Submitted:
23 April 2024
Posted:
23 April 2024
You are already at the latest version
Abstract
Carbapenems-resistant enterobacteria are considered multidrug-resistant, making the control of their nosocomial infections a major challenge in healthcare settings. It is important to know the incubation site and source of the spread of these strains, especially in neonatal intensive care units (NICU). Colonization of the colon of preterm neonates per se by multidrug-resistant bacterial strains could be the source of dissemination of these multidrug-resistant strains and their site of incubation. In this study, colonization of the colon of preterm neonates with enterobacteria and their multidrug-resistant strains was investigated. The presence of enterobacteria in the stool of preterm neonates was investigated on EMB (eosin-methylene blue) agar, followed by morphological and biochemical characterization and phylogenetic analysis of the 16S rRNA-encoding gene sequences. Antibiotic susceptibility of all isolated strains was tested using VITEK 2 technology. All cases examined at two weeks of age and older showed bacterial colonization of the colon. Klebsiella pneumoniae colonized the colon in 56.2% of the preterm neonates and Klebsiella quasipneumoniae was found in 6.3%, corresponding to 62.5% in total. The colonizing Klebsiella pneumoniae was multidrug-resistant in 2 of 9 cases. One strain of Klebsiella pneumoniae was resistant to carbapenems and another showed intermediate resistance to these antibiotics. These Klebsiella pneumoniae strains showed resistance to most of the other antibiotics tested. Some cases contained more than one genus, with Escherichia coli found in 50% of cases in preterm neonates. In contrast to Klebsiella pneumoniae, none of the E. coli strains colonizing the colon of preterm neonates in this study showed resistance to carbapenems. The results of this study suggest that Klebsiella pneumoniae and its carbapenems multidrug-resistant strains are well marked in preterm neonates colonizing their colon, which is the incubation site and source of such multidrug-resistant bacterial strains in NICU. This study highlights the importance of careful management of preterm neonates in the NICU and possible contamination with multidrug-resistant Klebsiella pneumoniae incubated in their colon. Further studies would be of interest to avoid the possibility of fecal-oral transmission and infection by these multidrug-resistant bacteria and their treatment in the colon, where their incubation site is located.
Keywords:
Carbapenems-resistant enterobacteria
; Escherichia coli
; Klebsiella pneumoniae
; Multidrug-resistant bacteria
; Preterm neonates
1. Introduction
Multidrug-resistant bacteria pose a growing challenge in combating their nosocomial infections, where only a limited range of antibiotics is available for effective treatment. Carbapenems, the most recently developed β-lactams, such as meropenem and imipenem (currently marketed as imipenem/cilastatin), have a broad-spectrum of antibacterial activity and are generally used as a last resort in the treatment of multidrug-resistant bacterial infections [1,2,3,4,5]. In nosocomial settings, where multidrug-resistant Gram-negative bacteria are already endemic, the spread of the carbapenems-resistant bacterial strains is particularly problematic [6]. The human colon is considered a microbial ecosystem whose microbial community is important for the health of the body [7,8,9]. The initial colonization of the colon in neonates with health-beneficial and commensal bacteria and the establishment of its microbiome is a complicated process [10,11,12], where the enterobacterium E. coli and lactic acid bacteria initial colonization of the colon occur in the first week of life in healthy neonates [13,14].
Because of the multiple metabolic functions it performs and its relevance to the human body’s health, the human gut microbial community is sometimes referred to as the forgotten metabolic organ [15,16,17,18]. In addition to its function in digestion of insoluble fibers, the human gut microbial community also protects the gut epithelium from pathogens by occupying the available niches of the intestinal lumen space and nutrients and plays a basic role in the development of the human body’s immune system [19,20,21]. The gut resident microbial community represent thouthands of species microbial genomes serving the human body in excess of the human genome. The human body contains trillions of commensal and health benificial microbial cells, belonging to thouthands of species, that outnumber the human body cells by around 10 fold [22,23] with an estimated composition of 100 trillion of microbial cells in the human gut [24]. The gut resident microbial community has a metabolic activity that is collectively equal to that of an acutal organ in the body [15].
Nosocomial infections by multidrug-resistant bacteria are posing a huge burden to healthcare systems worldwide and are associated with high morbidity and mortality [25,26,27,28,29]. The plumbing systems components and sanitary installations in the hospital restrooms could represent a reservoir and incubation sites for multidrug-resistant bacteria [29,30,31,32]. Several factors may increase the risk of nosocomial infections with multidrug-resistant bacteria, including overuse of broad-spectrum antibiotics, sterility of invasive devices, the intensive health care unit environment, and other factors. The abundance and continuity of the source of incubation of these multidrug-resistant bacteria is an important risk factor in the intensive care units. Therefore, it is important to know, the incubation site and source of spread of multidrug-resistant bacteria in the neonatal intensive care unit. Colonization of the colon of preterm neonates by multidrug-resistant bacterial strains could be the incubation site and source of spread of these multidrug-resistant strains. In this study, colonization of the colon of preterm neonates by normal resident enterobacteria and their multidrug-resistant strains was investigated to determine whether colonization of the colon of preterm neonates by multidrug-resistant enterobacteria is the site of incubation and the source of spread of these strains.
2. Materials and Methods
The colonization of preterm neonates’ colon by normal resident enterobacteria and their multidrug-resistant strains was investigated by exploring its occurrence in preterm neonates’ stool. This study was conducted according to approval from the ethics committee of the institutional review board (IRB), General Directorate of Health Affairs in Madinah, Kingdom of Saudi Arabia (KSA) [IRB committee Head, Dr. Yasmeen Talal AlJehani]. Informed consents were obtained from parents of preterm neonates enrolled in the study for the collection of the preterm neonate’s stool. Stool samples from preterm neonates were collected from the neonatal intensive care units in three hospitals in Madinah, KSA [Cases 1, 2, 3 were collected from Maternity and Children hospital; cases 4, 5, 6 from Saudi German hospital Madinah; cases 7 up to 21 from Ohud hospital]. The different preterm neonates were chosen randomly to obtain the stool samples for having a wide view of the study. Once the samples of preterm neonate stool normally obtained in diapers were collected, they were sent for microbiological analysis in the bacteriology laboratory at Taibah University and used directly for exploring the occurrence of enterobacteria and its multidrug-resistant strains. The enterobacteria occurrence in the stool of preterm neonates was followed using EMB agar.
Serial dilution of stool samples was performed and subsequently E. coli colonies with metallic green sheen or Klebsiella large, dome-shaped, pink to purple dark mucoid colonies or others were detected on the EMB agar after 24 h of inoculation and incubation at 37 °C and CFU (Colony Forming Units) was followed by MPN (Most Probable Number) techniques [33]. The EMB plates with low detectable number of bacterial colonies have been used to determine the CFU of the dominating bacterial colonies of similar morphological and biochemical characteristics from which an isolated bacterial strain was subcultured in EMB agar for obtaining a purified strain. Bacterial isolates were further purified on LB (Luria-Bertani) agar medium, cultured on LB broth at 37 °C for 24 h and preserved in same medium retaining 20% glycerol at -80 °C. Morphological and biochemical characterization of enterobacteria were conducted for identification as outlined in Bergey’s Manual [34] and by VITEK 2 technology. Further genetic identification of bacterial strains using molecular biological technology by 16S rRNA encoding gene sequence phylogenetic analysis. The antibiotic susceptibility test of all isolated bacterial strains was conducted using VITEK 2 technology automated susceptibility testing system in Al Madinah Al Munawarah Hospital in King Salman Bin Abdulaziz Medical City. The susceptibility of isolated bacterial stains to various antibiotics and thier resistance versus sensitivity were determined from the MIC (Minimum Inhibitory Concentration) followed by VITEK.2 device in Al Madinah Al Munawarah Hospital in King Salman Bin Abdulaziz Medical City.
2.1. Genetic Identification of Bacterial Strains by Phylogenetic Analysis of 16S rRNA Encoding Gene Sequence
The genetic identification of bacterial strains using molecular biological technology was conducted by phylogenetic analysis of the 16S rRNA encoding gene sequence of each bacterial strain.
2.1.1. PCR Amplification of the 16S rRNA Gene
Extraction and purification of the genomic DNA of bacterial cultures were conducted using Promega Wizard (Promega Corporation, Madison, WI, USA) genomic DNA purification kit according to instructions described by the manufacturer. The purified genomic DNA was used as a template for amplifying a nearly full-length nucleotide sequence of the 16S rRNA-encoding gene by polymerase chain reaction (PCR). The universal forward (27F, 5'-AGAGTTTGATC[A/C]TGGCTCAG-3') and reverse (1492R, 5'-G[C/T]TACCTTGTTACGACTT-3') primers were used for the PCR amplification of the nearly full-length sequence of the 16S rRNA-encoding gene [35]. The PCR amplification of the nearly full-length sequence of the 16S rRNA-encoding gene was conducted in a thermal cycler (model 2720; Applied Biosystems, Foster City, CA, USA) using a 25 µl reaction mixture (25 µl) with the following composition: 2.5 µl 10× Taq buffer (100 mM Tris-HCl, pH 8), 100 mM deoxynucleoside triphosphates (dNTPs) (Invitrogen, Carlsbad, CA, USA), 1.25 mM MgCl2, 1.2 mM forward and reverse primers, 0.5 U Taq DNA polymerase (Invitrogen, USA), and about 5 ng of the template bacterial genomic DNA. The following PCR program was followed for the amplification: Initial denaturation at 95°C for 5 min followed by 35 cycles of amplification [94°C (denaturation) for 1 min, 56°C (annealing) for 1 min, and 72°C (extension) for 1 min] and a final extension 10 min at 72°C. Analyzing the PCR amplification products was conducted using agarose gel electrophoresis using 1% agarose gels containing 5 µ/ml ethidium bromide in comparison to a DNA size marker (1 kb Plus DNA ladder; Invitrogen, USA).
2.1.2. Nucleotide Sequence Analysis
Purification and cycle sequencing of the PCR products were conducted at the Macrogen Korea sequencing facility located in Seoul, South Korea. The purified PCR products were subjected to direct cycle sequencing at the Macrogen Korea sequencing facility by the automated florescent dye terminator sequencing method [36] in the 3730XL DNA analyzer (Applied Biosystems, CA, USA) using the same forward (27F) and reverse (1492R) primers for sequencing in both directions. Using the NCBI (National Center for Biotechnology and Information) server’ nucleotide-nucleotide BLAST search tool, the sequence reads of the 16S rRNA-encoding gene of each isolated bacterial strain colonizing the preterm neonates’ colon were assembled and compared with the closest matches of bacterial strains recorded in the GenBank at www.ncbi.nlm.nih.gov/blast/Blast.cgi. Alignments of sequences of the 16S rRNA gene were conducted using Clustal W1.83 XP [37]. Using the neighbor-joining method [38], the derived phylogenetic tree of the 16S rRNA gene sequences was constructed by MEGA11 software [39]. Bacillus subtilis strain JCM 1465 (accession number NR_113265) was used as an outgroup.
2.1.3. Data Availability and Nucleotide Sequences Accession Numbers
The partial nucleotide sequences of the 16S rRNA-encoding gene of the bacterial strains colonizing the colon of preterm neonates were deposited in the GenBank under an accession number assigned for each strain in the NCBI’s GenBank nucleotide sequence database. The deposited 16S rRNA-encoding gene sequence accession numbers are listed in Table 1.
3. Results
In this study, the colonization of the preterm neonate’s colon by enterobacteria and its multidrug-resistant strains was investigated for exploring whether the colon of preterm neonates per se could be the source of dissemination of these multidrug-resistant strains and their site of incubation. This was conducted by following the occurrence of enterobacteria in the stool of preterm neonates (Table 1) on EMB agar followed by morphological and biochemical characterizations for identification and by VITEK 2 technology. The antibiotic susceptibility using VITEK 2 technology automated susceptibility testing system of all isolated bacterial strains was also conducted (Table 2 and Table 3). The gestational age of all investigated preterm neonate cases ranged between 25 and 33 weeks (Table 1).
All investigated preterm neonate cases were receiving antibiotics treatment intravenously in the intensive care unit as outlined in Table 1. The age of all investigated randomly selected preterm neonate cases ranged between 2 and 64 days (Table 1). No bacteria were detected in the stool of all investigated preterm neonate cases with ages 2 up to 8 (Table 1). All investigated preterm neonate cases with age two weeks and above (16 cases) showed bacterial colonization of their colon (Table 1).
After identification by morphological and biochemical characterizations and by using VITEK 2 technology, the molecular biological identification of all isolated strains of bacteria was also performed by phylogenetic analysis of each bacterial strain 16S rRNA-encoding gene sequence (Figure 1, Figure 2, Figure 3 and Figure 4). The near full-length nucleotide sequence of the 16S rRNA-encoding gene of all isolated bacterial strains deposited in the NCBI’s GenBank nucleotide sequence database, were used for phylogenetic analysis based identification of these bacterial strains in comparasion to the closest matches of bacterial strains recorded in the GenBank (Figure 1, Figure 2, Figure 3 and Figure 4).
Klebsiella pneumoniae (Figure 1) was found colonizing colon in 9 cases of preterm neonates in neonatal intensive care units (Table 1) representing 56.2% of the total colonized cases (16 cases) and the relevant bacterium Klebsiella quasipneumoniae (Figure 2) was found in one case representing 6.3% and so, both bacteria represent in total 62.5% of the total colonized cases. Some cases contained more than one genus (Table 1) where E. coli (Figure 3) was found colonizing colon in 8 cases representing 50 % of the total colonized cases of preterm neonates (16 cases) in neonatal intensive care units (Table 1).
For exploring the occurrence of multidrug-resistant Enterobacteria, the isolated bacterial stains susceptibility to various antibiotics and the resistance versus sensitivity of these bacterial stains were explored from the MIC (Table 2 and 3) followed by VITEK.2 device. Strains of Klebsiella pneumoniae (Figure 1) colonized the colon of the investigated preterm neonates showing extensive multidrug resistance was clear in two strains (Table 2). One strain Klebsiella pneumoniae EFTU110 was resistant to Carbapenems (Meropenem and Imipenem) and another Klebsiella pneumoniae EFTU113 showed intermediate resistance to these antibiotics. These two strains of Klebsiella pneumoniae showed resistance to most other tested antibiotics (Table 2). The occurence of Carbapenems resistant Klebsiella pneumoniae in stool of of preterm neonates (Table 2) refers to the ability of these multidrug-resistant bacterial strains to colonize the colon of preterm neonates where its incubation site is located in the neonatal intesnsive care units. In contrast to Klebsiella pneumoniae, none of the E. coli strains (Figure 3) colonizing the colon of the investigated preterm neonate cases in this study showed resistance to the Carbapenems tested Meropenem and Imipenem (Table 3).
Along with enterobacteria, a non-Enterobacterium Acinetobacter baumannii known for its active nosocomial infections was found in the stool of two cases representing 12.5% of the total colonized preterm neonate cases (Table 1) confirmed by identification using the phylogenetic analysis of the 16S rRNA-encoding gene sequence (Figure 4). The Acinetobacter baumannii strains colonizing the colon of preterm neonates isolated from the two cases did not show resistance to the Carbapenems tested Meropenem and Imipenem (Table 2) and thus Carbapenems multidrug-resistant strains were found only belonging to Klebsiella pneumoniae.
4. Discussion
This study was devoted to visualizing the incubation site and source of the spread of the multidrug-resistant enterobacteria strains in the neonatal intensive care units and investigating whether the colonization of the colon of the antibiotics intravenously treated preterm neonates per se by multidrug-resistant enterobacteria is such incubation site and source of the spread of these multidrug-resistant strains. The colonization of preterm neonates’ colon by normal resident enterobacteria and its multidrug-resistant strains was followed by exploring its occurrence in preterm neonates’ stool. All investigated cases with ages 2 up to 8 did not show any bacteria in their stool indicating a delay in colonization possibly due to antibiotic treatment and health care environment in the neonatal intensive care unit where bacterial colonization of preterm neonates’ colon appeared in all investigated preterm neonate cases with age two weeks and above. This is in contrast to a previous study of healthy neonates where the healthy neonates’ colon showed initial colonization with the normal colon resident enterobacterium E. coli at days 5 and 6 in the second half of the first week of their life in all healthy neonates investigated subjects despite the delay in neonates born by cesarean section in comparison to vaginal birth [14].
E. coli is a major member of the enteric bacteria of the normal microbial community in the human colon which plays several beneficial roles for the body health [40,41,42,43,44] and is one of the first bacterial community members initially colonizing the colon microbial ecosystem in the neonates [14]. Despite its frequent pathogenicity, the enteric E. coli as one of the normal microbiota in the human gut plays many interesting roles for the human body health [42,43]. The commensal E. coli as a normal resident has many healthy beneficial roles in the human colon including production of certain vitamins such as vitamin K2 (menaquinone) [40,45] and vitamin B1 [46,47]. Strains of E. coli includig the well-known probiotic E. coli strain Nissle 1917 marketed as Mutaflor [48,49] and the probiotic E. coli marketed as Symbioflor [50,51] have long been used as a probiotic with many health benefits [52,53,54,55,56,57,58]. The probiotic Symbioflor E. coli is less known than Mutaflor (E. coli Nissle 1917) and thier main differences is that Symbioflor probiotic contains six genotypes of E. coli comprising six probiotic E. coli strains which together constitute the probiotic product entitled Symbioflor 2 [50,59]. The enterobacterium E. coli was also suggested along with other facultative anaerobes in the colon to play an important role to sustain the installation of anaerobiosis in the large intestine as a basic requirement for the various fermentation processes where such fermentation processes in the colon is of importance to the human body health in many aspects [60,61].
Carbapenems-resistant gram-negative bacteria, such as Carbapenems-resistant Enterobacteriaceae are considered, multidrug-resistant bacteria. The occurrence of Carbapenems-resistant Enterobacterium Klebsiella pneumoniae in the stool of preterm neonates represents a spotlight on the incubation and possibly the creation site of these multidrug-resistant strains colonizing the colon in antibiotics intravenously treated preterm neonates in the neonatal intensive care units. The multidrug-resistant bacteria may lead to virtually untreatable infections where such pathogens might acquire resistance determinants for multiple antibiotic classes [62]. Carbapenems-resistant Enterobacterales infections represent an important therapeutic problem where other effective therapeutic alternatives are limited [63]. The Carbapenems class of antibiotics is usually reserved for the treatment of infections caused by known or suspected multidrug-resistant bacteria [4]. The appearance of resistant bacteria to this last resort effective therapeutic alternative antibiotics would be especially daunting in neonatal intensive care units. The presence of Acinetobacter baumannii, detected in 12.5 % of the investigated preterm neonate subjects indicates the ability of this non-Enterobacterium to colonize the colon of preterm neonates along with enterobacteria. This non-Enterobacteriaceae bacterium Acinetobacter baumannii was reported responsible for outbreaks in the neonatal intensive care units and many infections in neonates [64]. This opportunistic pathogen has a high ability for nosocomial infections [65] particularly among immunocompromised individuals in a prolonged hospital stay [66,67] and hence might represent a risk for preterm neonates with immature immune system. The colonization of preterm neonates’ colon by this bacterium should be visualized in the neonatal intensive care unit patients for treatment and to avoid this incubation source of such pathogen. The strains of Acinetobacter baumannii colonizing the preterm neonates’ colon in this study did not show resistance to Carbapenems in similar results to E. coli strains indicating that Carbapenems multidrug-resistant Klebsiella pneumoniae strains are well marked in preterm neonate cases and more likely to occur in the neonatal intensive care units. Klebsiella pneumonia represent opportunistic nosocomial pathogen and is reported as a cause of various healthcare-associated infections other than bacterial pneumonia which include bloodstream infections [68,69,70,71], meningitis [72,73] and others [74] where it is a major cause of neonatal sepsis [75,76]. The ability of multidrug-resistant Klebsiella pneumoniae to colonize the colon of preterm neonates might indicate a high risk of infection to preterm neonates in the neonatal intesnsive care units.
In conclusion, an analysis of the bacterial community in the stool of preterm neonates to screen the pathogenic bacteria colonizing the colon, especially multidrug-resistant bacteria, could be helpful to overcome the source of incubation of these pathogens by treatment and thus prevent healthcare-associated infections and hospital outbreaks in neonatal intensive care units. Such an analysis to determine colonization of the colon of preterm neonates by multidrug-resistant Enterobacteriaceae and other bacteria could be important to isolate the cases that incubate these strains in their colon and thus control nosocomial infections with multidrug-resistant bacteria in neonatal intensive care units.
In future applications, A frequent microbiology test of stool samples from preterm neonates in the neonatal intensive health care units, might be helpful for better followup and isolation of cases in which multidrug resistant strains of klebsiella neomonia or other bacteria apprear. This might also be helpful to determine the normality of the major colonization members of the microbial community in the preterm neonates gut in comparasion to a general figure of the microbial community colonizing the gut in healthy neonates of the same age. This would require further future studies to have a full picture for diagnosing the microbial community in the neonates gut ecosystem in this early critical stage of the human life.
Conflicts of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
All authors listed declare the absence of any commercial or financial relationships to the conducted research that could be construed as a potential conflict of interest. All authors listed contributed equally to this work and made a substantial contribution to the work.
References
- Nordmann, P.; Dortet, L.; Poirel, L. Carbapenem resistance in Enterobacteriaceae: Here is the storm! Trends Mol. Med. 2012, 18, 263–272. [Google Scholar] [CrossRef]
- Ellappan, K.; Belgode Narasimha, H.; Kumar, S. Coexistence of multidrug resistance mechanisms and virulence genes in carbapenem-resistant Pseudomonas aeruginosa strains from a tertiary care hospital in South India. J. Glob. Antimicrob. Resist. 2018, 12, 37–43. [Google Scholar] [CrossRef]
- Lee, Y.; Bradley, N. Overview and insights into Carbapenem allergy. Pharmacy 2019, 7, 110–116. [Google Scholar] [CrossRef]
- Elshamy, A.A.; Aboshanab, K.M. A review on bacterial resistance to carbapenems: epidemiology, detection and treatment options. Future Sci. OA 2020, 6, FSO438. [Google Scholar] [CrossRef]
- Mohamed, A.; Daef, E.; Nafie, A.; Shaban, L.; Ibrahim, M. Characteristics of Carbapenem-resistant Gram-negative Bacilli in patients with ventilator-associated pneumonia. Antibiotics 2021, 10, 1325. [Google Scholar] [CrossRef]
- Pitout, J.D.D.; Nordmann, P.; Poirel, L. Carbapenemase-producing Klebsiella pneumoniae, a key pathogen set for global nosocomial dominance. Antimicrob. Agents Chemother. 2015, 59, 5873–5884. [Google Scholar] [CrossRef]
- Grönlund, M.M.; Arvilommi, H.; Kero, P.; Lehtonen, O.P.; Isolauri, E. Importance of intestinal colonisation in the maturation of humoral immunity in early infancy: a prospective follow up study of healthy infants aged 0–6 months. Arch. Dis. Child. Fetal. Neonatal. Ed. 2000, 83, F186–F192. [Google Scholar] [CrossRef]
- MacPherson, A.J.; Harris, N.L. Interactions between commensal intestinal bacteria and the immune system. Nat. Rev. Immunol. 2004, 4, 478–485. [Google Scholar] [CrossRef]
- Stavropoulou, E.; Bezirtzoglou, E. Human microbiota in aging and infection: a review. Crit Rev Food Sci Nutr 2019, 59, 537–545. [Google Scholar] [CrossRef]
- Fanaro, S.; Chierici, R.; Guerrini, P.; Vigi, V. Intestinal microflora in early infancy: composition and development. Acta Paediatr. Suppl. 2003, 91, 48–55. [Google Scholar] [CrossRef]
- Faith, J.J.; Guruge, J.L.; Charbonneau, M.; Subramanian, S.; Seedorf, H.; Goodman, A.L.; Clemente, J.C.; Knight, R.; Heath, A.C.; Leibel, R.L.; et al. The long term stability of the human gut microbiota. Science 2013, 341, 1237439. [Google Scholar] [CrossRef]
- Sommer, F.; Anderson, J.M.; Bharti, R.; Raes, J.; Rosenstiel, P. The resilience of the intestinal microbiota influences health and disease. Nat. Rev. Microbiol. 2017, 15, 630–638. [Google Scholar] [CrossRef]
- Al-Balawi, M.; Morsy, F.M. Enterococcus faecalis is a better competitor than other lactic acid bacteria in the initial colonization of colon of healthy newborn babies at first week of their life. Front. Microbiol. 2020, 11, 2017. [Google Scholar] [CrossRef]
- Al-Balawi, M.; Morsy, F.M. Prenatal versus postnatal initial colonization of healthy neonates’ colon ecosystem by the enterobacterium Escherichia coli. Microbiol. Spectrum 2021, 9, e00379-21. [Google Scholar] [CrossRef]
- Bocci, V. The neglected organ: bacterial flora has a crucial immunostimulatory role. Perspect. Biol. Med. 1992, 35, 251–260. [Google Scholar] [CrossRef]
- O’Hara, A.; Shanahan, F. The gut flora as a forgotten organ. EMBO Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef]
- Guinane, C.M.; Cotter, P.D. Role of the gut microbiota in health and chronic gastrointestinal disease: understanding a hidden metabolic organ. Therap. Adv. Gastroenterol. 2013, 6, 295–308. [Google Scholar] [CrossRef]
- Van de Vliet, M.; Joossens, M. The resemblance between bacterial gut colonization in pigs and humans. Microorganisms 2022, 10, 1831. [Google Scholar] [CrossRef]
- Flint, H.J.; Scott, K.P.; Louis, P.; Duncan, S.H. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 577–589. [Google Scholar] [CrossRef]
- Tamburini, S.; Shen, N.; Wu, H.C.; Clemente, J.C. The microbiome in early life: Implications for health outcomes. Nat. Med. 2016, 22, 713–722. [Google Scholar] [CrossRef]
- Gentile, C.L.; Weir, T.L. The gut microbiota at the intersection of diet and human health. Science 2018, 362, 776–780. [Google Scholar] [CrossRef]
- Shanahan, F. The host–microbe interface within the gut. Best Pract. Res. Clin. Gastroenterol. 2002, 16, 915–931. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef]
- Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell 2006, 124, 837–848. [Google Scholar] [CrossRef]
- Sousa, S.A.; Feliciano, J.R.; Pita, T.; Soeiro, C.F.; Mendes, B.L.; Alves, L.G.; Leitão, J.H. Bacterial nosocomial infections: Multidrug resistance as a trigger for the development of novel antimicrobials. Antibiotics 2021, 10, 942. [Google Scholar] [CrossRef]
- Coque, T.M.; Cantón, R.; Pérez-Cobas, A.E.; Fernández-de-Bobadilla, M.D.; Baquero, F. Antimicrobial Resistance in the Global Health Network: Known Unknowns and Challenges for Efficient Responses in the 21st Century. Microorganisms 2023, 11, 1050. [Google Scholar] [CrossRef]
- Freitas, A.R.; Werner, G. Nosocomial Pathogens and Antimicrobial Resistance: Modern Challenges and Future Opportunities. Microorganisms 2023, 11, 1685. [Google Scholar] [CrossRef]
- Duan, Z.; Li, X.; Li, S.; Zhou, H.; Hu, L.; Xia, H.; Xie, L.; Xie, F. Nosocomial surveillance of multidrug-resistant Acinetobacter baumannii: a genomic epidemiological study. Microbiol Spectr. 2024, 12, e0220723. [Google Scholar] [CrossRef]
- Valzano, F.; Coda, A.R.D.; Liso, A.; Arena, F. Multidrug-resistant bacteria contaminating plumbing components and sanitary installations of hospital restrooms. Microorganisms 2024, 12, 136. [Google Scholar] [CrossRef]
- Hocquet, D.; Muller, A.; Bertrand, X. What Happens in Hospitals Does Not Stay in Hospitals: Antibiotic-Resistant Bacteria in Hospital Wastewater Systems. J. Hosp. Infect. 2016, 93, 395–402. [Google Scholar] [CrossRef]
- Kizny Gordon, A.E.; Mathers, A.J.; Cheong, E.Y.L.; Gottlieb, T.; Kotay, S.; Walker, A.S.; Peto, T.E.A.; Crook, D.W.; Stoesser, N. The Hospital Water Environment as a Reservoir for Carbapenem-Resistant Organisms Causing Hospital-Acquired Infections-A Systematic Review of the Literature. Clin. Infect. Dis. 2017, 64, 1435–1444. [Google Scholar] [CrossRef]
- Park, S.C.; Parikh, H.; Vegesana, K.; Stoesser, N.; Barry, K.E.; Kotay, S.M.; Dudley, S.; Peto, T.E.A.; Crook, D.W.; Walker, A.S.; et al. Risk Factors Associated with Carbapenemase-Producing Enterobacterales (CPE) Positivity in the Hospital Wastewater Environment. Appl. Environ. Microbiol. 2020, 86, e01715–e01720. [Google Scholar] [CrossRef]
- MacFaddin, J. Media for isolation-cultivation-identification-maintenance of medical bacteria, Baltimore, Maryland: Williams and Wilkins; 1985, vol. 1.
- Brenner, D.J.; Krieg, N.R.; Staley, J.T.; Garrity, G.M. Bergey’s manual of systematic bacteriology. 2005, 2nd edition, Vol. 2 (The Proteobacteria), part C (The Alpha-, Beta-, Delta-, and Epsilonproteobacteria), Springer, New York, NY.
- Lane, D.J. 16S/23S rRNA sequencing, 1991, p 115–175. In Stackebrandt E, Goodfellow M (ed), Nucleic acid techniques in bacterial systematics. John Wiley and Sons, NewYork, NY.
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-termination inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef]
- Thompson, D.; Gibson, J.; Plewinak, F.; Jeanmougin, F.; Higgins, G. The ClastalX windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nuc. Acids Res. 1997, 25, 4867–4887. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 38,3022–3027. [Google Scholar] [CrossRef]
- Bhattacharyya, D.K.; Kwon, O.; Meganathan, R. Vitamin K2 (menaquinone) biosynthesis in Escherichia coli: evidence for the presence of an essential histidine residue in o-succinylbenzoyl coenzyme A synthetase. J. Bacteriol. 1997, 179, 6061–6065. [Google Scholar] [CrossRef]
- Gao, Y.D.; Zhao, Y.; Huang, J. 2014. Metabolic modeling of common Escherichia coli strains in human gut microbiome. Biomed Res Int 2014, 694967. [Google Scholar] [CrossRef]
- Delmas, J.; Dalmasso, G.; Bonnet, R. Escherichia coli: the good, the bad and the ugly. Clin. Microbiol.2015, 4:2.
- Cools, P. The role of Escherichia coli in reproductive health: state of the art. Res. Microbiol. 2017, 168, 892–901. [Google Scholar] [CrossRef]
- Christofi, T.; Panayidou, S.; Dieronitou, I.; Michael, C.; Apidianakis, Y. Metabolic output defines Escherichia coli as a health-promoting microbe against intestinal Pseudomonas aeruginosa. Sci Rep 2019, 9, 14463. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Cao, Y.; Guo, Z.F.; Chen, M.; Chen, X.; Guo, Z. Menaquinone biosynthesis in Escherichia coli: identification of 2-succinyl-5-enolpyruvyl-6- hydroxy-3-cyclohexene-1-carboxylate as a novel intermediate and re-evaluation of MenD activity. Biochemistry 2007, 46, 10979–10989. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, R.; Fairhurst, S.A.; Kriek, M.; Lowe, D.J.; Roach, P.L. Thiamine biosynthesis in Escherichia coli: isolation and initial characterization of the ThiGH complex. FEBS Lett. 2003, 539, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Wang, H.; Xie, J. Thiamin (vitamin B1) biosynthesis and regulation: a rich source of antimicrobial drug targets? Int. J. Biol. Sci. 2011, 7, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Jacobi, C.A.; Malfertheiner, P. Escherichia coli Nissle 1917 (Mutaflor): new insights into an old probiotic bacterium. Dig Dis. 2011, 29, 600–607. [Google Scholar] [CrossRef]
- Sonnenborn, U. Escherichia coli strain Nissle 1917-from bench to bedside and back: history of a special Escherichia coli strain with probiotic properties. FEMS Microbiol. Lett. 2016, 363, fnw212. [Google Scholar] [CrossRef] [PubMed]
- Beimfohr, C. A review of research conducted with probiotic E. coli marketed as Symbioflor. Int. J. Bacteriol. 2016, 2016, 3535621. [Google Scholar] [CrossRef]
- Escribano-Vazquez, U.; Beimfohr, C.; Bellet, D.; Thomas, M.; Zimmermann, K.; Langella, P.; Cherbuy, C. Symbioflor2® Escherichia coli genotypes enhance ileal and colonic gene expression associated with mucosal defense in gnotobiotic mice. Microorganisms 2020, 8, 512. [Google Scholar] [CrossRef] [PubMed]
- Altenhoefer, A.; Oswald, S.; Sonnenborn, U.; Enders, C.; Schulze, J.; Hacker, J.; Oelschlaeger, T.A. The probiotic Escherichia coli strain Nissle 1917 interferes with invasion of human intestinal epithelial cells by different enteroinvasive bacterial pathogens. FEMS Immunol. Med. Microbiol. 2004, 40, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Westendorf, A.M.; Gunzer, F.; Deppenmeier, S.; Tapadar, D.; Hunger, J.K.; Schmidt, M.A.; Buer, J.; Bruder, D. Intestinal immunity of Escherichia coli NISSLE 1917: a safe carrier for therapeutic molecules. FEMS Immunol. Med. Microbiol. 2005, 43, 373–384. [Google Scholar] [CrossRef]
- Ukena, S.N.; Singh, A.; Dringenberg, U.; Engelhardt, R.; Seidler, U.; Hansen, W.; Bleich, A.; Bruder, D.; Franzke, A.; Rogler, G.; et al. Probiotic Escherichia coli Nissle 1917 inhibits leaky gut by enhancing mucosal integrity. PLoS ONE 2007, 2, e1308. [Google Scholar] [CrossRef]
- Mohsin, M.; Guenther, S.; Schierack, P.; Tedin, K.; Wieler, L.H. Probiotic Escherichia coli Nissle 1917 reduces growth, Shiga toxin expression, release and thus cytotoxicity of enterohemorrhagic Escherichia coli. Int. J. Med. Microbiol. 2015, 305, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Wassenaar, T.M. Insights from 100 years of research with probiotic. Eur. J. Microbiol. Immunol. 2016, 2016. 6, 147–161. [Google Scholar] [CrossRef]
- Zimmer, C.; Dorea, C. Enumeration of Escherichia coli in probiotic products. Microorganisms 2019, 7, 437. [Google Scholar] [CrossRef] [PubMed]
- Rebai, Y.; Wagner, L.; Gnaien, M.; Hammer, M.L.; Kapitan, M.; Niemiec, M.J.; Mami, W.; Mosbah, A.; Messadi, E.; Mardassi, H.; et al. Escherichia coli Nissle 1917 Antagonizes Candida albicans Growth and Protects Intestinal Cells from C. albicans-Mediated Damage. Microorganisms 2023, 11, 1929. [Google Scholar] [CrossRef]
- Zschüttig, A.; Auerbach, C.; Meltke, S.; Eichhorn, C.; Brandt, M.; Blom, J.; Goesmann, A.; Jarek, M.; Scharfe, M.; Zimmermann, K.; et al. Complete sequence of probiotic symbioflor 2 Escherichia coli strain G3/10 and draft sequences of symbioflor 2 E. coli strains G1/2, G4/9, G5, G6/7, and G8. Genome Announc. 2015, 3, e01330-14. [Google Scholar] [CrossRef]
- Miller, T.L.; Wolin, M.J. Fermentation by the human large intestine microbial community in an in vitro semi-continuous culture system. Appl. Environ. Microbiol. 1981, 42, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T.; Macfarlane, S. Fermentation in the human large intestine: its physiologic consequences and the potential contribution of prebiotics. J. Clin. Gastroenterol. 2011, 45, S120–S127. [Google Scholar] [CrossRef] [PubMed]
- Abat, C.; Fournier, P.E.; Jimeno, M.T.; Rolain, J.M.; Raoult, D. Extremely and pandrug-resistant bacteria extra-deaths: myth or reality? Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1687–1697. [Google Scholar] [CrossRef]
- Gajdács, M.; Ábrók, M.; Lázár, A.; Jánvári, L.; Tóth, Á.; Terhes, G.; Burián, K. Detection of VIM, NDM and OXA-48 producing carbapenem resistant Enterobacterales among clinical isolates in Southern Hungary. Acta Microbiol. Immunol. Hung. 2020, 67, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Zarrilli, R.; Bagattini, M.; Esposito, E.P.; Triassi, M. Acinetobacter Infections in Neonates. Curr. Infect. Dis. Rep. 2018, 10, 48. [Google Scholar] [CrossRef]
- Wisplinghoff, H.; Bischoff, T.; Tallent, S.M.; Seifert, H.; Wenzel, R.P.; Edmond, M.B. Nosocomial bloodstream infections in US hospitals: analysis of 24,179 cases from a prospective nationwide surveillance study. Clin. Infect. Dis. 2004, 39, 309–317. [Google Scholar] [CrossRef]
- Montefour, K.; Frieden, J.; Hurst, S.; Helmich, C.; Headley, D.; Martin, M.; Boyle, D.A. Acinetobacter baumannii: an emerging multidrug-resistant pathogen in critical care. Crit. Care Nurse 2008, 28, 15–26. [Google Scholar] [CrossRef]
- Howard, A.; O'Donoghue, M.; Feeney, A.; Sleator, R.D. Acinetobacter baumannii: an emerging opportunistic pathogen. Virulence 2012, 3, 243–250. [Google Scholar] [CrossRef]
- Meatherall, B.L.; Gregson, D.; Ross, T.; Pitout, J.D.; Laupland, K.B. Incidence, risk factors, and outcomes of Klebsiella pneumoniae bacteremia. Am. J. Med. 2009, 122, 866–873. [Google Scholar] [CrossRef]
- Girometti, N.; Lewis, R.E.; Giannella, M.; Ambretti, S.; Bartoletti, M.; Tedeschi, S.; Tumietto, F.; Cristini, F.; Trapani, F.; Gaibani, P.; et al. Klebsiella pneumoniae bloodstream infection: epidemiology and impact of inappropriate empirical therapy. Medicine 2014, 93, 298–309. [Google Scholar] [CrossRef]
- Durdu, B.; Hakyemez, I.N.; Bolukcu, S.; Okay, G.; Gultepe, B.; Aslan, T. Mortality markers in nosocomial Klebsiella pneumoniae bloodstream infection. SpringerPlus 2016, 5, 1892. [Google Scholar] [CrossRef]
- Li, L.; Huang, H. Risk factors of mortality in bloodstream infections caused by Klebsiella pneumonia: A single-center retrospective study in China. Medicine 2017, 96, e7924. [Google Scholar] [CrossRef]
- Tang, L.M.; Chen, S.T. Klebsiella pneumoniae meningitis: prognostic factors. Scand. J. Infect. Dis. 1994, 26, 95–102. [Google Scholar] [CrossRef]
- Ku, Y.H.; Chuang, Y.C.; Chen, C.C.; Lee, M.F.; Yang, Y.C.; Tang, H.J.; Yu, W.L. Klebsiella pneumoniae isolates from meningitis: Epidemiology, virulence and antibiotic resistance. Sci. Rep. 2017, 7, 6634. [Google Scholar] [CrossRef]
- Abbas, R.; Chakkour, M.; Zein El Dine, H.; Obaseki, E.F.; Obeid, S.T.; Jezzini, A.; Ghssein, G.; Ezzeddine, Z. General Overview of Klebsiella pneumonia: Epidemiology and the Role of Siderophores in Its Pathogenicity. Biology 2024, 13, 78. [Google Scholar] [CrossRef]
- Mukherjee, S.; Mitra, S.; Dutta, S.; Basu, S. Neonatal Sepsis: The Impact of Carbapenem-Resistant and Hypervirulent Klebsiella pneumoniae. Front. Med. 2021, 8, 634349. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Bhadury, P.; Mitra, S.; Naha, S.; Saha, B.; Dutta, S.; Basu, S. Hypervirulent Klebsiella pneumoniae Causing Neonatal Bloodstream Infections: Emergence of NDM-1-Producing Hypervirulent ST11-K2 and ST15-K54 Strains Possessing pLVPK-Associated Markers. Microbiol. Spectr. 2023, 11, e0412122. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic analysis of 16S rRNA encoding gene of Klebsiella pneumoniae strains isolated from preterm neoantes stool. The isolated Klebsiella pneumoniae strains phylogenetic tree shows the relationship with the closest neighbor strains of bacteria from NCBI. The accession numbers of 16S rRNA encoding genes of isolated Klebsiella pneumoniae strains are shown in boldface. The isolated Klebsiella pneumoniae strains and other bacterial strains neighborjoining tree was determined using 16S rDNA nearly full-length gene sequences and the frequency filter in the analysis package of MEGA 11 software. An out-group Bacillus subtilis JCM 1465 (accession number NR_113265) was used in the analysis. The scale bar shown indicates 2% estimated difference in sequence. The NCBI database accession numbers and strain names are shown in the phylogenetic tree and are also listed here: Klebsiella pneumoniae EFTU104, PP177307; Klebsiella pneumoniae EFTU113, PP204007; Klebsiella pneumoniae EFTU105, PP177367; Klebsiella pneumoniae EFTU110, PP177465; Klebsiella pneumoniae EFTU101, PP157088; Klebsiella pneumoniae KPLYC2, MT953921; Klebsiella pneumoniae E, OM090128; Klebsiella pneumoniae DJY-1, OK036428; Klebsiella pneumoniae LJH-D1, MK968716; Klebsiella pneumoniae EFTU116, PP177909; Klebsiella pneumoniae PSB B-10, OP750994; Klebsiella pneumoniae SDWH02, KX636139; Klebsiella pneumoniae EFTU3, PP157908; Klebsiella pneumoniae QLR-3, KM096435; Klebsiella pneumoniae EFTU103, PP177305; Klebsiella pneumoniae EFTU102, PP177257; Klebsiella pneumoniae ATCC 13883, NR_119278; Enterobacter cloacae ATCC 23373, NR_118011; Citrobacter freundii ATCC 8090, NR_028894; Enterobacter hormaechei 0992-77, NR_042154; Escherichia coli S484, MK951738; Bacillus subtilis JCM 1465, NR_113265.
Figure 1.
Phylogenetic analysis of 16S rRNA encoding gene of Klebsiella pneumoniae strains isolated from preterm neoantes stool. The isolated Klebsiella pneumoniae strains phylogenetic tree shows the relationship with the closest neighbor strains of bacteria from NCBI. The accession numbers of 16S rRNA encoding genes of isolated Klebsiella pneumoniae strains are shown in boldface. The isolated Klebsiella pneumoniae strains and other bacterial strains neighborjoining tree was determined using 16S rDNA nearly full-length gene sequences and the frequency filter in the analysis package of MEGA 11 software. An out-group Bacillus subtilis JCM 1465 (accession number NR_113265) was used in the analysis. The scale bar shown indicates 2% estimated difference in sequence. The NCBI database accession numbers and strain names are shown in the phylogenetic tree and are also listed here: Klebsiella pneumoniae EFTU104, PP177307; Klebsiella pneumoniae EFTU113, PP204007; Klebsiella pneumoniae EFTU105, PP177367; Klebsiella pneumoniae EFTU110, PP177465; Klebsiella pneumoniae EFTU101, PP157088; Klebsiella pneumoniae KPLYC2, MT953921; Klebsiella pneumoniae E, OM090128; Klebsiella pneumoniae DJY-1, OK036428; Klebsiella pneumoniae LJH-D1, MK968716; Klebsiella pneumoniae EFTU116, PP177909; Klebsiella pneumoniae PSB B-10, OP750994; Klebsiella pneumoniae SDWH02, KX636139; Klebsiella pneumoniae EFTU3, PP157908; Klebsiella pneumoniae QLR-3, KM096435; Klebsiella pneumoniae EFTU103, PP177305; Klebsiella pneumoniae EFTU102, PP177257; Klebsiella pneumoniae ATCC 13883, NR_119278; Enterobacter cloacae ATCC 23373, NR_118011; Citrobacter freundii ATCC 8090, NR_028894; Enterobacter hormaechei 0992-77, NR_042154; Escherichia coli S484, MK951738; Bacillus subtilis JCM 1465, NR_113265.
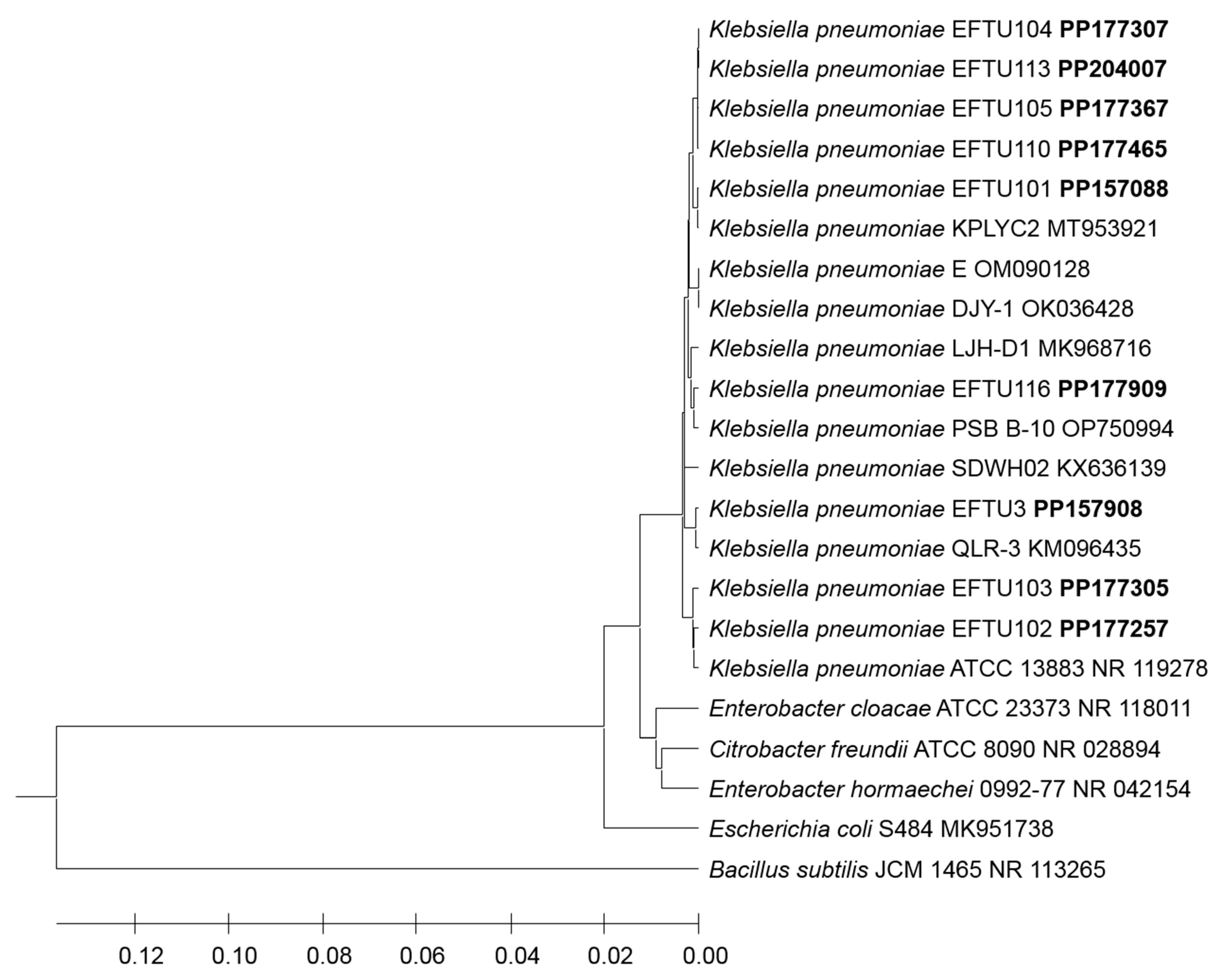
Figure 2.
Phylogenetic analysis of 16S rRNA encoding gene of Klebsiella quasipneumoniae strain EFTU115 isolated from preterm neoantes stool. The isolated Klebsiella quasipneumoniae strain EFTU115 phylogenetic tree shows the relationship with the closest neighbor strains of bacteria from NCBI. The accession numbers of 16S rRNA encoding genes of isolated Klebsiella quasipneumoniae strain EFTU115 is shown in boldface. The isolated Klebsiella quasipneumoniae strain EFTU115 and other bacterial strains neighborjoining tree was determined using 16S rDNA nearly full-length gene sequences and the frequency filter in the analysis package of MEGA 11 software. An out-group Bacillus subtilis JCM 1465 (accession number NR_113265) was used in the analysis. The scale bar shown indicates 2% estimated difference in sequence. The NCBI database accession numbers and strain names are shown in the phylogenetic tree and are also listed here: Klebsiella quasipneumoniae Kq-CW12, MK880381; Klebsiella quasipneumoniae. 2437, MT604862; Klebsiella quasipneumoniae A3GWS1-22(1)-E1, OR889330; Klebsiella quasipneumoniae EFTU115, PP177908; Klebsiella quasipneumoniae VITDJ2, MT826235; Klebsiella pneumoniae OOA, OR702992; Klebsiella pneumoniae ATCC 13883, NR_119278; Klebsiella pneumoniae UT09, OM978274; Enterobacter cloacae ATCC 23373, NR_118011; Citrobacter freundii ATCC 8090, NR_028894; Enterobacter hormaechei 0992-77, NR_042154; Escherichia coli S484, MK951738; Bacillus subtilis JCM 1465, NR_113265.
Figure 2.
Phylogenetic analysis of 16S rRNA encoding gene of Klebsiella quasipneumoniae strain EFTU115 isolated from preterm neoantes stool. The isolated Klebsiella quasipneumoniae strain EFTU115 phylogenetic tree shows the relationship with the closest neighbor strains of bacteria from NCBI. The accession numbers of 16S rRNA encoding genes of isolated Klebsiella quasipneumoniae strain EFTU115 is shown in boldface. The isolated Klebsiella quasipneumoniae strain EFTU115 and other bacterial strains neighborjoining tree was determined using 16S rDNA nearly full-length gene sequences and the frequency filter in the analysis package of MEGA 11 software. An out-group Bacillus subtilis JCM 1465 (accession number NR_113265) was used in the analysis. The scale bar shown indicates 2% estimated difference in sequence. The NCBI database accession numbers and strain names are shown in the phylogenetic tree and are also listed here: Klebsiella quasipneumoniae Kq-CW12, MK880381; Klebsiella quasipneumoniae. 2437, MT604862; Klebsiella quasipneumoniae A3GWS1-22(1)-E1, OR889330; Klebsiella quasipneumoniae EFTU115, PP177908; Klebsiella quasipneumoniae VITDJ2, MT826235; Klebsiella pneumoniae OOA, OR702992; Klebsiella pneumoniae ATCC 13883, NR_119278; Klebsiella pneumoniae UT09, OM978274; Enterobacter cloacae ATCC 23373, NR_118011; Citrobacter freundii ATCC 8090, NR_028894; Enterobacter hormaechei 0992-77, NR_042154; Escherichia coli S484, MK951738; Bacillus subtilis JCM 1465, NR_113265.

Figure 3.
Phylogenetic analysis of 16S rRNA encoding gene of Escherichia coli strains isolated from preterm neoantes stool. The isolated Escherichia coli strains phylogenetic tree shows the relationship with the closest neighbor strains of bacteria from NCBI. The accession numbers of 16S rRNA encoding genes of isolated Escherichia coli strains are shown in boldface. The isolated Escherichia coli strains and other bacterial strains neighborjoining tree was determined using 16S rDNA nearly full-length gene sequences and the frequency filter in the analysis package of MEGA 11 software. An out-group Bacillus subtilis JCM 1465 (accession number NR_113265) was used in the analysis. The scale bar shown indicates 2% estimated difference in sequence. The NCBI database accession numbers and strain names are shown in the phylogenetic tree and are also listed here: Escherichia coli EFTU7, PP188038; Escherichia coli EFTU8, PP188042; Escherichia coli FUA 1046, GQ222387; Escherichia coli S484, MK951738; Escherichia coli EGI292, MN704502; Escherichia coli EFTU9, PP188096; Escherichia coli LZ-11, OR717626; Escherichia coli UFV 478, MN557058; Escherichia coli EFTU2, PP157624; Escherichia coli EFTU1, PP156718; Escherichia coli EFTU6, PP187981; Escherichia coli EFTU4, PP182268; Escherichia coli EFTU5, PP187030; Escherichia coli 2p-64, KU682209; Klebsiella pneumoniae ATCC13883, NR_119278; Enterobacter cloacae ATCC 23373, NR_118011; Citrobacter freundii ATCC 8090, NR_028894; Enterobacter hormaechei 0992-77, NR_042154; Bacillus subtilis JCM 1465, NR_113265.
Figure 3.
Phylogenetic analysis of 16S rRNA encoding gene of Escherichia coli strains isolated from preterm neoantes stool. The isolated Escherichia coli strains phylogenetic tree shows the relationship with the closest neighbor strains of bacteria from NCBI. The accession numbers of 16S rRNA encoding genes of isolated Escherichia coli strains are shown in boldface. The isolated Escherichia coli strains and other bacterial strains neighborjoining tree was determined using 16S rDNA nearly full-length gene sequences and the frequency filter in the analysis package of MEGA 11 software. An out-group Bacillus subtilis JCM 1465 (accession number NR_113265) was used in the analysis. The scale bar shown indicates 2% estimated difference in sequence. The NCBI database accession numbers and strain names are shown in the phylogenetic tree and are also listed here: Escherichia coli EFTU7, PP188038; Escherichia coli EFTU8, PP188042; Escherichia coli FUA 1046, GQ222387; Escherichia coli S484, MK951738; Escherichia coli EGI292, MN704502; Escherichia coli EFTU9, PP188096; Escherichia coli LZ-11, OR717626; Escherichia coli UFV 478, MN557058; Escherichia coli EFTU2, PP157624; Escherichia coli EFTU1, PP156718; Escherichia coli EFTU6, PP187981; Escherichia coli EFTU4, PP182268; Escherichia coli EFTU5, PP187030; Escherichia coli 2p-64, KU682209; Klebsiella pneumoniae ATCC13883, NR_119278; Enterobacter cloacae ATCC 23373, NR_118011; Citrobacter freundii ATCC 8090, NR_028894; Enterobacter hormaechei 0992-77, NR_042154; Bacillus subtilis JCM 1465, NR_113265.
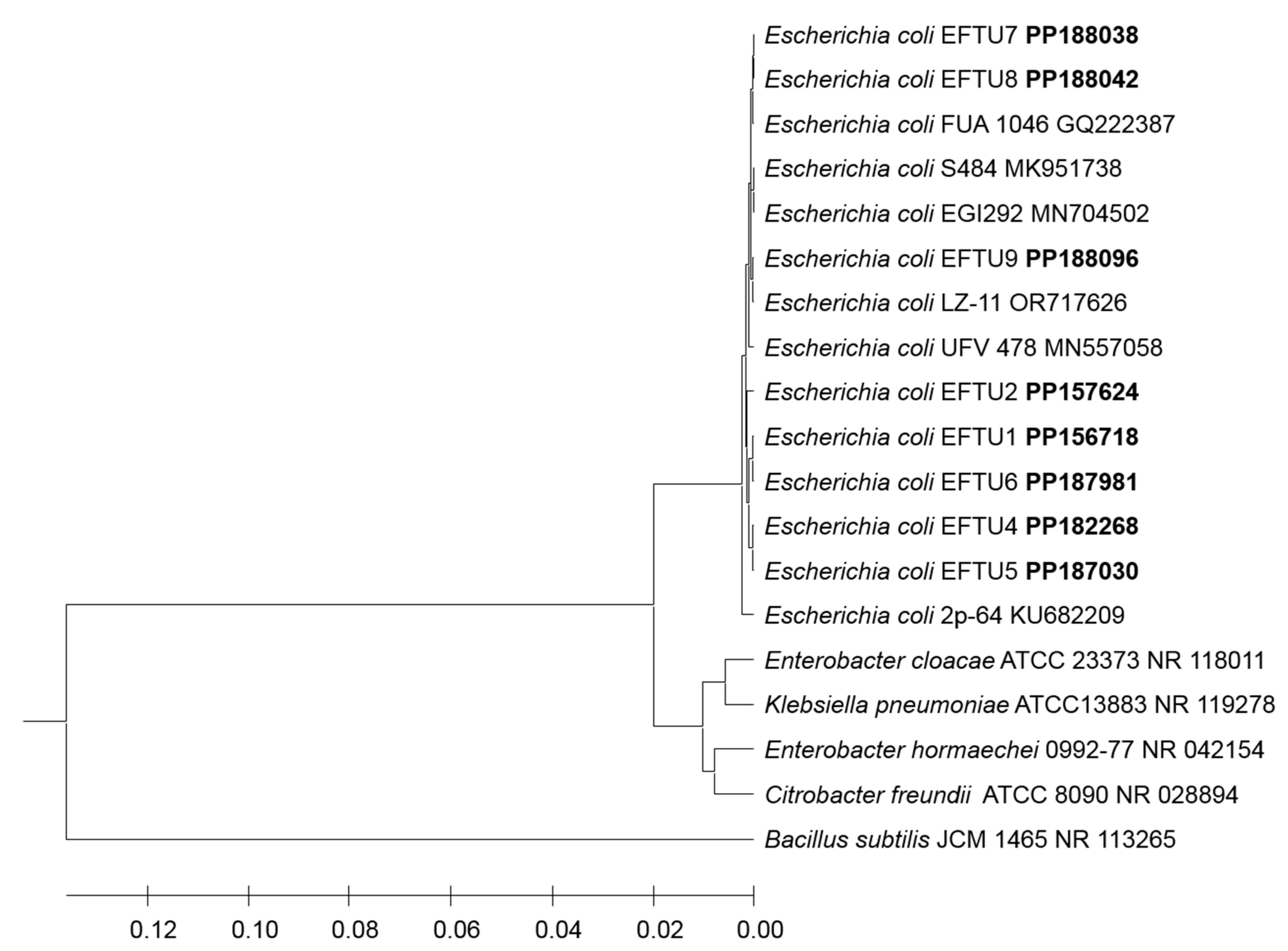
Figure 4.
Phylogenetic analysis of 16S rRNA encoding gene of Acinetobacter baumannii (a Non-Enterobacteriaceae bacterium) strains isolated from preterm neoantes stool. The isolated Acinetobacter baumannii strains phylogenetic tree shows the relationship with the closest neighbor strains of bacteria from NCBI. The accession numbers of 16S rRNA encoding genes of isolated Acinetobacter baumannii strains are shown in boldface. The isolated Acinetobacter baumannii strains and other bacterial strains neighborjoining tree was determined using 16S rDNA nearly full-length gene sequences and the frequency filter in the analysis package of MEGA 11 software. An out-group Bacillus subtilis JCM 1465 (accession number NR_113265) was used in the analysis. The scale bar shown indicates 2% estimated difference in sequence. The NCBI database accession numbers and strain names are shown in the phylogenetic tree and are also listed here: Acinetobacter baumannii C49, OQ405518; Acinetobacter baumannii Sn21, KM373317; Acinetobacter baumannii D4, MK799974; Acinetobacter baumannii EFTU111, PP177550; Acinetobacter baumannii HNNY-FY, OQ569359; Acinetobacter baumannii EFTU114, PP177613; Acinetobacter baumannii HNNY86G, OQ569357; Acinetobacter baumannii CGKV/U36d-2014, MK078538; Acinetobacter baumannii G21, OQ405624; Acinetobacter baumannii CR7, KR780411; Acinetobacter baumannii HAU425, OK483367; Acinetobacter baumannii rY25, MN173925; Acinetobacter baumannii B5, MG234437; Acinetobacter baumannii VrB1, LT963768; Moraxella catarrhalis Ne 11 ATCC 25238, NR_028669; Moraxella bovis L-3 ATCC 10900, NR_028668; Pseudomonas aeruginosa DSM 50071, NR_026078; Bacillus subtilis JCM 1465, NR_113265.
Figure 4.
Phylogenetic analysis of 16S rRNA encoding gene of Acinetobacter baumannii (a Non-Enterobacteriaceae bacterium) strains isolated from preterm neoantes stool. The isolated Acinetobacter baumannii strains phylogenetic tree shows the relationship with the closest neighbor strains of bacteria from NCBI. The accession numbers of 16S rRNA encoding genes of isolated Acinetobacter baumannii strains are shown in boldface. The isolated Acinetobacter baumannii strains and other bacterial strains neighborjoining tree was determined using 16S rDNA nearly full-length gene sequences and the frequency filter in the analysis package of MEGA 11 software. An out-group Bacillus subtilis JCM 1465 (accession number NR_113265) was used in the analysis. The scale bar shown indicates 2% estimated difference in sequence. The NCBI database accession numbers and strain names are shown in the phylogenetic tree and are also listed here: Acinetobacter baumannii C49, OQ405518; Acinetobacter baumannii Sn21, KM373317; Acinetobacter baumannii D4, MK799974; Acinetobacter baumannii EFTU111, PP177550; Acinetobacter baumannii HNNY-FY, OQ569359; Acinetobacter baumannii EFTU114, PP177613; Acinetobacter baumannii HNNY86G, OQ569357; Acinetobacter baumannii CGKV/U36d-2014, MK078538; Acinetobacter baumannii G21, OQ405624; Acinetobacter baumannii CR7, KR780411; Acinetobacter baumannii HAU425, OK483367; Acinetobacter baumannii rY25, MN173925; Acinetobacter baumannii B5, MG234437; Acinetobacter baumannii VrB1, LT963768; Moraxella catarrhalis Ne 11 ATCC 25238, NR_028669; Moraxella bovis L-3 ATCC 10900, NR_028668; Pseudomonas aeruginosa DSM 50071, NR_026078; Bacillus subtilis JCM 1465, NR_113265.
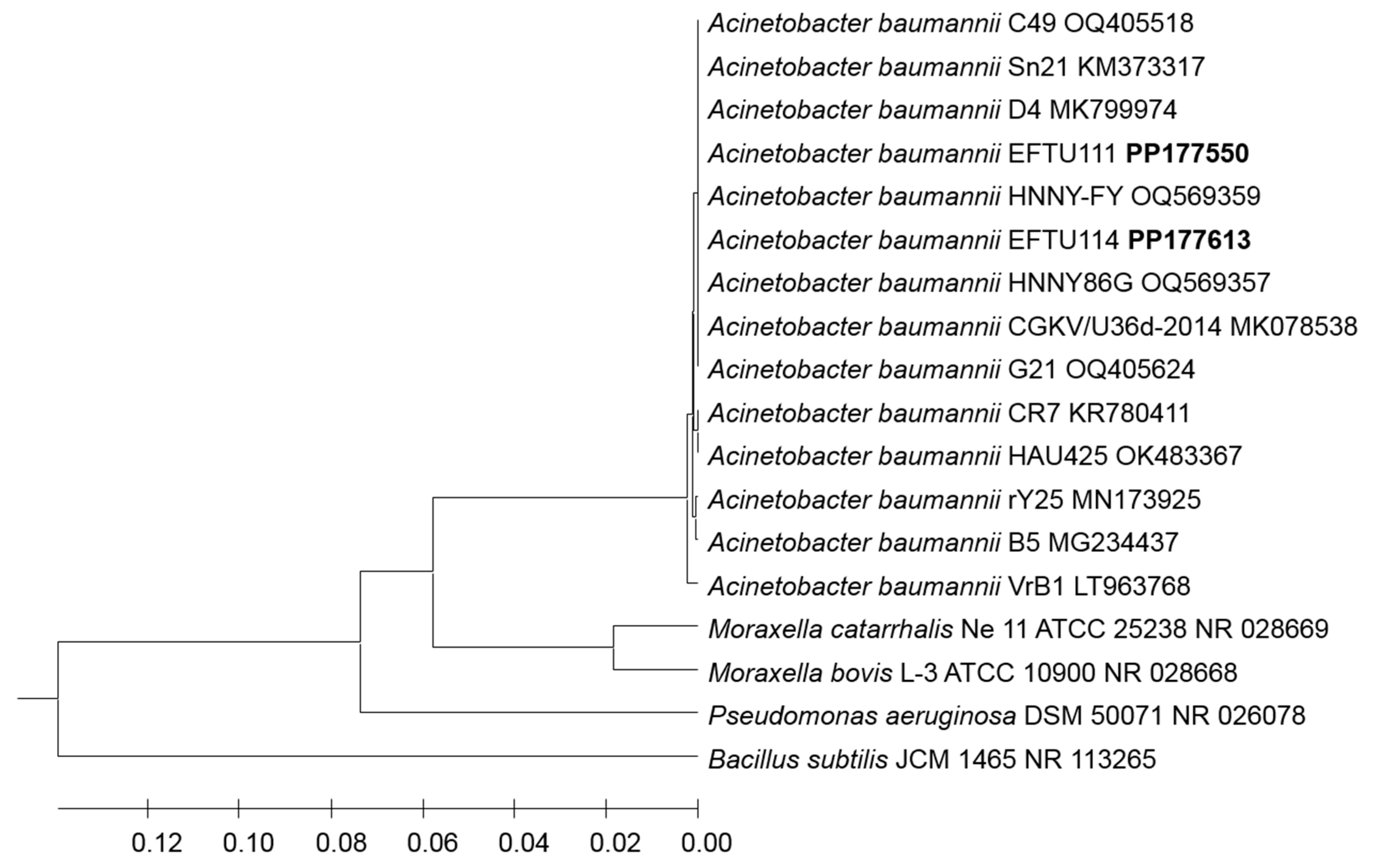

Table 1.
Colonization of colon by Klebsiella pneumoniae versus other Enterobacteria including the normal colon resident enterobacterium Escherichia coli as well as Acinetobacter baumannii (a Non-Enterobacteriaceae bacterium) in antibiotics intravenously treated preterm neonates.
Table 1.
Colonization of colon by Klebsiella pneumoniae versus other Enterobacteria including the normal colon resident enterobacterium Escherichia coli as well as Acinetobacter baumannii (a Non-Enterobacteriaceae bacterium) in antibiotics intravenously treated preterm neonates.
| Preterm Neonates subject # | Age days |
Gestational age (Weeks) | Types of antibiotics Treated with |
Bacterial strains Colonizing Preterm Neonates Colon | Accession number | |
|---|---|---|---|---|---|---|
| 1 st week | Case 3 | 2 | 31 | Ampicillin 50mg/ivq12h Cefotaxim50mg ivq12/h |
ND | - |
| Case 4 1st time | 5 | 32 | Gentamicin Ampicillin50mg/iv/q12h |
ND | - | |
| Case 5 1st time | 6 | 32 | Gentamicin Ampicillin50mg/iv/q12h |
ND | - | |
| Case 9 | 2 | 30 | Merobenem80mg q12h/iv Cefotaxime 80mg ivq12h |
ND | - | |
| Case 18 | 3 | 33 | Ampicillin 90Mg iv q 12h Gentamicin 8Mg iv q 36h |
ND | - | |
| Case 19 |
5 | 26 | Ampicillin 12Mg iv q 12h CEFOTAXIME --Mg iv q 48h |
ND | - | |
| > 1 week | Case 6 1st time | 8 | 28 | Ampicillin50mg/iv/q12h Cefotaxim50mg/iv/q12h |
ND | - |
| Case 1 | 22 | 27 | Vancomycin10mg/ivq36h | Escherichia coli EFTU1 | PP156718 | |
| Case 2 | 18 | 25 | Ampicillin 50mg/ivq12h Cefotaxim50mg ivq12/h Fucanazol13mg/ivq12 |
Klebsiella pneumoniae EFTU101 |
PP157088 | |
| Case 4 2nd time |
15 | 32 | Stop AB before 5 days |
E. coli EFTU2 |
PP157624 | |
| Case 5 2nd time | 16 | 33 | Gentamicin Ampicillin50mg/iv/q12h |
Klebsiella pneumoniae EFTU102 | PP177257 | |
| Case 6 2nd time | 18 | 28 | Ampicillin50mg/iv/q12h Cefotaxim50mg/iv/q12h |
Klebsiella pneumoniae EFTU103 |
PP177305 | |
| Case 7 | 44 | 26 | Vancomycin 10mg q12h/iv Merobenem20mg q12h/iv |
Klebsiella pneumoniae EFTU104 | PP177307 | |
| Case 8 | 44 | 26 | Vancomycin 10mg q12h/iv Merobenem20mg q12h/iv |
Klebsiella pneumoniae EFTU105 | PP177367 | |
| Case 10 | 44 | 29 | He was taking vancomycin 15mg iv q 12 h and stopping before 18day | Klebsiella pneumoniae EFTU3 | PP157908 | |
| Case 11 | 24 | 31 | No antibiotic | Escherichia coli EFTU4 | PP182268 | |
| Case 13 | 21 | 25 | Merobenem27mg q12h Colitin45mg q12h/iv |
Escherichia coli EFTU5 | PP187030 | |
| Case 14 | 16 | 33 | Gentamicin8mgivq36h Ampicillin76mg/iv/q12h |
Klebsiella pneumoniae EFTU110 | PP177465 | |
| Case 15 |
46 | 27 | Vancomycin 10mg q8h/iv Merobenem20mg q8h/iv |
Escherichia coli EFTU6 | PP187981 | |
| Klebsiella pneumoniae EFTU113 | PP204007 | |||||
| Acinetobacter baumannii EFTU111 | PP177550 |
|||||
| Case 16 | 29 | 29 | Ampicillin 65Mg iv q 12h Gentamicin 65Mg iv q 48h |
Acinetobacter baumannii EFTU114 | PP177613 |
|
| Case 17 | 32 | 29 | Ampicillin 60Mg iv q 12h CEFOTAXIME 60Mg iv q 48h |
Escherichia coli EFTU7 | PP188038 | |
| Klebsiella quasipneumoniae EFTU115 | PP177908 |
|||||
| Case 20 |
64 | 31 | meropenem 50Mg iv q 8h Colistin 50Mg iv q 8h |
Escherichia coli EFTU8 | PP188042 | |
| Klebsiella pneumoniae EFTU116 | PP177909 | |||||
| Case 21 | 14 | 31 | AMPCILLIN 67MGiv q 12h Gentamicin 6,7Mg iv q 36h stopping before six day |
Escherichia coli EFTU9 |
PP188096 |
ND, Not Detected.
Table 2.
In vitro antibiotics sensitivity of various Klebsiella pneumoniae and Klebsiella quasipneumoniae (Enterobacteriaceae ) strains and Acinetobacter baumannii (a Non-Enterobacteriaceae bacterium) colonizing preterm neonates’ colon.
Table 2.
In vitro antibiotics sensitivity of various Klebsiella pneumoniae and Klebsiella quasipneumoniae (Enterobacteriaceae ) strains and Acinetobacter baumannii (a Non-Enterobacteriaceae bacterium) colonizing preterm neonates’ colon.
| Bacterial strains | ESBL | Antibiotic | |||||||||||||||
| AMP | Amox | Tzp | CF | FOX | CAZ | CRO | FEP | ||||||||||
| MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | ||
| Klebsiella pneumoniae EFTU101 | P0S | >=32 | R | 4 | S | <=4 | S | >=64 | R | <=4 | S | 8 | R | >=64 | R | >=64 | R |
| Klebsiella pneumoniae EFTU102 | NEG | >=32 | R | <=2 | S | <=4 | S | <=2 | S | <=4 | S | <=1 | S | <=1 | S | <=1 | S |
| Klebsiella pneumoniae EFTU103 | NEG | >=32 | R | <=2 | S | <=4 | S | <=2 | S | <=4 | S | <=1 | S | <=1 | S | <=1 | S |
| Klebsiella pneumoniae EFTU104 | P0S | >=32 | R | 16 | i | 32 | i | >=64 | R | 16 | R | >=64 | R | >=64 | R | >=64 | R |
| Klebsiella pneumoniae EFTU105 | P0S | >=32 | R | 8 | S | <=4 | S | >=64 | R | <=4 | S | 8 | R | >=64 | R | 2 | R |
| Klebsiella pneumoniae EFTU3 | NEG | >=32 | R | 4 | S | <=4 | S | 16 | I | <=4 | S | <=1 | S | <=1 | S | <=1 | S |
| Klebsiella pneumoniae EFTU110 | NEG | >=32 | R | >=32 | R | ≥128 | R | >=64 | R | >=64 | R | 16 | R | >=64 | R | >=64 | R |
| Acinetobacter baumannii EFTU111 | NM | NM | - | NM | - | <=4 | S | NM | - | NM | - | 4 | S | 16 | R | 2 | S |
| Klebsiella pneumoniae EFTU113 | P0S | >=32 | R | >=32 | R | 3 | R | >=64 | R | <=4 | R | >=64 | R | >=64 | R | >=64 | R |
| Acinetobacter baumannii EFTU114 | NM | NM | - | NM | - | <=4 | S | NM | - | NM | - | 4 | S | 8 | R | 2 | S |
| Klebsiella quasipneumoniae EFTU115 | NM | NM | - | NM | - | <=4 | S | NM | - | NM | - | 4 | S | 8 | R | 2 | S |
| Klebsiella pneumoniae EFTU116 | NEG | >=32 | R | >=32 | R | 8 | S | >=64 | R | >=64 | R | 4 | S | <=1 | S | <=1 | S |
| Bacterial strains | Antibiotic | ||||||||||||||||
| IMI | MERO | AK | GM | CIP | TGC | FT | SXT | ||||||||||
| MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | ||
| Klebsiella pneumoniae EFTU101 | <=0,25 | S | <=0,25 | S | <=2 | S | <=1 | S | 0, 5 | S | 1 | S | 64 | I | 320 | R | |
| Klebsiella pneumoniae EFTU102 | <=0,25 | S | <=0,25 | S | <=2 | S | <=1 | S | <=0, 25 | S | =<0,5 | S | 32 | S | 20 | S | |
| Klebsiella pneumoniae EFTU103 | <=0,25 | S | <=0,25 | S | <=2 | S | <=1 | S | <=0, 25 | S | =<0,5 | S | 32 | S | 20 | S | |
| Klebsiella pneumoniae EFTU104 | <=0,25 | S | <=0,25 | S | <=2 | S | <=1 | S | 1 | S | 1 | S | 128 | R | >=320 | R | |
| Klebsiella pneumoniae EFTU105 | <=0,25 | S | <=0,25 | S | <=2 | S | <=1 | S | 1 | S | 2 | S | 64 | I | >=320 | R | |
| Klebsiella pneumoniae EFTU3 | <=0,25 | S | <=0,25 | S | <=2 | S | <=1 | S | <=0,25 | S | <=0,5 | S | <=16 | S | <=20 | S | |
| Klebsiella pneumoniae EFTU110 | >=16 | R | >=16 | R | ≥64 | R | <=1 | S | >=4 | R | 2 | S | 256 | R | 40 | S | |
| Acinetobacter baumannii EFTU111 | <=0,25 | S | <=0,25 | S | NM | - | <=1 | S | <=0,25 | S | <=0,5 | S | NM | - | <=20 | S | |
| Klebsiella pneumoniae EFTU113 | 2 | I | 2 | I | 4 | S | 2 | S | 2 | R | 2 | S | 128 | R | <=320 | R | |
| Acinetobacter baumannii EFTU114 | <=0,25 | S | <=0,25 | S | NM | - | <=1 | S | <=0,25 | S | <=0,5 | S | NM | - | <=20 | S | |
| Klebsiella quasipneumoniae EFTU115 | <=0,25 | S | <=0,25 | S | NM | - | <=1 | S | <=0,25 | S | <=0,5 | S | NM | - | <=20 | S | |
| Klebsiella pneumoniae EFTU116 | <=0,25 | S | <=0,25 | S | <=2 | S | <=1 | S | 1 | S | 2 | S | 128 | R | 320 | R | |
Interpretation, Intpr; Resistant, R; Sensitive, S; Intermediate, I; Minimum Inhibitory Concentration, MIC (µg/ml); Negative, Neg; Positive, Pos. Extended-spectrum beta-lactamases, ESBL; NM, Not measured; Ampicillin/Sulbactam, AMP; Amoxicillin/Clavulanic Acid, AMOX; Piperacillin/Tazobactam, TZP; Cefalotin, CF; Cefoxitin, FOX; Ceftazidime, CAZ; Ceftariaxone, CRO; Cefepime, FEP; Imipenem, IPM; Meropenem, MERO; Amikacin, AK; Gentamicin, GM; Ciprofloxacin, CIP; Tigecycline, TGC; Nitrofurantoin, FT; Trimethoprim/Sulfamethoxazole, SXT.
Table 3.
In vitro antibiotics sensitivity of various Escherichia coli strains colonizing preterm neonates’ colon.
Table 3.
In vitro antibiotics sensitivity of various Escherichia coli strains colonizing preterm neonates’ colon.
| Strains | ESBL |
Antibiotics | ||||||||||||||||||||||||||||
| AMP | Amox | Tzp | CF | FOX | CAZ | CRO | FEP | |||||||||||||||||||||||
| MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | |||||||||||||||
| E. coli EFTU 1 | NEG | >=32 | R | 4 | S | <=4 | S | 16 | I | <=4 | S | <=1 | S | <=1 | S | <=1 | S | |||||||||||||
| E. coli EFTU 2 | POS | >=32 | R | 4 | S | <=4 | S | >=64 | R | <=4 | S | 4 | R | <=64 | R | 2 | R | |||||||||||||
| E. coli EFTU 4 | NEG | >=32 | R | 4 | S | <=4 | S | 16 | I | <=4 | S | <=1 | S | <=1 | S | <=1 | S | |||||||||||||
| E. coli ETTU5 | NEG | <=2 | S | <=2 | S | <=4 | S | 4 | S | <=4 | S | <=1 | S | <=1 | S | <=1 | S | |||||||||||||
| E. coli EFTU6 | NEG | >=32 | R | 16 | I | <=4 | S | 16 | I | <=4 | S | <=1 | S | <=1 | S | <=1 | S | |||||||||||||
| E. coli EFTU7 | P0S | >=32 | R | 4 | S | <=4 | S | >=64 | R | <=4 | S | <=1 | R | >=64 | R | <=1 | R | |||||||||||||
| E. coli EFTU8 | P0S | >=32 | R | 4 | S | <=4 | S | >=64 | R | <=4 | S | 16 | R | >=64 | R | 8 | R | |||||||||||||
| E. coli EFTU9 | P0S | >=32 | R | 4 | S | <=4 | S | >=64 | R | <=4 | S | 16 | R | >=64 | R | >=64 | R | |||||||||||||
| Strains | Antibiotics | |||||||||||||||||||||||||||||
| IMI | MERO | AK | GM | CIP | TGC | FT | SXT | |||||||||||||||||||||||
| MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | MIC | Intpr | |||||||||||||||
| E. coli EFTU 1 | <=0,25 | S | <=0,25 | S | <=2 | S | <=1 | S | <=0,25 | S | <=0,5 | S | <=16 | S | <=20 | S | ||||||||||||||
| E. coli EFTU 2 | <=0,25 | S | <=0,25 | S | <=2 | S | <=1 | S | <=0,25 | S | <=0,5 | S | <=16 | S | <=20 | S | ||||||||||||||
| E. coli EFTU 4 | <=0,25 | S | <=0,25 | S | <=2 | S | <=1 | S | <=0,25 | S | <=0,5 | S | <=16 | S | <=20 | S | ||||||||||||||
| E. coli ETTU5 | <=0,25 | S | <=0,25 | S | <=2 | S | <=1 | S | <=0,25 | S | <=0, 5 | S | <=16 | S | <=20 | S | ||||||||||||||
| E. coli EFTU6 | <=0,25 | S | <=0,25 | S | <=2 | S | <=1 | S | <=0,25 | S | <=0,5 | S | <=16 | S | >=320 | R | ||||||||||||||
| E. coli EFTU7 | <=0,25 | S | <=0,25 | S | <=2 | S | <=1 | S | <=0,25 | S | <=0, 5 | S | <=16 | S | <=20 | S | ||||||||||||||
| E. coli EFTU8 | <=0,25 | S | <=0,25 | S | <=2 | S | <=1 | S | 0, 5 | S | <=0, 5 | S | <=16 | S | <=20 | S | ||||||||||||||
| E. coli EFTU9 | <=0,25 | S | <=0,25 | S | <=2 | S | <=1 | S | 0, 5 | S | <=0, 5 | S | <=16 | S | <=20 | S | ||||||||||||||
Interpretation, Intpr; Resistant, R; Sensitive, S; Intermediate, I; Minimum Inhibitory Concentration, MIC (µg/ml); Negative, Neg; Positive, Pos. Extended-spectrum beta-lactamases, ESBL; Ampicillin/Sulbactam, AMP; Amoxicillin/Clavulanic Acid, AMOX; Piperacillin/Tazobactam, TZP; Cefalotin, CF; Cefoxitin, FOX; Ceftazidime, CAZ; Ceftariaxone, CRO; Cefepime, FEP; Imipenem, IPM; Meropenem, MERO; Amikacin, AK; Gentamicin, GM; Ciprofloxacin, CIP; Tigecycline, TGC; Nitrofurantoin, FT; Trimethoprim/Sulfamethoxazole, SXT.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



