Submitted:
23 April 2024
Posted:
26 April 2024
You are already at the latest version
Abstract
Resistance against amitraz in varroa mite populations has become a subject of interest in recent years due to increasing reports of reduced field efficacy of amitraz treatments, especially from some beekeepers in France and the United States. The loss of amitraz as a reliable tool to effectively reduce varroa mite infestation in the field could severely worsen the position of beekeepers in the fight to keep varroa infestation rates in their colonies at low levels. In this publication, we present data from French apiaries, collected in the years 2020 and 2021. These data include field efficacy of an authorized amitraz-based varroa treatment (Apivar®) and results of laboratory sensitivity assays of varroa mites exposed towards the reference LC90 concentration of amitraz. In addition, a total of 240 varroa mites from Eastern, Central, and Southern regions in France that were previously classified as either “sensitive” or “resistant” towards amitraz in a laboratory sensitivity assay were genotyped. The genetic analyses of mite samples are focused on the β−adrenergic-like octopamine receptor, which is considered as the main target site for amitraz in varroa mites. Special attention was given to a single nucleotide polymorphism (SNP) at position 260 of the ORβ-2R-L gene that had previously been described by Hernández Rodríguez et al. (2021) as associated to the resistance to amitraz in French varroa mites. Our findings confirm that amitraz resistance occurs in patches or “islands of resistance” with a less severe reduction in treatment efficacy compared to pyrethroid resistance or coumaphos resistance in varroa mites. The results of our genetic analyses of varroa mites call into question the hypothesis of the SNP at position 260 of the ORβ-2R-L gene being directly responsible for amitraz resistance development.
Keywords:
varroa mite
; resistance
; variation
; single nucleotide polymorphism
; honey bee
; amitraz
1. Introduction
The efficient and sustainable control of varroa mites (Varroa destructor) in honey bee colonies (Apis mellifera) is characterized by ongoing challenges for beekeepers on a global scale [1]. Although several varroa treatments with different active ingredients have been available for several decades, high colony losses are periodically reported by beekeepers from different global regions [2,3,4]. Multiple factors such as environmental change (climate and human land use changes) [5], inter-seasonal variations in weather, available nectar and pollen sources, as well as honey bee and varroa population growth [6,7], and the development of acaricide resistance in varroa mites [8], can contribute to high colony losses due to varroosis. In addition, the complete lack of control options for bee viruses, which are transmitted by varroa mites, allows for ever-increasing viral loads in honey bee colonies that build up continuously over time [9].
In beekeeping, one of the most prominent examples of acaricide resistance development in varroa mites is pyrethroid resistance [10]. These molecules have been primarily formulated and applied in honey bee colonies as long-term treatments (over a period of several weeks) in the form of plastic strips. In the Lombardy region in Italy, tau-fluvalinate resistance has been reported at the beginning of the 1990s, only a few years after the authorization of a tau-fluvalinate-based varroa treatment in 1989 [11]. In the United States, tau-fluvalinate has been authorized since 1990 [12]. The first signs of resistance in varroa mite populations were detected in the 1990s, culminating in a severe reduction of treatment efficacy in the field towards the end of the decade [13]. Resistance against flumethrin, another pyrethroid that has been authorized for varroa control in countries all over the world, has been detected repeatedly over the last few decades – sometimes as cross resistance against both, tau-fluvalinate and flumethrin [14,15,16]. In these and other instances of varroa mite resistance against pyrethroids, resistance spread quickly once it occurred and led to severe reductions in treatment efficacy in the affected apiaries [10,11].
The underlying genetic mechanisms leading to pyrethroid resistance in varroa mites have been investigated extensively over the years [10,17,18,19]. Mite populations from different countries were genotyped to identify polymorphisms that could provide more information on possible resistance mechanisms on the phenotypical level. In several research studies, single nucleotide polymorphisms (SNPs) were identified as the source of tau-fluvalinate and/or flumethrin resistance in varroa mites. Although the location of SNPs in the mites’ genome varied geographically, the main target mechanism of the detected variations appeared to be the same: an alteration of the voltage-gated sodium channels, the main target site for pyrethroids [10].
For many years, amitraz (a formamidine) withstood any signs of resistance development in most regions where it was used for varroa control. At present, amitraz has been used effectively and for a long period of time as a varroa treatment in several countries [10,20,21]. In Poland, amitraz use in beekeeping dates to the beginning of the 1980s. Despite a now 40+ year treatment history of amitraz in the country’s bee hives, it is still applied today, resulting in successful treatment outcome [20]. Similarly, in Spain, amitraz-based varroa control products date back to the late 1990s. Until this day, the molecule remains an effective component of varroa control in Spanish beekeeping operations [21].
In recent years, data from the United States and France have raised concerns that amitraz resistance among varroa mite populations in these countries appears to be developing in more and more apiaries [22,23]. In the United States, many amitraz resistance reports are based on field bioassay data (also referred to as “Pettis test” in beekeeping, after Dr. Jeff Pettis) [23,24,25]. In France, laboratory sensitivity assays, detecting the susceptibility of varroa mite populations towards amitraz, have been utilized as the main tool to identify amitraz resistance in French varroa mite populations [22,24,26]. Interestingly, the occurrence of amitraz resistance in apiaries across both countries has been characterized much differently than pyrethroid resistance. Instead of a resistant varroa mite phenotype that spreads quickly within and across apiaries and leads to severe reductions in field efficacy, amitraz resistance appears to manifest in “patches” or “islands of resistance” with – at least initially – a less severe reduction in field efficacy within affected apiaries [23].
Concerning the genetic basis of varroa mite resistance towards amitraz, similarly as for pyrethroid resistance, single nucleotide polymorphisms have been proposed as potential manifestations of amitraz resistance in French and American varroa mite populations. By comparing the sequences of octopamine and tyramine receptors (the known targets of amitraz) genes obtained from mites collected in different apiaries with different treatment regimens in France and United States, Hernández Rodríguez et al. indeed identified two variations in the β-adrenergic-like octopamine receptor gene Orβ-2R-L potentially associated with amitraz treatment failure events [24]. The first variation located at position 260 in the Orβ-2R-L gene (substitution of A to G at nucleotide 260 within the ORF) was exclusively detected in French mites. The resulting amino acid substitution within the β-adrenergic-like octopamine receptor, N87S (asparagine (AAT) to serine (AGT) substitution at position 87) is positioned at the end of helix II just near the residues thought to constitute the binding site for octopamine. This variation is likely to disturb the correct folding of the protein and thus, the efficient binding of the octopamine and of its antagonist, the amitraz. The second variation identified (substitution of T to C at position 643 of the ORF) was detected only in the mites collected in the U.S. It results in a tyrosine (TAT) to histidine (CAT) substitution at position 215 (Y215H) within the octopamine receptor. Located in the fifth transmembrane segment of the protein, this substitution is predicted to strongly reduce the stability of the receptor and to affect the interaction with its ligand.
The topic of resistance development against active ingredients of acaricides in varroa mite populations has received increasing attention in recent years [10,22,23]. Rising concerns among beekeepers, veterinarians, bee inspectors, and honey bee health researchers are due to an increasing number of reports of amitraz resistance in varroa mite populations from the field. Most of these reports stem from beekeeping operations in the United States and France, with ongoing investigations in Canadian beekeeping operations underway [22,23,27].
Here, we present data relevant for assessing the scope and underlying genetic mechanism of amitraz resistance in French varroa mite populations, sampled in the years of 2020 and 2021. During this timeframe, 1. Treatment efficacy of an authorized varroa control product (Apivar®) in the field, 2. Laboratory sensitivity of varroa mites towards amitraz, and 3. Genotyping data from French varroa mite populations, were collected. The objective of the present study was twofold: The first goal was to gain information about the spread and magnitude of amitraz resistance in the field in French apiaries. The second goal was to investigate genetic mechanisms underlying amitraz resistance in French varroa mite populations, focusing on the single nucleotide polymorphism at position 260 in the Orβ-2R-L gene described by Hernández Rodríguez et al. (2021) given their identification in French varroa populations [24].
Our study focuses on variation rather than mutation, using the term “variant” to describe alterations that can be benign, pathogenic, or of unknown significance. “Variant” is increasingly replacing “mutation” in scientific discourse.
2. Materials and Methods
2.1. Apiary Locations
Varroa populations were gathered from two “organic” apiaries from Bretagne and Occitanie, where amitraz treatments had not been used to determine the reference LC90. These apiaries differ from the others included in the study in 2020 and 2021.
Varroa mite samples for the main susceptibility assays were collected in June and July 2020 in 12 apiaries spread over 6 French regions: Nouvelle-Aquitaine, Occitanie, Provence-Alpes-Côte d’Azur, Auvergne-Rhône-Alpes, Centre-Val de Loire and Grand Est. A sample is defined as a sample of varroa mites from one or several frames of capped brood from a single colony. Two or three colonies were randomly sampled in each apiary.
In 2021, our focus was on two regions crucial to national honey production in the South of France: Occitanie and Provence-Alpes-Côte d’Azur (PACA). We collected varroa mite samples from 10 apiaries in these regions, as well as from two additional “organic” apiaries in same regions.
2.2. Laboratory Sensitivity of Varroa Mites Exposed to Amitraz in 2020 and 2021
2.2.1. Determination of Reference LC90 Concentration of Amitraz
Prior to the laboratory sensitivity tests, the reference LC90 (Lethal Concentration for 90% of the study population) for amitraz in varroa mites had to be pre-determined to validate the method on susceptible varroa mite populations from different regions in France. To do this, varroa populations from two “organic” apiaries which have not been exposed to amitraz treatments during the last 5 years were selected to be sampled between May and early June 2020. In these two apiaries, one located in Bretagne, the other one in Occitanie, adult female varroa mites were collected from (preferably) drone brood (or, if necessary, worker brood). Samples were taken from 5 to 6 colonies from the same apiary and sent to LDA 39 for a determination of the baseline LC90 for amitraz. A minimum of 300 Varroa mites per apiary was necessary to obtain meaningful results for these preliminary assays.
The apiaries selected for the determination of the reference LC90 were required to:
- −
- show no signs of other diseases (such as American or European foulbrood, or nosemosis),
- −
- −
- not have a new queen introduced or been merged with colonies that have been treated with amitraz in the past 5 years,
- −
- have sufficient nutritional resources and include frames that have been renewed,
- −
- varroa mites were harvested the same day of reception and 24 hours later by opening each cell capped one by one. For this preliminary study, varroa mites were grouped by apiary, even if they came from different colonies. Only mature varroa females (founding females or young mature females) extracted from capped cells were used for the assay. Ten subjects, or just eight when the quantity was not sufficient, were placed in each capsule. Capsules and Petri dishes were placed in an incubator at 32.5°C, with a relative humidity (RH) of about 70%. After 1 hour, the mites were observed under a magnifying glass binocular, then transferred to a Petri dish with 2 or 3 new larvae/nymphs of worker bees, taken from capped cells from the same brood fragments the varroa mites were harvested from.
Since amitraz is a lipophilic molecule, the method adopted comes from a standard method from the Coloss Beebook recommended for fat-soluble substances acting by contact (§ 3.6.3.2 of the Coloss Beebook [30]). It is based on the work of Milani et al. which was carried out between 1995 and 2004 [31]. The tested acaricide is incorporated at different concentrations into paraffin capsules.
A range of twelve concentrations, to which the negative control group was added, was tested as follows:
- −
- 3 replicates x 10 varroa mites per apiary: 0 ppm 0.5 ppm 1 ppm 2 ppm 3 ppm 5 ppm 7.5 ppm 10 ppm,
- −
- 2 replicates x 10 varroa mites per apiary: 12.5 ppm,
- −
- 1 replicate x 10 varroa mites per apiary: 15 ppm, 20 ppm, 50 ppm, 100 ppm.
To have more values around the dose corresponding to a 90% mortality obtained in 2020, the dilution range was revised in 2021 as follows:
- −
- 3 replicates x 8 varroas: 0 ppm 1 ppm 5 ppm 7.5 ppm 10 ppm 12.5 ppm 15 ppm 20 ppm 25 ppm 30 ppm
- −
- 1 replicate x 8 varroas: 40 ppm 50 ppm 100 ppm
2.2.2. Preparation of Paraffin Capsules Coated with Amitraz
Paraffin capsules are prepared as described in Milani (1995) [31] and in La Santé de l’Abeille No. 277 • 1-2 / 2017 [32]. These capsules are formed using 2 Na-Ca glass discs (62 mm in diameter) and 1 stainless steel ring (56 mm inside diameter - 2 mm thick - 5 mm high). The interior of these capsules is entirely covered by a thin layer of paraffin (Merck ref. 1.07151.1000 – melting point 46-48°C) containing a known concentration of amitraz.
For each of the concentrations, 10 grams of paraffin were melted in a crystallizer placed in a water bath at 60°C to which the required quantity of amitraz was added (Amitraz Pestanal® Sigma-Aldrich – ref. 45323-250MG), dissolved in 2 mL hexane (n-Hexane – VWR – ref. 24577.323). For negative controls, hexane alone was added. The mixture was stirred continuously for 1 minute and then regularly for at least 10 minutes to allow the hexane to evaporate. The metal rings were immersed in melted paraffin and one side of the glass discs was covered by lowering the disc on the melted paraffin.
Three to four capsules were prepared for each of the concentrations. The capsules were identified on each of the 2 glass discs. Capsules poorly coated or whose paraffin weight is outside the range [1.6 – 2 g] were excluded from the experiment. Paraffin coated capsules were kept open (disc + ring on one side, 2nd disk alone on the other) for 24 hours at room temperature to allow the hexane residues to evaporate, then were closed and kept at room temperature. They were placed in an oven at 32.5°C before the test.
2.2.3. Laboratory Exposure of Varroa Mites Towards LC90 Concentration of Amitraz
The mites were brought into contact with the treated paraffin. The contact time with the acaricide was set at 1 hour. Mortality rates were then counted 24 hours later. The action of amitraz, which targets the mites’ octopamine receptors at the level of synaptic transmission and which leads to paralysis of the mites is rather sub-lethal [33,34]. The death of varroa mites occurs secondarily due to starvation related to paralysis. Therefore, mites that were paralyzed after 24 hours, unable to move, were counted in the same title as dead mites.
During observation under a binocular magnifying glass, the mites were classified into 3 categories:
- −
- mobile mites: when mites could move when put on their paws and after stimuli, if necessary (with the brush),
- −
- paralyzed mites: when mites could move one or more appendages but without being able to move (change location),
- −
- dead mites: when there was no reaction after three stimuli.
Lost or accidentally killed mites (caught between the metal ring and a glass disc, for example) were not counted.
The assessment occurs at the conclusion of exposure to the acaricide while transferring the mites from the capsules to the Petri dishes, and it’s repeated 24 hours later at T0+24 after being introduced into the capsule.
2.2.5. Laboratory Assays in 2020 and 2021
In 2020, Véto-pharma organized laboratory assays to detect varroa mite susceptibility towards amitraz in the laboratory which were carried out by the LDA39 (Laboratoire Départemental d’Analyses du Jura). Sampling of varroa mites was carried out between May and July 2020 from brood frames (of workers and drones) collected from different commercial apiaries across France and sent to the LDA39. Varroa mites were then exposed to a pre-determined concentration of amitraz (LC90: 90% lethal concentration), in laboratory tests. The sensitivity towards amitraz of varroa populations from different regions of France was measured using the mortality of varroa mites during the assay as an indicator of susceptibility.
The second laboratory assay in 2021 aims to consolidate the first results obtained both in the predetermination of LC90 and by carrying out additional sensitivity tests with varroa mite samples from 10 different beekeepers. When there were not enough varroa mites in the brood, the tests were adapted and carried out with 8 varroa mites instead of 10 per capsule replicate.
In addition, in 2021, these laboratory susceptibility tests were coupled with field efficacy tests with the authorized varroa treatment Apivar®.
Prior to the laboratory tests measuring mite sensitivity towards amitraz, the reference LC90 for the chosen method was pre-determined on susceptible varroa populations from organic apiaries. Varroa samples from organic apiaries in France that have not been exposed to amitraz treatments in the past 5 years were selected. In 2020, the tests carried out on two apiaries located in Bretagne and Occitanie had already established a reference LC90. To confirm these first results and increase the validity of the reference concentration, two additional apiaries were sampled in 2021, one in the PACA region (Provence-Alpes-Côte d’Azur) and one in Occitanie.
For the main varroa mite sensitivity test in the laboratory, in each of the regions, five apiaries were selected. The mite samples for a given apiary were collected from six brood frames from six colonies. The samples were taken over a period of 5 weeks between June 7 and July 6 at the rate of 2 apiaries per week. Due to a very low varroa infestation during this first period, additional samples were taken between July 19 and September 13. As it is a fact that sensitivity is not the same from one hive to another in the same apiary [23], sensitivity tests are carried out at the colony scale.
2.2.4. Data Analysis
The percentage of miticide effect (mortality/paralysis) was calculated as follows:
(no. of paralyzed varroas + no. of dead varroas – no. of varroas lost or accidentally dead)
(initial number of varroa mites – number of varroa mites lost or accidentally dead)
The % effect obtained for the different concentrations were corrected to subtract the “natural mortality” according to the Schneider-Orelli formula: (b – k)/(1 – k) with b = % effect in the test and k = % effect in the negative control [30].
LC90 calculations were obtained using the probit transformation method. The confidence interval of the LC90 is given with a 5% risk of error. The coefficient of determination R² represents the quality of prediction of the linear regression. The prediction is good when R² is close to 1, it is on the contrary weak for values close to 0. The correlation coefficient r represents the strength of correlation between the acaricide concentration and % effect variables. The correlation is good for values between 0.5 and 1. It is low for values between 0 and 0.5.
The trials for which the % effect in the negative controls was greater than 30% were invalidated according to Beebook method (cf.§ 3.6.3.2 in Beebook II).
Varroa mite populations were classified into 3 categories based on the % effect obtained at the LC90:
- -
- between 75% and 100%: sensitive population
- -
- between 60% and 75%: intermediate population
- -
- under 60%: resistant population
3. Genotyping of the ORβ-2R-L Gene and Potential Links to Amitraz Resistance
3.1. Genomic DNA Extraction
Genomic DNA extraction from the mites to be genotyped was carried out as described by González-Cabrera et al. (2013) [17]. Briefly, individualized mites transferred in 1.5 Eppendorf tubes were first washed with 1 mL of UP water to remove traces of ethanol in which they were kept until analysis. After water removing, they were incubated at 99°C for 10 min in 20 µL of 0.25 M NaOH. They were subsequently crushed using a plastic homogenizer and the solution was neutralized adding 10 µL of 0.25 M HCl, 5 µL of 0.5 M Tris-HCl and 5 µL of 2% Triton X-100 before a last incubation at 99°C for 10 min. Genomic DNA suspensions were stored at -20°C.
3.2. Genotyping by Sanger Sequencing
Amplification of the ORβ-2R-L gene by PCR was performed in a reaction mixture of 25 µL containing 1.5 µL of genomic DNA, 12.5 µL of AccuStart II PCR SuperMix 2X (Quantabio) and 0.2 M of each primer. PCR program consisted in an initial denaturation step at 95°C for 10 min, followed by 40 cycles of 95°C for 20 s, 55°C for 20 s and 70°C for 1 min, and a final elongation step at 70°C for 1 min. The primers used, ORβ-2R-L_Seq For (5’-GCTGATCTCGATCATATTGAC-3’) and ORβ-2R-L_Seq Rev (5’-CTCGAGTGGCTTGATGATCGC-3’), were designed with the program Primer 3 using as reference a sequence of the ORβ-2R-L gene available in the NCBI database (accession number: XM_022808962.1). They allow the amplification of a short fragment of the gene (304 bp) covering the targeted variation at position 260. They. After checking of their length by capillary electrophoresis, PCR products were purified using 2 µL of ExoSAP ITTM (Applied Biosystems) for 5 µL of gDNA. PCR Sanger reactions were performed using 3 µL of purified fragments and the primers that were used for the amplification step. Sanger PCR products were run on the SeqStudio (Applied Biosystems) and the obtained sequences were aligned the reference sequence of the gene (accession number: XM_022808962.1).
4. Treatment Efficacy of Apivar® in the Field
Treatment Efficacy of Apivar® - Protocol 2021 (PACA and Occitanie Region)
The field efficacy trial with Apivar® was carried out on two apiaries with 25 colonies each, located in Avignon (Department 84, PACA region) and Auzeville-Tolosane (Department 31, Occitanie), by the French commercial beekeeper associations ADAPI (Provence-Alpes-Côte d’Azur) and ADA Occitanie. The hives of the five beekeepers from each region were moved to these two apiaries at the end of August 2021. The installation of the Apivar® treatment was carried out on August 31 in Auzeville and on September 14 in Avignon.
Each hive has been equipped with a meshed floor, allowing for easy varroa mite counts on white papers on the bottom board. Counts were performed once or twice a week, depending on the phase of the test. Initial counts were conducted 11 to 14 days before the application of the Apivar treatment. Mite counts were then continued until up to three weeks after the application of the second control treatment (see below: Control treatments).
On the day of application of the Apivar® strips (Day 0 or D0) and on the day of their removal (Day 70 or D70), the colonies were evaluated with the ColEval method [35], weighed and sampled to assess the rate of phoretic varroa mites per 100 bees (PVM100bees). On day 35 of the treatment, the strips were cleaned with a hive tool and repositioned in the center of the brood nest (Figure 1).
Two control treatments were applied. When the Apivar® (D70) strips were removed, the queens were caged and four Bayvarol (flumethrin) strips were inserted between the brood frames. After 28 days, these strips were removed, the queens released and an Oxybee® (oxalic acid) treatment was applied at a rate of 5 mL per occupied interframe.
The treatment efficiencies measured at the colony correspond to the sum of the varroa falls during the 10 weeks of Apivar® treatment (Days 0 to 70 or D0-70) compared to the total number of varroa mites fallen during the entire device (Apivar® + Bayvarol® and Oxybee® control treatments; Days 0 to 112 or D0-112).

5. Results
5.1. Laboratory Sensitivity of Varroa Mites Exposed to Amitraz LC90 and Genotyping of Varroa Mites in 2020
5.1.1. Laboratory Sensitivity of Varroa Mites Towards the LC90 of Amitraz (2020)
The results of the preliminary test, determining the reference LC90 of amitraz in varroa mites, was carried out with mite samples from two different apiaries from Occitanie (46 capsules of 10 varroa mites) and Bretagne (17 capsules of 8 varroa mites). These assays resulted in the determination of an LC90 of 28 ppm [CI95: 22-36]. The coefficient of determination (R²) obtained was 0.8, indicating a good prediction of the linear regression. The correlation coefficient (r) is 0.9, indicating a high strength of correlation between the variables “acaricide concentration” and “% mite mortality”.
Out of the 18 colonies sampled for the main sensitivity test, valid amitraz sensitivity assay results could be obtained for n=751 varroa mites (excluding negative controls) from 17 colonies from 9 different apiaries (Figure 2) The results from one colony were excluded because the mortality in the negative control group of mites not exposed to amitraz was too high (>30%, see Table 1). Of these 17 colonies, mite samples from 15 colonies were tested susceptible towards amitraz as per the pre-defined mortality rate of at least 75% (Table 1). Varroa mites from 2/3 of these colonies (n=10) showed a mortality rate of 95% or higher during and after the exposure towards the amitraz LC90 in the laboratory. Varroa mite samples from three colonies demonstrated a mortality rate around the expected 90% mortality, between 89% and 94%. The remaining two mite samples that were classified as susceptible towards amitraz demonstrated a mortality rate of 77% and 78% (Table 1).
Figure 2.
Laboratory Sensitivity of Varroa Mites Exposed to Amitraz LC90 and Genotyping of Varroa Mites in 2020 distribution. A – Distribution map of varroa population phenotyping – by colony (A) and by apiary (B).
Figure 2.
Laboratory Sensitivity of Varroa Mites Exposed to Amitraz LC90 and Genotyping of Varroa Mites in 2020 distribution. A – Distribution map of varroa population phenotyping – by colony (A) and by apiary (B).
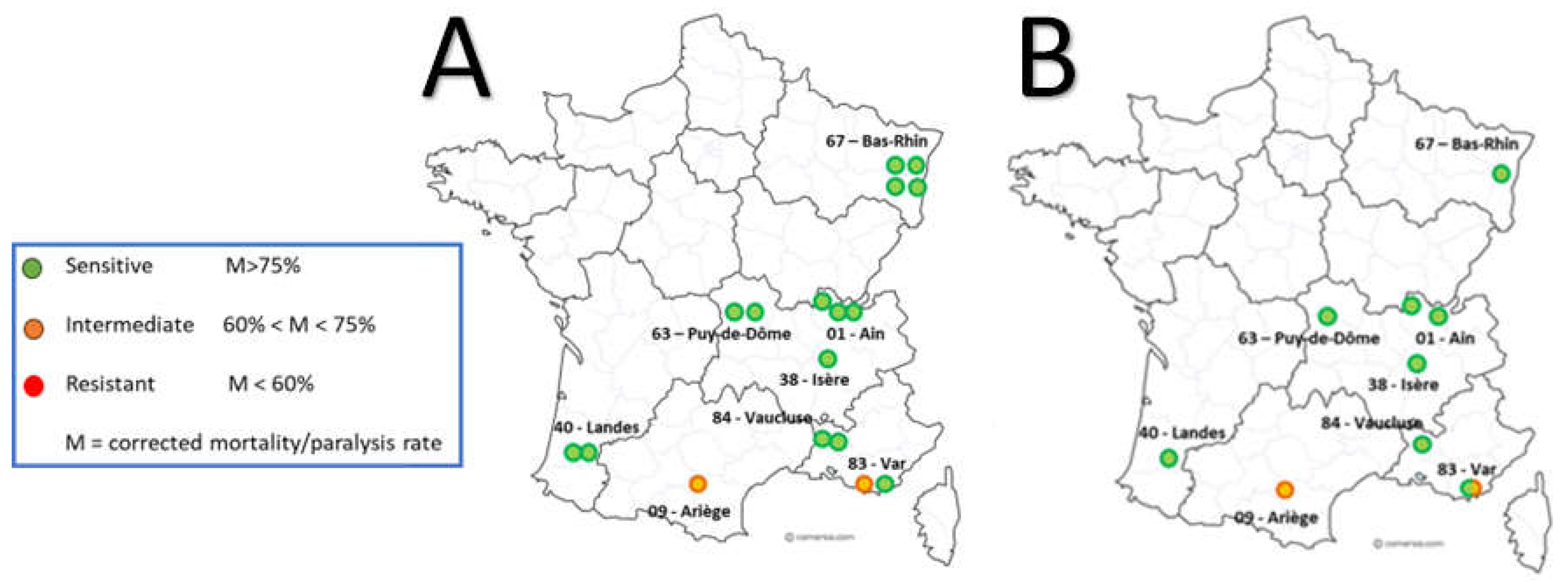

Table 1.
Overview of the mortality rate of varroa mites after exposure towards the LC90 of amitraz in the laboratory (2020). The sample size (n) represents the total [
number of mites tested over 6 French regions: Nouvelle-Aquitaine, Occitanie, Provence-Alpes-Côte d’Azur, Auvergne-Rhône-Alpes, Centre-Val de Loire and Grand Est. Mortality rates of those total samples are given in % for LC90 test assays and negative controls.
Table 1.
Overview of the mortality rate of varroa mites after exposure towards the LC90 of amitraz in the laboratory (2020). The sample size (n) represents the total [
number of mites tested over 6 French regions: Nouvelle-Aquitaine, Occitanie, Provence-Alpes-Côte d’Azur, Auvergne-Rhône-Alpes, Centre-Val de Loire and Grand Est. Mortality rates of those total samples are given in % for LC90 test assays and negative controls.
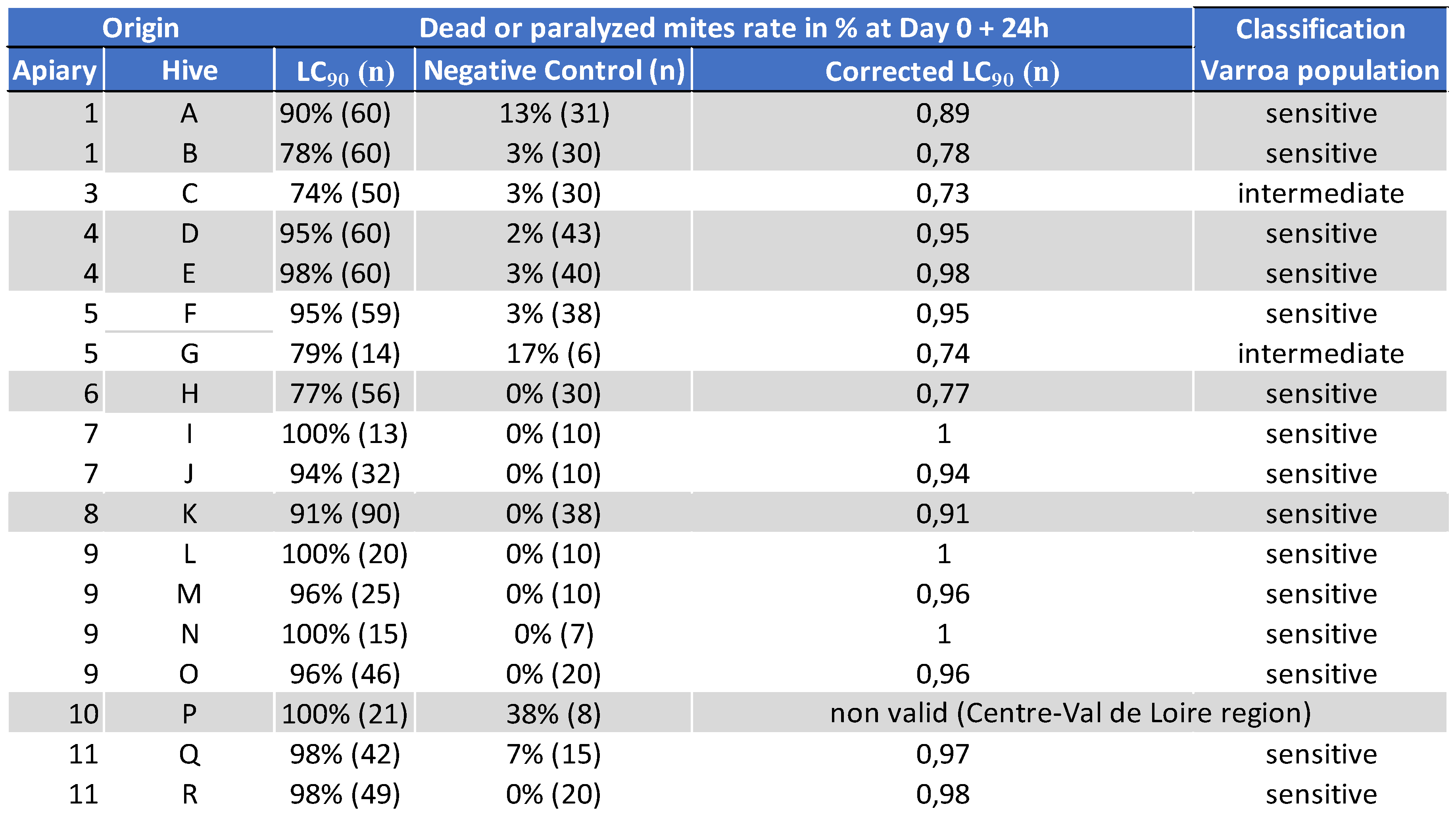
Varroa mites from two out of the 17 sampled colonies were classified as showing intermediate sensitivity (60-75% mortality rate) in response to amitraz exposure in the laboratory (Table 1). Not one of the mite samples - and therefore the associated varroa populations of the respective colonies - was classified as resistant against amitraz (Table 1).
The overall LC90 mortality/paralysis rate for susceptible populations is 92% (n = 751), very close to the 90% expected, which supports the estimate of the LC90 obtained in preliminary tests.
No correlation was found in this study between the infestation rate and level of sensitivity of varroa mites. The four populations with the lowest mite sensitivity results (between 73% and 78% mortality rate) come from hives with varroa brood infestation rate estimates between 1.9% and 5.4%. The 13 most sensitive mite populations (between 89% and 100% mortality rate) correspond to highly variable infestation rates in the respective honey bee colonies, with values between a minimum of 1% up to a maximum value of 22.8% varroa mite infestation.
5.1.2. Genotyping of Varroa Mites Previously Tested in the 2020 Laboratory Assay
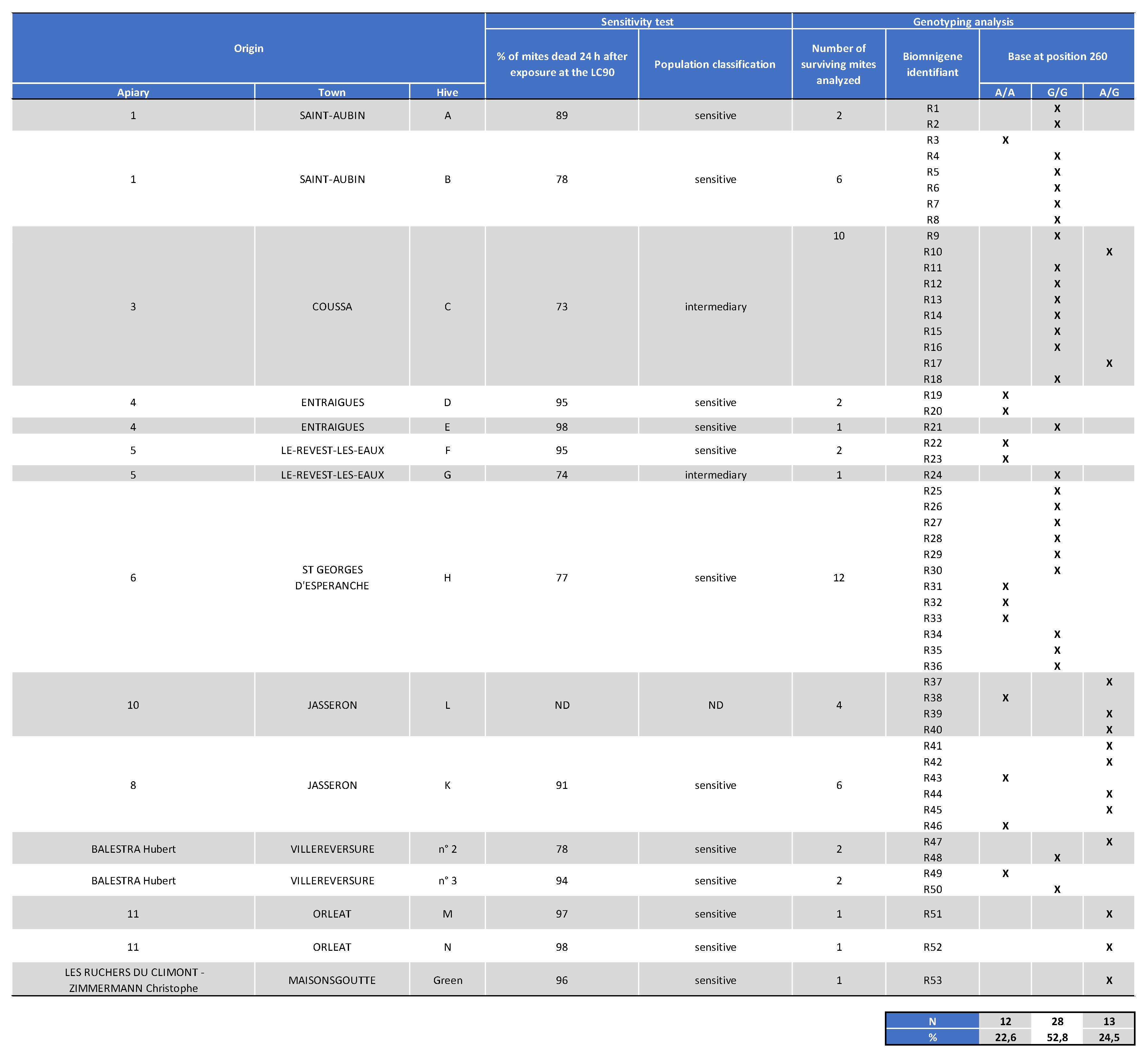
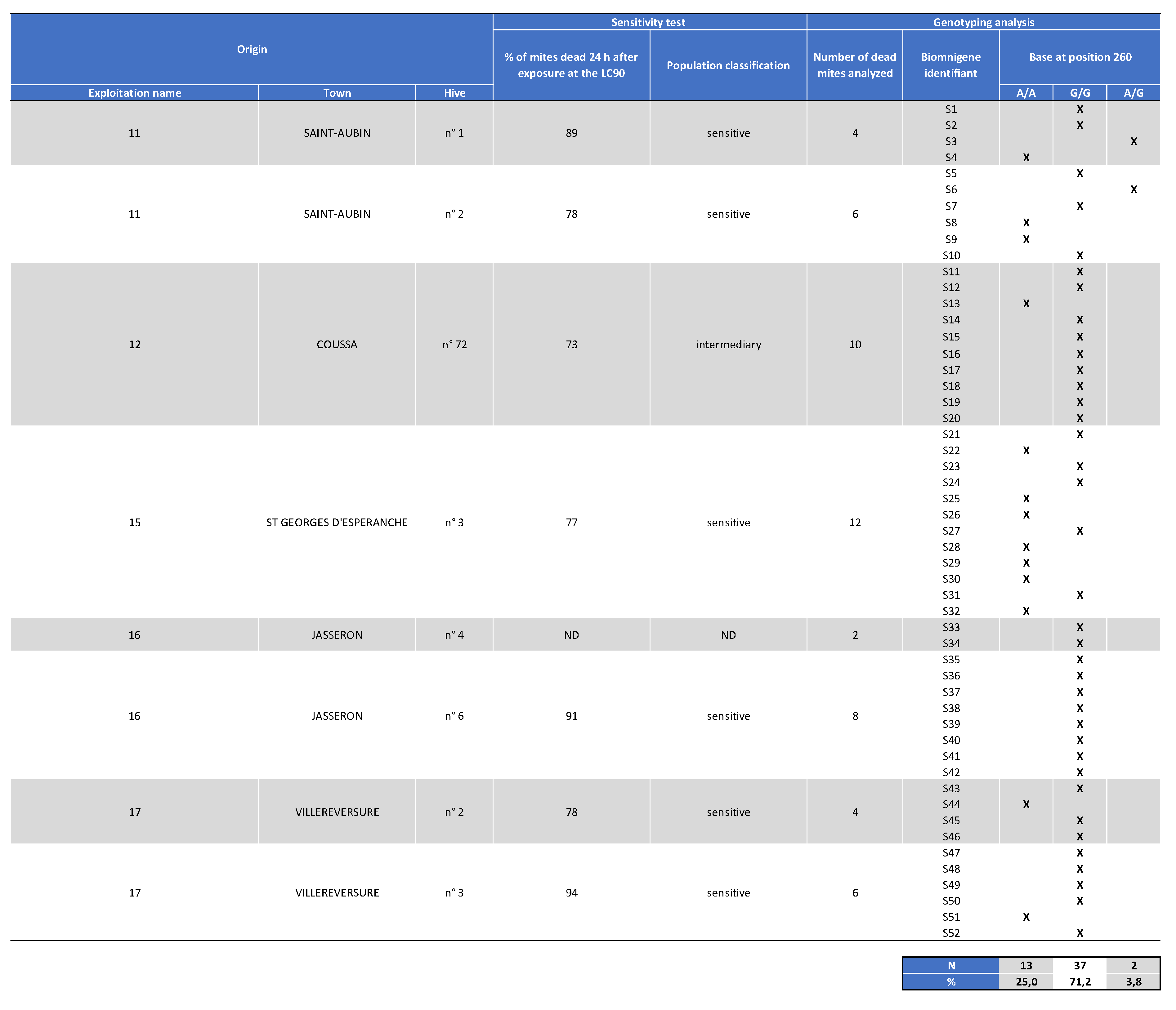
A total of 105 varroa mites from all mites that were tested in the 2020 laboratory assays (53 and 52 mites selected randomly among the dead and the surviving mites, respectively; see Appendix I and Appendix II for more details) were later genotyped, and grouped in one of three genotypes, depending on the base pair detected at location 260 of their ORβ-2R-L gene: 1. A/A (wild-type) (n=25), 2. A/G (heterozygous) (n=15), and 3. G/G (homozygous variation) (n=65). To note, genotyped mites originated from 15 different colonies, 12 that were classified as sensitive and two that were classified as showing intermediate sensitivity towards amitraz at the end of the laboratory sensitivity assay. The test did not give valid results for the remaining colony (Appendix I and Appendix II).
Thus, a total of 80 of the varroa mites previously tested in the laboratory assay carried the variation at position 260 of the ORβ-2R-L gene described by Hernández Rodríguez et al. [24], 65 of them presenting a homozygous genotype.
Looking at a potential link between varroa mite sensitivity towards amitraz and the genotypes determined for 105 of the assay-tested mites, we were unable to identify a direct relationship between the variation at position 260 and a resistant phenotype in mites that had survived the assay.
Table 2 shows that in both groups, mites that perished during or shortly after exposure to amitraz in the laboratory and mites that survived the assay, a majority of mites carried the variation identified by Hernández Rodríguez et al. [24]. The percentage of mites carrying the variation was very similar in both groups (75% in mites that had died after amitraz exposure and 77.3% in mites that had survived amitraz exposure), not showing significant difference (Chi2=0.0029765, df=1, p=0.9565) (Table 2). The only significant difference (Chi2=7.606, df=2, p=0.005817) between both groups (perished and surviving mites) is the distribution of homozygous and heterozygous individuals: 24.5% of surviving mites carried the A/G genotype, whereas the same was true for only 3.8% of perished mites (Table 2). Likewise, 52.8% of mites carried the homozygous G/G genotype in the group of surviving mites, against 71.2% among the perished mites.
Table 2.
Genotyping results for varroa mites, sampled after LC90 exposure towards amitraz. Results show the base at position 260 of the varroa mite ORβ-2R-L gene (2020).
Table 2.
Genotyping results for varroa mites, sampled after LC90 exposure towards amitraz. Results show the base at position 260 of the varroa mite ORβ-2R-L gene (2020).

5.2. Laboratory Sensitivity of Varroa Mites Exposed to Amitraz LC90, Treatment Efficacy of Apivar® in the Field, and Genotyping of Varroa Mites in 2021
5.2.1. Laboratory Sensitivity of Varroa Mites Towards the LC90 of Amitraz (2021)
The two varroa mite samples utilized for the determination of the LC90 were received on 27 May and 1 June 2021. The number of live varroa mites harvested from the two “organic” apiaries in the Occitanie and PACA regions for preliminary testing was 217 and 279, respectively - lower than the 300 initially envisaged. As a result, the tests were adapted and carried out with 8 varroa mites instead of 10 per capsule replicate, as stated in Materials and Methods, and, for the 1st apiary, two replicates instead of three were set up for the concentrations of 1, 5, 15 and 20 ppm.
The results obtained for these two apiaries were compiled with the results obtained for two other apiaries sampled in 2020 and resulted in an estimate of the LC90 of 25 ppm [CI95: 21-29]. The coefficient of determination (R²) obtained is 0.7 which indicates a good prediction of linear regression, and the correlation coefficient (r) is 0.8 indicating a high correlation strength between the variables acaricide concentration and % mortality effect.
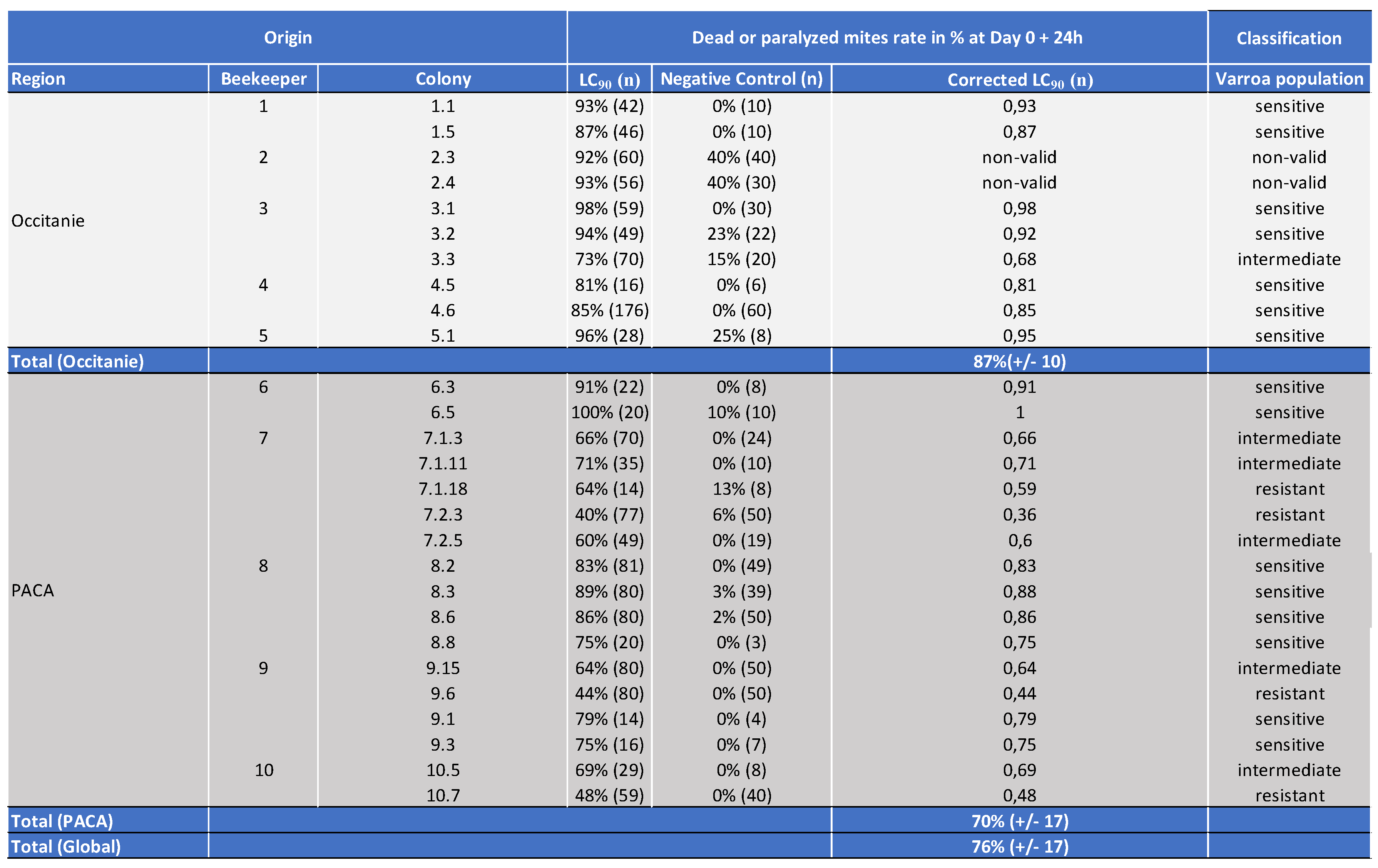
Although a total of 84 brood frames were sampled to test laboratory sensitivity of varroa mites towards amitraz, only 25 in vitro assays resulted in valid sensitivity rates (8 samples from Occitanie and 17 samples from PACA). The other samples showed too low infestation rates, problems related to excess mortality of varroa mites in the brood during transport or in negative controls during the tests.
The average level of susceptibility of varroa populations towards LC90 (25 ppm amitraz) exposure is 76 ± 17%. This sensitivity varies from 58 ± 13% to 96 ± 6% depending on the beekeeping operation with contrasting profiles depending on the region (87% ± 10% in Occitanie vs. 70 ± 17% in PACA) (Table 3). In total, of the 25 colonies sampled, 15 varroa populations were classified as sensitive towards amitraz (75-100% mortality), 6 were classified as intermediate (60-75% mortality) and 4 were classified as resistant (<60% mortality).
Table 3.
Overview of the mortality rate of varroa mites after exposure towards the LC90 of amitraz in the laboratory (2021). The sample size (n) represents the total number of mites tested. Mortality rates of those total samples are given in % for LC90 test assays and negative controls. (Géographic maps).
Table 3.
Overview of the mortality rate of varroa mites after exposure towards the LC90 of amitraz in the laboratory (2021). The sample size (n) represents the total number of mites tested. Mortality rates of those total samples are given in % for LC90 test assays and negative controls. (Géographic maps).
5.2.2. Genotyping of Varroa Mites Previously Tested in the 2021 Laboratory Assay
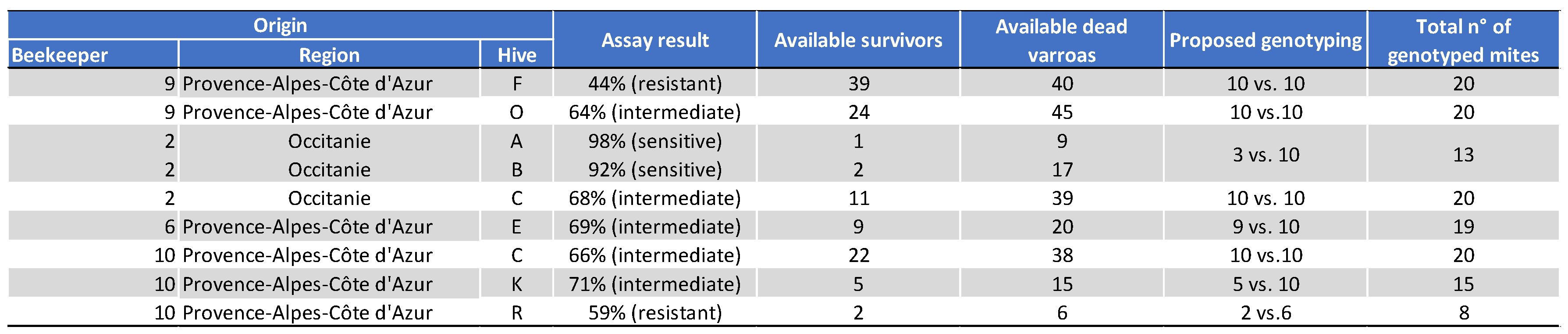
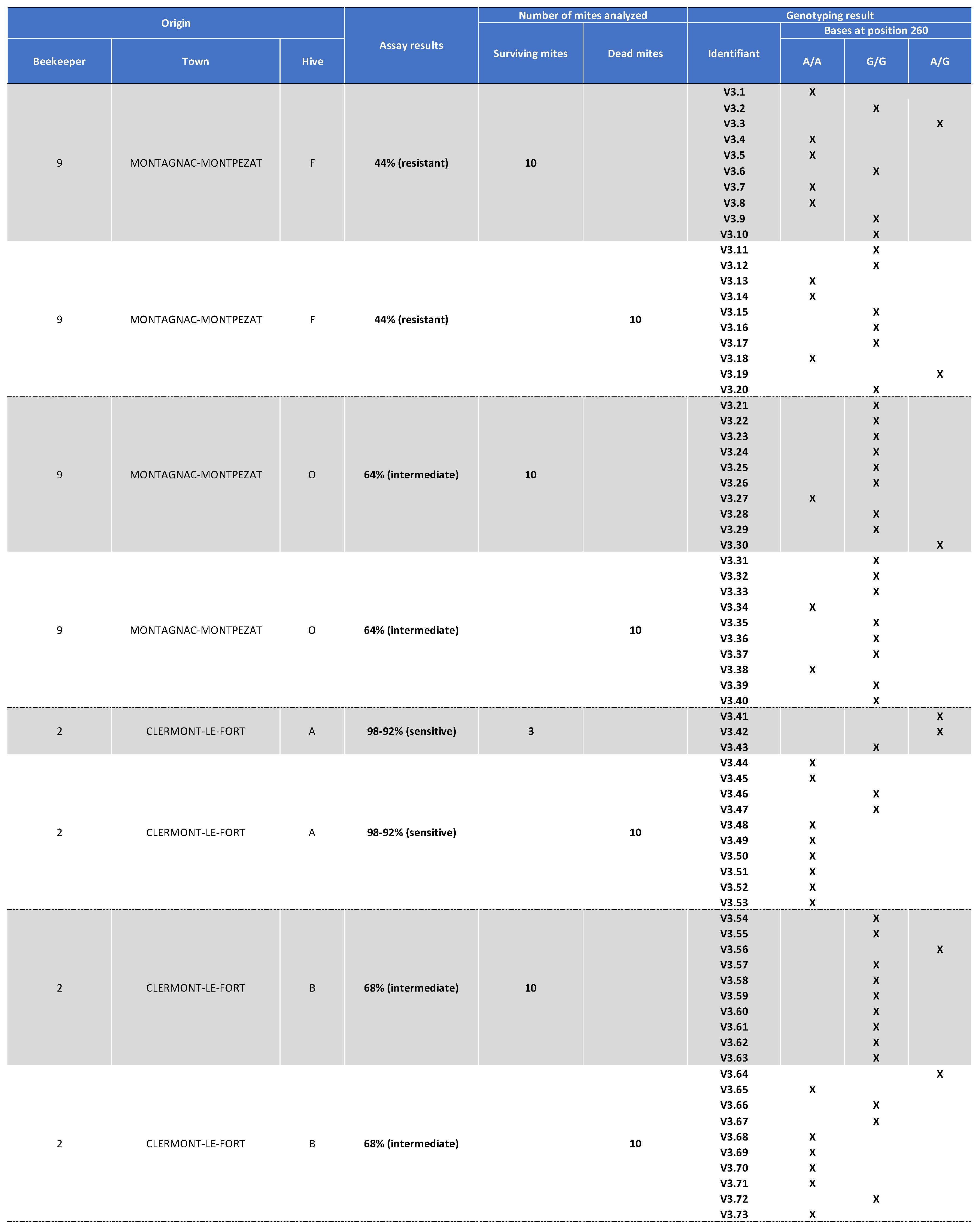
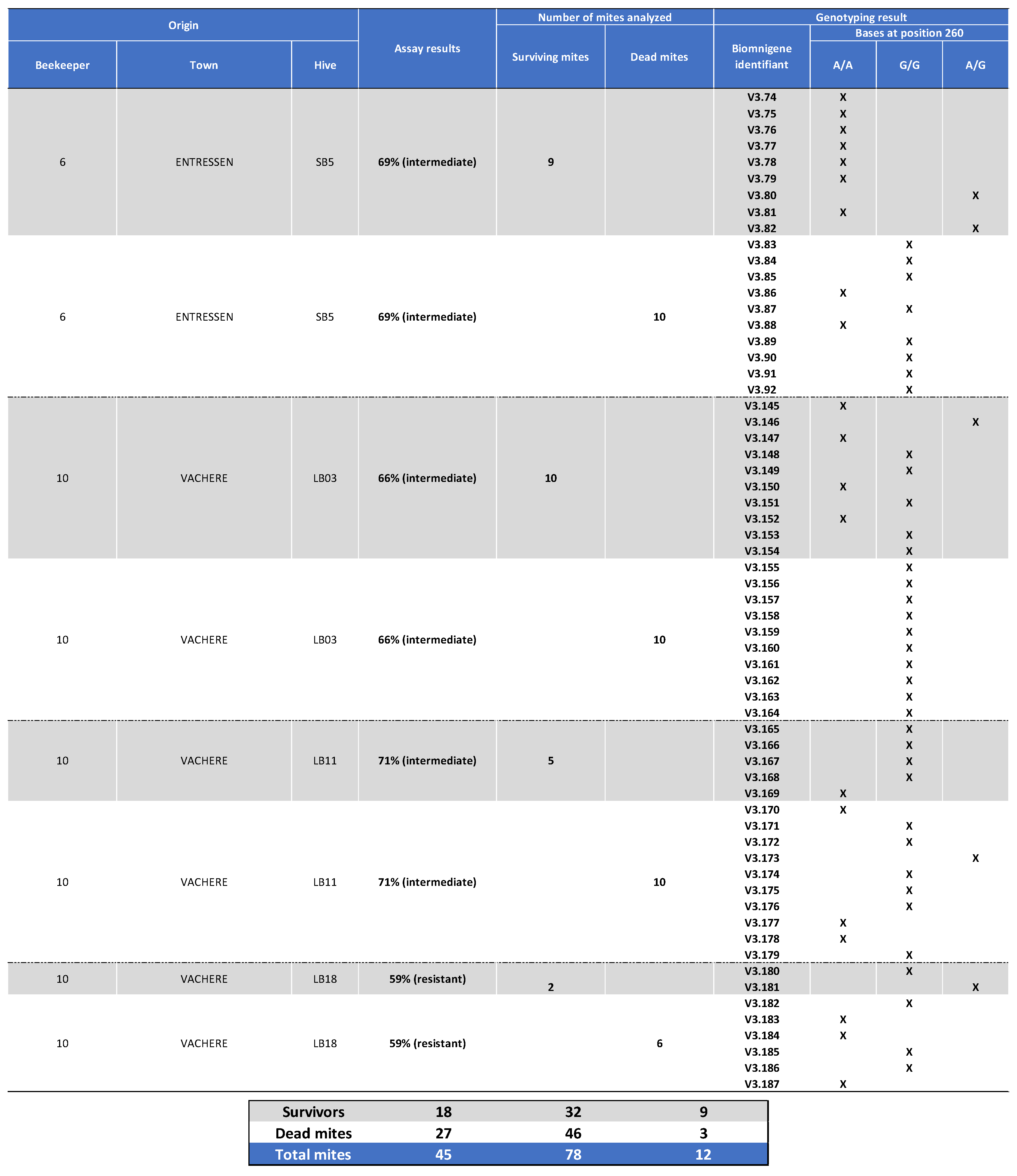
A total of 135 varroa mites (59 surviving mites and 76 dead mites at the end of the laboratory sensitive assay) from 8 different colonies sampled for the laboratory sensitivity test and included in the field efficacy test with Apivar® were genotyped in 2021. Two of the colonies had previously been classified as resistant in the laboratory sensitivity assay, five colonies were classified as showing intermediate sensitivity towards amitraz, and the remaining one was classified as sensitive towards amitraz (Table 4).
Table 4.
Overview of genotyped varroa mites from laboratory assay samples, presenting the mites’ origin, laboratory sensitivity result of the colony, and status of the mites after amitraz exposure in the laboratory (survivors vs. dead varroa mites).
Table 4.
Overview of genotyped varroa mites from laboratory assay samples, presenting the mites’ origin, laboratory sensitivity result of the colony, and status of the mites after amitraz exposure in the laboratory (survivors vs. dead varroa mites).
Like the genotyping results from the year 2020, the 2021 analyses show a higher percentage of mites carrying the homozygous G/G genotype (the variation, detected by Hernández Rodríguez et al.) compared to the homozygous wild-type A/A. This is true for varroa mites from both groups (surviving mites and mites that perished during amitraz exposure in the laboratory) (Table 5). Similarly (compared to the 2020 results), the group of heterozygous mites makes up the smallest portion of the three genotypes: 8.9% in 2021 vs. 14.3% in 2020 (Table 2 & Table 5).
Table 5.
Genotyping results for varroa mites, sampled after LC90 exposure towards amitraz. Results show the base at position 260 of the varroa mite ORβ-2R-L gene (2021).
Table 5.
Genotyping results for varroa mites, sampled after LC90 exposure towards amitraz. Results show the base at position 260 of the varroa mite ORβ-2R-L gene (2021).
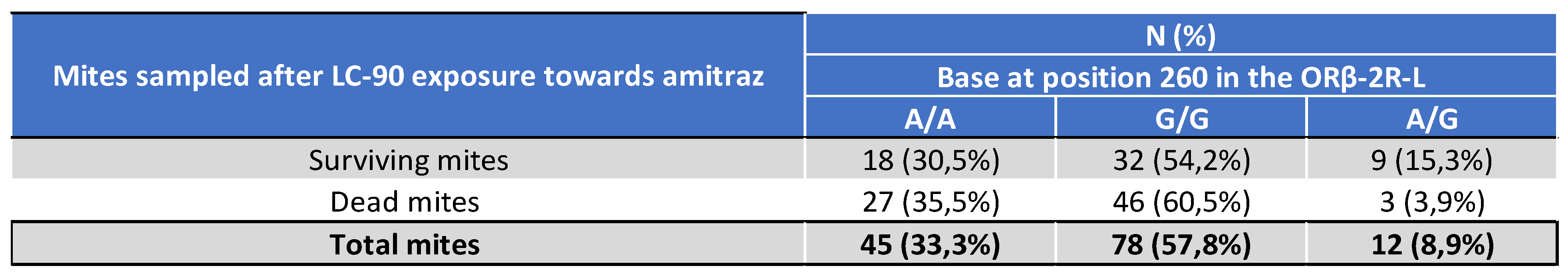
Between varroa mites which survived amitraz exposure in the laboratory and varroa mites which did not survive, there is only a slight difference (Chi2=2.5128, df=1, p=0.1129) in percentage of mites carrying the variation (G/G) with a homozygous genotype (Table 5). When it comes to the heterozygous genotype, as in 2020, surviving mites are more likely to carry this genotype with 15.3% compared to 3.9% heterozygous mites within the pool of mites that perished during the laboratory assay exposure to amitraz.
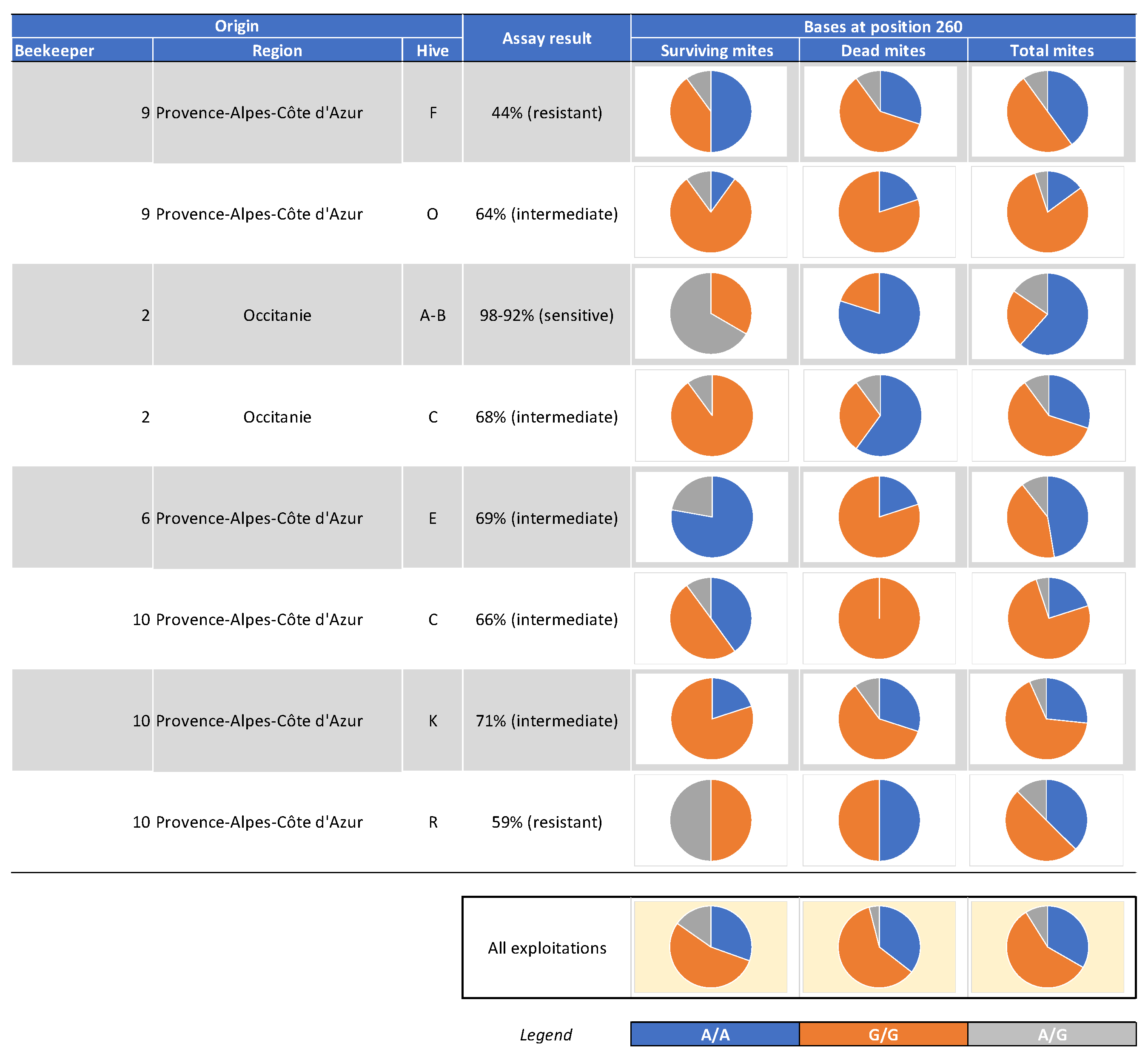
In a more detailed portrayal of the results, Figure 3 shows the partitioning of genotypes (A/A, G/G, and A/G) in individual mite samples (survivors vs. perished mites) for each individual colony as well as summarized across all genotyped mites (Figure 3).
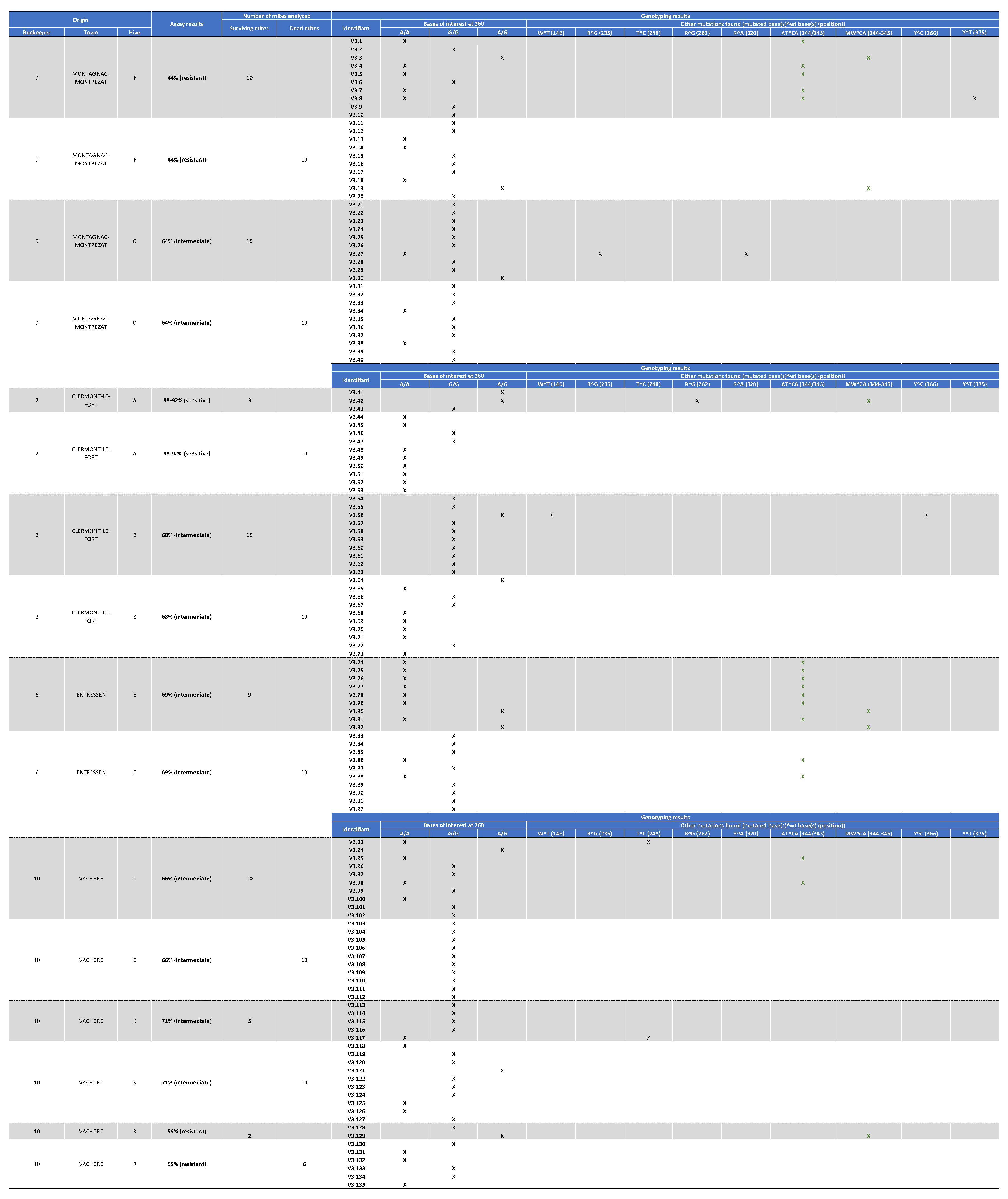
In addition, Appendix III presents the individual result for each genotyped varroa mite, including information about the origin, classification after the laboratory assay (sensitive, intermediate, or resistant) as well as the individual assay outcome (survived vs. perished) of the assay.
Additional variations were found in the ORβ-2R-L gene (nature and positions of the variations described in the table reported in Appendix IV). We notice that most of them, especially the variated bases AT at positions 344-345 (found in 19 out of the 59 surviving mites), were present in surviving mites on the same allele as the base A at position 260, suggesting an involvement in the acquisition of the resistance to amitraz. This hypothesis would merit further exploration.
5.2.3. Treatment Efficacy of Apivar® (2021) in the Occitanie and PACA Regions
A total of nine colonies were excluded from the final evaluation of the field efficacy, five in Occitanie and four in PACA, because the queen had to be renewed at the beginning of the test (n = 3), because the initial varroa infestation was too low (<300 varroa mites; n = 2) or because of significant weakening of the bee population at the end of treatment (“non-value” < 3000 bees; n = 4).
The mean efficacy in all tested colonies in both regions at the end of the 10-weeks treatment was 93% (SD±6.4) (n=41 colonies). Twenty-one of these colonies demonstrated an efficacy of 95% or higher, 7 of these in PACA and 14 in Occitanie. Of the 20 colonies below this threshold, 9 did not achieve 90% efficiency (Figure 4). The mean field efficacy after 10 weeks in the two different regions was 95.5% (SD±2.7) in Occitanie (n=14/20 > 95% Eff.; n = 2 < 90% Eff.) and 90.5% (SD±7.8) in PACA (n=7/21 > 95% Eff.; n=7 < 90% Eff.) (Table 6, Figure 5).
Figure 4.
Map presenting field efficacy and sensitivity results assessed per colony in each apiary.
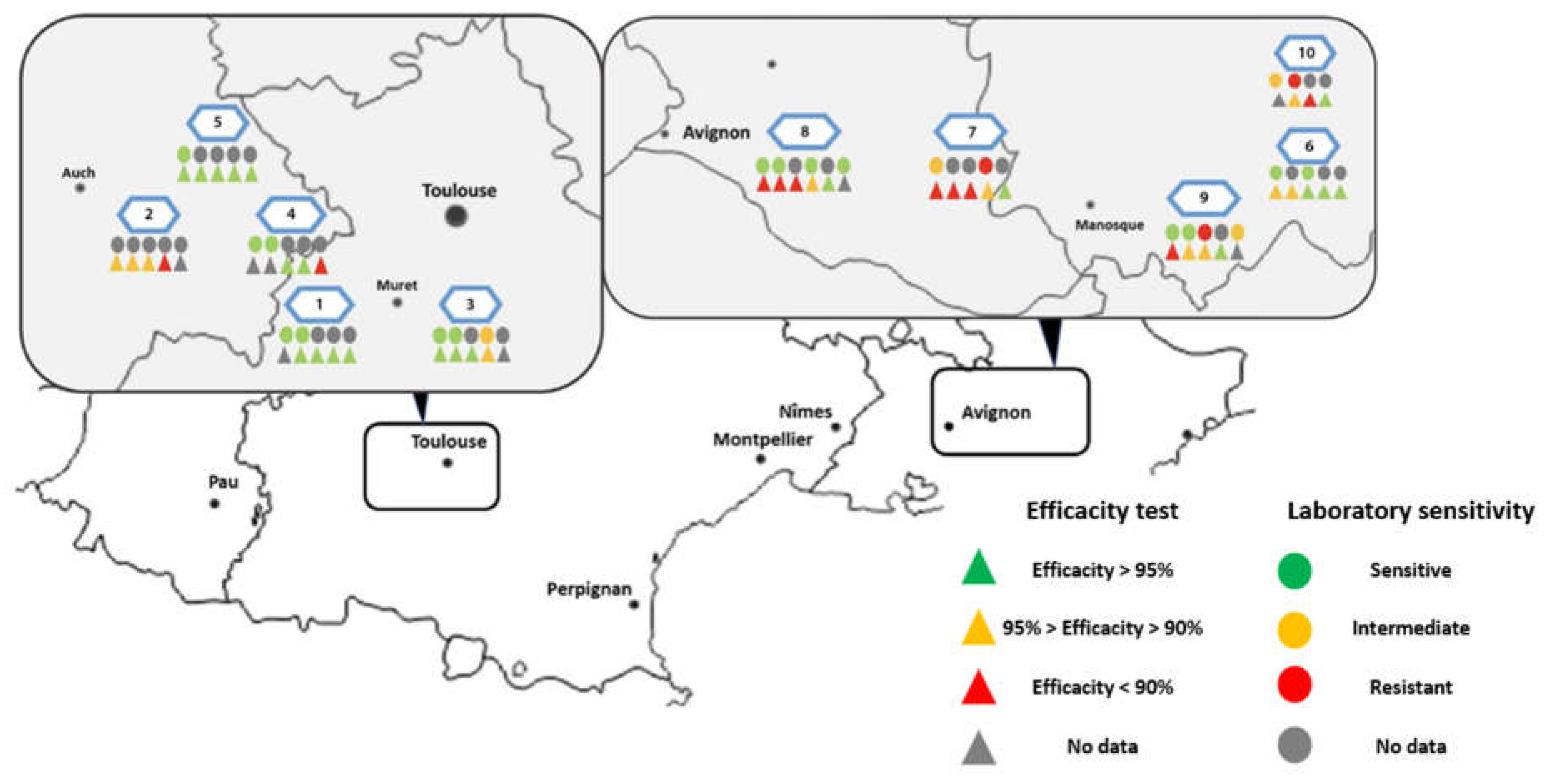
Table 6.
Summary table of efficacy results (SD averages both globally and segmented by region).
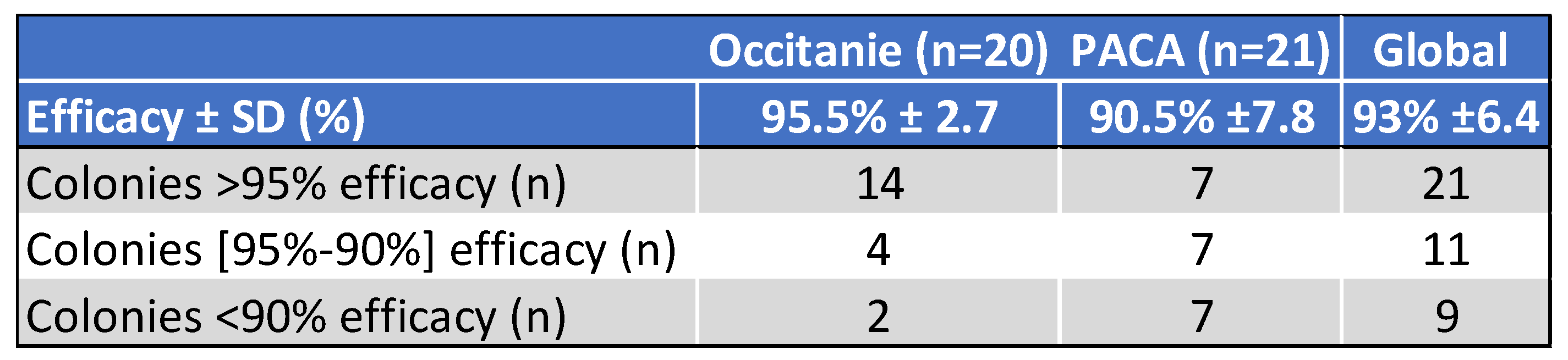
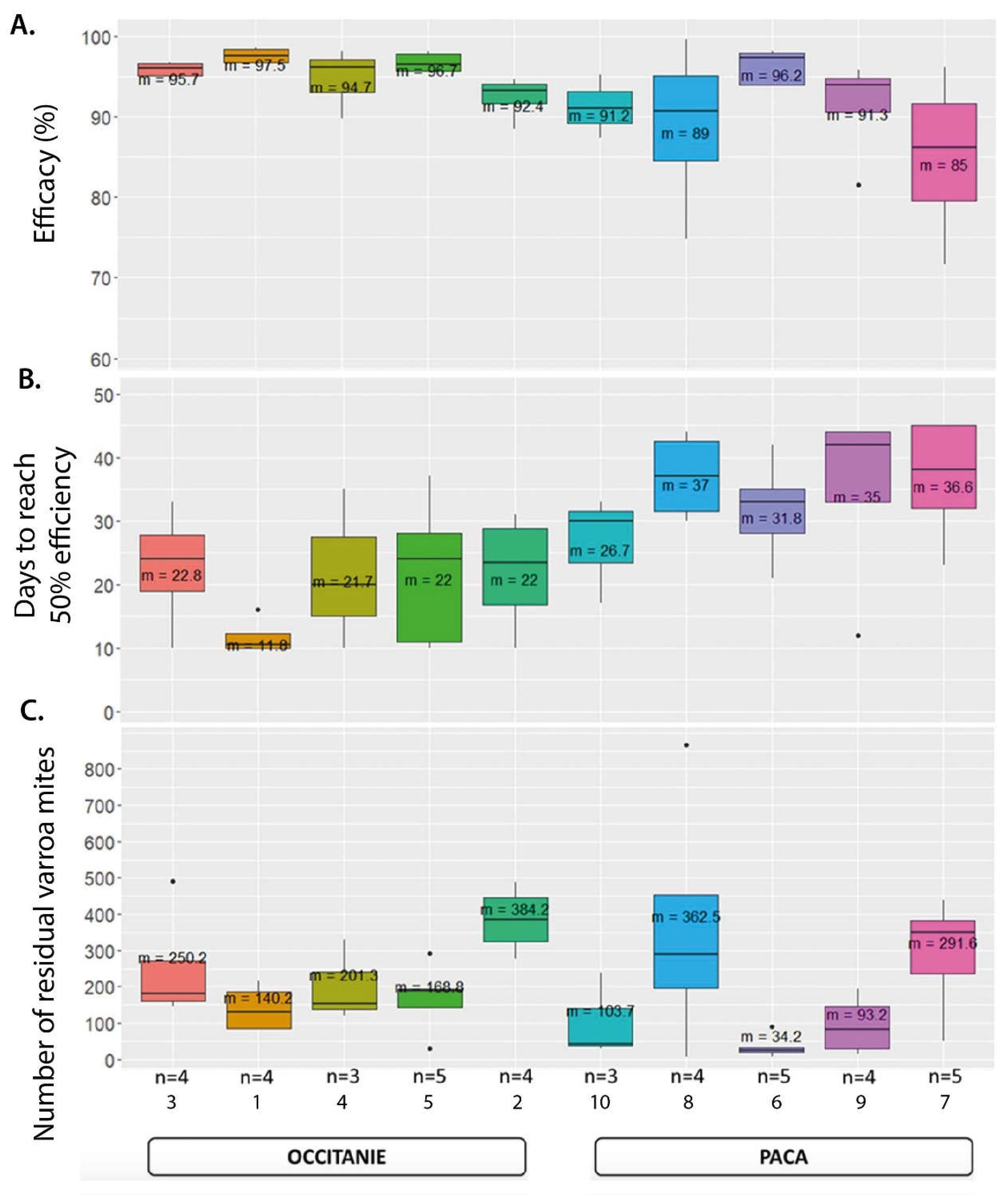
Figure 5.
Boxplots of calculated efficacy for the Apivar treatment (A), days to achieve 50% efficacy (B) and numbers of residual varroa mites after treatment (C). Numbers (n) of hive per apiary and means (m) are indicated.
Figure 5.
Boxplots of calculated efficacy for the Apivar treatment (A), days to achieve 50% efficacy (B) and numbers of residual varroa mites after treatment (C). Numbers (n) of hive per apiary and means (m) are indicated.
A systematic comparison of varroa populations with regards to amitraz sensitivity measured in the laboratory and efficacy of an Apivar treatment in the field on 50 colonies could not be fully realized. Despite the effort of sampling a total of 84 brood frames, only 25 in vitro assays resulted in valid varroa mite mortality rates (8 samples from Occitanie and 17 from PACA). The other samples showed a too low mite infestation, problems with excess mortality of varroa mites in the brood during transport or in negative controls during the tests.
The amitraz sensitivity - Apivar efficacy relationship could only be studied on 16 colonies (9 colonies were excluded from the field trial between varroa mite sampling and the end of the trial). Despite the small amount of data, the results show a tendency towards better treatment efficacy on colonies with more sensitive varroa populations (Spearman test, P<0.05, rho = 0.5). This correlation is no longer significant when only the 11 colonies monitored in PACA are considered, revealing a regional effect. This indicates mite sensitivity is likely not the only factor relevant for treatment efficacy in the field.
6. Discussion
6.1. Geographic Distribution and Spread and Magnitude of Amitraz Resistance Detected the Field in French Apiaries
Over the course of two years (2020-2021), laboratory sensitivity of varroa mites towards amitraz and field efficacy results after treatment of honey bee colonies with an amitraz-based veterinary medicine against Varroa destructor were collected from a total of 75 honey bee colonies, stemming from 19 different apiaries. The sampled apiaries were predominantly located in the Eastern, Central, and Southern regions of France. The main reasons for the focus on these geographic regions were that 1. amitraz resistance in French varroa mite populations was first reported from the AURA (Auvergne-Rhône-Alpes) region in the East of the country [22] and 2. The Southern regions of France offer a high density of honey bee colonies and commercial beekeeping operations.
The results of the laboratory assay in 2020, including varroa mites from 17 colonies of 9 different apiaries, do not indicate widespread amitraz resistance in the sampled regions of AURA, Bretagne, Grand Est and Occitanie. Varroa mite samples from 15 out of 17 sampled colonies were classified as “sensitive” towards amitraz. However, it is important to note that, due to shipment constraints during the COVID-19 pandemic in 2020 and resulting difficulties in testing varroa mites for the laboratory assays in time (when they were still alive), the total sample size of 17 colonies remains relatively low. On the other hand, the sampled mites tested in the assay were sourced from 9 apiaries across the country, including the AURA region in Eastern France and some apiaries where resistance had previously been reported. For the investigation of the genetic basis of amitraz resistance in Varroa destructor (see below), the number of mites available for genotyping was more than adequate.
In 2021, we decided to focus on two regions in the South of France with high importance for the national honey production [36]: Occitanie and Provence-Alpes-Côte d’Azur (short: PACA). For this project, a field efficacy trial, consisting of a 10-weeks application with the authorized amitraz-based product Apivar®, was added to the laboratory assays. We found that, in both tests (laboratory assay and field efficacy trial), mites from the Occitanie region were largely sensitive towards amitraz and test results (mortality rate in the assay, treatment efficacy in %) were superior compared with the PACA region. With a treatment efficacy of 95.5% in the field, the result for the Occitanie region was compliant with the required minimum varroa treatment efficacy of 95% for synthetic acaricides defined by the European Medicines Agency (EMA) [37]. The result of 90.5% efficacy in the PACA region was below the required minimum efficacy for synthetic treatments – the standard for organic varroa treatments is slightly lower with a required minimum efficacy of 90% [37]. The difference in field efficacy between the PACA and Occitanie regions may be explained with regional differences in treatment history. Participating beekeepers from the Occitanie region reported they had treated the honey bee colonies included in this trial with Apivar® strips for 10-14 weeks in previous years (maximum treatment period of Apivar® as per product label: 10 weeks). On the other hand, beekeepers from the PACA region reported they had treated their colonies included in the present trial with Apivar® for up to 24 weeks in previous years [38]. The prolonged, and possibly repeated exposure of varroa mite populations towards an amitraz-based treatment which, in France, is authorized for a maximum treatment period of 10 weeks could have contributed to desensitization of varroa mites in the PACA region towards amitraz. For a more detailed analysis of a correlation between off-label use of authorized and unauthorized amitraz-based varroa treatments, we suggest further investigations including a higher sample size of honey bee colonies with known treatment history.
Overall, the results of the present field efficacy trial support the hypotheses presented by Rinkevich (2020) [23] that 1. amitraz resistance occurs in “patches” or “islands” of resistance rather than affecting all honey bee colonies located in a single apiary or even broader geographic regions within a short period of time. 2. Treatment efficacy reduction in the field clearly develops more slowly than pyrethroid or coumaphos resistance in varroa mites [11,13,27]. Further, it might be closely related to apiary management practices such as compliance with treatment recommendations.
In addition, this trial demonstrated that, across regions, the measured mortality rate of mites in the laboratory assay was lower compared to the corresponding field efficacy result (in %) after treatment with Apivar®. This finding confirms similar results obtained in previous studies where field efficacy was compared to the mortality rate of varroa mites in sensitivity assays [14]. A global result of 93% field efficacy (n=41) corresponded to a laboratory mortality rate of 77.3% (n=16). Even when we consider the standard for treatment efficacy in the field is 95% mortality and treatments such as Apivar® have been formulated accordingly, while we exposed mites in the laboratory towards the LC90, the mean difference is slightly higher than expected. This could be due to different factors such as a higher natural mite mortality rate in honey bee colonies in the field [14] or a lack of long-term exposure of varroa mites towards amitraz in the laboratory that does not correspond to the weeks-long treatment exposure in the field and therefore does not result in comparable mite mortality. While conditions in a live honey bee colony are clearly more variable than the standardized conditions in the laboratory, the mites do have much more opportunity for avoidance behavior inside a honey bee colony, compared to a spatially limited paraffin capsule in the laboratory. This contradicts the observed reduced laboratory mortality compared to the exposure in a honey bee colony. Future studies on varroa mite resistance should consider the difference between laboratory mortality and field efficacy - which has been observed before – carefully.
6.2. Genotyping of the ORβ-2R-L Gene and Potential Links to Amitraz Resistance
The genotyping analyses carried out in 2020 and 2021 included a total of 240 varroa mites from 14 French apiaries in the Eastern, Central, and Southern regions of France. The sequences constituted for all mites were compared to the sequence of the ORβ-2R-L gene referenced in NCBI.
The substitution of A to G at position 260 described by Hernández Rodríguez et al. [24] was identified, with a strong representativeness (70.8%) in all samples analyzed (70.8%), and this whatever their geographic localization and the status of their original colony defined by laboratory sensitivity tests (sensitive, intermediate, or resistant). Nevertheless, we were not able to establish a direct link between the presence of this variation and the development of a resistance to amitraz. Indeed, although many of the surviving mites of laboratory and field bioassays clearly carried the variation, no clear pattern in the distribution of the genotypes between perished and surviving mites was possible. Surviving mites did not carry the variation more often than mites that had died from amitraz exposure. Noteworthy, out of the 170 mites carrying the variation at position 260, 143 (84.12%) were homozygous and 27 (15.88%) were heterozygous. As previously, it was difficult to establish a correlation between these two variated genotypes and a possible resistance to amitraz due to their similar distribution between the surviving and the dead mites.
Genotyping analyses allowed the identification of other, previously undescribed variations in the ORβ-2R-L gene (Appendix IV). Two of them particularly drew our attention: the variations located at positions 344-345 that were detected in the varroa mites analyzed in 2021 and were more frequently present in the “resistant” mites (ratio 19/3 between surviving and dead mites). Involvement of these variations in the resistance development would be inopportune given their low representativity (variations detected in 9.17% of the mites analyzed).
The genome of V. destructor was for long time believed to be particularly stable. Genetic bottleneck events, haplodiploid sex determination and the mite’s sibling system contribute to this low genetic diversity [39]. However, in recent years, several studies have demonstrated that the genetic variation of the varroa mite genome is higher than previously suspected, and that mite populations can differ significantly with respect to certain traits. Varroa destructor has as original obligate host the Asian honey bee, Apis cerana (Hymenoptera; Apidae). In 1952, it switched from Apis cerana to A. mellifera colonies in eastern Russia producing what was named the Korean haplotype of the mite [40]. Another switch event occurred around 1957, leading to the creation of a second haplotype, the Japanese haplotype [40]. Both haplotypes (with a dominance of the Korean haplotype) first colonized the west of their countries of origin [41,42,43,44] and they are now present all around the world. These host switch events did not occur without any genetic changes. Several variants of both haplotypes were identified in different aeras of Asia [44]. A more recent study suggests that the genomic diversity of the populations across the world could be even higher [45]. Whole genome analyses carried out by the authors on 63 V. destructor and V. jacobsoni mites collected in their native ranges from both their ancestral and novel hosts allowed the identification of previously undiscovered mitochondrial lineages on the novel host as well as the equivalent of tens of individuals involved in the initial host switch, demonstrating that modest gene flow remains between mites adapted to their host [45]. Analyses of the fine scale population structure of V. destructor in a managed apiary setting highlighted hierarchical genetic variation between apiaries, between colonies within an apiary and even within colonies [46] Authors also reported a modest increase in the amount of total variation over time within individuals, possibly due to de novo generation of diversity or more probably (due to the short time scales of the study) to horizontal transmission of mites between colonies [46]. It is also now well known that a few populations of A. mellifera can survive Varroa destructor infestation without treatments by means of natural selection. A study was designed in 2016 to investigate potential genetic changes in mite populations from Sweden occurring in response to their host adaptation [47]. Comparison of mites collected in 2009 and 2017-2018 from Varroa-resistant A. mellifera populations and from neighboring mite-susceptible colonies highlighted significant changes in the genetic structure of the mites during the time frame of the study, with more pronounced variations in the V. destructor population collected in the mite-resistant honeybee colonies. This suggests that, like other parasites such as the human lice [48], the human plasmodia or else mammal Leishmania [49], V. destructor adapts to its host adaptation, and reciprocally [47], and this coevolution is under the direct influence environmental factors (climatic conditions, food resources, exposure to pesticides, pollution). The variations that appeared in the genome of V. destructor following treatments with acaricides to develop resistance are perfect examples [24]. Another source of V. destructor genome variability is the possible hybridization with Varroa jacobsoni [50], another Varroa species mostly found within Apis cerana but with less damage compared to V destructor. Sharing 99.7% of their genome in common, both species are very similar genetically [51]. All events detailed above are all arguments that could explain the presence and the geographic specificity of the variation at position 260 (as well as the variations at positions 344-345), in addition to the development of a resistance to amitraz for with no correlation has been clearly established in this study.
7. Conclusions
The present results call into question the hypothesis of a single nucleotide polymorphism as the direct cause of amitraz resistance in varroa mites. While an association between amitraz resistance and the occurrence of the variation at position 260 cannot be completely ruled out, we suspect that such an association would have to include additional steps and perhaps other, interacting variations that would lead to a resistant phenotype.
Further, potential effects of off-label treatments (exposure towards higher doses, longer exposure period than recommended, use of unauthorized treatments etc.) on resistance development should be the focus of future investigations. It is crucial for honey bee health as well as the global beekeeping industry to determine the potential long-term outcomes of off-label use of varroa treatments and raise awareness of the consequences of risking desensitization of varroa mites against present and future treatment options.
Funding
not applicable.
Authors’ Contributions
UM was a major contributor in writing the manuscript, BR performed the genetic analysis, writing materials and methods and part of the results. AD performed and analyzed the genetics. AV analyzed and interpreted the sensibility data (LDA39 expert). MAR performed the study, interpretation and reviewing manuscript. AH participated to reviewing manuscript.
Data Availability Statement
The authors confirm that the data supporting the findings of this study are available within the article and/or its supplementary materials. authors used R to perform the Chi-square tests. RStudio 2023.09.0+463 “Desert Sunflower” Release (b51c81cc303d4b52b010767e5b30438beb904641, 2023-09-25) for windows. Mozilla/5.0 (Windows NT 10.0; Win64; x64) AppleWebKit/537.36 (KHTML, like Gecko) RStudio/2023.09.0+463 Chrome/114.0.5735.289 Electron/25.5.0 Safari/537.36.
Institutional Review Board Statement
not applicable.
Informed Consent Statement
not applicable.
Acknowledgments
We would like to thank Biomnigene for performing genotyping assays and statistical analyses and LDA39 for performing laboratory sensitivity tests. We also appreciate the collaboration of Dr. Alexandre Douablin, Dr. Bénédicte Rognon and Dr. Alain Viry for helping with the writing and paper review.
Conflicts of Interest
no conflicts of interest.
Appendix I. Table Showing the Mites’ Origin, Classification After the Laboratory Assay (Sensitive, Intermediate, or Resistant) as well as Genotyping Results for Varroa Surviving Mite Samples Collected in 2020.
Appendix II. Table Showing the Mites Origin, Classification after the Laboratory Assay (Sensitive, Intermediate, or Resistant) as well as Genotyping Results for Varroa Dead Mite Samples Collected in 2020.
Appendix III. Table Showing the mites’ Origin, Classification after the Laboratory Assay (Sensitive, Intermediate, or Resistant) as well as Genotyping Results for Varroa Mite Samples Collected in 2021.
Appendix IV. Table Reporting the Nature and Positions of the Variations Found in the 135 Sequenced Samples in 2021.
References
- Mondet F, Parejo M, Meixner MD, Costa C, Kryger P, Andonov S, Servin B, Basso B, Bieńkowska M, Bigio G, Căuia E, Cebotari V, Dahle B, Dražić MM, Hatjina F, Kovačić M, Kretavicius J, Lima AS, Panasiuk B, Pinto MA, Uzunov A, Wilde J, Büchler R. Evaluation of Suppressed Mite Reproduction (SMR) Reveals Potential for Varroa Resistance in European Honey Bees (Apis mellifera L.). Insects. 2020 Sep 3;11(9):595. [CrossRef] [PubMed]
- Van Engelsdorp D, Hayes J Jr, Underwood RM, Pettis J. A survey of honey bee colony losses in the U.S., fall 2007 to spring 2008. PLoS One. 2008;3(12):e4071. Epub 2008 Dec 30. [CrossRef] [PubMed]
- Stahlmann-Brown, P.; Hall, R.J.; Pragert, H.; Robertson, T. Varroa Appears to Drive Persistent Increases in New Zealand Colony Losses. Insects 2022, 13, 589. [Google Scholar] [CrossRef] [PubMed]
- Mutinelli F, Pinto A, Barzon L, Toson M. Some Considerations about Winter Colony Losses in Italy According to the Coloss Questionnaire. Insects. 2022 Nov 16;13(11):1059. [CrossRef] [PubMed]
- Richardson, Rodney & Conflitti, Ida & Labuschagne, Renata & Hoover, Shelley & Currie, Rob & Giovenazzo, Pierre & Guarna, M. & Pernal, Stephen & Foster, Leonard & Zayed, Amro. (2023). Land use changes associated with declining honey bee health across temperate North America. Environmental Research Letters. 18. [CrossRef]
- Smoliński S, Langowska A, Glazaczow A. Raised seasonal temperatures reinforce autumn Varroa destructor infestation in honey bee colonies. Sci Rep. 2021 Nov 15;11(1):22256. [CrossRef] [PubMed]
- Jack CJ, de Bem Oliveira I, Kimmel CB and Ellis JD (2023) Seasonal differences in Varroa destructor population growth in western honey bee (Apis mellifera) colonies. Front. Ecol. Evol. 11:1102457. [CrossRef]
- Rose A. McGruddy, Mariana Bulgarella, Antoine Felden, James W. Baty, John Haywood, Philip Stahlmann-Brown, Philip J. Lester. Are increasing honey bee colony losses attributed to Varroa destructor in New Zealand driven by miticide resistance? 2023. bioRxiv. 2023.03.22.533871. [CrossRef]
- Posada-Florez, F., Lamas, Z.S., Hawthorne, D.J. Ryabov, Eugene V.. Pupal cannibalism by worker honey bees contributes to the spread of deformed wing virus. Sci Rep 11, 8989 (2021). [CrossRef]
- Mitton, Giulia & Meroi Arcerito, Facundo & Cooley, Hazel & Fernandez de Landa, Gregorio & Eguaras, Martín & Ruffinengo, Sergio & Maggi, Matías. (2022). More than sixty years living with Varroa destructor: a review of acaricide resistance. International Journal of Pest Management. 1-18. [CrossRef]
- Lodesani, Marco, Mauro Colombo, and M. Spreafico. “Ineffectiveness of Apistan® treatment against the mite Varroa jacobsoni Oud in several districts of Lombardy (Italy).” Apidologie 26.1 (1995): 67-72. [CrossRef]
- Johnson, Reed M., Henry S. Pollock, and May R. Berenbaum. “Synergistic interactionsbetween in-hive miticides in Apis mellifera.” Journal of economic entomology 102.2 (2009): 474-479.
- Patti J. Elzen, Frank A. Eischen, James R. Baxter, Gary W. Elzen, William T. Wilson. “Detection of resistance in US Varroa jacobsoni Oud.(Mesostigmata: Varroidae) to the acaricide fluvalinate.” Apidologie 30.1 (1999): 13-17. [CrossRef]
- Trouiller, Jérôme. “Monitoring Varroa jacobsoni resistance to pyrethroids in western Europe.” Apidologie 29.6 (1998): 537-546.
- Helen M. Thompson, Michael A. Brown, Richard F. Ball, Medwin H. Bew. “First report of Varroa destructor resistance to pyrethroids in the UK.” Apidologie 33.4 (2002) : 357-366. [CrossRef]
- Mitton, G. A., Quintana, S., Giménez Martínez, P., Mendoza, Y., Ramallo, G., Brasesco, C., Villalba, A., Eguaras, M. J., Maggi, M. D., Ruffinengo, S. R. (2016). First record of resistance to flumethrin in a varroa population from Uruguay. Journal of Apicultural Research, 55(5), 422–427. [CrossRef]
- González-Cabrera J, Davies TG, Field LM, Kennedy PJ, Williamson MS. An amino acid substitution (L925V) associated with resistance to pyrethroids in Varroa destructor. PLoS One. 2013 Dec 18;8(12):e82941. [CrossRef] [PubMed]
- González-Cabrera J, Rodríguez-Vargas S, Davies TG, Field LM, Schmehl D, Ellis JD, Krieger K, Williamson MS. Novel Mutations in the Voltage-Gated Sodium Channel of Pyrethroid-Resistant Varroa destructor Populations from the Southeastern USA. PLoS One. 2016 May 18;11(5):e0155332. [CrossRef] [PubMed]
- Millán-Leiva A, Marín Ó, Christmon K, vanEngelsdorp D, González-Cabrera J. Mutations associated with pyrethroid resistance in Varroa mite, a parasite of honey bees, are widespread across the United States. Pest Manag Sci. 2021 Jul;77(7):3241-3249. Epub 2021 Apr 1. [CrossRef] [PubMed]
- Semkiw, Piotr & Skubida, Piotr & Pohorecka, Krystyna. (2013). The Amitraz Strips Efficacy in Control of Varroa Destructor After Many Years Application of Amitraz in Apiaries. Journal of Apicultural Science. 57. 107-121. [CrossRef]
- Higes M, Martín-Hernández R, Hernández-Rodríguez CS, González-Cabrera J. Assessing the resistance to acaricides in Varroa destructor from several Spanish locations. Parasitol Res. 2020 Nov;119(11):3595-3601. Epub 2020 Sep 16. [CrossRef] [PubMed]
- Almecija G, Poirot B, Cochard P, Suppo C. Inventory of Varroa destructor susceptibility to amitraz and tau-fluvalinate in France. Exp Appl Acarol. 2020 Sep;82(1):1-16. Epub 2020 Aug 18. [CrossRef] [PubMed]
- Rinkevich, Frank D. “Detection of amitraz resistance and reduced treatment efficacy in the Varroa Mite, Varroa destructor, within commercial beekeeping operations.” PloS one 15.1 (2020): e0227264.
- Hernández-Rodríguez, C.S., Moreno-Martí, S., Almecija, G., Christmon K., Johnson, J. D., Ventelon, M., vanEngelsdorp, D., Cook, S. C., González-Cabrera, J., Resistance to amitraz in the parasitic honey bee mite Varroa destructor is associated with mutations in the β-adrenergic-like octopamine receptor. J Pest Sci 95, 1179–1195 (2022). [CrossRef]
- Pettis, J. S., H. Shimanuki, and M. F. Feldlaufer. “Detection of varroa resistant to fluvalinate.” Vida Apicola (España) (1999).
- Almecija, Gabrielle. “Résistances de Varroa destructor aux acaricides : conséquences sur l’efficacité des traitements. Application au tau-fluvalinate et à l’amitraze.” (2021).
- Morfin, N., Rawn, D., Petukhova, T., Kozak, P., Eccles, L., Chaput, J., Pasma, T., and Guzman-Novoa, E. 2022. Surveillance of synthetic acaricide efficacy against Varroa destructor in Ontario, Canada. The Canadian Entomologist. [CrossRef]
- Milani, Norberto & Vedova, Giorgio. Decline in the proportion of mites resistant to fluvalinate in a population of Varroa destructor not treated with pyrethroids. (2002). Apidologie 33(4):417-422.
- Elzen, Patti & Westervelt, David. A scientific note on reversion of fluvalinate resistance to a degree of susceptibility in Varroa destructor. (2004). Apidologie 35(5):519-520.
- Dietemann, V; Ellis, J D; Neumann, P. EDS. The COLOSS BEEBOOK, Volume I: standard methods for Apis mellifera research. International Bee Research Association; Cardiff, UK. 636 pp. ISBN 978-0-86098-274-6 (2013).
- Milani, N. The resistance of Varroa jacobsoni Oud. To pyrethroids – a laboratory assay. Apidologie 26 : 415–429 (1995).
- Sandon Y., Viry, A. Lutte contre le Varroa – Etude de sensibilité/résistance à l’amitraze chez Varroa destructor – La Santé de l’Abeille N°277: 47-56 (2017).
- Giles, David P., Rothwell David N. The sub-lethal activity of amidines on insects and acarids. Pesticide Science 14, (3), 303–312 (1983).
- Evans P.D., Gee J.D. Action of formamidine pesticides on octopamine receptors. Nature 1980; 287 (5777): 60-62. [CrossRef]
- Alban Maisonnasse, J. Hernandez, C. Le Quintec, Marianne Cousin, Constance Beri, et al. Evaluation de la structure des colonies d’abeilles, création et utilisation de la méthode ColEval (Colony Evaluation). Innovations Agronomiques, 2016, 53, pp.27-37.
- Observatoire de la production de miel et de gelée royale - Edition juillet 2022 © FranceAgriMer 2022.
- Guideline on veterinary medicinal products controlling Varroa destructor parasitosis in bees EMA/CVMP/EWP/459883/2008-Rev.1.
- Rapport d’étude Evaluation de l’efficacité du traitement Apivar® en lien avec la sensibilité des populations de varroas à l’amitraze (2021). (Microsoft Word - Rapport_EfficacitéApivar2021_v04082022 (adaoccitanie.org)).
- Rosenkranz P, Aumeier P, Ziegelmann B. Biology and control of Varroa destructor. J Invertebr Pathol. 2010; 103:S96–S119. [PubMed: 19909970].
- Oldroyd, B. P. 1999. Coevolution while you wait: Varroa jacobsoni, a new parasite of western honeybees. Trends Ecol. Evol. 14: 312–315.
- Ayan, A., O. S. Aldemır, and Z. Selamoglu. 2017. Analysis of COI gene region of Varroa destructor in honey bees (Apis mellifera) in province of Siirt. Turkish J. Vet. Res. 1: 20–23.
- Hajializadeh, Z., M. Asadi, and H. Kavousi. 2018. First report of Varroa genotype in western Asia based on genotype identification of Iranian Varroa destructor populations (Mesostigmata: Varroidae) using RAPD marker. Syst. Appl. Acarol. 23: 199–205.
- Ogihara, M. H., M. Yoshiyama, N. Morimoto, and K. Kimura. 2020. Dominant honeybee colony infestation by Varroa destructor (Acari: Varroidae) K haplotype in Japan. Appl. Entomol. Zool. 55: 189–197.
- Navajas, M., D. L. Anderson, L. I. De Guzman, Z. Y. Huang, J. Clement, T. Zhou, and Y. Le Conte. 2010. New Asian types of Varroa destructor: a potential new threat for world apiculture. Apidologie. 41: 181–193.
- Techer, M. A., J. M. K. Roberts, R. A. Cartwright, and A. S. Mikheyev. 2021. The first steps toward a global pandemic: Reconstructing the demographic history of parasite host switches in its native range. Mol Ecol. [CrossRef]
- Dynes TL, et al. Fine scale population genetic structure of Varroa destructor, an ectoparasitic mite of the honey bee (Apis mellifera) Apidologie. 2017;48:93–101. [CrossRef]
- Beaurepaire A.L., Moro A., Mondet F., Le Conte Y., Neumann P., Locke B. Population genetics of ectoparasitic mites suggest arms race with honeybee hosts. Sci. Rep. 2019;9:11355. [CrossRef]
- Ascunce, M. S., Toups, M. A., Kassu, G., Fane, J., Scholl, K., & Reed, D. L. (2013). Nuclear genetic diversity in human lice (Pediculus humanus) reveals continental differences and high inbreeding among worldwide populations. PLoS One, 8(2), e57619. [CrossRef]
- Nozais J.P. The origin and dispersion of human parasitic diseases in the old world (Africa, Europe and Madagascar) Mem Inst Oswaldo Cruz. 2003;98(Suppl. I):13–19.
- Dietemann, V. , Beaurepaire, A. , Page, P. , Yañez, O. , Buawangpong, N. , Chantawannakul, P. , & Neumann, P. (2019). Population genetics of ectoparasitic mites Varroa spp. in Eastern and Western honey bees. Parasitology, 146, 1429–1439.
- Techer, M. A., R. V. Rane, M. L. Grau, J. M. K. Roberts, S. T. Sullivan, I. Liachko, A. K. Childers, J. D. Evans, and A. S. Mikheyev. 2019. Divergent evolutionary trajectories following speciation in two ectoparasitic honey bee mites. Commun. Biol. 2: 357.
Figure 1.
Time schedule of Apivar® application in the field in the late summer of 2021.
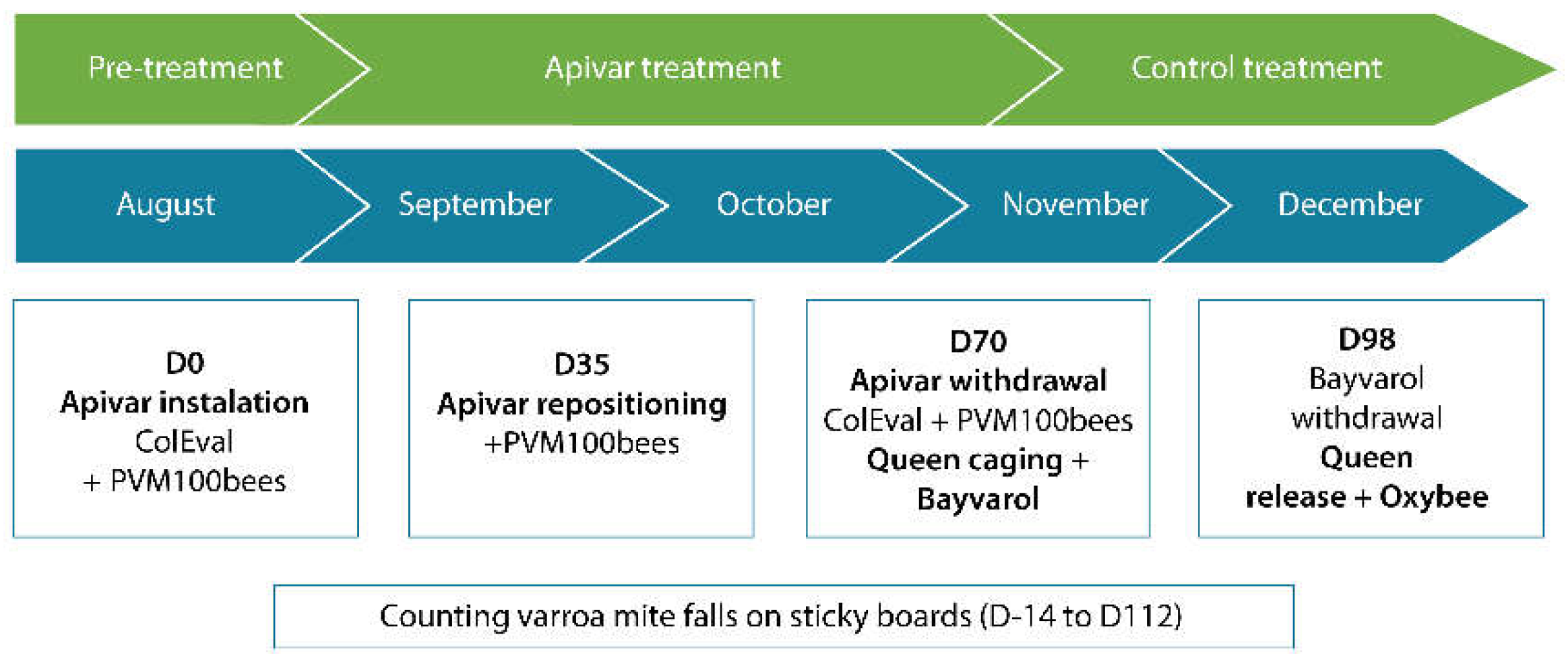
Figure 3.
Partitioning of genotypes of varroa mite samples collected in 2021 after mites exposure towards the LC90 of amitraz in the laboratory.
Figure 3.
Partitioning of genotypes of varroa mite samples collected in 2021 after mites exposure towards the LC90 of amitraz in the laboratory.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



