Submitted:
10 April 2024
Posted:
10 April 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Currently, SARS-CoV-2 has evolved into various variants, including the numerous highly mutated Omicron sub-lineages, significantly increasing immune evasion ability. The development raises concerns about possibly diminished effectiveness of available vaccines and antibody-based therapeutics. Here, we describe those representative categories of broadly neutralizing antibodies (bnAbs) that retain prominent effectiveness against emerging variants including Omicron sub-lineages. The molecular characteristics, epitope conservation, and resistance mechanisms of these antibodies are further detailed, aiming to offer suggestion or direction for the development of therapeutic antibodies, and facilitate the vaccine design with broad-spectrum potential.
Keywords:
SARS-CoV-2 variants
; mutations
; broadly neutralizing antibodies
; epitopes
1. Introduction
On January 30, 2020, the World Health Organization (WHO) declared the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) epidemic as a public health emergency of international concern. Subsequently, on February 11, 2020, the International Committee on Taxonomy of Viruses (ICTV) named the virus as SARS-CoV-2[1], and the WHO officially designated the outbreak of coronavirus disease as Coronavirus Disease-2019, commonly known as COVID-19[2]. As of March, 2024, the WHO has reports over 774 million infections and 7 million deaths globally (https://covid19.who.int/).
The genome analysis reveals that SARS-CoV-2 is a novel beta-coronavirus[3, 4]. The viral genome consists of a about 30 kb, positive-sense, single-stranded RNA, encoding nonstructural proteins (NSPs), accessory proteins, and structural proteins. The 16 NSPs (nsp1-nsp16) are required for viral RNA synthesis. Nine accessory proteins provide a selective advantage in the infected host. The four structural proteins: spike protein (S), membrane protein (M), envelope protein (E) and nucleocapsid protein (N), are necessary for assembly and production of the whole viral particles[5] (Figure 1B). Among them, the S protein belongs to class I fusion transmembrane protein, mediating virus attachment to and fusion with the host cell membrane[6]. The M protein serves as the primary structural protein in the envelope, playing a vital role in virus assembly, budding, envelope formation, and pathogenesis[7, 8]. The E protein is a minor structural protein, forming pentameric protein-lipid channels within host cell membrane, crucial for intracellular trafficking[8, 9]. The N protein interacts with the RNA genome, providing protection for the RNA and facilitating the processes of RNA replication and transcription[8, 10].
The S protein exists as a homotrimer, comprising three protomers (Figure 2A). Each protomer contains S1 and S2 subunits, with furin cleavage sites (PRRAR) at the junction of S1/S2 subunits[11]. The furin proprotein convertase cleaves the S protein into S1 and S2 subunits in the infected cells, which greatly enhances the efficiency of viral entry[6]. The distally located S1 subunit, comprises two domains, N-terminal domain (NTD) and C-terminal domain (CTD) functioning as a receptor-binding domain (RBD)[12, 13]. The S2 is a membrane-anchored subunit that consists of a fusion peptide (FP), a heptad repeat 1 (HR1), a heptad repeat 2 (HR2), a transmembrane region (TM), and an intracellular domain (IC)[14] (Figure 1A). The RBDs are located at the apex of S trimer, adopting either receptor-accessible “up” conformation or the receptor-inaccessible “down” conformation [6]. When the virus enters host cell, the RBD firstly binds to the angiotensin-converting enzyme 2 (ACE2) receptor on the target cell, triggering conformational shift of S protein from the inactive prefusion state to the active postfusion state. The shift leads to the dissociation of the S1 subunit from the S2 subunit, exposing the S2’ cleavage site within the S2 subunit[6]. Subsequently, the S2’ site is cleaved by the TMPRSS2 or cathepsin, exposing the FP. Then, HR1 drives FP to insert into the target cell membrane, initiating membrane fusion. Finally, the viral RNA is released into the host cell via fusion pores formed during the merging of viral and cellular membranes[6, 15] (Figure 1C). Given the crucial role of the S protein in the viral entry process, it has been identified as a key target for the development of neutralizing antibodies (nAbs), new drugs, and vaccine design.
2. Cumulative Mutations in SARS-CoV-2 Spike
SARS-CoV-2 has undergone multiple mutations, resulting in diverse variants. Given the growing risk to global public health, WHO has classified the SARS-CoV-2 strains as variants of high consequence (VOHC), variants of concern (VOC), variants of interest (VOI), and variants under monitoring (VUM). Among these, the VOCs, including Alpha (B.1.1.7), Beta (B.1.351), Gamma (P.1), Delta (B.1.617.2), and Omicron (B.1.1.529), are the main drivers of multiple waves of the global COVID-19 pandemic[16]. In late December 2020, Alpha variant was first reported in the United Kingdom. Alpha variant contains 10 amino acid mutations within the NTD, RBD and furin cleavage sites of S protein, notably including Δ69-70, Δ144, N501Y and P681H[8, 17]. In the following weeks, the Beta and Gamma variants rapidly spread in South Africa and Brazil, harboring significant mutations in the S protein[8, 17]. In May 2021, the Delta variant emerged in India as a VOC, quickly replacing other VOCs and significantly increasing global cases. The Delta variant carries 10 amino acid mutations within the S protein, including T19R, G142D, Δ156-157, R158G located in the NTD, L452R, T478K in the RBD, and P681R in the furin cleavage sites [8, 17]. In November 2021, the Omicron variant, first detected in South Africa and Botswana, led to a new global surge in infections and becoming the dominant strain[17, 18]. Over time, Omicron continues to evolve into many different sub-lineages, such as BA.1, BA.2, BA.3, BA.4/5, BF.7, BQ.1, XBB, and EG.5.1, and these new subvariants have shown enhanced ability to transmit and evade antibodies[16, 19]. Meanwhile, WHO has recently identified Omicron sub-lineages XBB.1.5, XBB.1.16 and EG.5 as VOIs (Table 1). Unlike the Alpha, Beta, Gamma, and Delta variants which are associated with a rise in hospitalization and mortality rates compared to the original strain, the Omicron lineage tends to manifest in a less severe form of the illness[17, 20, 21]. However, the Omicron variant is found over 30 mutations in the S protein (Table 1), resulting in a significant reduction or even a loss of neutralizing effectiveness for most nAbs[8, 16, 22, 23]. Specially, studies suggest that BQ and XBB sub-lineages pose serious threats to the efficacy of existing COVID-19 vaccines, and render all authorized antibodies ineffective[24].
Key mutations and widespread transmission have led to the emergence of viral variants that can evade nAbs, as well as the immunity developed from natural infections and vaccinations. The D614G, the first mutation identified in SARS-CoV-2, is universally present across in all VOCs[8]. The emergence and fixation of D614G yielded the variant with enhanced ACE2 binding, transmissibility, and replication, which greatly facilitates virus adaptability[25, 26]. In the S protein, aside from the D614G mutation, the RBD, NTD, and the furin cleavage site are the primary regions undergoing substantial variations that significantly affect viral behavior[8]. Notably, the Δ69-70 deletion in the NTD, found in diverse variants such as Alpha and Omicron, potentially facilitates viral transmission, but does not primarily contribute to nAbs escape[27, 28]. Other mutations in the NTD, including L18F, D80A, H146Y, D253G/Y, and S255F, are associated with reduced or eliminated recognition of NTD-specific antibodies[28]. The N501Y mutation, present in the RBD of Alpha, Beta, Gamma and Omicron variants, enhances the affinity for ACE2, while simultaneously diminishes the effectiveness of nAbs[29, 30]. The variation of E484 site is observed in the RBD of Beta, Gamma, and Omicron variants. Specifically, both Beta and Gamma variants carry the E484K mutation, which slightly enhances the binding between RBD and ACE2[8]. The Omicron variant contains the E484A mutation, which leads to a reduction in binding affinity[8, 31, 32]. The K417 site variation appears in Beta, Gamma and Omicron variants. Both Beta and Omicron variants carry the K417N mutation, while Gamma variant displays K417T mutation. The two different substitutions, K417N/T, similarly affect the variants by reducing the affinity between RBD and ACE2[33]. However, K417N/T mutations usually co-occur with the N501Y and E484K mutations, which are adequate to compensate for the loss of affinity caused by K417N/T[8]. The L452R mutation, unique to the Delta variant, markedly increases viral infectivity and facilitates immune escape by enhancing affinity for ACE2[8, 34, 35]. Various single mutations of SARS-CoV-2 variants can impair the efficacy of nAbs that target diverse epitopes[36]. Generally, mutations including K417N, G446S, E484A, and Q493R enable the SARS-CoV-2 variants to escape most class 1 and class 2 antibodies targeting the ACE2 binding site[36-38]. Similarly, mutations G339D, N440K, and S371L make several class 3 and class 4 nAbs ineffective[36, 38]. Moreover, the P681H mutation, observed within furin cleavage sites in Alpha and Omicron variants, slightly enhances S1/S2 cleavage[39]. The Delta variant, carrying P681R mutation, significantly facilitates the S protein cleavage and cell fusion[40]. In summary, the substantial number of mutations in the S protein result in extensive antibodies escape, posing a significant challenge to the efficiency of current vaccines and antibody treatments [16,19,41].
3. Potent bnAbs against SARS-CoV-2
It is critical to continue searching for nAbs that exhibit efficacy against emerging SARS-CoV-2 variants, particularly those with the capacity for broad neutralization and prevention of infections from VOCs, including the Omicron sub-lineages. The currently reported nAbs against the SARS-CoV-2 mainly target the S protein, with the majority of highly effective antibodies focusing on the RBD (Figure 2A). Furthermore, antibodies targeting the NTD and S2 of the S protein have also shown potential in neutralizing emerging SARS-CoV-2 variants. The properties of these antibodies are primarily described as follows.
3.1. RBD Antibodies
Antibodies targeting RBD of SARS-CoV-2 demonstrate significant neutralizing potency, therefore, the majority of antibodies currently undergoing clinical trials are RBD nAbs. These antibodies are further classified based on the specific epitopes they target. In this review, we will primarily adopt the Barnes classification to elucidate these categories [38] (Figure 2B).
3.1.1. Class 1 Antibodies
The class 1 antibodies specifically recognize the “up” conformation of the RBD. The epitope targeted by class 1 antibodies overlaps with the receptor-binding motif (RBM) (Figure 2B and 2C), allowing these antibodies to effectively neutralize SARS-CoV-2 by blocking the interaction between RBD and ACE2[38]. However, prevalent mutations within the RBD, such as K417N, E484K, and N501Y, can weaken the affinity of these antibodies for the RBD, facilitating the viral escape neutralization[42]. Although the most class 1 antibodies display limited neutralization breadth across different coronavirus, a minority of antibodies demonstrate broad and effective neutralizing activity against multiple SARS-CoV-2 variants (Table 2).
Firstly, a series of bnAbs with F486 as a critical central residue have been identified. The S2E12, a typical of these antibodies, extensively packs the F486 within a cavity formed by aromatic residues at the interface between heavy chain and light chain of antibody, while mutation residues E484 and S477 are located at the epitope edge. S2E12 exhibits broad neutralizing activity, showing remarkable efficacy against Alpha, Beta, Gamma, Delta, and BA.1, BA.2 variants, with inhibitory concentrations (IC50) values ranging from 0.6 to 137.2 ng/mL[43-45]. Further structural and functional analysis revealed that antibodies A23-58.1, B1-182.1, Cv2.1169 and 87G7 share a similar RBD recognition pattern with S2E12[46-48]. Both A23-58.1 and B1-182.1 can neutralize the Alpha, Beta, Gamma, Delta, and BA.1 variants with IC50 values ranging from 1.6 to 231.0 ng/mL and 0.6 to 281.0 ng/mL, respectively[44, 47]. Cv2.1169 and 87G7 are also effective against SARS-CoV-2 VOCs, Omicron sub-lineages BA.1 and BA.2[46, 48]. Moreover, the human nAb 17T2, sharing a high sequence identity with S2E12, but engages a larger interaction region with the RBM compared to S2E12. This enhanced area contributes to its high affinity, guaranteeing a complete blockade of RBD, thereby enabling 17T2 to effectively neutralize a broad spectrum of SARS-CoV-2 variants, including multiple Omicron sub-lineages, BA.1, BA.2, BA.2.86, BA.4/5, BQ.1.1, XBB.1.5, and XBB.1.16[45]. Importantly, the prophylactic and therapeutic application of 17T2 significantly reduced microscopic lung lesions in a mouse model infected with the Omicron variant[45]. Additionally, the AZD8895, which forms a hydrogen bond network around the residue F486, effectively neutralize a range of variants, including VOCs, and BA.1 with IC50 values ranging from 1.4 to 269.0 ng/mL[44, 49, 50]. The antibodies exhibit a high degree of resistance against viral escape due to the significant detrimental effects of F486 mutation, which reduce the binding of RBD to ACE2 and the viral replicative fitness[43, 51]. However, in subsequent subvariants containing the F486S mutation, such as BA.2.75.2, the neutralizing capacity may be impaired[52].
Another, the ACE2-mimic antibodies, such as S2K146, with a binding site highly similar to ACE2, effectively blocks receptor attachment[53]. As a human cross-neutralizing antibody, S2K146 shows considerable neutralizing breadth against sarbecoviruses including SARS-CoV-2 variants[53]. Research has confirmed that S2K146 neutralizes VOCs, and maintains effectiveness against the Omicron sub-lineages, including BA.1, BA.1.1, BA.2, BA.3, BA.2.12.1, BA.4/5, BQ.1, and BQ.1.1[15, 53, 54]. The GAR05, exhibiting a binding mode similar to that of S2K146, can effectively neutralize BA.1, BA.2, BA.5, and protect K18-hACE2 mice from original SARS-CoV-2 and Delta challenge[55]. The antibody P2C-1F11 shows significant neutralizing effectiveness against VOCs, and Omicron sub-lineages including BA.2, BA.2.75, BA.4/5, BF.7 by mimicking ACE2 and triggering shedding of S1[56, 57]. Craig et al. reported the isolation of the human nAb P4J15 from a convalescent donor, noting its exceptional breadth and neutralization potential against SARS-CoV-2 VOCs, XBB.2.3 and EG.5.1 sub-lineages[58]. Structural analyses revealed that the epitope of P4J15 shares about 93% of its buried surface region with the RBM[58]. In live virus challenge studies, P4J15 offers protection to hamsters against the Omicron BA.5 infection and to monkeys against the XBB.1.5 variant[58]. Moreover, a group of nAbs, including P2-1B1, P5-1C8, P5S-2B10, P5S-2B6, and P5-1H1, isolated from the peripheral memory B cells, also bind to RBD by mimicking ACE2[59]. These antibodies exhibit broad neutralizing capability against a range of SARS-CoV-2 variants, including Alpha, Beta, Gamma, Delta, BA.1, BA.2, BA.2.12.1, BA.2.75, BA.3, and BA.4/5[59].
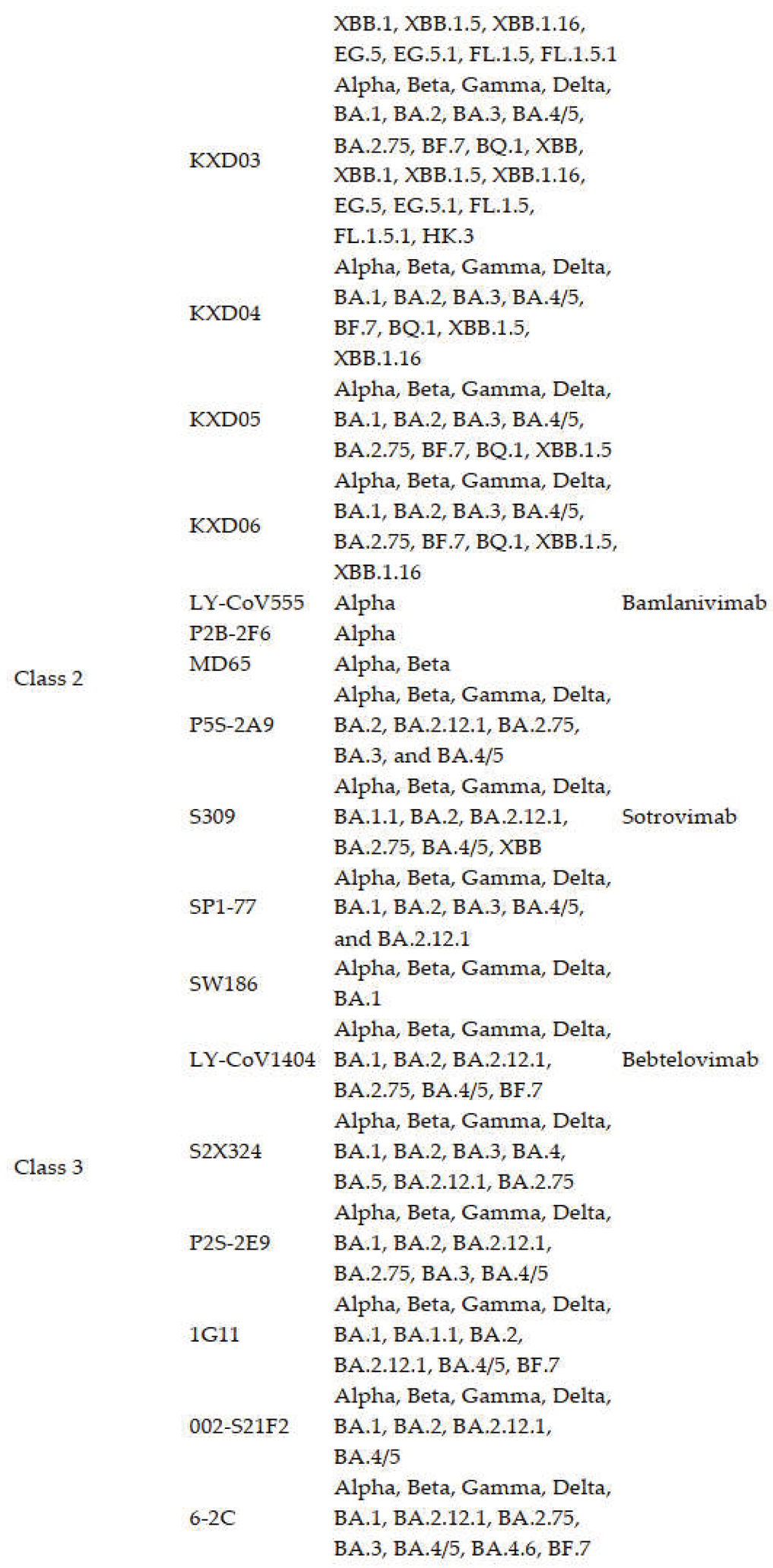
Besides, the antibody 10-5B, isolated from individual immunized the inactivated vaccine, demonstrates remarkable efficacy against the most SARS-CoV-2 variants, including Omicron subvariants BA.1, BA.2, BA.2.12.1, BA.3, with IC50 values ranging from 2.0 to 46.0 ng/mL[60]. The 10-5B epitope largely overlaps with the RBM site, directly competing with ACE2 for RBD binding[60]. However, the potency of 10-5B significantly diminishes against Omicron BA.4/5 and related subvariants due to mutations S477N, E484A, F486V, which affect the formation of hydrogen bonds between RBD and 10-5B[60]. Another group of class 1 antibodies, KXD01, KXD02, KXD03, KXD04, KXD05 and KXD06 are cloned from B cells of convalescent donor that probably infected SARS-CoV-2 BA.5 or BF.7[57]. Remarkably, the antibodies KXD01-06 can broadly neutralize all the VOCs, including Omicron XBB and BQ sub-lineages, and even KDX01-03 show neutralizing effectiveness against the recent EG.5 and FL.1.5 variants[57]. Notably, these nAbs are common found in the convalescent individuals, and their considerable neutralization breadth is attributed to extensive affinity maturation[57].
Thess class 1 antibodies are characterized by their potent and broad neutralizing capability, which enable them effectively to neutralize emerging SARS-COV-2 variants. These antibodies could be used either alone or in combination with other classes to manufacture therapeutic antibodies and also offer valuable insights for the development of next generation vaccine design.
3.1.2. Class 2 Antibodies
The class 2 antibodies bind to RBM in both “up” and “down” RBD, and neutralize by blocking interaction between ACE2 and RBD, which is similar to class 1 antibodies[38](Figure 2B and 2D). Research shows that the K417N and E484K substitutions diminish the binding and neutralization capability of several class 2 antibodies[61]. The therapeutic antibody LY-CoV555 (bamlanivimab) experiences complete inhibition of binding due to the E484K mutation[62]. Furthermore, this mutation also decreases the binding affinity of P2B-2F6 to the S protein, thereby restricting the neutralization capability of LY-CoV555 and P2B-2F6 to only the Alpha VOC[63-65]. In contrast, another antibody MD65, showing a similar binding profile to LY-CoV555, is not impacted in its binding efficacy by the E484K mutation[66]. MD65 exhibits efficacy against Alpha, Beta and Gamma variants in vivo assessment on K18-hACE2 transgenic mice[66]. Additionally, P5S-2A9, isolated from peripheral memory B cells, displays broad neutralizing efficacy against multiple VOCs, including BA.2, BA.2.12.1, BA.2.75, BA.3, and BA.4/5[59] (Table 2). Remarkably, despite P5S-2A9 exhibits low neutralization potency, such broad neutralization is rare among class 2 antibodies, suggesting the potential for developing broadly reactive class 2 nAbs[59].
3.1.3. Class 3 Antibodies
The class 3 antibodies recognize non-RBM epitopes on the RBD in both “up” and “down” conformation[38](Figure 2B and 2E). The majority of class 3 antibodies target a relatively conserved and buried region, maintaining considerable neutralizing activity against Omicron sub-lineages[67]. However, the emergence of XBB, BQ.1, and subsequent variants has led to the loss of conservation in this region, likely attributed to new mutations including R346T, K444T, V445P and N460K, which have been identified as key factors in severe immune escape[67, 68].
The typical class 3 nAb, S309(sotrovimab), specially binds to an epitope containing a glycan universally conserved across the sarbecovirus, without blocking ACE2 attachment[69]. Identified in a patient infected with SARS-CoV in 2003 and subsequently isolated in 2013, S309 exhibits a strong affinity for SARS-CoV-2 RBD[69]. Additionally, it demonstrates broad neutralization against SARS-CoV-2 VOCs, and Omicron subvariants BA.1.1, BA.2, BA.2.12.1, BA.2.75, BA.4/5, XBB[67]. In May 2021, sotrovimab, a stabilized derivative of S309 optimized for enhanced Fc receptor affinity, was granted emergency use authorization as a therapeutic antibody to prevent disease progression in high-risk, early-stage COVID-19 patients[70, 71]. Although S309 has a high neutralizing breadth, its neutralizing efficacy against several variants is relatively low. Particularly, the mutations R346T, K444T, and V445P in the RBD notably increase immune evasion in the BF.7, BQ, and XBB sub-lineages[67]. The SP1-77, an antibody derived from a single human VH-rearranging mouse, interacts with the N343 glycosylation site in a manner similar to S309 and exhibits a superior neutralizing effectiveness against VOCs, BA.1, BA.2, BA.3, BA.4/5, and BA.2.12.1 variants, with IC50 values ranging from 6.5 to 76.0 ng/mL[72]. Unlike class 1 and class 2 antibodies, S309 and SP1-77 do not inhibit attachment between SARS-CoV-2 and the host cell. Instead, S309 blocks the virus through mechanisms like S trimer cross-linking, steric hindrance or virion aggregation, and SP1-77 prevents fusion of viral and cellular membranes by blocking S1 dissociation, impacting the subsequent steps of the SARS-CoV-2 infection process[69, 72]. The antibody SW186 recognizes a conserved epitope outside the RBM, allowing it to maintain neutralizing effectiveness against various SARS-CoV-2 variants, such as Alpha, Beta, Gamma, Delta, and BA.1[73]. The SW186 epitope consists of conserved amino acids, particularly the N343 glycosylation site, which plays a pivotal role in the transition of the RBD from a glycan-shielded “down” conformation to an accessible “up” conformation, crucial for viral entry into host cells. Despite SW186 does not directly bind to the RBM, it still exhibits partial inhibition of the binding interaction between the RBD and ACE2[73].
Another antibody LY-CoV1404 (bebtelovimab), isolated from a convalescent donor, demonstrates strong neutralization capability for prevalent VOCs, and Omicron subvariants BA.1, BA.2, BA.2.12.1, BA.2.75, BA.4/5, BF.7 with IC50 values between 0.7 and 2.2 ng/mL[67, 74]. The LY-CoV1404 targets a region accessible in both the “up” and “down” conformations of the RBD, partially overlapping with the RBM[74]. Consequently, LY-CoV1404 impedes the binding of the ACE2, explaining the strong mechanism for its potent neutralizing activity[74]. Although this property suggests that LY-CoV1404 is a class 2 antibody, the location of its epitope is closer to the canonical class 3 antibody S309[74]. Further analysis demonstrates that aside from the common mutations N439 and N501, the contact residues of the LY-CoV1404 epitope are highly conserved. However, the binding and neutralizing capability of LY-CoV1404 remains unaffected by N439K and N501Y mutations[74]. The S2X324 recognizes a similar region to LY-CoV1404, which partially overlaps with the RBM[75]. The S2X324 exhibits cross-reactivity and effectively neutralizes Omicron subvariants BA.1, BA.2, BA.3, BA.4, BA.5, BA.2.12.1, and BA.2.75 with IC50 values below 10.0 ng/mL, except BA.2.75, with IC50 value 18.0 ng/mL[75]. Biochemical experiments reveal that S2X324 primarily inhibits SARS-CoV-2 infection by blocking ACE2 binding[75]. Additionally, the antibody P2S-2E9, with extensive overlap in binding footprints with LY-CoV1404, effectively neutralize various SARS-CoV-2 variants, including VOCs, and Omicron subvariants BA.1, BA.2, BA.2.12.1, BA.2.75, BA.3, and BA.4/5[59].
The antibody 1G11 specially targets an epitope outside the N343 glycan site and the RBM, thereby positioning it uniquely between the binding domains of S309 and LY-CoV1404[67]. Notably, 1G11 demonstrates robust neutralizing activity against pseudotyped SARS-CoV-2 VOCs, maintaining high level against Omicron subvariants BA.1, BA.1.1, BA.2, BA.2.12.1, BA.4/5, and BF.7[67]. 1G11 does not interfere binding of RBD with ACE2, instead, it promotes cross-linking of spikes after binding to the RBD, clarifying its mechanism for potent ACE2 blockade and exceptional neutralization efficacy[67]. Moreover, the 002-S21F2 recognizes a conformationally conserved epitope located on the external surface of the RBD. It has shown consistent neutralizing efficacy against a range of SARS-CoV-2 variants, including VOCs, BA.1, BA.2, BA.2.12.1, BA.4, and BA.5, with IC50 values between 20.0 and 130.0 ng/mL[76]. Additionally, the antibody 6-2C, derived from individual immunized with the inactivated vaccine, can broadly neutralized VOCs, and the variants highly evasive to antibodies, such as XBB.1.5, XBB, BQ.1.1, and BA.2.75.2[60] (Table 2). The epitope targeted by 6-2C is mutationally constrained in terms of folding and expression, which accounts for its broad neutralization potential[60].
3.1.4. Class 4 Antibodies
The Class 4 antibodies bind to non-RBM epitopes in “up” RBD[38, 77] (Figure 2B and 2F). The majority of antibodies previously described are cross-reactive but weakly neutralizing[78, 79].
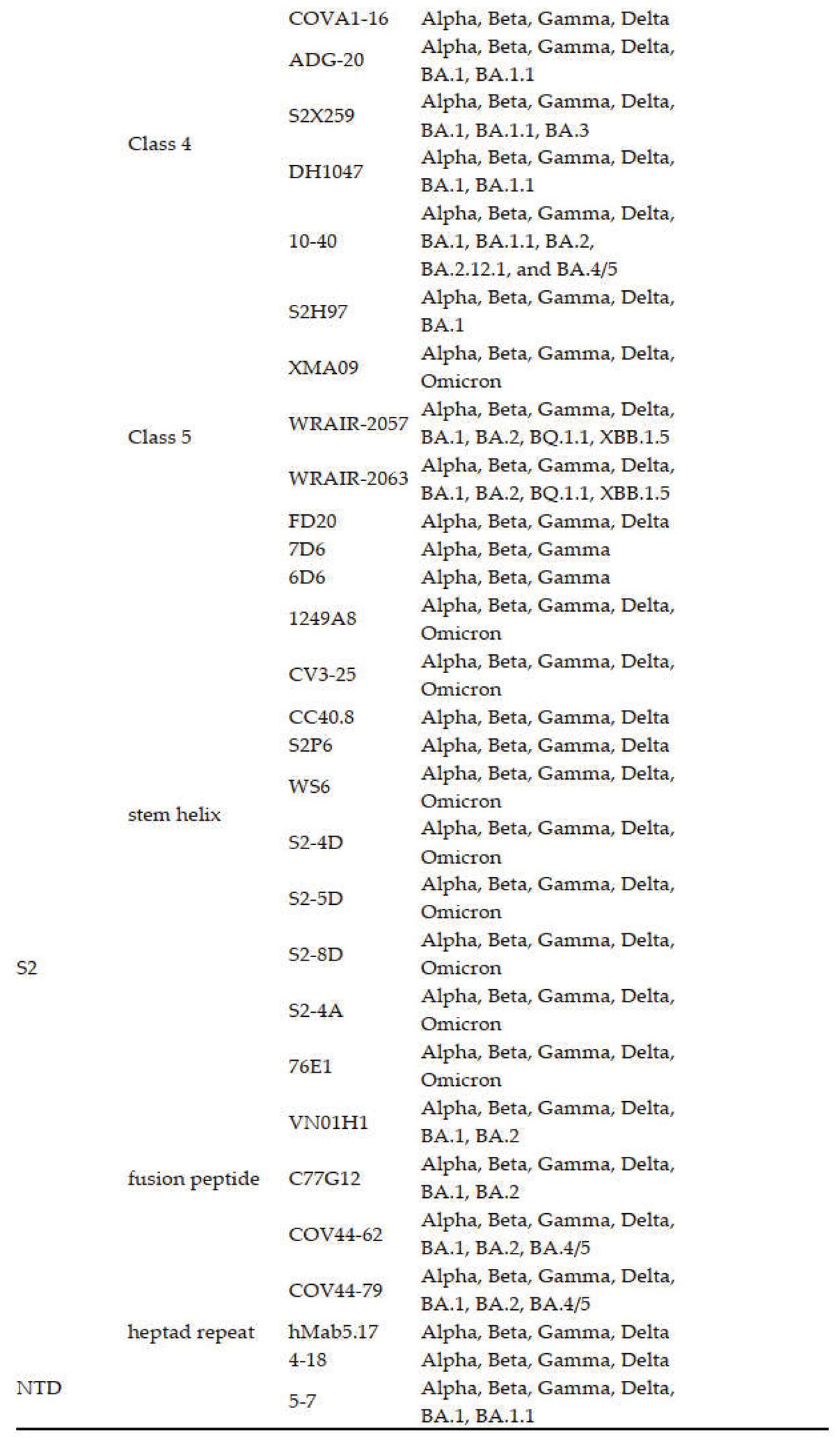
The classic class 4 antibody CR3022, isolated from a convalescent SARS patient, specifically recognizes a highly conserved epitope shared between SARS-CoV-2 and SARS-CoV, distinct from the RBM[77]. Further structural analysis demonstrates that CR3022 epitope is accessible only when at least two RBDs of the S trimer are in “up” conformation with a slight rotation[77]. The epitope of CR3022 is inaccessible in the prefusion state of spike, suggesting that its binding facilitates transition to the postfusion state[80]. Cryogenic electron microscopy analysis confirms that incubation of CR3022 Fab with spike protein results in disruption of the prefusion trimer[80]. Despite the emergence of multiple VOCs and Omicron sub-lineages, only one residue mutation within the CR3022 epitope is observed, underlining its potential in therapeutic applications[81]. However, antibodies targeting the CR3022 epitope display a broad binding breadth but low neutralization strength against SARS-CoV-2[78]. Specifically, COVA1-16, which also targets this epitope, loses its neutralization activity against the Omicron variant[82]. The diminished effectiveness can be attributed to multiple factors: insufficient affinity, the relative inaccessibility of the epitope, or a direct competition failure with the ACE2 receptor[78]. Notably, nAb ADG-20, an affinity-matured derivative of ADI-55688, interacting with the CR3022 epitope, demonstrates neutralizing efficacy against BA.1 and BA.1.1 by competing with ACE2[78].
The nAb S2X259, derived from the memory B cells of a COVID-19 convalescent donor, recognizes a cryptic conserved epitope of the RBD and demonstrates cross-reactivity with spike proteins of all sarbecovirus[79]. It can broadly neutralize SARS-CoV-2 VOCs, Omicron subvariants BA.1, BA.1.1, BA.3, and a range of sarbecoviruses, by inhibiting ACE2 binding[19, 79]. Remarkably, the S2X259 epitope is conserved across currently circulating strains and does not comprise prevalent RBD mutations, such as S477N, N439K or L452R[79]. Further analyses demonstrate that the rare G504D substitution is the only escape mutation for S2X259, suggesting this nAb might have a high barrier against the SARS-CoV-2 escape mutants[79]. Another cross-reactive antibody, DH1047, has been proven its efficacy in neutralizing BA.1, BA.1.1 by targeting a conserved epitope within RBD[19, 83]. Similarly, the nAbs 10-40 exhibits broad neutralizing capability against a range of variants of SARS-CoV-2, including Omicron subvariants BA.1, BA.1.1, BA.2, BA.2.12.1, BA.4/5, and also SARS-CoV[19, 84] (Table 2). The epitopes of 10-40 and DH1047 significantly overlap, however, angles that they approach the targeted site are different, highlighting the targeted site is a viable target for developing a universal sarbecovirus vaccine[84].
The class 3 and class 4 epitope exhibits a high level of conservation across sarbecoviruses, suggesting their potential as effective targets for bnAbs, and indicating that the epitopes maintain functional stability and have a reduced association with immune escape mechanisms[38]. Additionally, class 3 and class 4 antibodies enrich the SARS-CoV-2 antibody repertoire, probably offering effective use in therapeutic combinations with class 1 and class 2 nAbs, which lead to additive neutralization effects and simultaneously curb viral escape mechanisms.
3.1.5. Class 5 Antibodies
The class 5 antibodies target a cryptic region, previously described as the “E465 patch”, that is highly conserved among diverse sarbecoviruses, including SARS-CoV-2[31, 51]. The class 5 epitope substantially overlaps with the NTD-interacting region and is exposed only when the spike protein adopts the “up” conformation with at least one RBD accessible[51, 85] (Figure 2B and 2G).
S2H97 exhibits high-affinity binding to a cryptical conserved epitope across all sarbecovirus clades, which is designated as site 5 (Figure 2G). This binding requires a substantial opening of the RBD to expose its specific epitope, accelerating the transmission of spike protein to the postfusion conformation and effectively inhibiting viral entry. S2H97 effectively neutralizes diverse sarbecoviruses and SARS-CoV-2 variants, including Omicron BA.1[51]. The antiboody XMA09 exhibits high epitope similarity with S2H97 and maintains comparable neutralizing capability against the Omicron pseudovirus to that of D614G[86]. ION_300 binds to a distinctive region on the opposite side of the RBM, obscured by the NTD of an adjacent S1 polypeptide chain when the RBD is in the closed conformation[87]. Analysis of epitope sites predicts that this antibody is expected to maintain binding and neutralization potency against prevalent RBD mutations, such as K417N/T, E484K, and N501Y[87]. The nAbs WRAIR-2057 and WRAIR-2063, isolated from a convalescent donor, can potently neutralize SARS-CoV and SARS-CoV-2. Moreover, they retain neutralization against various Omicron subvariants, including BA.1, BA.2, BQ.1.1, and XBB.1.5[81, 88]. The epitope of WRAIR-2057 exhibits 48% overlap with S2H97 and 70% with ION-300[81]. The epitope of WRAIR-2063 is accessible only when one or more RBDs is in the “up” conformation, which may facilitate the shift of spike protein from prefusion to postfusion, resulting in the rapid disintegration of the S trimer[85,89]. The superimposition of the WRAIR-2063-RBD complex onto the RBD of the S trimer reveals no significant steric clashes with RBDs of the adjacent protomers, suggesting that multiple WRAIR-2063 antibody could bind to the S trimer[85]. The antibodies FD20, 7D6 and 6D6 show a similar neutralization mechanism with that of WRAIR-2063, which involves disassembling the spike protein[85, 89, 90]. The FD20, isolated from the COVID-19 convalescent patient can neutralizes Alpha, Beta, Gamma and Delta[85, 89]. The murine cross-neutralizing antibodies 7D6 and 6D6 showed almost consistent neutralizing efficacy against Alpha, Beta and Gamma variants. The binding of 7D6 and 6D6 with the RBD causes steric hindrance with the adjacent NTD, thereby disrupting the viral spike[85, 89]. Furthermore, research demonstrates that the interaction of the S protein with CR3022 and other antibodies recognizing cryptic epitopes is notably improved when WRAIR-2063 is present, suggesting that engagement with WRAIR-2063 induces conformational alterations in the S trimer, thereby affecting accessibility of these cryptic sites[85]. This may indicate that binding of class 5 antibodies leads to more significant conformational rearrangements in the S trimer, ultimately impacting neutralization effectiveness and breadth of antibodies that aim at associated epitopes.
Despite the continuous emergence of VOCs including Omicron, none has yet revealed mutations within the class 5 epitope, highlighting the therapeutic potential of these antibodies[81]. The class 5 antibodies exhibit a spectrum of neutralization capability and a considerable breadth against a variety of sarbecoviruses (Table 2), emphasizing their critical role in the development of vaccine and therapeutics.
3.2. NTD Antibodies
The limited immunogenicity of the SARS-CoV-2 NTD in COVID-19 patients has been attributed to its extensive N-linked glycan shielding[91, 92]. However, recent studies report the isolation of NTD antibodies that show neutralizing capability against SARS-CoV-2 infection in vitro, indicating their potential utility in COVID-19 prophylaxis or treatment[28] (Table 2). The first nAb targeting SARS-CoV-2 NTD, 4A8, was reported by Chi et al., exhibits neutralizing potency with an IC50 value of 0.39μg/mL against the authentic SARS-CoV-2 virus[93]. Matthew et al. described an antigenic map of the heavily glycosylated SARS-CoV-2 NTD by characterizing 41 NTD human antibodies, in which six antigenic sites (i-vi) were designed[28]. Most potently nAbs, including 4A8, competed for binding to the NTD site i, which is identified a supersite. NTD is highly glycosylated, and its supersite, positioned at the periphery of the spike protein, represents the largest glycan-free area on the NTD[94]. The three regions, including NTD N-terminal region, residues 14-20, a β-hairpin formed by residues 140-158, and a loop spanning residues 245-264, targeted by antibodies S2L28, S2M28, and S2X333, together constitute an antigenic supersite at the apex of the NTD, on the side distal to the viral membrane. Among these, S2X333 inhibits the fusion between cells, activates effector functions, and offers protection to hamsters against SARS-CoV-2 challenge[28]. However, SARS-CoV-2 VOCs possess a large number of mutations within the NTD supersite, potentially constraining the recognition and neutralizing capability of nAbs at supersite[28, 95, 96]. The antibody 4-18, targeting supersite of the NTD lose neutralizing activity against all Omicron subvariants[94, 97]. Another potent nAb 5-7, recognizing NTD outside of supersite, retains neutralization potency against VOCs, including BA.1 and BA.1.1, with binding site remote from most mutations[19, 98]. In general, nAbs targeting the NTD achieve neutralization by locking the S protein thereby inhibiting conformational transitions to postfusion state, rather than by directly blocking ACE2 receptor[99].
Generally, the NTD region exhibits significant variability across various coronaviruses, making it an unlikely target for cross-neutralizing antibodies. Nevertheless, combining NTD nAbs with RBD antibodies may enhance the efficacy of antibody cocktails.
3.3. S2 Antibodies
Compared to the S1 subunit, the amino acid sequence of the S2 subunit are relatively conserved, which is consistent with its conserved function[100]. The S2 subunit plays an important role in facilitating the fusion of viruses with host cell membranes, suggesting a low tolerance for variants without affecting the viral fitness. Studies have found that S2 contains immunodominant epitopes capable of eliciting nAbs[101, 102]. Despite the limited production of S2-specific antibodies following natural SARS-CoV-2 infection or vaccination, and only a few antibodies show neutralizing activity, those targeting specific S2 epitopes tend to show broad neutralizing potential[103-105] (Table 2).
Notably, the S2 stem helix region within residues 1141-1160 of S protein demonstrates conservation across various SARS-CoV-2 VOCs[106]. Therefore, it represents an optimal target for the development of bnAbs. The S2 stem helix-specific antibody 1249A8 can universally recognize all human β-coronavirus and effectively neutralize SARS-CoV-2, SARS-CoV, and MERS-CoV by inhibiting the post-attachment fusion process. Furthermore, it also shows prophylactic activities against SARS-CoV-2 WA-1, Beta and Omicron strains in K18 hACE2 mice[105]. Competition surface plasmon resonance (SPR) results reveal that human nAbs CV3-25, CC40.8, and S2P6 target the epitope overlapping with that of 1249A8. They can neutralize various SARS-CoV-2 VOCs, by interfering the membrane fusion crucial for viral infection[107-111]. However, only CV3-25 is capable of neutralizing the Omicron variant, possibly because its binding epitope extends more towards the C-terminally[110, 111]. The antibody WS6, isolated from mice immunized with mRNA encoding the S protein of SARS-CoV-2, could neutralize Alph, Beta, Gamma, Delta, Omicron, and SARS-CoV, by inhibiting the conformational changes of the S protein[108]. Similarly, a group of murine antibodies, S2-4A, S2-4D, S2-5D and S2-8D, also exhibit broad neutralizing activity against these VOCs[112]. Furthermore, the FP sequence within the S2 subunit is completely conserved across all SARS-CoV-2 VOCs. The FP-specific antibody 76E1, derived from a COVID-19 convalescent donor, effectively neutralizes the SARS-CoV-2 VOCs, exhibiting no residue mutations in the 76E1 epitope across current SARS-CoV-2 VOCs[113]. Structural analysis revealed that the epitope of 76E1 is buried within the prefusion S trimer, and a conformational change from the prefusion to the postfusion state leads to the exposure of the epitope to allow binding of 76E1[113]. Then, 76E1 suppresses membrane fusion by blocking the cleavage of S2’[113]. The binding of ACE2 facilitates the exposure of the S2’ site and the fusion peptide, hence 76E1 has a synergistic effect with ACE2 in preventing SARS-CoV-2 infection, suggesting that a potential combination of 76E1 with ACE2 or RBD antibodies could prevent SARS-CoV-2 infection. The antibodies VN01H1 and C77G12 can neutralize the Omicron subvariants BA.1 and BA.2[114]. Similar to 76E1, their binding sites are located within the prefusion S trimer, which allows them to also work synergistically with ACE2 or ACE2-mimicking antibodies to enhance the neutralizing effect[114]. The other two FP-specific antibodies, COV44-62 and COV44-79, demonstrate a broad capacity to neutralize SARS-CoV-2 VOCs, and Omicron subvariants BA.1, BA.2, BA.4/5, by inhibiting virus and host membrane fusion[115]. The epitope mapping reveals that COV44-62 and COV44-79 bind to the same region of the FP helix from distinct angles, with part of their epitope accessible in the prefusion spike protein[115]. Furthermore, binding analysis shows that COV44-62 and COV44-79 exhibit higher affinity for the S2 subunit compared to the prefusion S protein, suggesting that their binding epitopes might be buried by the S1 subunit. This might also explain why FP-specific antibodies are only produced in a subset of COVID-19 patients and rarely during natural infection or vaccination[115]. The hMab5.17, a humanized antibody, targets the HR2 region and is capable of effectively neutralizing both SARS-CoV and SARS-CoV-2 VOCs[116]. Given that the HR2 region exhibits nearly 100% conservation across SARS-CoV-2 VOCs, it suggests a potential for hMab5.17 to elicit bnAbs against SARS-CoV-2[106].
Despite antibodies targeting S2 subunit display broadly neutralizing activity, they tend to exhibit lower neutralizing titers compared to those targeting RBD[106]. Therefore, enhancing the potency of antibodies specific to the S2 subunit remains a significant challenge. Moreover, combination of S1 and S2 antibodies may represent a promising strategy for COVID-19 treatment, potentially preventing the emergence of immune escape strains.
4. Conclusions
The emerging SARS-CoV-2 variants constantly escape neutralization provided by antibodies from current vaccines and natural infections. Upon the advent of the BQ.1, BQ.1.1, and XBB.1 lineages, all the approved therapeutic antibodies have completely lost their effectiveness[117]. Moreover, under the selective immune pressure resulting from the widespread prevalence of viruses circulating in the general population, the SARS-CoV-2 sounds continuing to evolve along. Therefore, it is necessary to develop new antibodies to combat the currently circulating variants and prepare for future strains. Only few antibodies targeting the RBD, NTD, and S2 have shown broad and potent efficacy against SARS-CoV-2 VOCs, including Omicron. Combining these neutralizing antibodies targeting different epitopes or designing bispecific antibodies for cocktail therapy are promising treatment options for patients with severe COVID-19. Specifically, antibody therapy offers potential clinical benefits for individuals at high risk with compromised immune functions, as they are unable to generate a protective humoral immune response following vaccination[118,119]. Furthermore, vaccines specifically targeting the epitopes of these broadly neutralizing antibodies may suggest an excellent strategy for the preparedness to curb next SARS-CoV-2 pandemic.
Author Contributions
S.L. and T.L. designed and funded the study. L.C. and T.L. wrote the original draft, reviewed and edited the manuscript, W.X., S.Z., H.W., H.L., Y.G., N.X., T.L. and S.L. participated in discussion and interpretation of the manuscript.
Data Availability Statement
All relevant data are within the manuscript.
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (grant no: 82001756, 81991491), China Postdoctoral Science Foundation (2023T160381), CAMS Innovation Fund for Medical Sciences of China (grant no: 2019RU022), and the Fundamental Research Funds for the Central Universities (20720220004, 20720220006).
Conflicts of Interests
No potential conflict of interest was reported by the authors.
References
- Hu, B.; et al. Characteristics of SARS-CoV-2 and COVID-19. Nature Reviews Microbiology 2021, 19, 141–154. [Google Scholar] [CrossRef]
- Wang, M.Y.; et al. SARS-CoV-2: Structure, Biology, and Structure-Based Therapeutics Development. Front Cell Infect Microbiol 2020, 10, 587269. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef]
- Lu, R.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding. The Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef]
- Bai, C.; Zhong, Q.; Gao, G.F. . Overview of SARS-CoV-2 genome-encoded proteins. Sci China Life Sci 2022, 65, 280–294. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.B.; et al. Mechanisms of SARS-CoV-2 entry into cells. Nat Rev Mol Cell Biol 2022, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Schoeman, D.; Fielding, B.C. Coronavirus envelope protein: current knowledge. Virology Journal 2019, 16, 69. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ye, Q. . The Key Site Variation and Immune Challenges in SARS-CoV-2 Evolution. Vaccines (Basel) 2023, 11. [Google Scholar] [CrossRef]
- Masters, P.S. The Molecular Biology of Coronaviruses, in Advances in Virus Research; Academic Press, 2006; pp. 193–292. [Google Scholar]
- Chang, C.-k.; et al. The SARS coronavirus nucleocapsid protein – Forms and functions. Antiviral Research 2014, 103, 39–50. [Google Scholar] [CrossRef]
- Yadav, R.; et al. Role of Structural and Non-Structural Proteins and Therapeutic Targets of SARS-CoV-2 for COVID-19. Cells 2021, 10. [Google Scholar] [CrossRef]
- Ke, Z.; et al. Structures and distributions of SARS-CoV-2 spike proteins on intact virions. Nature 2020, 588, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef]
- V'Kovski, P.; et al. Coronavirus biology and replication: implications for SARS-CoV-2. Nat Rev Microbiol 2021, 19, 155–170. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Y.; Zhou, H. Potent antibodies against immune invasive SARS-CoV-2 Omicron subvariants. Int J Biol Macromol 2023, 249, 125997. [Google Scholar] [CrossRef]
- Wang, L.; et al. Immune evasion of neutralizing antibodies by SARS-CoV-2 Omicron. Cytokine Growth Factor Rev 2023, 70, 13–25. [Google Scholar] [CrossRef]
- Markov, P.V.; et al. The evolution of SARS-CoV-2. Nature Reviews Microbiology 2023, 21, 361–379. [Google Scholar] [CrossRef]
- Viana, R.; et al. Rapid epidemic expansion of the SARS-CoV-2 Omicron variant in southern Africa. Nature 2022, 603, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; et al. Antibody evasion by SARS-CoV-2 Omicron subvariants BA.2.12.1, BA.4 and BA.5. Nature 2022, 608, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Challen, R.; et al. Risk of mortality in patients infected with SARS-CoV-2 variant of concern 202012/1: matched cohort study. Bmj 2021, 372, n579. [Google Scholar] [CrossRef]
- Tegally, H.; et al. Detection of a SARS-CoV-2 variant of concern in South Africa. Nature, 2021, 592, 438–443. [Google Scholar] [CrossRef]
- Iketani, S.; et al. Antibody evasion properties of SARS-CoV-2 Omicron sublineages. Nature 2022, 604, 553–556. [Google Scholar] [CrossRef] [PubMed]
- Planas, D.; et al. Considerable escape of SARS-CoV-2 Omicron to antibody neutralization. Nature 2022, 602, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; et al. Alarming antibody evasion properties of rising SARS-CoV-2 BQ and XBB subvariants. Cell 2023, 186, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Omotuyi, I.O.; et al. Atomistic simulation reveals structural mechanisms underlying D614G spike glycoprotein-enhanced fitness in SARS-COV-2. J Comput Chem 2020, 41, 2158–2161. [Google Scholar] [CrossRef]
- Yurkovetskiy, L.; et al. Structural and Functional Analysis of the D614G SARS-CoV-2 Spike Protein Variant. Cell 2020, 183, 739–751. [Google Scholar] [CrossRef]
- Meng, B.; et al. Recurrent emergence of SARS-CoV-2 spike deletion H69/V70 and its role in the Alpha variant B.1.1.7. Cell Rep 2021, 35, 109292. [Google Scholar] [CrossRef] [PubMed]
- McCallum, M.; et al. N-terminal domain antigenic mapping reveals a site of vulnerability for SARS-CoV-2. Cell 2021, 184, 2332–2347. [Google Scholar] [CrossRef]
- Liu, H.; et al. The basis of a more contagious 501Y.V1 variant of SARS-CoV-2. Cell Res 2021, 31, 720–722. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; et al. SARS-CoV-2 and Emerging Variants: Unmasking Structure, Function, Infection, and Immune Escape Mechanisms. Front Cell Infect Microbiol 2022, 12, 869832. [Google Scholar] [CrossRef]
- Starr, T.N.; et al. Deep Mutational Scanning of SARS-CoV-2 Receptor Binding Domain Reveals Constraints on Folding and ACE2 Binding. Cell 2020, 182, 1295–1310. [Google Scholar] [CrossRef]
- Han, P.; et al. Receptor binding and complex structures of human ACE2 to spike RBD from omicron and delta SARS-CoV-2. Cell 2022, 185, 630–640. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; et al. Higher infectivity of the SARS-CoV-2 new variants is associated with K417N/T, E484K, and N501Y mutants: An insight from structural data. J Cell Physiol 2021, 236, 7045–7057. [Google Scholar] [CrossRef] [PubMed]
- Motozono, C.; et al. SARS-CoV-2 spike L452R variant evades cellular immunity and increases infectivity. Cell Host Microbe 2021, 29, 1124–1136. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; et al. Transmission, infectivity, and neutralization of a spike L452R SARS-CoV-2 variant. Cell 2021, 184, 3426–3437. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; et al. Omicron escapes the majority of existing SARS-CoV-2 neutralizing antibodies. Nature 2022, 602, 657–663. [Google Scholar] [CrossRef]
- West, A.P., Jr.; et al. Detection and characterization of the SARS-CoV-2 lineage B.1.526 in New York. Nat Commun 2021, 12, 4886. [Google Scholar] [CrossRef] [PubMed]
- Barnes, C.O.; et al. SARS-CoV-2 neutralizing antibody structures inform therapeutic strategies. Nature 2020, 588, 682–687. [Google Scholar] [CrossRef] [PubMed]
- Lubinski, B.; et al. Functional evaluation of the P681H mutation on the proteolytic activation of the SARS-CoV-2 variant B.1.1.7 (Alpha) spike. iScience 2022, 25, 103589. [Google Scholar] [CrossRef] [PubMed]
- Saito, A.; et al. Enhanced fusogenicity and pathogenicity of SARS-CoV-2 Delta P681R mutation. Nature 2022, 602, 300–306. [Google Scholar] [CrossRef]
- Shah, M.; Woo, H.G. . Omicron: A Heavily Mutated SARS-CoV-2 Variant Exhibits Stronger Binding to ACE2 and Potently Escapes Approved COVID-19 Therapeutic Antibodies. Front Immunol 2021, 12, 830527. [Google Scholar] [CrossRef]
- Miller, N.L.; et al. Insights on the mutational landscape of the SARS-CoV-2 Omicron variant receptor-binding domain. Cell Rep Med 2022, 3, 100527. [Google Scholar] [CrossRef] [PubMed]
- Tortorici, M.A.; et al. Ultrapotent human antibodies protect against SARS-CoV-2 challenge via multiple mechanisms. Science 2020, 370, 950–957. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; et al. Structural basis for potent antibody neutralization of SARS-CoV-2 variants including B.1.1.529. Science 2022, 376, eabn8897. [Google Scholar] [CrossRef] [PubMed]
- de Campos-Mata, L.; et al. A monoclonal antibody targeting a large surface of the receptor binding motif shows pan-neutralizing SARS-CoV-2 activity. Nat Commun 2024, 15, 1051. [Google Scholar] [CrossRef] [PubMed]
- Planchais, C.; et al. Potent human broadly SARS-CoV-2-neutralizing IgA and IgG antibodies effective against Omicron BA.1 and BA.2. J Exp Med 2022, 219. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; et al. Ultrapotent antibodies against diverse and highly transmissible SARS-CoV-2 variants. Science 2021, 373, eabh1766. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; et al. An ACE2-blocking antibody confers broad neutralization and protection against Omicron and other SARS-CoV-2 variants of concern. Sci Immunol 2022, 7, eabp9312. [Google Scholar] [CrossRef]
- Dong, J.; et al. Genetic and structural basis for SARS-CoV-2 variant neutralization by a two-antibody cocktail. Nat Microbiol 2021, 6, 1233–1244. [Google Scholar] [CrossRef] [PubMed]
- Zost, S.J.; et al. Potently neutralizing and protective human antibodies against SARS-CoV-2. Nature 2020, 584, 443–449. [Google Scholar] [CrossRef]
- Starr, T.N.; et al. SARS-CoV-2 RBD antibodies that maximize breadth and resistance to escape. Nature 2021, 597, 97–102. [Google Scholar] [CrossRef]
- Qu, P.; et al. Enhanced neutralization resistance of SARS-CoV-2 Omicron subvariants BQ.1, BQ.1.1, BA.4.6, BF.7, and BA.2.75.2. Cell Host Microbe 2023, 31, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.J.; et al. Antibody-mediated broad sarbecovirus neutralization through ACE2 molecular mimicry. Science 2022, 375, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Cameroni, E.; et al. Broadly neutralizing antibodies overcome SARS-CoV-2 Omicron antigenic shift. Nature 2022, 602, 664–670. [Google Scholar] [CrossRef]
- Rouet, R.; et al. Broadly neutralizing SARS-CoV-2 antibodies through epitope-based selection from convalescent patients. Nat Commun 2023, 14, 687. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; et al. Antibody neutralization of SARS-CoV-2 through ACE2 receptor mimicry. Nat Commun 2021, 12, 250. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; et al. Breakthrough infection elicits hypermutated IGHV3-53/3-66 public antibodies with broad and potent neutralizing activity against SARS-CoV-2 variants including the emerging EG.5 lineages. PLOS PATHOGENS 2023, 19. [Google Scholar] [CrossRef]
- Fenwick, C.; et al. Broadly potent anti-SARS-CoV-2 antibody shares 93% of epitope with ACE2 and provides full protection in monkeys. Journal of Infection 2023, 87, 524–537. [Google Scholar] [CrossRef]
- Ju, B.; et al. Infection with wild-type SARS-CoV-2 elicits broadly neutralizing and protective antibodies against omicron subvariants. Nat Immunol 2023, 24, 690–699. [Google Scholar] [CrossRef]
- Liu, Y.; et al. Inactivated vaccine-elicited potent antibodies can broadly neutralize SARS-CoV-2 circulating variants. Nat Commun 2023, 14, 2179. [Google Scholar] [CrossRef]
- Yuan, M.; et al. Structural and functional ramifications of antigenic drift in recent SARS-CoV-2 variants. Science 2021, 373, 818–823. [Google Scholar] [CrossRef]
- Jones, B.E.; et al. The neutralizing antibody, LY-CoV555, protects against SARS-CoV-2 infection in nonhuman primates. Sci Transl Med 2021, 13. [Google Scholar] [CrossRef]
- Chen, J.; et al. Rational optimization of a human neutralizing antibody of SARS-CoV-2. Comput Biol Med 2021, 135, 104550. [Google Scholar] [CrossRef] [PubMed]
- Ju, B.; et al. Human neutralizing antibodies elicited by SARS-CoV-2 infection. Nature 2020, 584, 115–119. [Google Scholar] [CrossRef]
- Patrick, C.; et al. Biophysical Fitness Landscape of the SARS-CoV-2 Delta Variant Receptor Binding Domain. J Mol Biol 2022, 434, 167622. [Google Scholar] [CrossRef]
- Makdasi, E.; et al. The neutralization potency of anti-SARS-CoV-2 therapeutic human monoclonal antibodies is retained against viral variants. Cell Rep 2021, 36, 109679. [Google Scholar] [CrossRef]
- Sun, H.; et al. Structural basis for broad neutralization of human antibody against Omicron sublineages and evasion by XBB variant. J Virol 2023, 97, e0113723. [Google Scholar] [CrossRef] [PubMed]
- Yue, C.; et al. ACE2 binding and antibody evasion in enhanced transmissibility of XBB.1.5. Lancet Infect Dis 2023, 23, 278–280. [Google Scholar] [CrossRef] [PubMed]
- Pinto, D.; et al. Cross-neutralization of SARS-CoV-2 by a human monoclonal SARS-CoV antibody. Nature 2020, 583, 290–295. [Google Scholar] [CrossRef]
- Gupta, A.; et al. Early Treatment for Covid-19 with SARS-CoV-2 Neutralizing Antibody Sotrovimab. New England Journal of Medicine 2021, 385, 1941–1950. [Google Scholar] [CrossRef]
- Magnus, C.L.; et al. Targeted escape of SARS-CoV-2 in vitro from monoclonal antibody S309, the precursor of sotrovimab. Frontiers in Immunology 2022, 13. [Google Scholar] [CrossRef]
- Luo, S.; et al. An antibody from single human VH-rearranging mouse neutralizes all SARS-CoV-2 variants through BA.5 by inhibiting membrane fusion. Science Immunology 2022, 7, eadd5446. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; et al. An antibody that neutralizes SARS-CoV-1 and SARS-CoV-2 by binding to a conserved spike epitope outside the receptor binding motif. Science Immunology 2022, 7, eabp9962. [Google Scholar] [CrossRef] [PubMed]
- Westendorf, K.; et al. LY-CoV1404 (bebtelovimab) potently neutralizes SARS-CoV-2 variants. Cell Rep 2022, 39, 110812. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-J.; et al. Imprinted antibody responses against SARS-CoV-2 Omicron sublineages. Science 2022, 378, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; et al. Structural insights for neutralization of Omicron variants BA.1, BA.2, BA.4, and BA.5 by a broadly neutralizing SARS-CoV-2 antibody. Science Advances 2022, 8, eadd2032. [Google Scholar] [CrossRef]
- Yuan, M.; et al. A highly conserved cryptic epitope in the receptor binding domains of SARS-CoV-2 and SARS-CoV. Science 2020, 368, 630–633. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; et al. A broad and potent neutralization epitope in SARS-related coronaviruses. Proc Natl Acad Sci U S A 2022, 119, e2205784119. [Google Scholar] [CrossRef]
- Tortorici, M.A.; et al. Broad sarbecovirus neutralization by a human monoclonal antibody. Nature 2021, 597, 103–108. [Google Scholar] [CrossRef]
- Huo, J.; et al. Neutralization of SARS-CoV-2 by Destruction of the Prefusion Spike. Cell Host & Microbe 2020, 28, 497. [Google Scholar]
- Sankhala, R.S.; et al. Antibody targeting of conserved sites of vulnerability on the SARS-CoV-2 spike receptor-binding domain. Structure 2024, 32, 131–147. [Google Scholar] [CrossRef]
- Liu, H.; et al. Cross-Neutralization of a SARS-CoV-2 Antibody to a Functionally Conserved Site Is Mediated by Avidity. Immunity 2020, 53, 1272–1280. [Google Scholar] [CrossRef]
- Li, D.; et al. In vitro and in vivo functions of SARS-CoV-2 infection-enhancing and neutralizing antibodies. Cell 2021, 184, 4203–4219. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; et al. An antibody class with a common CDRH3 motif broadly neutralizes sarbecoviruses. Science Translational Medicine 2022, 14, eabn6859. [Google Scholar] [CrossRef] [PubMed]
- Jensen, J.L.; et al. Targeting the Spike Receptor Binding Domain Class V Cryptic Epitope by an Antibody with Pan-Sarbecovirus Activity. J Virol 2023, 97, e0159622. [Google Scholar] [CrossRef]
- Wang, S.; et al. Three SARS-CoV-2 antibodies provide broad and synergistic neutralization against variants of concern, including Omicron. Cell Rep 2022, 39, 110862. [Google Scholar] [CrossRef] [PubMed]
- Bullen, G.; et al. Cross-Reactive SARS-CoV-2 Neutralizing Antibodies From Deep Mining of Early Patient Responses. Front Immunol 2021, 12, 678570. [Google Scholar] [CrossRef]
- Dussupt, V.; et al. Low-dose in vivo protection and neutralization across SARS-CoV-2 variants by monoclonal antibody combinations. Nature Immunology 2021, 22, 1503–1514. [Google Scholar] [CrossRef]
- Li, T.; et al. Uncovering a conserved vulnerability site in SARS-CoV-2 by a human antibody. EMBO Molecular Medicine 2021, 13, e14544. [Google Scholar] [CrossRef]
- Li, T.; et al. Cross-neutralizing antibodies bind a SARS-CoV-2 cryptic site and resist circulating variants. Nat Commun 2021, 12, 5652. [Google Scholar] [CrossRef]
- Olukitibi, T.A.; et al. Significance of Conserved Regions in Coronavirus Spike Protein for Developing a Novel Vaccine against SARS-CoV-2 Infection. Vaccines 2023, 11. [Google Scholar] [CrossRef]
- Shrock, E.; et al. Viral epitope profiling of COVID-19 patients reveals cross-reactivity and correlates of severity. Science 2020, 370, eabd4250. [Google Scholar] [CrossRef] [PubMed]
- Ladner, J.T.; et al. Epitope-resolved profiling of the SARS-CoV-2 antibody response identifies cross-reactivity with endemic human coronaviruses. Cell Reports Medicine 2021, 2, 100189. [Google Scholar] [CrossRef] [PubMed]
- Crowley, A.R.; et al. Boosting of Cross-Reactive Antibodies to Endemic Coronaviruses by SARS-CoV-2 Infection but not Vaccination with Stabilized Spike. medRxiv 2021. [Google Scholar]
- Pannus, P.; et al. Poor Antibody Response to BioNTech/Pfizer Coronavirus Disease 2019 Vaccination in Severe Acute Respiratory Syndrome Coronavirus 2-Naive Residents of Nursing Homes. Clin Infect Dis 2022, 75, e695–e704. [Google Scholar] [CrossRef] [PubMed]
- Piepenbrink, M.S.; et al. Potent universal beta-coronavirus therapeutic activity mediated by direct respiratory administration of a Spike S2 domain-specific human neutralizing monoclonal antibody. PLoS Pathog 2022, 18, e1010691. [Google Scholar] [CrossRef] [PubMed]
- Li, C.J.; Chang, S.C. . SARS-CoV-2 spike S2-specific neutralizing antibodies. Emerg Microbes Infect 2023, 12, 2220582. [Google Scholar] [CrossRef] [PubMed]
- Pinto, D.; et al. Broad betacoronavirus neutralization by a stem helix–specific human antibody. Science 2021, 373, 1109–1116. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; et al. Vaccine-elicited murine antibody WS6 neutralizes diverse beta-coronaviruses by recognizing a helical stem supersite of vulnerability. Structure 2022, 30, 1233–1244. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; et al. A human antibody reveals a conserved site on beta-coronavirus spike proteins and confers protection against SARS-CoV-2 infection. Science Translational Medicine 2022, 14, eabi9215. [Google Scholar] [CrossRef]
- Hurlburt, N.K.; et al. Structural definition of a pan-sarbecovirus neutralizing epitope on the spike S2 subunit. Communications Biology 2022, 5, 342. [Google Scholar] [CrossRef]
- Jennewein, M.F.; et al. Isolation and characterization of cross-neutralizing coronavirus antibodies from COVID-19+ subjects. Cell Reports 2021, 36, 109353. [Google Scholar] [CrossRef] [PubMed]
- Li, C.J.; et al. Neutralizing Monoclonal Antibodies Inhibit SARS-CoV-2 Infection through Blocking Membrane Fusion. Microbiol Spectr 2022, 10, e0181421. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; et al. Neutralization mechanism of a human antibody with pan-coronavirus reactivity including SARS-CoV-2. Nature Microbiology 2022, 7, 1063–1074. [Google Scholar] [CrossRef] [PubMed]
- Low, J.S.; et al. ACE2-binding exposes the SARS-CoV-2 fusion peptide to broadly neutralizing coronavirus antibodies. Science 2022, 377, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Dacon, C.; et al. Broadly neutralizing antibodies target the coronavirus fusion peptide. Science 2022, 377, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.L.; et al. Monoclonal antibody targeting the conserved region of the SARS-CoV-2 spike protein to overcome viral variants. JCI Insight 2022, 7. [Google Scholar] [CrossRef] [PubMed]
- Piccoli, L.; et al. Mapping Neutralizing and Immunodominant Sites on the SARS-CoV-2 Spike Receptor-Binding Domain by Structure-Guided High-Resolution Serology. Cell 2020, 183, 1024–1042. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.C.; et al. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292. [Google Scholar] [CrossRef]
- Chi, X.; et al. A neutralizing human antibody binds to the N-terminal domain of the Spike protein of SARS-CoV-2. Science 2020, 369, 650–655. [Google Scholar] [CrossRef]
- Cerutti, G.; et al. Potent SARS-CoV-2 neutralizing antibodies directed against spike N-terminal domain target a single supersite. Cell Host & Microbe 2021, 29, 819–833. [Google Scholar]
- Souza, P.F.N.; et al. The spike glycoprotein of SARS-CoV-2: A review of how mutations of spike glycoproteins have driven the emergence of variants with high transmissibility and immune escape. Int J Biol Macromol 2022, 208, 105–125. [Google Scholar] [CrossRef]
- Lok, S.-M. An NTD supersite of attack. Cell Host & Microbe 2021, 29, 744–746. [Google Scholar]
- Liu, L.; et al. Potent neutralizing antibodies against multiple epitopes on SARS-CoV-2 spike. Nature 2020, 584, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, G.; et al. Neutralizing antibody 5-7 defines a distinct site of vulnerability in SARS-CoV-2 spike N-terminal domain. Cell Reports 2021, 37, 109928. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; et al. SARS-CoV-2 Neutralizing Antibodies for COVID-19 Prevention and Treatment. Annual Review of Medicine 2022, 73, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; et al. Imprinted SARS-CoV-2 humoral immunity induces convergent Omicron RBD evolution. Nature 2023, 614, 521–529. [Google Scholar] [CrossRef]
- Casadevall, A.; Focosi, D. SARS-CoV-2 variants resistant to monoclonal antibodies in immunocompromised patients constitute a public health concern. J Clin Invest 2023, 133. [Google Scholar] [CrossRef]
- Levin, M.J.; et al. LB5. PROVENT: Phase 3 Study of Efficacy and Safety of AZD7442 (Tixagevimab/Cilgavimab) for Pre-exposure Prophylaxis of COVID-19 in Adults. Open Forum Infectious Diseases 2021, 8 (Supplement_1), S810–S810. [Google Scholar] [CrossRef]
Figure 1.
(A) Schematic representation of SARS-COV-2 spike protein, highlighting key domains including N-terminal domain (NTD), receptor-binding domain (RBD), fusion peptide (FP), heptad repeat 1 (HR1), heptad repeat 2 (HR2), transmembrane domain (TM), intracellular domain (IC). (B) Structure of SARS-COV-2 virus. S, spike protein; M, membrane protein; E, envelope protein; N, nucleocapsid protein. (C) The process of viral entry mediated by the SARS-CoV-2 spike protein.
Figure 1.
(A) Schematic representation of SARS-COV-2 spike protein, highlighting key domains including N-terminal domain (NTD), receptor-binding domain (RBD), fusion peptide (FP), heptad repeat 1 (HR1), heptad repeat 2 (HR2), transmembrane domain (TM), intracellular domain (IC). (B) Structure of SARS-COV-2 virus. S, spike protein; M, membrane protein; E, envelope protein; N, nucleocapsid protein. (C) The process of viral entry mediated by the SARS-CoV-2 spike protein.
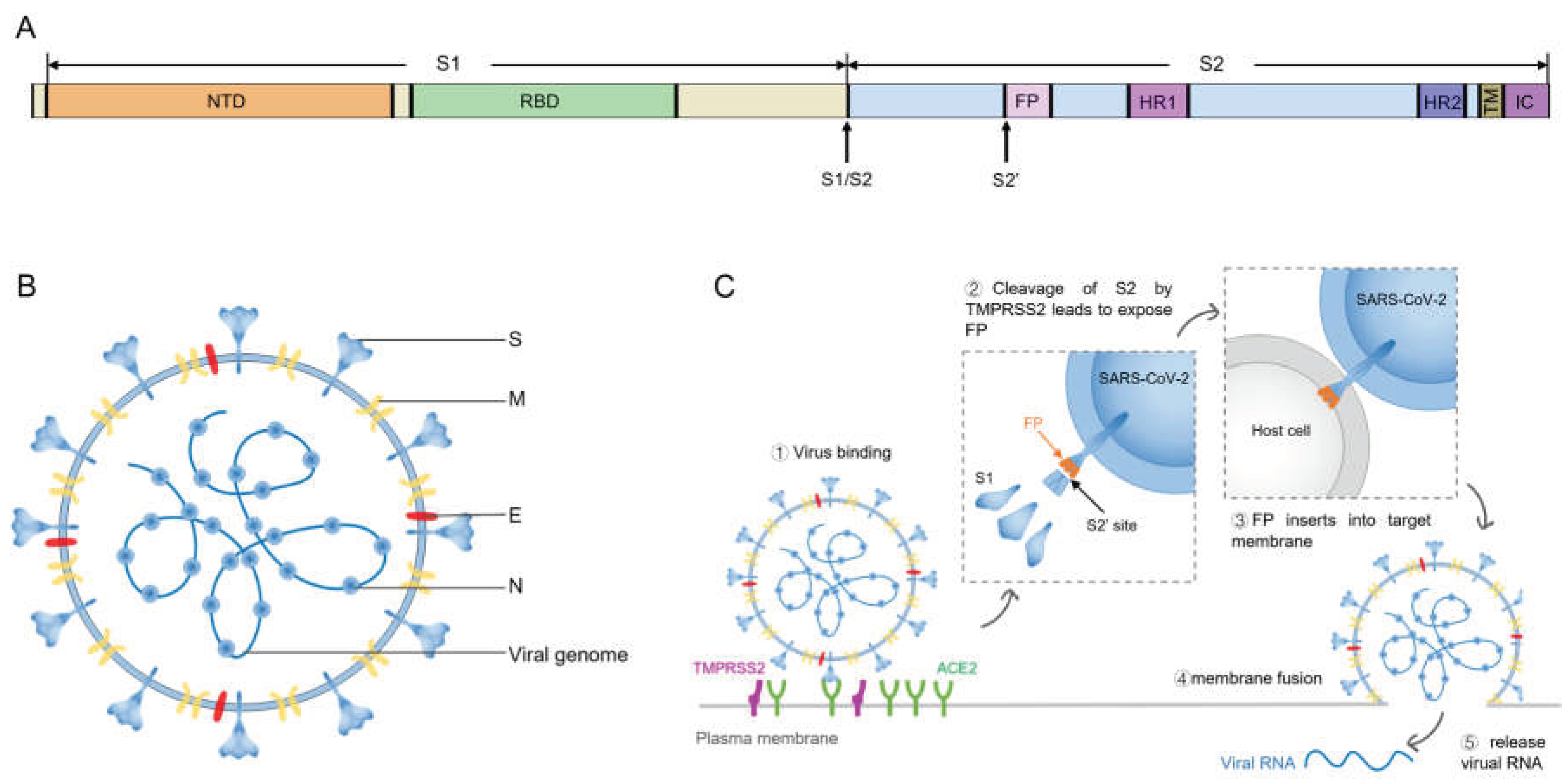
Figure 2.
Spike structure and epitopes of class 1-5 RBD antibodies. (A) The spike trimer (PDB: 7DX1) is shown using a bimodal representation. Two of the protomers are displayed with their molecular surfaces in dark and grey, the third protomer is illustrated as a ribbon. (B) Overlay of RBD structures (light grey) alongside Class 1-5 representative antibodies, S2E12(PDB: 7R6X), P2B-2F6(PDB: 8DCC), S309(PDB: 7R6X), CR3022(PDB: 8FAH), S2H97(PDB: 7M7W), showcasing distinct binding orientations. The receptor-binding motif (RBM) is highlighted by rosy brown. (C)-(G) The localization of class 1-5 representative antibodies on the epitopes within RBD.
Figure 2.
Spike structure and epitopes of class 1-5 RBD antibodies. (A) The spike trimer (PDB: 7DX1) is shown using a bimodal representation. Two of the protomers are displayed with their molecular surfaces in dark and grey, the third protomer is illustrated as a ribbon. (B) Overlay of RBD structures (light grey) alongside Class 1-5 representative antibodies, S2E12(PDB: 7R6X), P2B-2F6(PDB: 8DCC), S309(PDB: 7R6X), CR3022(PDB: 8FAH), S2H97(PDB: 7M7W), showcasing distinct binding orientations. The receptor-binding motif (RBM) is highlighted by rosy brown. (C)-(G) The localization of class 1-5 representative antibodies on the epitopes within RBD.
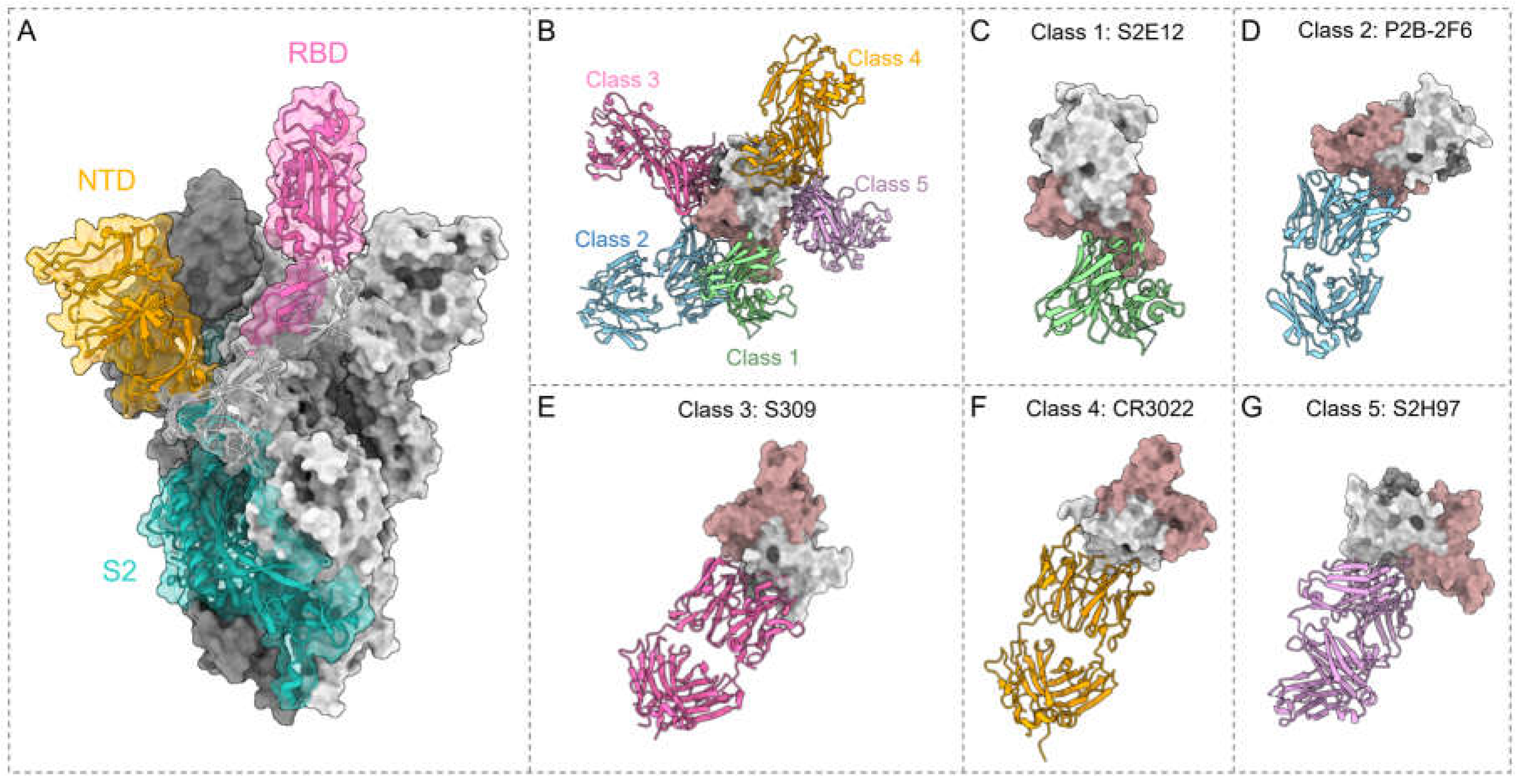

Table 1.
Mutations table in the S protein of SARS-CoV-2 variants[8]. (https://outbreak.info/situation-reports).
Table 1.
Mutations table in the S protein of SARS-CoV-2 variants[8]. (https://outbreak.info/situation-reports).
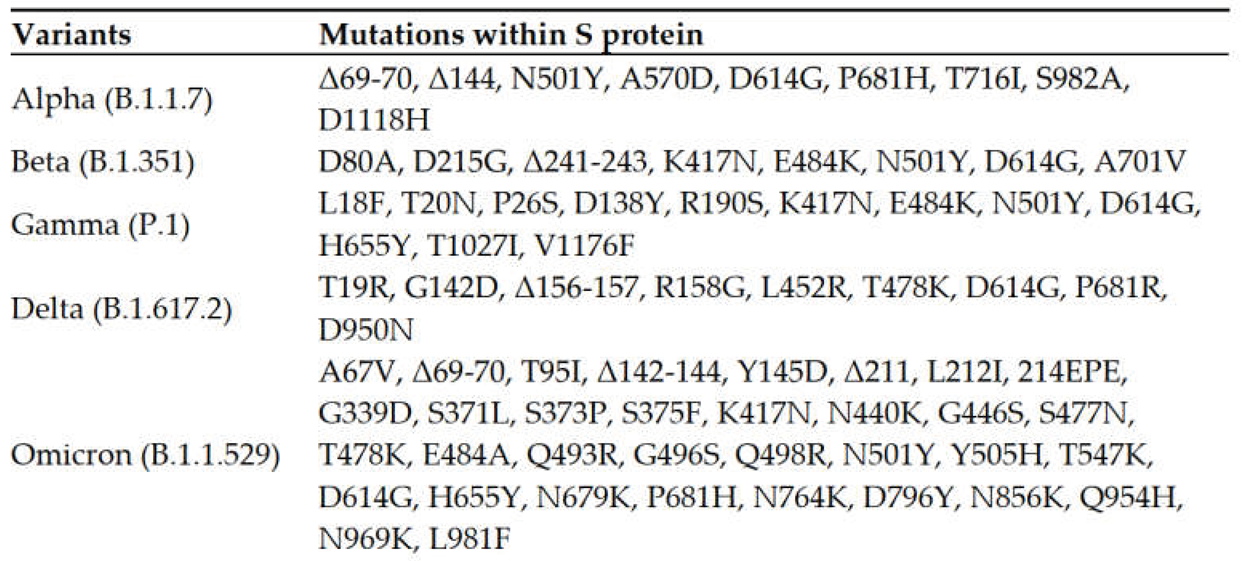

Table 2.
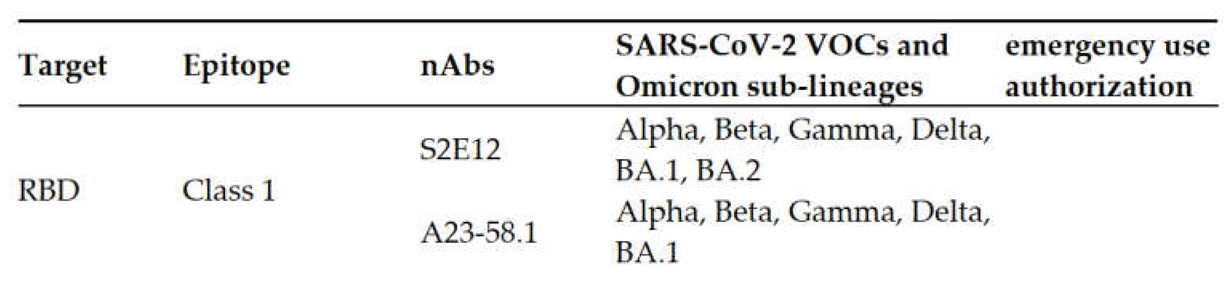
Neutralizing antibodies with potent neutralizing effectiveness against SARS-CoV-2 variants.
Table 2.
Neutralizing antibodies with potent neutralizing effectiveness against SARS-CoV-2 variants.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



