Submitted:
23 April 2024
Posted:
24 April 2024
You are already at the latest version
Abstract
We have studied the inheritance of mitochondria in Pelargonium section Ciconium using interspecific crosses. We generated 36 different interspecific crosses. We designed KAPS markers targeting four individual mitochondrial markers, belonging to the four main crossing parents, that discriminate between an individual species and the other section Ciconium species. We found that maternal inheritance of mitochondria is most frequent, but we find occasional occurrences of paternal inheritance, while biparental inheritance is rare. For the P. multibracteatum crossing series we found ambiguous results. Our results confirm what was found before, namely that paternal inheritance of mitochondria can occur in P. sect Ciconium but that the instance is rare and much less common than is the case for chloroplasts.
Keywords:
Pelargonium
; mitochondrion
; inheritance
; CNI
; Ciconium
1. Introduction
Species from genus Pelargonium (Geraniaceae) have been used regularly as a model organism for studying Cyto-Nuclear Incompatibility (CNI), e.g., [1,2,3,4,5,6,7]. The species of the genus are renowned for being relatively easy to cross, at least on the intrasectional level and CNI between interspecific hybrids is a common occurrence [5,6,7]. Cyto-Nuclear Incompatibility is the (partial) failure or breakdown in communication between nuclear and organellar genomes. It occurs when populations, derived from a single ancestor, and having become separated in space and time, undergo secondary contact. Such populations may have acquired mutations independently from each other, creating possible reproductive barriers. This is referred to as the Bateson-Dobzhansky-Muller (BDM) model of speciation [8,9,10] and is thought to underly and explain the occurrence of CNI. Cyto-nuclear incompatibility can be caused by nuclear mismatch with mitochondria (mCNI) as well as with chloroplasts (pCNI).Whereas mCNI manifests itself as dwarf growth or (partial) male sterility [11], pCNI on the other hand, presents itself as bleaching of the leaves (chlorosis), which is a regularly occurring phenomenon in F1 hybrids of interspecific crosses in Pelargonium [Geraniaceae; 1,5,6]. Both types of CNI seem to occur in Pelargonium crosses, but pCNI has been studied in far greater detail [1,2,3,4,5,6,7,12,13] than the inheritance of mitochondria and mCNI. However, tantalizing evidence of possible biparental/paternal inheritance of mitochondria in Pelargonium has been reported [13,14,15]. These observations can be explained by direct inheritance of mitochondria, but recombination of mitomes at some point has also been invoked as an explanation for the apparent heteroplasmy of hybrid offspring [16]. In this paper heteroplasmy defined as the occurrence of two or more genotypes in one organism or even individual cell and which can, but not necessarily do, result in different CNI phenotypes.
Recombination can occur during cell division if conditions allow see [17] (meiosis or mitosis). It could also occur during gamete fusion when pollen and embryo content merge as mitochondria (and plastids) were demonstrated to be physically transmitted by pollen, as are plastids [14]. Both methods could be ‘common’ and allow for recombination to occur each time a cell divides or gametes fuse. Recombination could also have occurred historically (on any time scale) and have been a ‘on off event’ if neutral or even beneficial (under specific circumstances). This could have resulted in fixation of the variants and effectively result in as we define “stable heteroplasmy”). Another explanation for observed heteroplasmy is intracellular gene transfer between organelles, as was demonstrated to occur in Geranium [18], which is a sister genus of Pelargonium. If there are selective penalties or benefits to heteroplasmy any of these scenarios could have been played a role in heteroplasmy becoming stable or recurring in Pelargonium at some point during the evolution of a lineage or population. Here, we take advantage of the great number of hybrids generated by our previous studies [6,7] to verify the inheritance of mitochondria in Pelargonium section Ciconium interspecific hybrids.
2. Materials and Methods
2.1. Plant material, DNA Extraction and Sequencing
The list of plant material, DNA extraction protocol, and standard Illumina HiSeq sequencing protocol are the same as those reported in [7]. Additional plant material was collected from herbaria and from living collections (Table 1, copied with permission from [19] and these were subjected to the same treatment with respect to DNA extraction and sequencing. For the sake of convenience, throughout the text four letter acronyms for each accession will be used. The corresponding species names and their acronyms can be found in Table 1. A total of 36 different F1 crosses consisting of 179 accessions were evaluated and of these 12 were wildtype plants (i.e., parental material). Resulting in a total of 163 F1 accessions for evaluation.
2.2. Mitome Assembly
From the Illumina sequence read libraries we assembled mitome fragments. We used getOrganelle v1.6.2e [20] and started assembly with known mitochondrial fragments available for Pelargonium [21,22]. After a first round of assembly, we evaluated the results, which we then took and used, as a more specific target to start our next fragment assembly on. We accepted an assembly with a base coverage >10 reads/base position as ‘good’. Although for practical purposes we sometimes accepted an assembly with a base coverage of >5<10. We then only focused on the exons as it was not possible to achieve homologous noncoding sequences for all accessions.
2.3. Organellar Genotyping Using PCR and KASP Markers
We employed a Competitive Allele Specific PCR (currently named ‘Kompetitive Allele Specific PCR’, or KASPTM) from Kbioscience or LGC Genomics (http://www.lgcgenomics.com) for mitotyping. KASP was proven to be a reliable and fast technique for genotyping material and is now considered a benchmark technique for SNP calling [23,24,25,26]. KASP is a PCR-based assay designed to detect SNP variants by using two forward primers (containing the SNP) and one reverse primer. Our F1 and F2 progeny had two parents that differed by SNPs at various positions. The mother of each crossing series was considered the ‘target’ and the paternal parent can be any other species from section Ciconium and is therefore referred to as ‘Ciconium’ in our assays. KASP primers were designed on (partial) assemblies of mitome exons as outlined above. Using introns for marker development is standard, as these are generally more variable yet close to conserved exon sequences. But, in the case of Pelargonium, mitome introns have been found to be absent from nad1 [22,27,28,29] but instead that exon silent nucleotide substitution rates were found to be significantly increased [21,27,29,30] and therefore may be good candidates for SNP marker development. The following exons were used as a source: cox2, cox3, cytb, NAD1-exon 1, nad7, NAD5 and atp1. We selected SNPs unique to a target, if possible, to be able to distinguish it from the other Ciconium species. We did this by comparing the sequences, obtained from the mitome assembly procedure described above, in separate alignments and using the SNP’s as targets for the KASP primers. Our KASP thermo profile was as follows: 5 minutes denaturation at 94°C, 10 cycles of 94°C 20s, 61-55°C annealing and extension for 60s (dropping 0.6°C for the annealing temperature each cycle), followed by 30 cycles of 94°C and 55°C extension for final amplification. We added 10µM F1 primer + 10µM F2 primer and 20µM R primer.. These were mixed with the KASP 2x master mix (LGC Genomics. (2013) and mQ water. Template DNA concentrations must be in the range of 0.2-10 ng/µL and these need to be added to the mix per each reaction. For KASP we used two positive control samples which contained only the maternal and paternal genotype, and non-template controls (NTC’) to be able to discriminate fluorescence signal caused by primer interactions from those with the templates. As stated above, KASP is a PCR-based technique that can determine the relative concentration of each genotype as well. In order to test if there are differences in concentrations between alleles, we mixed DNA extracts of one parent with a ‘Ciconium’ allele (set to 0,2 ng/µL) with the extracts of a sample with the target (e.g., ‘HORT’) allele to obtain series of known concentration ratios (10:90, 25:75, 50:50, 75:25 & 90:10.) This allowed us to determine if there were different ratios of mitochondria of either parent present in the hybrid accessions and thus enable quantification after 30 cycles. Because markers were designed for four different targets, hybrids based on two targeted parents could be reciprocally evaluated and we have done so for offspring that was the result of: ‘MULT, ACET, BARK and HORT’ crosses. The full results are visualized in Figure 1 as allelic discrimination plots. Mitome type calling was done as follows: A plant can have a maternally (M), paternally (P), biparental (B) or conflicting mitome type (B/H/R). The last category can occur if the parental plants happen to be heteroplasmic themselves or if the mitomes are recombining, effectively resulting in a heteroplasmic offspring. The KASP primers can be found in OSM.
3. Results
3.1. Confirmed F1 Hybrids
The resulting hybrids are described in our previous studies [6,7,19], but a short summary is given here for convenience. These studies established a total of 30 different F1 interspecific hybrids and hundreds of plants. The majority of these came from two (nearly) comprehensive attempts of crossing series with P. x hortorum (‘Pinto White’ diploid cultivar) X Ciconium spp. and P. multibracteatum X Ciconium spp. Other successful F1 interspecific hybrids series were from P. barklyi X Ciconium spp. and P. acetosum X Ciconium spp. Thus, a wide range of crosses were established covering four clades from the section, as estimated based on plastome comparison by [31].
3.2. Mitome Assemblies
We were able to assemble mitome fragments with a read depth >10 for most accessions (see OMS1). For some accessions however, markers were assembled with a base coverage of 5-10 and these are considered moderately reliable. These moderately reliable sequences were incorporated for practical and comparative purposes, but not used for primer design. Our final dataset, only counting the concatenated exons, was 4,353 bp long and the assembly lengths for each fragment were: COX2: 687 bp; COX3: 1028 bp; cytb: 487 bp; nad1 exon 1: 311 bp; nad5: 502 bp; nad7: 582 bp and atp1: 756 bp. Length differences were observed in nad1 exon 1 and nad5 (OSM with alignments). For P. yemenense, the cytb sequence remained incomplete despite repeated attempts at assembly.
3.3. Mitotyping
We have successfully determined the mitome type of 179 plants. All results are summarized in Table 2 and visualized in Figure 1. Excluding the MULT X Ciconium series (see below) and the wildtype plants (the F0 parental populations), there were 117 F1 plants in total. Of these, 89% displayed the maternal mitotype, 8.5% the paternal type, 0.8% (one plant) displayed a conflicting signal, but this involved a ‘bridge cross’ in which an F1 plant was crossed with a third species (an F1 HORT x ZONA plant was crossed with P. aridum Table 2), and 0.8% (one plant) displayed evidence of biparental inheritance (F1 TONG X ACET). This last example was confirmed by a second reaction with another KASP marker (shown in Table 2) confirming the result. Finally, Four F1 plants failed to yield any result. For the P. multibracteatum series, 10 F1 crosses (55 plants) were evaluated. Of these, two crosses displayed apparent biparental genotypes, eight others displayed an apparent paternal mitotype. We tested the wildtype plants with all markers available and these displayed conflicting results (biparental/heterozygous).
Mito-typing results are visualised as KASP allelic discriminant plots in Figure 1. Plants with predominantly maternal mitotypes with a non-target Ciconium mother and a target Ciconium father (e.g., FRUT x BARK) nearly always ended up in quadrant I (Figure 1 A-C) as expected and those with the target as mother and non-target Ciconium as father in quadrant IV (Figure 1 A-C). One population (F1 HORT x QUIN) displayed discrete bi-parental inheritance with plants displaying either the QUIN (paternal) or HORT (maternal) genotype. One accession, (F1 of TONG x ACET), has a KASP result close to the non-target ‘Ciconium’: target ‘acetosum’ ratio of 25:75 (indicated by a star in Figure 1B). In this single case there is more of the paternal type (~75%), but the maternal type is also present. This plant appears to be the exception to the rule and may be one of those rare individuals which show heteroplasmy at significant levels for mitochondrial types. Three F1 samples show a strictly paternal genotype: These are F1 BARK x QUIN, F1 ACET x ZONA and F1 BARK x INQU. Two F2 plants of HORT x ACET also displayed a paternal (ACET) genotype
Figure 1. KASP allelic discrimination plots.
4. Discussion
Our results support the long-held notion that mitochondria inherit both paternally and maternally in Pelargonium section Ciconium [7,13,14,15]. If we exclude the P. multibracteatum population and its derived hybrids, the instance of heteroplasmy in hybrid offspring seems to be less common than in the case of plastids, for which heteroplasmy was found more frequently ]6,7]. Our results do point to maternal inheritance as the prevailing mode of inheritance. This phenomenon has intrigued people since long. And it has been considered the result of some imperfect ‘sorting mechanism’ in which paternal leakage is occurring at low frequency [13,32] preferentially selecting a certain type. For a review of proposed mechanisms see [33], or for an example [17]. We hypothesize that the high incidence of maternal inheritance of mitomes means that, even if these mechanisms are in place in P. sect Ciconium, they do not function perfectly and the amount of paternally inherited mitochondria is more than mere ‘parental leakage’. These studies mainly target plastid inheritance and plastome variation, but the same or similar mechanisms may underly mitochondrial inheritance and selection as well [13]. Another explanation for the stronger maternal effect could be that mCNI is much stronger than chloroplast effects, especially because our controlled growth environment is optimized for the survival of every chlorotic plant, but not for embryos. Even though we did employ embryo rescue, this does not salvage any CNI effects that would occur directly upon fertilization. Plastids are undeveloped during and directly after fertilization and seed development, whereas the mitochondria are active during these phases. So, any mCNI effect would be stronger than pCNI at these crucial early developmental stages. This would explain the high number of aborted embryos and empty seeds found on all our F1 plants [6]. Given that the mother plant is ‘responsible’ for supplying energy to the development of the seeds, it is logical that there is a strong maternal bias. However, plastids which are introduced to the embryo via the pollen [14,34]) are sorted out and removed early in development as well [13,32]. Nevertheless, they can be present in all tissue early on [15] before most of them are removed eventually.
Finally, recombination has also been invoked as a, partial, explanation for observed patterns of heteroplasmy and cannot be excluded to play a role as well [16,35]. Basically, recombination is thought to affect occurrence of varying mitome types in a cell or plant and occasionally male sterile phenotypes and result in a more random distribution of mitome variants. We think that this explanation is less likely as this would have been a more rare occurrence. The relatively high occurrence of both paternal and maternal (and even bi-parental) modes inheritance across the section as well as persistence throughout generations (see [6] for an example from plastids), evidenced by the recovery of our SNP markers in offspring, are easier explained by simple inheritance.
The Case of P. multibracteatum
The P. multibracteatum crossing series showed ambiguous results. The reciprocal comparison of the results for the other markers with the MULT marker were contradictory (see Table 2 & OSM1). The subsequent reciprocal comparison between the other markers (ACET, BARK and HORT) clearly showed that these three confirmed each other, leaving us with the MULT markers’ ambiguous results. To assess performance issues of the MULT KASP marker, we re-analyzed the P. multibracteatum series with a second KASP marker and we included another P. multibracteatum accession (not shown). This again resulted in a near universal ‘Ciconium’ call and this time the positive control also resulted in a ‘Ciconium’ call. We therefore conclude that we do not have consistent results for the mito-typing of the P. multibracteatum series. Rather, we believe that the patterns we see is caused by innate heteroplasmy of P. multibracteatum). The full overview of all reactions and raw data fluorophore measurements can be found in the supporting materials.
Evolutionary Effects of mCNI
Our results lend support for both the idea that bi-parental inheritance of organelles could provide an escape from CNI [36] and the hypothesis that organellar changes, resulting in CNI, have a profound influence of speciation ([37,38]. Further support for these two hypotheses comes the fact that second generation of plants segregate for chlorosis [6,7] with only one plastid type present, showing that selection for organelle management and expression genes acts immediately after the first generation of hybridization [6,37]. Different organelle types induce different CNI in crosses with equal nuclear genomic backgrounds (as can be seen for the chloroplast in P. x hortorum crosses published in [7]). The preference for one type, as well as preferentially backcrossing with one of the parents (introgression), after a historical hybridization event could explain the problematic position of taxa in phylogenetic trees due to conflict between plastid and nuclear genomic markers. For instance, the four-petalled P. nanum which is currently not assigned to a section [39] was suspected to be an ancient relict of a now extinct group (section) of species because of the unique floral morphology and its ‘single branch’ status in current phylogenetic trees (e.g., [31]) usually grouping as a sister to clade A2, based on the clade systematics established by [40]. Other cases can be seen in P. sect. Hoarea where the occurrence of non-monophyletic species has been attributed to ‘chloroplast capture’ [41]. Such taxa would have retained the organelle of one species, while displaying the morphology and nuclear genomic type of another. Further testing of such incongruencies could be done by using more markers from the nuclear genomes. For instance, the repeatome appears promising as a source of phylogenetic markers [42,43]) as it provides resolution at a low taxonomic level and provides a genome-wide overview represented by the most abundant parts of the non-coding DNA (repeats).
Naturally occurring hybrids in Pelargonium are rarely found, (pers. Comm. Powrie Kirstenbosch RSA), but not unheard off [44,45]. This is logical given the reduced fitness characteristic of most hybrid offspring which will result in lower chances of surviving to the reproductive life stage. However, despite this post-zygotic barrier our study also shows that species can be highly compatible as we obtained many (~30) interspecific crosses, some of which are fully green and fertile. Therefore, we do not exclude that hybridization does play an additional, if minor, role in Pelargonium evolution. Two cases of possible natural hybrids from section Ciconium are known. The first is an herbarium specimen of a wild hybrid between P. peltatum and P. alchemilloides at RBGE (M. Gibby pers. comm.). The second case is P. x salmoneum (from our own collections). We have analyzed the plastome and mitome of P. x salmoneum and found that it carries the P. inquinans plastome and a mix of P. inquinans and P. acetosum mitochondrial genotypes (OSM1), potentially due to mitome recombination [16] or historical biparental inheritance. The morphology of P. x salmoneum is intermediate between P. inquinans and P. acetosum. Pelargonium x salmoneum is a fully fertile, green plant which segregates for numerous traits such as plants size (an indication of possible mitochondrial CNI effects), flower shape, leaf shape indicating it is not a ‘stable’ species (yet), but a hybrid. We propose that P. x salmoneum, irrespective of whether it arose naturally or was the result of human crossing activities, is a genuine interspecific hybrid with equal fitness to either of its proposed parents and that it contains possible traces of mitochondrial recombination. We hypothesize that embryonic organelle sorting is absent or impaired in Pelargonium section Ciconium and conclude that the sorting of mitochondria is stronger than that of plastids. Heteroplasmy is rare for mitochondria, in all P. sect. Ciconium species, but seems to be innate to the P. multibracteatum population used in this study. This may imply that selective barriers are stronger for mitochondria than for chloroplasts.
Supplementary Materials
The following supporting information can be downloaded at website of this paper posted on Preprints.org, OSM1: Mitome fragment assembly results. Plant lists with mitotypes per plant and KASP primers. OSM2: Fasta alignments of mitome fragments.
Author Contributions
Conceived the study: FCB, FTB, MES. Wrote the manuscript: FCB, FTB. Carried out the analysis: FCB. Peptide analysis FCB, CV. Experimental design PCR: FCB, FTB, rpo typing: FCB, FTB. Experimental design crossing and embryo rescue: FCB, RCS, MES, MS-S. Experimental design PCR: FCB, JWK. Laboratory work: FCB, JWK. All authors read the draft and gave feedback. This research was funded by the Dutch Foundations for applied scientific research (TTW). Grant number: 14531 “Pelargonium genomics for overcoming cytonuclear incompatibility and bridging species barriers” of the Green Genetics program NOW. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
This research was funded by the Dutch Foundations for applied scientific research (TTW). Grant number: 14531 “Pelargonium genomics for overcoming cytonuclear incompatibility and bridging species barriers” of the Green Genetics program
Data Availability Statement
SRA’s on which the mitome fragments are based have been published as part of the PhD theis by FCB entitled: “Exploring patterns of cytonuclear incompatibility in Pelargonium section Ciconium”
Acknowledgments
The crews at Syngenta for caring for the plants. Tony Lokkers (Syngenta) for pollinations and Mireia Sancho-Such (Syngenta) for the embryo rescue.
Conflicts of Interest
RCS is employed at Syngenta NV the other authors have no conflicting interests to declare.
References
- Baur E. Das Wesen und die Earblichkeitsverhältnisse der ‘varietates albomarginatae hort’ von Pelargonium zonale. Z. Indukt. Abstammungs-Vererbungsl 1909, 1, 330–351. [CrossRef]
- Tilney-Bassett, R. A. E. The control of plastid inheritance in Pelargonium II. Heredity 1973, 30, 1–13. [CrossRef]
- Tilney-Bassett, R. A. E. The control of plastid inheritance in Pelargonium III. Heredity 1974, 33, 353–360. [CrossRef]
- Tilney-Bassett, R. A. E. “Genetics of variegated plants,” in Genetics and biogenesis of mitochondria and chloroplasts. Eds. C. W.Birky P. S Perlman, and T. J Byers (Columbus: Ohio State University Press), 1975, 268–308.
- Horn, W. Interspecific crossability and inheritance in Pelargonium. Plant Breeding 1994, 113, 3–17. [CrossRef]
- Breman, F.C.; Snijder, R.C.; Korver, J.W.; Pelzer, S.; Sancho-Such, M.; Schranz, M.E.; Bakker, F.T. Interspecific Hybrids Between Pelargonium × hortorum and species From P. section Ciconium reveal biparental plastid inheritance and multi-locus cyto-nuclear incompatibility. Front. Plant Sci. 2020, 11:614871. [CrossRef]
- Breman, F.C. Exploring patterns of cytonuclear incompatibility in Pelargonium section Ciconium. PhD thesis, Wageningen UR, the Netherlands; 2021. [CrossRef]
- Bateson, W. Heredity and variation in modern lights. In Darwin and modern science (ed. A. C. Seward), pp. 85– 101. Cambridge, UK: 1909. Cambridge University Press.
- Dobzhansky T. Studies on hybrid sterility. II. Localization of sterility factors in Drosophila pseudoobscura hybrids. Genetics 1936, 21, 113–135.
- Müller, HJ. Isolating mechanisms, evolution, and temperature. Biol. Symp. 1942, 6, 71–125.
- Schnable, P.S.; Wise, R.P. The molecular basis of cytoplasmic male sterility and fertility restoration. Trends Plant Sci 1998, 3:175–180. [CrossRef]
- Tilney-Bassett, R.A.E.; Almouslem, A.B.; Amoate, H.M. Complementary genes control biparental plastid inheritance in Pelargonium. Theor. Appl. Genet. 1992, 85:317-324. [CrossRef]
- Weihe, A.; Apitz, J.; Salinas, A.; Pohlheim, F.; Börner, T. Biparental inheritance of plastidial and mitochondrial DNA and hybrid variegation in Pelargonium. Mol. Genet. Genomics 2009, 282:587–593. [CrossRef]
- Kuroiwa, T.; Kawazu, T.; Ushida, H.; Ohta, T. Kuroiwa H. Direct evidence of plastid DNA and mitochondrial DNA in sperm cells in relation to biparental inheritance of organelle DNA in Pelargonium zonale by fluorescence/electron microscopy. Eur. J. Cell Biol. 1993, 62:307–313.
- Guo, F.L.; Hu, S.Y. Cytological evidence of biparental inheritance of plastids and mitochondria in Pelargonium. Protoplasma. 1995, 186:201-207. [CrossRef]
- Apitz, J.; Weihe, A.; Pohlheim, F.; Börner, T. Biparental inheritance of organelles in Pelargonium: evidence for intergenomic recombination of mitochondrial DNA. Planta 2013, 237:509–515. [CrossRef]
- Sobanski, J.; Giavalisco, P.; Fischer, A.; Kreiner, J.M.; Walther, D.; Schöttler, M.A.; Pellizzer, T.; Golczyk, H.; Obata, T.; Bock, R.; Sears, B.B.; Greiner, S. Chloroplast competition is controlled by lipid biosynthesis in evening primroses. PNAS 2019 Doi: www.pnas.org/cgi/doi/10.1073/pnas.1811661116.
- Park, S.; Grewe, F.; Zhu, A.; et al. Dynamic evolution of Geranium mitochondrial genomes through multiple horizontal and intracellular gene transfers. The New Phytologist. 2015, 208(2):570-583. [CrossRef]
- Breman, F.C.; Korver, J.W.; Snijder, R.C.; Bakker, F.T. Plastid-encoded RNA polymerase variation in Pelargonium sect. Ciconium. HORTIC. ADV. 2024, 2. 1. [CrossRef]
- Jin, J.J.; Yu, W.B.; Yang, J.B.; et al. GetOrganelle: a fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [CrossRef]
- Bakker, F.T.; Breman, F.C.; Merckx, V. DNA sequence evolution in fast evolving mitochondrial DNA nad1 exons in Geraniaceae and Plantaginaceae. Taxon 2006, 55:887–896. [CrossRef]
- Grewe, F.; Zhu, A.; Mower, J.P. Loss of a trans-splicing nad1 intron from Geraniaceae and transfer of the maturase gene matR to the nucleus in Pelargonium. Genome Biol Evol. 2016, 30;8(10):3193-3201. [CrossRef]
- Hiremath, P.J.; Kumar, A.; Penmetsa, R.V.; Farmer, A.; Schlueter, J.A.; et al. Large-scale development of cost- effective SNP marker assays for diversity assessment and genetic mapping in chickpea and comparative mapping in legumes. Plant Biotechnol. J. 2012, 10:716–32. [CrossRef]
- Semagn, K.; Babu, R.; Hearne, S. Single nucleotide polymorphism genotyping using Kompetitive Allele Specific PCR (KASP): Over view of the technology and its application in crop improvement. Mol. Breed. 2014, 33:1–14. [CrossRef]
- Islam, M.S.; Thyssen, G.N.; Jenkins, J.N.; Fang, D.D. Detection, validation, and application of genotyping-by-sequencing based single nucleotide polymorphisms in upland Cotton. Plant Genome 2014, 8:1–10. [CrossRef]
- Ka S, Quinton-Tulloch MJ, Amgai RB, et al. Accelerating public sector rice breeding with high-density KASP markers derived from whole genome sequencing of indica rice. Mol. Breed. 2018, 38:38. [CrossRef]
- Bakker F.T.; Culham A.; Pankhurst C.E.; Gibby M. Mitochondrial and chloroplast DNA-based phylogeny of Pelargonium (Geraniaceae). Amer. J. Bot. 2000, 87: 727– 734. [CrossRef]
- Palmer, J.D.; Adams, K.L.; Cho, Y.; Parkinson, C.L.; Qiu, Y.L.; Song, K. Dynamic evolution of plant mitochondrial genomes: mobile genes and introns and highly variable mutation rates. Proc. Natl. Acad. Sci. U.S.A. 2000, 97: 6960–6966. [CrossRef]
- Choi K.; Weng M-L.; Ruhlman T.A.; Jansen R.K. Extensive variation in nucleotide substitution rate and gene/intron loss in mitochondrial genomes of Pelargonium. Mol. Phyl. and Evol. 2021, 155, (106986). [CrossRef]
- Mower J.P.; Touzet P.; Gummow J.S.; Delph L.F.; Palmer J.D.; Extensive variation in synonymous substitution rates in mitochondrial genes of seed plants. BMC Evol. Biol. 2007, 7:135. [CrossRef]
- Kerke, van de S.J.; Shrestha B.; Ruhlman T.A.; Weng M-L.; Jansen R.K.; Jones C.S.; Schlichting C.D.; et al. Plastome based phylogenetics and younger crown node age in Pelargonium. Mol. Phyl. and Evol. 2019, 137:33–43. [CrossRef]
- Kirk J.T.O.; Tilney-Bassett R.A.E. The plastids. Freeman and Co., London 1967, United Kingdom.
- Greiner S.; Sobanski J.; Bock R. Why are most organelle genomes transmitted maternally? Bioessays 2015, 37, 80–94. [CrossRef]
- Kuroiwa, H.; Kuroiwa, T. Giant mitochondria in the mature egg cell of Pelargonium zonale. Protoplasma 1992, 168, 184–188. [CrossRef]
- Maréchal A.; Brisson N. Recombination and the maintenance of plant organelle genome stability. New Phytol. 2010, 186: 299–317.
- Barnard-Kubow, K.B.; McCoy, M.A.; Galloway, L.F. Biparental chloroplast inheritance leads to rescue from cytonuclear incompatibility. New Phytol. 2017, 213(3):1466-1476. [CrossRef]
- Barnard-Kubow, K.B.; So, N.; Galloway, L.F. Cytonuclear incompatibility contributes to the early stages of speciation. Evolution 2016, 70(12):2752-2766. [CrossRef]
- Greiner, S.; Bock, R. Tuning a ménage à trois: Co-evolution and co-adaptation of nuclear and organellar genomes in plants. Bioessays 2013, 35: 354–365. [CrossRef]
- Röschenbleck, J.; Albers, F.; Müller, K.; Weinl, S.; Kudla, J. Phylogenetics, character evolution and a subgeneric revision of the genus Pelargonium (Geraniaceae). Phytotaxa 2014, 159(2), 31–76. [CrossRef]
- Bakker, F.T.; Culham, A.; Daugherty, L.C.; Gibby, M. A trnL-F based phylogeny for species of (Geraniaceae) with small chromosomes in Pelargonium. Plant Species Biol. 1999, 216, 309–324.
- Bakker F.T.; A. Culham A.; Marais E.M.; Gibby M. Nested radiation in Cape Pelargonium. Pp. 75–100 in Bakker FT, Chartrou LW, Gravendeel B, Pielser PB, eds. Plant species-level systematics: new perspectives on pattern and process. A. R. G. Ganter Verlag K. G., Ruggell, Liechtstein 2005.
- Dodsworth S.; Jang T.S.; Struebig M.; Chase M.W.; Weiss-Schneeweiss H.; et al. Genome-wide repeat dynamics reflect phylogenetic distance in closely related allotetraploid Nicotiana (Solanaceae). Pl. Syst. and Evol. 2017. [CrossRef]
- Breman, F.C.; Chen G.; Snijder R.C.; Schranz M.E.; F.T. Repeatome-Based Phylogenetics in Pelargonium Section Ciconium (Sweet) Harvey, Genome Biol. and Evol., 2021, 13(12), evab269. [CrossRef]
- Knuth R. Geraniaceae. - In Encler A., (Ed.): Das Pflanzenreich 4, 1–9. – Leipzig (1912), Germany Engelmann.
- Van der Walt J.J.A.; Albers F.; Gibby M. Delimitation of Pelargonium sect. Glaucophyllum. PI. Syst. Evol. 1990, 171: 15—26. [CrossRef]
Figure 1.
KASP allelic discrimination plots for F1 hybrid accessions from crosses involving P. x hortorum (A) P. acetosum (B), P. barklyi (C) and P. multibracteatum (D)F1 hybrid samples. Each data point represents the fluorescence signal of an individual DNA sample, with the x-axis indicating the amount of FAM fluorescence and the y-axis the HEX fluorescence for each sample. Negative control samples are expected to be placed around the origin of the plot. Samples of the same mitotype will have generated similar levels of fluorescence and will therefore cluster together on the plot. Each graph is divided into four quadrants (I-IV). Here, the y-axis displays the values assigned to the non-target Ciconium genotypes. Hence quadrant I holds the non-target Ciconium genotypes. Quadrant IV holds the target genotypes with target fluorescence values displayed on the x-axis. Quadrant III either contains the control samples or samples that displayed neither genotype. The star in 5B indicates biparental, F1 TONG x ACET progeny. ‘sample’= , ‘mult/hort//bark/acet’- , ‘cico’ = , ‘mix mult 10: cico 90’ = ,a mix of the multbreacteatum target and the non-target Ciconium in ratios of 10/90 (or other) ‘NTC’ = non target control (parent), ‘H2O’ is the water control.
Figure 1.
KASP allelic discrimination plots for F1 hybrid accessions from crosses involving P. x hortorum (A) P. acetosum (B), P. barklyi (C) and P. multibracteatum (D)F1 hybrid samples. Each data point represents the fluorescence signal of an individual DNA sample, with the x-axis indicating the amount of FAM fluorescence and the y-axis the HEX fluorescence for each sample. Negative control samples are expected to be placed around the origin of the plot. Samples of the same mitotype will have generated similar levels of fluorescence and will therefore cluster together on the plot. Each graph is divided into four quadrants (I-IV). Here, the y-axis displays the values assigned to the non-target Ciconium genotypes. Hence quadrant I holds the non-target Ciconium genotypes. Quadrant IV holds the target genotypes with target fluorescence values displayed on the x-axis. Quadrant III either contains the control samples or samples that displayed neither genotype. The star in 5B indicates biparental, F1 TONG x ACET progeny. ‘sample’= , ‘mult/hort//bark/acet’- , ‘cico’ = , ‘mix mult 10: cico 90’ = ,a mix of the multbreacteatum target and the non-target Ciconium in ratios of 10/90 (or other) ‘NTC’ = non target control (parent), ‘H2O’ is the water control.
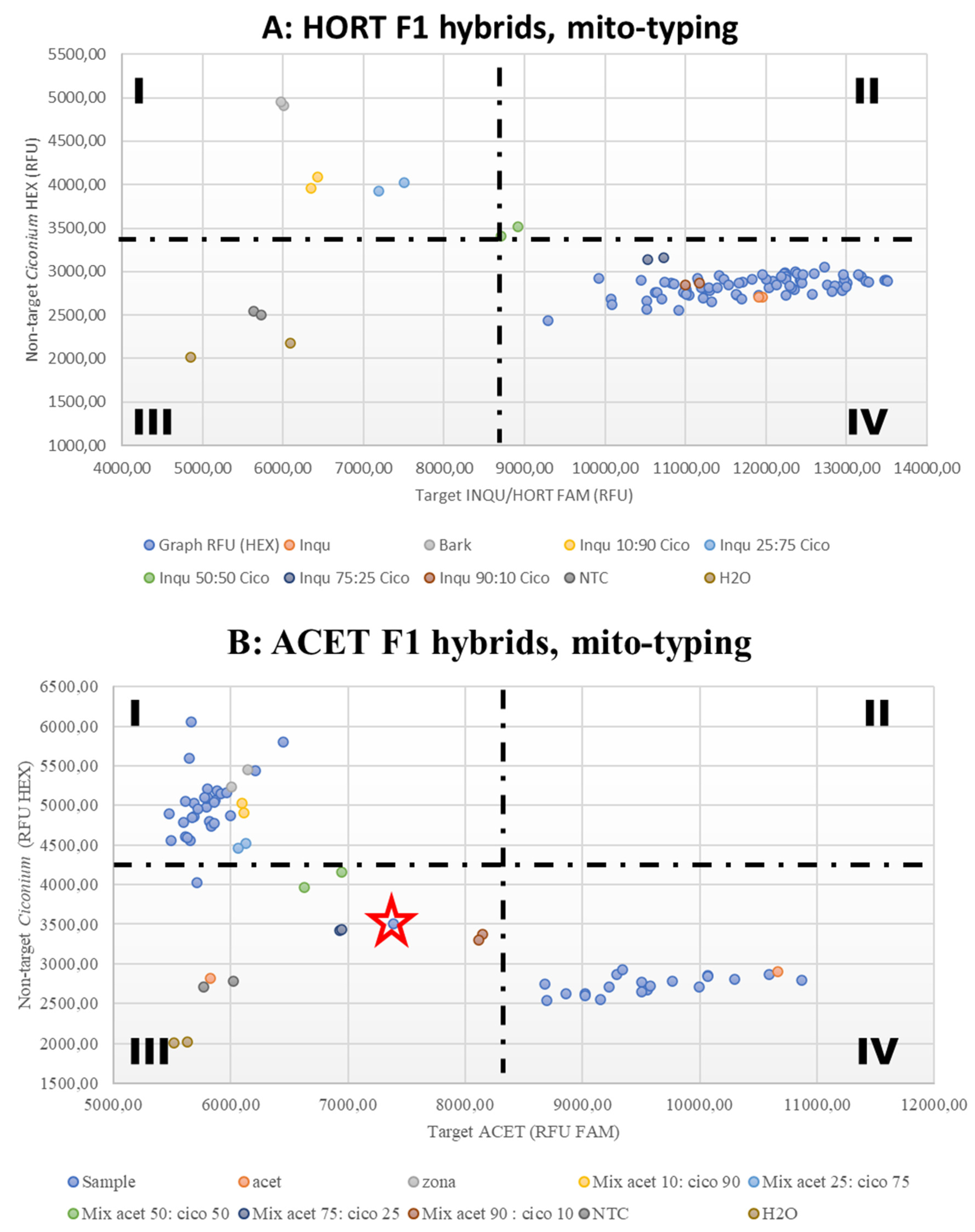
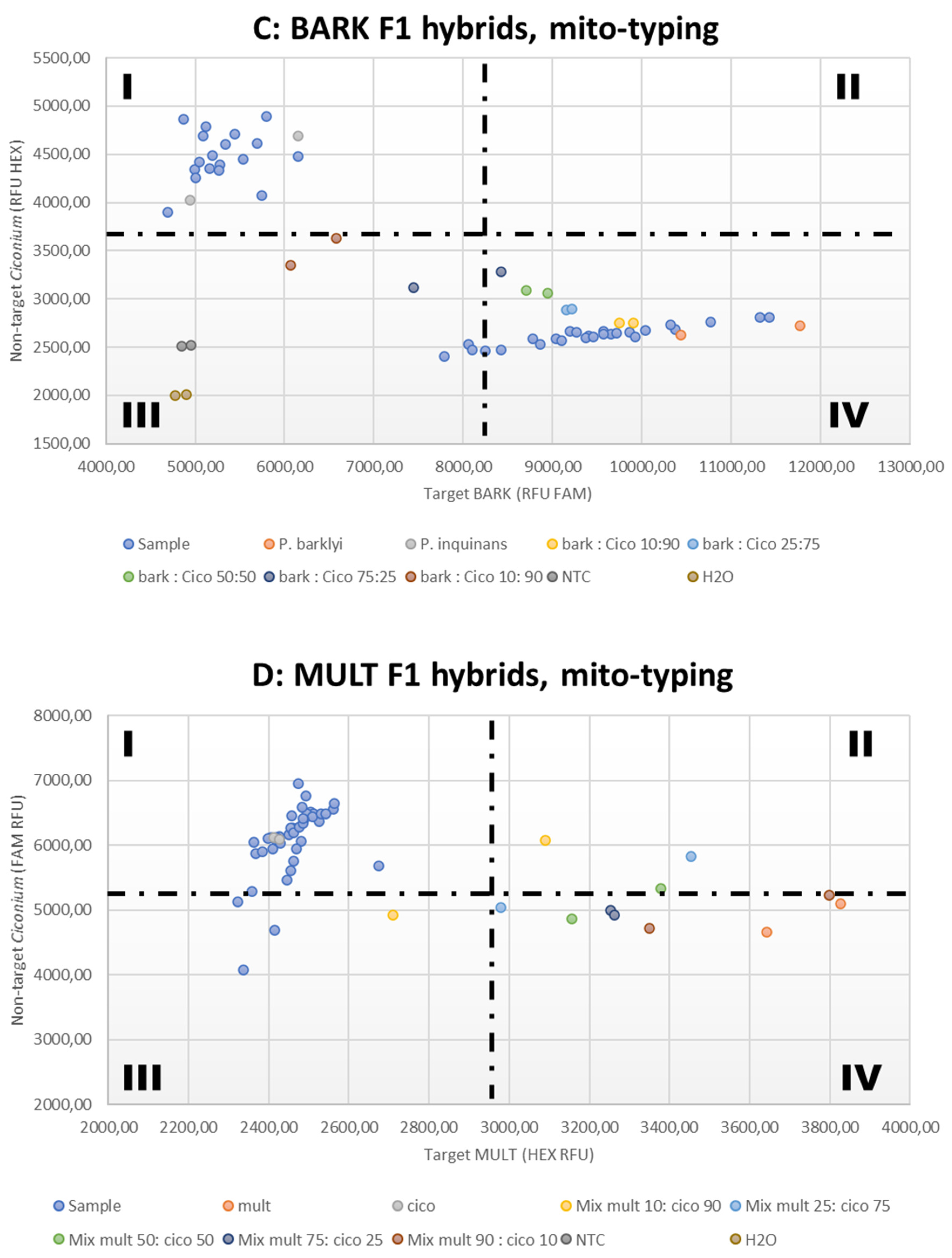

Table 1.
Plant materials used in this study, along with herbarium voucher information.
| species | Herbarium Voucher accession | Institute1 | Acronym used in text |
|---|---|---|---|
| P. acetosum | 1243 | STEU | ACET |
| P. acraeum | 1975 | STEU | ACRA |
| P. alchemilloides | 1885 | STEU | ALCH2x |
| P. alchemilloides | 1882 | STEU | ALCH4x |
| P. articulatum | 1972055 | WAG | ARTI |
| P. barklyi | 1972061 | WAG | BARK |
| P. frutetorum | 0754 | STEU | FRUT |
| P. inquinans | 0682 | STEU | INQU |
| P. multibracteatum | 2902 | STEU | MULT |
| P. peltatum | 1890 | STEU | PELT |
| P. quinquelobatum | 1972049 | WAG | QUIN |
| P. ranuncolophyllum | A3651 | MSUN* | RANU |
| P. tongaense | 3074 | STEU | TONG |
| P. zonale | 1896 | STEU | ZONA |
| P. elongatum | 0854 | STEU | ELON |
| P. aridum | 1847 | STEU | ARID |
| P. insularis | 19990489 | RBGE | INSU |
| P. yemenense sp. nov | 1972037 | WAG | YEME |
| P. omanense sp. nov | 2184 | RBGE | OMAN |
| P. somalense | V-067490 | V | SOMA |
1STEU = Stellenbosch University, RSA; AL = Albers/MSUN=Münster & *Bakker et al. 2004. WAG= National Herbarium of the Netherlands. V=Uppsala herbarium.
Table 2.
Mitome type determined per F1 cross of P. sect Ciconium species ‘M’ and ‘P’ denote maternally and paternally inherited mitome marker, respectively, B denotes biparental inheritance of the mitochondrial marker(s), ‘B/H/R’ denotes plants which showed biparental inheritance, but one of the parents could also have been heteroplasmic or there could even have been recombination. ‘WT’ refers to parental stocks displaying the wildtype marker. * are F1 plants for which (one of the) parents may have been heteroplasmic. ° plant treated as wildtype, but the result of a hybridisation process. WT plants are excluded from the final calculation of total and percentages. The fertility phenotypes are given as F which is fully fertile; P: partial fertile (pollen observed); MS: male sterile / no pollen observed, ‘- -’ could not be evaluated because the plants did not flower.
Table 2.
Mitome type determined per F1 cross of P. sect Ciconium species ‘M’ and ‘P’ denote maternally and paternally inherited mitome marker, respectively, B denotes biparental inheritance of the mitochondrial marker(s), ‘B/H/R’ denotes plants which showed biparental inheritance, but one of the parents could also have been heteroplasmic or there could even have been recombination. ‘WT’ refers to parental stocks displaying the wildtype marker. * are F1 plants for which (one of the) parents may have been heteroplasmic. ° plant treated as wildtype, but the result of a hybridisation process. WT plants are excluded from the final calculation of total and percentages. The fertility phenotypes are given as F which is fully fertile; P: partial fertile (pollen observed); MS: male sterile / no pollen observed, ‘- -’ could not be evaluated because the plants did not flower.
| F1 types | # plants/cross | # marker pairs /cross | fertility phenotype | (M), (P), (B), (H), (R), (WT) |
|---|---|---|---|---|
| acet_x_frut | 2 | 1 | P | M |
| acet_x_inqu | 1 | 2 | P | M |
| acet_x_zona | 12 | 1 | -- | M (11) P (1) |
| alch(4x)_x_bark | 1 | 1 | -- | M |
| alch(4x)_x_yeme | 2 | 1 | P | P |
| bark_x_frut | 3 | 1 | MS | M |
| bark_x_inqu | 1 | 2 | -- | P |
| bark_x_mult | 3 | 2 | MS | M |
| bark_x_quin | 2 | 1 | -- | M(2) P(1) |
| frut_x_acet | 1 | 1 | P | M |
| frut_x_bark | 3 | 1 | MS | M |
| hort(4x)_x_arti(4x) | 8 | 1 | MS | M |
| (hort_x_zona)_x_arid | 5 | 1 | MS | M |
| hort_x_acet | 3 | 2 | P | M |
| hort_x_acra | 1 | 1 | P | M |
| hort_x_alch | 1 | 1 | MS | M |
| hort_x_arid | 6 | 2 | MS | M |
| hort_x_bark | 2 | 1 | -- | M |
| hort_x_frut | 1 | 1 | F | M |
| hort_x_mult | 1 | 1 | MS | M |
| hort_x_quin | 15 | 1 | MS | M(8) P(7) |
| hort_x_tong | 8 | 1 | P | M |
| hort_x_tong(4x) | 1 | 1 | P | M |
| hort_x_zona | 26 | 1 | P | M |
| tong_x_acet | 7 | 1-2 | P | B(1)/P |
| yeme_x_alch(4x) | 1 | 1 | P | P |
| P. inquinans | 1 | 1 | F | WT |
| P. peltatum | 1 | 1 | F | WT |
| P. salmoneum | 1 | 2 | F | WT |
| P. x hortorum_4x° | 1 | 1 | P | WT* |
| P. quinquelobatum | 1 | 3 | F | WT |
| P. yemenense | 1 | 3 | F | WT |
| P. barklyi | 1 | 3 | F | |
| P. aridum | 1 | 3 | F | WT |
| P. quinquelobatum | 1 | 3 | F | WT |
| P. alchemilloides | 1 | 3 | F | WT |
| P. tongaense | 1 | 1 | F | WT |
| P. articulatum | 1 | 1 | F | WT |
| P. multibreacteatum | 1 | 2 | F | WT/H |
| mult_x_acet | 8 | 2-3 | -- | B/H* |
| mult_x_alch | 14 | 2 | P | P/H* |
| mult_x_arid | 3 | 1 | MS | P/H* |
| mult_x_bark | 5 | 2 | MS | B/H* |
| mult_x_inqu | 3 | 1 | -- | P/H* |
| mult_x_pelt | 3 | 1 | MS | P/H* |
| mult_x_quin | 6 | 2 | P | P/H* |
| mult_x_ranu | 9 | 1-2 | P | P/H* |
| mult_x_tong | 2 | 1 | MS | P/H* |
| mult_x_zona | 2 | 2 | MS | P/H* |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



