in the chicken pretectum. J. Comp. Neurol. 2009, 517, 405–451. [Google Scholar] [CrossRef] [PubMed]
- Flames, N.; Pla, R.; Gelman, D.M.; Rubenstein, J.L.R.; Puelles, L.; Marín, O. Delineation of Multiple Subpallial Progenitor Domains by the Combinatorial Expression of Transcriptional Codes. J. Neurosci. 2007, 27, 9682–9695. [Google Scholar] [CrossRef]
- García-Calero E, Fernández-Garre P, Martínez S, Puelles L. 2008. Early mamillary pouch specification in the course of prechordal ventralization of the forebrain tegmentum. Devel Biol 320:366-377.
- Garcia-Calero, E.; Martinez-De-La-Torre, M.; Puelles, L. A radial histogenetic model of the mouse pallial amygdala. Anat. Embryol. 2020, 225, 1921–1956. [Google Scholar] [CrossRef]
- García-Guillén, I.M.; Martínez-De-La-Torre, M.; Puelles, L.; Aroca, P.; Marín, F. Molecular Segmentation of the Spinal Trigeminal Nucleus in the Adult Mouse Brain. Front. Neuroanat. 2021, 15, 785840. [Google Scholar] [CrossRef]
- Herrick, C.J. The morphology of the forebrain in amphibia and reptilia. J. Comp. Neurol. Psychol. 1910, 20, 413–547. [Google Scholar] [CrossRef]
- His, W. 1893. Über das frontale Ende des Gehirnrohres. Arch Anat EntwGesch 1893:157-171.
- Ju, M.; Aroca, P.; Luo, J.; Puelles, L.; Redies, C. Molecular profiling indicates avian branchiomotor nuclei invade the hindbrain alar plate. Neuroscience 2004, 128, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Krieg, W.J.S. The hypothalamus of the albino rat. J. Comp. Neurol. 1932, 55, 19–89. [Google Scholar] [CrossRef]
- Locy, W.A. Contribution to the structure and development of the vertebrate head. J. Morphol. 1895, 11, 497–594. [Google Scholar] [CrossRef]
- López-González, L.; Alonso, A.; García-Calero, E.; de Puelles, E.; Puelles, L. Tangential Intrahypothalamic Migration of the Mouse Ventral Premamillary Nucleus and Fgf8 Signaling. Front. Cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef]
- Lorente-Cánovas, B.; Marín, F.; Corral-San-Miguel, R.; Hidalgo-Sánchez, M.; Ferrán, J.L.; Puelles, L.; Aroca, P. Multiple origins, migratory paths and molecular profiles of cells populating the avian interpeduncular nucleus. Dev. Biol. 2012, 361, 12–26. [Google Scholar] [CrossRef]
- Marín, F.; Aroca, P.; Puelles, L. Hox gene colinear expression in the avian medulla oblongata is correlated with pseudorhombomeric domains. Dev. Biol. 2008, 323, 230–247. [Google Scholar] [CrossRef] [PubMed]
- Masland, RH. 2004. Neuronal cell types. Curr Evol 14:497-500.
- Mathis L, Bonnerot C, Puelles L, Nicolas J-F. 1997. Retrospective clonal analysis of the cerebellum using genetic laacZ/lacZ mouse mutants. Devel 124:4089-4104.
- McClure, C.F.W. The segmentation of the primitive vertebrate brain. J. Morphol. 1890, 4, 35–56. [Google Scholar] [CrossRef]
- Nieuwenhuys, R.; Puelles, L. Towards a New Neuromorphology; Springer Science and Business Media LLC: Dordrecht, GX, Netherlands, 2016. [Google Scholar]
- Nieuwenhuys, R. Principles of Current Vertebrate Neuromorphology. Brain, Behav. Evol. 2017, 90, 117–130. [Google Scholar] [CrossRef]
- Nieuwenhuys, R.; Broere, C.A.J.; Cerliani, L. A new myeloarchitectonic map of the human neocortex based on data from the Vogt–Vogt school. Anat. Embryol. 2014, 220, 2551–2573. [Google Scholar] [CrossRef]
- Orr, H. 1887. Contribution to the embryology of the lizard. J Morphol 1:311-372. [CrossRef]
- Puelles, L. 2013. Plan of the developing vertebrate nervous system relating embryology to the adult nervous system (prosomere model, overview of brain organization). In: “Comprehensive Developmental Neuroscience: Patterning and Cell Type Specification in the Developing CNS and PNS”, pp 187-209, Rubenstein JLR and Rakic P (eds), Amsterdam: Academic Press.
- Puelles, L. 2017. Role of secondary organizers in the evolution of forebrain development in vertebrates. In: Handbook of Evolutionary Neuroscience. (S.V. Shepherd, ed) Blackwell-Wiley: Chichester, UK. pp. 350-387.
- Puelles, L. Developmental studies of avian brain organization. Int. J. Dev. Biol. 2018, 62, 207–224. [Google Scholar] [CrossRef]
- Puelles, L. Functional Implications of the Prosomeric Brain Model. Biomolecules 2024, 14, 331. [Google Scholar] [CrossRef]
- Puelles, L.; Rubenstein, J.L. Expression patterns of homeobox and other putative regulatory genes in the embryonic mouse forebrain suggest a neuromeric organization. Trends Neurosci. 1993, 16, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Puelles, L.; Rubenstein, J.L. Forebrain gene expression domains and the evolving prosomeric model. Trends Neurosci. 2003, 26, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Puelles, L. and Martinez S. 2013. Patterning of the diencephalon. In: “Comprehensive Developmental Neuroscience: Patterning and Cell Type Specification in the Developing CNS and PNS”, pp. 151-172, Rubenstein JLR and Rakic P (eds), Amsterdam: Academic Press.
- Puelles, L.; Rubenstein, J.L.R. A new scenario of hypothalamic organization: rationale of new hypotheses introduced in the updated prosomeric model. Front. Neuroanat. 2015, 9, 27–27. [Google Scholar] [CrossRef]
- Puelles, L.; Hidalgo-Sánchez, M. The Midbrain Preisthmus: A Poorly Known Effect of the Isthmic Organizer. Int. J. Mol. Sci. 2023, 24, 9769. [Google Scholar] [CrossRef]
- Puelles L, Martinez-de-la-Torre M, Bardet S, Rubenstein, JLR. 2012a. Hypothalamus. In “The Mouse Nervous System”, pp221-312, Watson C, Paxinos G, Puelles L (eds), New York: Academic Press/Elsevier.
- Puelles E, Martinez-de-la-Torre M, Watson C, Puelles L. 2012b. Midbrain. In «The Mouse Nervous System », pp 337-359, Watson C, Paxinos G, Puelles L (eds), New York: Academic Press/Elsevier.
- Puelles L, Fernández B, Martinez-de-la-Torre M. 2015. Neuromeric landmarks in the rat midbrain, diencephalon and hypothalamus, compared with acetylcholinesterase histochemistry. In: The Rat Nervous System, Fourth Edition (Paxinos G., ed), New York, Academic Press/Elsevier pp.25-43.
- Puelles, L.; Tvrdik, P.; Martínez-De-La-Torre, M. The Postmigratory Alar Topography of Visceral Cranial Nerve Efferents Challenges the Classical Model of Hindbrain Columns. Anat. Rec. 2018, 302, 485–504. [Google Scholar] [CrossRef] [PubMed]
- Puelles, L.; Diaz, C.; Stuhmer, T.; Ferran, J.L.; la Torre, M.M.-D.; Rubenstein, J.L.R. LacZ-reporter mapping of Dlx5/6 expression and genoarchitectural analysis of the postnatal mouse prethalamus. J. Comp. Neurol. 2021, 529, 367–420. [Google Scholar] [CrossRef]
- Rubenstein, J.L.R.; Martinez, S.; Shimamura, K.; Puelles, L. The Embryonic Vertebrate Forebrain: the Prosomeric Model. Science 1994, 266, 578–580. [Google Scholar] [CrossRef]
- Sánchez-Arrones, L.; Ferrán, J.L.; Rodríguez-Gallardo, L.; Puelles, L. Incipient forebrain boundaries traced by differential gene expression and fate mapping in the chick neural plate. Dev. Biol. 2009, 335, 43–65. [Google Scholar] [CrossRef]
- Swanson, LW. 2004. Brain Maps: Structure of the Rat Brain (Third Edition). Elsevier Academic Press, San Diego, CA.
- Swanson, LW. 2012. Brain Architecture. Understanding the Basic Plan. (Second edition) Oxford: Oxford University Press.
- Tomás-Roca, L.; Corral-San-Miguel, R.; Aroca, P.; Puelles, L.; Marín, F. Crypto-rhombomeres of the mouse medulla oblongata, defined by molecular and morphological features. Anat. Embryol. 2014, 221, 815–838. [Google Scholar] [CrossRef] [PubMed]
- Vaage, S. The segmentation of the primitive neural tube in chick embryos (Gallus domesticus). A morphological, histochemical and autoradiographical investigation. Adv Anat Embryol Cell Biol. 1969, 41, 3–87. [Google Scholar]
- Von Kupffer, C. 1906. Die Morphogenesis des Centralnervensystems. In: Handbuch der vergleichenden und experimentellen Entwicklungslehre der Wirbeltiere (Hertwig O, Ed), Bd.2, Teil 3. Jena:Gustav Fisher.
- Watson C, Shimogori T, Puelles L. 2017a. Mouse FGF8-Cre lineage analysis defines the territory of the postnatal mammalian isthmus. J Comp Neurol 525:2782-2799.
Figure 1.
Models mentioned in the text. a, Columnar model of Herrick (1910), postulating an unbent length axis ending in the telencephalon. Note the notochord lies only under the brainstem (Mes, Rhomb), but the postulated longitudinal columns also extend into the forebrain (Dienc, Tel). The rostral ends of the floor and roof domains were not indicated. b,c,d, Cross-sections through Tel, Dienc and Rhomb in a, illustrating the postulated four columns at each level. e, Modification of Herrick’s columnar model proposed by Swanson (2004, 2012). Essentially, a general division into basal (cream) and alar (blue) plates is proposed, again without topographic correlation with the notochord. Roof and floor are not mapped precisely, but the postulated floor possibly reaches the anterior commissure (ac), implying a prechordal part. f, Neuromeric model of Bergquist and Källén (1954), encompassing also His’s (1893, 1904) longitudinal zones (names at caudal end) and alar-basal boundary (red dashes). Note Tel forms a unit with the hypothalamus, the secondary prosencephalon (green). The diencephalon lies caudal to that and consists of three transverse neuromeres, thought to represent more faithfully the domains with identical color codes in a,c. All neuromeres extend from roof to floor, whose rostral ends are not represented. The intersection of longitudinal and transversal limits creates a number of quadrangular ‘migration areas’. M indicates mamillary body. g, Neuromeric (prosomeric) model of Puelles et al. (2012; updated significantly relative to earlier versions). There are three parallel axial references: 1) the floor (blue) ending rostrally at mamillary body, 2) the alar-basal boundary (red dash-line; determined molecularly in the forebrain by Shh and Nkx2.2 markers), and 3) the roof plate (yellow), fate-mapped in several vertebrate species to end at the anterior commissure (ac). The concept of unitary secondary prosencephalon from f is maintained, subdivided into two hypothalamo-telencephalic prosomeres (hp1, hp2), whose floor is retromamillary (RM) or mamillary (M). The respective parts of hypothalamus are named ‘peduncular hypothalamus’ (PHy) and ‘terminal hypothalamus (THy). Hp2 ends rostrally at the acroterminal rostromedian domain (green), stretched between rostral roof and rostral floor, which displays unique formations such as the alar preoptic lamina terminalis and the optic chiasma, and the basal infundibular tuberal region with the neurohypophysis. The dienc is trineuromeric, like in f. The midbrain has two neuromeres (m1, m2) and ends at the isthmo-mesencephalic boundary (thick black dashes). The hindbrain shows 12 rhombomeres, some of which are cryptic (detected only molecularly and experimentally). The hindbrain has four proneuromeres (prepontine: pp, pontine: pons, retropontine: rpon, and medulla: med; limits as intermediate dashes). The spinal cord (beyond the rhombospinal boundary; thick black dashes) has myelomeres.
Figure 1.
Models mentioned in the text. a, Columnar model of Herrick (1910), postulating an unbent length axis ending in the telencephalon. Note the notochord lies only under the brainstem (Mes, Rhomb), but the postulated longitudinal columns also extend into the forebrain (Dienc, Tel). The rostral ends of the floor and roof domains were not indicated. b,c,d, Cross-sections through Tel, Dienc and Rhomb in a, illustrating the postulated four columns at each level. e, Modification of Herrick’s columnar model proposed by Swanson (2004, 2012). Essentially, a general division into basal (cream) and alar (blue) plates is proposed, again without topographic correlation with the notochord. Roof and floor are not mapped precisely, but the postulated floor possibly reaches the anterior commissure (ac), implying a prechordal part. f, Neuromeric model of Bergquist and Källén (1954), encompassing also His’s (1893, 1904) longitudinal zones (names at caudal end) and alar-basal boundary (red dashes). Note Tel forms a unit with the hypothalamus, the secondary prosencephalon (green). The diencephalon lies caudal to that and consists of three transverse neuromeres, thought to represent more faithfully the domains with identical color codes in a,c. All neuromeres extend from roof to floor, whose rostral ends are not represented. The intersection of longitudinal and transversal limits creates a number of quadrangular ‘migration areas’. M indicates mamillary body. g, Neuromeric (prosomeric) model of Puelles et al. (2012; updated significantly relative to earlier versions). There are three parallel axial references: 1) the floor (blue) ending rostrally at mamillary body, 2) the alar-basal boundary (red dash-line; determined molecularly in the forebrain by Shh and Nkx2.2 markers), and 3) the roof plate (yellow), fate-mapped in several vertebrate species to end at the anterior commissure (ac). The concept of unitary secondary prosencephalon from f is maintained, subdivided into two hypothalamo-telencephalic prosomeres (hp1, hp2), whose floor is retromamillary (RM) or mamillary (M). The respective parts of hypothalamus are named ‘peduncular hypothalamus’ (PHy) and ‘terminal hypothalamus (THy). Hp2 ends rostrally at the acroterminal rostromedian domain (green), stretched between rostral roof and rostral floor, which displays unique formations such as the alar preoptic lamina terminalis and the optic chiasma, and the basal infundibular tuberal region with the neurohypophysis. The dienc is trineuromeric, like in f. The midbrain has two neuromeres (m1, m2) and ends at the isthmo-mesencephalic boundary (thick black dashes). The hindbrain shows 12 rhombomeres, some of which are cryptic (detected only molecularly and experimentally). The hindbrain has four proneuromeres (prepontine: pp, pontine: pons, retropontine: rpon, and medulla: med; limits as intermediate dashes). The spinal cord (beyond the rhombospinal boundary; thick black dashes) has myelomeres.
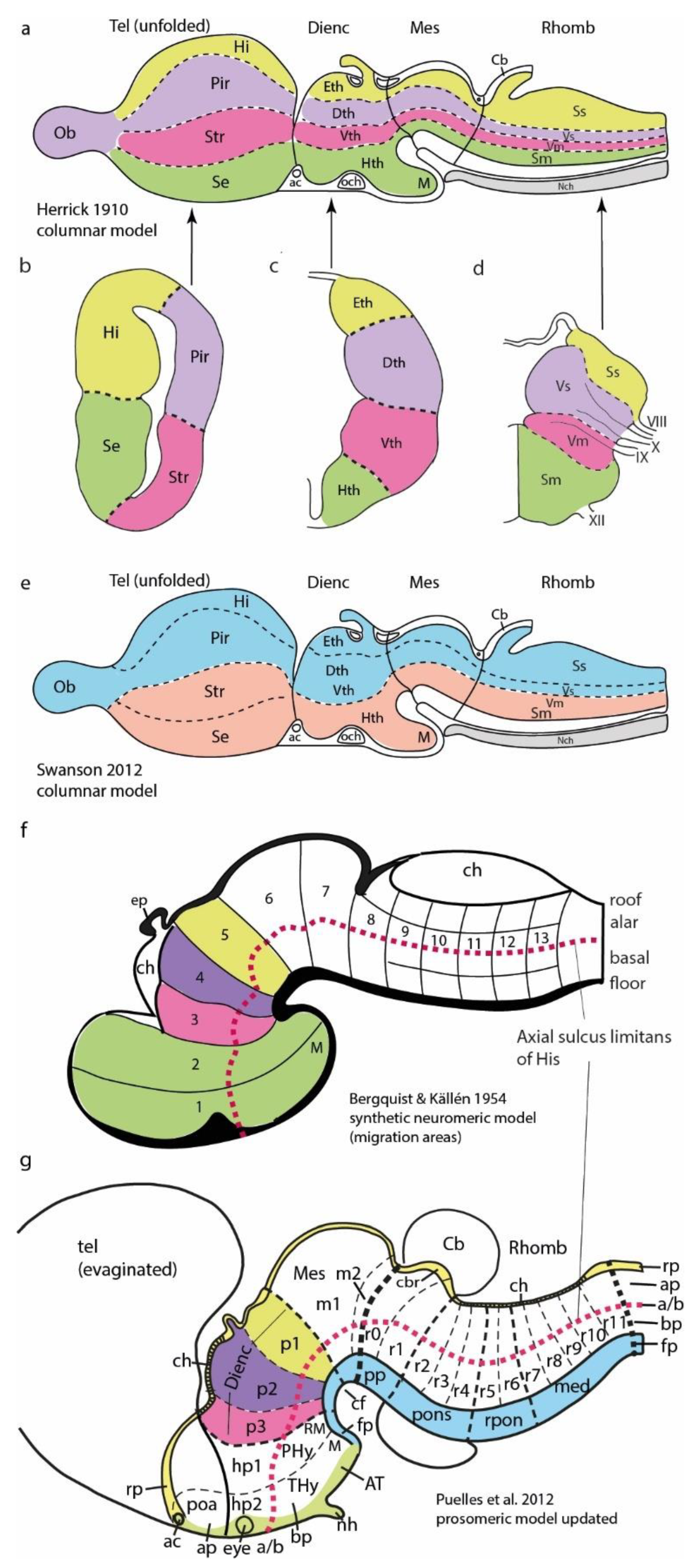
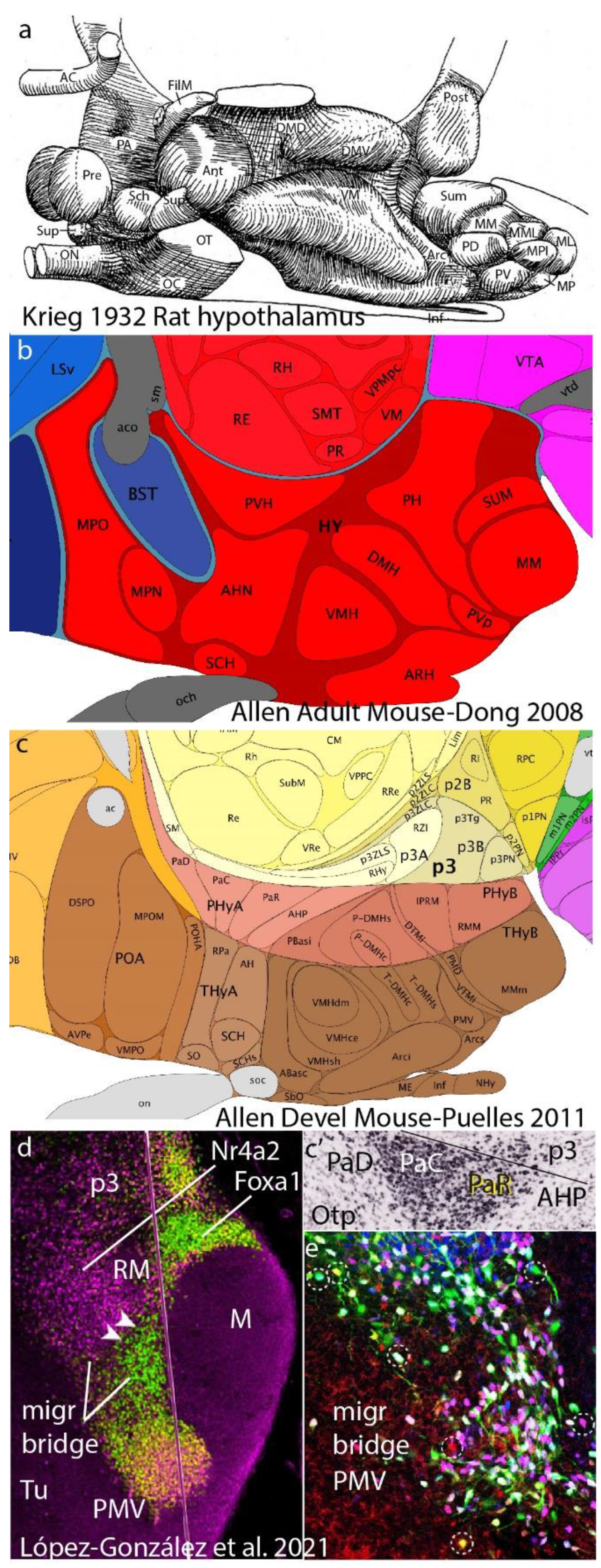
Figure 2.
Krieg’s (1932) nuclear structure of the rat hypothalamus. Krieg produced this relatively precise 3D view of hypothalamic nuclear variety in his Figure 21 (note the hypothalamus was one of Herrick’s diencephalic columns). There is no apparent order in the nuclear arrangement. The columnar caudorostral axis runs from right to left, so that the bottom of the reconstruction (Inf) was regarded as the floor of the diencephalon. We see the optic nerve (ON) and the anterior commissure (AC) at the rostral end. AC anterior commissure, Ant anterior nucleus, Arc arcuate nucleus, DMD dorsal part, dorsomedial nucleus, DMV ventral part, dorsomedial nucleus, Inf infundibulum, ML lateral mamillary nucleus, MM medial mamillary nucleus, MML mediomedial mamillary nucleus, MPI posterior intermediate mamillary nucleus, MP posterior mamillary nucleus, PA preoptic area, FilM filiform (paraventricular) nucleus, PD dorsal premamillary nucleus, Post posterior hypothalamic nucleus, Pre preoptic nucleus, PV ventral premamillary nucleus, OC optic chiasma, ON optic nerve, OT optic tract, Sch suprachiasmatic nucleus, Sum supramamillary nucleus, Sup supraoptic nucleus, VM ventromedial nucleus.
Figure 2.
Krieg’s (1932) nuclear structure of the rat hypothalamus. Krieg produced this relatively precise 3D view of hypothalamic nuclear variety in his Figure 21 (note the hypothalamus was one of Herrick’s diencephalic columns). There is no apparent order in the nuclear arrangement. The columnar caudorostral axis runs from right to left, so that the bottom of the reconstruction (Inf) was regarded as the floor of the diencephalon. We see the optic nerve (ON) and the anterior commissure (AC) at the rostral end. AC anterior commissure, Ant anterior nucleus, Arc arcuate nucleus, DMD dorsal part, dorsomedial nucleus, DMV ventral part, dorsomedial nucleus, Inf infundibulum, ML lateral mamillary nucleus, MM medial mamillary nucleus, MML mediomedial mamillary nucleus, MPI posterior intermediate mamillary nucleus, MP posterior mamillary nucleus, PA preoptic area, FilM filiform (paraventricular) nucleus, PD dorsal premamillary nucleus, Post posterior hypothalamic nucleus, Pre preoptic nucleus, PV ventral premamillary nucleus, OC optic chiasma, ON optic nerve, OT optic tract, Sch suprachiasmatic nucleus, Sum supramamillary nucleus, Sup supraoptic nucleus, VM ventromedial nucleus.
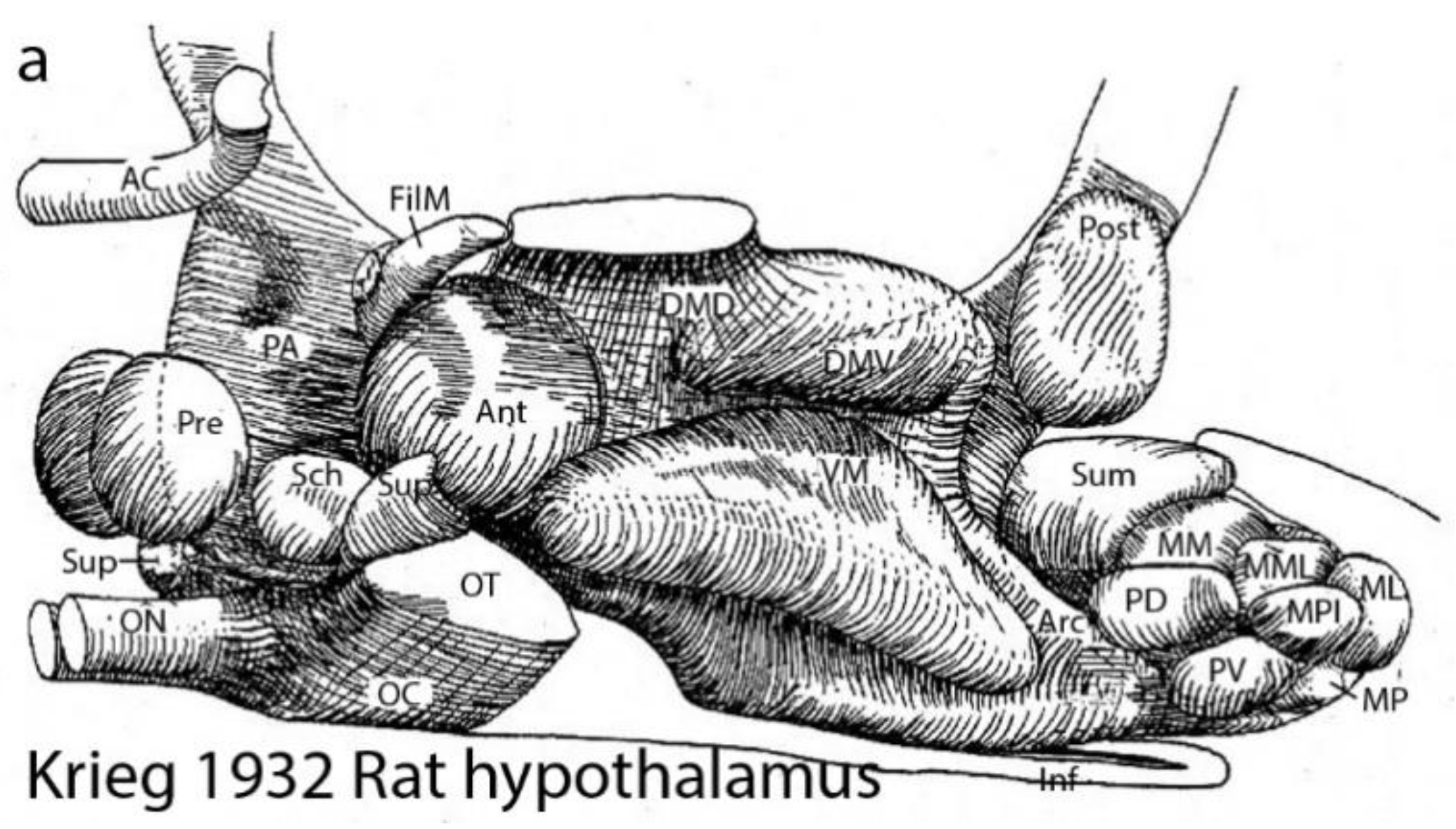
Figure 3.
The wiring of the human brain (Figure used in a 1992 talk, reproduced from Nieuwenhuys and Puelles 2016). Schema displaying sensory and motor pathways jointly with some interconnections.
Figure 3.
The wiring of the human brain (Figure used in a 1992 talk, reproduced from Nieuwenhuys and Puelles 2016). Schema displaying sensory and motor pathways jointly with some interconnections.
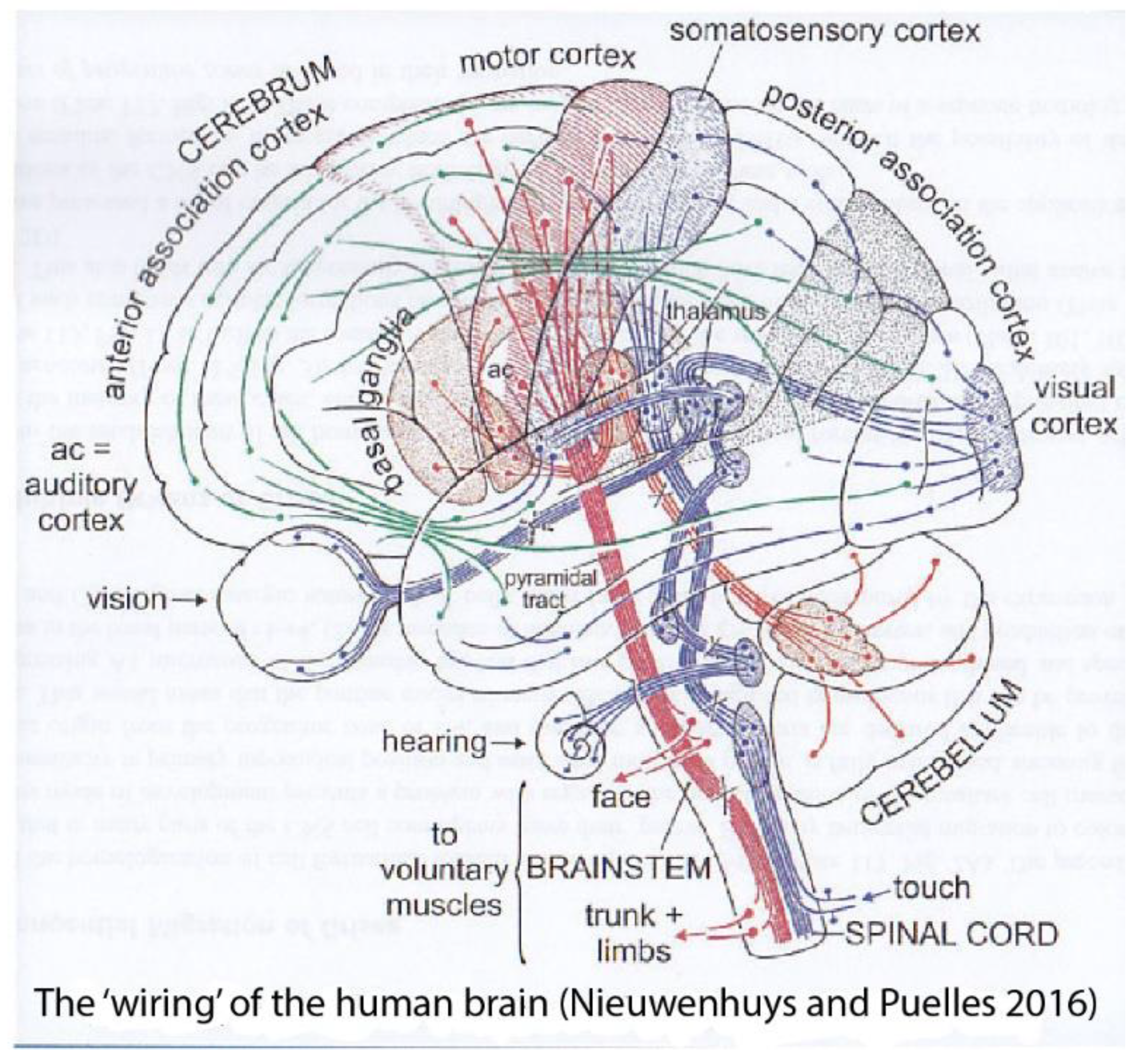
Figure 4.
The model of His (1893) and its DV elements, in modern perspective. a, Schema of His (1893) with his precise DV subdivision into floor, basal, alar, and roof longitudinal zones (including the limiting and appropriately bent alar-basal boundary sulcus ending at the optic stalk). His also entered relatively imprecise AP partitions (Roman/Arabic tags) now substituted by neuromeres. He was the first to postulate the isthmic domain (tagged III1/III2) as a separate brain segment (see Figure 5c). b, Mouse E10.5 brain showing the alar-basal band of Nkx2.2 expression (downstream of notochordal SHH signal) which approximates the sulcus of His in a. The neuromeric alar ventricular concavities of the three diencephalic segments are visible (p1, p2, p3). The zona limitans interthalamica (zl) is a transverse singularity of the p2/p3 boundary, caused by a separate Shh enhancer, secondarily affecting Nkx2.2. c,d, Floor plate labeling at E11.5 and E13.5 with Ntn1 (from Puelles et al. 2012; Allen Institute data). The floor stops rostrally over the tip of the notochord at the mamillary pouch (mam). Note also slight Ntn1 signal at E13.5 at the locus of the anterior commissure (fate-mapped rostralmost roof plate). e, Three-day old chick embryo brain reacted wholemount for AChE (from Amat et al. 2022). This marker identifies postmitotic neurons and is negative generally in progenitor cells (labeling at the zl -p2/p3 interneuromeric limit as in b- contrarily marks radial glia or progenitors). The neurogenetically precocious basal zone is massively full of neurons, while the retarded alar zone is relatively unpopulated at this stage. Note some transverse neuromeric borders, orthogonal to the alar-basal limit. The telencephalon shows a fully alar pattern, falsating columnar assumptions. f,g, Selective expression of Dlk1 at the mouse acroterminal domain at E13.5 and E15.5 (data from the Allen Developmental Mouse Brain Atlas).
Figure 4.
The model of His (1893) and its DV elements, in modern perspective. a, Schema of His (1893) with his precise DV subdivision into floor, basal, alar, and roof longitudinal zones (including the limiting and appropriately bent alar-basal boundary sulcus ending at the optic stalk). His also entered relatively imprecise AP partitions (Roman/Arabic tags) now substituted by neuromeres. He was the first to postulate the isthmic domain (tagged III1/III2) as a separate brain segment (see Figure 5c). b, Mouse E10.5 brain showing the alar-basal band of Nkx2.2 expression (downstream of notochordal SHH signal) which approximates the sulcus of His in a. The neuromeric alar ventricular concavities of the three diencephalic segments are visible (p1, p2, p3). The zona limitans interthalamica (zl) is a transverse singularity of the p2/p3 boundary, caused by a separate Shh enhancer, secondarily affecting Nkx2.2. c,d, Floor plate labeling at E11.5 and E13.5 with Ntn1 (from Puelles et al. 2012; Allen Institute data). The floor stops rostrally over the tip of the notochord at the mamillary pouch (mam). Note also slight Ntn1 signal at E13.5 at the locus of the anterior commissure (fate-mapped rostralmost roof plate). e, Three-day old chick embryo brain reacted wholemount for AChE (from Amat et al. 2022). This marker identifies postmitotic neurons and is negative generally in progenitor cells (labeling at the zl -p2/p3 interneuromeric limit as in b- contrarily marks radial glia or progenitors). The neurogenetically precocious basal zone is massively full of neurons, while the retarded alar zone is relatively unpopulated at this stage. Note some transverse neuromeric borders, orthogonal to the alar-basal limit. The telencephalon shows a fully alar pattern, falsating columnar assumptions. f,g, Selective expression of Dlk1 at the mouse acroterminal domain at E13.5 and E15.5 (data from the Allen Developmental Mouse Brain Atlas).
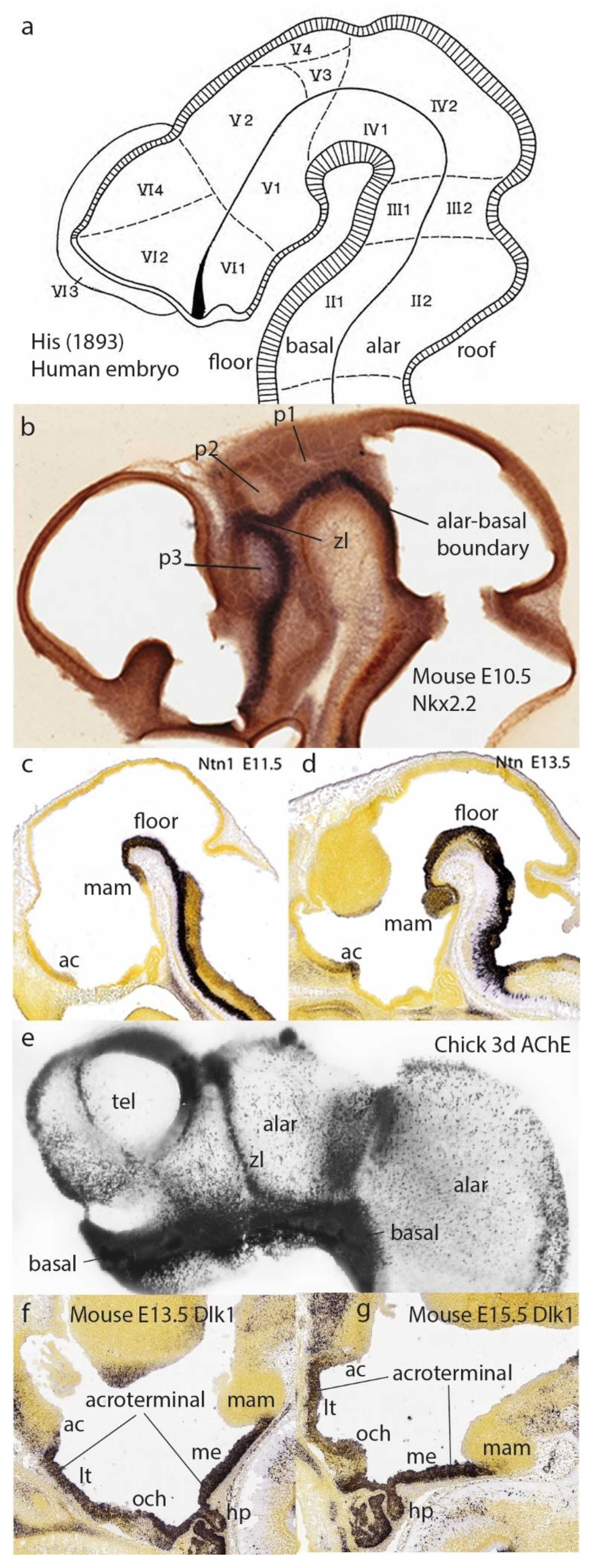
Figure 5.
Neuromeres, their DV subdivisions, plus underlying migratory and clonal aspects. a, Forebrain and hindbrain neuromeric bulges in the shark Squalus acanthias, drawn by Locy (1895). The hindbrain shows 5 preotic rhombomeres comprising prepontine and pontine units (now called r0-r4) and 3 postotic units (r5-r7) corresponding to retropontine (r5,r6) and the first medullary unit (r7). b, Graphic reconstruction of forebrain prosomeres in an E11.5 rat brain, created out of a sagittal section series. The postulated neuromeric cavities of hp1/hp2 (hypothalamus), p1-p3 (diencephalon) and m1/m2 (midbrain) were captured (Puelles et al. 2015); see the original for individual sections and correlated description of wholemount AChE-stained postmitotic neurons. c,d, Coronal and sagittal sections through transgenic adult mouse brains carrying a Fgf8-LacZ construct that labels all progeny derived from progenitors that expressed early on the Fgf8 gene (a marker typical of the embryonic isthmic rhombomere r0). The observed blue LacZ reaction demonstrates that the neuromere-derived adult brain part continues being transversal, limits with the caudal midbrain, participates in the vermal cerebellum, and is complete from ventricle to pia; i.e., the neuromere is still there with conserved boundaries, but has transformed into its adult counterpart (some blue-labeled cell populations seem to have migrated out of the isthmus into neighboring areas). Analogous material exists for other neuromeres. e,f, Schemata of fine microzonal regionalization subdividing the basal and alar domains, observed with molecular markers and experimentation at the spinal cord (e) and hindbrain neuromeres (f). Usually there are 5 basal microzones and 6-7 alar ones. Different neuronal populations are produced at each locus. The spinal DL domain (e) forms the layered dorsal horn, mixing derivatives from several microzones. In the basal plate, the MN microzone forms the motoneurons, whereas the others form different sorts of interneurons. In the hindbrain (f) the alar microzones form the sensory columns (color-coded) and the rhombic lip, dorsally, from where most neurons migrate subpially ventralwards, forming e.g. the pontine nuclei. Violet colored motoneurons form three separate columns, two of them migrated into the VL alar plate (these were classified classically as visceromotor, ignoring that all motoneurons -different subtypes- come from the same MN microzone). g, Radial migration in the cerebellar cortex of a transgenically LacZ-labeled clone composed of different neuron types (Purkinje: P, Golgi: G, Molecular: M), derived from a single very rare LacZ-reconstitutive event in a progenitor cell labeled by a non-functional LaacZ construct (Mathis et al. 1997). h, Pax7-LacZ progeny derives exclusively from the alar plate, so that one expects in the spinal cord only massive blue derivatives in the dorsal horn; however, a number of blue alar neurons apparently migrate into the ventral horn (basal plate), probably representing interneurons.
Figure 5.
Neuromeres, their DV subdivisions, plus underlying migratory and clonal aspects. a, Forebrain and hindbrain neuromeric bulges in the shark Squalus acanthias, drawn by Locy (1895). The hindbrain shows 5 preotic rhombomeres comprising prepontine and pontine units (now called r0-r4) and 3 postotic units (r5-r7) corresponding to retropontine (r5,r6) and the first medullary unit (r7). b, Graphic reconstruction of forebrain prosomeres in an E11.5 rat brain, created out of a sagittal section series. The postulated neuromeric cavities of hp1/hp2 (hypothalamus), p1-p3 (diencephalon) and m1/m2 (midbrain) were captured (Puelles et al. 2015); see the original for individual sections and correlated description of wholemount AChE-stained postmitotic neurons. c,d, Coronal and sagittal sections through transgenic adult mouse brains carrying a Fgf8-LacZ construct that labels all progeny derived from progenitors that expressed early on the Fgf8 gene (a marker typical of the embryonic isthmic rhombomere r0). The observed blue LacZ reaction demonstrates that the neuromere-derived adult brain part continues being transversal, limits with the caudal midbrain, participates in the vermal cerebellum, and is complete from ventricle to pia; i.e., the neuromere is still there with conserved boundaries, but has transformed into its adult counterpart (some blue-labeled cell populations seem to have migrated out of the isthmus into neighboring areas). Analogous material exists for other neuromeres. e,f, Schemata of fine microzonal regionalization subdividing the basal and alar domains, observed with molecular markers and experimentation at the spinal cord (e) and hindbrain neuromeres (f). Usually there are 5 basal microzones and 6-7 alar ones. Different neuronal populations are produced at each locus. The spinal DL domain (e) forms the layered dorsal horn, mixing derivatives from several microzones. In the basal plate, the MN microzone forms the motoneurons, whereas the others form different sorts of interneurons. In the hindbrain (f) the alar microzones form the sensory columns (color-coded) and the rhombic lip, dorsally, from where most neurons migrate subpially ventralwards, forming e.g. the pontine nuclei. Violet colored motoneurons form three separate columns, two of them migrated into the VL alar plate (these were classified classically as visceromotor, ignoring that all motoneurons -different subtypes- come from the same MN microzone). g, Radial migration in the cerebellar cortex of a transgenically LacZ-labeled clone composed of different neuron types (Purkinje: P, Golgi: G, Molecular: M), derived from a single very rare LacZ-reconstitutive event in a progenitor cell labeled by a non-functional LaacZ construct (Mathis et al. 1997). h, Pax7-LacZ progeny derives exclusively from the alar plate, so that one expects in the spinal cord only massive blue derivatives in the dorsal horn; however, a number of blue alar neurons apparently migrate into the ventral horn (basal plate), probably representing interneurons.
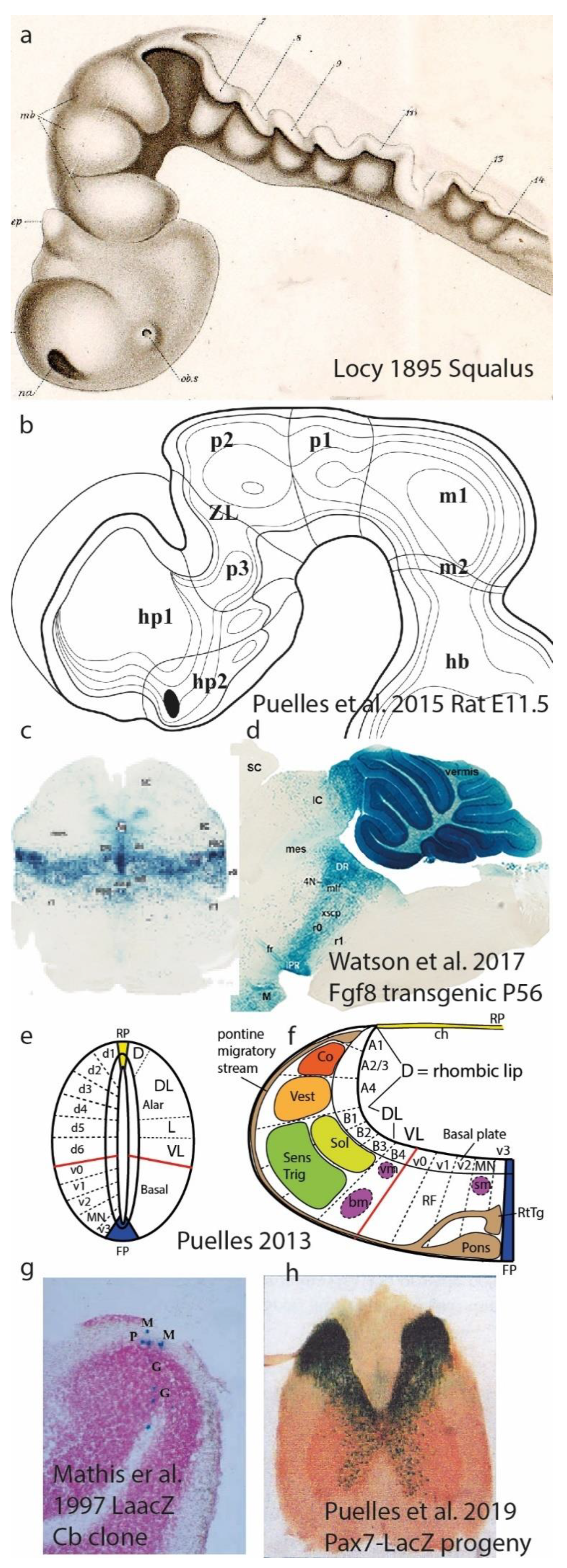
Figure 6.
Secondary (advanced) DV and AP microzonal regionalization of neuromeres. a, Prosomeric map of advanced DV patterning reported at different places of the neural tube, collected from literature and personal results by Nieuwenhuys and Puelles (2016). The mapping was already incomplete at forebrain levels at that date, due to hypothalamic data of Puelles et al. 2012 and unrepresented pretectal data from Ferran et al. 2009. However, it allows to appreciate the commonality of the pattern observed in spinal cord and hindbrain. b,c, These panels show partial molecular mapping data in the hypothalamus that suggest a systematic alar and basal DV subdivision into roughly parallel (longitudinal) molecularly diverse microzonal compartments, both at the lateral hypothalamus (b; Diaz et al. 2022) and in the general bineuromeric distribution of peptidergic neurons of several subtypes (c; Puelles et al. 2012). Several single cell transcriptomic studies corroborate multiple cell types of various sorts in these areas. Our studies correlated this distribution with our molecular characterization of the early progenitor domains. Note the optic chiasma roughly marks the alar-basal boundary. d-f, Illustration of advanced (secondary) AP microzonal tripartite pattern in the pretectal alar plate (p1 prosomere: precommissural PcP, juxtacommissural JcP, and commissural CoP AP compartments; Ferran et al. 2009). d shows an early stage previous to DV microzonation, while e displays the added DV subdivision. f presents a horizontal section in a 9d chick embryo (plane indicated in e by a blue bar), where the pretectum (alar p1; boundaries as dashed black lines) is seen lying intercalated between the thalamus (in alar p2) and the midbrain tectum (in alar m1). Foxp2 signal characterizes selectively intermediate and superficial strata of the rostral PcP subdomain, while PAX7 immunoreaction marks likewise diverse stratified CoP derivatives; the intermediate JcP subdomain is negative for these two markers, but stains specifically, e.g., for Six3. This result simultaneously shows the radially complete AP partition and reveals that each microzone forms different neuronal subtypes that occupy different layers of the microzonal mantle. Each of these derivatives is fated to produce different nuclei or cell layers, with differential connective properties. The m1 midbrain tectum also shows in this image a more thinly layered rostral microzonal component found next to the CoP, and a more thickly layered caudal component (the optic tectum). The signal-free boundary area between thalamus and PcP may represent the missing third thalamic subdivision (consistent with all neuromeres having this pattern).
Figure 6.
Secondary (advanced) DV and AP microzonal regionalization of neuromeres. a, Prosomeric map of advanced DV patterning reported at different places of the neural tube, collected from literature and personal results by Nieuwenhuys and Puelles (2016). The mapping was already incomplete at forebrain levels at that date, due to hypothalamic data of Puelles et al. 2012 and unrepresented pretectal data from Ferran et al. 2009. However, it allows to appreciate the commonality of the pattern observed in spinal cord and hindbrain. b,c, These panels show partial molecular mapping data in the hypothalamus that suggest a systematic alar and basal DV subdivision into roughly parallel (longitudinal) molecularly diverse microzonal compartments, both at the lateral hypothalamus (b; Diaz et al. 2022) and in the general bineuromeric distribution of peptidergic neurons of several subtypes (c; Puelles et al. 2012). Several single cell transcriptomic studies corroborate multiple cell types of various sorts in these areas. Our studies correlated this distribution with our molecular characterization of the early progenitor domains. Note the optic chiasma roughly marks the alar-basal boundary. d-f, Illustration of advanced (secondary) AP microzonal tripartite pattern in the pretectal alar plate (p1 prosomere: precommissural PcP, juxtacommissural JcP, and commissural CoP AP compartments; Ferran et al. 2009). d shows an early stage previous to DV microzonation, while e displays the added DV subdivision. f presents a horizontal section in a 9d chick embryo (plane indicated in e by a blue bar), where the pretectum (alar p1; boundaries as dashed black lines) is seen lying intercalated between the thalamus (in alar p2) and the midbrain tectum (in alar m1). Foxp2 signal characterizes selectively intermediate and superficial strata of the rostral PcP subdomain, while PAX7 immunoreaction marks likewise diverse stratified CoP derivatives; the intermediate JcP subdomain is negative for these two markers, but stains specifically, e.g., for Six3. This result simultaneously shows the radially complete AP partition and reveals that each microzone forms different neuronal subtypes that occupy different layers of the microzonal mantle. Each of these derivatives is fated to produce different nuclei or cell layers, with differential connective properties. The m1 midbrain tectum also shows in this image a more thinly layered rostral microzonal component found next to the CoP, and a more thickly layered caudal component (the optic tectum). The signal-free boundary area between thalamus and PcP may represent the missing third thalamic subdivision (consistent with all neuromeres having this pattern).
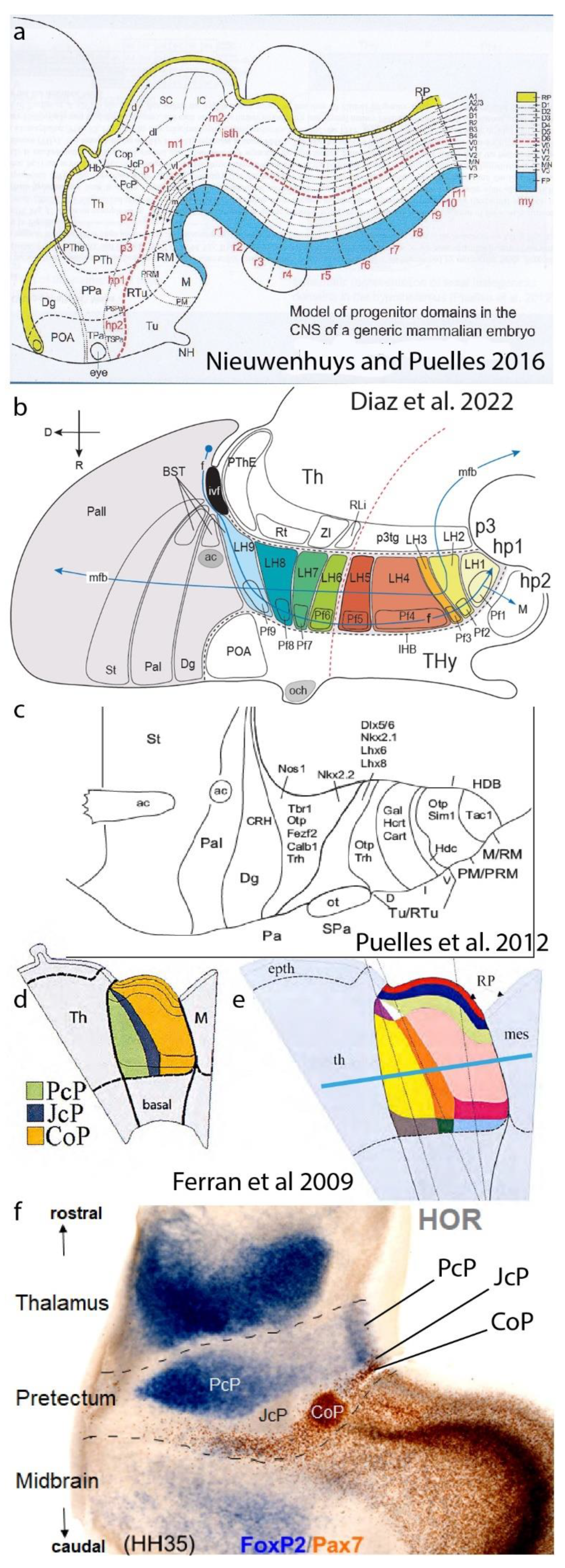
Figure 7.
Recapitulation of our argument using the hypothalamus as example. a, b. These panels show two examples of columnar order-less description of nuclei in the hypothalamus, by Krieg (1932) in a, and Dong (2008) in b, representing examples of what we have called ‘potato-sack morphology’, that is, description without any sort of classificatory or explaining concepts. c, Here LP, working in collaboration with the Allen Institute on the Ontology and Reference Atlases of the Developing Mouse Brain Atlas (2008-2011), proposed AP partition of the P56 hypothalamus into two hypothalamic neuromeres, defining their respective floor, basal, alar and roof domains plus DV microzonal divisions. The known adult nuclei fell naturally into the resulting intersectional pigeon-holes, and were consistent with gene patterns analyzed. This effort using some 4,000 genes at the Allen Institute helped evolving the more complete, updated prosomeric model of Puelles et al. 2012a. d,e, Jumping now to phenomena in a single microzone from López-González et al. 2021, we show a sagittal image of the retromamillary area (RM; a basal plate area of hp1 next to the floor, understood within columnar theory as ‘supramamillary area’) where we see distinct ventral and dorsal subareas labeled differentially for Foxa1 (green) and Nr4a2 (fuchsia). Both populations generate cells that enter a rostralward migration stream that courses above the mamillary body into hp2, and enters the suprajacent ventral tuberal area. As the cells approach rostrally the acroterminal area (which may have attracted them) they stop migrating, forming an oval-shaped nucleus, the classically known ventral premamillary nucleus. There is very little cell mixing with cells derived from hp2, whose cells often express quite different gene markers. In contrast, the PMV migration is composed of intermixed green- and fuchsia-labeled RM cells, which subdivide into several intermixed subgroups according to which of the two markers they express. The final PMV nucleus has a molecularly distinguishable core and shell structure with multiple cell types.
Figure 7.
Recapitulation of our argument using the hypothalamus as example. a, b. These panels show two examples of columnar order-less description of nuclei in the hypothalamus, by Krieg (1932) in a, and Dong (2008) in b, representing examples of what we have called ‘potato-sack morphology’, that is, description without any sort of classificatory or explaining concepts. c, Here LP, working in collaboration with the Allen Institute on the Ontology and Reference Atlases of the Developing Mouse Brain Atlas (2008-2011), proposed AP partition of the P56 hypothalamus into two hypothalamic neuromeres, defining their respective floor, basal, alar and roof domains plus DV microzonal divisions. The known adult nuclei fell naturally into the resulting intersectional pigeon-holes, and were consistent with gene patterns analyzed. This effort using some 4,000 genes at the Allen Institute helped evolving the more complete, updated prosomeric model of Puelles et al. 2012a. d,e, Jumping now to phenomena in a single microzone from López-González et al. 2021, we show a sagittal image of the retromamillary area (RM; a basal plate area of hp1 next to the floor, understood within columnar theory as ‘supramamillary area’) where we see distinct ventral and dorsal subareas labeled differentially for Foxa1 (green) and Nr4a2 (fuchsia). Both populations generate cells that enter a rostralward migration stream that courses above the mamillary body into hp2, and enters the suprajacent ventral tuberal area. As the cells approach rostrally the acroterminal area (which may have attracted them) they stop migrating, forming an oval-shaped nucleus, the classically known ventral premamillary nucleus. There is very little cell mixing with cells derived from hp2, whose cells often express quite different gene markers. In contrast, the PMV migration is composed of intermixed green- and fuchsia-labeled RM cells, which subdivide into several intermixed subgroups according to which of the two markers they express. The final PMV nucleus has a molecularly distinguishable core and shell structure with multiple cell types.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



