Submitted:
21 April 2024
Posted:
22 April 2024
You are already at the latest version
Abstract
Ixodes ricinus ticks are widespread in Europe and important vectors for multiple pathogens. The purpose of the study was to collect ticks in a mountainous area in Greece and screen those of I. ricinus species for several pathogens. I. ricinus was the predominant tick species identified (398/440, 90.45%). Following pooling (1-17 ticks per pool), homogenization and extraction of genetic material of the ticks, a real time PCR was applied for the detection of a panel of tick-borne pathogens, while additional conventional PCRs combined with Sanger sequencing were applied for detection of viruses and for typing of Rickettsia and Borrelia species. At least one pathogen was detected in 60 of the 80 (75%) pools. Rickettsia spp. predominated as it was detected in 63.75% of the tick pools (51/80), followed by Borrelia spp. (35 pools, 45%); A. phagocytophilum was detected in 2 (2.5%) pools, phleboviruses in 3 (3.75%) and nairoviruses in 7 (8.75%) pools. All the Rickettsia sequences taken belonged to R. monacensis, while among Borrelia spp., B. garinii and B. valaisiana were detected. Regarding viruses, Norway phlebovirus 1, Sulina and Grotenhout nairoviruses were detected. Although a small number of ticks from only one area in Greece was tested, a variety of pathogens and recently identified viruses were detected, prompting for further studies in ticks and surveillance studies in humans.
Keywords:
Ixodes ricinus
; tick-borne pathogens
; Rickettsiae
; Greece
1. Introduction
Ixodes ricinus is a widespread tick species in Europe and known vector for a wide range of pathogens, including Borrelia, Anaplasma, Rickettsia, and Babesia species, as well as tick-borne encephalitis virus (TBEV). Ticks of this species undergo a three-host life cycle, and primarily inhabit wooded and forested regions where they feed on various mammalian and avian hosts [1]. The distribution of I. ricinus is shaped by multiple factors, like environmental and vegetation patterns, and availability of suitable hosts [2]. The likelihood of pathogen transmission to humans and animals through tick bites is associated with high density and prevalence of infected ticks alongside with hosts' activities [3].
Several studies have been conducted in Greece aiming to identify the tick species and/or to detect pathogens in ticks removed from humans or livestock (sheep, goats and cattle). Results from a study in 519 ticks removed from humans showed that 81.5% were Rhipicephalus sanguineus, while I. ricinus accounted for only 1% [4]. The tick prevalence and distribution vary among studies on ticks collected from domestic animals depending on the location, animal host and seasonality. As an example, a study on 11,620 ticks collected during 1983-1986 from domestic animals in the Macedonia Region of Greece, identified 18 tick species and subspecies, with Rhipicephalus bursa being the most common tick (36.3%); I. ricinus accounted for 6.8% of the ticks, and was found mainly during autumn and winter in the biotopes of the attenuated mesomediterranean and submediterranean bioclimates, and especially in deciduous forest areas [5]. Similarly, another study on 2,108 ixodid ticks collected from small ruminants in Greece showed that Rhipicephalus sanguineus s.l. and R. bursa were the most prevalent tick species (64.8% and 25.9%, respectively), whereas the least frequently collected species were I. ricinus, H. punctata and H. impeltatum (0.04% each) [6]. However, different results were taken from a targeted study in northern Greece where the collection was performed only during April–July and September–December, and only on livestock (mainly goats) grazing in hilly areas neighboring with forests of deciduous and evergreen vegetation, where the average humidity was approximately 80%; in that study ticks were collected from 3,249 animals, and I. ricinus was the predominant tick species (44.57%) [7].
Application of 16S rRNA next generation sequencing in ticks belonging to four genera (Ixodes, Rhipicephalus, Dermacentor, Haemaphyssalis) showed that the predominant phylum in I. ricinus ticks was the Proteobacteria; bacteria of the Rickettsiaceae (Rickettsia and Anaplasma species) and “Candidatus Midichloriaceae” families were detected [8]. Another small-scaled study showed that 66.6% and 33.3% pools of I. ricinus ticks collected from goats were positive for Rickettsia monacensis and Anaplasma platys, respectively [9].
Regarding viruses, Greek goat encephalitis virus (GGEV) from the tick-borne encephalitis group, was isolated in 1969 from the brain of a newborn goat with neurological symptoms [10]. To identify the tick vector of GGEV, Ixodidae ticks collected from goats and sheep in rural areas of northern Greece were tested, and GGEV was detected in two pools of I. ricinus ticks [7]. The virus was then isolated from one of the positive pools [11]. Few human TBE cases have been reported in Greece during the last years (article in preparation), suggesting that the virus is present in the country and needs further attention and surveillance studies.
To gain a better insight into the pathogens carried by I. ricinus ticks in Greece, a mountainous area with an ecosystem favorable for this tick species was selected for tick collection and screening for various pathogens.
2. Materials and Methods
2.1. Study Area
Mount Vermio (40°31'32.9"N 22°00'37.4"E) is located in the central part of northern Greece, having a north-south orientation. The study area included 83 locations in the municipalities of Veria, Naoussa, and Edessa (Figure 1); the collective population of the area is 199,116 according to the most recent census data [12]. The slopes facing to the East are influenced by pluvial aerial masses coming from the Aegean Sea; as a result, the ecosystem is more productive than the West-facing sites. The mountain is bounded to the East by the Central Macedonian plain, characterized by intensive cultivation of fruit-bearing trees. Its southern and northern boundaries are delineated by Aliakmonas and Agras rivers, respectively. During the last decades the gradual abandonment of transhumance in the highlands of the mountain resulted in a relative increase of forested areas. In addition, the area is undergoing considerable land use change, marked by the implementation of renewable energy projects, such as wind farms and solar parks. The climate of the area is temperate Mediterranean, characterized by rainy winters and warm summers. The total annual rainfall in the area is 1500 mm, whereas the mean annual precipitation is approximately 699.83 mm, and the mean annual air temperature is 15°C. The minimum rainfall occurs during July-August; however, the atmosphere remains humid due to its proximity to the archipelagos.
2.2. Sampling
During April 2021 to March 2022, drag sampling was conducted by using a 1m2 flannel cloth. An attempt was made to revisit the sites surveyed in spring and summer 2021, with subsequent sampling extending in autumn, winter, and spring of 2021 and 2022, resulting in a cumulative total of 135 sampling occurrences. Sampling was conducted in a diverse range of ecological niches, including deciduous forests, characterized mainly by Carpinus orientalis, Ostrya carpinifolia, Quercus frainetto, Q. pubescens, Q. trojana, Castanea sativa, Tilia tomentosa, Fagus sylvatica, occurring individually or in various combinations. Furthermore, the investigations were extended to coniferous evergreen forested areas where Pinus nigra predominated, and to typical maquis vegetation, where the dominant shrub species was Quercus coccifera, followed by Buxus sempervirens. Survey locations encompassed a spectrum of settings, including forested sites adjacent to agricultural and grazing zones, local recreational areas, hiking trails, hunting grounds, and areas designated under the NATURA 2000 conservation program within the mountainous terrain. The altitudinal range for the samplings spanned from 65 to 1900 meters above sea level.
2.3. Tick Identification and Transportation
The identification of ticks was performed under a stereomicroscope on an ice pack using morphological identification keys [13]. Then, they were transferred to the laboratory in dry ice and kept in -80oC until processing.
2.4. RNA Extraction and Pathogens Detection
Ticks were grouped into pools (1-17 ticks per pool), based on the collection date and site. Following washing with distilled water, they were homogenized by vortexing for 5 minutes in phosphate buffered saline using glass beads (diameter 150-212 μm). Total RNA was extracted from 200 μl of the supernatants using the QIAamp cador Pathogen Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. The CerTest VIASURE Tick Borne Diseases Real Time PCR Detection kit (Abacus dx, Australia) was used for the detection of TBEV (region of the 3’UTR), Rickettsia spp. (23S rRNA gene), Babesia microti (CCT-eta gene), Babesia divergens (hsp70 gene), Ehrlichia chafeensis and Ehrlichia muris (GroEl gene), Borrelia burgdorferi sensu lato (s.l.), Borrelia miyamotoi and/or Borrelia hermsii, (23S rRNA gene), Anaplasma phagocytophilum (msp2 gene) and Coxiella burnetii (IS1111 gene).
To identify Borrelia spp. and Rickettsia spp. at the genospecies level, the positive pools with Ct value <28 were further tested individually by in house PCR assays which amplify the 5S-23S rRNA intergenic spacer of B. burgdoferi s.l. [14], and the citrate synthase gene of Rickettsia spp. [15]. Additional in-house PCR assays were applied for the detection of partial segment of the NS5 gene of flaviviruses [16], and partial L segment of phleboviruses (family Phenuiviridae) [17] and nairoviruses (family Nairoviridae) [18].
2.5. Sanger Sequencing and Analysis
The PCR products of the in-house PCRs were Sanger sequenced in a 3130 ABI Genetic Analyzer (Applied Biosystems, Foster City, CA). Nucleotide sequences were compared with respective ones available in GenBank using the National Center for Biotechnology Information (NCBI) Basic Local Alignment Sequence Tool (BLAST) search engine to identify the best fit.
3. Results
3.1. Ticks
I. ricinus accounted for 90.45 (398/440) of the total number of collected ticks, most of them (87.2%) adults (Table 1). They were found in 47.4% (64/135) of the sampling locations, at least once in 51.8% (43/83) of the sampled locations, mainly in deciduous forests (all developmental stages), but also in typical maquis vegetation (only adults). No ticks were present in Pinus nigra forested areas. Adult I. ricinus ticks were the predominant developmental stage irrespective of the period of the year. The altitudinal activity pattern of I. ricinus varied from 295m to 1580 m; it was most commonly present above 600m.
3.2. Pathogen Detection
At least one pathogen was detected in 60 of the 80 (75%) pools. Rickettsia spp. predominated as it was detected in 63.75% of the tick pools (51/80). Borrelia spp. was detected in 36 pools (45%), A. phagocytophilum in 2 (2.5%), phleboviruses in 3 (3.75%) and nairoviruses in 7 (8.75%) pools (Table 2). Further analysis through individually tick testing from the positive pools revealed that all Rickettsia sequences presenting 100% identity with R. monacensis, IrR/Munich strain (GenBank Accession number LN794217). Among six Borrelia-positive individual ticks, four were carrying B. garinii sequences and two B. valaisiana sequences, both belonging to the B. burgdorferi sl. complex. Regarding B. garinii, one sample carried sequences 100% identical with the isolate B12F6Swallowcliffe17 (Acc. No. OL848409), two samples carried sequences 100% identical with the isolate B13D2Donheads15 (Acc. No. OL848423), while the sequences of the fourth tick presented 99.2% identity with the isolate B13D2Donheads15. The two B. valaisiana sequences were identical to those of the isolate B13E4GCommon (Acc. No. OL848426) [19].
Regarding the three phlebovirus-positive tick pools, all were carrying sequences belonging to the tick-specific Ixovirus genus in the Phenuiviridae family, differing from the Norway phlebovirus 1 (or Fairhair virus) isolate NOR/A2/Bronnoya/2014 (Acc. No. NC_055434) by 1.9-2.6% and 0.7-2.1% at nucleotide and amino acid level, respectively [20].
Among the seven nairovirus positive tick pools, one was carrying sequences presenting 90.3% identity to Sulina virus (unclassified Orthonairovirus in the Nairoviridae family) (Acc. No. NC_078999) detected in 2016 in I. ricinus ticks collected at the Danube Delta in Romania [21], while the rest six tick pools were carrying sequences presenting 94.74-98.03% (mean 96.6%) identity to Grotenhout virus isolate Gierle-1 (genus Nairovirus in the Nairoviridae family) detected in 2009 in I. ricinus ticks collected in Belgium (Acc. No. NC_078265) [22]. At amino acid level all nairovirus sequences were 100% identical.
Co-infections were detected in 30 (37.5%) pools presenting a variety of combinations, with most common (26.25%) that of Rickettsia and Borrelia species (Table 3).
4. Discussion
In contrast to most of the previous studies conducted on ticks collected from livestock in Greece, I. ricinus was the predominant tick species (90.45%) detected in the present study, suggesting that areas with deciduous forests at altitudes above 600m is a favorable ecosystem for this tick species in the country, resembling that in temperate Europe. The only previous study in Greece in which I. ricinus predominated (44.57%), was the one conducted during specific months of the year in humid hilly areas neighboring with forests [7]. The fact that ticks were not found in Pinus nigra forested areas is in accordance with a study in Belgium where the abundance of ticks of all stages was higher in oak stands compared to pine stands, and increased with increasing shrub cover, probably related to the main hosts habitat preference [23].
Rickettsia spp. displayed the highest prevalence, and all obtained sequences were identical to R. monacensis IrR/Munich isolate (Acc.No. LN794217) which was identified in May 1998 in adult I. ricinus ticks in Munich, Germany [24]. R. monacensis is a spotted fever group rickettsia and is rarely associated with disease in humans [25,26,27]. The most common Rickettsia species of this group in Greece (as in all Mediterranean countries) is R. conorii conorii which is transmitted mainly by R. sanguineus ticks [28]
I. ricinus also serves as vector for species of B. burgdorferi s.l. group, which are spirochete bacteria responsible for Lyme borreliosis, the most prevalent tick-borne disease in the northern hemisphere [29]. In the present study B. burgdorferi (s.l.) was detected in almost half (45%) of the tick pools. Despite the significant prevalence, which is similar to that detected in northern and central European countries, only few Lyme borreliosis cases have been laboratory diagnosed in Greece (unpublished data). A probable explanation might be that few areas in Greece fulfill the criteria for suitable habitats for I. ricinus, combined with presence of key tick hosts, while underdiagnosis cannot be excluded. In Europe, Lyme borreliosis is mainly caused by three genospecies: B. garinii, associated with neuroborreliosis, B. afzelii related to atrophic chronic acrodermatitis, and B. burgdorferi sensu stricto (s.s.) related to arthritis [30]. B. garinni was detected in the present study, followed by B. valaisiana. In a recent study in Spain, B. garinni and B. valaisiana predominated among the detected Borrelia species [31].
A. phagocytophilum, an obligate intracellular bacterium causing human granulocytic anaplasmosis in humans was detected in only two tick pools; both pools were co-infected with Rickettsia spp., while in one of them Borrelia species was also present. Similar coinfections are often described in Europe where these pathogens are prevalent [32].
Phleboviruses are transmitted mainly by phlebotomine sandflies. However, various phleboviruses have been detected in ticks in several countries, including Greece [33,34]. For this reason, they were included in the testing, and Norway phlebovirus 1 was detected in 3 pools. Similar viruses have been detected in Belgium and France [20]. In addition, two different nairoviruses were detected in the ticks: Sulina virus, which was initially detected in Danube Delta in Romania and recently in France, and Grotenhout virus, which is also present (with various names) in other European countries [20,21,22,35,36]. Further studies are needed to find out whether the detected phleboviruses and nairoviruses are important for the public and animal health or they play any role in the life cycle of the ticks.
5. Conclusions
The present study shows that there is an established I. ricinus population in specific areas in Greece, such as Vermion mountain, which is infected with various pathogens (mostly Rickettsia spp.) suggesting that country-wide studies are needed, firstly to identify the temporal and spatial distribution and abundance of I. ricinus ticks, and secondly to estimate the rate of ticks infected with various pathogens. Although TBEV was not detected in the present study, it should be included in the screening list of pathogens as there are reports of human cases in Greece. It is expected that the climate change together with various environmental and anthropogenic factors will result in alterations of the distribution of vectors, including ticks; therefore, surveillance data have to be often updated to ensure the efficient design of control and prevention strategies to mitigate the risk associated with tick-borne pathogens.
Author Contributions
Conceptualization, A.S. and A.P.; methodology, A.P. and A.S.; validation, A.P.; formal analysis, A.P.; investigation, K.T., A.S. and S.P.; resources, A.S. and A.P.; data curation, A.P., A.S. and K.T.; writing—original draft preparation, K.T. and A.S.; writing—review and editing, A.P.; visualization, A.P..; supervision, A.P.; project administration, A.S. and A.P.; funding acquisition, A.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by European Union’s Horizon 2020 research and innovation programme under Grant number 874735 (VEO).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gray, J.S.; Kahl, O.; Lane, R.S.; Levin, M.L.; Tsao, J.I. Diapause in ticks of the medically important Ixodes ricinus species complex. Ticks Tick Borne Dis. 2016, 7, 992–1003. [Google Scholar] [CrossRef]
- Estrada-Pena, A.; Ayllon, N.; de la Fuente, J. Impact of climate trends on tick-borne pathogen transmission. Front Physiol. 2012, 3, 64. [Google Scholar] [CrossRef]
- Heyman, P.; Cochez, C.; Hofhuis, A.; van der Giessen, J.; Sprong, H.; Porter, S.R.; et al. A clear and present danger: tick-borne diseases in Europe. Expert Rev Anti Infect Ther. 2010, 8, 33–50. [Google Scholar] [CrossRef]
- Papa, A.; Chaligiannis, I.; Xanthopoulou, K.; Papaioakim, M.; Papanastasiou, S.; Sotiraki, S. Ticks parasitizing humans in Greece. Vector Borne Zoonotic Dis. 2011, 11, 539–542. [Google Scholar] [CrossRef]
- Papadopoulos, B.; Morel, P.C.; Aeschlimann, A. Ticks of domestic animals in the Macedonia region of Greece. Vet Parasitol. 1996, 63, 25–40. [Google Scholar] [CrossRef]
- Chaligiannis, I.; Musella, V.; Rinaldi, L.; Cringoli, G.; de la Fuente, J.; Papa, A.; et al. Species diversity and spatial distribution of ixodid ticks on small ruminants in Greece. Parasitol Res. 2016, 115, 4673–4680. [Google Scholar] [CrossRef]
- Pavlidou, V.; Gerou, S.; Kahrimanidou, M.; Papa, A. Ticks infesting domestic animals in northern Greece. Exp Appl Acarol. 2008; 45, 195–198. [Google Scholar] [CrossRef]
- Papa, A.; Tsioka, K.; Daskou, M.A.; Minti, F.; Papadopoulou, E.; Melidou, A.; et al. Application of 16S rRNA next generation sequencing in ticks in Greece. Heliyon. 2020, 6, e04542. [Google Scholar] [CrossRef]
- Papa, A.; Tsioka, K.; Kontana, A.; Papadopoulos, C.; Giadinis, N. Bacterial pathogens and endosymbionts in ticks. Ticks Tick Borne Dis. 2017, 8, 31–35. [Google Scholar] [CrossRef]
- Papadopoulos, O.; Paschaleri-Papadopoulou, E.; Deligaris, N.; Doukas, G. Isolation of tick borne encephalitis virus from a flock of goats with abortions and fatal disease (preliminary report). Vet News Greece 1971, 3, 112–114. [Google Scholar]
- Papa, A.; Pavlidou, V.; Antoniadis, A. Greek goat encephalitis virus strain isolated from Ixodes ricinus, Greece. Emerg Infect Dis. 2008, 14, 330–332. [Google Scholar] [CrossRef]
- ELSTAT. 2021:https://elstat-outsourcers.statistics.gr/Census2022_GR.pdf.
- Estrada-Peña, A.; Mihalca, A.D.; Petney, T.N. Ticks of Europe and North Africa; Springer Cham: N.Y., USA.
- Mysterud, A.; Stigum, V.M.; Jaarsma, R.I.; Sprong, H. Genospecies of Borrelia burgdorferi sensu lato detected in 16 mammal species and questing ticks from northern Europe. Sci Rep. 2019, 9, 5088. [Google Scholar] [CrossRef]
- Regnery, R.L.; Spruill, C.L.; Plikaytis, B.D. Genotypic identification of rickettsiae and estimation of intraspecies sequence divergence for portions of two rickettsial genes. J Bacteriol. 1991, 173, 1576–1589. [Google Scholar] [CrossRef]
- Scaramozzino, N.; Crance, J.M.; Jouan, A.; DeBriel, D.A.; Stoll, F.; Garin, D. Comparison of flavivirus universal primer pairs and development of a rapid, highly sensitive heminested reverse transcription-PCR assay for detection of flaviviruses targeted to a conserved region of the NS5 gene sequences. J Clin Microbiol. 2001, 39, 1922–1927. [Google Scholar] [CrossRef]
- Klimentov, A.S.; Butenko, A.M.; Khutoretskaya, N.V.; Shustova, E.Y.; Larichev, V.F.; Isaeva, O.V. Development of pan-phlebovirus RT-PCR assay. J Virol Methods 2016, 232, 29–32. [Google Scholar] [CrossRef]
- Honig, J.E.; Osborne, J.C.; Nichol, S.T. The high genetic variation of viruses of the genus Nairovirus reflects the diversity of their predominant tick hosts. Virology. 2004, 318, 10–16. [Google Scholar] [CrossRef]
- Hansford, K.M.; McGinley, L.; Wheeler, B.W.; Tschirren, B.; Medlock, J.M. Ixodes ricinus density, Borrelia prevalence and the density of infected nymphs along an urban-rural gradient in southern England. Zoonoses Public Health. 2023, 70, 304–314. [Google Scholar] [CrossRef]
- Pettersson, J.H.; Shi, M.; Bohlin, J.; Eldholm, V.; Brynildsrud, O.B.; Paulsen, K.M.; et al. Characterizing the virome of Ixodes ricinus ticks from northern Europe. Sci Rep. 2017, 7, 10870. [Google Scholar] [CrossRef]
- Tomazatos, A.; von Possel, R.; Pekarek, N.; Holm, T.; Rieger, T.; Baum, H.; et al. Discovery and genetic characterization of a novel orthonairovirus in Ixodes ricinus ticks from Danube Delta. Infect Genet Evol. 2021, 88, 104704. [Google Scholar] [CrossRef]
- Vanmechelen, B.; Laenen, L.; Vergote, V.; Maes, P. Grotenhout Virus, a Novel Nairovirus Found in Ixodes ricinus in Belgium. Genome Announc. 2017, 5, 21. [Google Scholar] [CrossRef]
- Tack, W.; Madder, M.; Baeten, L.; De Frenne, P.; Verheyen, K. The abundance of Ixodes ricinus ticks depends on tree species composition and shrub cover. Parasitology. 2012, 139, 1273–1281. [Google Scholar] [CrossRef]
- Simser, J.A.; Palmer, A.T.; Fingerle, V.; Wilske, B.; Kurtti, T.J.; Munderloh, U.G. Rickettsia monacensis sp. nov., a spotted fever group Rickettsia, from ticks (Ixodes ricinus) collected in a European city park. Appl Environ Microbiol. 2002, 68, 4559–4566. [Google Scholar] [CrossRef]
- de Sousa, R.; Dos Santos, M.L.; Cruz, C.; Almeida, V.; Garrote, A.R.; Ramirez, F.; et al. Rare Case of Rickettsiosis Caused by Rickettsia monacensis, Portugal, 2021. Emerg Infect Dis. 2022, 28, 1068–1071. [Google Scholar] [CrossRef]
- Jado, I.; Oteo, J.A.; Aldamiz, M.; Gil, H.; Escudero, R.; Ibarra, V.; et al. Rickettsia monacensis and human disease, Spain. Emerg Infect Dis. 2007, 13, 1405–1407. [Google Scholar] [CrossRef]
- Madeddu, G.; Mancini, F.; Caddeo, A.; Ciervo, A.; Babudieri, S.; Maida, I.; et al. Rickettsia monacensis as cause of Mediterranean spotted fever-like illness, Italy. Emerg Infect Dis. 2012, 18, 702–704. [Google Scholar] [CrossRef]
- Papa, A.; Dalla, V.; Petala, A.; Maltezou, H.C.; Maltezos, E. Fatal Mediterranean spotted fever in Greece. Clin Microbiol Infect. 2010, 16, 589–592. [Google Scholar] [CrossRef]
- Stanek, G.; Strle, F. Lyme borreliosis-from tick bite to diagnosis and treatment. FEMS Microbiol Rev. 2018, 42, 233–258. [Google Scholar] [CrossRef]
- Stanek, G.; Wormser, G.P.; Gray, J.; Strle, F. Lyme borreliosis. Lancet. 2012, 379, 461–473. [Google Scholar] [CrossRef]
- Diaz, P.; Arnal, J.L.; Remesar, S.; Perez-Creo, A.; Venzal, J.M.; Vazquez-Lopez, M.E.; et al. Molecular identification of Borrelia spirochetes in questing Ixodes ricinus from northwestern Spain. Parasit Vectors. 2017, 10, 615. [Google Scholar] [CrossRef]
- Glass, A.; Springer, A.; Raulf, M.K.; Fingerle, V.; Strube, C. 15-year Borrelia prevalence and species distribution monitoring in Ixodes ricinus/inopinatus populations in the city of Hanover, Germany. Ticks Tick Borne Dis. 2023, 14, 102074. [Google Scholar] [CrossRef]
- Papa, A.; Kontana, A.; Tsioka, K.; Chaligiannis, I.; Sotiraki, S. Novel phleboviruses detected in ticks, Greece. Ticks Tick Borne Dis. 2016, 7, 690–693. [Google Scholar] [CrossRef]
- Papa, A.; Kontana, A.; Tsioka, K.; Saratsis, A.; Sotiraki, S. Novel phlebovirus detected in Haemaphysalis parva ticks in a Greek island. Ticks Tick Borne Dis. 2017, 8, 157–160. [Google Scholar] [CrossRef]
- Sameroff, S.; Tokarz, R.; Vucelja, M.; Jain, K.; Oleynik, A.; Boljfetic, M.; et al. Virome of Ixodes ricinus, Dermacentor reticulatus, and Haemaphysalis concinna Ticks from Croatia. Viruses 2022, 14, 5. [Google Scholar] [CrossRef]
- Moutailler, S.; Popovici, I.; Devillers, E.; Vayssier-Taussat, M.; Eloit, M. Diversity of viruses in Ixodes ricinus, and characterization of a neurotropic strain of Eyach virus. New Microbes New Infect. 2016, 11, 71–81. [Google Scholar] [CrossRef]
Figure 1.
Locations in Mount Vermion, northern Greece, where Ixodes ricinus ticks were collected. The location of the area in Greece is seen in the inset.
Figure 1.
Locations in Mount Vermion, northern Greece, where Ixodes ricinus ticks were collected. The location of the area in Greece is seen in the inset.
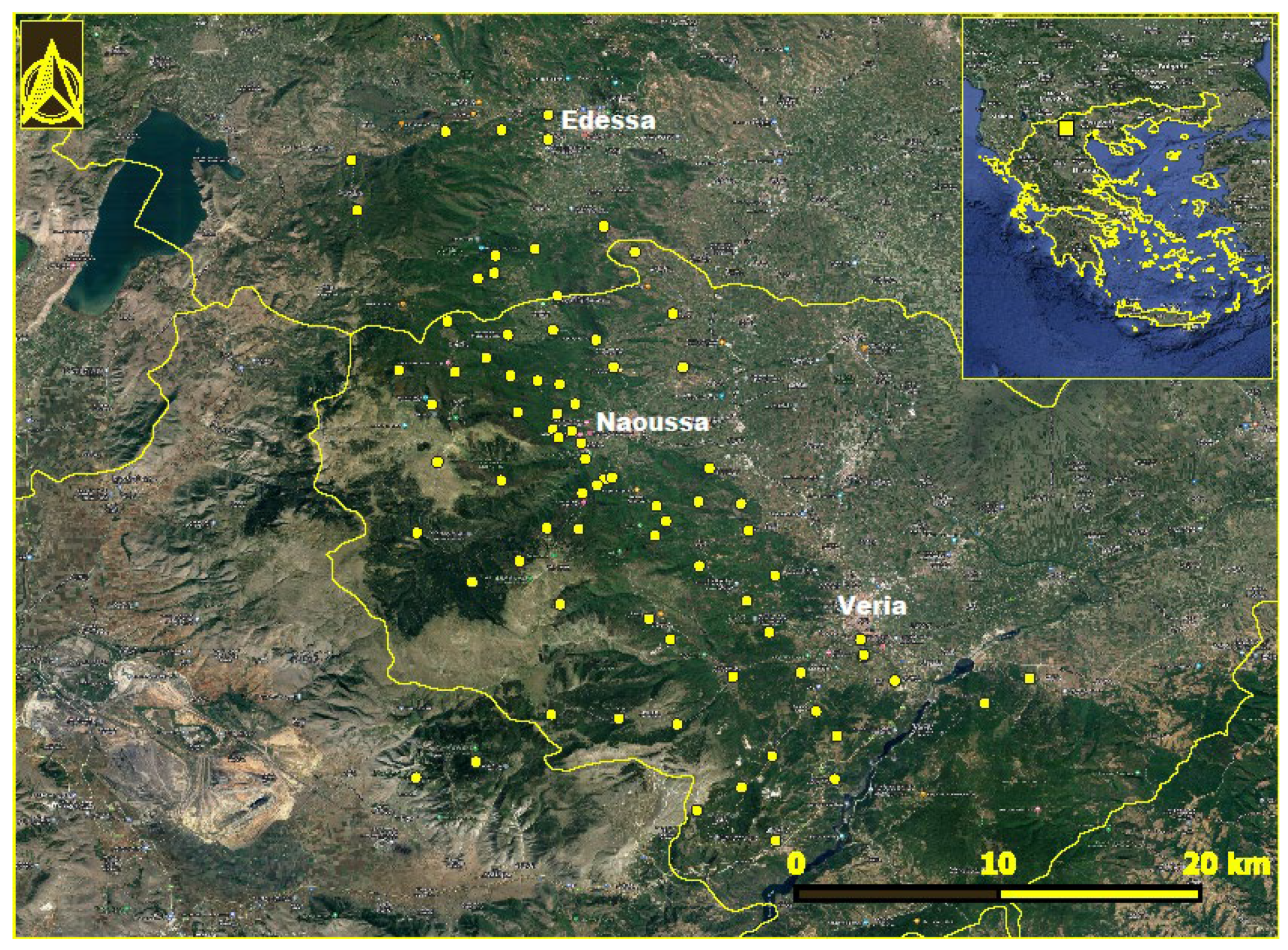

Table 1.
Tick species collected in Mount Vermion during April 2021-March 2022.
| Tick species | Number (%) |
|---|---|
| I. ricinus (female) | 157 (35.7) |
| I. ricinus (male) | 190 (43.2) |
| I. ricinus (nymph) | 49 (11.1) |
| I. ricinus (larva) | 2 (0.5) |
| Dermacentor marginatus (female) | 5 (1.1) |
| Dermacentor marginatus (male) | 4 (0.9) |
| Haemahysalis punctata (female) | 2 (0.5) |
| Haemahysalis punctata (male) | 1 (0.2) |
| Haemaphysalis inermis (female) | 9 (2.0) |
| Haemaphysalis inermis (male) | 2 (0.5) |
| Haemaphysalis inermis (nymph) | 2 (0.5) |
| Haemaphysalis parva (female) | 2 (0.5) |
| Haemaphysalis parva (male) | 2 (0.5) |
| I. frontalis (nymph) | 6 (1.4) |
| I. gibbosus (female) | 2 (0.5) |
| Rhipicephalus sanguineus s.l (male) | 1 (0.2) |
| Rhipicephalus turanicus s.l (female) | 2 (0.5) |
| Rhipicephalus turanicus s.l (male) | 2 (0.5) |
| Total | 440 (100) |
Table 2.
Bacteria and viruses detected in Ixodes ricinus ticks collected in Mount Vermion.
| Microorganism | No of pools (%) | |
|---|---|---|
| Bacteria | Rickettsia spp. | 51 (63.75) |
| Borrelia spp. | 36 (45.0) | |
| A. phagocytophilum | 2 (2.5) | |
| Viruses | Phleboviruses | 3 (3.75) |
| Orthonairoviruses | 7 (8.75) | |
Table 3.
Bacteria and viruses detected in Ixodes ricinus ticks collected in Vermion Mountain, individually and as co-infections. .
Table 3.
Bacteria and viruses detected in Ixodes ricinus ticks collected in Vermion Mountain, individually and as co-infections. .
| Microorganism | No of pools (%) |
|---|---|
| Rickettsia spp. | 22 (27.5) |
| Borrelia spp. | 8 (10.0) |
| Rickettsia spp. + Borrelia spp. | 21 (26.25) |
| Rickettsia spp + A. phagocytophilum | 1 (1.25) |
| Borrelia spp.+Nairovirus | 1 (1.25) |
| Rickettsia spp + Borrelia spp.+A. phagocytophilum | 1 (1.25) |
| Rickettsia spp.+phlebovirus+Orthonairovirus | 1 (1.25) |
| Rickettsia spp + Borrelia spp.+ phlebovirus+ Orthonairovirus | 2 (2.5) |
| Rickettsia spp. + Borrelia spp. + Orthonairovirus | 3 (3.75) |
| Negative | 20 (25.0) |
| Total | 80 (100.0) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



