
Submitted:
29 April 2024
Posted:
01 May 2024
You are already at the latest version
Abstract
Prostate cancer, according to the World Health Organization (WHO), ranks as the fourth most prevalent type in the population, even though it is highly frequent when detected early and promptly treated, resulting in a low mortality rate. However, in some cases, despite adherence to proper clinical protocols, there is a progression to castration-resistant prostate cancer (CRPC). In these cases, as outlined in clinical guidelines such as those developed by the National Comprehensive Cancer Network (NCCN), one approach involves the utilization of the pharmaceutical agent abiraterone to implement androgen deprivation therapy (ADT), aimed at suppressing the production of hormones directly implicated in disease progression. Another noteworthy aspect is the limited literature on clinical protocols and their efficacy concerning the Amazonian indigenous population, which often exhibits unique genomic profiles, with descriptions of variants not yet documented in the literature. Building upon these considerations, this study aims to survey the variants identified in genes associated with the abiraterone signaling pathway, their frequencies in the Amazonian indigenous population, and the correlation of these frequencies with those observed in other global populations. The objective is to contribute to the refinement of clinical practices in the treatment of prostate cancer within this population.
Keywords:
abiraterone
; prostate
; amazonian
; indigenous
; cancer
1. Introduction
The worldwide incidence of prostate cancer, as reported by the World Health Organization, reached 1.2 million individuals, securing the fourth position in terms of case numbers. The mortality rate stands at approximately 40%, with studies suggesting that poorer prognoses are more closely associated with the European population [21].
The clinical approach to addressing this type of cancer depends on the stage of the disease. In more advanced stages, according to the National Comprehensive Cancer Network (NCCN), one of the approaches involves implementing androgen deprivation therapy (ADT). In this context, one of the drugs used is abiraterone [17], which acts as a selective inhibitor of the enzyme CYP17.
Even after the implementation of ADT protocols, some patients develop a more aggressive form of the disease called castration-resistant prostate cancer (CRPC), in which there is high expression of steroid hormones that ultimately promote disease recurrence and shorter survival time.
After studies, it has been observed that individuals who progress to CRPC exhibit variants in the HSD3B1 gene. This gene is already described as a biomarker for the occurrence of CRPC and has clinical indications on the PharmGKBD portal [1,3] and regulatory bodies such as the FDA.
Focusing on the ADT protocol, the signaling pathway of the drug abiraterone was examined, revealing five related genes: SLCO2B1, SULT2A1, CYP3A4, HSD3B1, HSD3B2, and UGT1A4. As can be seen, the HSD3B1 gene, associated with CRPC occurrence, is present in this pathway.
The aim of this article is to assess variants found in genes of the abiraterone signaling pathway in the indigenous population of the Amazon, including the variant of the HSD3B1 gene related as a biomarker for CRPC, and compare the frequencies found with those already verified in other global populations. The analysis also seeks to validate the unique character of the indigenous population studied, reinforcing the need for a specific approach to precision medicine and hormone therapy to benefit this population.
2. Materials and Methods
2.1 Population Analyzed in the Study
The analyzed indigenous population comprises indigenous peoples from the Amazon, totaling 64 individuals who are part of 12 indigenous tribes from the northern region of Brazil: Asurini from the Xingu and Tocantins localities, Arara, Araweté, Awa-Guajá, Juruna, Kayapó, Xikrin, Karipuna, Munduruku, Phurere, Wajãpi, and Zo’é. These tribes were collectively grouped as the Indigenous (INDG) population for statistical analyses. Genetic data were obtained with the consent of all individuals and their leaders, who completed and signed a specific form for this purpose. The entire study and information gathering were approved by the national ethics committee (CONEP) and the ethics committee of the Center for Tropical Medicine at the Federal University of Para (CAE: 20654313.6.0000.5172).
The frequencies of indigenous peoples were compared with those of other continental populations: Europe (EUR), Africa (AFR), East Asia (EAS), South Asia (SAS), and the Americas (AMR). These datasets were sourced from the 1000 Genomes Database, version 3 (available at: http://www.1000genomes.org; accessed on Feb 2, 2024). The study included 503 individuals from Europe, 661 from Africa, 504 from East Asia, 489 from South Asia, and 347 from the Americas.
2.2. DNA Extraction and Exome Analysis
DNA extraction was conducted following the Phenol-Chloroform method [19] with modifications. The extracted product's quantity was measured using a Nanodrop-8000 spectrophotometer (Thermo Fisher Scientific Inc., Wilmington, DE, USA), and the prospective analysis of the extracted material's quality was performed through 2% agarose gel electrophoresis.
2.3. Gene Selection
Gene selection was performed by consulting the PharmGKB database (https://www.pharmgkb.org/pathway/PA166310681; accessed on March 20, 2024). The five selected genes (SLCO2B1, SULT2A1, CYP3A4, HSD3B1, HSD3B2, and UGT1A4) are related to the signaling pathway of the drug abiraterone, which is the focus of this study in prostate cancer treatment.
2.4. Statistical and Bioinformatic Analysis
The allele frequency of the studied population was obtained by gene counting and compared with other major populations already investigated (EUR, AMR, EAS, SAS, and AFR). Fisher's exact test was used to assess the statistical significance in frequency differentiation between populations. The population variability of polymorphisms was observed through Wright's fixation index (FST). A p-value ≤ 0.05 was considered significant. The entire investigation was conducted using RStudio v.3.5.1. Bioinformatic analyses followed the procedures described by Cohen-Paes et al., 2022 [22].
2.5. Inclusion Criteria
The inclusion criteria for SNPs were as follows: (i) a minimum of 10 coverage reads (fastx_tools v.0.13 http://hannonlab.cshl.edu/fastx_toolkit/, accessed in January 2024); (ii) variant impact: modifier, moderate, or high (SNPeff classification (https://pcingola.github.io/SnpEff/, accessed in March 2024); and (iii) allelic and genomic frequency in worldwide populations (http://www.1000genomes.org, accessed in March 2024).
3. Results
One of the treatment approaches for prostate cancer involves androgen deprivation therapy (ADT), aiming to reduce testosterone production [1]. Among the drugs used to implement this protocol are those containing the substance abiraterone [2]. However, in some cases, despite the application of this protocol, some patients develop castration-resistant prostate cancer (CRPC). Upon investigating possible mechanisms underlying this resistance, a mutation in the HSD3B1 gene was discovered, which has been identified as a potential biomarker for poor prognosis when ADT is implemented [4].
Taking the gene identified as the biomarker HSD3B1 and other genes involved in the drug signaling pathway, including SLCO2B1, SULT2A1, CYP3A4, HSD3B1, HSD3B2, and UGT1A4, as a starting point, we investigated the described variants, their distribution across the global population. Our objective was to identify which of these variants are present in the indigenous population of the Amazon, with the aim of mapping whether variants associated with the HSD3B1 gene and other genes in the pathway are also prevalent in this demographic.
Based on the genes outlined in the signaling pathway illustrated in Figure 01, 46 variants were identified, among which a mutation was found that had only been described in the indigenous population, initially indicated as having low impact. As shown in Figure 02, there is a predominance of variants with modifier impact, comprising 64.3% of the total variants, followed by 21.4% of variants with moderate impact and 14.3% of variants with low impact. Upon analyzing the genes separately, it can be observed that the SULT2A1 gene has the highest percentage of variants with moderate impact, accounting for 80% of all variants described for this gene, followed by the UGT1A4 gene (75.86%), CYP3A4 (66.67%), SLCO2B1 (60%), and HSD3B1 (22.22%).
Out of a total of 56 variants, 48 were selected with modifier and moderate impacts, and one variant not described in the databases used, initially classified as having low impact. In terms of variant allocation by gene, there are three for the CYP3A4 gene, five for the HSD3B1 gene, ten for the SLCO2B1 gene, five for the SULT2A1 gene, and twenty-six for the UGT1A4 gene. The new variant found in the indigenous population, associated with rfseq NM_017460.5, SNV type, in the CDS region, was identified in the CYP3A4 gene.
Table 1 presents the characteristics of these variants, including their reference number, chromosomal region, nucleotide exchange, predicted impact by the SNPeff software, and allelic frequency related to the indigenous group (IND) and the five continental populations present in the 1000 Genomes Program (AFR, AMR, EAS, EUR, and SAS).
Regarding variant types, thirty-six variants are predicted to have a modifier impact, and twelve have a moderate impact. In terms of location, thirty-three were found in intronic regions, thirteen in the CDS region, two in the 5'UTR region, and one in the 3'UTR region. It was observed that variants predicted to have a modifier impact are all located in the 3'UTR, 5'UTR, and intronic regions, while all variants with a moderate impact are located in the CDS region.
Focusing on the HSD3B1 gene, which is identified as a biomarker for increased CPRC recurrence, only one variant was found in the indigenous population.
Data for all identified variants will be available in a supplementary table.
Table 2 presents all variants identified in the indigenous population in a detailed manner, including: gene, refseq, dbsnp, VarType, Impact and detailed region.
Through multidimensional scaling analysis (MDS) (Figure 3), using the FST values (supplementary table) for the 46 variants related to the genes SLCO2B1, SULT2A1, CYP3A4, HSD3B1, HSD3B2, and UGT1A4, the distribution of the five population groups was evidenced (Figure 2). Based on the distribution of populations, the African population is isolated, as expected due to genetic diversity. The American and European populations show significant similarity, followed by populations from South Asia and East Asia with little divergence. An important point to note is the indigenous population, which is also isolated from all other populations, indicating a significant genetic distance from them. The isolation of the studied Amazonian indigenous population contributes to the existence of the variant described only in this population.
4. Discussion
According to the World Health Organization (WHO), in 2020 the global incidence of prostate cancer was 1.2 million individuals, ranking fourth in terms of the number of cases and resulting in 375,000 deaths. Despite its high incidence, the survival rate is 78%, achieved through effective therapeutic approaches including early diagnosis and protocols based on androgen deprivation therapy (ADT) and prostatectomy procedures.
In cases with a poor prognosis, such as castration-resistant prostate cancer (CRPC), individuals may develop resistance to treatment despite adherence to protocols. This resistance often leads to high levels of steroid hormones, such as testosterone, even after ADT and prostatectomy protocols are implemented. Increased steroid hormone expression is a key indicator of CRPC progression and is associated with a shorter survival time.
To understand the mechanisms underlying resistance, genetic studies have focused on genes involved in steroid hormone regulation linked to CRPC incidence. Tissue analysis of these tumors revealed variants in the HSB3D1 gene, which plays a crucial role in steroid hormone production regulation. Due to the significance of this gene and the recurrent mutations observed in individuals with CRPC, it has been identified as a biomarker for poor prognosis in ADT protocols.
In accordance with the guidelines for prostate cancer management provided by the National Comprehensive Cancer Network (NCCN), the primary medication used for treating CRPC is abiraterone acetate. Building upon this protocol, an examination of mutations in this gene and other genes implicated in its signaling pathway was conducted, along with an assessment of the prevalence of these variants in global populations. Additionally, the study delved into the evaluation of these variants within indigenous populations of the Amazon, renowned for their extreme genetic isolation, which often yields divergent treatment responses compared to other populations, including heightened drug toxicity and diminished efficacy.
The distinctive and secluded genetic profile of the indigenous population is highlighted through the multidimensional scaling analysis (MDS) graph (Figure 3), where this population emerges as distinctly separate from others in terms of the identified variants within the studied genes. Notably, the population most genetically akin to the indigenous population is the American population, owing to greater amalgamation with Latino populations.
This genetic distinctiveness underscores the imperative for a highly tailored approach in precision medicine. Analysis of the variants present in the target genes revealed occurrences across all genes studied. For the HSD3B1 gene, three occurrences were identified, two initially classified as having low impact (rs33937873 and rs6203), yet recent studies indicate they serve as risk factors for prostate cancer development and recurrence. Additionally, a variant with moderate impact (rs6205) was identified.
In the SLCO2B1 gene, seven variants were detected, all of which play a pivotal role in ADT protocols by facilitating the transport of abiraterone. Variants within this gene have been associated with cancer progression and poor response to ADT.
Among the five variants described in the SULT2A1 gene, three were found within the indigenous population, all characterized by a modifier impact. Variants within this gene have been linked to an increased risk of prostate cancer development.
The gene harboring the greatest number of described variants within the studied pathway was UGT1A4, with twenty-six variants described, ten of which were identified within the indigenous population. These variants have been associated with the recurrence of localized prostate cancer following radical prostatectomy.
The CYP3A4 gene exhibited the fewest occurrences, with three detected across all studied populations and two within the indigenous population. One occurrence pertained to a new variant lacking description within the examined databases, initially classified as having a low impact. This discovery underscores the genetic distinctiveness of the Amazon indigenous population, as evidenced by the multidimensional scaling analysis (MDS).
Overall, the indigenous population lacks the variant associated with HSD3B1-mediated CRPC, suggesting a potentially more favorable prognosis for prostate cancer treatment. However, variants indicating a risk of developing this type of cancer were uncovered, accentuating the importance of implementing strategies for early diagnosis to enhance treatment efficacy. The identification of a new variant within the CYP3A4 gene underscores the uniqueness of the Amazon indigenous population in genomic studies, rendering this study singular in its field.
These findings and discussions contribute to the development of novel clinical protocols for precision medicine in hormone therapy, aimed at facilitating more efficacious management of this population and mixed populations.5. Conclusions
This section is not mandatory but can be added to the manuscript if the discussion is unusually long or complex.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Shiota M, Narita S, Akamatsu S, et al. Association of Missense Polymorphism in HSD3B1 With Outcomes Among Men With Prostate Cancer Treated With Androgen-Deprivation Therapy or Abiraterone. JAMA Netw Open. 2019;2(2):e190115. doi:10.1001/jamanetworkopen.2019.0115. [CrossRef]
- Alex AB, Pal SK, Agarwal N. CYP17 inhibitors in prostate cancer: latest evidence and clinical potential. Ther Adv Med Oncol. 2016 Jul;8(4):267-75. doi: 10.1177/1758834016642370. Epub 2016 Apr 19. PMID: 27482286; PMCID: PMC4952018. [CrossRef]
- Hearn JWD, Xie W, Nakabayashi M, et al. Association of HSD3B1 Genotype With Response to Androgen-Deprivation Therapy for Biochemical Recurrence After Radiotherapy for Localized Prostate Cancer. JAMA Oncol. 2018;4(4):558–562. doi:10.1001/jamaoncol.2017.3164. [CrossRef]
- Almassi N, Reichard C, Li J, et al. HSD3B1 and Response to a Nonsteroidal CYP17A1 Inhibitor in Castration-Resistant Prostate Cancer. JAMA Oncol. 2018;4(4):554–557. doi:10.1001/jamaoncol.2017.3159. [CrossRef]
- Sai Harisha Rajanala, Anna Plym, Jane B Vaselkiv, Ericka M Ebot, Konstantina Matsoukas, Zhike Lin, Goutam Chakraborty, Sarah C Markt, Kathryn L Penney, Gwo-Shu M Lee, Lorelei A Mucci, Philip W Kantoff, Konrad H Stopsack, SLCO1B3 and SLCO2B1 genotypes, androgen deprivation therapy, and prostate cancer outcomes: a prospective cohort study and meta-analysis, Carcinogenesis, Volume 45, Issue 1-2, January-February 2024, Pages 35–44, https://doi.org/10.1093/carcin/bgad075. [CrossRef]
- Andrew W Hahn et al., Germline variant alleles in rs12422149 of SLCO2B1 and response to abiraterone acetate (AA) in men with metastatic castration-resistant prostate cancer (mCRPC).. JCO 36, 5076-5076(2018). DOI:10.1200/JCO.2018.36.15_suppl.5076. [CrossRef]
- Fujimoto N, Kubo T, Inatomi H, Bui HT, Shiota M, Sho T, Matsumoto T. Polymorphisms of the androgen transporting gene SLCO2B1 may influence the castration resistance of prostate cancer and the racial differences in response to androgen deprivation. Prostate Cancer Prostatic Dis. 2013 Dec;16(4):336-40. doi: 10.1038/pcan.2013.23. Epub 2013 Jul 30. PMID: 23896625. [CrossRef]
- Fujimoto N, Kubo T, Inatomi H, Bui HT, Shiota M, Sho T, Matsumoto T. Polymorphisms of the androgen transporting gene SLCO2B1 may influence the castration resistance of prostate cancer and the racial differences in response to androgen deprivation. Prostate Cancer Prostatic Dis. 2013 Dec;16(4):336-40. doi: 10.1038/pcan.2013.23. Epub 2013 Jul 30. PMID: 23896625. [CrossRef]
- Wilborn, Teresa & Lang, Nicholas & Johnson, Michelle & Meleth, Sreelatha & Falany, Charles. (2006). Association of SULT2A1 allelic variants with plasma adrenal androgens and prostate cancer in African American men. The Journal of steroid biochemistry and molecular biology. 99. 209-14. 10.1016/j.jsbmb.2006.01.006. [CrossRef]
- Kurogi K, Rasool MI, Alherz FA, El Daibani AA, Bairam AF, Abunnaja MS, Yasuda S, Wilson LJ, Hui Y, Liu MC. SULT genetic polymorphisms: physiological, pharmacological and clinical implications. Expert Opin Drug Metab Toxicol. 2021 Jul;17(7):767-784. doi: 10.1080/17425255.2021.1940952. Epub 2021 Jun 30. PMID: 34107842; PMCID: PMC8369464. [CrossRef]
- Teresa W. Wilborn, Nicholas P. Lang, Michelle Smith, Sreelatha Meleth, Charles N. Falany, Association of SULT2A1 allelic variants with plasma adrenal androgens and prostate cancer in African American men, The Journal of Steroid Biochemistry and Molecular Biology, Volume 99, Issues 4–5, 2006, Pages 209-214, ISSN 0960-0760, https://doi.org/10.1016/j.jsbmb.2006.01.006. [CrossRef]
- Laverdière I, Flageole C, Audet-Walsh É, Caron P, Fradet Y, Lacombe L, Lévesque É, Guillemette C. The UGT1 locus is a determinant of prostate cancer recurrence after prostatectomy. Endocr Relat Cancer. 2015 Feb;22(1):77-85. doi: 10.1530/ERC-14-0423. Epub 2014 Dec 1. PMID: 25452636. [CrossRef]
- Amrousy YM, Haffez H, Abdou DM, Atya HB. Role of single nucleotide polymorphisms of the HSD3B1 gene (rs6203 and rs33937873) in the prediction of prostate cancer risk. Mol Med Rep. 2022 Aug;26(2):271. doi: 10.3892/mmr.2022.12787. Epub 2022 Jul 7. PMID: 35795973; PMCID: PMC9309536. [CrossRef]
- National Library of Medicine, National Center for Biotechnology information, acessed 16 March 2024 <https://www.ncbi.nlm.nih.gov/gene/1576>.
- World Health Organization, Health Topics, acessed 12 March 2024 https://www.who.int/news-room/fact-sheets/detail/cancer.
- Juliana Carla Gomes Rodrigues et al., Identification of pharmacogenomic variants associated with oncology treatments in Brazilian Amazonian Amerindians. JCO 40, e15082-e15082(2022). DOI:10.1200/JCO.2022.40.16_suppl.e15082. [CrossRef]
- National Comprehensive Cancer Network, NCCN Guidelines, Prostate Cancer, acessed 18 March 2024 https://www.nccn.org/guidelines/guidelines-detail?category=1&id=1459.
- Rodrigues, J.C.G.; Fernandes, M.R.; Ribeiro-dos-Santos, A.M.; de Araújo, G.S.; de Souza, S.J.; Guerreiro, J.F.; Ribeiro-dos-Santos, Â.; de Assumpção, P.P.; Santos, N.P.C.d.; Santos, S. Pharmacogenomic Profile of Amazonian Amerindians. J. Pers. Med. 2022, 12, 952. https://doi.org/10.3390/jpm12060952. [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring: Harbor, NY, USA, 1989.
- M. Whirl-Carrillo1, R. Huddart1, L. Gong, K. Sangkuhl, C.F. Thorn, R. Whaley and T.E. Klein. "An evidence-based framework for evaluating pharmacogenomics knowledge for personalized medicine" Clinical Pharmacology & Therapeutics (2021) online ahead of print.In Progress: Abiraterone Pathway, Pharmacokinetics. acessed 20 March 2024 https://www.pharmgkb.org/pathway/PA166310681.
- Jason W D Hearn*, Ghada AbuAli*, Chad A Reichard, Chandana A Reddy, Cristina Magi-Galluzzi, Kai-Hsiung Chang, Rachel Carlson,Laureano Rangel, Kevin Reagan, Brian J Davis, R Jeffrey Karnes, Manish Kohli, Donald Tindall, Eric A Klein, Nima Sharif. HSD3B1 and resistance to androgen-deprivation therapy in prostate cancer: a retrospective, multicohort study. www.thelancet.com/oncology Published online August 26, 2016 http://dx.doi.org/10.1016/S1470-2045(16)30227-3. [CrossRef]
- Cohen-Paes, A.d.N.; de Carvalho, D.C.; Pastana, L.F.; Dobbin, E.A.F.; Moreira, F.C.; de Souza, T.P.; Fernandes, M.R.; Leal, D.F.d.V.B.; de Sá, R.B.A.; de Alcântara, A.L.; et al. Characterization of PCLO gene in Amazonian Native American populations.Genes 2022, 13, 499.Author 1, A.; Author 2, B. Title of the chapter. In Book Title, 2nd ed.; Editor 1, A., Editor 2, B., Eds.; Publisher: Publisher Location, Country, 2007; Volume 3, pp. 154–196.
Figure 1.
Genes present in the signaling pathway of the drug abiraterone.

Figure 2.
Distribution of mutations across the genes present in the signaling pathway of the studied drug.
Figure 2.
Distribution of mutations across the genes present in the signaling pathway of the studied drug.
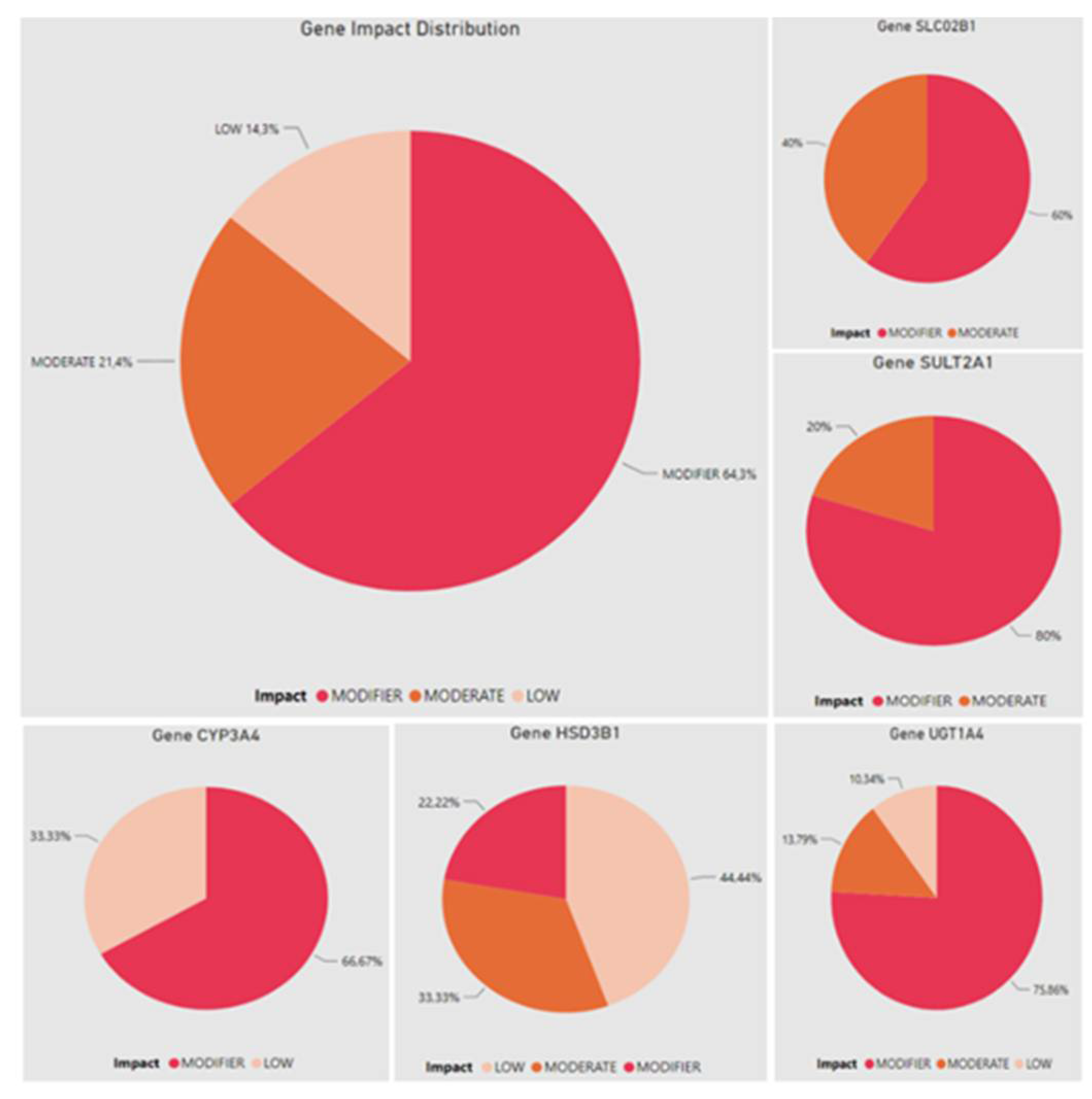
Figure 3.
Multidimensional scaling analysis (MDS).
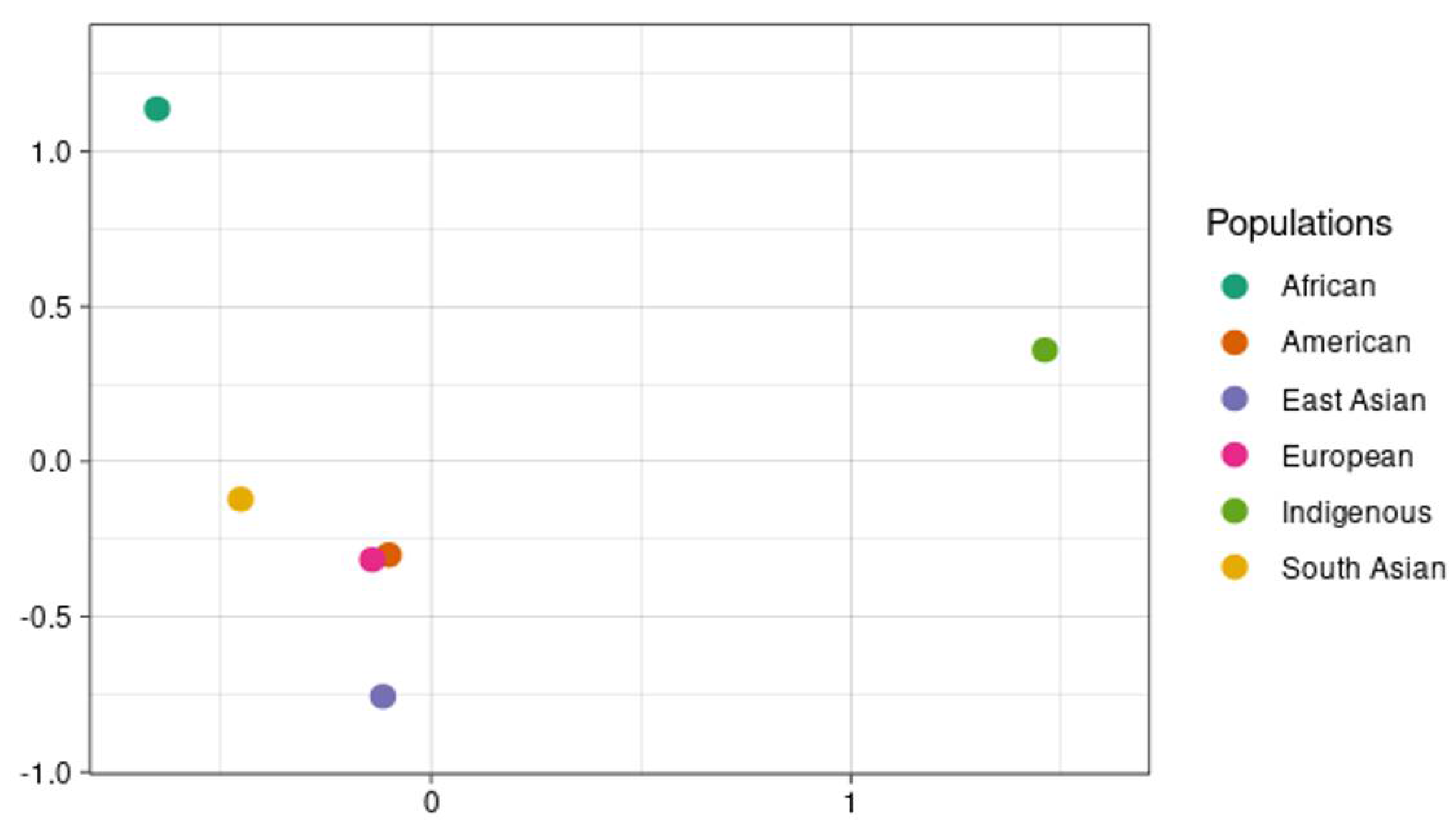

Table 1.
- Variants found and their distribution in the global and indigenous populations.
| Gene | SNP ID | Region | Impact | IND | AFR | AMR | EAS | EUR | SAS |
|---|---|---|---|---|---|---|---|---|---|
| CYP3A4 | . | cds | low | 0,0179 | 0 | 0 | 0 | 0 | 0 |
| CYP3A4 | rs12721620 | Intronic | Modifier | 0,0161 | 0,2671 | 0,0118 | 0 | 0,0006 | 0,008 |
| CYP3A4 | rs2687116 | Intronic | Modifier | 0 | 0,3782 | 0,9165 | 0,9982 | 0,9624 | 0,9629 |
| HSD3B1 | rs1047303 | Cds | Moderate | 0 | 0,8897 | 0,8599 | 0,9376 | 0,6797 | 0,8118 |
| HSD3B1 | rs6201 | Cds | Moderate | 0 | 0,33 | 0,0794 | 0,0412 | 0,003 | 0,0557 |
| HSD3B1 | rs6205 | Cds | Moderate | 0,0391 | 0,39 | 0,0844 | 0,0428 | 0,0034 | 0,0561 |
| HSD3B1 | rs6671149 | Intronic | Modifier | 0 | 0,151 | 0,046 | 0,055 | 0,003 | 0,049 |
| HSD3B1 | rs6673653 | Intronic | Modifier | 0 | 0,1 | 0,04 | 0,054 | 0,002 | 0,05 |
| SLCO2B1 | rs12422149 | Cds | Moderate | 0,6667 | 0,0953 | 0,4573 | 0,3336 | 0,1094 | 0,2231 |
| SLCO2B1 | rs149765874 | Cds | Moderate | 0 | 0 | 0,0001 | 0 | 0,0002 | 0,001 |
| SLCO2B1 | rs2306168 | Cds | Moderate | 0,1349 | 0,3431 | 0,0972 | 0,2308 | 0,0266 | 0,0619 |
| SLCO2B1 | rs60113013 | Cds | Moderate | 0 | 0,0038 | 0,0618 | 0,1107 | 0,0211 | 0,0379 |
| SLCO2B1 | rs12287059 | Intronic | Modifier | 0,0135 | 0,1543 | 0,0074 | 0 | 0,001 | 0,0002 |
| SLCO2B1 | rs1944612 | 5utr | Modifier | 1 | 0,9999 | 0,9994 | 0,8861 | 0,9992 | 0,9961 |
| SLCO2B1 | rs2851069 | 5utr | Modifier | 0,0676 | 0,144 | 0,372 | 0,268 | 0,622 | 0,555 |
| SLCO2B1 | rs7125268 | Intronic | Modifier | 0,1429 | 0,4186 | 0,5222 | 0,3276 | 0,2235 | 0,3283 |
| SLCO2B1 | rs74885054 | Intronic | Modifier | 0,1087 | 0,089 | 0,386 | 0,311 | 0,116 | 0,302 |
| SLCO2B1 | rs995893327 | Intronic | Modifier | 0 | 0 | 0,0002 | 0 | 0 | 0 |
| SULT2A1 | rs11569679 | Cds | Moderate | 0 | 0,1248 | 0,0048 | 0 | 0,0003 | 0,0007 |
| SULT2A1 | rs11569678 | 3utr | Modifier | 0,0161 | 0,049 | 0,049 | 0 | 0 | 0 |
| SULT2A1 | rs2547238 | Intronic | Modifier | 0,0833 | 0,026 | 0,367 | 0,506 | 0,284 | 0,355 |
| SULT2A1 | rs62531056 | Intronic | Modifier | 0,0405 | 0,0007 | 0,0627 | 0,0003 | 0 | 0,0007 |
| SULT2A1 | rs767511533 | Intronic | Modifier | 0 | 0 | 0,0001 | 0 | 0 | 0 |
| UGT1A4 | rs2011425 | cds | Moderate | 0 | 0,0986 | 0,1275 | 0,2065 | 0,0907 | 0,1996 |
| UGT1A4 | rs3892221 | cds | Moderate | 0 | 0,0429 | 0,0011 | 0,005 | 0,0011 | 0,0006 |
| UGT1A4 | rs45540231 | cds | Moderate | 0 | 0,0641 | 0,0032 | 0,0001 | 0,0002 | |
| UGT1A4 | rs6755571 | cds | Moderate | 0 | 0,0162 | 0,0215 | 0,0001 | 0,0553 | 0,0102 |
| UGT1A4 | rs10929301 | Intronic | Modifier | 0 | 0,7048 | 0,4579 | 0,3323 | 0,4511 | 0,6108 |
| UGT1A4 | rs12466997 | Intronic | Modifier | 0,0811 | 0,288 | 0,107 | 0,225 | 0,069 | 0,201 |
| UGT1A4 | rs12471326 | Intronic | Modifier | 0,1111 | 0,0489 | 0,1074 | 0,0038 | 0,0296 | 0,0174 |
| UGT1A4 | rs199892897 | Intronic | Modifier | 0 | 0,086 | 0,004 | 0 | 0 | 0 |
| UGT1A4 | rs2011219 | Intronic | Modifier | 0 | 0,0519 | 0,1106 | 0,1979 | 0,0598 | 0,1759 |
| UGT1A4 | rs2302538 | Intronic | Modifier | 0 | 0,425 | 0,097 | 0,052 | 0,128 | 0,157 |
| UGT1A4 | rs2361501 | Intronic | Modifier | 0 | 0,8134 | 0,4585 | 0,2834 | 0,4469 | 0,6085 |
| UGT1A4 | rs28898618 | Intronic | Modifier | 0,0208 | 0,0172 | 0,0005 | 0 | 0 | 0,0001 |
| UGT1A4 | rs28900402 | Intronic | Modifier | 0 | 0,104 | 0,006 | 0 | 0 | 0,066 |
| UGT1A4 | rs34547608 | Intronic | Modifier | 0 | 0,099 | 0,007 | 0,001 | 0 | 0 |
| UGT1A4 | rs34622615 | Intronic | Modifier | 0 | 0,0519 | 0,0165 | 0,0017 | 0,0307 | 0,0194 |
| UGT1A4 | rs34650714 | Intronic | Modifier | 0 | 0,1218 | 0,0064 | 0,0012 | 0,0008 | 0,0004 |
| UGT1A4 | rs377453564 | Intronic | Modifier | 0 | 0,0001 | 0,0001 | 0,0001 | 0,0002 | 0 |
| UGT1A4 | rs3821242 | Intronic | Modifier | 0,3684 | 0,6401 | 0,4537 | 0,3342 | 0,4453 | 0,61 |
| UGT1A4 | rs4148323 | Intronic | Modifier | 0,0172 | 0,0009 | 0,026 | 0,1524 | 0,0036 | 0,019 |
| UGT1A4 | rs45449995 | Intronic | Modifier | 0,0159 | 0,0485 | 0,0165 | 0,0018 | 0,0306 | 0,0193 |
| UGT1A4 | rs570829500 | Intronic | Modifier | 0 | 0,0003 | 0,0002 | 0,0001 | 0 | 0 |
| UGT1A4 | rs62191918 | Intronic | Modifier | 0,1016 | 0,0946 | 0,1192 | 0,2103 | 0,0615 | 0,1818 |
| UGT1A4 | rs6431625 | Intronic | Modifier | 0,4531 | 0,6698 | 0,3398 | 0,1206 | 0,3867 | 0,4283 |
| UGT1A4 | rs6706232 | Intronic | Modifier | 0,582 | 0,7644 | 0,4583 | 0,3345 | 0,4473 | 0,61 |
| UGT1A4 | rs7574296 | Intronic | Modifier | 0,5192 | 0,7646 | 0,4587 | 0,3322 | 0,4478 | 0,6098 |
| UGT1A4 | rs77053267 | Intronic | Modifier | 0 | 0,0149 | 0,0004 | 0,0001 | 0,0001 |
Table 2.
- Description of variants found in the indigenous population.
| Gene | Refseq | dbSNP | Var Type | Impact | Region Detailed |
|---|---|---|---|---|---|
| CYP3A4 | NM_017460.5 | . | SNV | LOW | SYNON_COD |
| SULT2A1 | NM_003167.3 | rs11569678 | SNV | MODIFIER | UTR_3_PRIME |
| SLCO2B1 | NM_007256.4 | rs12287059 | SNV | MODIFIER | INTRON |
| SLCO2B1 | NM_007256.4 | rs12422149 | SNV | MODERATE | N_SYNON_COD |
| UGT1A4 | NM_007120.2 | rs12466997 | SNV | MODIFIER | INTRON |
| UGT1A4 | NM_007120.2 | rs12471326 | SNV | MODIFIER | INTRON |
| CYP3A4 | NM_017460.5 | rs12721620 | SNV | MODIFIER | INTRON |
| SLCO2B1 | NM_007256.4 | rs1944612 | SNV | MODIFIER | UTR_5_PRIME |
| UGT1A4 | NM_007120.2 | rs2011404 | SNV | LOW | SYNON_COD |
| SLCO2B1 | NM_007256.4 | rs2306168 | SNV | MODERATE | N_SYNON_COD |
| SULT2A1 | NM_003167.3 | rs2547238 | SNV | MODIFIER | INTRON |
| SLCO2B1 | NM_007256.4 | rs2851069 | SNV | MODIFIER | UTR_5_PRIME |
| UGT1A4 | NM_007120.2 | rs28898618 | SNV | MODIFIER | INTRON |
| HSD3B1 | NM_000862.2 | rs33937873 | SNV | LOW | SYNON_COD |
| UGT1A4 | NM_007120.2 | rs3732217 | SNV | LOW | SYNON_COD |
| UGT1A4 | NM_007120.2 | rs3821242 | SNV | MODIFIER | INTRON |
| UGT1A4 | NM_007120.2 | rs4148323 | SNV | MODIFIER | INTRON |
| UGT1A4 | NM_007120.2 | rs45449995 | SNV | MODIFIER | INTRON |
| HSD3B1 | NM_000862.2 | rs6203 | SNV | LOW | SYNON_COD |
| HSD3B1 | NM_000862.2 | rs6205 | SNV | MODERATE | N_SYNON_COD |
| UGT1A4 | NM_007120.2 | rs62191918 | SNV | MODIFIER | INTRON |
| SULT2A1 | NM_003167.3 | rs62531056 | SNV | MODIFIER | INTRON |
| UGT1A4 | NM_007120.2 | rs6431625 | SNV | MODIFIER | INTRON |
| UGT1A4 | NM_007120.2 | rs6706232 | SNV | MODIFIER | INTRON |
| SLCO2B1 | NM_007256.4 | rs7125268 | SNV | MODIFIER | INTRON |
| SLCO2B1 | NM_007256.4 | rs74885054 | INDEL | MODIFIER | INTRON |
| UGT1A4 | NM_007120.2 | rs7574296 | SNV | MODIFIER | INTRON |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



