Submitted:
15 April 2024
Posted:
15 April 2024
You are already at the latest version
Abstract
The pharmacological effects of the fruiting body of Ophiocordyceps sinensis (Os) exerted antioxidant, anti-virus, and immunomodulatory, whereas the anti-inflammatory effects and active components of Os mycelial submerged culture (OMSC) are not known. This study aimed to investigate the active compounds in the fermented liquid (FLOS), hot water (WEOS), and 50-95% (EEOS-50, EEOS-95) ethanol extracts of OMSC and their anti-inflammatory effects and potential mechanisms in LPS-stimulated microglial BV2 cells. The results demonstrated that all of the OMSC extracts could inhibit NO production in BV2 cells. EEOS-95 exhibited the strongest inhibitory effects (41 % inhibitory ability at 500 µg/mL), and its ergosterol, GABA, total phenolic, and total flavonoid contents were significantly higher than those of the other extracts (18.60, 18.60, 2.28, and 2.14 mg/g, p < 0.05, respectively). EEOS-95 also has a strong inhibitory ability against IL-6, IL-1β and TNF-α with IC50 of 617, 277 and 507 µg/mL, respectively, which is higher than that of 1 mM melatonin. The anti-inflammatory mechanism of EEOS-95 seems to be associated with up-regulation of PPAR-γ/Nrf-2/HO-1 antioxidant-related expression and down-regulation of NF-κB/COX-2/iNOS pro-inflammatory expression signaling. Therefore, we hypothesized that EEOS-95 can prevent neurodegenerative diseases, but the effect on the application in vivo should be further investigated.
Keywords:
Ophiocordyceps sinensis
; neurodegenerative disease
; microglial
; anti-inflammatory
; cytokines
; submerged culture
1. Introduction
The caterpillar fungus Ophiocordyceps sinensis (Os), known as Yarshagumba in Nepal and Dong-chong-xia-cao in Chinese, is found in the alpine regions of Bhutan, India, China, and Nepal [1,2]. O. sinensis has been treasured for centuries in traditional medicine. Primarily used as a tonic to boost immunity, it holds immense potential for various therapeutic applications. Several studies have documented its diverse beneficial properties, making it a fascinating subject for scientific exploration. Previous research has highlighted the potential of Os in addressing a multitude of health concerns. Some of its notable attributes in nervous system diseases, immunomodulatory, renal, liver, cardiovascular; especially, it inhibited the severe acute respiratory syndrome virus (SARS virus) 2003 in China [3,4]. Besides, Os exerted an excellent source of phytochemicals such as phenolic acids, amino acids, fatty acids, sterols, polysaccharides, nucleosides, etc… which have contributed to various beneficial nutritional or pharmacological properties [4]. Os traditionally consumed in powdered form, mixed with milk or water, or added to tea or soup. The high price of Os is mainly due to its medicinal value, which has led to an increasing demand from consumers and a gradual decrease in the production of natural wild species (fruiting bodies). Wei et al. warn that climate change further threatens its wild distribution [5]. Based on the above reasons, some researchers have carried out mycelium isolation technology and used the isolated Os mycelial for submerged culture [3]. The benefits of this new approach are twofold: it reduces pressure on wild populations and promotes ecological sustainability, while providing a more economically viable source of Os. Although many studies have been conducted in the past to demonstrate the effects of fruiting body of Os on human health, the functionality of submerged cultures of Os mycelia and the active compounds in mycelia and fermentation still need to be further verified.
Neurodegenerative diseases (NDs) are a group of progressive conditions that affect millions of people worldwide, predominantly in their later years. These devastating disorders attack the nervous system, particularly the brain, leading to a decline in cognitive and motor function. Alzheimer's disease, Parkinson's disease, Huntington's disease, and amyotrophic lateral sclerosis (ALS) are just a few examples of the many faces of NDs. The rising number of ND cases puts immense pressure on healthcare systems, families, and communities [6,7,8,9,10]. While the specific causes of each NDs vary, researchers have identified several key factors that contribute to their development, these include oxidative stress (an imbalance between free radicals and antioxidants) leading to cellular damage, protein aggregation (the abnormal buildup of misfolded proteins in the brain, forming toxic clumps), neuro-inflammation (chronic inflammation in the nervous system damaging brain cells), neurotransmission impairment (disruption of chemical communication between neurons), mitochondrial dysfunction (impaired energy production in brain cells), excitotoxicity (excessive stimulation of brain cells, leading to their death) [11].
Neuroinflammation is a defensive response of the brain to injury or infection, occurring in both chronic and acute neurodegenerative disorders. Microglia, the innate immune cells of the central nervous system, become rapidly activated when the brain is infected, injured, or damaged. They regulate the production of proinflammatory cytokines and immune response mediators, including nitric oxide (NO), tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), reactive oxygen species (ROS), and various neurotoxic mediators [12,13]. Therefore, inhibiting the activation of microglia and neuroinflammation is a potential therapeutic target for reducing neurodegenerative diseases and neuronal damage.
The important bioactive compounds detected in Os mycelia are as follows: adenosine, polysaccharides, ergosterol, cordycepin, etc. [14,15]. These compounds have been reported to have various biological and pharmacological effects such as immunomodulatory, anti-inflammatory, and antioxidant. [3]. Li et al. reported that bioactive compounds of mycelia isolated from wild Os had neuroprotective effects on PC12 cells and prevented hydrogen peroxide-induced nerve damage in rats [16]. However, to date, no analytical data are available on the effects of OMSC on BV2 microglia and the analysis of the bioactive components of their mycelial extracts.
Thus, in the present report, we explored the contents of active compounds of fermented liquid (FLOS), and various mycelial extracts (hot water -WEOS, 50 % -EEOS-50, and 95 % -EEOS-95 ethanol extracts) of Os mycelial submerged culture (OMSC) by RP-HPLC assayed and evaluated their potential on cytotoxicity and mediator NO production. Then, their anti-inflammatory effects and possible mechanisms of OMSC in microglial BV2 cells induced by lipopolysaccharides (LPS) will be studied.
2. Materials and Methods
2.1. Chemicals
The mouse microglial BV2 cell line was purchased from the Food Industry Research and Development Institute (Hsinchu, Taiwan, R.O.C). Lipopolysaccharide (LPS; Escherichia coli O26:B6) and bovine serum albumin were bought from Sigma-Aldrich Co. (St Louis, MO, USA). RPMI 1640, and fetal bovine serum (FBS) from Gibco by Life Technologies (Frederick, MD, USA). Chemicals and reagents are of high quality. Tips, dishes, test tubes, etc. for cell culture were bought from Thermo Fisher Scientific (Waltham, MA, USA).
2.2. Preparation of Hot Water and Ethanol Extracts from Ophiocordyceps Sinensis Submerged Culture
Ophiocordyceps sinensis (BCRC 930166) were collected from wild Ophiocordyceps sinensis (Qinghai Province, China), and identified by Bioresource Collection and Research Center (Hsinchu, Taiwan, R.O.C.). This experiment used the dried mycelia (Os H101) kindly provided by Professor Hsu Tai-Hao. Os H101 were cultured at pH 7.0, 100×g, 18 °C, and 15 days in a 2-L Erlenmeyer flask containing (3% glucose, 0.5% peptone, 0.3% yeast extract, 0.1% H2PO4, and 0.05% MgSO4.7 H2O) on a rotary shaker (120 rpm). At the end of the fermentation process, the mycelial submerged culture of Os H101 (OMSC) was harvested. OMSC was extracted with a 1:20 ratio (w/v) of 121 ± 2 ◦C hot water for 15 mins; or extracted with a 1:20 ratio of 50%, 95% ethanol by soaking at room temperature for 24 h. The extracts were centrifuged (1500 rpm, 10 min) and concentrated at reduced pressure by using a rotary evaporator. Then, the OMSC extracts were lyophilized to obtain fermented liquid of Os (FLOS), 90°C hot water extract of Os (WEOS), 50 % ethanol extract of Os (EEOS-50), 95 % ethanol extract of Os (EEOS-95), respectively. Finally, samples were stored at 4°C for the following analysis. The extraction yield of WEOS (32.9%), EEOS-50 (25.1%), EEOS-95 (7.4%), respectively.
2.3. Determination of Bioactive Compounds
The bioactive compounds of OMSC were carried out using an Agilent 1200 reversed phase High-performance liquid chromatography coupled with a diode-array detector (Hitachi, Chiyoda City, Japan, Chromaster 5430). A HIQ Sil C18W reversed-phase column was used (4.6 mm × 250 mm, 5 µm). The results were expressed in mg/g. All solvents were of HPLC grade and filtered before HPLC analysis.
2.3.1. Measurement of Adenosine
Adenosine was determined by applying the method described in Chang et al., [17] with a slightly modified. The sample or standard was dissolved in 1 mL distilled water, and ultrasound-assisted (ultrasonic power 100 W) extracted for 1 h. The mobile phase consisted of methanol:0.02 M KH2PO4 (15:85, v/v). The flow rate was 0.7 mL/min, absorbance was measured at 254 nm, and the injection volume was 20 µL.
2.3.2. Measurement of Ergosterol
Ergosterol was detected as described by Yuan et al., [18]. Sample or standard (10 mg) was dissolved in 1 mL methanol/ dichloromethane (75: 25, v/v), and shaken for 1 h. The mobile phase consisted of solvents A (80% methanol) and B (75% methanol in dichloromethane) with gradient elution as follows: 0–5 min (100% A); 5–19 min (0%–100% B); 20 min (100% B); 34-35 min (100% B). The flow rate was 1.0 mL/min, absorbance was measured at 280 nm, and the injection volume was 20 µL.
2.3.3. Measurement of GABA
Measurement of GABA was performed as previously described by Rog´erio da Silva Moraes et al., [19]. The mobile phase was A: composed of 50 mM sodium acetate, 5% methanol, and 2-propanol (pH 5.67) and B: 70% methanol. The gradient is as follows: 0–10 min: 100% A, 10–20 min: 70% A, 20–25 min: 50% A, 25 min: 100% A. The flow rate was 0.8 mL/min, absorbance was measured at 338 nm, and the injection volume was 20 µL.
Prepare chemical as follows: derivatization working solution (OPA-NAC complex) to allow reaction between OPA and NAC thiol group: 16.3 mg N-acetylcysteine (NAC), 13 mg o-phthalaldehyde (OPA), 300 µL methanol was react for 30 min, room temperature, dark, stored in plastic test tubes at 4°C, in dark (used within 24 hours after preparation); and borate buffer (pH 9.6).
GABA standard or samples were mixed: 60 µL standard or sample, 40 µL borate buffer, and 10 µL OPA-NAC complex; after 10 min of reaction, vortex this final solution prior to HPLC analysis.
2.3.4. Measurement of TPC, TFC, and EPS Contents
Extracellular polysaccharides (EPS) were detected following the phenol-sulfuric colorimetric method described by Jiménez et al., [20].
Total phenolic content (TPC) and total flavonoid content (TFC) were measured by applying the Folin-ciocalteu and aluminum chloride colorimetric methods according to the method presented in Yang et al., [21].
2.4. Cytotoxicity Test of OMSC on BV2 Microglial
The mouse microglial BV2 cell line was purchased from the Food Industry Research and Development Institute (Hsinchu, Taiwan, R.O.C). Cells were cultured in RPMI 1640, containing 2 mM L-glutamine, 1.5 g/L sodium bicarbonate, 10% fetal bovine serum, and 1% antibiotic penicillin/streptomycin. The medium for cells used in the experiments was changed every 2 days, incubated at 37 °C and 5% CO2.
The cytotoxicity evaluation of OMSC on BV2 microglial cells was determined by the MTT method described in Vo et al., [22]. Cells were treated with various concentrations (10–1000 µg/mL) of FLOS, WEOS, EEOS-50, and EEOS-95; and cell viability was detected at 490 nm using an ELISA reader (Synergy HTX, BioTek, Winooski, VT, USA).
2.5. Measurement of Nitrite Production
The evaluation effect of OMSC on NO level was carried out by measuring nitrite accumulation as described in Vo et al. [22] with minor modifications. BV2 cells (5 × 105 cells/mL) were seeded in 24 well plates for 2 h. Cells were treated with 10–1000 µg/mL of FLOS, WEOS, EEOS-50, and EEOS-95 for 24 h, respectively. LPS-stimulated (1 µg/mL) for 24 h, and nitrite levels were detected at 540 nm using an ELISA reader (Synergy HTX, BioTek, Winooski, VT, USA).
2.6. Anti-Inflammatory Activities of 95% Ethanol Extract OMSC (EEOS-95)
2.6.1. Measurement of Pro-Inflammatory Cytokines Level
BV2 cells (5 × 105 cells/well) were seeded in 24 well plates for 2 h and treated with different concentrations (50-500 µg/mL) EEOS-95 for 24 h, then incubated for 24 h with 1 µg/mL LPS to induce the secretion of inflammatory-related cytokines. After incubation, the concentration of cytokines in the culture medium was determined using an ELISA kit according to the manufacturer’s instructions. Cytokines were measured as IL-1β, IL-6, TNF-α (Invitrogen Co. Camarillo, CA, USA), and PGE2 (Life Technologies Corp. Frederick, MD, USA).
2.6.2. Western Blotting Analysis
Cell pellets were harvested and washed with cold phosphate buffer saline (pH 7.4). The cytoplasmic proteins 50 µg were electrophoresed on 10% sodium dodecyl sulfate-polyacrylamide gels (SDS-PAGE) and transferred to a poly-vinylidene fluoride membrane. The membrane was blotted and incubated with specific primary antibodies overnight at 4 °C, followed by incubation with horseradish peroxidase-conjugated secondary antibody. Finally, the blots were probed using enhanced chemiluminescence and autoradiographed. The relative density of protein expression was quantified using ImageJ software, developed by Wayne Rasband at the National Institutes of Health and the Laboratory for Optical and Computational Instrumentation (LOCI, University of Wisconsin, USA). The protein content in the supernatant was determined using the BCA protein assay kit (Thermo, Rockford, IL, USA). Western blot analysis detected the protein expression of iNOS, COX-2, NF-κB, HO-1, Nrf-2, PARP-γ, and β-actin in the BV2 microglial cells.
2.7. Statistical Analysis
All statistical analyses were performed using SPSS for Windows, version 18 (SPSS Inc., Chicago, IL, USA). Data are expressed as means ± standard deviation and analyzed using one-way ANOVA followed by Duncan’s multiple range test. p < 0.05 is considered statistically significant.
3. Results
3.1. Bioactive Compounds in Extract of OMSC
As shown in Figure 1, the HPLC profile standard compounds of adenosine, ergosterol, and GABA; and various extracts of Os mycelia were detected by the HPC-DAD-UV/Vis system. Figure 1A,E, the chromatogram shows that adenosine and GABA were found in all extracts of OMSC following FLOS, WEOS, EEOS-50, and EEOS-95. The full wavelength of the sample is compared with that of the standard and has the same waveform. (Figure 1B,F). However, the ergosterol peak was only detected in EEOS-95 (Figure 1C), as evidenced by the ergosterol standard full-wavelength waveform. (Figure 1D). As shown in Table 1, the dried matter of the fermented liquid of OMSC (FLOS) was rich in adenosine and polysaccharides (2.12 and 107.60 mg/g, respectively). Except for adenosine and polysaccharides (2.83 and 156.30 mg/g, respectively), GABA was also determined in the hot water extract of OMSC (WEOS) (12.60 mg/g), and the content was significantly lower than that of EEOS-95 (18.60 mg/g) (p < 0.05). It is noteworthy that EEOS-95 was significantly higher (p < 0.05) in all active compounds except adenosine and polysaccharides which were lower than the other extracts, especially ergosterol (18.60 mg/g), while its total polyphenols, total flavonoids, and GABA were 2.28, 2.14, and 18.6 mg/g, respectively.
3.2. Cytotoxicity Evaluation of OMSC
The cytotoxicity of OMSC mycelia extracts and FLOS (50-1000 µg/mL) in BV2 microglial cells was evaluated using MTT analysis after 24 h incubation. Table 2 indicated that treatment of all extracts at concentrations up to 1000 (µg/mL) was not effective on cell survival of BV2 cells (p < 0.05) (cell viability > 90%). Thus, there was no toxic effect when BV2 cells were treated with 50 to 1000 (µg/mL) of OMSC extracts. The results also indicated that below a concentration of 500 µg/mL, the highest percentage of proliferation increased by EEOS-95 (about 68.5%). At a concentration of 1000 µg/mL, the cell viability of all the samples began to show a decreasing trend.
3.3. Effect of Os Mycelia on Nitric Oxide
We examined the effect of various extracts of OMSC on LPS-induced NO production by Griess reagent assay (Table 3). BV2 cells were stimulated with LPS (1 μg/mL), and the NO level significantly increased (0.78 nmol/106 cells) compared with the control (0.33 nmol/106 cells) (p < 0.05). Our results also indicated that cells pre-treated with different concentrations (50-500 µg/mL) of extracts from OMSC mycelia-WEOS, EEOS-50, and EEOS-95 significantly suppressed the NO production by LPS-stimulated BV2 cells in a dose-dependent manner (p < 0.05), the inhibition effects at 500 µg/mL were 25, 26 and 41%, and EEOS-95 had the highest inhibition effect on NO production. FLOS also inhibited 25% of NO levels; however, there was no significant difference among treatment doses. (p < 0.05).
3.4. Anti-Inflammation Effect of EEOS-95 on Lipopolysaccharide-Induced BV2 Microglial Cells
We further tested EEOS-95, which affected the cytokine production induced by LPS in BV2 microglial cells. The level of IL-1β, TNF-α, IL-6, and PGE2 was determined by ELISA kit. As shown in Table 4, LPS markedly increased IL-1β, IL-6, TNF-α, and PGE2 levels, as compared with untreated controls (p < 0.05); however, all of these pro-inflammatory cytokines were significantly decreased by EEOS-95 (50-500 µg/mL) in a concentration-dependent manner. In addition, at a concentration of 500 µg/mL, the inhibitory effects of EEOS-95 on IL-1β, IL-6, TNF-α and PGE2 production (96.60, 70.33, 49.83, and 43.57%, respectively, p < 0.05) were significantly better than that of 1 mM MT (30.22, 61.60, 11.40 and 6.70%, respectively, p < 0.05). The inhibition ratio of EEOS-95 on different cytokines showed that EEOS-95 had the best inhibition effect on PGE2, followed by IL-1β, TNF-α, and IL-6, with inhibition rates of 90, 60, 50, and 40%, respectively, p < 0.05, at a concentration of 500 µg/mL (Figure 2).
3.5. Effect of EEOS-95 on the Expression of Inflammatory Proteins in LPS-Induced BV2 Microglial
Western blot analyses were performed to determine whether EEOS-95 has a direct effect on the pro-inflammatory factor in LPS-induced BV2 microglial. As shown in Figure 3A, LPS significantly induced the expression of inflammation-associated proteins such as iNOS, COX-2, and NF-κB, which were dose-dependently (50-500 µg/mL) reduced by EEOS-95 (p < 0.05). At a concentration of 500 µg/mL, EEOS-95 showed a 34, 55, and 20% decrease in protein expression of iNOS, COX-2, and NF-κB, respectively, compared to the LPS-treated group alone (Figure 3B,C,D, p < 0.05). Pretreatment of BV2 cells with 1 mM melatonin also reduced the expression levels of 12% of iNOS and COX-2 and did not affect NF-κB compared with the LPS group only.
3.6. Effect of EEOS-95 on the Expression of Anti-Inflammatory Protein in LPS-Induced BV2 Microglial
As the results were shown in Figure 4, cells incubated with EEOS-95 significantly increased the expression levels of antioxidant factors (HO-1, Nrf-2, PPAR-γ) in the LPS-induced BV2 microglial. The protein expression level of HO-1, Nrf-2, and PPAR-γ in BV2 cells treated with EEOS-95 at 25-250 μg/mL showed a significant increase (p < 0.05) compared with the control group and the LPS-treated group. In contrast, EEOS-95 at a concentration of 500 µg/mL increased the expression of the control group by more than 50% and was superior to that of the 1 mM MT-treated group.
4. Discussion
The wild caterpillar fungus Os is a traditional Chinese medicine, almost found in Asia for its rich bioactive profile and diverse therapeutic applications, also has over 30 bioactive compounds that exhibit numerous beneficial effects, including anti-inflammatory, antioxidant, anti-tumor, immunomodulatory, and anti-osteoporotic activities, proving effective in tackling ailments like diabetes, liver cancer, and kidney cancer [15,23,24]. Overharvesting of wild Os has significantly reduced its annual production, causing a gap between rising demand and declining supply. This has fueled research into sustainable alternatives, including artificial cultivation using C. militaris and submerged culture techniques for natural Os [24,25,26,27,28]. These approaches focus on optimizing culture conditions, maximizing mycelial biomass production, and exploring polysaccharide extraction. While substantial research has investigated the pharmacological potential of OMSC, a vital knowledge gap remains concerning the detailed analysis of their extracted bioactive components. Our study employed aqueous and ethanolic solvents to extract bioactive compounds from OMSC to identify their main bioactive components and anti-inflammatory potential.
Our quantitative analysis of the dried powder of OMSC extract using RP-HPLC revealed six bioactive ingredients: adenosine, ergosterol, polysaccharide, TPC, TFC, and GABA. Briefly, Adenosine belongs to nucleoside, which is an essential bioactive compound in the organism. The content of adenosine is 3.29 mg/g (EEOS-50), which was similar to previous reports (3.06 mg/g) and remarkably higher than that of natural Os [15]. Zhang et al. also pointed out that adenosine is neuronal protection in NDs such as Alzheimer’s and Parkinson’s disease, suggesting its value as a marker for quality control in Os submerged fermentation [29]. However, the presence of these diverse bioactive compounds warrants further investigation into their potential health benefits and applications.
Ergosterol is an essential component of fungal cell membranes called main fungal sterol; its function is similar to cholesterol in animals, which can be converted to vitamin D2 under ultraviolet radiation or sunlight. The vegetarian body needs every day to provide vitamin D through a source of mushrooms; thus, ergosterol is interesting in further research [30]. The pharmacological effects of ergosterol have also been reported as an antioxidant, anti-inflammatory, anti-neurodegenerative, antimicrobial, anticancer, antidiabetic, etc... [31,32,33,34,35,36,37,38,39]. As reported by Peng and colleagues, ergosterol is the active compound of cultured mycelium C. sinensis, which has inhibited liver fibrosis [38]. Herein, we confirm that ergosterol is only present in EEOS-95 (18.60 mg/g), which is 6 times higher than other published data in the literature (3.20 mg/g) [14]. Thus, the potential of phytol-ergosterol from the OMSC will be interesting for future biomedical applications.
GABA is γ-aminobutyric acid, known as a non-protein amino acid, which has been biosynthesized from glutamic acid in the human body. GABA is considered the major inhibitory neurotransmitter in the central nervous system. GABA exhibited its inhibitory effect through various parts of the nervous system, including the cerebellum, hippocampus, hypothalamus, striatum, and spinal cord [40]. A previous study pointed out that biosynthetic GABA intake may have a beneficial effect on stress reduction and sleep improvement [41]. Surprisingly, the content of GABA in EEOS-95 (18.60 mg/g) is much higher than those of C. militaris (0.0686–0.180 mg/g) and C. sinensis (0.220 mg/g) [24,42]. Thus, the GABA-rich OMSC will be a potential source of natural neuroprotection.
Previous studies have addressed Cordyceps (Ophiocordyceps) sinensis, which contain polysaccharide compositions including arabinose, xylose, galacturonic acid, glucose, mannose, galactose, rhamnose [43,44]. These polysaccharides have exerted different pharmacological effects, involving antioxidant, anti-inflammatory, immunomodulatory, anti-cancer, and neuroprotection [15]. Nguyen et al. demonstrated that polysaccharides extracted from Os mycelia possess strong antioxidant effects and against hepatotoxicity in rats [45]. Additionally, three kinds of polysaccharides- glucose, mannose, and galactose isolated from Os, have strong antioxidation activity and neuroprotective properties against the free radical-induced neuronal cell toxicity on the cultured rat PC12 cells [16]. The content of polysaccharides in WEOS is 156.30 mg/g, which is similar to artificial cordyceps mycelia polysaccharide of submerged fermentation by previous reports (162.30 mg/g) [14]. Given their numerous potential health benefits, polysaccharides from cordyceps mycelia of submerged fermentation continue to be the focus of intense research and investigation in natural products with medicinal chemistry.
Evidence indicates polysaccharides can promote cell proliferation, scavenge cellular ROS, and downregulate the secretion of pro-inflammatory cytokines [46,47]. In the cell viability test, we also found that FLOS, WEOS, and EEOS-50 EEOS-95 promoted cell proliferation, which was presumably related to the polysaccharides contained in the samples. However, polysaccharides were not detected in EEOS-95, but their cell viability was higher than that of other extracts, suggesting that EEOS-95 may contain other cell proliferation components. Ergosterol is regarded as a “fungal hormone” that can stimulate growth and proliferation. Many studies have shown that ergosterol is essential for mitochondrial DNA maintenance in fungi, as cholesterol does in humans [48,49,50]. Ergosterol also exerts a differential effect on Androgen-dependent LNCaP and Androgen-independent DU-145 cancer cells; that is, ergosterol showed an antiproliferative effect on LNCaP and proliferative effect on DU-145. Thus, the promoted cell proliferative effect of EEOS-95 in BV2 cells (Androgen-independent cells) should contribute from the ergosterol.
Natural phenolic or flavonoid compounds, a class of bioactive compounds found in medicinal plants and mushrooms, are known for their antioxidant properties. These properties are directly attributed to their scavenging ability, provided by the hydroxyl groups within their structures, and contribute to improvements in human health. Our study demonstrated that flavonoids were extracted from medicinal plants (ex., daidzein, genistein, genistin, cajanol, and biochanin A), which exhibit several pharmacological properties on antioxidant, oxidative stress anti-inflammation, anti-glycemic, and apoptosis [22,51]. TPC and TFC contents of fermented liquid (FLOS) and all mycelia extracts of Os submerged culture were 1.57~2.28 mg/g (TPC) and 1.17~2.14 mg/g (TFC). Tran et al [52] pointed out that the amounts of gallic acid, quercetin, quercitrin, and hesperidin dominated the Ophiocordyceps Sobolifera extracts at 193.60 142.07; 544.53 and 110.08 µg/g, respectively, out of a total of 990.27 µg/g dry weight active phenolic fraction, and these phenolic compounds of Ophiocordyceps Sobolifera extract were responsible for renal injury prevention [52]. Thus, we suggested that TPC and TFC present in OMSC extracts enhance the benefit of pharmacological properties of OMSC for functional foods. Our results revealed that EEOS-95 was the most potent extract in OMSC by inhibiting NO production induced by LPS in BV2 cells, which is related to their high contents of active compounds (ergosterol, TPC, TFC, and GABA).
Activation of microglia leads to the production of excessive inflammatory molecules and deleterious consequences leading to neuronal death, which has been thought to contribute to the pathogenesis of neurodegenerative diseases; cause of acute injuries (stroke, traumatic brain injury) or chronic neurodegeneration (such as Alzheimer’s disease, Parkinson’s disease, and chronic traumatic encephalopathy) [53]. Recent studies have reported that some compounds such as nucleoside, ergosterol, GABA, polysaccharides, and cordycepin were isolated from Cordyceps militaris exerted inhibited the production of NO, reduced pro-inflammatory cytokines; and they possessed an effect of neuroprotection by inhibiting microglia-mediated inflammation in LPS-induced microglia BV2 cells [35,54]. In the present study, our results demonstrated that EEOS-95 has significantly inhibited the levels of cytokines (IL-1β, IL-6, TNF-α, and PGE2) secreted and decreased the production of NO in LPS-stimulated BV2 cells. Therefore, EEOS-95 is possible for natural ingredients to be used as therapeutics for neurodegenerative disorders with neuro-inflammation.
Peroxisome proliferator-activated receptor gamma (PPAR-γ) is a subunit of the PPAR and is a ligand-activated nuclear transcription factor; if brain injury, PPAR-γ can be “key” of cytoprotective stress responses and enhance the chances of cellular survival [53]. Furthermore, several recent studies showed that PPAR-γ is also capable of inhibiting other transcription factors, and has not only been implicated in the downregulation transcription and expression of related genes involved in proinflammatory cytokines (IL-1β, IL-6, and TNF-α), and neuro-inflammatory genes (COX-2 and iNOS); as well as it can be inhibiting active-NF-κB signals; playing a neuroprotective role of microglia [53,55].
Luteoloside (a flavonoid compound) significantly upregulated PPAR-γ and Nrf-2 and decreased the release of proinflammatory cytokines in focal cerebral ischemia on middle cerebral artery occlusion (MCAO) rats by inhibiting the NF-κB pathway [56]. Ergosterol isolated from mushrooms (Auricularia polytricha, Cordyceps militaris) attenuates bisphenol A or LPS-induced BV2 microglial cell inflammation [57,58]. Zheng et al. indicated that GABA-enriched Moringa oleifera leaves (MLFB) fermentation broth also could effectively alleviate LPS-induced inflammatory response by inhibiting the secretions of pro-inflammatory cytokines and the anti-inflammatory activity might be related to the relatively high contents of GABA, flavonoids, phenolics and organic acids in MLFB, its mechanism might be associated with the inhibition of TLR-4/NF-κB inflammatory signaling pathway activation [59]. Actually, treatment of EEOS-95 significantly inhibited the expression of NF-κB, iNOS, and COX-2; also enhanced upregulated of Nrf-2 and HO-1; so, we suggested that EEOS-95, an effective activator of PPAR-γ, will be an attenuated neuro-inflammation through by modulating the PPAR-γ/Nrf-2/HO-1/NF-κB signaling pathway, and ergosterol, GABA, flavonoids and polyphenol may be responsible for their activity.
Additionally, the nuclear factor Nrf-2 is known as another transcription factor and master regulator of detoxification and antioxidant regulation, which may play a main role in neuroprotective function [60]. Some documents suggested that it correlates with PPAR-γ and Nrf-2, they exerted against oxidative stress, which effectively reduced the inflammatory response by inhibiting NF-κB signals [61,62]. Besides, Duan et al., pointed out the synergistic effect of PPAR-γ and Nrf-2 pathway to upregulate the expression of related genes and inhibit ferroptosis-induced neuronal injury in intracerebral hemorrhage rats in vitro and in vivo [62]. Furthermore, pro-oxidant HO-1 expression is upregulated by oxidative stress, nitric oxide, CO, and hypoxia. Choi and colleagues also highlighted that HO-1 is present and its role in neurovascular diseases, such as age-related macular degeneration (AMD), ischemia-reperfusion injury, traumatic brain injury, and Alzheimer’s disease [63]. The abnormal HO-1 levels with Nrf-2 dysfunction are implicated in the pathogenesis of neurovascular systems related to ischemia, trauma, and aging; thus, Nrf-2/ HO-1 signal mechanism is involved in the development, oxidative stress responses, and anti-inflammation [64].
This research mainly studied the effect of EEOS-95 on PPAR- γ, an activator that inhibits oxidative stress, and against neuronal inflammation through the synergistic actions of the expression- PPAR-γ, Nrf-2, and HO-1 pathways in LPS-stimulated in the microglia BV2 cells (Figure 5). Furthermore, EEOS-95 has exhibited strong pharmacological properties and exerted a potential neuro-inflammatory effect; so, we will continue to investigate deeply explore the molecular mechanism by which PPAR-γ/Nrf-2/HO-1 pathway to neuro-protection in vivo, safe-dose, as well as preclinical and clinical in future studies of 95% ethanol extract of OMSC mycelia.
5. Conclusions
The findings demonstrate the potential of EEOS-95 to be developed into functional food due to its against neuro-inflammatory by upregulated PPAR-γ/Nrf-2/HO-1 inhibited NF-κB/COX-2/iNOS pathways to decrease pro-inflammatory cytokines (IL-1β, IL-6, and TNF-α). We also found six types of bioactive components involving adenosine, ergosterol, polysaccharide, GABA, TPC, and TFC present in the extracts of OMSC. The synergistic of these phytochemicals would contribute to pharmacological properties in neuroprotection on LPS-induced BV2 microglia cells. The further identified bioactive ingredients components and proposals of possible mechanisms to obtain those bioactive compounds in the molecular mechanism will continue the further investigation. Thus, OMSC possesses a potential source of neuroprotective and also contributes to the function of food or nutraceutical pharmacy.
Author Contributions
Conceptualization, Y.-J.S, H.-C.H., and T.-H.H; methodology and software, Y.-J.S, H.-C.H, T.-L.T.V., T.-H.H., and T.-Y.S.; formal analysis, Y.-J.S, H.-C.H, and T.-L.T.V.; investigation, H.-C.H.; data curation, Y.-J.S.; writing—original draft preparation, H.-C.H, T.-L.T.V., T.-Y.S.; writing—review and editing, T.-L.T.V., and T.-Y.S.; supervision, T.-Y.S.; project administration, T.-Y.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science and Technology Council in Taiwan, grant number MOST 104-2632-E-212-001 and MOST 105-2320-B-212-002. The APC was funded by Da-Yeh University.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chakraborty, S.; Chowdhury, S.; and Nandi, G. Review on Yarsagumba (Cordyceps sinensis)- an exotic medicinal mushroom. Int J PharmacognPhytochem Res 2014, 6(2), 339–346. [Google Scholar]
- Pandey, K. A review of the medicinal value of Ophiocordyceps sinensis (Yarshagumba). Asian J Pharmacognosy 2022, 6(1), 6–9. [Google Scholar]
- Yan, J.K.; Wang, W.Q.; Wu, J.Y. Recent advances in Cordyceps sinensis polysaccharides: Mycelial fermentation, isolation, structure, and bioactivities: a review. J Funct Foods 2014, 6, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; Karati, D.; Priyadarshini, R.; Dua, T.K.; Paul, P.; Sahu, R.; Nandi, G. Cordyceps sinensis (yarsagumba): Pharmacological properties of a mushroom. Pharmacol Res Mod Chin Med 2023, 8, 100294.
- Wei, Y.; Zhang, L.; Wang, J.; Wang, W.; Niyati, N.; Guo, Y.; Wang, X. Chinese caterpillar fungus (Ophiocordyceps sinensis) in China: Current distribution, trading, and futures under climate change and overexploitation. Sci Total Environ 2021, 755, 142548. [Google Scholar] [CrossRef]
- Knopman, D.S.; Petersen, R.C. Mild cognitive impairment and mild dementia: a clinical perspective. Mayo Clin Proc 2014, 89(10), 1452–1459. [Google Scholar] [CrossRef] [PubMed]
- Hague, S.; Klaffke, S.; Bandmann, O. Neurodegenerative disorders: Parkinson’s disease and Huntington’s disease. J Neurol Neurosurg Psychiatry 2005, 76, 1058–1063. [Google Scholar] [CrossRef] [PubMed]
- Teoh, H.L.; Carey, K.; Sampaio, H.; Mowat, D.; Roscioli, T.; Farrar, M. Inherited Paediatric Motor Neuron Disorders: Beyond Spinal Muscular Atrophy. Neural Plast 2017, 2017, 6509493. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.D.; Mao, S.S.; Lin, L.; Bai, G.N.; Liu, B.J.; Mao, J.H. Stress granules in the spinal muscular atrophy and amyotrophic lateral sclerosis: The correlation and promising therapy. Neurobiol Dis 2022, 170, 105749. [Google Scholar] [CrossRef] [PubMed]
- Klockgether, T; Mariotti, C.; Paulson, H.L. Spinocerebellar ataxia. Nat Rev Dis Primers 2019, 5(1), 24. [Google Scholar] [CrossRef] [PubMed]
- Moreira, J.; Machado, M.; Dias-Teixeira, M.; Ferraz, R.; Delerue-Matos, C.; Grosso, C. The neuroprotective effect of traditional Chinese medicinal plants—A critical review. Acta Pharm Sin B 2023, 13(8), 3208–3237. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Yang, J.; Wang, X.; Yang, P.; Zhao, Y.; Li, K.; Chen, Y.; Xu, Y. Herbal Compounds Play a Role in Neuroprotection through the Inhibition of Microglial Activation. J Immunol Res 2018, 2018, 9348046. [Google Scholar] [CrossRef]
- Gupta, D.P.; Park, S.H.; Lee, Y.S.; Lee, S.; Lim, S.; Byun, J.; Cho, I.H.; Song, G.J. Daphne genkwa flower extract promotes the neuroprotective effects of microglia. Phytomedicine 2023, 108, 154486. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.Q.; Wang, C.Y.; Zhao, R.; Li, H.H.; Zhang, Q.H.; Cai, D. Artificial cordyceps mycelium by submerged fermentation of Hirsutella sinensis HS 1201 using rice bran hydrolysate as substrate. Environ Qual Manag 2021, 31(1), 109–118. [Google Scholar] [CrossRef]
- Chatnarin, S. and Thirabunyanon, M. Potential bioactivities via anticancer, antioxidant, and immunomodulatory properties of cultured mycelial enriched β-D-glucan polysaccharides from a novel fungus Ophiocordyceps sinensis OS8. Front Immunol 2023, 14, 1150287. Front Immunol, 2023; 14, 1150287.
- Li, S.P.; Zhao, K.J.; Ji, Z.N.; Song, Z.H.; Dong, T.T.; Lo, C.K.; Cheung, J.K.; Zhu, S.Q.; Tsim, K.W. A polysaccharide isolated from Cordyceps sinensis, a traditional Chinese medicine, protects PC12 cells against hydrogen peroxide-induced injury. Life Sci 2003, 73(19), 2503–2513. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.Y.; Lue, M.Y.; and Pan, T.M. Determination of adenosine, cordycepin and ergosterol contents in cultivated Antrodia camphorata by HPLC method. J Food Drug Anal 2005, 13(4), 338–342. [Google Scholar] [CrossRef]
- Yuan, J.P.; Wang, J.H.; Liu, X.; Kuang, H.C.; and Zhao, S.Y. Simultaneous determination of free ergosterol and ergosteryl esters in Cordyceps sinensis by HPLC. Food Chem 2007, 105, 1755–1759. [Google Scholar] [CrossRef]
- Rogério da Silva Moraes, E.; Santos-Silva, M.; Grisólia, A.A.; Braga, D.V.; Reis Leão, L.K.; Bahia, C.P.; Soares de Moraes, S.A.; Passos, A.F.; de Jesus Oliveira Batista, E.; Herculano, A.M.; Matos Oliveira, K.R.H. High performance liquid chromatography-based method to analyze activity of GABA transporters in central nervous system. Neurochem Int 2022, 158, 105359. [Google Scholar] [CrossRef] [PubMed]
- Quero-Jiménez, P.C.; Montenegro, O.N.; Sosa, R.; de la Torre, J.B.; Valero Acosta, J.; López Pérez, D.; Santana Rodríguez, A.; Ramos Méndez, R.; Calvo Alonso, A.; Corrales, A.J.; Broche Hernández, N. Total carbohydrates concentration evaluation in products of microbial origin. Afinidad Journal of Chemical Engineering Theoretical and Applied Chemistry 2019, 76, 195–203. [Google Scholar]
- Yang, S.E.; Vo, T.L.T.; Chen, C.L.; Yang, N.C.; Chen, C.I.; Song, T.Y. Nutritional Composition, Bioactive Compounds and Functional Evaluation of Various Parts of Cajanus cajan (L.) Millsp. Agriculture 2020, 10, 55. [Google Scholar] [CrossRef]
- Vo, T.L.T.; Yang, N.C.; Yang, S.E.; Chen, C.L.; Wu, C.H.; Song, T.Y. Effects of Cajanus cajan (L.) millsp. Roots extracts on the antioxidant and anti-inflammatory activities. Chin J Physiol 2020, 63, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.D.; Wu, C.K.; Yuan, F.; Wang, Y.B.; Huang, L.D.; Chen, Z.H.; Zeng, W.B.; Wang, Y.; Yang, Z.L.; Zeng, P.S.; Lemetti, P.; Mo, X.X.; Yu, H. Evolutionary biogeography on Ophiocordyceps sinensis: An indicator of molecular phylogeny to geochronological and ecological exchanges. Geoscience Frontiers 2020, 11(3), 807–820. [Google Scholar] [CrossRef]
- Lo, H.C.; Hsieh, C.; Lin, F.Y.; Hsu, T.H. A systematic review of the mysterious caterpillar fungus Ophiocordyceps sinensis in DongChongXiaCao and related bioactive ingredients. J Tradit Complementary Med 2013, 3(1), 16–32. [Google Scholar] [CrossRef]
- Kim, H.O.; Yun, J.W. A comparative study on the production of exopolysaccharides between two entomopathogenic fungi Cordyceps militaris and Cordyceps sinensis in submerged mycelial cultures. J Appl Microbiol 2005, 99(4), 728–738. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.H.; Yao, Y.J. Nutritional requirements of mycelial growth of Cordyceps sinensis in submerged culture. J Appl Microbiol 2005, 99(3), 483–492. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.H.; Lim, J.S.; Yoon, C.S.; Koh, J.H.; Chang, H.I.; Kim, S.W. Production of mycelia and exo-biopolymer from molasses by Cordyceps sinensis 16 in submerged culture. Bioresour Technol 2007, 98(1), 165–168. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Liu, G.Q.; Zhu, C.Y.; Zhou, G.Y.; Kuang, S.M. Enhanced production of mycelial biomass and extracellular polysaccharides in caterpillar-shaped medicinal mushroom Cordyceps sinensis CS001 by the addition of palmitic acid. J Med Plant Res 2011, 5(13), 2873–2878. [Google Scholar]
- Zhang, Y.; Cao, H.; Qiu, X.; Xu, D.; Chen, Y.; Barnes, G.N.; Tu, Y.; Gyabaah, A.T.; Gharbal, A.H.A.A.; Peng, C.; Cai, J.; and Cai, X. Neuroprotective Effects of Adenosine A1 Receptor Signaling on Cognitive Impairment Induced by Chronic Intermittent Hypoxia in Mice. Front Cell Neurosci 2020, 14, 202. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Zhang, M.; Mujumdar, A.S. UV induced conversion during drying of ergosterol to vitamin D in various mushrooms: Effect of different drying conditions. Trends Food Sci Technol 2020, 105, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lin, C.; Wang, T.; Zhang, P.; Liu, Z.; Lu, C. Ergosterol attenuates LPS-induced myocardial injury by modulating oxidative stress and apoptosis in rats. Cell Physiol Biochem 2018, 48(2), 583–592. [Google Scholar] [CrossRef]
- Liu, C.; Zhao, S.; Zhu, C.; Gao, Q.; Bai, J.; Si, J.; Chen, Y. Ergosterol ameliorates renal inflammatory responses in mice model of diabetic nephropathy. Biomed Pharmacother 2020, 128, 110252. [Google Scholar] [CrossRef] [PubMed]
- Kushairi, N.; Tarmizi, N.A.K.A.; Phan, C.W.; Macreadie, I.; Sabaratnam, V.; Naidu, M.; David, P. Modulation of neuroinflammatory pathways by medicinal mushrooms, with particular relevance to Alzheimer’s disease. Trends Food Sci Technol 2020, 104, 153–162. [Google Scholar] [CrossRef]
- Ano, Y.; Kutsukake, T.; Hoshi, A.; Yoshida, A.; Nakayama, H. Identification of a novel dehydroergosterol enhancing microglial anti-inflammatory activity in a dairy product fermented with Penicillium candidum. PLoS ONE 2015, 10(3), e0116598. [Google Scholar] [CrossRef]
- Nallathamby, N.; Guan-Serm, L.; Vidyadaran, S.; Abd Malek, S.N.; Raman, J.; Sabaratnam, V. Ergosterol of Cordyceps militaris attenuates LPS induced inflammation in BV2 microglia cells. Nat Prod Commun 2015, 10(6), 885–886. [Google Scholar] [CrossRef] [PubMed]
- Sillapachaiyaporn, C.; Chuchawankul, S.; Nilkhet, S.; Moungkote, N.; Sarachana, T.; Ung, A.T.; Baek, S.J.; Tencomnao, T. Ergosterol isolated from cloud ear mushroom (Auricularia polytricha) attenuates bisphenol A-induced BV2 microglial cell inflammation. Food Res Int 2022, 157, 111433. [Google Scholar] [CrossRef] [PubMed]
- Sillapachaiyaporn, C.; Mongkolpobsin, K.; Chuchawankul, S.; Tencomnao, T.; Baek, S.J. Neuroprotective effects of ergosterol against TNF-α-induced HT-22 hippocampal cell injury. Biomed Pharmacother 2022, 154, 113596. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Tao, Y.; Wang, Q.; Shen, L.; Yang, T.; Liu, Z.; Liu, C. Ergosterol Is the Active Compound of Cultured Mycelium Cordyceps sinensis on Antiliver Fibrosis. Evid Based Complement Alternat Med 2014, 2014, 537234. [Google Scholar] [CrossRef]
- Xiong, M.; Huang, Y.; Liu, Y.; Huang, M.; Song, G.; Ming, Q.; Ma, X.; Yang, J.; Deng, S.; Wen, Y.; et al. Antidiabetic activity of ergosterol from Pleurotus ostreatus in KK-A(y) mice with spontaneous type 2 diabetes mellitus. Mol Nutr Food Res 2018, 62(3), 1700444. [Google Scholar] [CrossRef] [PubMed]
- Boonstra, E.; de Kleijn, R.; Colzato, L.S.; Alkemade, A.; Forstmann, B.U.; Nieuwenhuis, S. Neurotransmitters as Food Supplements: The Effects of GABA on Brain and Behavior. Front Psychol 2015, 6, 1520. [Google Scholar] [CrossRef] [PubMed]
- Hepsomali, P.; Groeger, J.A.; Nishihira, J.; Scholey, A. Effects of Oral Gamma-Aminobutyric Acid (GABA) Administration on Stress and Sleep in Humans: A Systematic Review. Front Neurosci 2020, 14, 923. [Google Scholar] [CrossRef] [PubMed]
- Jędrejko, K.J.; Lazur, J.; Muszyńska, B. Cordyceps militaris: An Overview of Its Chemical Constituents in Relation to Biological Activity. Foods 2021, 10(11), 2634. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, F.; Xu, Z.; Ding, Z. Bioactive Mushroom Polysaccharides: A Review on Monosaccharide Composition, Biosynthesis and Regulation. Molecules 2017, 22(6), 955. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.H.; Xie, F.; Tan, J.; Yuan, Y.; Mei, H.; Zheng, Y.; Sheng, R. Extraction, structure and pharmacological effects of the polysaccharides from Cordyceps sinensis: A review. J Funct Foods 2022, 89, 104909. [Google Scholar] [CrossRef]
- Nguyen, Q.V.; Vu, T.T.; Tran, M.T.; Ho, P.T.T.; Le, T.H.T.; et al. Antioxidant activity and hepatoprotective effect of exopolysaccharides from cultivated ophiocordyceps sinensis against CCl4-induced liver damages. Nat Prod Commun 2021, 16(2), 1–9. [Google Scholar] [CrossRef]
- Chen, Z.; Cheng, L.; He, Y.; Wei, X. Extraction, characterization, utilization as wound dressing and drug delivery of Bletilla striata polysaccharide: A review. Int. J. Biol. Macromol. 2018, 120, 2076–2085. [Google Scholar] [CrossRef] [PubMed]
- Sung, T.J.; Wang, Y.Y.; Liu, K.L.; Chou, C.H.; Lai, P.S.; Hsieh, C.W. Pholiota nameko polysaccharides promotes cell proliferation and migration and reduces ros content in h2o2-induced l929 cells. Antioxidants (Basel). 2020, 9(1), 65. [Google Scholar] [CrossRef] [PubMed]
- Cirigliano, A.; Macone, A.; Bianchi, M.M.; Oliaro-Bosso, S.; Balliano, G.; Negri, R.; Rinaldi, T. Ergosterol reduction impairs mitochondrial DNA maintenance in S. cerevisiae. Biochim. Biophys. Acta Mol. Cell Biol. Lipids. 2019, 1864, 290–303. [Google Scholar] [CrossRef] [PubMed]
- Jordá, T. and Puig, S. Regulation of Ergosterol Biosynthesis in Saccharomyces cerevisiae. Genes (Basel). 2020, 11(7), 795. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Fonseca, M.B.; Vidal-Limon, A.; Fernández-Pomares, C.; Rojas-Durán, F.; Hernández-Aguilar, M.E.; Espinoza, C.; Trigos, A.; Suárez-Medellín, J. Ergosterol exerts a differential effect on AR-dependent LNCaP and AR-independent DU-145 cancer cells. Natural Product Research 2021, 35(22), 4857–4860. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.L.T.; Cai, X.M.; Liao, J.W.; Huang, L.G.; Chen, C.L.; Wu, C.H.; Song, T.Y. Safety Assessment and Hepatic–Renal Protection of Cajanus cajan (L.) Millsp. Root and Its Soy Isoflavone Contents. Nutrients 2023, 15, 3963. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.V.T.; Tran, V.K.; Le, L.S.; Tran, T.M.; Nguyen, D.G.C.; Trinh, T.K.; Nguyen, Q.V.; Nguyen, C.C.; Le, T.H. Phenolic content and antioxidant activity of Ophiocordyceps Sobolifera extract for renal injury prevention. Process Biochemistry 2022, 121, 322–329. [Google Scholar]
- Shao, F.; Wang, X.; Wu, H.; Wu, Q.; Zhang, J. Microglia and Neuroinflammation: Crucial Pathological Mechanisms in Traumatic Brain Injury-Induced Neurodegeneration. Front Aging Neurosci 2022, 14, 825086. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Wang, P.; Ge, H.; Qu, X.; Jin, X. Effects of cordycepin on the microglia-overactivation-induced impairments of growth and development of hippocampal cultured neurons. PLoS One 2015, 10(5), e0125902. [Google Scholar] [CrossRef]
- Ding, Y.; Kang, J.; Liu, S.; Xu, Y.; Shao, B. The Protective Effects of Peroxisome Proliferator-Activated Receptor Gamma in Cerebral Ischemia-Reperfusion Injury. Front Neurol 2020, 11, 588516. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Tian, Z.; Wang, M.; Kou, J.; Wang, C.; Rong, X.; Li, J.; Xie, X. , Pang, X. Luteoloside attenuates neuroinflammation in focal cerebral ischemia in rats via regulation of the PPARγ/Nrf2/NF-κB signaling pathway. Int Immunopharmacol 2019, 66, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Sillapachaiyaporn, C.; Chuchawankul, S.; Nilkhet, S.; Moungkote, N.; Sarachana, T.; Ung, A.T.; Baek, S.J.; Tencomnao, T. Ergosterol isolated from cloud ear mushroom (Auricularia polytricha) attenuates bisphenol A-induced BV2 microglial cell inflammation. Food Research International 2022, 157, 111433. [Google Scholar] [CrossRef] [PubMed]
- Nallathamby, N.; Guan-Serm, L.; Vidyadaran, S.; Malek, S.N.A.; Raman, J.; Sabaratnam, V. Ergosterol of Cordyceps militaris Attenuates LPS Induced Inflammation in BV2 Microglia Cells. Nat Prod Commun. 2015, 10(6), 885–6. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Lu, X.; Yang, S.; Zou, Y.; Zeng, F.; Xiong, S.; Cao, Y.; Zhou, W. The anti-inflammatory activity of GABA-enriched Moringa oleifera leaves produced by fermentation with Lactobacillus plantarum LK-1. Front. Nutr. 2023, 10, 1–12. [Google Scholar]
- Zhao, X.R.; Gonzales, N.; Aronowski, J. Pleiotropic role of PPARγ in intracerebral hemorrhage: an intricate system involving Nrf2, RXR, and NF-κB. CNS Neurosci Ther 2015, 21(4), 357–366. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Zhao, T.; Xiao, H. The Implication of Oxidative Stress and AMPK-Nrf2 Antioxidative Signaling in Pneumonia Pathogenesis. Front Endocrinol (Lausanne) 2020, 11, 400. [Google Scholar] [CrossRef] [PubMed]
- Duan, C.; Jiao, D.; Wang, H.; Wu, Q.; Men, W.; Yan, H.; Li, C. Activation of the PPARγ Prevents Ferroptosis-Induced Neuronal Loss in Response to Intracerebral Hemorrhage Through Synergistic Actions With the Nrf2. Front Pharmacol 2022, 13, 869300. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.K.; Kim, Y.M. Beneficial and Detrimental Roles of Heme Oxygenase-1 in the Neurovascular System. Int J Mol Sci 2022, 23, 7041. [Google Scholar] [CrossRef] [PubMed]
- Nitti, M.; Piras, S.; Brondolo, L.; Marinari, U.M.; Pronzato, M.A.; Furfaro, A.L. Heme Oxygenase 1 in the Nervous System: Does It Favor Neuronal Cell Survival or Induce Neurodegeneration? Int J Mol Sci 2018, 19, 2260. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
High-performance liquid chromatography chromatograms and wavelengths of Adenosine, Ergosterol, GABA compounds, and extracts of the mycelial submerged culture of Os H101 (OMSC). Chromatograms (A, C, and E) show standards of adenosine (A), ergosterol (C), GABA (E), and various extracts of OMSC, respectively. A diode array detector (DAD) was used to detect the full-wavelength (200–400 nm) comparison waveforms of standards and different extracts of OMSC (B, D, F). FLOS: fermented liquid of OMSC, WEOS: hot water extract of OMSC, EEOS-50: 50 % ethanol extract of OMSC, EEOS-95: 95 % ethanol extract of OMSC.
Figure 1.
High-performance liquid chromatography chromatograms and wavelengths of Adenosine, Ergosterol, GABA compounds, and extracts of the mycelial submerged culture of Os H101 (OMSC). Chromatograms (A, C, and E) show standards of adenosine (A), ergosterol (C), GABA (E), and various extracts of OMSC, respectively. A diode array detector (DAD) was used to detect the full-wavelength (200–400 nm) comparison waveforms of standards and different extracts of OMSC (B, D, F). FLOS: fermented liquid of OMSC, WEOS: hot water extract of OMSC, EEOS-50: 50 % ethanol extract of OMSC, EEOS-95: 95 % ethanol extract of OMSC.
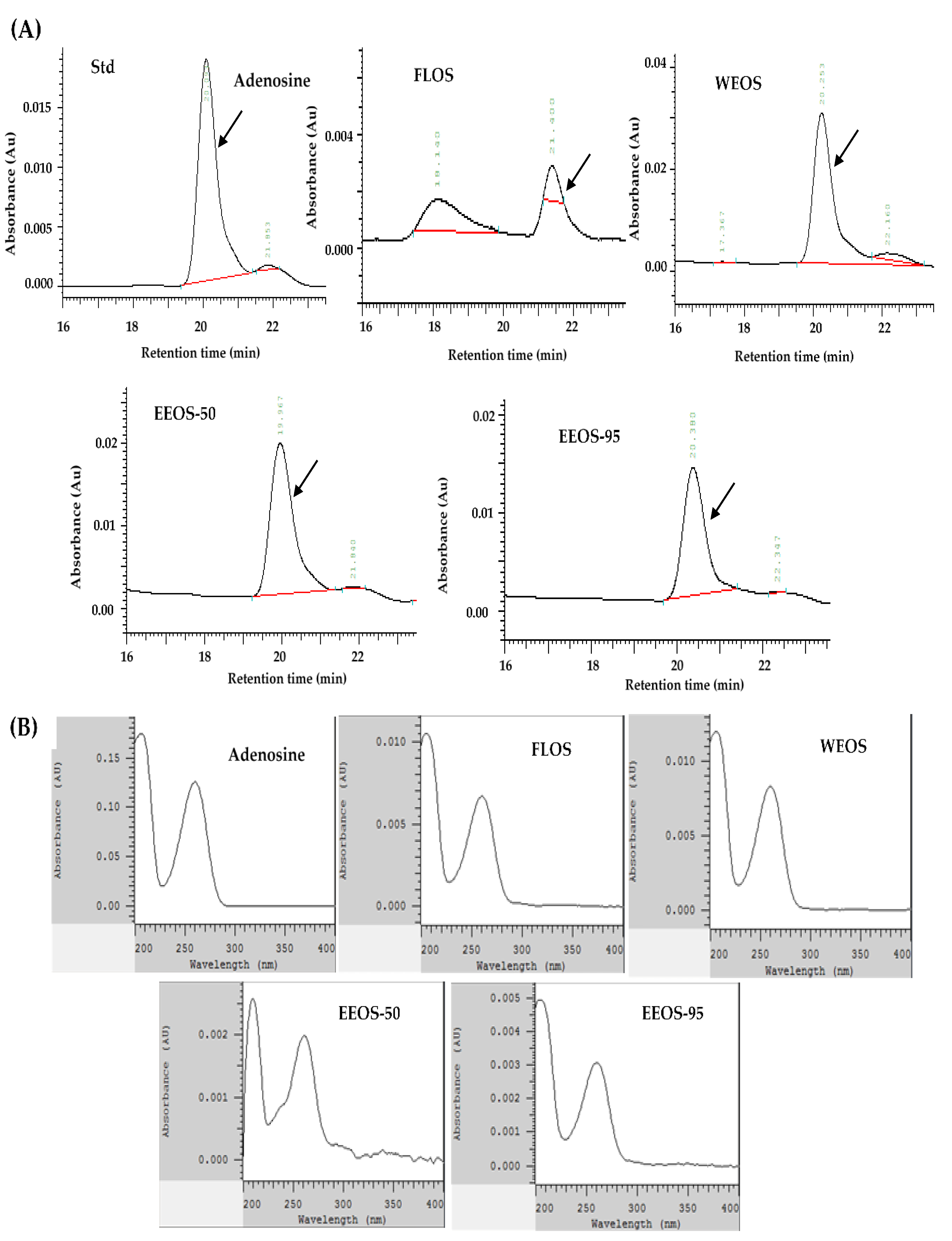
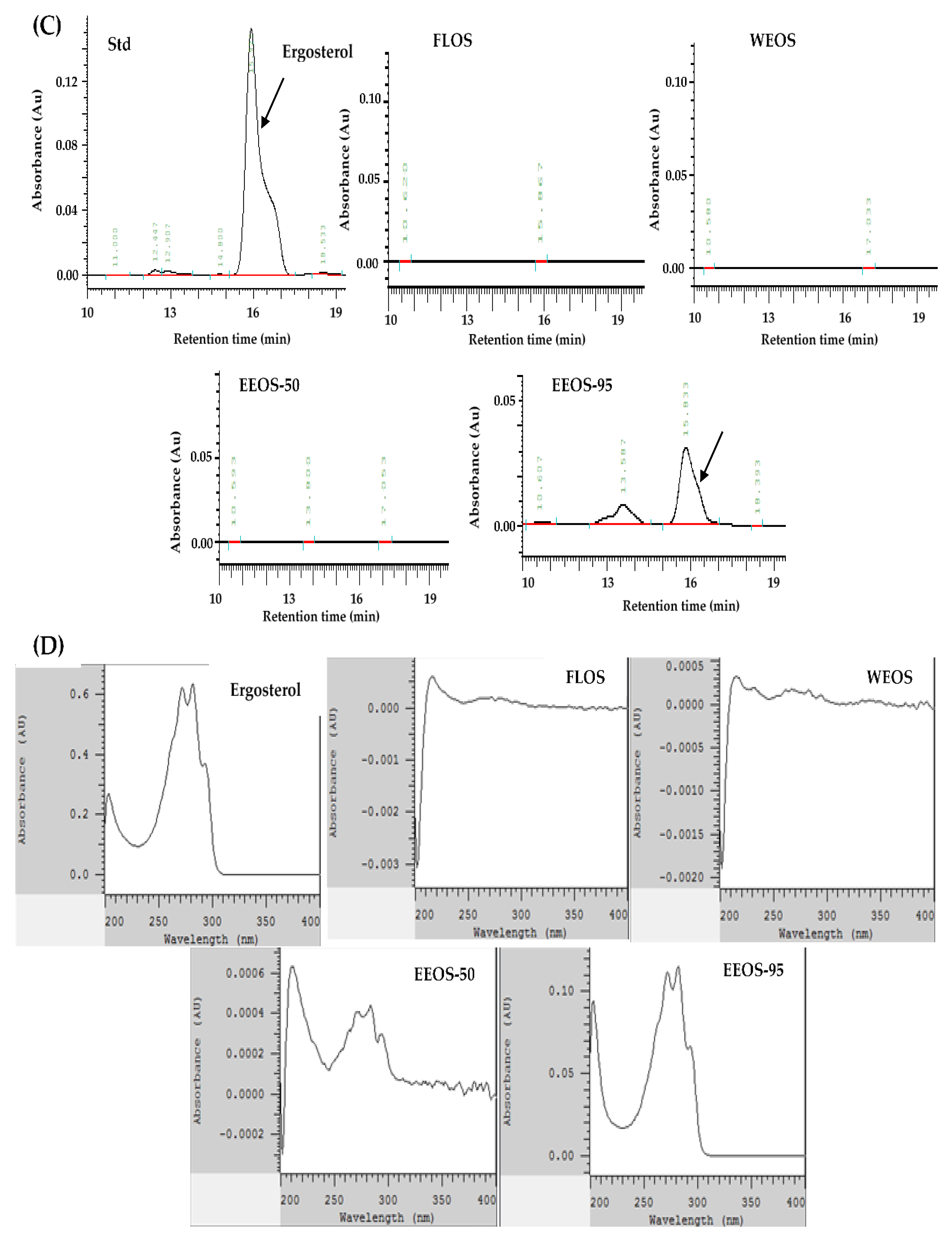
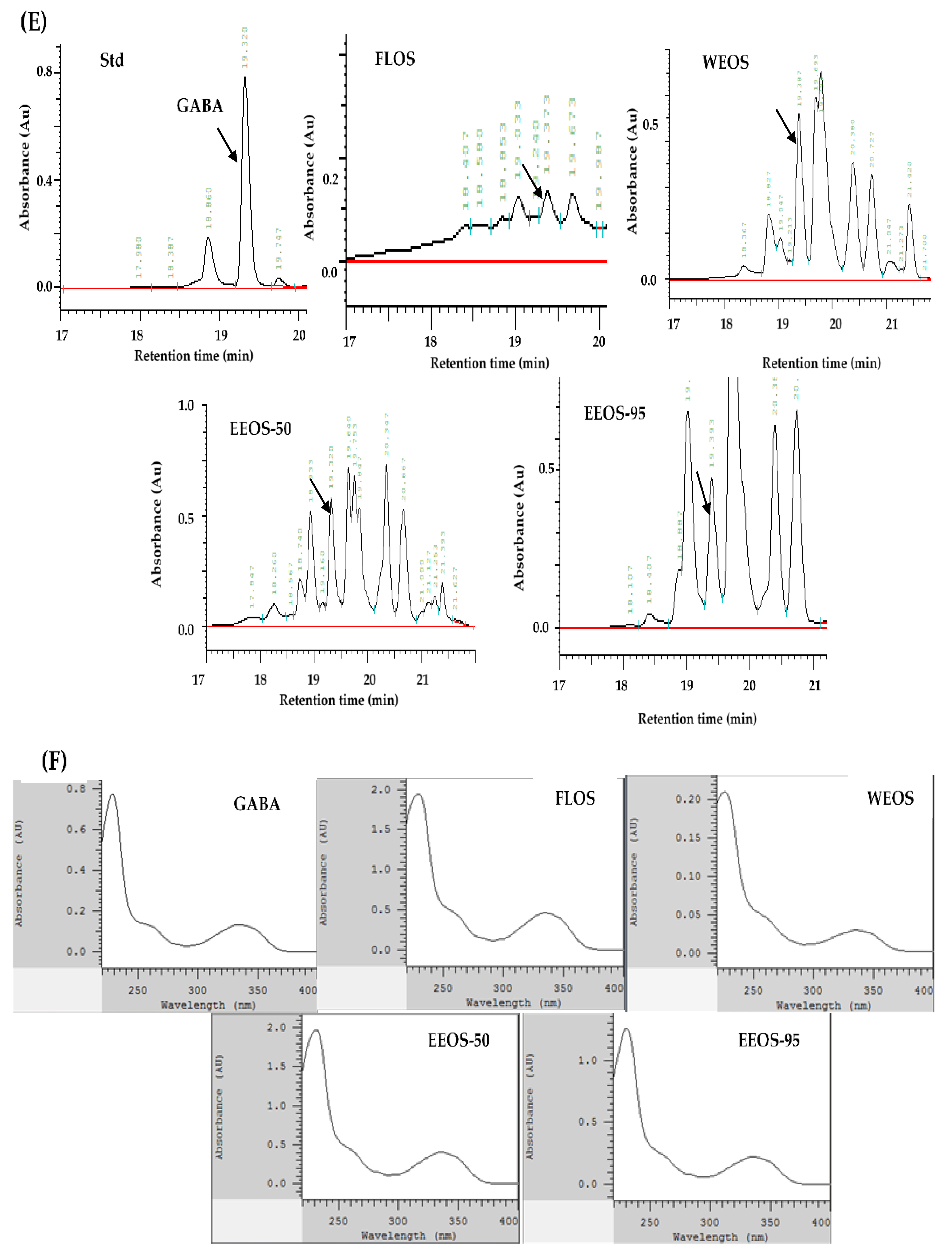
Figure 2.
The inhibitory ratio of EEOS-95 on lipopolysaccharide-induced cytokines production in BV2 microglial cells. Values (means ± SD, n = 3 for the test groups) not sharing the same superscript letter are significantly different (p < 0.05).
Figure 2.
The inhibitory ratio of EEOS-95 on lipopolysaccharide-induced cytokines production in BV2 microglial cells. Values (means ± SD, n = 3 for the test groups) not sharing the same superscript letter are significantly different (p < 0.05).
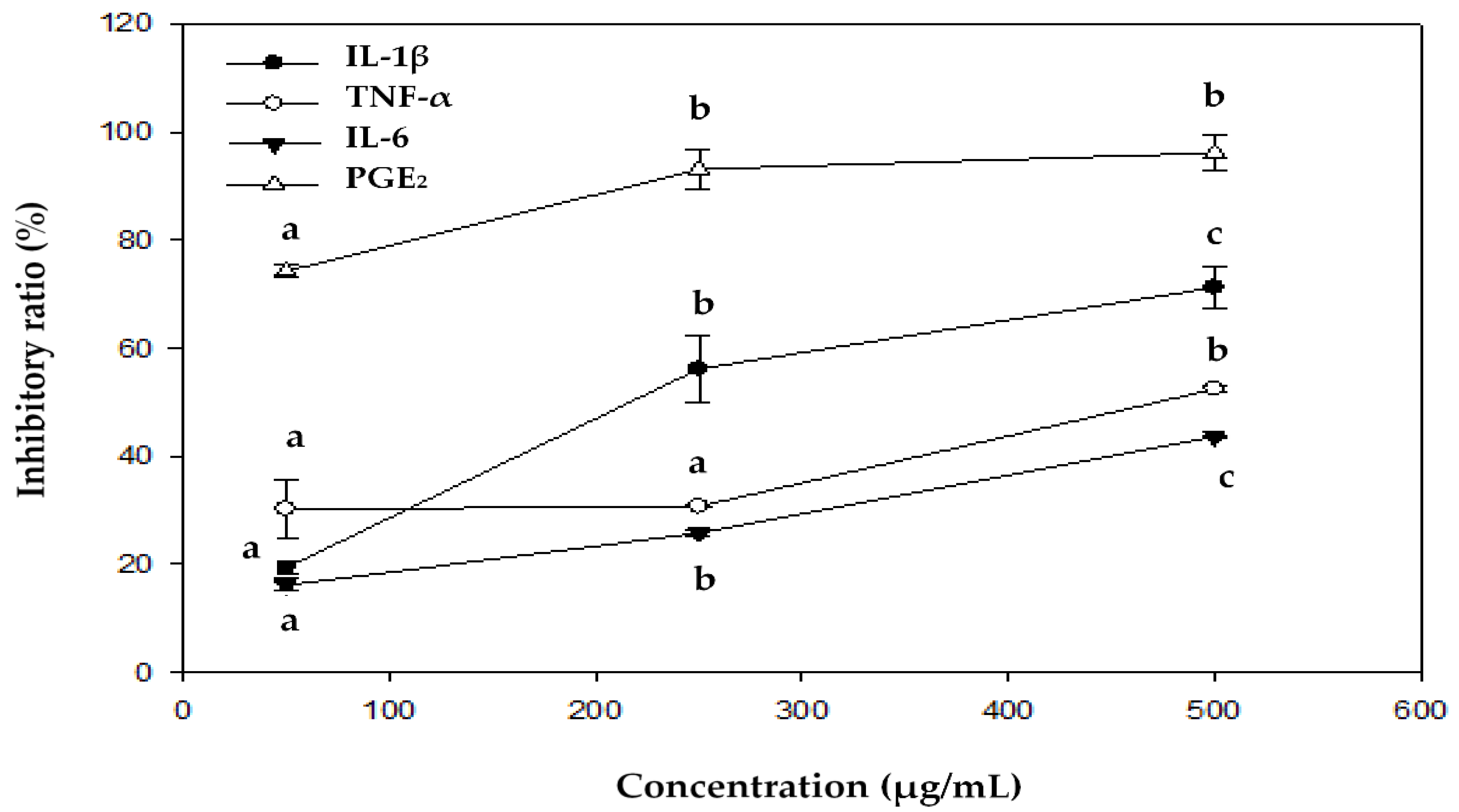
Figure 3.
Effect of EEOS-95 on the expression levels of pro-inflammation factors in lipopolysaccharide-induced BV2 microglial. Cells were treated with EEOS95 (50-500 μg/mL) and MT (1 mM Melatonin) for 24 h and incubated with lipopolysaccharide (LPS, 1 µg/mL) for 24 h. (A) protein expression of inducible nitric oxide synthase (iNOS), cyclooxygenase-2 (COX-2), nuclear factor-κB (NF-ĸB) and β-actin were detected in the cytoplasm by Western blotting assay. The quantitative of (B) iNOS, (C) COX-2, and (D) NF-ĸB were analyzed by ImageJ software. Values (means ± SD, n = 3 for the test groups) not sharing the same superscript letter are significantly different (p < 0.05).
Figure 3.
Effect of EEOS-95 on the expression levels of pro-inflammation factors in lipopolysaccharide-induced BV2 microglial. Cells were treated with EEOS95 (50-500 μg/mL) and MT (1 mM Melatonin) for 24 h and incubated with lipopolysaccharide (LPS, 1 µg/mL) for 24 h. (A) protein expression of inducible nitric oxide synthase (iNOS), cyclooxygenase-2 (COX-2), nuclear factor-κB (NF-ĸB) and β-actin were detected in the cytoplasm by Western blotting assay. The quantitative of (B) iNOS, (C) COX-2, and (D) NF-ĸB were analyzed by ImageJ software. Values (means ± SD, n = 3 for the test groups) not sharing the same superscript letter are significantly different (p < 0.05).
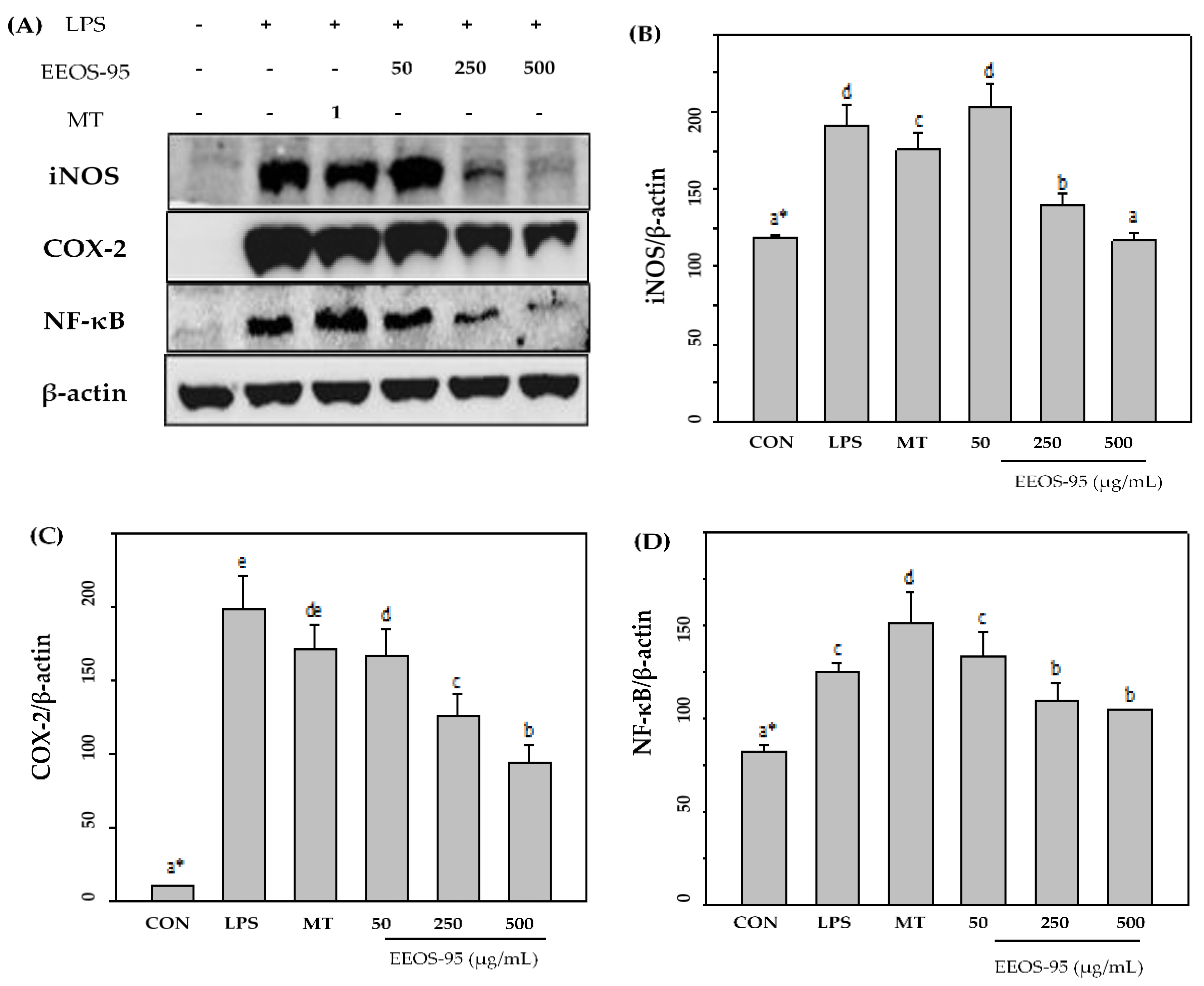
Figure 4.
Effect of EEOS-95 on expression levels of antioxidant factors in lipopolysaccharide-induced BV2 microglial. Cells were treated with EEOS95 (50-500 μg/mL) and MT (Melatonin 1 mM) for 24 h and incubated with lipopolysaccharide (1 µg/mL) for 24 h. (A) protein expression of heme oxygenase-1 (HO-1), nuclear-related factor 2 (Nrf-2), peroxisome proliferator-activated receptors-γ (PPAR-γ), and β-actin were detected in the cytoplasm by Western blotting assay. The quantitative of (B) HO-1, (C) Nrf-2 and (D) PPAR-γ were analyzed by ImageJ software. Values (means ± SD, n = 3 for the test groups) not sharing the same superscript letter are significantly different (p < 0.05).
Figure 4.
Effect of EEOS-95 on expression levels of antioxidant factors in lipopolysaccharide-induced BV2 microglial. Cells were treated with EEOS95 (50-500 μg/mL) and MT (Melatonin 1 mM) for 24 h and incubated with lipopolysaccharide (1 µg/mL) for 24 h. (A) protein expression of heme oxygenase-1 (HO-1), nuclear-related factor 2 (Nrf-2), peroxisome proliferator-activated receptors-γ (PPAR-γ), and β-actin were detected in the cytoplasm by Western blotting assay. The quantitative of (B) HO-1, (C) Nrf-2 and (D) PPAR-γ were analyzed by ImageJ software. Values (means ± SD, n = 3 for the test groups) not sharing the same superscript letter are significantly different (p < 0.05).
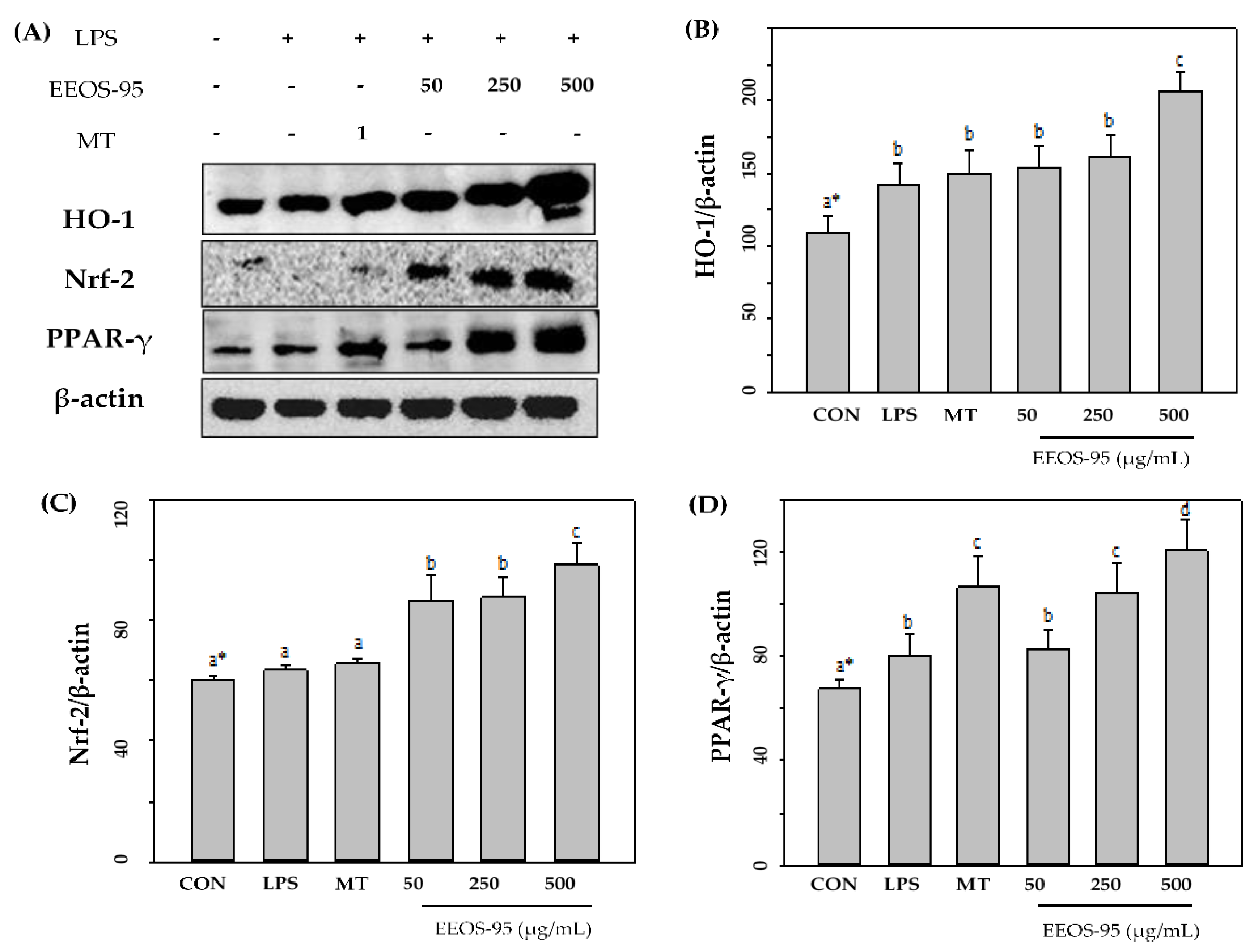
Figure 5.
Schematic summaries of the effects of OMSC and EEOS-95 on lipopolysaccharide-stimulated BV2 microglial cells. FLOS: fermented liquid of OMSC, WEOS: hot water extract of OMSC, EEOS-50: 50 % ethanol extract of OMSC, EEOS-95: 95 % ethanol extract of OMSC, mycelial submerged culture of Os H101 (OMSC).
Figure 5.
Schematic summaries of the effects of OMSC and EEOS-95 on lipopolysaccharide-stimulated BV2 microglial cells. FLOS: fermented liquid of OMSC, WEOS: hot water extract of OMSC, EEOS-50: 50 % ethanol extract of OMSC, EEOS-95: 95 % ethanol extract of OMSC, mycelial submerged culture of Os H101 (OMSC).
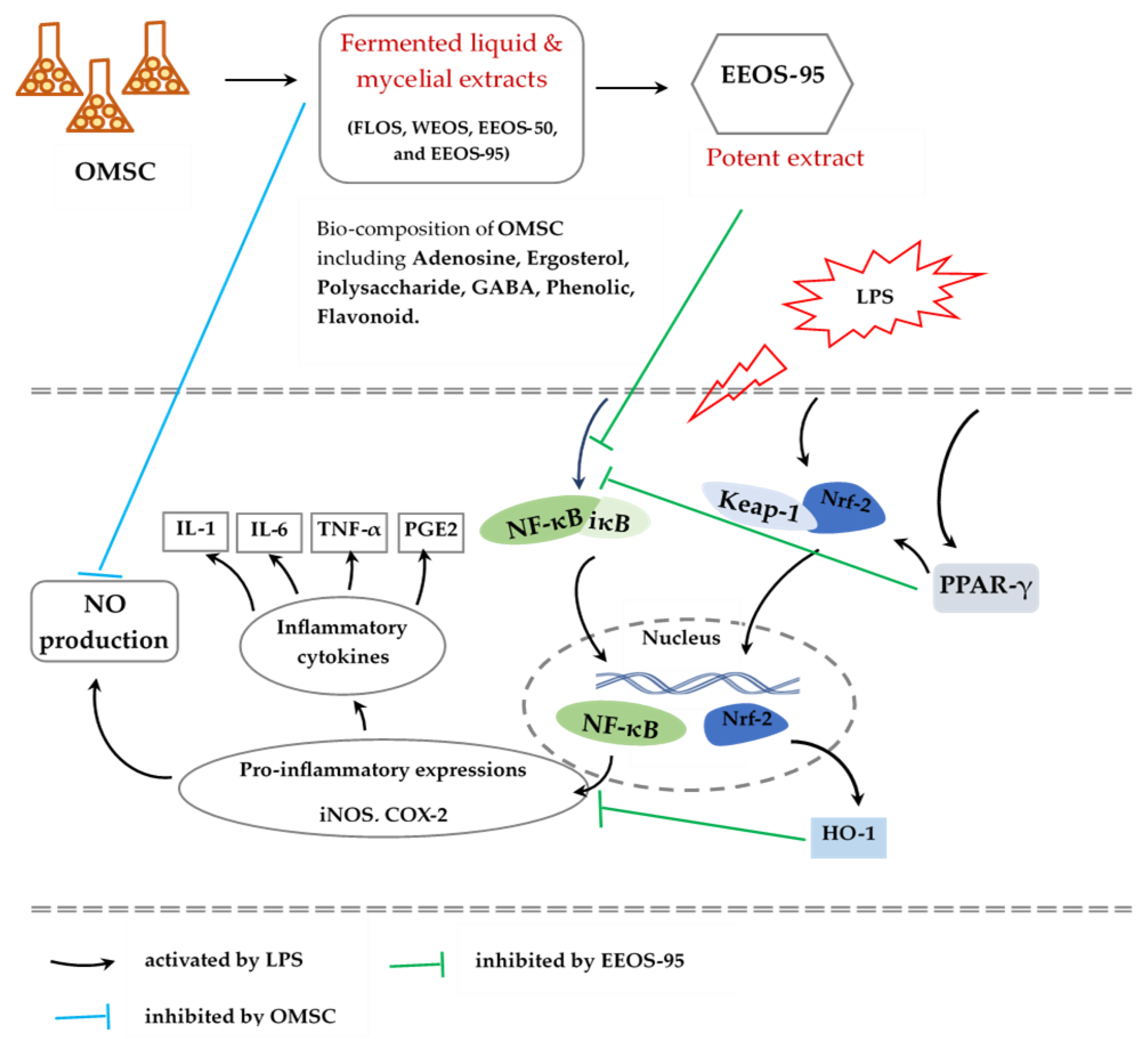

Table 1.
Bioactive ingredients of various extracts from OMSC.
| Compounds (mg/g) | Extracts1 | |||
| FLOS | WEOS | EEOS-50 | EEOS-95 | |
| Adenosine | 2.12 ± 0.09b2 | 2.83 ± 0.05c | 3.29 ± 0.29d | 0.19 ± 0.01a |
| Ergosterol | ND | ND | ND | 18.60 ± 0.70 |
| Polysaccharide | 107.60 ± 10.20b | 156.30 ± 7.10c | 28.50 ± 7.70a | ND |
| Total Polyphenols | 1.57 ± 0.09a | 1.71 ± 0.07bc | 1.77 ± 0.15c | 2.28 ± 0.05d |
| Total Flavonoids | 1.34 ± 0.01b | 1.17 ± 0.02a | 1.65 ± 0.01c | 2.14 ± 0.06d |
| GABA | 3.70 ± 0.30a | 12.60 ± 0.80b | 13.20 ± 0.60b | 18.60 ± 0.50c |
1FLOS: fermented liquid of OMSC, WEOS: hot water extract of OMSC, EEOS-50: 50 % ethanol extract of OMSC, EEOS-95: 95 % ethanol extract of OMSC. ND: Non-detected. 2Values (means ± SD, n = 3 for the test groups) not sharing the same superscript letter in a row are significantly different (p < 0.05).
Table 2.
Effect of various extracts of OMSC on cytotoxicity in BV2 microglial cells.
| Treatments1 | Cell viability (% of CON) | |||
| FLOS2 | WEOS | EEOS-50 | EEOS-95 | |
| 50 | 89.80 ± 13.20a3 | 95.90 ± 7.20a | 94.30 ± 11.90a | 116.60 ± 8.60a |
| 100 | 106.40 ± 25.60a | 96.70 ± 1.40a | 102.20 ± 6.00a | 137.30 ± 12.60b |
| 250 | 119.40 ± 18.30ab | 108.80 ± 5.10a | 116.80 ± 6.20ab | 171.70 ± 12.90cd |
| 500 | 134.20 ± 11.30b | 123.70 ± 11.90b | 125.70 ± 4.40b | 196.50 ± 13.80d |
| 1000 | 114.00 ± 23.40ab | 116.50 ± 8.40ab | 123.00 ± 1.00b | 175.90 ± 3.00c |
1Cells were pretreated with various extracts of OMSC (50-1000 µg/mL) for 24 h. The viability was measured by MTT assay. 2FLOS: fermented liquid of OMSC, WEOS: hot water extract of OMSC, EEOS-50: 50 % ethanol extract of OMSC, EEOS-95: 95 % ethanol extract of OMSC. 3Values (means ± SD, n = 3 for the test groups) not sharing the same superscript letter in a column are significantly different (p < 0.05).
Table 3.
Effect of various extracts of OMSC on lipopolysaccharide-induced nitrite production in BV2 microglial.
Table 3.
Effect of various extracts of OMSC on lipopolysaccharide-induced nitrite production in BV2 microglial.
| Treatments1 | Nitrite concentration (nmol/106 cells) | ||||
| 50 | 250 | 500 | |||
| CON | 0.33 ± 0.05a4 | - | - | - | |
| LPS (1 µg/mL) | 0.78 ± 0.12d | - | - | - | |
| MT (1mM)2 | + LPS | 0.45 ± 0.03b | - | - | - |
| FLOS3 | - | 0.55 ± 0.00c | 0.59 ± 0.07c | 0.59 ± 0.07c | |
| WEOS | - | 0.75 ± 0.02d | 0.74 ± 0.02d | 0.59 ± 0.05c | |
| EEOS-50 | - | 0.76 ± 0.11d | 0.66 ± 0.05cd | 0.58 ± 0.10c | |
| EEOS-95 | - | 0.64 ± 0.16cd | 0.53 ± 0.09c | 0.46 ± 0.02b | |
1Cells were pretreated with 1 mM melatonin (MT) or various extracts of OMSC (µg/mL) for 24 h and then incubated with lipopolysaccharide (LPS, 1 µg/mL) for 24 h. 2Melatonin (MT, N-acetyl-5-methoxytryptamine) is an animal hormone that exhibits physiological functions such as improving sleep, delaying aging, etc… MT is a positive control. 3FLOS: fermented liquid of OMSC, WEOS: hot water extract of OMSC, EEOS-50: 50 % ethanol extract of OMSC, EEOS-95: 95 % ethanol extract of OMSC. 4Values (means ± SD, n = 3 for the test groups) not sharing the same superscript letter in a column are significantly different (p < 0.05).
Table 4.
Effect of EEOS-95 on lipopolysaccharide-induced cytokines in BV2 microglial.
| Treatments1 | Cytokines | ||||
| IL-1β (pg/mL) | TNF-α (ng/mL) | IL-6 (ng/mL) | PGE2 (pg/mL) | ||
| CON | 4.29 ± 1.30a3 | 13.71 ± 0.69a | 0.00 ± 0.00a | 97.60 ± 5.50a | |
| LPS (1 µg/mL) | 76.66 ± 3.47f | 70.54 ± 4.89d | 608.22 ± 17.40e | 933.90 ± 12.30e | |
| MT (1 mM)2 | + LPS | 54.79 ± 0.94d | 35.53 ± 3.58b | 538.87 ± 28.22de | 877.90 ± 10.30d |
| 50 | 62.86 ± 3.90e | 49.58 ± 0.75c | 510.21 ± 7.09cd | 308.20 ± 9.80c | |
| 250 | 36.59 ± 1.54c | 53.10 ± 3.53c | 451.58 ± 2.73c | 155.40 ± 30.40b | |
| 500 | 25.76 ± 0.61b | 42.22 ± 3.22bc | 343.20 ± 1.86b | 126.00 ± 27.90a | |
1Cells were pretreated with 1 mM melatonin or 50-500 (µg/mL) of EEOS-95: 95 % ethanol extracts of OMSC for 24 h and then incubated with lipopolysaccharide (LPS, 1 µg/mL) for 24 h. 2Melatonin (MT) is a positive control. 3Values (means ± SD, n = 3 for the test groups) not sharing the same superscript letter in a column are significantly different (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



