Submitted:
10 April 2024
Posted:
11 April 2024
You are already at the latest version
Abstract
Hypertensive diseases of pregnancy (HDP) represent a global clinical challenge, affecting 5-10% of women and leading to complications for both maternal well-being and fetal development. At the heart of these complications is endothelial dysfunction, with oxidative stress emerging as a pivotal causative factor. The reduction in nitric oxide (NO) bioavailability is a vital indicator of this dysfunction, culminating in blood pressure dysregulation. In the therapeutic context, although antihypertensive medications are commonly used, they come with inherent concerns related to maternal-fetal safety, and a percentage of women do not respond to these therapies. Therefore, alternative strategies that directly address the pathophysiology of HDP are required. This article focuses on the potential of the nitrate-nitrite-NO pathway, abundantly present in dark leafy greens and beetroot, as an alternative approach to treating HDP. The objective of this review is to discuss the prospective antioxidant role of nitrate. We hope our discussion paves the way for using nitrate to improve endothelial dysfunction and control oxidative stress, offering a potential therapy for managing HDP.
Keywords:
Endothelial cell
; pregnancy
; nitric oxide
; nitrate
; oxidative stress
; pre-eclampsia.
Potential Antioxidant Mechanisms of the Nitrate-Nitrite-Nitric Oxide Pathway: Dietary nitrate is converted to nitrite by commensal bacteria. Nitrite is subsequently transformed into nitric oxide (NO). The nitrate surpasses that of nitrite, with nitrite levels exceeding those of NO. The pathway elucidates various antioxidant effects, including blocking NADPH oxidase activity, xanthine oxidase inhibition, mitochondrial production of reactive oxygen species (ROS), sGC activation leading to cGMP production, and promoting Keap1 dissociation. Solid arrows represent established pathways, while dashed arrows indicate potential pathways. CAT: catalase; cGMP: cyclic guanosine monophosphate; GPx: glutathione peroxidase; GTP: guanosine triphosphate; Keap1: Kelch-like ECH-associated protein 1; Nrf2: nuclear factor erythroid 2-related factor 2; NO: nitric oxide; NO3-: nitrate; NO2-: nitrite; ROS: reactive oxygen species sGC: soluble guanylate cyclase; SOD: superoxide dismutase; XOR: xanthine oxidoreductase
1. Introduction
A significant percentage of pregnant women worldwide suffer from hypertension, a common and potentially fatal pregnancy condition. It is estimated that approximately 5% to 10% of all pregnancies are complicated by hypertensive disorders of pregnancy (HDP), making it one of the leading causes of maternal and fetal morbidity and mortality [1,2]. HDP is categorized into four main categories: gestational hypertension, pre-eclampsia/eclampsia, chronic hypertension, and chronic hypertension with pre-eclampsia superimposed. Each has distinct characteristics and implications for the health of the mother and fetus [3].
HDP can harm the placenta and affect the baby's growth, leading to problems such as low birth weight, restricted growth, and premature birth [4,5]. A malfunctioning placenta is also a significant contributor to the development of pre-eclampsia, placing both the mother and the baby at risk [5]. As a consequence, children born from pregnancies complicated by HDP are more prone to conditions like heart disease and metabolic disorders later in life [6]. For mothers, the repercussions included renal and liver damage, thrombosis, and, in severe cases, cerebral hemorrhage [7].
Although HDP is a common and potentially serious issue for both mothers and babies, there are limited therapeutic options. Managing conditions related to HDP typically involves controlling blood pressure and preventing complications [8]. Commonly prescribed antihypertensive drugs include methyldopa, labetalol, and nifedipine [9], and aspirin supplementation may be recommended for specific cases [9]. In the most severe cases, when there is a threat to the life of the mother or the fetus, early delivery may be necessary, even if the baby has not yet reached full gestation maturity.
Managing high blood pressure during pregnancy requires reliable and effective therapeutic intervention. Research into complementary strategies, like dietary interventions, is essential alongside conventional treatments to ensure a safer and healthier pregnancy. Vitamins, minerals, and amino acids have been extensively studied for their potential effects on HDP [10,11,12,13].
Among this exploration, dietary nitrate intake from natural sources shows promising potential. Many studies center on nitrates' ability to elevate nitric oxide (NO) levels via the nitrate-nitrite-NO pathway, reducing blood pressure [14,15,16]. The nitrate-nitrite-NO pathway is recognized for its influence on vascular relaxation, and reviews published to date usually focus on these vasodilatory properties. However, the antioxidant effects of the nitrate-nitrite-NO pathway in the context of HDP still need to be explored. Therefore, this review discusses the potential implications of the nitrate-nitrite-NO pathway on HDP, emphasizing its prospective antioxidant role and underlying mechanisms of the redox system.
Initially, we will discuss the foundational concepts of HDP pathophysiology: endothelial dysfunction and oxidative stress. From there, we will delve into the core of our discussion—the nitrate-nitrite-NO pathway's potential as an antioxidant, exploring possible links with the pathophysiology of HDP. We hope to uncover this pathway's potential therapeutic value in a context beyond vasodilation, providing a deeper understanding of the mechanisms that might involve the modulation of the redox system.
2. Endothelial Dysfunction, Oxidative Stress and Hypertensive Disorders of Pregnancy
The endothelium, a monolayer of endothelial cells lining the inner surface of blood vessels, regulates fluid and electrolyte flow between the bloodstream and tissues, aids vasodilation, and prevents blood clots in a healthy state [17]. Endothelial dysfunction, characterized by its inability to maintain vascular functions, is a hallmark of HDP [18,19]. More than being a consequence, emerging evidence suggests that endothelial dysfunction may act as a primary etiology of HDP [19]. Characteristic manifestations of this dysfunction encompass reduced NO bioavailability, increased inflammatory processes, and enhanced oxidative stress, forming a self-potentiating feedback cycle.
Oxidative stress occurs when there is an imbalance between reactive oxygen species (ROS) production and the body's capacity to neutralize their harmful effects. Oxidative stress is associated with numerous cardiovascular diseases, including HDP [20]. During pregnancy, oxidative stress plays a pivotal role in vascular physiological adaptations due to the increasing oxygen and metabolic demands of the growing fetus [21,22]. At physiological levels, ROS are important in cellular signaling and the modulation of vascular function [21] and contribute to the maturation and functionality of the placenta [23]. However, when levels become excessive during pregnancy, that can impair vascular equilibrium and disrupt endothelial harmony, resulting in the endothelial dysfunction characteristic of HDP.
The endothelium's vulnerability to elevated ROS levels is derived from various metabolic pathways activated in blood vessels during pregnancy, leading to free radical generation. The NADPH oxidase enzyme family is particularly identified as one of the primary sources of superoxide (O2•-) within the endothelium [24], and there is an upregulation in the expression of NADPH oxidase and its subunits in the pre-eclampsia [25,26,27]. As the superoxide production driven by NADPH oxidase increases, this free radical rapidly reacts with vascular NO, forming peroxynitrite (ONOO-), an even more toxic radical [28]. More importantly, this fast reaction between superoxide anion and NO reduces NO bioavailability, feeding a detrimental cycle. Indeed, studies assessing peroxynitrite concentrations in pregnant women with hypertensive disorders have shown that elevated levels of peroxynitrite are related to increased vascular damage [29,30].
The uncoupling endothelial nitric oxide synthase (eNOS) adds complexity to that scenario [31]. Typically, eNOS converts L-arginine into the vasodilator NO. Under stress conditions, however, its activity is redirected towards producing superoxide anion [32]. This shift is accentuated by the upregulation of NADPH oxidase, leading to excessive superoxide anions. These radicals, known for oxidizing tetrahydrobiopterin (BH4) – an indispensable cofactor for eNOS – can contribute to the onset of pre-eclampsia by affecting BH4 levels and thereby disrupting eNOS regulation [33]. A study by Chatre et al. (2022) showed the importance of BH4 to eNOS function, demonstrating that BH4 supplementation might promote eNOS coupling and potentially ameliorate pre-eclampsia symptoms in a mice model [34]. The oxidation of BH4 blocks its contribution to NO synthesis and further drives eNOS towards uncoupling, propelling the system further into a pronounced prooxidative state. In a rat model of pregnancy-induced hypertension, the authors observed increased superoxide generation and BH4 oxidation, indicating the connection between that pathway [35].
Xanthine dehydrogenase (XDH) and xanthine oxidase (XO) are interconvertible forms of an enzyme essential for purine metabolism. Ordinarily, XDH predominates, catalyzing the conversion of hypoxanthine to xanthine and, subsequently, to uric acid, producing NADH in the process [36]. Stressors, such as ischemia, can induce a transition from XDH to XO, which then utilizes molecular oxygen as its electron acceptor, producing superoxide and hydrogen peroxide [37]. Both pre-eclampsia and gestational hypertension have been linked to increased XO activity [38,39].
Lastly, mitochondria, essential to cellular bioenergetics, serve as an established source of ROS, particularly within the placental environment [40]. Acting as the cellular site for aerobic ATP synthesis via oxidative phosphorylation, the importance of mitochondrial function becomes accentuated during pregnancy due to the placenta's higher metabolic demands [40,41]. The increased mitochondrial activity in the placenta directly correlates with a rise in ROS generation [42]. Inefficient electron transfer can facilitate partial oxygen reduction, leading to an excessive generation of superoxide radicals, as evidenced in HDP [43,44].
A robust maternal antioxidant defense system regulates metabolic pathways, reducing the harmful effects of oxidative stress and protecting against endothelial dysfunction [45]. However, this appears not to be the case in pregnant women with HDP. While some studies have reported a decline in maternal antioxidant capacity in HDP, characterized by a decrease in the activity of enzymes such as catalase (CAT), superoxide dismutase (SOD), and glutathione peroxidase (GPx) [46-48], others observed an upregulated antioxidant response [49,50], suggesting a compensatory mechanism to the heightened release of ROS. The indisputable fact is that there is an increase in ROS levels in HDP conditions. Additionally, elevated superoxide anions reduce the bioavailability of NO and have profound implications for endothelium-dependent vasodilation. Beyond these redox system alterations, evidence points to reduced nitrite and nitrate levels in the HDP [51,52]. Contemporary research increasingly postulates that a combination of endothelial dysfunction, oxidative stress, and failure of the NO system coordinate the onset and progression of pregnancy-associated hypertensive disorders.
A fundamental proposition of this review is that the nitrate-nitrite-NO pathway could exhibit antioxidant properties and play a role in modulating oxidative stress. Although this perspective is little explored in the literature, it can have a significant impact on treating HDP.
3. Nitrate-Nitrite-NO Pathway: Antioxidant Dynamics and Therapeutic Potential
NO is traditionally synthesized enzymatically via the nitric oxide synthase (NOS) pathway. However, research breakthroughs alternative routes of NO generation, most notably by reducing nitrate and nitrite [53,54] (Figure 1). The nitrate-nitrite-NO pathway is increasingly recognized for its critical function in the NO dynamic modulation [55]. Because of the volatile nature of NO and the myriad physiological processes that can deplete NO levels in the body, nitrate and nitrite are now perceived as functional reservoirs for stored NO.
An equilibrium in NO is pivotal for maintaining vascular health, but it becomes vulnerable during detrimental conditions like endothelial dysfunction and oxidative stress. Reviews of clinical trials have underscored the role of nitrate in improving endothelial dysfunction and lowering blood pressure [56,57]. HDPs, such as pre-eclampsia, often exhibit reduced NO levels in the circulatory system, exacerbating vascular complications and elevating blood pressure [58,59]. A deficit in NO has been associated with endothelial dysfunction and overall oxidative stress. As a result, therapeutic interventions based on dietary modifications or supplements aimed at restoring NO balance could hold promise in managing HDPs by addressing endothelial dysfunction.
In exploring the nitrate-nitrite-NO pathway, it is essential to recognize the importance of nitrite, even though dietary sources often contain higher concentrations of nitrate. Following consumption, a substantial portion of the ingested nitrate is reduced to nitrite within the body [55]. Therefore, nitrite acts as an essential intermediate in the body's nitrate metabolic pathway. To comprehensively discuss the nitrate-nitrite-NO pathway's antioxidant properties, we must adopt a viewpoint encompassing nitrite-centric findings, understanding the synergistic relationship between nitrate and nitrite in the antioxidant mechanism.
The first study demonstrating a pronounced antioxidant effect resulting from sodium nitrite treatment was conducted in hypertensive animals by Montenegro and colleagues in 2011 [60]. By that time, the emerging role of the novel nitrate-nitrite-NO pathway was taking shape, and the first evidence that this newer path could be used to impact human blood pressure was shown in 2008 by Andrew Webb and his collaborators [61]. In an elegant study, they demonstrated the influence of organic nitrate from a dietary source (beetroot juice) on human blood pressure and the role of the bioconversion of nitrate to nitrite [61]. However, only healthy volunteers were included. Montenegro’s study initially questioned the role of the newer pathway in disease scenarios such as hypertension, where the increased oxidative stress could limit NO bioavailability. They used an experimental model of hypertension 2K1C known for its high activation of the renin-angiotensin system that leads to overexpression of NADPH oxidase, increasing the superoxide production [60]. Surprisingly, in addition to antihypertensive effects induced by nitrite, confirming the potential of the newer pathway even during high oxidative stress conditions induced by hypertension, nitrite treatment dramatically reduced oxidative stress, including in the sham-treated animals [60]. It highlights the dual effect of nitrite anion: A nitrite oxide donor and a potent antioxidant [56]. Further investigation depicts the role of nitrite in inhibiting the vascular NADPH oxidase, and these experimental findings have been confirmed in several subsequent studies [62,63,64].
Experimental studies offer insights into the potential mechanisms of nitrate and nitrite in controlling oxidative stress [64,65,66]. Nitrate has been shown to reduce ROS production, decreasing oxidative markers tied to lipid oxidation, such as TBARS adducts and 4-hydroxynonenal (4-HNE) [67,68,69]. Moreover, short-term nitrite therapy has shown promise in reversing age-related vascular endothelial dysfunction and reducing oxidative stress. This is evident from the restoration of endothelium-dependent dilation and significant mitigation of arterial superoxide production and inflammation in aged male C57BL6J mice [70].
Beyond the effects on lipid oxidation, nitrate and nitrite have exhibited protective mechanisms against protein oxidations – a crucial aspect of oxidative stress. Such modifications can lead to irreversible changes to protein side chains, often compromising cellular function by affecting protein stability and enzymatic activity [71]. In clinical and experimental studies, nitrate and nitrite supplementation has demonstrated a dual benefit: it improves endothelial function and results in a notable decrease in nitrotyrosine levels – a marker of protein oxidation [72,73,74].
In this regard, the potential therapeutic benefits of nitrate for HDP appear particularly promising. The ability of nitrate-nitrite-NO to mitigate oxidative stress markers suggests its potential to reduce such markers and alleviate endothelial dysfunction, placing nitrate supplementation as a viable path for HDP management.
Elevated markers of lipid and protein oxidation in pregnant women with pre-eclampsia and gestational hypertension are central contributors to the endothelial dysfunction characteristic of HDP. Studies carried out in women with pre-eclampsia show a pronounced elevation of lipid peroxidation markers [75,76,77,78,79]. Similarly, plasma from HDP-afflicted women consistently reveals heightened levels of protein oxidation, contrasting with samples from women experiencing normotensive pregnancies [80]. Although DNA oxidation markers are less frequently examined, they are also altered in women with HDP [81,82].
The exact mechanism by which nitrate-nitrite-NO exerts these protective roles remains under investigation. Nonetheless, based on existing studies, it seems these compounds regulate the production of the superoxide radical, targeting its primary sources, namely NADPH oxidase, XOR, and mitochondria. Additionally, considering nitrate and nitrite as reservoirs for NO, it is essential to emphasize their potential to enhance NO levels or their bioavailability.
Research across various animal models has consistently shown that the potential mechanism behind nitrate's action and its intermediary nitrite might reside in the inhibition of NADPH oxidase activity [60,64,65]. Although direct neutralization of free radicals by these compounds has been suggested, the potential therapeutic role of the nitrate-nitrite-NO pathway seems to be derived from blocking NADPH oxidase activity and the synthesis of O2•- [60,62]. These results align with in vitro studies; in macrophages activated with LPS, the NO derived from nitrite reduced the NOX-dependent superoxide production, further emphasizing the role of nitrite-NO in this mechanism [83]. Such results become especially relevant considering the elevated NADPH oxidase activity observed in HDP contexts [84].
Hypotheses about the underlying mechanisms by which the nitrate-nitrite-NO pathway reduces NADPH oxidase activity and superoxide production have gained prominence in the scientific community. A central hypothesis delves into the role of NO in facilitating protein modifications via S-nitrosylation. S-nitrosylation represents a post-translational protein modification in which a NO group is covalently attached to a cysteine residue in a protein, and this process plays a pivotal role in how NO regulates various proteins [85]. Intriguingly, S-nitrosylation is a crucial mechanism by which NO might govern NADPH oxidase activity. In a cellular model, Qian et al. (2012) uncovered that NO could facilitate the reversible post-translational modification of Nox5, leading to reduced ROS production [86].
Beyond the influence of nitrate and nitrite on NADPH oxidase, we cannot dismiss the potential mechanisms for controlling oxidative stress via XOR. Amaral et al. (2015) utilized the deoxycorticosterone-salt (DOCA-salt) hypertensive model and found that sodium nitrite supplementation displayed antioxidant properties [62]. In a separate study centered on the activated macrophage, nitrite supplementation appeared to suppress NADPH oxidase activity, although not through the previously hypothesized S-nitrosation mechanism [87]. The authors suggest that this result might be attributed to changes in Nox2 and XOR [87]. The decrease in XOR activity emphasizes the evolving understanding of the effects of the nitrate-nitrite-NO pathway.
The results of nitrate as an antioxidant are also promising regarding mitochondria control. A study involving rats assessed dietary nitrate supplementation's ability to mitigate oxidative stress in a hypoxia model. In this research, nitrate provided protection against the reduction in complex I activity and alleviated oxidative stress, thereby boosting NO bioavailability within the myocardium [88].
Interestingly, even though nitrate is bioactivated to nitrite, which exerts antioxidant effects, nitrite and nitrate may also regulate eNOS activity and affect endogenous NO formation [89]. Nitrate supplementation dose-dependently reduced vascular eNOS Ser1177 phosphorylation and increased Thr495 phosphorylation, thus decreasing global eNOS activity, with corresponding attenuation of vascular responses [89]. Similar alterations were observed when nitrite was directly added to vascular preparations, indicating a direct inhibitory effect of nitrite on the endogenous NO formation [89]. In contrast, there is evidence that nitrite may directly promote eNOS-derived NO formation in hypertensive disorders. Nitrite administration improved endothelium-dependent relaxation in spontaneously hypertensive rats by a mechanism involving eNOS activation to produce NO [90]. These studies lead to contrasting conclusions. However, both nitrate and nitrite may exert opposite vascular effects when we compare healthy [89] and hypertensive [90] vessels, with significant differences concerning redox conditions, so nitrite may exert beneficial effects in disease conditions. Adding more complexity to nitrite/nitrate biology, it is also possible that nitrate may inhibit some of the responses found with nitrite, as previously shown [91]. XOR bioactivates both nitrate and nitrite, and they compete for the catalytic site of the enzyme. These effects may translate nitrate protection against excess nitrite bioactivation to NO by XOR [91].
The body's defense against oxidative stress is complex, and nitrate emerges as a potential contributor to enhancing antioxidant capacities. A study by Cui et al. (2019), utilizing a model of ischemia-reperfusion injury-induced post-skin flap in mice, showed that nitrate restored the enzymatic activity of SOD, GPx, and CAT [92]. Extending these findings to humans, research has explored nitrate's impact on the enzymatic defense system, especially in exercise, given NO's vasodilatory role [93]. Another study involving patients with metabolic syndrome noted that acute exercise, combined with the intake of nitrate-rich juice, amplified the expression of MnSOD, GPx, and CAT [94].
In HDP conditions, increased oxidative stress can overload antioxidant enzymes, causing the antioxidant system to operate at or beyond its capacity to neutralize the excessive free radicals, leading to an apparent decline in enzyme activity. Women diagnosed with pre-eclampsia typically display lowered plasma concentrations of enzymes like GPx and SOD, a fact that is observed at the mRNA level for these enzymes [95]. Furthermore, CAT activity appears to be suboptimal in women with gestational hypertension than in those with normotensive pregnancies [96]. Building on this, a study by Qiu et al. (2021) highlighted reduced activities of CAT, SOD, and GPx in preeclamptic placental tissues compared to their regular counterparts [97]. While many studies report a decline in antioxidant activity, findings indicate increased enzyme activity as the body attempts to restore normal homeostasis. That idea is supported by the work of Caldeiras-Dias et al. (2021), where women with pre-eclampsia demonstrated heightened activity in the antioxidant response element (ARE) compared to normal pregnancies [98]. Intriguingly, resveratrol, a well-known antioxidant, enhanced the ARE activity [98]. This underscores that, regardless of whether there is a decline or increase in activity, adding compounds with antioxidant potential can bolster antioxidant capabilities.
Delving into the molecular complexity of the antioxidant defense, the Nrf2 pathway needs attention. The ARE region is a direct downstream target of Nrf2, encompassing various enzymes essential for antioxidant and detoxification processes [99]. Nrf2 is pivotal for protecting against oxidative stress, orchestrating the coordinated expression of various antioxidant genes [99]. Under basal conditions, Nrf2 is anchored in the cytoplasm, bound closely to the Kelch-like ECH-associated protein 1 (Keap1) [99]. In oxidative stress, Nrf2 detaches from Keap1, allowing it to translocate to the nucleus, initiating its protective transcriptional program. Given the conceptual role of Nrf2, one might expect its pathway to be increased in conditions like HDP. However, studies indicate that the oxidative stress signaling pathway, mediated by Nrf2-Keap1, is compromised in the placentas of women with pre-eclampsia [100-102]. Studies in animals have shown the potential of nitrites to influence Nrf2 [73,103]. Fascinatingly, Amaral et al. (2019), utilizing a two-kidney one-clip hypertensive rat model, highlighted nitrite's capability – as an NO donor – to modulate the transcription factor Nrf2 [103]. Mechanistic investigations suggest that NO might enact its effect-inducing modifications in Keap1, subsequently releasing Nrf2 or through the NO-cGMP pathway, bolstering the notion of NO's central role in orchestrating the antioxidant response.
In light of evidence highlighting the antioxidant abilities of the nitrate-nitrite-NO pathway and the potential influence on transcription factors like Nrf2, it seems pertinent to explore the therapeutic potential of these compounds in the context of HDP. The idea of supplementing with nitrate or nitrite to enhance antioxidant defenses in HDP warrants thorough investigation. Such an approach might not only mitigate oxidative stress. Still, it could also offer a new path in managing HDP by addressing the core oxidative perturbations intrinsic to these conditions. Achieving control over oxidative stress provides a dual advantage: 1) augments the bioavailability of NO and 2) prevents the combination of superoxide with NO, which would lead to the production of the potent oxidant peroxynitrite. In the HDP context, enhancing vasodilation through NO, especially when considering nitrate and nitrite supplementation, is pivotal for blood pressure control and improving endothelial dysfunction. This idea is supported by an intriguing study on a pregnant eNOS−/− mice model, where supplementation with beetroot juice – a known rich nitrate source– reduced blood pressure and improved endothelial function [104]. This outcome is in line with the inhibitory role of nitrite on NADPH oxidase, which reduces oxidative stress, increasing thus NO availability.
Drawing upon dietary sources for nitrate consumption, rather than synthetic derivatives, presents both safety and bioavailability benefits. Foods naturally rich in nitrates, like beetroot and green vegetables, may confer a synergistic health advantage, given their many nutritive constituents. Although the literature has explored the antihypertensive facets of nitrate-rich foods, especially in the HDP context [105,106], there still needs to be more in-depth research on the antioxidant potential of dietary nitrate in this specific cohort.
Endothelial dysfunction and oxidative stress are unmistakable hallmarks of HDP. However, most human studies concerning nitrate or dietary sources thereof are predominantly concerned with their capacity to modulate blood pressure. We intend to maximize the importance of blood pressure control in HDP conditions. Instead, we want to discuss its role beyond the vasodilator action. The outcomes of animal studies addressed in this review reinforce the antioxidant potential of the nitrate-nitrite-NO pathway, as summarized in Table 1.
A topic warranting brief exploration is the association of nitrate and nitrite with the formation of nitrosamines – compounds linked to cancer [107]. Often, nitrate and nitrite are utilized as preservatives in processed foods, including sausages, ham, and specific meat types [108]. Consuming high quantities of meat and processed foods has been associated with increased cancer risk [109]. On the other hand, the Mediterranean diet, lauded for its numerous health benefits, is rich in nitrates from leafy green vegetables and beetroot [110]. Beyond nitrate content, these dietary patterns also have high amounts of antioxidants, fiber, and other beneficial compounds that contribute to a balanced redox environment.
It is necessary to consider the intrinsic nature of nitrate versus its source and intake conditions. The nitrate debate calls for caution, considering the importance of dietary and physiological context. An illustrative example comes from a clinical study on using isolated compounds: while antioxidants like vitamin C and E might be beneficial when consumed through natural sources like fruits – due to their interactions with other compounds – their isolated forms did not yield positive results in pre-eclampsia patients [111]. This discrepancy underscores the complexity of bioactive compounds and their interactions. The mechanisms of action of these antioxidants, including potential synergies with other compounds in whole foods, might play a pivotal role in their beneficial effects. Therefore, rather than viewing nitrate as a singular antagonist, it is more judicious to analyze the contexts in which it is introduced and the potential synergistic or antagonistic outcomes that may arise.
4. Perspectives and Conclusions
In HDP, the vascular endothelium's role in ensuring vasodilator, anti-inflammatory, and anti-thrombotic functions becomes severely compromised. A healthy endothelium produces NO in a well-regulated manner. However, this synthesis is hampered in hypertensive conditions. With the increased oxidative stress and endothelial dysfunction that characterize HDP, the health and well-being of both mother and fetus are jeopardized. The potential of nitrate, mainly from natural food sources, to reduce oxidative stress, improve endothelial function, and possibly lower blood pressure through the nitrate-nitrite-NO pathway presents a promising avenue for exploration in HDP.
The potential of the nitrate-nitrite-NO pathway holds considerable promise regarding safety and bioavailability. Natural nitrate-rich foods, such as beetroot and leafy greens, provide a matrix of nutrients and compounds that may improve dietary nitrate uptake and metabolic conversion to bioactive forms. Aside from the intrinsic safety of dietary consumption, there is the added benefit that certain foods may provide synergistic health benefits due to the presence of other nutritional constituents. While preliminary evidence is promising [105,106], more comprehensive, well-designed clinical trials are required before recommending nitrate-rich diets or supplements as a complementary treatment in HDP. The physiological alterations during pregnancy may affect the impact of nitrate and nitrite. Dosage, administration, frequency, and the overall nutritional environment are pivotal factors determining efficacy and safety. Additionally, the long-term effects on fetal development and maternal health post-partum demand extensive research.
Finally, caution notes regarding nitrate or nitrite use during pregnancy should be considered. To our knowledge, no information is available concerning the amounts of nitrate and nitrite that may cross the placental barrier and interact with the biology of the fetus. Nitrate from dietary sources leads to high plasma concentration (90 – 152 μM), and nitrite is finely controlled and is kept at lower levels [112]. No previous study has simultaneously assessed nitrite and nitrate concentrations in the mother and the newborn. However, it is known that fetal hemoglobin is more easily oxidized to form methemoglobin by nitrite than adult hemoglobin [113]. Methemoglobin is generated when the ferrous component (Fe2+) of hemoglobin is oxidized by oxidants, rendering it incapable of binding to the O2 [114]. Moreover, there is evidence that NADH-methemoglobin reductase activity, which reconverts methemoglobin back to hemoglobin, is lower in infants than in adults [115], which could be looked at with caution. Whether maternal nitrate therapy increases newborns' methemoglobin concentrations remains to be determined, and further studies are needed.
In conclusion, this review exploits a novel perspective of the nitrate-nitrite NO pathway's potential as an antioxidant agent in the context of HDP - a topic previously underexplored. Our work shines a light on future investigations against the context of limited research on the interplay between nitrate supplementation and its interplay with controlling oxidative stress, endothelial dysfunction, and HDP.
Author Contributions
Conceptualization, P.O.B., and V.C.S.; Writing – Original Draft Preparation, P.O.B., J.E.T.S., and V.C.S.; Writing and critical review & Editing, P.O.B., R.C.C., V.C.S., T.B., and M.F.M.; Supervision, R.C.C., and V.C.S.; Funding Acquisition, V.C.S. All authors have read and agreed to submit the current version of the manuscript for publication.
Funding
This research was supported by the São Paulo Research Foundation, Brazil (FAPESP) grant numbers 21/12010-7 and 23/07589-1.
Acknowledgments
The authors thank Dr Priscila Rezeck Nunes for reading this manuscript and commenting critically. All figures were generated using BioRender.com.
References
- Gestational Hypertension and Preeclampsia: ACOG Practice Bulletin, Number 222. Obstet Gynecol 2020, 135, e237–e260. [CrossRef] [PubMed]
- Regitz-Zagrosek, V.; Roos-Hesselink, J.W.; Bauersachs, J.; Blomstrom-Lundqvist, C.; Cifkova, R.; De Bonis, M.; Iung, B.; Johnson, M.R.; Kintscher, U.; Kranke, P. , et al. 2018 ESC Guidelines for the management of cardiovascular diseases during pregnancy. Eur Heart J 2018, 39, 3165–3241. [Google Scholar] [CrossRef] [PubMed]
- Coggins, N.; Lai, S. Hypertensive Disorders of Pregnancy. Emerg Med Clin North Am 2023, 41, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, N.; An, H.; Li, Z.; Zhang, L.; Li, H.; Zhang, Y.; Ye, R. Impact of gestational hypertension and preeclampsia on low birthweight and small-for-gestational-age infants in China: A large prospective cohort study. J Clin Hypertens (Greenwich) 2021, 23, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Di Martino, D.D.; Avagliano, L.; Ferrazzi, E.; Fuse, F.; Sterpi, V.; Parasiliti, M.; Stampalija, T.; Zullino, S.; Farina, A.; Bulfamante, G.P. , et al. Hypertensive Disorders of Pregnancy and Fetal Growth Restriction: Clinical Characteristics and Placental Lesions and Possible Preventive Nutritional Targets. Nutrients 2022, 14. [Google Scholar] [CrossRef]
- Crispi, F.; Miranda, J.; Gratacos, E. Long-term cardiovascular consequences of fetal growth restriction: biology, clinical implications, and opportunities for prevention of adult disease. Am J Obstet Gynecol 2018, 218, S869–S879. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, M.W., Jr.; LaMarca, B. Risk of cardiovascular disease, end-stage renal disease, and stroke in postpartum women and their fetuses after a hypertensive pregnancy. Am J Physiol Regul Integr Comp Physiol 2018, 315, R521–R528. [Google Scholar] [CrossRef] [PubMed]
- Metoki, H.; Iwama, N.; Hamada, H.; Satoh, M.; Murakami, T.; Ishikuro, M.; Obara, T. Hypertensive disorders of pregnancy: definition, management, and out-of-office blood pressure measurement. Hypertens Res 2022, 45, 1298–1309. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.M.; Garovic, V.D. Drug treatment of hypertension in pregnancy. Drugs 2014, 74, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Xiaomang, J.; Yanling, W. Effect of vitamin D3 supplementation during pregnancy on high risk factors - a randomized controlled trial. J Perinat Med 2021, 49, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Camarena Pulido, E.E.; Garcia Benavides, L.; Panduro Baron, J.G.; Pascoe Gonzalez, S.; Madrigal Saray, A.J.; Garcia Padilla, F.E.; Totsuka Sutto, S.E. Efficacy of L-arginine for preventing preeclampsia in high-risk pregnancies: A double-blind, randomized, clinical trial. Hypertens Pregnancy 2016, 35, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Monari, F.; Menichini, D.; Pignatti, L.; Basile, L.; Facchinetti, F.; Neri, I. Effect of L-arginine supplementation in pregnant women with chronic hypertension and previous placenta vascular disorders receiving Aspirin prophylaxis: a randomized control trial. Minerva Obstet Gynecol 2021, 73, 782–789. [Google Scholar] [CrossRef]
- Mesdaghinia, E.; Shahin, F.; Ghaderi, A.; Shahin, D.; Shariat, M.; Banafshe, H. The Effect of Selenium Supplementation on Clinical Outcomes, Metabolic Profiles, and Pulsatility Index of the Uterine Artery in High-Risk Mothers in Terms of Preeclampsia Screening with Quadruple Test: a Randomized, Double-Blind, Placebo-Controlled Clinical Trial : Selenium and preeclampsia. Biol Trace Elem Res 2023, 201, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.H.; Bondonno, C.P.; Croft, K.D.; Puddey, I.B.; Woodman, R.J.; Rich, L.; Ward, N.C.; Vita, J.A.; Hodgson, J.M. Effects of a nitrate-rich meal on arterial stiffness and blood pressure in healthy volunteers. Nitric Oxide 2013, 35, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Jovanovski, E.; Bosco, L.; Khan, K.; Au-Yeung, F.; Ho, H.; Zurbau, A.; Jenkins, A.L.; Vuksan, V. Effect of Spinach, a High Dietary Nitrate Source, on Arterial Stiffness and Related Hemodynamic Measures: A Randomized, Controlled Trial in Healthy Adults. Clin Nutr Res 2015, 4, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Stanaway, L.; Rutherfurd-Markwick, K.; Page, R.; Wong, M.; Jirangrat, W.; Teh, K.H.; Ali, A. Acute Supplementation with Nitrate-Rich Beetroot Juice Causes a Greater Increase in Plasma Nitrite and Reduction in Blood Pressure of Older Compared to Younger Adults. Nutrients 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Aird, W.C. Endothelium as an organ system. Crit Care Med 2004, 32, S271–279. [Google Scholar] [CrossRef] [PubMed]
- Endemann, D.H.; Schiffrin, E.L. Endothelial dysfunction. J Am Soc Nephrol 2004, 15, 1983–1992. [Google Scholar] [CrossRef] [PubMed]
- Possomato-Vieira, J.S.; Khalil, R.A. Mechanisms of Endothelial Dysfunction in Hypertensive Pregnancy and Preeclampsia. Adv Pharmacol 2016, 77, 361–431. [Google Scholar] [CrossRef]
- Sanchez-Aranguren, L.C.; Prada, C.E.; Riano-Medina, C.E.; Lopez, M. Endothelial dysfunction and preeclampsia: role of oxidative stress. Front Physiol 2014, 5, 372. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.D.; De Long, N.E.; Wang, R.C.; Yazdi, F.T.; Holloway, A.C.; Raha, S. Angiogenesis in the placenta: the role of reactive oxygen species signaling. Biomed Res Int 2015, 2015, 814543. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Murtaza, G.; Metwally, E.; Kalhoro, D.H.; Kalhoro, M.S.; Rahu, B.A.; Sahito, R.G.A.; Yin, Y.; Yang, H.; Chughtai, M.I. , et al. The Role of Oxidative Stress and Antioxidant Balance in Pregnancy. Mediators Inflamm 2021, 2021, 9962860. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Jin, H.; Qiu, Y.; Liu, Y.; Wen, L.; Fu, Y.; Qi, H.; Baker, P.N.; Tong, C. Reactive Oxygen Species are Essential for Placental Angiogenesis During Early Gestation. Oxid Med Cell Longev 2022, 2022, 4290922. [Google Scholar] [CrossRef]
- Aouache, R.; Biquard, L.; Vaiman, D.; Miralles, F. Oxidative Stress in Preeclampsia and Placental Diseases. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Dechend, R.; Viedt, C.; Muller, D.N.; Ugele, B.; Brandes, R.P.; Wallukat, G.; Park, J.K.; Janke, J.; Barta, P.; Theuer, J. , et al. AT1 receptor agonistic antibodies from preeclamptic patients stimulate NADPH oxidase. Circulation 2003, 107, 1632–1639. [Google Scholar] [CrossRef] [PubMed]
- Raijmakers, M.T.; Dechend, R.; Poston, L. Oxidative stress and preeclampsia: rationale for antioxidant clinical trials. Hypertension 2004, 44, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.L.; Brockman, D.; Campos, B.; Myatt, L. Expression of NADPH oxidase isoform 1 (Nox1) in human placenta: involvement in preeclampsia. Placenta 2006, 27, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Schulz, E.; Gori, T.; Munzel, T. Oxidative stress and endothelial dysfunction in hypertension. Hypertens Res 2011, 34, 665–673. [Google Scholar] [CrossRef]
- Myatt, L.; Rosenfield, R.B.; Eis, A.L.; Brockman, D.E.; Greer, I.; Lyall, F. Nitrotyrosine residues in placenta. Evidence of peroxynitrite formation and action. Hypertension 1996, 28, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Roggensack, A.M.; Zhang, Y.; Davidge, S.T. Evidence for peroxynitrite formation in the vasculature of women with preeclampsia. Hypertension 1999, 33, 83–89. [Google Scholar] [CrossRef]
- Janaszak-Jasiecka, A.; Ploska, A.; Wieronska, J.M.; Dobrucki, L.W.; Kalinowski, L. Endothelial dysfunction due to eNOS uncoupling: molecular mechanisms as potential therapeutic targets. Cell Mol Biol Lett 2023, 28, 21. [Google Scholar] [CrossRef] [PubMed]
- Phoswa, W.N.; Khaliq, O.P. The Role of Oxidative Stress in Hypertensive Disorders of Pregnancy (Preeclampsia, Gestational Hypertension) and Metabolic Disorder of Pregnancy (Gestational Diabetes Mellitus). Oxid Med Cell Longev 2021, 2021, 5581570. [Google Scholar] [CrossRef] [PubMed]
- Kukor, Z.; Valent, S.; Toth, M. Regulation of nitric oxide synthase activity by tetrahydrobiopterin in human placentae from normal and pre-eclamptic pregnancies. Placenta 2000, 21, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Chatre, L.; Ducat, A.; Spradley, F.T.; Palei, A.C.; Chereau, C.; Couderc, B.; Thomas, K.C.; Wilson, A.R.; Amaral, L.M.; Gaillard, I. , et al. Increased NOS coupling by the metabolite tetrahydrobiopterin (BH4) reduces preeclampsia/IUGR consequences. Redox Biol 2022, 55, 102406. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, B.M.; Cook, L.G.; Danchuk, S.; Puschett, J.B. Uncoupled endothelial nitric oxide synthase and oxidative stress in a rat model of pregnancy-induced hypertension. Am J Hypertens 2007, 20, 1297–1304. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.Y.; Baek, B.S.; Song, S.H.; Kim, M.S.; Huh, J.I.; Shim, K.H.; Kim, K.W.; Lee, K.H. Xanthine dehydrogenase/xanthine oxidase and oxidative stress. Age (Omaha) 1997, 20, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Bortolotti, M.; Polito, L.; Battelli, M.G.; Bolognesi, A. Xanthine oxidoreductase: One enzyme for multiple physiological tasks. Redox Biol 2021, 41, 101882. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, I.; Talosi, G.; Papp, A.; Boda, D. Xanthine oxidase activation in mild gestational hypertension. Hypertens Pregnancy 2002, 21, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Karabulut, A.B.; Kafkasli, A.; Burak, F.; Gozukara, E.M. Maternal and fetal plasma adenosine deaminase, xanthine oxidase and malondialdehyde levels in pre-eclampsia. Cell Biochem Funct 2005, 23, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Holland, O.J.; Hickey, A.J.R.; Alvsaker, A.; Moran, S.; Hedges, C.; Chamley, L.W.; Perkins, A.V. Changes in mitochondrial respiration in the human placenta over gestation. Placenta 2017, 57, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Rosario, F.J.; Gupta, M.B.; Myatt, L.; Powell, T.L.; Glenn, J.P.; Cox, L.; Jansson, T. Mechanistic Target of Rapamycin Complex 1 Promotes the Expression of Genes Encoding Electron Transport Chain Proteins and Stimulates Oxidative Phosphorylation in Primary Human Trophoblast Cells by Regulating Mitochondrial Biogenesis. Sci Rep 2019, 9, 246. [Google Scholar] [CrossRef] [PubMed]
- Aye, I.L.M.H.; Aiken, C.E.; Charnock-Jones, D.S.; Smith, G.C.S. Placental energy metabolism in health and disease-significance of development and implications for preeclampsia. Am J Obstet Gynecol 2022, 226, S928–S944. [Google Scholar] [CrossRef]
- Holland, O.J.; Cuffe, J.S.M.; Dekker Nitert, M.; Callaway, L.; Kwan Cheung, K.A.; Radenkovic, F.; Perkins, A.V. Placental mitochondrial adaptations in preeclampsia associated with progression to term delivery. Cell Death Dis 2018, 9, 1150. [Google Scholar] [CrossRef] [PubMed]
- Marin, R.; Chiarello, D.I.; Abad, C.; Rojas, D.; Toledo, F.; Sobrevia, L. Oxidative stress and mitochondrial dysfunction in early-onset and late-onset preeclampsia. Biochim Biophys Acta Mol Basis Dis 2020, 1866, 165961. [Google Scholar] [CrossRef] [PubMed]
- Al-Gubory, K.H.; Fowler, P.A.; Garrel, C. The roles of cellular reactive oxygen species, oxidative stress and antioxidants in pregnancy outcomes. Int J Biochem Cell Biol 2010, 42, 1634–1650. [Google Scholar] [CrossRef] [PubMed]
- Freire, V.A.F.; Melo, A.D.; Santos, H.L.; Barros-Pinheiro, M. Evaluation of oxidative stress markers in subtypes of preeclampsia: A systematic review and meta-analysis. Placenta 2023, 132, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Yalcin, S.; Ulas, T.; Eren, M.A.; Aydogan, H.; Camuzcuoglu, A.; Kucuk, A.; Yuce, H.H.; Demir, M.E.; Vural, M.; Aksoy, N. Relationship between oxidative stress parameters and cystatin C levels in patients with severe preeclampsia. Medicina (Kaunas) 2013, 49, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Shaarawy, M.; Aref, A.; Salem, M.E.; Sheiba, M. Radical-scavenging antioxidants in pre-eclampsia and eclampsia. Int J Gynaecol Obstet 1998, 60, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Gomes, H.F.; Palei, A.C.; Machado, J.S.; da Silva, L.M.; Montenegro, M.F.; Jordao, A.A.; Duarte, G.; Tanus-Santos, J.E.; Cavalli, R.C.; Sandrim, V.C. Assessment of oxidative status markers and NO bioavailability in hypertensive disorders of pregnancy. J Hum Hypertens 2013, 27, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Adiga, U.; D'Souza, V.; Kamath, A.; Mangalore, N. Antioxidant activity and lipid peroxidation in preeclampsia. J Chin Med Assoc 2007, 70, 435–438. [Google Scholar] [CrossRef] [PubMed]
- Sandrim, V.C.; Palei, A.C.; Metzger, I.F.; Cavalli, R.C.; Duarte, G.; Tanus-Santos, J.E. Interethnic differences in ADMA concentrations and negative association with nitric oxide formation in preeclampsia. Clin Chim Acta 2010, 411, 1457–1460. [Google Scholar] [CrossRef] [PubMed]
- Socha, M.W.; Stankiewicz, M.; Zolniezewicz, K.; Puk, O.; Wartega, M. Decrease in Nitric Oxide Production as a Key Mediator in the Pathogenesis of Preeclampsia and a Potential Therapeutic Target: A Case-Control Study. Biomedicines 2022, 10. [Google Scholar] [CrossRef]
- Sobko, T.; Reinders, C.I.; Jansson, E.; Norin, E.; Midtvedt, T.; Lundberg, J.O. Gastrointestinal bacteria generate nitric oxide from nitrate and nitrite. Nitric Oxide 2005, 13, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Cosby, K.; Partovi, K.S.; Crawford, J.H.; Patel, R.P.; Reiter, C.D.; Martyr, S.; Yang, B.K.; Waclawiw, M.A.; Zalos, G.; Xu, X. , et al. Nitrite reduction to nitric oxide by deoxyhemoglobin vasodilates the human circulation. Nat Med 2003, 9, 1498–1505. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, J.O.; Weitzberg, E.; Gladwin, M.T. The nitrate-nitrite-nitric oxide pathway in physiology and therapeutics. Nat Rev Drug Discov 2008, 7, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Carlstrom, M.; Montenegro, M.F. Therapeutic value of stimulating the nitrate-nitrite-nitric oxide pathway to attenuate oxidative stress and restore nitric oxide bioavailability in cardiorenal disease. J Intern Med 2019, 285, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Kapil, V.; Milsom, A.B.; Okorie, M.; Maleki-Toyserkani, S.; Akram, F.; Rehman, F.; Arghandawi, S.; Pearl, V.; Benjamin, N.; Loukogeorgakis, S. , et al. Inorganic nitrate supplementation lowers blood pressure in humans: role for nitrite-derived NO. Hypertension 2010, 56, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Im, M.W.; Pai, S.H. Nitric oxide production increases during normal pregnancy and decreases in preeclampsia. Ann Clin Lab Sci 2002, 32, 257–263. [Google Scholar] [PubMed]
- Schiessl, B.; Strasburger, C.; Bidlingmaier, M.; Mylonas, I.; Jeschke, U.; Kainer, F.; Friese, K. Plasma- and urine concentrations of nitrite/nitrate and cyclic Guanosinemonophosphate in intrauterine growth restricted and preeclamptic pregnancies. Arch Gynecol Obstet 2006, 274, 150–154. [Google Scholar] [CrossRef]
- Montenegro, M.F.; Amaral, J.H.; Pinheiro, L.C.; Sakamoto, E.K.; Ferreira, G.C.; Reis, R.I.; Marcal, D.M.; Pereira, R.P.; Tanus-Santos, J.E. Sodium nitrite downregulates vascular NADPH oxidase and exerts antihypertensive effects in hypertension. Free Radic Biol Med 2011, 51, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Webb, A.J.; Patel, N.; Loukogeorgakis, S.; Okorie, M.; Aboud, Z.; Misra, S.; Rashid, R.; Miall, P.; Deanfield, J.; Benjamin, N. , et al. Acute blood pressure lowering, vasoprotective, and antiplatelet properties of dietary nitrate via bioconversion to nitrite. Hypertension 2008, 51, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Amaral, J.H.; Ferreira, G.C.; Pinheiro, L.C.; Montenegro, M.F.; Tanus-Santos, J.E. Consistent antioxidant and antihypertensive effects of oral sodium nitrite in DOCA-salt hypertension. Redox Biol 2015, 5, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.; Tian, R.; Lu, N. NADPH oxidase is a primary target for antioxidant effects by inorganic nitrite in lipopolysaccharide-induced oxidative stress in mice and in macrophage cells. Nitric Oxide 2019, 89, 46–53. [Google Scholar] [CrossRef]
- Gao, X.; Yang, T.; Liu, M.; Peleli, M.; Zollbrecht, C.; Weitzberg, E.; Lundberg, J.O.; Persson, A.E.; Carlstrom, M. NADPH oxidase in the renal microvasculature is a primary target for blood pressure-lowering effects by inorganic nitrate and nitrite. Hypertension 2015, 65, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Zhang, X.M.; Tarnawski, L.; Peleli, M.; Zhuge, Z.; Terrando, N.; Harris, R.A.; Olofsson, P.S.; Larsson, E.; Persson, A.E.G. , et al. Dietary nitrate attenuates renal ischemia-reperfusion injuries by modulation of immune responses and reduction of oxidative stress. Redox Biol 2017, 13, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Cordero-Herrera, I.; Kozyra, M.; Zhuge, Z.; McCann Haworth, S.; Moretti, C.; Peleli, M.; Caldeira-Dias, M.; Jahandideh, A.; Huirong, H.; Cruz, J.C. , et al. AMP-activated protein kinase activation and NADPH oxidase inhibition by inorganic nitrate and nitrite prevent liver steatosis. Proc Natl Acad Sci U S A 2019, 116, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Brunetta, H.S.; Politis-Barber, V.; Petrick, H.L.; Dennis, K.; Kirsh, A.J.; Barbeau, P.A.; Nunes, E.A.; Holloway, G.P. Nitrate attenuates high fat diet-induced glucose intolerance in association with reduced epididymal adipose tissue inflammation and mitochondrial reactive oxygen species emission. J Physiol 2020, 598, 3357–3371. [Google Scholar] [CrossRef] [PubMed]
- DesOrmeaux, G.J.; Petrick, H.L.; Brunetta, H.S.; Holloway, G.P. Independent of mitochondrial respiratory function, dietary nitrate attenuates HFD-induced lipid accumulation and mitochondrial ROS emission within the liver. Am J Physiol Endocrinol Metab 2021, 321, E217–E228. [Google Scholar] [CrossRef] [PubMed]
- Carlstrom, M.; Persson, A.E.; Larsson, E.; Hezel, M.; Scheffer, P.G.; Teerlink, T.; Weitzberg, E.; Lundberg, J.O. Dietary nitrate attenuates oxidative stress, prevents cardiac and renal injuries, and reduces blood pressure in salt-induced hypertension. Cardiovasc Res 2011, 89, 574–585. [Google Scholar] [CrossRef] [PubMed]
- Sindler, A.L.; Fleenor, B.S.; Calvert, J.W.; Marshall, K.D.; Zigler, M.L.; Lefer, D.J.; Seals, D.R. Nitrite supplementation reverses vascular endothelial dysfunction and large elastic artery stiffness with aging. Aging Cell 2011, 10, 429–437. [Google Scholar] [CrossRef]
- Davies, M.J. Protein oxidation and peroxidation. Biochem J 2016, 473, 805–825. [Google Scholar] [CrossRef] [PubMed]
- Ling, W.C.; Mustafa, M.R.; Vanhoutte, P.M.; Murugan, D.D. Chronic administration of sodium nitrite prevents hypertension and protects arterial endothelial function by reducing oxidative stress in angiotensin II-infused mice. Vascul Pharmacol 2018, 102, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Rossman, M.J.; Gioscia-Ryan, R.A.; Santos-Parker, J.R.; Ziemba, B.P.; Lubieniecki, K.L.; Johnson, L.C.; Poliektov, N.E.; Bispham, N.Z.; Woodward, K.A.; Nagy, E.E. , et al. Inorganic Nitrite Supplementation Improves Endothelial Function With Aging: Translational Evidence for Suppression of Mitochondria-Derived Oxidative Stress. Hypertension 2021, 77, 1212–1222. [Google Scholar] [CrossRef] [PubMed]
- Ashor, A.W.; Chowdhury, S.; Oggioni, C.; Qadir, O.; Brandt, K.; Ishaq, A.; Mathers, J.C.; Saretzki, G.; Siervo, M. Inorganic Nitrate Supplementation in Young and Old Obese Adults Does Not Affect Acute Glucose and Insulin Responses but Lowers Oxidative Stress. J Nutr 2016, 146, 2224–2232. [Google Scholar] [CrossRef] [PubMed]
- Chamy, V.M.; Lepe, J.; Catalan, A.; Retamal, D.; Escobar, J.A.; Madrid, E.M. Oxidative stress is closely related to clinical severity of pre-eclampsia. Biol Res 2006, 39, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Surmiak, P.; Wojnarowicz, O.; Szymkowiak, M. Malondialdehyde and Neutrophil Gelatinase-Associated Lipocalin as Markers of Oxidative Stress in Small for Gestational Age Newborns from Hypertensive and Preeclamptic Pregnancies. Biomed Res Int 2022, 2022, 9246233. [Google Scholar] [CrossRef] [PubMed]
- Gratacos, E.; Casals, E.; Deulofeu, R.; Cararach, V.; Alonso, P.L.; Fortuny, A. Lipid peroxide and vitamin E patterns in pregnant women with different types of hypertension in pregnancy. Am J Obstet Gynecol 1998, 178, 1072–1076. [Google Scholar] [CrossRef] [PubMed]
- Noris, M.; Todeschini, M.; Cassis, P.; Pasta, F.; Cappellini, A.; Bonazzola, S.; Macconi, D.; Maucci, R.; Porrati, F.; Benigni, A. , et al. L-arginine depletion in preeclampsia orients nitric oxide synthase toward oxidant species. Hypertension 2004, 43, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.W.; Vaughan, J.E.; Wang, Y.; Roberts, L.J., 2nd. Placental isoprostane is significantly increased in preeclampsia. FASEB J 2000, 14, 1289–1296. [Google Scholar] [CrossRef] [PubMed]
- D'Souza J, M.P.; Harish, S.; Pai, V.R.; Shriyan, C. Increased Oxidatively Modified Forms of Albumin in Association with Decreased Total Antioxidant Activity in Different Types of Hypertensive Disorders of Pregnancy. Indian J Clin Biochem 2017, 32, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Kimura, C.; Watanabe, K.; Iwasaki, A.; Mori, T.; Matsushita, H.; Shinohara, K.; Wakatsuki, A. The severity of hypoxic changes and oxidative DNA damage in the placenta of early-onset preeclamptic women and fetal growth restriction. J Matern Fetal Neonatal Med 2013, 26, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Hilali, N.; Kocyigit, A.; Demir, M.; Camuzcuoglu, A.; Incebiyik, A.; Camuzcuoglu, H.; Vural, M.; Taskin, A. DNA damage and oxidative stress in patients with mild preeclampsia and offspring. Eur J Obstet Gynecol Reprod Biol 2013, 170, 377–380. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Peleli, M.; Zollbrecht, C.; Giulietti, A.; Terrando, N.; Lundberg, J.O.; Weitzberg, E.; Carlstrom, M. Inorganic nitrite attenuates NADPH oxidase-derived superoxide generation in activated macrophages via a nitric oxide-dependent mechanism. Free Radic Biol Med 2015, 83, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Lee, V.M.; Quinn, P.A.; Jennings, S.C.; Ng, L.L. NADPH oxidase activity in preeclampsia with immortalized lymphoblasts used as models. Hypertension 2003, 41, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Rezeck Nunes, P.; Cezar Pinheiro, L.; Zanetoni Martins, L.; Alan Dias-Junior, C.; Carolina Taveiros Palei, A.; Cristina Sandrim, V. A new look at the role of nitric oxide in preeclampsia: Protein S-nitrosylation. Pregnancy Hypertens 2022, 29, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Chen, F.; Kovalenkov, Y.; Pandey, D.; Moseley, M.A.; Foster, M.W.; Black, S.M.; Venema, R.C.; Stepp, D.W.; Fulton, D.J. Nitric oxide reduces NADPH oxidase 5 (Nox5) activity by reversible S-nitrosylation. Free Radic Biol Med 2012, 52, 1806–1819. [Google Scholar] [CrossRef] [PubMed]
- Zollbrecht, C.; Persson, A.E.; Lundberg, J.O.; Weitzberg, E.; Carlstrom, M. Nitrite-mediated reduction of macrophage NADPH oxidase activity is dependent on xanthine oxidoreductase-derived nitric oxide but independent of S-nitrosation. Redox Biol 2016, 10, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Ashmore, T.; Fernandez, B.O.; Branco-Price, C.; West, J.A.; Cowburn, A.S.; Heather, L.C.; Griffin, J.L.; Johnson, R.S.; Feelisch, M.; Murray, A.J. Dietary nitrate increases arginine availability and protects mitochondrial complex I and energetics in the hypoxic rat heart. J Physiol 2014, 592, 4715–4731. [Google Scholar] [CrossRef] [PubMed]
- Carlstrom, M.; Liu, M.; Yang, T.; Zollbrecht, C.; Huang, L.; Peleli, M.; Borniquel, S.; Kishikawa, H.; Hezel, M.; Persson, A.E. , et al. Cross-talk Between Nitrate-Nitrite-NO and NO Synthase Pathways in Control of Vascular NO Homeostasis. Antioxid Redox Signal 2015, 23, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Ling, W.C.; Murugan, D.D.; Lau, Y.S.; Vanhoutte, P.M.; Mustafa, M.R. Sodium nitrite exerts an antihypertensive effect and improves endothelial function through activation of eNOS in the SHR. Sci Rep 2016, 6, 33048. [Google Scholar] [CrossRef]
- Damacena-Angelis, C.; Oliveira-Paula, G.H.; Pinheiro, L.C.; Crevelin, E.J.; Portella, R.L.; Moraes, L.A.B.; Tanus-Santos, J.E. Nitrate decreases xanthine oxidoreductase-mediated nitrite reductase activity and attenuates vascular and blood pressure responses to nitrite. Redox Biol 2017, 12, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Feng, Y.; Shu, C.; Yuan, R.; Bu, L.; Jia, M.; Pang, B. Dietary Nitrate Protects Against Skin Flap Ischemia-Reperfusion Injury in Rats via Modulation of Antioxidative Action and Reduction of Inflammatory Responses. Front Pharmacol 2019, 10, 1605. [Google Scholar] [CrossRef] [PubMed]
- Menezes, E.F.; Peixoto, L.G.; Teixeira, R.R.; Justino, A.B.; Puga, G.M.; Espindola, F.S. Potential Benefits of Nitrate Supplementation on Antioxidant Defense System and Blood Pressure Responses after Exercise Performance. Oxid Med Cell Longev 2019, 2019, 7218936. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, M.D.; Capo, X.; Reynes, C.; Quetglas, M.; Salaberry, E.; Tonolo, F.; Suau, R.; Mari, B.; Tur, J.A.; Sureda, A. , et al. Dietary Sodium Nitrate Activates Antioxidant and Mitochondrial Dynamics Genes after Moderate Intensity Acute Exercise in Metabolic Syndrome Patients. J Clin Med 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Walsh, S.W. Antioxidant activities and mRNA expression of superoxide dismutase, catalase, and glutathione peroxidase in normal and preeclamptic placentas. J Soc Gynecol Investig 1996, 3, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Osunkalu, V.O.; Taiwo, I.A.; Makwe, C.C.; Akinsola, O.J.; Quao, R.A. Methylenetetrahydrofolate Reductase Enzyme Level and Antioxidant Activity in Women with Gestational Hypertension and Pre-eclampsia in Lagos, Nigeria. J Obstet Gynaecol India 2019, 69, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Qiu, D.; Wu, J.; Li, M.; Wang, L.; Zhu, X.; Chen, Y. Impaction of factors associated with oxidative stress on the pathogenesis of gestational hypertension and preeclampsia: A Chinese patients based study. Medicine (Baltimore) 2021, 100, e23666. [Google Scholar] [CrossRef] [PubMed]
- Caldeira-Dias, M.; Viana-Mattioli, S.; de Souza Rangel Machado, J.; Carlstrom, M.; de Carvalho Cavalli, R.; Sandrim, V.C. Resveratrol and grape juice: Effects on redox status and nitric oxide production of endothelial cells in in vitro preeclampsia model. Pregnancy Hypertens 2021, 23, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Ngo, V.; Duennwald, M.L. Nrf2 and Oxidative Stress: A General Overview of Mechanisms and Implications in Human Disease. Antioxidants (Basel) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Kweider, N.; Huppertz, B.; Wruck, C.J.; Beckmann, R.; Rath, W.; Pufe, T.; Kadyrov, M. A role for Nrf2 in redox signalling of the invasive extravillous trophoblast in severe early onset IUGR associated with preeclampsia. PLoS One 2012, 7, e47055. [Google Scholar] [CrossRef] [PubMed]
- Chigusa, Y.; Tatsumi, K.; Kondoh, E.; Fujita, K.; Nishimura, F.; Mogami, H.; Konishi, I. Decreased lectin-like oxidized LDL receptor 1 (LOX-1) and low Nrf2 activation in placenta are involved in preeclampsia. J Clin Endocrinol Metab 2012, 97, E1862–1870. [Google Scholar] [CrossRef]
- Khadir, F.; Rahimi, Z.; Ghanbarpour, A.; Vaisi-Raygani, A. Nrf2 rs6721961 and Oxidative Stress in Preeclampsia: Association with the Risk of Preeclampsia and Early-Onset Preeclampsia. Int J Mol Cell Med 2022, 11, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Amaral, J.H.; Rizzi, E.S.; Alves-Lopes, R.; Pinheiro, L.C.; Tostes, R.C.; Tanus-Santos, J.E. Antioxidant and antihypertensive responses to oral nitrite involves activation of the Nrf2 pathway. Free Radic Biol Med 2019, 141, 261–268. [Google Scholar] [CrossRef]
- Tropea, T.; Renshall, L.J.; Nihlen, C.; Weitzberg, E.; Lundberg, J.O.; David, A.L.; Tsatsaris, V.; Stuckey, D.J.; Wareing, M.; Greenwood, S.L. , et al. Beetroot juice lowers blood pressure and improves endothelial function in pregnant eNOS(-/-) mice: importance of nitrate-independent effects. J Physiol 2020, 598, 4079–4092. [Google Scholar] [CrossRef]
- Ormesher, L.; Myers, J.E.; Chmiel, C.; Wareing, M.; Greenwood, S.L.; Tropea, T.; Lundberg, J.O.; Weitzberg, E.; Nihlen, C.; Sibley, C.P. , et al. Effects of dietary nitrate supplementation, from beetroot juice, on blood pressure in hypertensive pregnant women: A randomised, double-blind, placebo-controlled feasibility trial. Nitric Oxide 2018, 80, 37–44. [Google Scholar] [CrossRef]
- Willmott, T.; Ormesher, L.; McBain, A.J.; Humphreys, G.J.; Myers, J.E.; Singh, G.; Lundberg, J.O.; Weitzberg, E.; Nihlen, C.; Cottrell, E.C. Altered Oral Nitrate Reduction and Bacterial Profiles in Hypertensive Women Predict Blood Pressure Lowering Following Acute Dietary Nitrate Supplementation. Hypertension 2023. [Google Scholar] [CrossRef]
- Said Abasse, K.; Essien, E.E.; Abbas, M.; Yu, X.; Xie, W.; Sun, J.; Akter, L.; Cote, A. Association between Dietary Nitrate, Nitrite Intake, and Site-Specific Cancer Risk: A Systematic Review and Meta-Analysis. Nutrients 2022, 14. [Google Scholar] [CrossRef]
- Bernardo, P.; Patarata, L.; Lorenzo, J.M.; Fraqueza, M.J. Nitrate Is Nitrate: The Status Quo of Using Nitrate through Vegetable Extracts in Meat Products. Foods 2021, 10. [Google Scholar] [CrossRef]
- Huang, Y.; Cao, D.; Chen, Z.; Chen, B.; Li, J.; Guo, J.; Dong, Q.; Liu, L.; Wei, Q. Red and processed meat consumption and cancer outcomes: Umbrella review. Food Chem 2021, 356, 129697. [Google Scholar] [CrossRef] [PubMed]
- Shannon, O.M.; Stephan, B.C.M.; Minihane, A.M.; Mathers, J.C.; Siervo, M. Nitric Oxide Boosting Effects of the Mediterranean Diet: A Potential Mechanism of Action. J Gerontol A Biol Sci Med Sci 2018, 73, 902–904. [Google Scholar] [CrossRef] [PubMed]
- Spinnato, J.A., 2nd; Freire, S.; Pinto, E.S.J.L.; Cunha Rudge, M.V.; Martins-Costa, S.; Koch, M.A.; Goco, N.; Santos Cde, B.; Cecatti, J.G.; Costa, R. , et al. Antioxidant therapy to prevent preeclampsia: a randomized controlled trial. Obstet Gynecol 2007, 110, 1311–1318. [Google Scholar] [CrossRef] [PubMed]
- Bondonno, C.P.; Liu, A.H.; Croft, K.D.; Ward, N.C.; Yang, X.; Considine, M.J.; Puddey, I.B.; Woodman, R.J.; Hodgson, J.M. Short-term effects of nitrate-rich green leafy vegetables on blood pressure and arterial stiffness in individuals with high-normal blood pressure. Free Radic Biol Med 2014, 77, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Gladwin, M.T.; Kim-Shapiro, D.B. The functional nitrite reductase activity of the heme-globins. Blood 2008, 112, 2636–2647. [Google Scholar] [CrossRef] [PubMed]
- Martinez, A.; Sanchez-Valverde, F.; Gil, F.; Clerigue, N.; Aznal, E.; Etayo, V.; Vitoria, I.; Oscoz, M. Methemoglobinemia induced by vegetable intake in infants in northern Spain. J Pediatr Gastroenterol Nutr 2013, 56, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Yip, L.; Spyker, D.A. NADH-methemoglobin reductase activity: adult versus child. Clin Toxicol (Phila) 2018, 56, 866–868. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Canonical and non-canonical pathways for nitric oxide (NO) production. The canonical nitric oxide pathway involves the enzymatic conversion of L-arginine to NO and citrulline by endothelial nitric oxide synthase (eNOS), predominantly in vascular endothelial cells. On the other hand, the non-canonical nitric oxide pathway highlights the stepwise reduction of dietary nitrate (NO3-) to nitrite (NO2-) and subsequently to NO in various tissues.
Figure 1.
Canonical and non-canonical pathways for nitric oxide (NO) production. The canonical nitric oxide pathway involves the enzymatic conversion of L-arginine to NO and citrulline by endothelial nitric oxide synthase (eNOS), predominantly in vascular endothelial cells. On the other hand, the non-canonical nitric oxide pathway highlights the stepwise reduction of dietary nitrate (NO3-) to nitrite (NO2-) and subsequently to NO in various tissues.
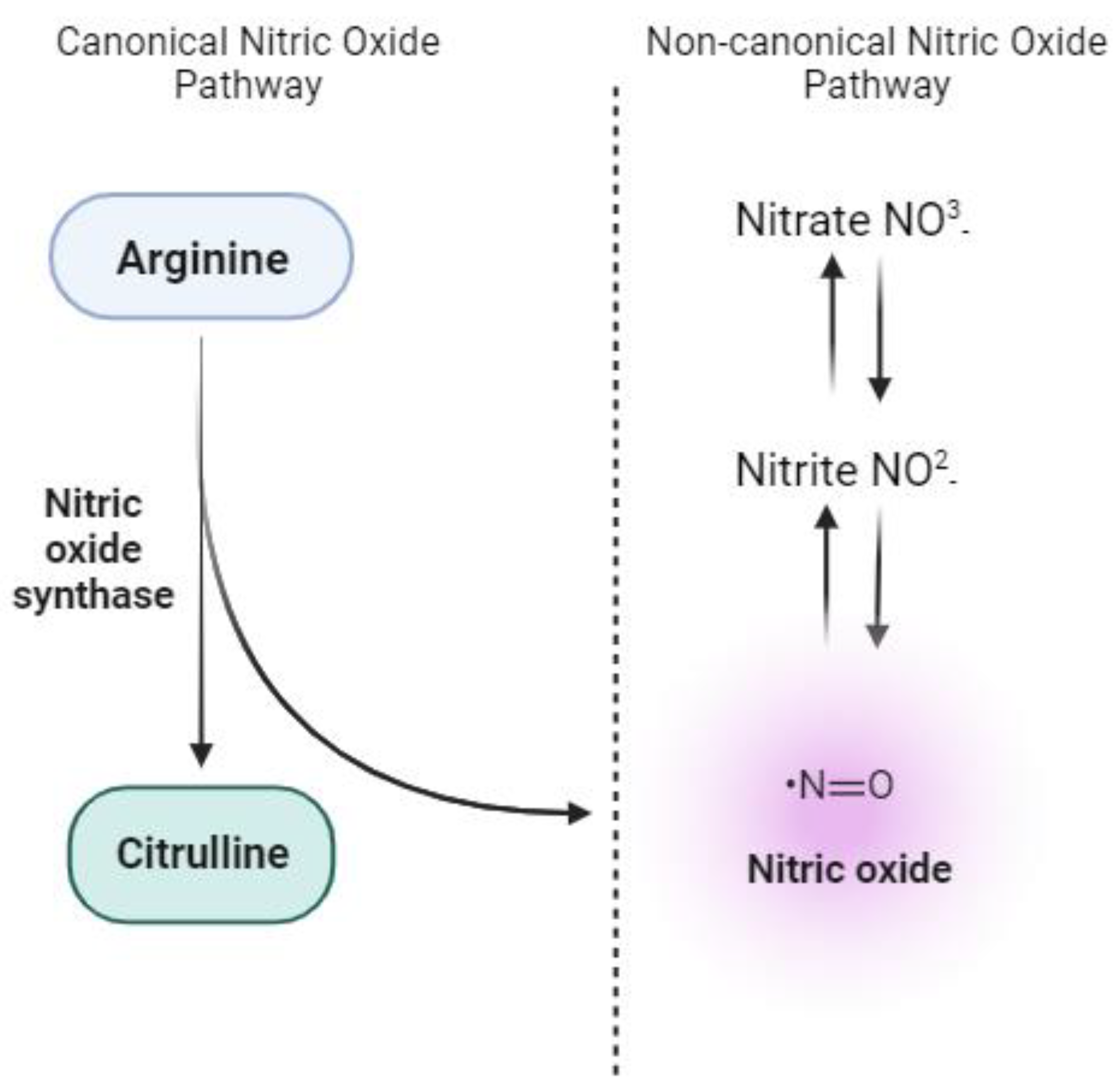

Table 1.
Summary of antioxidant effects on animal studies.
| Model | Source | Dose | Time | Mainly outcomes | Reference |
|---|---|---|---|---|---|
| Oxidative Stress Hypertension | Nitrite Nitrate |
10-5 mol/L 10-2 mol/L |
30 minutes 1 week |
↓ NADPH oxidase activity ↓ Hypertension |
[64] |
| Renal Ischaemia-Reperfusion | Nitrate | 1 mmol/kg/day | 2 weeks | ↓ O2•− levels ↓ IL-12p70, IL-1β, IL-6 |
[65] |
| High-fat diet | Nitrate | 1.0 mmol.kg-1.d-1 | 7 weeks | ↓ NADPH oxidase-derived superoxide and hydrogen peroxide ↓ p67phox protein expression |
[66] |
| High-fat diet | Nitrate | 4 mM | 8 weeks |
↓ Mitochondrial ROS ↓ 4-HNE |
[67] |
| High-fat diet | Nitrate | 4 mM | 8 weeks | ↓ H2O2 mitochondrial emission ↓ TBARs adducts ↓ N3 |
[68] |
| Renal and cardiovascular disease | Nitrate | 0.14 or 1.4 NaNO3 kg -1 | 11 weeks | ↓ Hypertension ↓ Cardiac and renal damage ↓ MDA, 8-OHdG and iPF2α |
[69] |
| Aging | Nitrite | 50 mg/L | 3 weeks | ↑ eNOS expression ↓ Nitrotyrosine levels ↓ IL-1B, IL-6, IFNg and TNF-α |
[70] |
| Hypertension | Nitrite | 50 mg/L | 2 weeks | ↓ Blood pressure ↓ ROS levels ↓ Superoxide Anion ↓ Nitrotyrosine ↓ Nox-4 protein |
[72] |
| Age-associated vascular endothelial dysfunction | Nitrite | 50 mg/L | 8 weeks | ↓ Mitochondrial ROS production ↑ Nrf2 |
[73] |
| 2K1C hypertension | Nitrite | 0.5, 5 and 50mM | 4 weeks | ↓ hypertension ↓ MDA, 8-isoprostane ↓ vascular ROS production ↓ NADPH oxidase activity |
[60] |
| DOCA-salt hypertension | Nitrite | 15 mg/kg | 4 weeks | ↓ hypertension ↓ MDA, 8-isoprostane ↓ NADPH oxidase activity ↓ XOR activity |
[62] |
| Hypoxia | Nitrate | 0.7 mmol-1 | 2 weeks | ↑ Complex I activity ↓ Protein Carbonyls ↓Nitroso-compounds |
[88] |
| IR injury Skin Flap | Nitrate | 5 mmol/L | 7 days before surgery | ↓ histological lesions and protected cells from apoptosis ↑ SOD, GSH-Px, CAT activity ↓MDA ↓ TNF-α, IL-6 |
[92] |
| 2K1C hypertension | Nitrite | 15 mg/kg | 4 weeks | ↓ Hypertension ↓ ROS levels ↑ Nrf2 ↑ SOD1, CAT, GPx2, TRDX-1, TRDX-2 mRNA expression ↑SOD and GPx activity |
[103] |
Legend: 2K1C: 2-kidney, 1-clip; 4-HNE: 4-hydroxynonenal; 8-OHdG: 8-hydroxy-2’-deoxyguanosine; CAT: catalase; DOCA-salt: deoxycorticosterone-salt; GPx: glutathione; GPx2: glutathione peroxidase 2; IFNg: interferon-gamma; IL-12p70: interleukin 12p70; IL-1β: interleukin 1 beta; IL-6: interleukin-6; iPF2α: isoprostane; IR: ischaemia-reperfusion; MDA: malondialdehyde; N3: 3-nitrotyrosine; ROS: reactive oxygen species; SOD: superoxide dismutase; TBARs: thiobarbituric acid reactive substance; TNF-α: tumour necrosis factor-alpha; TRDX-1: thioredoxin-1; TRDX-2: thioredoxin-2; XOR: xanthine oxidoreductase.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



