Submitted:
30 April 2024
Posted:
30 April 2024
You are already at the latest version
Abstract
The early elongation events in transcription or translational control in the N-terminal of genes are assuming critical roles in their heterologous expression. The secretory expression of the alkaline pectin lyase APL in B. subtilis were studied to explore the effects of the N-terminal 5-7 amino acids sequences of different signal peptides on gene transcription and protein secretion in this study. The first 5 amino acid sequences of the N-terminal of the signal peptide LipA were identified to play an important role in promoting the expression of APL for the first time. Meanwhile, it also revealed that the signal peptide LipA could lead to higher secretory expression than other signal peptides not because of its sequence structure associated with secretion is stronger but because the N-terminal sequences showed greater advantages in gene transcription. At this basis, the recombinant strain structed in this work showed a new record for the highest extracellular yields (12 295 U/ml) of APL in B. subtilis which was 1.9 times higher than that expressed in the recombinant Escherichia coli strain we had reported before. The novel theories revealed in this study would assuming significance roles for improving expression of foreign proteins in or out of the cells.
Keywords:
signal peptide
; secretion
; transcription
; extracellular expression
; N-terminal amino acids sequence
; alkaline pectinase
1. Introduction
Bacillus subtilis has many attributes such as food security, easy genetic manipulation, and efficient protein secretion ability, which make it a popular host for recombinant protein production [1,2,3]. It also has long been used as a model organism for molecular research, as well as an industrial workhorse for the production of valuable enzymes [4]. Many strategies including host strain modification, promoter and signal peptide optimization, and fermentation optimization have been used in the construction of cell factory of B. subtilis to improve the production level of individual proteins [1,5,6,7,8]. Even so, as the expression of heterologous protein is a complex process in which cells have their own unknown regulation law in every step of protein synthesis, most of the heterologous proteins are produced at very low levels or even inexpression in the recombinant B. subtilis strains [1,9,10,11].
Generally, except codon optimization, the initial steps to enhance the yield of enzymes are to optimize molecular tools of the expression system, such as promoters and signal peptides (SPs). However, the optimal expression molecular tools for any given secretory protein cannot currently be predicted through experience or in silico so far [1]. N-terminal signal peptides (SPs) are responsible for guiding the target proteins into the typical secretion pathways (Sec and TAT) in B. subtilis [12,13,14]. Many studies have been performed to improve the extracellular expression of the specific protein by screening an efficient signal peptide [15,16,17,18,19]. However, no research had pay attention to another property of signal peptide is that the N-terminal SP sequence is also the initiation of transcription and translation of the target genes so far. While the transcription and translation initiation are the major rate-limiting steps for protein synthesis [20]. Verma et al. had report that the N-terminal amino acids encoded by codons 3 to 5 impact protein yield [20]. Besides, the effect is independent of tRNA abundance, translation initiation efficiency, or overall mRNA structure [20]. According to this theory, we could bold assume that some low efficient secretion SPs for one target protein identified through detection of extracellular protein content may not about its specific secretory structure but limited by its transcription and translation initiation of the N-terminal sequences. Likewise, the optimum SP for a target protein determined by detecting extracellular protein content may concurrently has suitable N-terminal sequences with more efficient transcription and translation initiation.
Based on the hypothesis, after identifying the optimum SP (LipA) of five SPs belonging two different secretion pathways (Sec and TAT) for the extracellular expression of an alkaline pectin lyase APL in B. subtilis, the N-terminal first 5 and 7 amino acids sequences were substituted to reveal their effects on the expression and secretion of APL. It is verified by a series of experimental designs that LipA guiding to relatively higher extracellular expression of APL in B. subtilis not only because its secretion ability but also accounting for its persistently high transcriptional level of the N-terminal amino acids. The highest extracellular APL activity produced by the engineered B. subtilis strain was up to 12,295 U/ml whcih is the highest heterologous expression yield of alkaline pectinase in prokaryotic expression system as far as reported before.
2. Materials and Methods
2.1. Bacterial Strains, Plasmids and Chemicals
E. coli DH5 which eposited in our laboratory was used for construction and storage of the recombinant plasmids. Bacillus subtilis SCK6 was used as the expression host can be obtained from Bacillus Genetic Stock Center (http://www.bgsc.org) with an accession code of 1A976 [21]. The mature peptide encoding sequence of a recombinant alkaline pectin lyase pelNK93I (APL) in our previous report [22] was used as the target expression gene in B. subltilis. The pMA5 was used as a vector for heterologous expression of target genes in B. subtilis SCK6. The restriction endonucleases and DNA polymerase were commercially supplied by Thermo Fisher Scientific Co., Ltd. All other chemicals and solutions with analytical reagent grade purity were purchased through commercial in China.
2.2. Screening of the Signal Peptides for Extracellualr Producing APL
The encoding genes of five signal peptides (Table 1) were inserted into the downstream of the promoter of pMA0911 at the N-terminal of the mature peptide encoding gene of APL, respectively. The recombinant plasmids were constructed using seamless cloning kit (GenScript Co., Ltd., Nanjing, China) according to manufacturer's protocols [4]. After sequencing verification, the corrected recombinant plasmids were transformed into the competent cells of B. subtilis SCK6 according to the method described in our previous work [4]. The positive clones of the recombinant strains were cultured in SR medium (30 g/L tryptone, 50 g/L yeast extract, and 6 g/L K2HPO4) for shake flask cultivation at 37 °C and 220 rpm for 48 h. After cultivation, the culture was centrifugated at 12,000 rpm for 10 min. The supernatant was used for APL enzyme activity determination and protein electrophoresis.
2.3. Construction of the Recombinant Signal Peptides
Based on the original sequences of LipA, YncM, and AmyX, substituted the encoding genes of the N-terminal first 5 and 7 amino acids of YncM and AmyX by the corresponding sequences of LipA, respectively. Meanwhile, the encoding genes of the N-terminal first 5 and 7 amino acids of YncM were also used to substitute the e corresponding sequences of LipA, respectively. All of the original and recombinant sequences (DNA and amino acids) of the signal peptides used in this work were list in Table 1. The recombinant signal peptide encoding genes were constructed by overlap-PCR using the primers in Table S1 and inserted into pMA0911 by Megaprimer PCR to obtain the recombinant plasmids according to our previous report [23]. The recombinant plasmids were transformed into the competent cells of B. subtilis SCK6 and detecting the extracellular expression of APL as described in 2.2. Here, the APL activity and protein expression in intracellular and inclusion bodies were also determined. The cells were treated with ultrasonication at 50 Hz for 10 min in ice water obtain intracellular products. During the shake flask fermentation process, sample testing was performed at regular intervals. The secretion rate was calculated as the proportion of extracellular APL activity to the sum of intracellular and extracellular APL activity. The biomass was displayed by measuring the dry cell weight of 1 ml culture dried for 24 h after centrifugating at 12,000 rpm for 10 min.
2.4. Enzyme Assay and SDS-PAGE
The APL activity was detected as our previous report [22]. One unit was defined as enzyme amount producing 1 μmol unsaturated polygalacturonic acid per minute from 0.2% pectin substrate at pH 9.0 and 60 °C for 10 min. The product yields were detected using an ELISA (BioTek, Epoch2TC) with the extinction coefficient of 4600.0 L/ (mol cm).
The target protein expression in extracellular, intracellular, and inclusion bodies were measured using sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) method as reported previously [4,10]. The electrophoresis was running in 5% stacking gel and 12% separating gel successively. The protein samples were dissolved in 4×loading buffler and loaded 10 μL of the mixed solution in the loading hole. Coomassie Brilliant Blue G250 was used for staining of the proteins.
2.5. Transcription Detection by Quantitative Real-Time PCR (qRT-PCR)
Appropriate amount of culture solution was taken from each test point and centrifuged at 12,000 rpm at 4 ℃ for 2 minutes to collect strain cells. The precipitated strain cells were resuspended using 100 μL TE solution (containing 3 mg/ml of lysozyme) and incubated at room temperature for 10 min. RNA isolation procedure and gene expression quantification were performed as previous reports [24,25]. The total RNA was extracted using a Simply P Total RNA Extraction Kit (BioFlux, China). The quantitation was performed using the Nanodrop 2000 spectrophotometer (Thermo Fisher Scientific). The DNA in the mRNA extract samples was removed by DNase I [25]. The cDNA was synthesized using the PrimeScript RT reagent Kit (TaKaRa, Japan) [25]. Primers used in qRT-PCR analysis to determine the expression levels of DanN (in the genome of B. subtilis SCK6) and APL (the recombinant plasmids) were listed in Table S1. The relative expression level of APL was normalized to the DanN gene.
2.6. Fed-Batch Fermentation of the Engineered Strain to Produce Extracellualr APL
Four hundred microlitres of original seed strains (frozen at -80℃) were inoculated into 400 mL LB medium (Kan) and cultured at 220 rpm at 37 ℃ for 12 h. The cultured seed solution was inoculated into a 5 L fermenter containing 3 L fermentation medium for fermentation. Fermentation conditions were at constant temperature 37 ℃, pH 7.0 (pH adjusted with ammonia and lactic acid), ventilation of 400 L/h, initial speed of 400 rpm (DO 30% was set as the threshold point for speed adjustment, 400-850rpm). Feeding begins when dissolved oxygen begins to rise during fermentation. The dry weight of strain cells and APL activity in fermentation liquid were measured every 6 h. The fermentation ends when both the dry weight and APL activity of the bacteria no longer rise. The fermentation medium contained sucrose (10 g/L), yeast extract powder (30 g/L), NaCl (8 g/L), KH2PO4 (3 g/L), MgCl2 (1 g/L), CaCl2·2H2O (0.3 g/L), MnSO4·2H2O (0.2 g/L), and ZnSO4·7H2O (0.02 g/L), and FeSO4·7H2O (0.02 g/L). The fed-batch feeding solution contained 400 g/L sucrose and 100 g/L yeast extract powder. Biomass was meaning as cell dry weight measured by weighting the deposit bacteria after centrifuging 1 ml of bacterial solution and dried at 80 oC for 24 h.
3. Results
3.1. Effects of Different Signal Peptides on the Secretory Expression of APL in B. subtilis
To screen the optimum signal peptide for secretory expression of the alkaline pectin lyase APL in B. subtilis, four commonly used signal peptides LipA, WapA, AmyX, and YncM were selected to guide the secretion of APL comparing with a pectinase signal peptide PelB. The results showed that the signal peptide YncM belonging to the Sec pathway made a little higher extracellular activity of APL than that of PelB after 48 h fermentation (Figure 1A). The signal peptides LipA and WapA which belonged to the TAT pathway made 4.8 and 3.7 times higher extracellular activity of APL than that of PelB, respectively (Figure 1A). However, the signal peptide AymX made almostly no APL expression in B. subtilis (Figure 1A and B). The wet cell weights of all the test strains showed similar values (Figure 1A). The extracellular target protein yields of the recombinant strains after 48 h fermentation showed the same relative yield trends with their extracellular activities (Figure 1A and B). As shown in Figure 1, LipA was the optimum signal peptides for the secretory expression of APL in B. subtilis in this study.
3.2. Effects of the N-Terminal 5-7 Amino Acids Sequences of the Signal Peptides on the Secretory Expression of APL in B. subtilis
Based on the above results, the N-terminal 5 or 7 amino acids of the optimum signal peptide LipA were used to substitute the N-terminal amino acids of AmyX and YncM, respectively. At the same time, the N-terminal 5 or 7 amino acids of YncM were used to substitute the N-terminal amino acids of LipA. The nucleotide and amino acid sequences of the recombinant signal peptides were showed in Table 1. As the results shown in Figure 2A, when the N-terminal 5 and 7 amino acids of YncM were substituted with those of LipA, the extracellular activities of APL were increased by 3.7 and 1.6 times, respectively. Besides, when the N-terminal 5 and 7 amino acids of AmyX were substituted with those of LipA, both the extracellular and intracellular activities of APL were improved remarkably (Figure 2A). Meanwhile, the results of the SDS-PAGE showed that the recombinant signal peptides AmyX (LipA N5) and AmyX (LipA N7) made higher expression level of APL in extracellular, intracellular and inclusion bodies, comparing with those guided by AmyX, especially in intracellular (Figure 2B–D). However, when the N-terminal 5 and 7 amino acids of LipA were substituted with those of YncM, both the extracellular and intracellular activities of APL were decreased obviously (Figure 2A). The extracellular protein yields of APL were also decreased at a similar scale of the enzyme activity (Figure 2A,B). Moreover, any of the substitute N-terminal amino acids could not influence the secretion ratio on a remarkable scale comparing with their original signal peptdies (Figure 2A).
The recombinant signal peptides YncM (LipA N5) and AmyX (LipA N5) were selected to detect the expression level both in extracellular and intracellular in the fermentation processing during 36 h with LipA as a control. The recombinant signal peptide YncM (LipA N5) made gradually increased extracellular activity of APL, while the intracellular activity remained at a steady low level (Figure 3A). However, using AmyX (LipA N5) as the signal peptide, the intracellular activity of APL was gradually increased with nearly no extracellular activity was detected before 12 h fermenatation. After 15 h fermentation, both extracellular and intracellular activity were gradually increased, while the intracellular activity was kept higher than the extracellular activity during the whole fermentation process (Figure 3A). Besides, the growth state of the three strains did not show significant difference in the whole fermentation process (Figure 3B).
The N-terminal 5 amino acids of LipA were used to directly attach to the N-terminal of the mature peptide of APL. It showed the similar effects both on the extracellular and intracellular activities with those of the recombinant signal peptides AmyX (LipA N5) during 48 h fermentation (Figure 4A and Figure 3A). Similarly, the growths of the strains were not been affected (Figure 4B).
3.3. Effects of the N-Terminal 5 Amino Acids Sequences of the Signal Peptides on the Gene Transcript in B. subtilis
The recombinant signal peptides LipA (YncM N5), YncM (LipA N5), AmyX (LipA N5), and APL (LipA N5) were used to compare with their corresponding original signal peptides LipA, YncM, AmyX, and APL for the gene transcript and the protein expression of APL in B. subtilis. During 48 h fermentation, LipA (YncM N5) led to remarkably decreased expression of APL than LipA and made the similar expression level guided by YncM (Figure 5A). The total expression level of APL guided by YncM (LipA N5) was higher than that guided by its original signal peptide YncM with almost no change on the secretion ratio (Figure 5A). Besides, both the recombinant signal peptides AmyX (LipA N5) and APL (LipA N5) could make remarkably higher expression of APL than their original signal peptides (Figure 5A). The total expression levels of APL guided by AmyX (LipA N5) or APL (LipA N5) were even higher than that guided by LipA (Figure 5A). Meanwhile, the growth of the eight strains at each time of the whole fermentation process showed no obvious difference (Figure 5B). At the basis, the relative transcript level of each strain at different fermentation times was determined by qRT-PCR. At the early expression time of 6 h, all of the recombinant strains showed the highest transcript level during the whole expression process, except the engineered strain containing signal peptide APL (LipA N5) which showed the highest transcript level at 12 h fermentation (Figure 5C). Besides, the engineered strain containing signal peptide AmyX showed remarkably lower transcript level than others at 6 h fermentation and kept relatively lower transcript level during the whole expression process (Figure 5C). The engineered strains containing signal peptide YncM or APL showed the highest transcript level at 6 h fermentation, but after that the transcript level rapidly declined and kept the relative lower transcript level during the whole expression process (Figure 5 C). However, the engineered strains containing the recombinant signal peptide (AmyX (LipA N5) and APL (LipA N5)) and the original signal peptide LipA kept relatively higher transcript level during the whole expression process (Figure 5C). The engineered strains containing the recombinant signal peptide LipA (YncM N5) and YncM (LipA N5) showed rapidly declined in their transcript level after 12 h fermentation (Figure 5C).
3.4. High Yield of APL through Fermentation of the Recombiant Strain in a 5-L Reactor
The engineered strain containing signal peptide LipA which showed the highest extracellular expression of APL was used to perform a scale up fermentation in a 5-L reactor. During 72 h fermentation, the engineered strain produced the highest extracellular activity of APL was up to 12295 U/ml at 60 h fermentation (Figure 6) which was 7.4 times increased from that produced by shake-flask fermentation. The highest wet cell weight (188 g/L) was also obtained at 60 h fermentation (Figure 6). The extracellular APL productivity of the engineered strain guided by signal peptide LipA was 2.05 x 105 U/L/h.
4. Discussion
B. subtilis is generally considered as a good prokaryotic expression host for heterologous proteins based on its advances in extracellular proteins secretion. Over the past decades, in order to make the efficient secretory expression of foreign proteins at a large scale in B. subtilis, more and more signal peptides were explored to improve the extracellular expression of different proteins. In this study, five signal peptides which had been verified to have efficient secretion rate were detected to guide the extracellular expression of APL in B. subtilis. Among them, the signal peptides YncM and PelB are belonging to the typical Sec pathway, while LipA, WapA, and AmyX are belonging to the TAT pathway with characteristic double arginines at the N-terminal of their sequences (Table 1). Based on comparing the extracellular APL activities of the recombinant strains connecting different signal peptides at the N-terminal of mature peptide of APL, the TAT-type signal peptides LipA and WapA showed remarkably higher secretory expression than the Sec-type signal peptides YncM and PelB (Figure 1). While another TAT-type signal peptide AmyX made obviously low expression both in and out cells (Figure 1). Thus, it could not simply estimate any secretion pathway or signal peptide type is more suitable for the secretory expression of APL in B. subtilis. Meanwhile, because the signal peptides were on the N-terminal of the target proteins, the beginning of the gene transcript and protein translation of the target proteins would also be related to the sequence properties of different signal peptides. As is well-known that the initiation of transcript and translation is a major rate-limiting step for protein synthesis [20]. So, the N-terminal signal peptides theoretically may not only determine the secretion rate but also make critical roles in the expression level of the target proteins. However, the effects of the N-terminal signal peptides on the expression level of the heterologous genes have few reported studies so far.
Verma et al. had employed a library of more than 250,000 reporters combined with in vitro and in vivo protein expression assays to assess the influence of early elongation on protein synthesis. They reported that the identity of the amino acids encoded by codons 3 to 5 impact protein yield [20]. Inspired by that study, we selectively switched the N-terminal first 5 amino acid sequences of different signal peptides to assess their influences both on the expression and secretion of the target protein APL (Table 1). Besides, in order to also evaluate the effects of typical double arginines on the protein secretion, the N-terminal first 7 amino acid sequences substitutions were designed simultaneously (Table 1). When the N-terminal 5 and 7 amino acids of LipA were substituted with that of YncM, respectively, the extracellular activities of APL were declined remarkably (Figure 2A). Besides, the recombinant signal peptide LipA (YncM N7) which broken the typical double-arginine sequence made relatively higher extracellular expression than the recombinant signal peptide LipA (YncM N5) which kept the typical double-arginine sequence of the TAT-type signal peptide (Figure 2A, B). It indicated that the specific N-terminal amino acids sequence is the critical factor for the extracellular expression of the target protein rather than the secretion type. Meanwhile, the N-terminal 5 and 7 amino acids of YncM and AmyX were substituted by that of LipA led to increasing of the extracellular activities of APL (Figure 2A). First of all, it could conclude that the N-terminal sequence of LipA has even greater advantages than those of YncM and AmyX in the synthesis of APL in B. subtilis. Besides, the N-terminal 5 and 7 amino acids sequence was not the only factor affecting the expression of the target proteins. When it combined with the remnant sequences of YncM and AmyX led to significant different effect on the gene expression of APL (Figure 2). The recombinant signal peptides AmyX (LipA N5) and AmyX (LipA N7) made very high expression levels of APL which even higher than that guided by LipA (Figure 2). This interesting finding would make a significant strategy for improving different heterologous proteins synthesis in B. subtilis. However, based on current research results in this study, the N-terminal amino acids sequence of the signal peptides did not make significant effect on the extracellular secretion rate of the target proteins (Figure 2A).
As shown in Figure 2A, YncM (LipA N5) and AmyX (LipA N5) made relatively more significant influences in the expression level of APL than YncM (LipA N7) and AmyX (LipA N7), respectively. Thus, the engineered strains containing the recombinant signal peptide YncM (LipA N5) and AmyX (LipA N5) were chose to further evaluate the APL expression difference during the whole fermentation process. As shown in Figure 3, LipA and YncM (LipA N5) mainly produced extracellular APL activities even at the beginning fermentation phase at 6 h, while AmyX (LipA N5) mainly produced intracellular APL activities during the whole fermentation process and the extracellular activity began increasing after 15 h fermentation. Inferred from this result that LipA N5 just has the effect on improving the gene expression and it may destroy the typic double arginines specific sequence thus led to AmyX (LipA N5) lost secretory function. As for a portion of extracellular APL activity detected, it is likely caused by cell lysis in the late fermentation period. To test this hypothesis, LipA N5 was directly attached to the N-terminal of the mature peptide of APL. As shown in Figure 4, LipA N5 indeed remarkably enhanced the expression of APL. As the strain containing the mature peptide of APL has been detected extracellular APL activity after 12 h fermentation, thus as the same the extracellular activity of the strain containing APL (LipA N5) is also due to cell lysis after 12 h fermentation (Figure 4). So, the hypothesis that LipA N5 only having the influence on the expression of target proteins was proved.
To sum up above results, the recombinant signal peptide YncM (LipA N5) made increased expression with no effect on its secretion, while the recombinant signal peptides AmyX (LipA N5) made remarkably increased expression with secretion function lost (Figure 5A). Through detecting the relative transcriptional level of each engineered strain, the recombinant proteins with LipA N5 sequence at their N-terminal showed relatively higher transcriptional level than their control strains (Figure 5C). Moreover, the high expression level strains also showed consistently high transcription levels during the fermentation process (Figure 5C). Thus, the specific N-terminal sequence LipA N5 was identified for the potential of improving the expression of heterologous proteins in B. subtilis. Meanwhile, it is also suggested that researchers should also consider the effect of N-terminal sequence on expression when selecting appropriate signal peptides in B. subtilis.
LipA was identified the optimum signal peptide for the heterologous extracellular expression of APL with both high expression level based on its N-terminal specific sequence and efficient secretion rate though TAT pathway in B. subtilis. Fed-batch culture in a 5-L reactor was performed to test the industrial fermentation capacity of the engineered strain. The highest extracellular APL activity (12,295 U/ml) obtained at 60 h fermentation which further exceeded the total soluble APL activity (10,181 U/mL) expressed in the recombinant Escherichia coli strain we had structed [22]. So far, it is the highest heterologous expression yield of alkaline pectinase in prokaryotic expression system as far as reported before [22].
5. Conclusions
Collectively, our data underscore the critical role of the first N-terminal 5-7 amino acids of different signal peptides in the expression of heterologous APL in B. subtilis. Revealed for the first time that the first N-terminal 5 amino acids of signal peptide LipA could remarkably improving the expression of the target protein APL due to enhanced transcriptional level in B. subtilis. It will open a new perspective of signal peptide screening for heterologous proteins expression. Moreover, this study set a new high yield record for the extracellular production of APL in B. subtilis (12,295 U/ml).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Primers used in this study.
Author Contributions
Conceptualization, H.Z. and M.Z.; methodology, J.Z.; software, M.Z. and J.T.; validation, X.Z., J.T., and X.F.; formal analysis, J.Z.; investigation, H.Z.; resources, J.Z., and J.X.; data curation, M.Z.; writing—original draft preparation, M.Z.; writing—review and editing, H.Z.; visualization, J.Z.; supervision, H.S., and H.Z.; project administration, H.S.; funding acquisition, H.Z., M.Z., Y.Z., and W.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the State Key Research and Development Program of China, grant number 2021YFC2100405; Heilongjiang Province Fundamental Research Funds, grant number 145209322; Science and Technology Partnership Program, Ministry of Science and Technology of China; grant number KY202001017; Tianjin Synthetic Biotechnology Innovation Capacity Improvement Project, grant number TSBICIP-IJCP-001-02, TSBICIP-PTJJ-007-13, and TSBICIP-KJGG-009-0202; and Tianjin Municipal Education Commission Scientific Research Project, grant number 2022KJ024.
Data Availability Statement
All data generated or analyzed during this study are included in this published article and its supplementary information files. All further data will be provided by the corresponding author at any time upon request.
Acknowledgments
The authors thank Dr. Zhanping Zhou and Jiangning Song for their kind donation of the encoding sequence of pelNK93I.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhang, K.; Su, L.; Wu, J. Enhancing extracellular pullulanase production in Bacillus subtilis through dltB disruption and signal peptide optimization. Appl Biochem Biotechnol 2022, 194, 1206-1220. [CrossRef]
- Sun, X.; Yang, J.; Fu, X.; Zhao, X.; Zhen, J.; Song, H.; Xu, J.; Zheng, H.; Bai, W. Trehalose Production Using Three Extracellular Enzymes Produced via One-Step Fermentation of an Engineered Bacillus subtilis Strain. Bioengineering (Basel) 2023, 10. [CrossRef]
- Xu, Y.; Xuan, X.; Gao, R.; Xie, G. Increased Expression Levels of Thermophilic Serine Protease TTHA0724 through Signal Peptide Screening in Bacillus subtilis and Applications of the Enzyme. Int J Mol Sci 2023, 24. [CrossRef]
- Yang, Y.; Fu, X.; Zhao, X.; Xu, J.; Liu, Y.; Zheng, H.; Bai, W.; Song, H. Overexpression of a thermostable α-amylase through genome integration in Bacillus subtilis. Fermentation 2023, 9, 139. [CrossRef]
- Yu, W.; Jin, K.; Wu, Y.; Zhang, Q.; Liu, Y.; Li, J.; Du, G.; Chen, J.; Lv, X.; Ledesma-Amaro, R.; et al. A pathway independent multi-modular ordered control system based on thermosensors and CRISPRi improves bioproduction in Bacillus subtilis. Nucleic Acids Res 2022, 50, 6587-6600. [CrossRef]
- Liu, Y.; Cheng, H.; Li, H.; Zhang, Y.; Wang, M. A Programmable CRISPR/Cas9 Toolkit Improves Lycopene Production in Bacillus subtilis. Appl Environ Microbiol 2023, 89, e0023023. [CrossRef]
- Xu, L.; Zhang, Y.; Dong, Y.; Qin, G.; Zhao, X.; Shen, Y. Enhanced extracellular beta-mannanase production by overexpressing PrsA lipoprotein in Bacillus subtilis and optimizing culture conditions. J Basic Microbiol 2022, 62, 815-823. [CrossRef]
- Lu, J.; Zhao, Y.; Cheng, Y.; Hu, R.; Fang, Y.; Lyu, M.; Wang, S.; Lu, Z. Optimal Secretory Expression of Acetaldehyde Dehydrogenase from Issatchenkia terricola in Bacillus subtilis through a Combined Strategy. Molecules 2022, 27. [CrossRef]
- Harwood, C.R.; Cranenburgh, R. Bacillus protein secretion: an unfolding story. Trends Microbiol 2008, 16, 73-79. [CrossRef]
- Liu, P.; Guo, J.; Miao, L.; Liu, H. Enhancing the secretion of a feruloyl esterase in Bacillus subtilis by signal peptide screening and rational design. Protein Expr Purif 2022, 200, 106165. [CrossRef]
- Gustafsson, C.; Minshull, J.; Govindarajan, S.; Ness, J.; Villalobos, A.; Welch, M. Engineering genes for predictable protein expression. Protein Expr Purif 2012, 83, 37-46. [CrossRef]
- Grasso, S.; Dabene, V.; Hendriks, M.; Zwartjens, P.; Pellaux, R.; Held, M.; Panke, S.; van Dijl, J.M.; Meyer, A.; van Rij, T. Signal Peptide Efficiency: From High-Throughput Data to Prediction and Explanation. ACS Synth Biol 2023, 12, 390-404. [CrossRef]
- Huang, X.; Cao, L.; Qin, Z.; Li, S.; Kong, W.; Liu, Y. Tat-Independent Secretion of Polyethylene Terephthalate Hydrolase PETase in Bacillus subtilis 168 Mediated by Its Native Signal Peptide. J Agric Food Chem 2018, 66, 13217-13227. [CrossRef]
- Liu, R.; Zuo, Z.; Xu, Y.; Song, C.; Jiang, H.; Qiao, C.; Xu, P.; Zhou, Q.; Yang, C. Twin-arginine signal peptide of Bacillus subtilis YwbN can direct Tat-dependent secretion of methyl parathion hydrolase. J Agric Food Chem 2014, 62, 2913-2918. [CrossRef]
- Su, L.; Li, Y.; Wu, J. Efficient secretory expression of Bacillus stearothermophilus alpha/beta-cyclodextrin glycosyltransferase in Bacillus subtilis. J Biotechnol 2021, 331, 74-82. [CrossRef]
- Wang, S.; Yang, Z.; Li, Z.; Tian, Y. Heterologous Expression of Recombinant Transglutaminase in Bacillus subtilis SCK6 with Optimized Signal Peptide and Codon, and Its Impact on Gelatin Properties. J Microbiol Biotechnol 2020, 30, 1082-1091. [CrossRef]
- Wang, N.; Guan, F.; Lv, X.; Han, D.; Zhang, Y.; Wu, N.; Xia, X.; Tian, J. Enhancing secretion of polyethylene terephthalate hydrolase PETase in Bacillus subtilis WB600 mediated by the SP(amy) signal peptide. Lett Appl Microbiol 2020, 71, 235-241. [CrossRef]
- Kang, X.M.; Cai, X.; Huang, Z.H.; Liu, Z.Q.; Zheng, Y.G. Construction of a highly active secretory expression system in Bacillus subtilis of a recombinant amidase by promoter and signal peptide engineering. Int J Biol Macromol 2020, 143, 833-841. [CrossRef]
- Heinrich, J.; Drewniok, C.; Neugebauer, E.; Kellner, H.; Wiegert, T. The YoaW signal peptide directs efficient secretion of different heterologous proteins fused to a StrepII-SUMO tag in Bacillus subtilis. Microb Cell Fact 2019, 18, 31. [CrossRef]
- Verma, M.; Choi, J.; Cottrell, K.A.; Lavagnino, Z.; Thomas, E.N.; Pavlovic-Djuranovic, S.; Szczesny, P.; Piston, D.W.; Zaher, H.S.; Puglisi, J.D.; et al. A short translational ramp determines the efficiency of protein synthesis. Nat Commun 2019, 10, 5774. [CrossRef]
- Zhang, X.Z.; You, C.; Zhang, Y.H. Transformation of Bacillus subtilis. Methods Mol Biol 2014, 1151, 95-101. [CrossRef]
- Zhen, J.; Tan, M.; Fu, X.; Shu, W.; Zhao, X.; Yang, S.; Xu, J.; Ma, Y.; Zheng, H.; Song, H. High-level extracellular production of an alkaline pectate lyase in E. coli BL21 (DE3) and its application in bioscouring of cotton fabric. 3 Biotech 2020, 10, 49. [CrossRef]
- Zhao, X.; Zheng, H.; Zhen, J.; Shu, W.; Yang, S.; Xu, J.; Song, H.; Ma, Y. Multiplex genetic engineering improves endogenous expression of mesophilic alpha-amylase gene in a wild strain Bacillus amyloliquefaciens 205. Int J Biol Macromol 2020, 165, 609-618. [CrossRef]
- Zhang, J.; Xu, X.; Li, X.; Chen, X.; Zhou, C.; Liu, Y.; Li, Y.; Lu, F. Reducing the cell lysis to enhance yield of acid-stable alpha amylase by deletion of multiple peptidoglycan hydrolase-related genes in Bacillus amyloliquefaciens. Int J Biol Macromol 2021, 167, 777-786. [CrossRef]
- Niu, T.; Lv, X.; Liu, Y.; Li, J.; Du, G.; Ledesma-Amaro, R.; Liu, L. The elucidation of phosphosugar stress response in Bacillus subtilis guides strain engineering for high N-acetylglucosamine production. Biotechnol Bioeng 2020. [CrossRef]
Figure 1.
Effects of different signal peptides on the extracellular expression of APL. (A) APL activities in the extracellular and intracellular, respectively. All values are expressed as mean ± SD (n = 3); (B) Extracellular proteins detected by SDS-PAGE.
Figure 1.
Effects of different signal peptides on the extracellular expression of APL. (A) APL activities in the extracellular and intracellular, respectively. All values are expressed as mean ± SD (n = 3); (B) Extracellular proteins detected by SDS-PAGE.
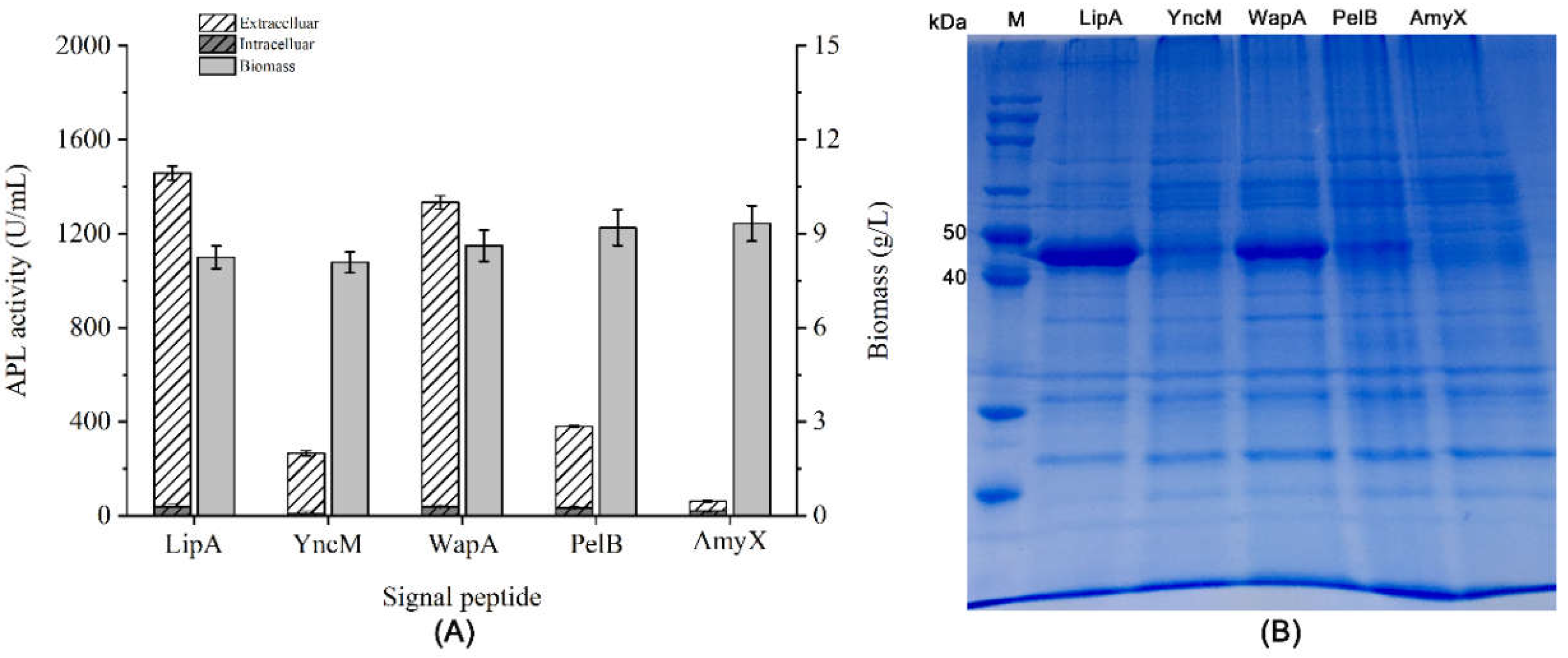
Figure 2.
Effects of the recombinant signal peptides on the expression and secretion of APL. (A) Activities and secretion rates of APL guided by different signal peptides. All values are expressed as mean ± SD (n = 3); (B) Extracellular proteins detected by SDS-PAGE; (C) Intracellular proteins detected by SDS-PAGE; (D) Inclusion bodies detected by SDS-PAGE. M: the protein molecular weight markers, the bands from top to bottom represent 180kDa, 130kDa, 95kDa, 70kDa, 55kDa, 43kDa, 33kDa, 25kDa, 17kDa, and 10kDa, respectively; Lane 1-9 represented APL expression strains harbouring different signal peptides. Lane 1: LipA; Lane 2: LipA (YncM N5); Lane 3: LipA (YncM N7); Lane 4: YncM; Lane 5: YncM (LipA N5); Lane 6: YncM (LipA N7); Lane 7: AmyX; Lane 8: AmyX (LipA N5); Lane 9: AmyX (LipA N7).
Figure 2.
Effects of the recombinant signal peptides on the expression and secretion of APL. (A) Activities and secretion rates of APL guided by different signal peptides. All values are expressed as mean ± SD (n = 3); (B) Extracellular proteins detected by SDS-PAGE; (C) Intracellular proteins detected by SDS-PAGE; (D) Inclusion bodies detected by SDS-PAGE. M: the protein molecular weight markers, the bands from top to bottom represent 180kDa, 130kDa, 95kDa, 70kDa, 55kDa, 43kDa, 33kDa, 25kDa, 17kDa, and 10kDa, respectively; Lane 1-9 represented APL expression strains harbouring different signal peptides. Lane 1: LipA; Lane 2: LipA (YncM N5); Lane 3: LipA (YncM N7); Lane 4: YncM; Lane 5: YncM (LipA N5); Lane 6: YncM (LipA N7); Lane 7: AmyX; Lane 8: AmyX (LipA N5); Lane 9: AmyX (LipA N7).
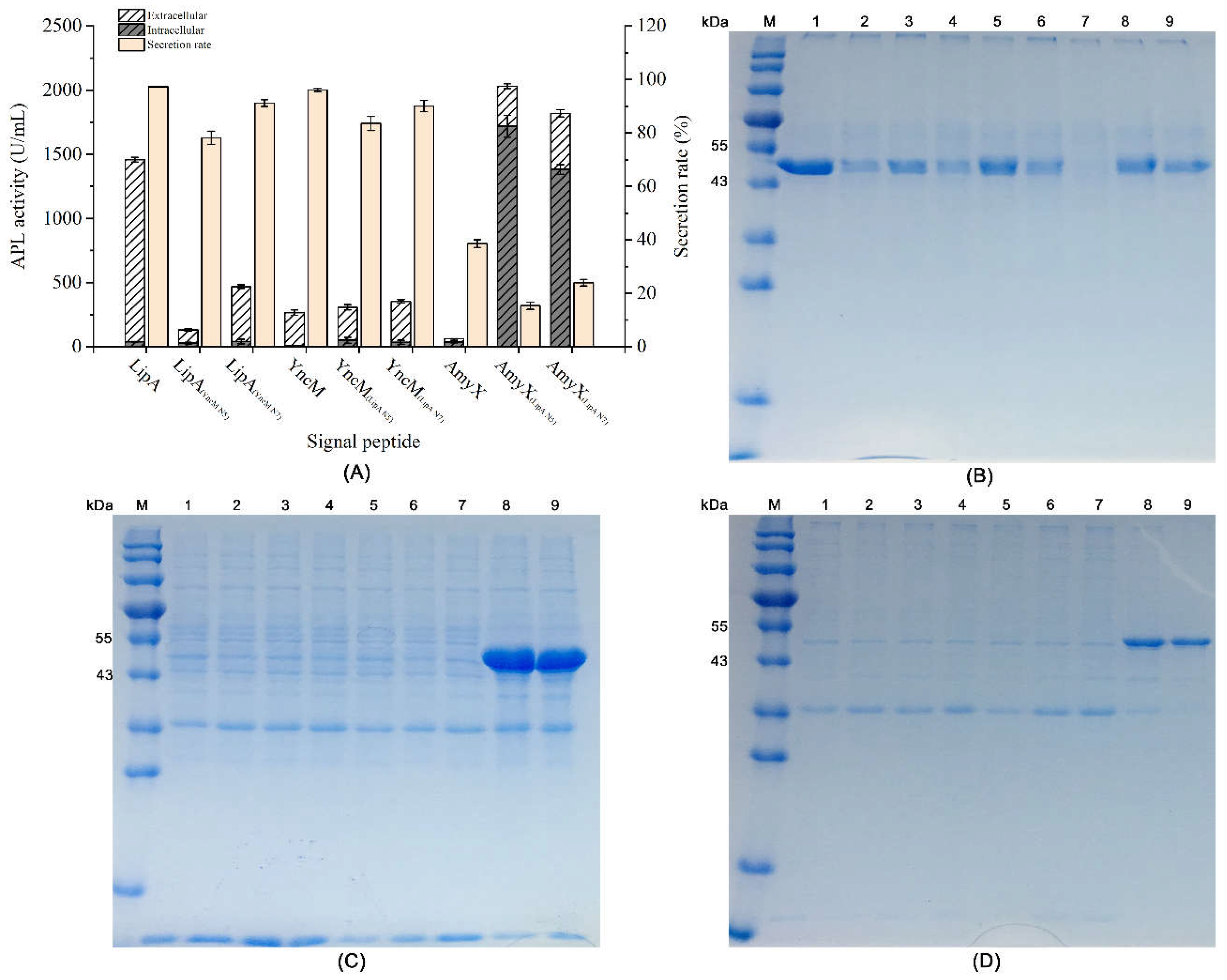
Figure 3.
The APL activity profiles (A) and strain growth profiles (B) of the recombinant strains containing plasmids pMA0911-LipA-APL, pMA0911-YncM (LipA N5)-APL, and pMA0911-AmyX (LipA N5)-APL, respectively. All values are expressed as mean ± SD (n = 3).
Figure 3.
The APL activity profiles (A) and strain growth profiles (B) of the recombinant strains containing plasmids pMA0911-LipA-APL, pMA0911-YncM (LipA N5)-APL, and pMA0911-AmyX (LipA N5)-APL, respectively. All values are expressed as mean ± SD (n = 3).
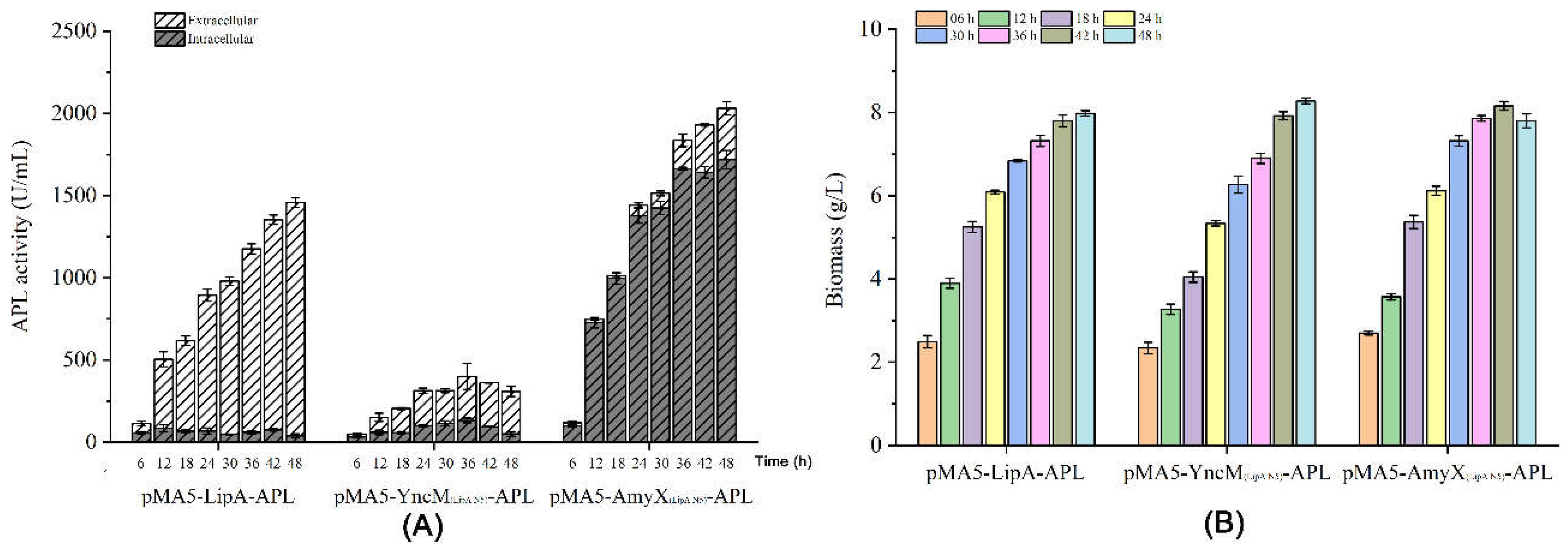
Figure 4.
The APL activity profiles (A) and strain growth profiles (B) of the recombinant strains containing plasmids pMA0911-APL (without signal peptide) and pMA0911-LipA N5-APL (with only five amino acids in the N-terminal), respectively. All values are expressed as mean ± SD (n = 3).
Figure 4.
The APL activity profiles (A) and strain growth profiles (B) of the recombinant strains containing plasmids pMA0911-APL (without signal peptide) and pMA0911-LipA N5-APL (with only five amino acids in the N-terminal), respectively. All values are expressed as mean ± SD (n = 3).
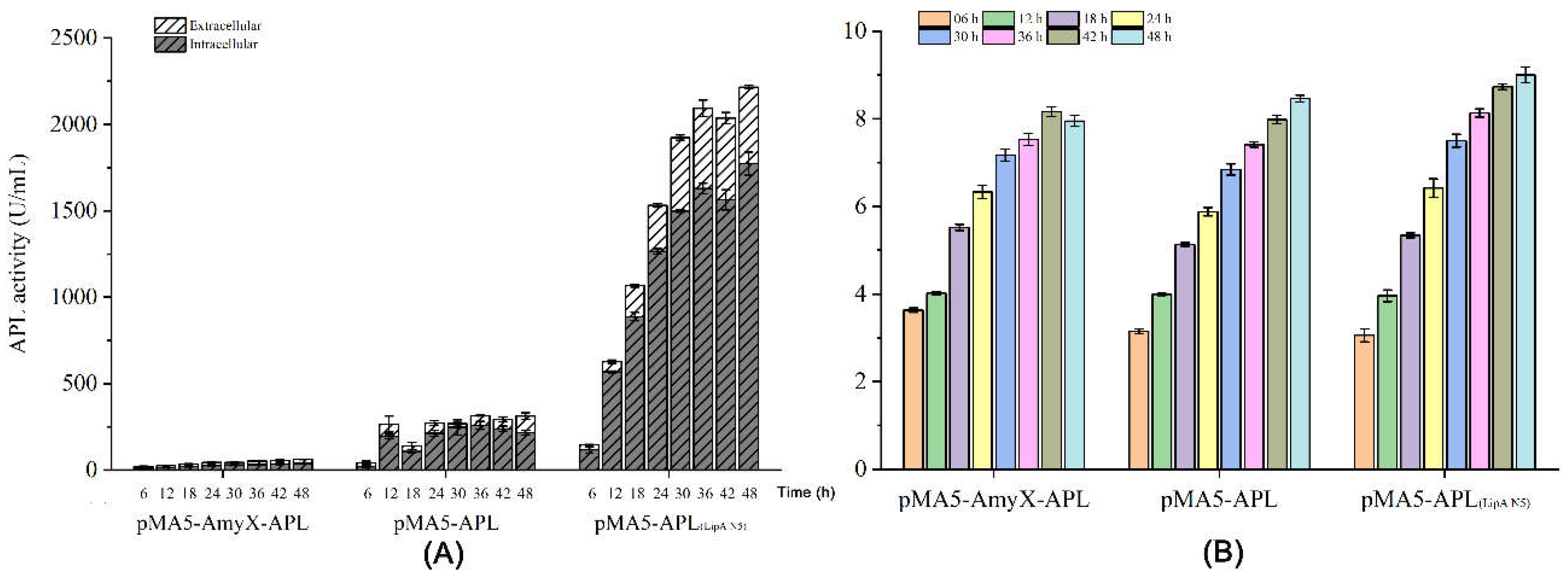
Figure 5.
The APL activity profiles (A), strain growth profiles (B), and transcriptional level profiles (C) of all the recombinant strains constructed in this work. All values are expressed as mean ± SD (n = 3).
Figure 5.
The APL activity profiles (A), strain growth profiles (B), and transcriptional level profiles (C) of all the recombinant strains constructed in this work. All values are expressed as mean ± SD (n = 3).
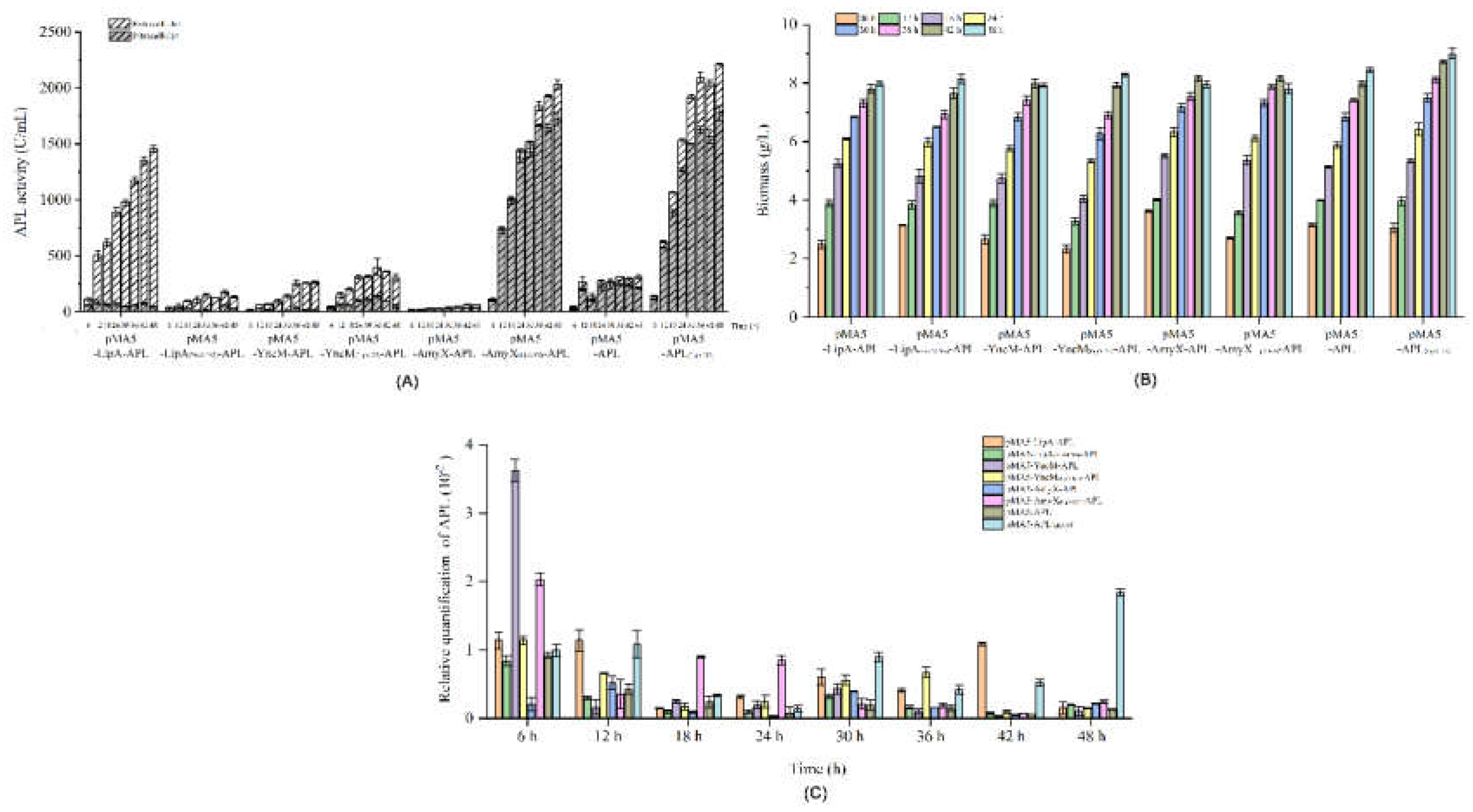
Figure 6.
Scale up fermentation in a 5 L reactor of the engineered strain B. subtilis SCK6 (pMA0911-LipA-APL). All values are expressed as mean ± SD (n = 3).
Figure 6.
Scale up fermentation in a 5 L reactor of the engineered strain B. subtilis SCK6 (pMA0911-LipA-APL). All values are expressed as mean ± SD (n = 3).
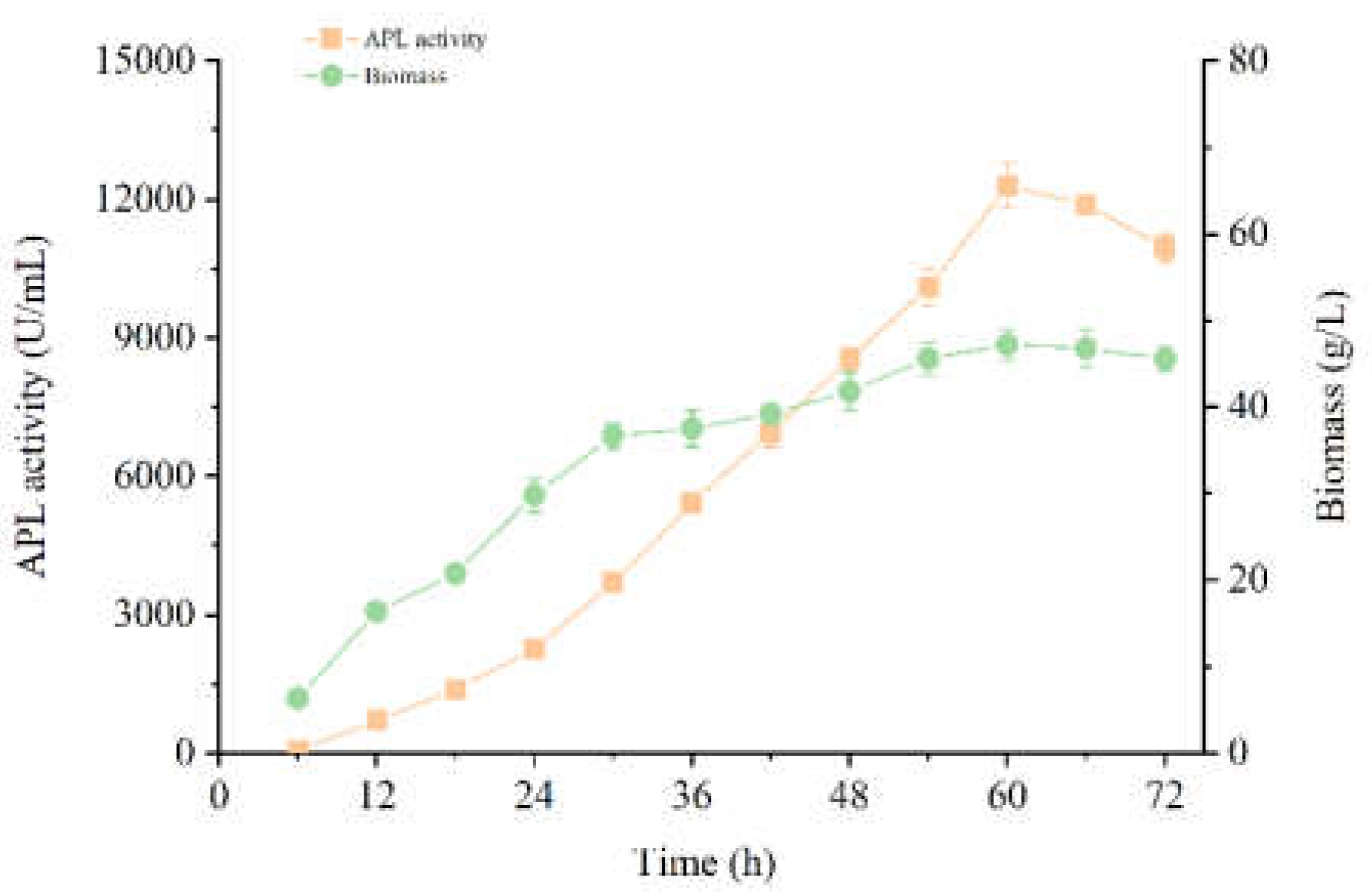

Table 1.
Nucleotide and amino acids sequences of the signal peptides used in this work.
| Signal Peptide | Nucleotide Sequence | Amino Acid Sequence |
|---|---|---|
| LipA | ATGAAATTTGTGAAACGCAGAATTATTGCGCTGGTGACAATTCTGATGCTGAGCGTGACAAGCCTGTTTGCGCTGCAACCGAGCGCGAAAGCG | MKFVKRRIIALVTILMLSVTSLFALQPSAKA |
| YncM | ATGGCTAAACCGCTGTCAAAAGGCGGCATTCTGGTTAAAAAAGTTCTGATTGCAGGCGCAGTTGGCACAGCAGTCCTGTTTGGCACGCTGAGTAGCGGCATTCCGGGACTGCCAGCAGCTGATGCG | MAKPLSKGGILVKKVLIAGAVGTAVLFGTLSSGIPGLPAADA |
| WapA | ATGAAAAAACGCAAACGCAGAAATTTTAAACGCTTTATTGCGGCGTTTCTGGTTCTGGCGCTGATGATTAGCCTGGTTCCGGCGGATGTGCTGGCG | MKKRKRRNFKRFIAAFLVLALMISLVPADVLA |
| PelB | ATGAAATACCTGCTGCCGACCGCTGCTGCTGGTCTGCTGCTCCTCGCTGCCCAGCCGGCGATGGCC | MKYLLPTAAAGLLLLAAQPAMA |
| AmyX | ATGGTCAGCATCCGCCGCAGCTTCGAAGCGTATGTCGATGACATGAATATCATTACTGTTCTGATTCCTGCTGAACAAAAGGAAATCATGACACCGCCG | MVSIRRSFEAYVDDMNIITVLIPAEQKEIMTPP |
| LipA(YncM N5) | ATGGCTAAACCGCTGCGCAGAATTATTGCGCTGGTGACAATTCTGATGCTGAGCGTGACAAGCCTGTTTGCGCTGCAACCGAGCGCGAAAGCG | MAKPLRRIIALVTILMLSVTSLFALQPSAKA |
| LipA(YncMN7) | ATGGCTAAACCGCTGTCAAAAATTATTGCGCTGGTGACAATTCTGATGCTGAGCGTGACAAGCCTGTTTGCGCTGCAACCGAGCGCGAAAGCG | MAKPLSKIIALVTILMLSVTSLFALQPSAKA |
| YncM(LipAN5) | ATGAAATTTGTGAAATCAAAAGGCGGCATTCTGGTTAAAAAAGTTCTGATTGCAGGCGCAGTTGGCACAGCAGTCCTGTTTGGCACGCTGAGTAGCGGCATTCCGGGACTGCCAGCAGCTGATGCG | MKFVKSKGGILVKKVLIAGAVGTAVLFGTLSSGIPGLPAADA |
| YncM(LipAN7) | ATGAAATTTGTGAAACGCAGAGGCGGCATTCTGGTTAAAAAAGTTCTGATTGCAGGCGCAGTTGGCACAGCAGTCCTGTTTGGCACGCTGAGTAGCGGCATTCCGGGACTGCCAGCAGCTGATGCG | MKFVKRRGGILVKKVLIAGAVGTAVLFGTLSSGIPGLPAADA |
| AmyX(LipA N5) | ATGAAATTTGTGAAACGCAGCTTCGAAGCGTATGTCGATGACATGAATATCATTACTGTTCTGATTCCTGCTGAACAAAAGGAAATCATGACACCGCCG | MKFVKRSFEAYVDDMNIITVLIPAEQKEIMTPP |
| AmyX(LipA N7) | ATGAAATTTGTGAAACGCAGATTCGAAGCGTATGTCGATGACATGAATATCATTACTGTTCTGATTCCTGCTGAACAAAAGGAAATCATGACACCGCCG | MKFVKRRFEAYVDDMNIITVLIPAEQKEIMTPP |
Note: The bold font represents the substitute sequences.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



