Submitted:
19 April 2024
Posted:
23 April 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Retroviruses perpetuate their survival by incorporating a copy of their genome into the host cell, a critical step catalyzed by the virally encoded integrase. The viral capsid plays an important role during the viral life-cycle, including nuclear importation in case of lentiviruses and integration targeting events, hence, targeting the integrase and the viral capsid is a favorable therapeutic strategy. While integrase strand transfer inhibitors (INSTIs) are recommended as first-line regimens given their high efficacy and tolerability, lenacapavir is the first capsid inhibitor and the newest addition to the HIV treatment arsenal. These inhibitors are however designed for treatment of HIV-1 infection, and their efficacy against HIV-2 remains widely understudied and inconclusive, supported only by few limited phenotypic susceptibility studies. We therefore carried out inhibition profiling of a panel of second-generation INSTIs and lenacapavir against HIV-2 in cell culture, utilizing pseudovirion inhibition profiling assays. Our results show that the tested INSTIs and lenacapavir exerted excellent efficacy against ROD-based HIV-2 integrase, and interestingly, raltegravir also showed modest antiviral activity against the SARS-CoV-2 Omicron variant in antiviral assays carried out in Vero E6 cells.
Keywords:
HIV-2
; integrase strand transfer inhibitor (INSTIs)
; HIV
; integrase
; Lenacapavir
; SARs-CoV-2
1. Introduction
Of the approximately 39 million people living with HIV worldwide, HIV-1 infection accounts for the majority of the cases [1]. Statistical data on the prevalence of HIV-2 remain largely unknown, with earlier estimations over a decade ago suggesting that around 2-3 million people are infected with HIV-2, a descendent of the simian immunodeficiency virus of sooty mangabeys (SIV/sm), which is endemic in West Africa [2]. Since then, significant number of cases have been reported in Europe, most prominently in France, Portugal, Spain and Italy [3], owing to colonial and economic ties, and migration from central and Western African countries [4]. Compared to HIV-1, infection with HIV-2 is characterized by protracted latency, lower viral load, and higher CD4+ cell count, however, in due time, the infection may progress to AIDS if left untreated; albeit at a much slower rate than that of its counterpart [5].
Combined antiretroviral therapy (ART) targeting different steps in the viral life-cycle has revolutionized the management of HIV infection, rendering it a chronic, yet manageable infection. Daily medication burden and compliance are however major issues resulting in treatment failure and drug resistance [6]. This has prompted the development of long-acting oral or injectable drugs with potential for monotherapy, and allow for pre-exposure prophylaxis and reduced intake intervals. Such recently approved drugs include cabotegravir and lenacapavir, belonging to the integrase strand transfer inhibitors (INSTIs) and capsid inhibitors (CIs) groups, respectively [7].
To date, all of the approved drugs were designed for HIV-1, and their efficacy against HIV-2 remains largely unknown. Bearing in mind the presence of inherent polymorphisms in its genetic sequence and difference in its replication dynamics, analyzing the efficacy of ART in the context of HIV-2 is indeed important. It was shown that HIV-2 is intrinsically resistant to non-nucleoside reverse transcriptase inhibitors [8], and many protease inhibitors [9], while being widely susceptible to nucleoside reverse transcriptase inhibitors (NRTIs) and INSTIs [10].
The retroviral integrase is derived from the Gag-Pol polyprotein as a 32 kDa protein composed of three functional domains: the amino zinc binding terminal domain comprising of histidine and cysteine residues, the DNA binding carboxyl domain, and the enzyme active central catalytic core [11]. The integrase mediates the 3′-end processing and strand transfer, which are key steps in integration. Conversely, INSTIs compete for the enzyme active binding sites including a divalent ion impeding merging of viral and host cell DNA and thereby, blocking integration [12]. To date, five INSTIs are approved for clinical use, classified into first-generation inhibitors: raltegravir and elvitegravir, and the second-generation ones: dolutegravir, bictegravir, & cabotegravir [13]. Initially, INSTIs exhibited potent efficacy against limited strains of HIV-2, however, this was short lived as resistance-associated mutations emerged in raltegravir and elvitegravir-treated patients [14]. Second-generation inhibitors soon followed, with a change from bicyclic to tricyclic ring at the central pharmacore of the drugs, boosting their potency. Dolutegravir showed higher stability, tolerability and efficacy against mutants resistant to raltegravir and elvitegravir, as reported in an in vitro study against HIV-2 in patients with resistance to first generation inhibitors [15]. A five amino acid insertion at the carboxy-terminal domain of HIV-2 integrase resulted in high level resistance to raltegravir and elvitegravir, and moderate resistance to dolutegravir [16,17,18]. Bictegravir showed high potency against HIV-2 in a single cycle assay, with EC50 between 1 nM – 5 nM, although, there is no published data on its clinical use in treatment-naïve individuals with HIV-2 infection [19]. Cabotegravir, approved in 2021 for use in combination with rilpivirine as a once daily oral tablet, or as a long-acting monthly injection, was reported to have an EC50 of 0.12 nM against four HIV-2 isolates [20]. A collection of studies analyzing efficacy of INSTIs against HIV-2 and their reported IC50/EC50 values is presented in supplementary Table S1.
The novel capsid inhibitor lenacapavir was approved in 2022 as a long-acting injectable inhibitor streamlined for use in patients failing multiple therapy and as pre-exposure prophylaxis (PrEP). Lenacapavir targets multiple phases of early and late viral life-cycle events by blocking nuclear import of pre-integration complex, proper capsid core formation and production of new virions [21]. Ongoing trials in HIV-1 patients reported its effectiveness at very low dosage in both treatment experienced and naïve patients. At the time of writing of this manuscript, there were no planned clinical trials for HIV-2 infected patients to assess its efficacy against HIV-2. To our knowledge, it has only been assessed in two clinical isolates from human peripheral blood mononuclear cells with an EC50 value of 885 pM [22].
On a separate note, in the midst of the coronavirus disease of 2019 (COVID-19) pandemic, HIV antiretroviral drugs became, amongst others, the focus of extensive research, given the lack of novel drugs against the severe acute respiratory syndrome -2 virus (SARS-CoV-2) at the time. Among the compounds evaluated against the SARs-CoV-2 main protease (mPro) and RNA-dependent RNA polymerase were the protease [23] and integrase inhibitors raltegravir and dolutegravir. Based on the binding energies, molecular docking studies reported raltegravir as a promising candidate for the treatment of COVID-19 [24], which prompted the need for further investigations to consolidate this finding.
Susceptibility of HIV-2 to the inhibitors remains widely understudied and a treatment protocol is yet to be established. Very limited number of studies were done to assay for the inhibitors against HIV-2, moreover, the majority of those studies were phenotypic assays using virions isolated from patients experiencing virological failure to ART or examined the efficacy of inhibitors in INSTIs-based regimens [25,26,27]. The lack of a standardized methodology to assay for the inhibitor’s efficacy further hinders the interpretation of the results; therefore, we set out to test the efficacy of lenacapavir and the INSTIs against ROD-based HIV-2 pseudovirions in cell culture-based inhibition profiling assays, analyzing the inhibitory potential of the inhibitors in the context of a unified integrase sequence. Furthermore, utilizing computational methods of molecular docking, we created a model structure of HIV-2 integrase and analyzed its binding with bictegravir and cabotegravir. Additionally, in order to verify the findings of previous in silico studies, we analyzed the efficacy of raltegravir against the ancestral (Wuhan) and omicron variants of SARS-CoV-2.
2. Materials and Methods
2.1. Plasmid Vectors and Inhibitors
We utilized 2nd generation lentiviral vectors for the production of HIV-1 and HIV-2 pseudovirions. For HIV-1, we used psPAX2 as a packaging plasmid, a kind gift from Dr. D. Trono (University of Geneva Medical School, Geneva, Switzerland); a modified pWOX expressing mCherry as transfer vector [28], and pMDG; encoding for the envelope protein of vesicular stomatitis virus (VSV-G). For HIV-2, the plasmids included HIV-2 CGP (a ROD-based HIV-2 protein expression vector encoding HIV-2 genes), HIV-2 CRU5SINCSW; a transfer plasmid, and pMDG plasmid [29]. HIV-2 CRU5SINCSW was modified to encode mCherry fluorescent protein under a CMV promoter in between BamHI and ECoRI restriction sites. Success of cloning and transduction efficiency were confirmed by restriction reactions, PCR, and transduction assays. HIV-2 vectors were a kind gift from Joseph P. Dougherty from the Robert Wood Johnson Medical School, (NJ, USA).
All the inhibitors were obtained from MedChem Express (MCE, NJ, USA) and diluted in DMSO in concentrations ranging from 1 nM to 1 µM.
2.2. Production of HIV-1 and HIV-2 Pseudovirions
We carried out pseudovirion production in 293T human embryonic kidney (HEK-293T) cells (Invitrogen, Carlsbad, USA). On day 1, the cells were seeded in T-75 flask in 15 ml full DMEM containing 10 % fetal bovine serum (FBS), 1 % L-glutamine and 1 % penicillin-streptomycin. Cells achieved about 70 % confluency (5-6×106 cells/ml) after 24 hrs. The following day, transfection was carried out using polyethylenimine (PEI) (Sigma-Aldrich, St. Louis, Missouri Unites States) and 10 µg of the plasmids and the transfected cells were incubated at 37 °C, 5 % CO2 in 5 ml antibiotics free DMEM containing 1% FBS after adding the PEI-DNA solution for 5-6 hours. Thereafter, the media was replaced with 10 ml full DMEM and the supernatant containing virions was collected after 24 hrs for three consecutive days and filtered through a 0.45 μm polyvinylidene fluoride filter (Merck Millipore, Darmstadt, Germany). The collected supernatant was concentrated by ultracentrifugation (100 000 x g, 2 hours, 4 °C), and the viral pellet was collected in 200 μl phosphate-buffered saline (PBS), aliquoted, and stored at -70 °C. ELISA-based colorimetric reverse transcriptase (RT) assay to measure the RT activity of the viruses (Roche Applied Science, Mannheim), was then used to measure the concentration of the pseudovirions in a nanograms of reverse-transcriptase (RT)/well unit.
2.3. Virus Production to Assay for Effects of Lenacapavir on HIV-2 Capsid Formation
Using the same protocol as above, viruses were produced in the presence of different concentrations of lenacapavir before transduction experiments. On day 1, three million HEK-293T cells were seeded in T-75 flasks in 10 ml full DMEM. On day 2, the cells were treated for 3 hrs with 1 nM, 10 nM and 100 nM lenacapavir and thereafter transfected with 10 µg of HIV-1 and HIV-2 vectors in 5 ml of 1 % DMEM. After 5 hrs of incubation, medium was changed with fresh full DMEM supplemented with the same concentrations of lenacapavir, followed by incubation for two days. The virion-containing supernatant was then collected and concentrated with Amicon Ultracel 100K (Merck Millipore Ltd., Tullagreen, Ireland), and ELISA-based colorimetric RT assay was used to measure concentration of virions, followed by transduction assays.
2.4. MTT Cell Proliferation Assay
96-well plates were seeded with 35,000 HEK-293T cells in full DMEM (10 % FBS, 1 % L-glutamine, 1 % penicillin/streptomicin). The following day, cells were treated with a serial dilution of the inhibitors and were incubated for two days. MTT (Thermo Fisher Scientific, Massachusetts United States) assay was then carried out to assess viability of the treated cells following the manufacturer’s protocol.
2.5. Inhibition Profiling in Jurkat Cells
48-well plates were seeded with 35 000 Jurkat cells (ATCC, Virginia United States) in 200 µl of RPMI medium complemented with 10 % FBS, 1 % L-glutamine) to reach a 50-70 % confluency the next day. Cells were then treated with a serial dilution of the inhibitors ranging from 1 nM to 1 µM of INSTIs and 100 pMol to 100 nM of lenacapavir in fresh medium without antibiotics and incubated at 37 °C for 3 hours, thereafter, the cells were transduced with 4 ng equivalent to RT/well activity of HIV-1 or HIV- 2 in the presence of 8 µg/ml polybrene/well, and further incubated at 37 °C for 48 hours. Cells were collected in 400 µl cold sterile PBS and quantitative analysis of the transduction efficiency in the presence of the inhibitors was assessed by flow cytometry (FACS caliber, BD Biosciences, NJ, USA). The results were analyzed by FlowJo Software Version 10 (Becton, Dickinson and Company; 2019) to determine the percentage of GFP or mCherry fluorescence in 5000 cells, indicating transduction by HIV-2 or HIV-1 respectively. GraphPad Prism 5.0 (GraphPad Software, Inc) performed the IC50 calculations and graphs.
2.6. Raltegravir Inhibition Assay against SARs-CoV-2
VeroE6-GFP cells (provided by M. van Loock, Janssen Pharmaceutica, Beerse, Belgium) were seeded at a density of 25 000 cells/well in 96-well plates (Greiner Bio One, catalogue no. 655090) and pre-treated with three-fold serial dilutions of raltegravir overnight in presence of the MDR1-inhibitor CP-100356 (final concentration 0.5 μM). On the next day (day 0), cells were infected with the SARS-CoV-2 inoculum at a multiplicity of infection (MOI) of 0.001 median tissue culture infectious dose (TCID50) per cell. The number of fluorescent pixels of GFP signal determined by High-Content Imaging (HCI) on day 4 post-infection (p.i.) was used as a read-out. Percentage of inhibition was calculated by subtracting background (number of fluorescent pixels in the untreated-infected control wells) and normalizing to the untreated-uninfected control wells (also background subtracted). The 50 % effective concentration (EC50, the concentration of raltegravir required for fifty percent recovery of cell-induced fluorescence) was determined using logarithmic interpolation. A similar protocol was used to determine antiviral activity in A549-Dual™ hACE2-TMPRSS2 cells, but no MDR1-inhibitor CP-100356 was used and the cell viability was determined 4 days p.i. using viability staining with MTS (3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium). The percentage of antiviral activity was calculated by subtracting the background and normalizing to the untreated-uninfected control wells, and the EC50 was determined using logarithmic interpolation. In both cell lines, potential toxicity of raltegravir was assessed in a similar set-up in treated-uninfected cultures where metabolic activity was quantified at day 5 using the MTS assay as described earlier [30]. The 50 % cytotoxic concentration (CC50, the concentration at which cell viability reduced to 50 %) was calculated by logarithmic interpolation.
SARS-CoV-2 GHB (EPI ISL407976|2020-02-03) was recovered from a nasopharyngeal swab taken from an asymptomatic patient returning from Wuhan, China. Virus stocks were inoculated and passaged first in HuH-7 cells and then passaged 7 times on VeroE6-eGFP cells. GHB-03021 has a ΔTQTNS deletion at 676–680 residues that is typical for SARS-CoV-2 strains that have been passaged several times on VeroE6 cells (Boudewijns et al. 2020).
SARS-CoV-2 variants B.1.1.7 (GISAID ID: EPI_ISL_791333) and Omicron BA.2 (GISAID ID: EPI_ISL_10654979) were recovered from nasopharyngeal swabs of RT-qPCR-confirmed human cases. Virus stocks were generated by passaging the virus in Calu-3 cells followed by production of a screening virus stock on A549+hACE2+hTMPRSS2 cells.
All SARS-CoV-2 manipulations were performed in biosafety level 3 (BSL-3) and 3+ (CAPs-IT) facilities at the Rega Institute for Medical Research, KU Leuven, according to institutional guidelines.
2.7. Molecular Docking
We modeled HIV-2 integrase structure based on the available Simian immunodeficiency virus red-capped mangabeys (SIVrcm) intasome structure in complex with bictegravir and DAN substrate using SWISS-MODEL (PDB ID: 6RWM) [31]. The original SIV DNA sequence was modified to represent our available HIV-2 sequence. Nucleotide transmutation was carried out with Chimera [32], which was also applied to examine the histidine protonation states. The protonation states of titratable residues were evaluated with PROPKA [33,34]. The structure was minimized with the Amber16 software [35,36,37], with ff14SB [38] and DNA OL15 [39] force fields applied, also applying the Li/Merz ion parameters (12-6-4 set) for the divalent Mg2+ ions [40], along with solvation in TIP3P [41] water molecules. Ions were added to approximate a 0.15 M NaCl concentration, modified in accordance with the charge split protocol [42]. Bictegravir and cabotegravir inhibitors were optimized with Gaussian09 software [43], applying the M06-2X [44] method and the 6-311+G (d, p) basis set for optimization, followed by RESP [45] charge calculation with antechamber [46]. Docking was carried out with the PLANTS [47,48] software, to subunit A of the integrase complex, that also included the bound DNA strands, the Zn2+ ion, the Mg2+ ions and three water molecules coordinated by the Mg2+ ions. The center of the docking was defined as the geometric point halfway between the Mg2+ ions. The binding site radius was 12 Å. The structure with lowest energy was accepted.
3. Results
3.1. Susceptibility of HIV-2 to INSTIs
In this work, using a standard protocol, we confirmed that all the integrase inhibitors were effective against ROD-based HIV-2 integrase at low nanomolar concentrations (Table 1 and Figure 1), similarly to HIV-1, and as reported elsewhere in the literature for some HIV-2 isolates in clinical and in vitro cell culture studies.
3.2. In Silico Analysis
We designed a HIV-2 integrase model, based on SIV integrase, and applying the PLANTS molecular docking software we successfully docked bictegravir and cabotegravir at the integrase active sites. Visual comparison of the docked bictegravir conformation with the bictegravir bound in SIV intasome (PDB ID: 6RWM) showed significant resemblance, confirming the possibility of inhibitor binding at the active site.
Results of the docking are presented in Figure 2.
3.3. Efficacy of Lenacapavir against HIV-2
We evaluated the inhibitory effects of lenacapavir on HIV-2 transduction efficiency and virion formation. Following treatment of cells with the inhibitor, we observed that lenacapavir inhibited pseudovirion transduction in picomolar concentration, as previously shown for HIV-1. In order to assess the inhibitory effects of lenacapavir against formation of HIV-2 pseudovirions, we produced the virions in the presence of various concentrations of lenacapavir (1, 10 and 100 nM). RT ELISA-based colorometric assay was carried out on the produced virions to assess for the activity, and we did not detect significant difference in RT activity between HIV-2 virions produced in the presence of lenacapavir and that of the control. However, when we transduced cells with pseudovirions produced in the presence of lenacapavir, transduction efficiency was severely compromized (Figure 3), indicating that the virions were defective despite having normal RT activity.
3.4. Anti-SARS-CoV-2 Activity of Raltegravir
Inhibition profiling of raltegravir in Vero E6 cells and A549 cells against SARS-CoV-2 showed no antiviral activity against the prototypical Wuhan-Hu-1 (GBH) (IC50 > 100 uM), while exerting a modest yet noticable inhibition of the Omicron BA.2 variant (IC50 35.9 +/- 2.7) (Figure 4).
4. Discussion
In our study, we conducted cell culture-based inhibition profiling assays of INSTIs against ROD-based HIV-2 psedovirions, comparing it to HIV-1. Our results show that all of the commercially available INSTIs exhibited efficacy against HIV-2 in nanomolar concentration. Raltegravir inhibited HIV-2 integrase with an IC50 value of 2.1 nM, comparable to an in vitro phenotypic study done soon after its approval against HIV-2 ROD clinical isolates [49]. Several clinical trials have demonstrated the effectiveness of raltegravir in suppressing viral load in treatment-naïve and experienced HIV-2 infected patients, although, N155H resistance mutation quickly emerged [32]. Elvitegravir was shown to have an IC50 of 0.7 nM [49], comparable to a cell culture study by Zheng et al. that reported elvitegravir to have an IC50 ranging from 0.3-0.9 nM in clinical isolates from integrase naïve patients [50]. In our study, elvitegravir inhibited HIV-2 with an IC50 of 2.6 nM which was slightly higher but nonetheless comparable.
Dolutegravir was reported to be an active integrase inhibitor in the treatment-naïve patients [15], and similary, an observational study in India showed that INSTIs naïve HIV-2 patients on dolutegravir regimen had undetectable viral load [17]. In our study, dolutegravir showed inhibition with an IC50 of 1.1 nM. Bictegravir, cabotegravir and lenacapavir are the latest approved inhibitors, and there is limited data on their efficacy against HIV-2. Based on dolutegravir, bictegravir was shown to be very potent at inhibiting HIV-1, resulting in low rate of resistance mutations compared to raltegravir, elvitegravir and dolutegravir [50]. In a recently published in vitro study, HIV-2 was sensitive to cabotegravir with low EC50 of 1.8 - 2.6 nM in single cycle spreading infection assay [20]. A study by Hingrat et. al. reported bictegravir to be more superior to cabotegravir [16]. In our study, bictegravir and cabotegravir were both effective against HIV-2 ROD-based pseudovirions with an IC50 of 1.8 nM and 0.8 nM, respectively.
As the only one of its class, lenacapavir is the latest addition to the arsenal in the fight against HIV, inhibiting the early phase of viral life cycle and capsid formation [51]. Lenacapavir was effective against HIV-1 in our study with an IC50 of 89 pM, comparable to cell culture studies in HIV-1 which reported a potency in the picomolar range (EC50 of 50-314 pM) [52]. In vitro cell culture studies by Link O. et. al. reported an EC50 of 105 pM against HIV-1 in MT-4 cells, 56 pM in macrophages, 32 pM in primary CD4 cells, and in 23 clinical HIV-1 isolates ranging from 20 - 160 pM in human peripheral blood mononuclear cells. In addition, this study also reported efficacy of the inhibitor against two HIV-2 isolates [22]. At the time of writing this manuscript, that was the only study evaluating the efficacy of lenacapavir against HIV-2. In our study, lenacapavir inhibited HIV-2 with an IC50 of 50.6 pM, a similar efficacy to that exibited agasint HIV-1.
Molecular docking of bictegravir and cabotegravir to our HIV-2 integrase model provides theoretical evidence of the inhibitors binding to the enzyme. Structural analysis suggests that the active site Mg2+ ions have a crucial role in inhibitor binding, forming interactions that were described as almost covalent between bictegravir and the SIVrcm integrase [49]. In that study, resistance mutations of SIVrcm integrase were shown to modulate the charge distribution of the Mg2+ ions, raising the possibility of similar effects in HIV-2, as well as HIV-1 integrase. Given the lack of studies on resistance mutations in the case o HIV-2 integrase, it would be important to analyse the effects of the mutations in the context of this coordinated binding to the Mg2+ ions in future studies.
Conclusion
To our knowledge, no study has evaluated the efficacy of all of the available INSTIs against HIV-2 integrase. Our analysis is perhaps advantegeous because we analyzed the efficacy of the inhibitors against the same ROD-based integrase sequence, utilizing a standardized cell culture-based methodology, without influence or interference from other drugs. Only a few studies have reported on the efficacy of bictegravir, cabotegravir against HIV-2, and in regards to lenacapavir, it was only evaluated against two isolates. On a separate note, rather surprisingly, raltegravir was shown to exhibit a modest, yet consistent inhibition of the Omicron variant of SARS-CoV-2. Although the IC50 observed was beyond concentrations normally achievable in the serum, further exploration of the mechanistic features of this inhibition deserves further attention, given the limed availabilty of novel inhibitors against SARS-CoV-2.
Author Contributions
IWK: methodology, investigation, writing original draft; GH: methodology, software, writing original draft; ML, PL, JN: investigation, writing original draft; JT: resources, project administration, finding, review and editing of manuscript; MM: conceptualization, analysis, investigation, supervision, review and editing of manuscript.
Funding
This work was supported by the Thematic Excellence Programme TKP2021-EGA-20 (Biotechnology) of the Ministry for Innovation and Technology in Hungary and Stipendium Hungaricum scholarship, and by ERINHA through the ERINHA-Advance project (funding from the European Union’s Horizon 2020 Research & Innovation Program, Grant agreement no. 824061). ÚNKP-23-3-II-DE-456 funding by the New National Excellence Program of the Ministry of Culture and Innovation (to G. H.) was also utilized. We acknowledge the Digital Success Backbone Network (DJG) developed and maintained by the KIFÜ for awarding us access to resource based in Hungary.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All required data are available in the manuscript. Any additional data can be provided upon request.
Acknowledgments
We are grateful to Joseph P. Dougherty from the Robert Wood Johnson Medical School, NJ, USA for providing us with the HIV-2 vectors. Authors would like to extend their gratitude to the staff of the Laboratory of Retroviral Biochemistry for their continued support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- UNAIDS. Global HIV/AIDS Data and Statistics. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on.
- Campbell-Yesufu, O.T.; Gandhi, R.T. Update on human immunodeficiency virus (HIV)-2 infection. Clinical infectious diseases: an official publication of the Infectious Diseases Society of America 2011, 52, 780–787. [Google Scholar] [CrossRef]
- Gottlieb, G.S.; Raugi, D.N.; Smith, R.A. 90-90-90 for HIV-2? Ending the HIV-2 epidemic by enhancing care and clinical management of patients infected with HIV-2. The lancet. HIV 2018, 5, e390–e399. [Google Scholar] [CrossRef]
- Ceccarelli, G.; Giovanetti, M.; Sagnelli, C.; Ciccozzi, A.; d’Ettorre, G.; Angeletti, S.; Borsetti, A.; Ciccozzi, M. Human Immunodeficiency Virus Type 2: The Neglected Threat. Pathogens 2021, 10. [Google Scholar] [CrossRef]
- Esbjornsson, J.; Mansson, F.; Kvist, A.; da Silva, Z.J.; Andersson, S.; Fenyo, E.M.; Isberg, P.E.; Biague, A.J.; Lindman, J.; Palm, A.A.; et al. Long-term follow-up of HIV-2-related AIDS and mortality in Guinea-Bissau: a prospective open cohort study. The lancet. HIV 2018. [Google Scholar] [CrossRef]
- Nachega, J.B.; Marconi, V.C.; van Zyl, G.U.; Gardner, E.M.; Preiser, W.; Hong, S.Y.; Mills, E.J.; Gross, R. HIV treatment adherence, drug resistance, virologic failure: evolving concepts. Infectious disorders drug targets 2011, 11, 167–174. [Google Scholar] [CrossRef]
- Slama, L.; Porcher, R.; Linard, F.; Chakvetadze, C.; Cros, A.; Carillon, S.; Gallardo, L.; Viard, J.P.; Molina, J.M. Injectable long acting antiretroviral for HIV treatment and prevention: perspectives of potential users. BMC infectious diseases 2023, 23, 98. [Google Scholar] [CrossRef]
- Witvrouw, M.; Pannecouque, C.; Van Laethem, K.; Desmyter, J.; De Clercq, E.; Vandamme, A.M. Activity of non-nucleoside reverse transcriptase inhibitors against HIV-2 and SIV. Aids 1999, 13, 1477–1483. [Google Scholar] [CrossRef]
- Desbois, D.; Roquebert, B.; Peytavin, G.; Damond, F.; Collin, G.; Benard, A.; Campa, P.; Matheron, S.; Chene, G.; Brun-Vezinet, F.; et al. In vitro phenotypic susceptibility of human immunodeficiency virus type 2 clinical isolates to protease inhibitors. Antimicrobial agents and chemotherapy 2008, 52, 1545–1548. [Google Scholar] [CrossRef] [PubMed]
- Menendez-Arias, L.; Alvarez, M. Antiretroviral therapy and drug resistance in human immunodeficiency virus type 2 infection. Antiviral research 2014, 102, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Craigie, R. The molecular biology of HIV integrase. Future virology 2012, 7, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Jozwik, I.K.; Passos, D.O.; Lyumkis, D. Structural Biology of HIV Integrase Strand Transfer Inhibitors. Trends in pharmacological sciences 2020, 41, 611–626. [Google Scholar] [CrossRef]
- Trivedi, J.; Mahajan, D.; Jaffe, R.J.; Acharya, A.; Mitra, D.; Byrareddy, S.N. Recent Advances in the Development of Integrase Inhibitors for HIV Treatment. Current HIV/AIDS reports 2020, 17, 63–75. [Google Scholar] [CrossRef]
- Smith, R.A.; Raugi, D.N.; Pan, C.; Coyne, M.; Hernandez, A.; Church, B.; Parker, K.; Mullins, J.I.; Sow, P.S.; Gottlieb, G.S.; et al. Three main mutational pathways in HIV-2 lead to high-level raltegravir and elvitegravir resistance: implications for emerging HIV-2 treatment regimens. PloS one 2012, 7, e45372. [Google Scholar] [CrossRef]
- Descamps, D.; Peytavin, G.; Visseaux, B.; Tubiana, R.; Damond, F.; Campa, P.; Charpentier, C.; Khuong-Josses, M.A.; Duvivier, C.; Karmochkine, M.; et al. Dolutegravir in HIV-2-Infected Patients With Resistant Virus to First-line Integrase Inhibitors From the French Named Patient Program. Clinical infectious diseases: an official publication of the Infectious Diseases Society of America 2015, 60, 1521–1527. [Google Scholar] [CrossRef] [PubMed]
- Le Hingrat, Q.; Collin, G.; Le, M.; Peytavin, G.; Visseaux, B.; Bertine, M.; Tubiana, R.; Karmochkine, M.; Valin, N.; Collin, F.; et al. A New Mechanism of Resistance of Human Immunodeficiency Virus Type 2 to Integrase Inhibitors: A 5-Amino-Acid Insertion in the Integrase C-Terminal Domain. Clinical infectious diseases: an official publication of the Infectious Diseases Society of America 2019, 69, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Pujari, S.; Patel, A.; Gaikwad, S.; Patel, K.; Dabhade, D.; Chitalikar, A.; Joshi, K.; Bele, V. Effectiveness of dolutegravir-based antiretroviral treatment for HIV-2 infection: retrospective observational study from Western India. The Journal of antimicrobial chemotherapy 2020, 75, 1950–1954. [Google Scholar] [CrossRef]
- Smith, R.A.; Raugi, D.N.; Pan, C.; Sow, P.S.; Seydi, M.; Mullins, J.I.; Gottlieb, G.S.; University of Washington-Dakar, H.I.V.S.G. In vitro activity of dolutegravir against wild-type and integrase inhibitor-resistant HIV-2. Retrovirology 2015, 12, 10. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.A.; Raugi, D.N.; Wu, V.H.; Zavala, C.G.; Song, J.; Diallo, K.M.; Seydi, M.; Gottlieb, G.S.; University of Washington-Dakar, H.I.V.S.G. Comparison of the Antiviral Activity of Bictegravir against HIV-1 and HIV-2 Isolates and Integrase Inhibitor-Resistant HIV-2 Mutants. Antimicrobial agents and chemotherapy 2019, 63. [Google Scholar] [CrossRef]
- Smith, R.A.; Wu, V.H.; Zavala, C.G.; Raugi, D.N.; Ba, S.; Seydi, M.; Gottlieb, G.S.; University of Washington-Dakar, H.I.V.S.G. In Vitro Antiviral Activity of Cabotegravir against HIV-2. Antimicrobial agents and chemotherapy 2018, 62. [Google Scholar] [CrossRef]
- Gilead Sciences, I. Sunlenca® (lenacapavir) Receives FDA Approval as a First-in-Class, Twice-Yearly Treatment Option for People Living With Multi-Drug Resistant HIV. Available online: . (accessed on 22.12.2022).
- Link, J.O.; Rhee, M.S.; Tse, W.C.; Zheng, J.; Somoza, J.R.; Rowe, W.; Begley, R.; Chiu, A.; Mulato, A.; Hansen, D.; et al. Clinical targeting of HIV capsid protein with a long-acting small molecule. Nature 2020, 584, 614–618. [Google Scholar] [CrossRef]
- Mahdi, M.; Motyan, J.A.; Szojka, Z.I.; Golda, M.; Miczi, M.; Tozser, J. Analysis of the efficacy of HIV protease inhibitors against SARS-CoV-2’s main protease. Virology journal 2020, 17, 190. [Google Scholar] [CrossRef]
- Indu, P.; Rameshkumar, M.R.; Arunagirinathan, N.; Al-Dhabi, N.A.; Valan Arasu, M.; Ignacimuthu, S. Raltegravir, Indinavir, Tipranavir, Dolutegravir, and Etravirine against main protease and RNA-dependent RNA polymerase of SARS-CoV-2: A molecular docking and drug repurposing approach. Journal of infection and public health 2020, 13, 1856–1861. [Google Scholar] [CrossRef] [PubMed]
- Cavaco-Silva, J.; Abecasis, A.; Miranda, A.C.; Pocas, J.; Narciso, J.; Aguas, M.J.; Maltez, F.; Almeida, I.; Germano, I.; Diniz, A.; et al. HIV-2 integrase polymorphisms and longitudinal genotypic analysis of HIV-2 infected patients failing a raltegravir-containing regimen. PloS one 2014, 9, e92747. [Google Scholar] [CrossRef] [PubMed]
- Hutapea, H.M.L.; Maladan, Y.; Widodo. Relationship between HIV integrase polymorphisms and integrase inhibitor susceptibility: An in silico analysis. Heliyon 2018, 4, e00956. [Google Scholar] [CrossRef] [PubMed]
- Bercoff, D.P.; Triqueneaux, P.; Lambert, C.; Oumar, A.A.; Ternes, A.M.; Dao, S.; Goubau, P.; Schmit, J.C.; Ruelle, J. Polymorphisms of HIV-2 integrase and selection of resistance to raltegravir. Retrovirology 2010, 7, 98. [Google Scholar] [CrossRef]
- Miklossy, G.; Tozser, J.; Kadas, J.; Ishima, R.; Louis, J.M.; Bagossi, P. Novel macromolecular inhibitors of human immunodeficiency virus-1 protease. Protein engineering, design & selection : PEDS 2008, 21, 453–461. [Google Scholar] [CrossRef]
- Mahdi, M.; Matuz, K.; Toth, F.; Tozser, J. A modular system to evaluate the efficacy of protease inhibitors against HIV-2. PloS one 2014, 9, e113221. [Google Scholar] [CrossRef] [PubMed]
- Jochmans, D.; Leyssen, P.; Neyts, J. A novel method for high-throughput screening to quantify antiviral activity against viruses that induce limited CPE. Journal of virological methods 2012, 183, 176–179. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic acids research 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. Journal of computational chemistry 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Sondergaard, C.R.; Olsson, M.H.; Rostkowski, M.; Jensen, J.H. Improved Treatment of Ligands and Coupling Effects in Empirical Calculation and Rationalization of pKa Values. Journal of chemical theory and computation 2011, 7, 2284–2295. [Google Scholar] [CrossRef] [PubMed]
- Olsson, M.H.; Sondergaard, C.R.; Rostkowski, M.; Jensen, J.H. PROPKA3: Consistent Treatment of Internal and Surface Residues in Empirical pKa Predictions. Journal of chemical theory and computation 2011, 7, 525–537. [Google Scholar] [CrossRef] [PubMed]
- D.A. Case, R.M.B., D. D.A. Case, R.M.B., D.S. Cerutti, T.E. Cheatham, III, T.A. Darden, R.E. Duke, T.J. Giese, H. Gohlke, A.W. Goetz, N. Homeyer, S. Izadi, P. Janowski, J. Kaus, A. Kovalenko, T.S. Lee, S. LeGrand, P. Li, C. Lin, T. Luchko, R. Luo, B. Madej, D. Mermelstein, K.M. Merz, G. Monard, H. Nguyen, H.T. Nguyen, I. Omelyan, A. Onufriev, D.R. Roe, A. Roitberg, C. Sagui, C.L. Simmerling, W.M. Botello-Smith, J. Swails, R.C. Walker, J. Wang, R.M. Wolf, X. Wu, L. Xiao and P.A. Kollman AMBER 2016, University of California, San Francisco.
- Scott Le Grand, A.W.G. , Ross C. Walker. SPFP: Speed without compromise—A mixed precision model for GPU accelerated molecular dynamics simulations. Computer Physics Communications 2013, 184, 374–380. [Google Scholar] [CrossRef]
- Salomon-Ferrer, R.; Gotz, A.W.; Poole, D.; Le Grand, S.; Walker, R.C. Routine Microsecond Molecular Dynamics Simulations with AMBER on GPUs. 2. Explicit Solvent Particle Mesh Ewald. Journal of chemical theory and computation 2013, 9, 3878–3888. [Google Scholar] [CrossRef]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the Accuracy of Protein Side Chain and Backbone Parameters from ff99SB. Journal of chemical theory and computation 2015, 11, 3696–3713. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Murillo, R.; Robertson, J.C.; Zgarbova, M.; Sponer, J.; Otyepka, M.; Jurecka, P.; Cheatham, T.E. , 3rd. Assessing the Current State of Amber Force Field Modifications for DNA. Journal of chemical theory and computation 2016, 12, 4114–4127. [Google Scholar] [CrossRef]
- Li, P.; Merz, K.M., Jr. Taking into Account the Ion-induced Dipole Interaction in the Nonbonded Model of Ions. Journal of chemical theory and computation 2014, 10, 289–297. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of Simple Potential Functions for Simulating Liquid Water. J Chem Phys 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Machado, M.R.; Pantano, S. Split the Charge Difference in Two! A Rule of Thumb for Adding Proper Amounts of Ions in MD Simulations. Journal of chemical theory and computation 2020, 16, 1367–1372. [Google Scholar] [CrossRef]
- M. J. Frisch, G.W.T., H. M. J. Frisch, G.W.T., H. B. Schlegel, G. E. Scuseria, M. A. Robb, J. R. Cheeseman, G. Scalmani, V. Barone, G. A. Petersson, H. Nakatsuji, X. Li, M. Caricato, A. Marenich, J. Bloino, B. G. Janesko, R. Gomperts, B. Mennucci, H. P. Hratchian, J. V. Ortiz, A. F. Izmaylov, J. L. Sonnenberg, D. Williams-Young, F. Ding, F. Lipparini, F. Egidi, J. Goings, B. Peng, A. Petrone, T. Henderson, D. Ranasinghe, V. G. Zakrzewski, J. Gao, N. Rega, G. Zheng, W. Liang, M. Hada, M. Ehara, K. Toyota, R. Fukuda, J. Hasegawa, M. Ishida, T. Nakajima, Y. Honda, O. Kitao, H. Nakai, T. Vreven, K. Throssell, J. A. Montgomery, Jr., J. E. Peralta, F. Ogliaro, M. Bearpark, J. J. Heyd, E. Brothers, K. N. Kudin, V. N. Staroverov, T. Keith, R. Kobayashi, J. Normand, K. Raghavachari, A. Rendell, J. C. Burant, S. S. Iyengar, J. Tomasi, M. Cossi, J. M. Millam, M. Klene, C. Adamo, R. Cammi, J. W. Ochterski, R. L. Martin, K. Morokuma, O. Farkas, J. B. Foresman, and D. J. Fox Gaussian 09, Gaussian, Inc., Wallingford CT: 2016.
- Zhao, Y.; Truhlar, D.G. The M06 suite of density functionals for main group thermochemistry, thermochemical kinetics, noncovalent interactions, excited states, and transition elements: two new functionals and systematic testing of four M06-class functionals and 12 other functionals. Theoretical Chemistry Accounts 2008, 120, 215–241. [Google Scholar] [CrossRef]
- Bayly, C.I.; Cieplak, P.; Cornell, W.; Kollman, P.A. A well-behaved electrostatic potential based method using charge restraints for deriving atomic charges: the RESP model. The Journal of Physical Chemistry 1993, 97, 10269–10280. [Google Scholar] [CrossRef]
- Wang, J.; Wang, W.; Kollman, P.A.; Case, D.A. Automatic atom type and bond type perception in molecular mechanical calculations. Journal of molecular graphics & modelling 2006, 25, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Korb, O.; Stützle, T.; Exner, T.E. An ant colony optimization approach to flexible protein–ligand docking. Swarm Intelligence 2007, 1, 115–134. [Google Scholar] [CrossRef]
- Korb, O.; Stützle, T.; Exner, T.E. PLANTS: Application of Ant Colony Optimization to Structure-Based Drug Design. In Proceedings of the Ant Colony Optimization and Swarm Intelligence, Berlin, Heidelberg, 2006; 2006//; pp. 247–258. [Google Scholar]
- Cook, N.J.; Li, W.; Berta, D.; Badaoui, M.; Ballandras-Colas, A.; Nans, A.; Kotecha, A.; Rosta, E.; Engelman, A.N.; Cherepanov, P. Structural basis of second-generation HIV integrase inhibitor action and viral resistance. Science 2020, 367, 806–810. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Lambert, C.; Arendt, V.; Seguin-Devaux, C. Virological and immunological outcomes of elvitegravir-based regimen in a treatment-naive HIV-2-infected patient. Aids 2014, 28, 2329–2331. [Google Scholar] [CrossRef] [PubMed]
- Prather, C.; Lee, A.; Yen, C. Lenacapavir: A first-in-class capsid inhibitor for the treatment of highly treatment-resistant HIV. American journal of health-system pharmacy: AJHP: official journal of the American Society of Health-System Pharmacists. [CrossRef]
- Bester, S.M.; Wei, G.; Zhao, H.; Adu-Ampratwum, D.; Iqbal, N.; Courouble, V.V.; Francis, A.C.; Annamalai, A.S.; Singh, P.K.; Shkriabai, N.; et al. Structural and mechanistic bases for a potent HIV-1 capsid inhibitor. Science 2020, 370, 360–364. [Google Scholar] [CrossRef]
- Roquebert, B.; Damond, F.; Collin, G.; Matheron, S.; Peytavin, G.; Benard, A.; Campa, P.; Chene, G.; Brun-Vezinet, F.; Descamps, D.; et al. HIV-2 integrase gene polymorphism and phenotypic susceptibility of HIV-2 clinical isolates to the integrase inhibitors raltegravir and elvitegravir in vitro. The Journal of antimicrobial chemotherapy 2008, 62, 914–920. [Google Scholar] [CrossRef]
Figure 1.
Inhibition profiling in cell culture. Four-parameter dose-response curves are displayed. y axis indicates percentage of fluorescence compared to negative control (cell treated with DMSO without the inhibitor), x axis is the logarithmic transformation of the inhibitor’s concentration. Results represent triplicate measurements.
Figure 1.
Inhibition profiling in cell culture. Four-parameter dose-response curves are displayed. y axis indicates percentage of fluorescence compared to negative control (cell treated with DMSO without the inhibitor), x axis is the logarithmic transformation of the inhibitor’s concentration. Results represent triplicate measurements.
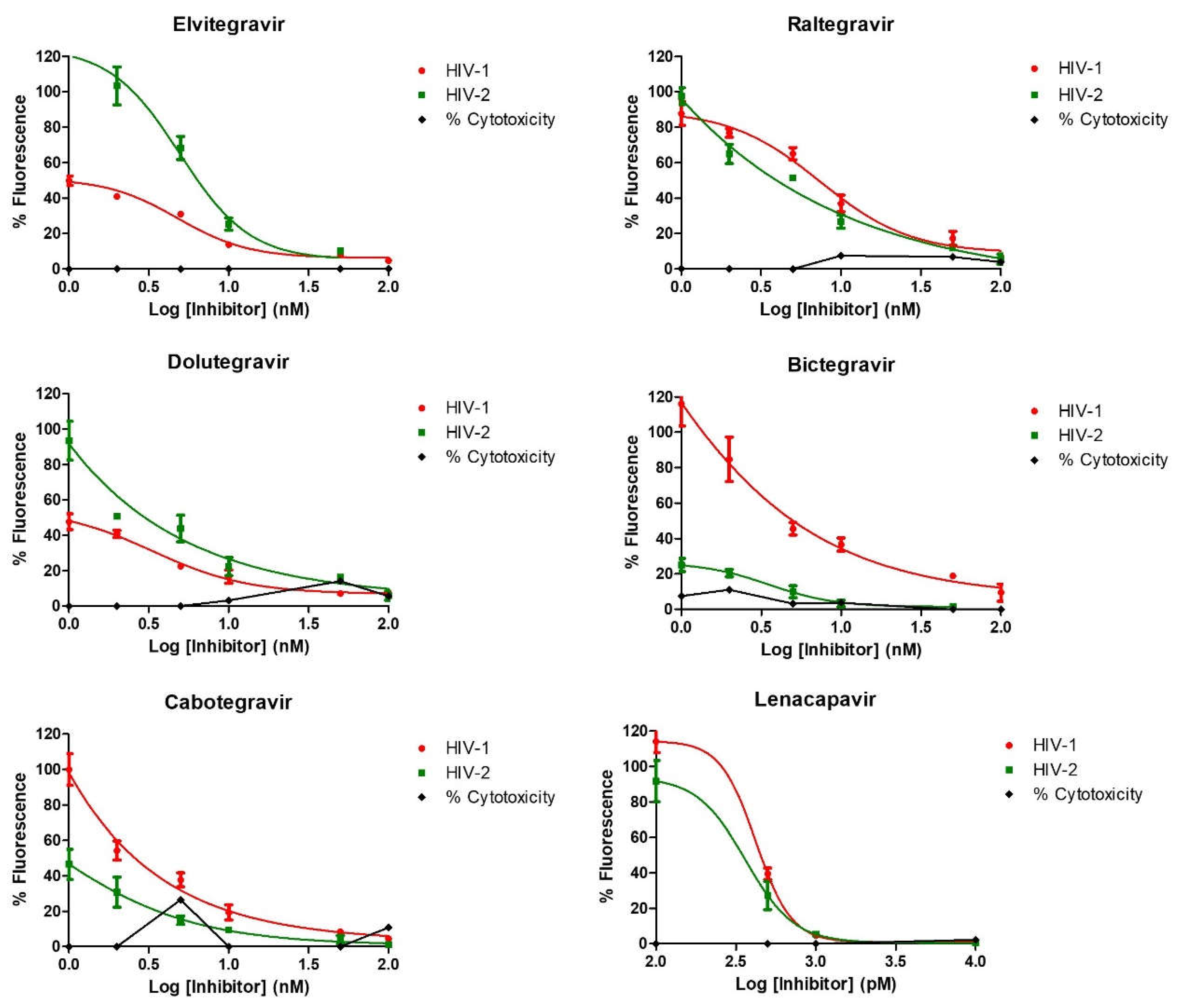
Figure 2.
In silico docking. Structure of (a) bictegravir and (b) cabotegravir docked by PLANTS in the active site of HIV-2 integrase. The inhibitor (teal), the DNA substrate (grey), and the Mg2+ ions along with the coordinated water molecules represented as spheres.
Figure 2.
In silico docking. Structure of (a) bictegravir and (b) cabotegravir docked by PLANTS in the active site of HIV-2 integrase. The inhibitor (teal), the DNA substrate (grey), and the Mg2+ ions along with the coordinated water molecules represented as spheres.
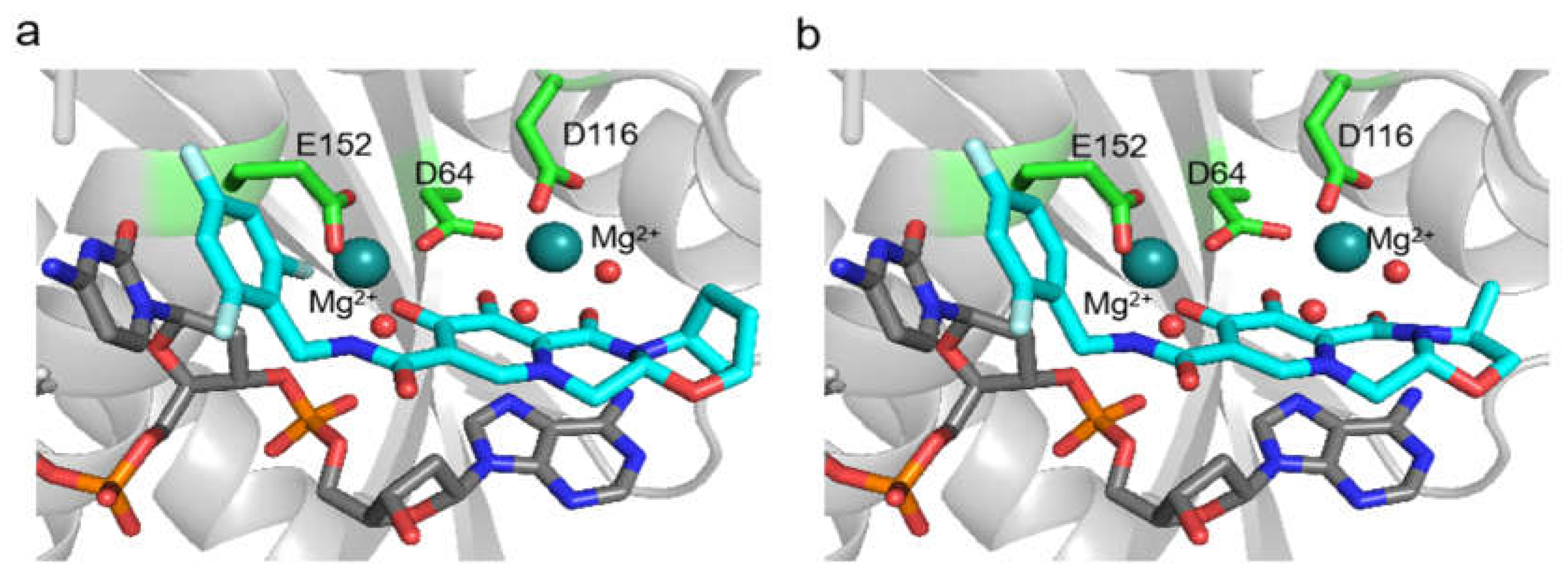
Figure 3.
Inhibition of HIV-1 and 2 pseudovirion production by lenacapavir. Pseudovirions were produced in the presence of 1, 10 and 100 nM of lenacapavir. y axis indicates percentage of transduction efficiency (percentage of fluorescence), x axis denotes the concentrations of lenacapavir used to treat the cells. Control cells were only treated with DMSO. Results are concluded from triplicate measurements.
Figure 3.
Inhibition of HIV-1 and 2 pseudovirion production by lenacapavir. Pseudovirions were produced in the presence of 1, 10 and 100 nM of lenacapavir. y axis indicates percentage of transduction efficiency (percentage of fluorescence), x axis denotes the concentrations of lenacapavir used to treat the cells. Control cells were only treated with DMSO. Results are concluded from triplicate measurements.
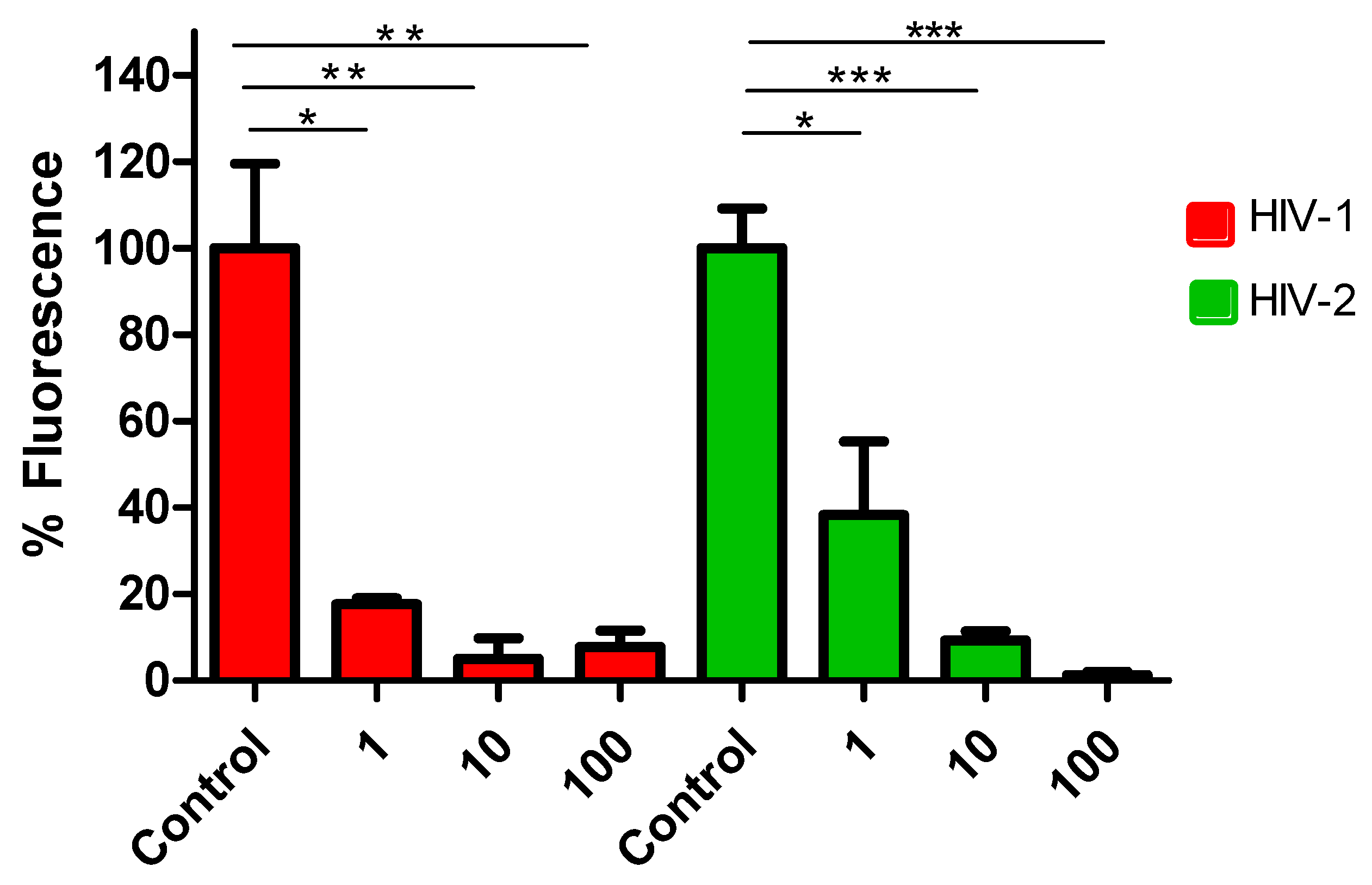
Figure 4.
Inhibition profiling of Raltegravir against SARs-CoV-2. Efficacy against the prototypical Wuhan-Hu-1 (GBH) is shown in green, while that agasint the omicron BA.2 is shown in red. Cytotoxycity is indicated in black. Inhibition profiling was carried out in Vero E6 cells and then confirmed in A549-Dual™ hACE2-TMPRSS2 cells. y axis indicates percentage of inhibition, x axis is the logarithmic transformation of the inhibitor’s concentration. Results represent at least triplicate measurements.
Figure 4.
Inhibition profiling of Raltegravir against SARs-CoV-2. Efficacy against the prototypical Wuhan-Hu-1 (GBH) is shown in green, while that agasint the omicron BA.2 is shown in red. Cytotoxycity is indicated in black. Inhibition profiling was carried out in Vero E6 cells and then confirmed in A549-Dual™ hACE2-TMPRSS2 cells. y axis indicates percentage of inhibition, x axis is the logarithmic transformation of the inhibitor’s concentration. Results represent at least triplicate measurements.
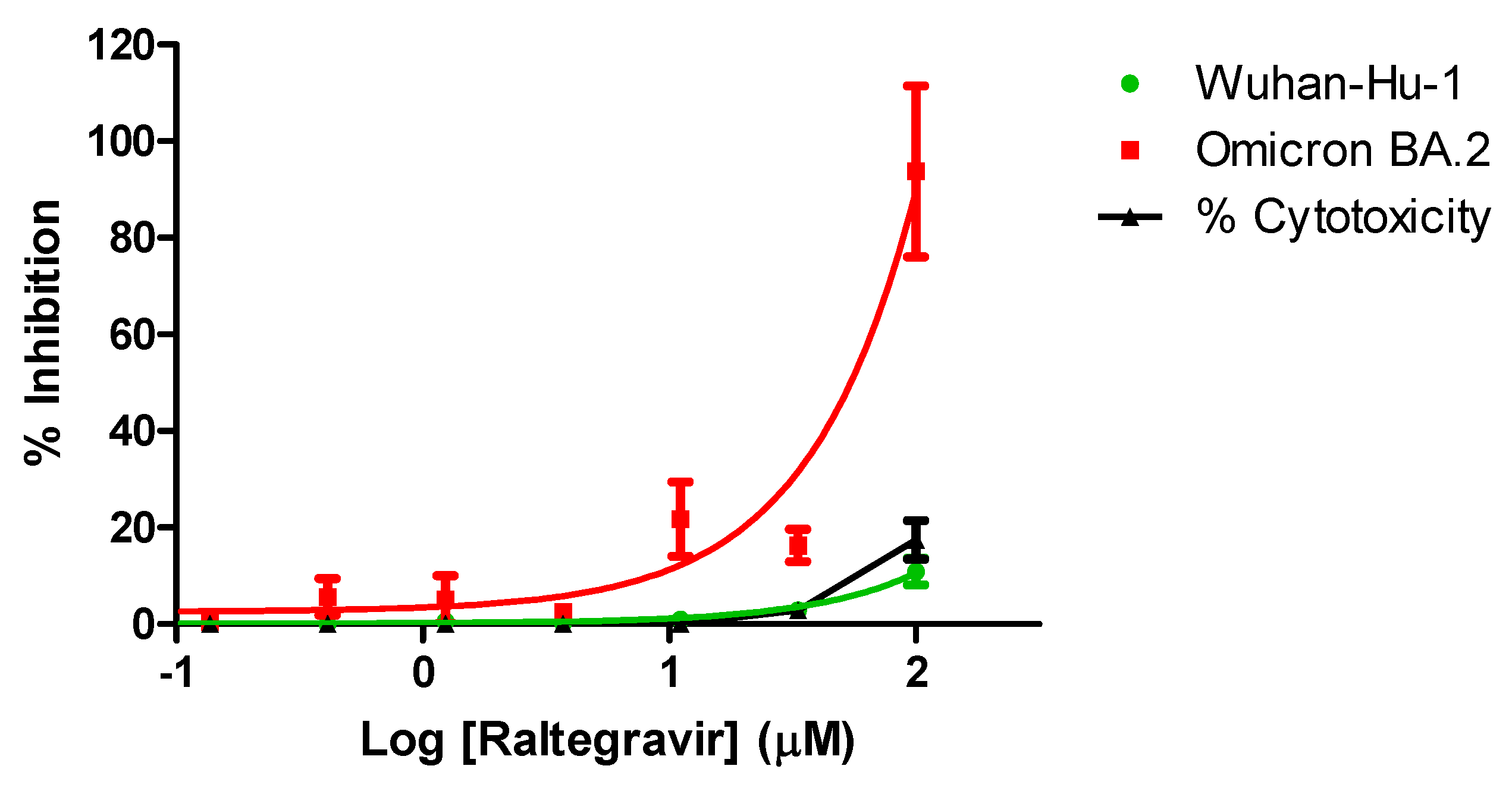

Table 1.
Results of inhibition profiling in cell culture. ± indicate standard error (SE) displayed as logIC50 value of the inhibitor. Results are concluded from triplicate measurements.
Table 1.
Results of inhibition profiling in cell culture. ± indicate standard error (SE) displayed as logIC50 value of the inhibitor. Results are concluded from triplicate measurements.
| IC50 (nM) | ||
|---|---|---|
| INSTI | HIV-1 | HIV-2 |
| Elvitegravir | 2.5 (±0.1) | 2.6 (±0.4) |
| Raltegravir | 6.9 (±0.1) | 2.1 (±0.1) |
| Dolutegravir | 2.2 (±0.1) | 1.1 (±0.3) |
| Bictegravir | 1.2 (±0.2) | 1.8 (±0.3) |
| Cabotegravir | 0.4 (±0.3) | 0.8 (±0.4) |
| Capsid Inhibitor | IC50 (pM) | |
| Lenacapavir | 89 (±0.2) | 50.6 (±0.6) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



