
Submitted:
04 April 2024
Posted:
07 April 2024
You are already at the latest version
Abstract
An imbalance between production and excretion of amyloid β peptide (Aβ) in the brain tissues of Alzheimer’s disease (AD) patients leads to Aβ accumulation and formation of noxious Aβ oligomers/plaques. A promising approach to AD prevention is reduction of free Aβ level by directed enhancement of Aβ binding to its natural depot, human serum albumin (HSA). We previously demonstrated the ability of specific low-molecular-weight ligands (LMWLs) of HSA to improve its affinity for Aβ. Here we develop this approach by bioinformatic search for the clinically approved AD-related LMWLs of HSA, followed by classification of the candidates according to the predicted location of their binding sites on HSA surface, ranking of the candidates, and selective experimental validation of their impact on HSA affinity for Aβ. The top 100 candidate LMWLs were classified into the five clusters. The specific representatives of the different clusters exhibit dramatically different behavior, with 3- to 13-fold changes in equilibrium dissociation constants for the HSA-Aβ40 interaction: prednisone favors HSA-Aβ interaction, mefenamic acid shows the opposite effect, while levothyroxine exhibits the bidirectional effects. Overall, the LMWLs of HSA chosen here provide a basis for drug repurposing for AD prevention, and for search for the medications promoting AD progression.
Keywords:
Alzheimer’s disease
; amyloid β peptide
; human serum albumin
; low-molecular-weight ligand
; protein-ligand interaction
; surface plasmon resonance spectroscopy
1. Introduction
Human serum albumin (HSA) is the predominant protein in blood plasma, accounting for approximately 60% of the total protein content [1]. HSA is a 66 kDa protein, containing 585 amino acid residues organized into three domains [2]. It possesses seven fatty acid (FA) binding sites and the two major sites specific to small molecules [3,4]. HSA exhibits a remarkable ability to bind and transport through the bloodstream numerous endogenous and exogenous ligands [5,6]. Thereby HSA serves as a depot and delivery vehicle for small molecules in the blood, such as FAs, hormones, bilirubin, hemin, drugs, etc. [6,7,8,9]. The long plasma half-life of HSA (12.7-18.2 days [2]) is widely used to prolong the half-life of therapeutic peptides/proteins by their covalent modification with FAs or other substances with high affinity for HSA [10]. Some of them are used for treatment of diabetes mellitus (type 1 and 2) and/or obesity [11,12]: insulin detemir (Levemir®), insulin degludec (Tresiba®), liraglutide (Victoza®/Saxenda®), and semaglutide (Ozempic®, Rybelsus®, Wegovy®).
HSA is a natural depot for amyloid β peptide (Aβ) [13,14], one of the key factors in the development of Alzheimer's disease (AD) [15,16]. HSA binds about 90% of Aβ in blood serum [17] and, according to various estimates, from 40% to 94% in cerebrospinal fluid [18]. HSA inhibits Aβ aggregation and lowers the risk of AD and its progression [19,20,21,22,23]. As a component of the interstitial fluid, HSA is presented in the intercellular space of the brain parenchyma [24]. Although HSA is primarily synthesized in the liver, brain microglial cells also synthesize and secrete HSA, especially upon their stimulation with Aβ or lipopolysaccharide [25]. Finally, HSA is included in amyloid deposits (plaques) in the brain of AD patients [26]. These facts indicate significance of HSA in Aβ metabolism and potential of HSA usage for therapy of AD.
Preliminary clinical trials confirm the effectiveness of AD treatment by replacement of the patient's serum albumin with its pharmacological preparation via plasmapheresis (plasma exchange (PE)) [19,23]. This approach is related with a risk of cardiovascular and respiratory complications, anaphylactoid reactions, infections, and hemorrhage [27]. The necessity to live with a catheter inserted in the chest and metabolic alterations related to PE can trigger psychiatric symptoms in the PE-treated AD patients [23]. Nevertheless, this approach is considered relatively safe, but requires participation of the highly trained medical personnel [23,28,29].
The potentially less harmful approach is to increase HSA affinity for Aβ via allosteric action of endogenous or exogenous HSA ligands, initially demonstrated for linoleic and arachidonic acids [30]. We further showed the even more pronounced effects for serotonin [31] and ibuprofen [32]. Moreover, ibuprofen enhances HSA ability to inhibit Aβ fibrillation [32]. These data are in line with clinical observations and the results derived from animal models of AD [33,34,35].
Despite the first encouraging results, a systematic search of the HSA ligands that can affect its interaction with Aβ has not been yet been carried out. To fill this gap, in the present work, we systematically search for the clinically approved low-molecular-weight HSA ligands related to AD progression that could modify HSA affinity for Aβ, and selectively test the candidate ligands from this perspective after their careful structural systematization and ranking.
2. Material and Methods
2.1. Materials
Recombinant human Aβ40 was expressed in E. coli and purified as previously described [32]. Catalytic core of ubiquitin carboxyl-terminal hydrolase 2 (Usp2-cc) was prepared mainly as described in ref. [36]. FA-free HSA prepared under non-denaturing conditions [37] was from Merck (cat. #126654, Darmstadt, Germany). Protein concentrations were measured spectrophotometrically at pH 7.4–8.0 and calculated using molar extinction coefficients at 280 nm estimated according to ref. [38]: 34,445 M−1cm−1 for HSA, 41,370 M−1cm−1 for Usp2-cc and 1,490 M−1cm−1 for Aβ40.
Propranolol (cat. #P913470), levothyroxine (L-thyroxine) (J62606.03), prednisone (PHR1042), warfarin (A2250), and mefenamic acid (M4267) were at least 98% pure and purchased from Macklin, Thermo Fisher Scientific (Waltham, Massachusetts, U.S.) and Sigma-Aldrich (Burlington, Massachusetts, U.S.) (for the last three compounds), respectively. Ultra-grade Tris and 2-mercaptoethanol (2-ME) were purchased from Amresco® LLC (Vienna, Austria). Urea, imidazole, sodium chloride, sodium hydroxide, sodium dodecyl sulfate (SDS), DL-dithiothreitol (DTT), and glycerol were from Panreac AppliChem (Darmstadt, Germany). Ethylenediaminetetraacetic acid (EDTA), acetonitrile, N-Ethyl-N′-(3-dimethylaminopropyl)carbodiimide hydrochloride (EDAC), N-hydroxysulfosuccinimide sodium salt (sulfo-NHS), and polyethylene glycol sorbitan monolaurate (TWEEN®) 20 were from Sigma-Aldrich (St. Louis, MO, USA). Calcium chloride was from Fluka (Buchs, Switzerland). Ethanolamine and ProfinityTM IMAC resin were bought from Bio-Rad Laboratories (Hercules, USA). Hydrochloric acid was from Sigma Tec LLC (Moscow, Russia). Dimethyl sulfoxide (DMSO) was from Helicon (Moscow, Russia). Trifluoroacetic acid (TFA) was purchased from Fisher Scientific (Pittsburgh, Pennsylvania, U.S.). Potassium chloride and sodium azide were from Dia-M (Moscow, Russia). Acetic acid and ammonium hydroxide were from Chimmed and Component-reaktiv (Moscow, Russia).
2.2. Bioinformatic Selection and Structural Analysis of the Therapeutic Low-Molecular-Weight HSA Ligands Associated with AD
The general workflow used for selection of the clinically approved LMWL of HSA related to AD and for their structural analysis is shown in Figure 1.
The list of the substances directly related to HSA (UniProt [39] (https://www.uniprot.org, accessed on 14 March 2024) ID P02768) was extracted from DrugBank (https://go.drugbank.com/bio_entities/BE0000530, accessed on 10 June 2023), a database containing chemical, pharmacological and pharmaceutical data on the clinically approved endogenous and exogenous substances [40,41]. The compounds with molecular mass (average mass, field “Structure”), above 900 Da and less than 100 Da [42] were excluded from consideration. The resulting low-molecular-weight ligands (LMWLs) of HSA were filtered according to the following requirements:
- water solubility of the ligand (Experimental Water Solubility / Calculated Water Solubility, field “Properties” of DrugBank) should exceed 1 μM to ensure the possibility of efficient HSA loading with the ligand;
- blood-brain barrier (BBB) penetration of the ligand (the field “Predicted ADMET Features” of DrugBank) should exceed 50% to ensure its efficient transfer from the bloodstream into the brain;
- plasma ligand concentration (manually collected from Pubmed (https://pubmed.ncbi.nlm.nih.gov, accessed on 14 March 2024) and DrugBank (field “Absorption”) should exceed 0.5 nM, which corresponds to the total plasma Aβ40 concentration [20].
The FAs were excluded from further consideration, since they had been studied in the previous works [30].
The association of the resulting LMWLs with AD was examined for each of them using the query «“Alzheimer's disease” + “substance name”» in the Alzforum online resource (https://www.alzforum.org/papers, accessed on 10 October 2023). The substances with less than 2 references on Alzforum were excluded from the further analysis, resulting in a total number of the candidates of 100.
To classify the selected LMWLs according to expected location of their binding sites on HSA surface, molecular docking of HSA and the ligands was performed using AutoDock Vina [43] (https://vina.scripps.edu, accessed on 14 March 2024). The crystal structure of HSA was taken from Protein Data Bank (PDB) [44] (https://www.rcsb.org, accessed on 14 March 2024): chain A of entry 1UOR. AutoDockTools software (https://autodocksuite.scripps.edu/adt/, accessed on 14 March 2024) was used for preparation of the PDB structure for the docking process, including removal of water molecules and addition of lacking hydrogen atoms. The three-dimensional structures of the LMWLs were taken from PubChem server (https://www.ncbi.nlm.nih.gov/pccompound, accessed on 14 March 2024) in a structure-data format (filename extension .sdf) and converted to PDB format using PyMOL (https://pymol.org/, accessed on 14 March 2024). In the case of stereoisomers, the biologically active (predominantly R) isomer was used for the modeling (for ibuprofen and for warfarin the more active (S)-isomer and the both enantiomers were used). The docking model corresponding to the lowest energy of the HSA-ligand complex was chosen. The protein-ligand complexes were visualized using PyMOL v.1.6. The HSA-ligand interactions were analyzed using the protein-ligand interaction profiler PLIP [45] (https://plip-tool.biotec.tu-dresden.de/plip-web/plip/index, accessed on 14 March 2024). The numbering of the amino acid residues corresponds to PDB entry 1UOR.
Dynamic time warping algorithms implemented in the ‘dtw’ library [46] written in R language (https://www.r-project.org/, accessed on 14 March 2024) were used to hierarchically cluster the AD-associated LMWLs based on the predicted location of their binding sites on HSA molecules. The rank of a ligand in a cluster, R, was calculated for all clusters using the Equation (1):
where faa is a frequency of occurrence of an amino acid in the binding site within the cluster, calculated as a number of ligands of the cluster whose binding site contains this amino acid; a is a number of amino acids of the cluster, calculated as a sum of all non-repetitive amino acids forming binding sites for all ligands of the cluster; n is a number of ligands in the cluster; k is a coefficient equal to 0 if the amino acid is absent in the binding site, and equal to 1 if the amino acid is present in the binding site.
The aforementioned algorithms for search, collection, alignment, representation and analysis of the data were implemented using the Python 3 (https://www.python.org/, accessed on 14 March 2024) programming language in PyCharm v.2018 environment (https://www.jetbrains.com/pycharm/, accessed on 14 March 2024). The specialized Python libraries (Requests, BeautifulSoup) were used to form HTTP requests and parse the web pages, search and collect the data into a local database.
2.3. Preparation of Recombinant Aβ
The human Aβ40 samples were pretreated prior to experimental studies essentially as described in ref. [32]. The freeze-dried Aβ40 samples were dissolved in neat TFA at a concentration of 0.5 mg/mL, followed by sonication for 30 s and TFA evaporation using an Eppendorf Concentrator plus. The dried Aβ40 samples were dissolved in DMSO at a concentration of 2 mg/mL and stored at -20 °C.
2.4. Solubility of HSA Ligands
Solubility of propranolol, prednisone, warfarin, levothyroxine, and mefenamic acid in 20 mM HEPES(Tris)-HCl, 150 mM NaCl, pH 7.4 buffer was determined by sequential addition of the ligand with constant stirring until appearance of the precipitate. DMSO (for prednisone, levothyroxine and propranolol) or ethanol (for warfarin and mefenamic acid) was added to the solution at a concentration of up to 4-5% (v/v) to improve solubility of the ligand.
2.5. Surface Plasmon Resonance Studies
Surface plasmon resonance (SPR) measurements of HSA interaction with monomeric Aβ40 were performed at 25 °C using a Bio-Rad ProteOn™ XPR36 instrument mainly according to ref. [30]. Ligand (50 µg/mL Aβ40 in 10 mM sodium acetate, pH 4.5 buffer) was immobilized on a ProteOn GLH sensor chip surface by amine coupling up to 10,000–14,000 resonance units, RUs. The remaining activated amine groups on the chip surface were blocked by 1 M ethanolamine solution. The noncovalently bound Aβ40 molecules were washed off the chip surface with a 0.5% SDS water solution until stabilization of the SPR signal. Analyte (2.5-40 µM HSA) in the running buffer (20 mM Tris-HCl, 150 mM NaCl, pH 7.4) in the presence/absence of prednisone (2.5 mM)/warfarin (1 mM)/mefenamic acid (250 µM)/levothyroxine (15 µM)/propranolol (1 mM) was passed over the chip at a rate of 30 µL/min for 300 s, followed by flushing the chip with the running buffer for 2,400 s. The sensor chip surface was regenerated by passage of 0.5% SDS water solution for 100 s. The kinetic and equilibrium association/dissociation constants for the HSA-Aβ40 interaction in the absence of the ligands were determined with or without addition of 5% DMSO/ethanol.
The kinetic SPR data were corrected for baseline drift and non-specific binding, and described using a heterogeneous ligand model (Equation (2)) (Aβ40 and HSA serve as a ligand (L) and an analyte (A), respectively):
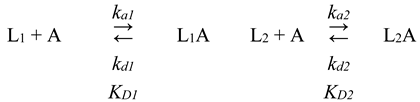 where ka and kd are kinetic association and dissociation constants, respectively; KD are equilibrium dissociation constants. The ka, kd and KD values were estimated using Bio-Rad ProteOn Manager™ v.3.1 software (Bio-Rad Laboratories, Inc.). The estimates were performed for each analyte concentrations, followed by their averaging (standard deviations are indicated). The free energy change accompanying HSA-Aβ40 interaction (ΔG) was calculated as follows: ΔGi = – RT ln(55.3/KDi), i=1,2.
where ka and kd are kinetic association and dissociation constants, respectively; KD are equilibrium dissociation constants. The ka, kd and KD values were estimated using Bio-Rad ProteOn Manager™ v.3.1 software (Bio-Rad Laboratories, Inc.). The estimates were performed for each analyte concentrations, followed by their averaging (standard deviations are indicated). The free energy change accompanying HSA-Aβ40 interaction (ΔG) was calculated as follows: ΔGi = – RT ln(55.3/KDi), i=1,2.
3. Results
3.1. Bioinformatic Selection of the Therapeutic Low-Molecular-Weight HSA Ligands Associated with AD
DrugBank database [40,41] was used as a source of the clinically approved HSA ligands related to AD. For each of the HSA ligands we have collected a set of the characteristics necessary for their further filtering: molecular mass values of experimental and theoretical water solubility, BBB penetration. Information on plasma concentrations of the substances was manually collected from Pubmed and DrugBank. The association of the substances with AD was assessed by the number of the relevant literature sources found on the Alzforum online resource. The filtering of the candidates by a set of parameters: molecular mass above 100 Da and less than 900 Da, water solubility above 1 μM, predicted BBB penetration exceeding 50%, plasma concentration above 0.5 nM, and more than 1 reference on Alzforum, resulted in 100 LMWLs of HSA associated with AD (Table S1), after removal of the FAs studied in our previous works [9,30,47,48]. The candidate compounds belong to different drug classes, including such common ones as antidiabetic drugs (rosiglitazone), non-steroidal anti-inflammatory drugs (ibuprofen, meloxicam), neuroleptics (risperidone), vitamins (vitamin A, thiamine), antibiotics (tetracycline, ampicillin), and hormones (testosterone, estradiol).
3.2. Classification and Ranking of the Selected LMWLs According to the Expected Location of Their Binding Sites on HSA
To predict location of the binding sites for the 100 AD-related LMWLs on HSA surface, three-dimensional structures of the HSA-LMWL complexes were built using Auto Dock Vina [43]. Note that the binding sites were previously experimentally localized by X-RAY only for a few members of our panel, namely ibuprofen [3], warfarin [3], indomethacin [3], halothane [49], propofol [49]. The binding sites predicted for these ligands coincide with those localized experimentally for all ligands, except for ibuprofen (the predicted site is located nearby the secondary binding site). The resulting binding sites were hierarchically classified into five clusters based on the dynamic time warping algorithms as implemented in the R language library ‘dtw’, used for classification and clustering of general number series (Table S2) [46]. Figure 2 illustrates location of the key residues constituting these clusters on HSA molecule. The cluster I is located between HSA subdomains IIA and IIB and is predicted to be specific for the 34 AD-associated LMWLs. The clusters II (subdomain IIIA), III (subdomain IB) and V (subdomain IIA) are predicted to bind 24, 13 and 28 ligands, respectively. Levothyroxine (DB00451) was attributed to a distinct cluster IV located between the cleft and subdomain IB.
The amino acid residues most commonly involved in recognition of the ligands (the residues with highest faa values, see Equation (1), Figure 2):
- Cluster I: P447 (faa = 84.4%), R222 (78.1%), K444 (62.5%), D451 (59.4 %), E292 (46.9%);
- Cluster II: Y452 (faa = 40.5%), K436 (32.4%), K195 (32.4%), A191 (27.0%) and K432 (27.0%);
- Cluster III: R117 (faa = 39.0%), E141 (34.8%), R145 (30.4%), E86, P118 (26%) and Y140 (26%);
- Cluster IV: R145, E425, N109, Q459, H146, R114, K525, R186;
- Cluster V: E292 (faa = 55.6%), R257 (47.2%), K199 (44.4%), R218 (41.7%), L219 (38.9%).
To select the most representative ligands in each cluster, the ranks of the ligands within each cluster (R) were calculated according to Equation (1), followed by choice of the ligands with high R values (Figure 2). The resulting ligands are most often recognized by the aforementioned residues, thereby serving as the most representative candidates for further experimental examination of their influence on HSA-Aβ interaction. Among them, only one candidate from each cluster was taken for further consideration, with an emphasis on the well-studied, socially significant drugs (Table 1).
3.3. Experimental Validation of the Ability of the Top-Ranked AD-Related LMWLs to Affect HSA Affinity for Aβ40
The effect on HSA interaction with Aβ40 of the following selected representatives of the clusters I-V was studied using SPR spectroscopy: prednisone (cluster I), warfarin (cluster II), mefenamic acid (cluster III), levothyroxine (cluster IV), propranolol (cluster V). The ligand concentrations were chosen to ensure their efficient binding to HSA, based on the literature data on HSA affinity for the ligands (Table 1). With the exception of levothyroxine, the estimated occupancy of HSA binding sites exceeds 70%. Due to the low solubility of levothyroxine in the assay buffers (15 μM), the achievable occupancy of HSA binding sites ranges from 9% to 41%. The resulting SPR sensograms for 20/40 μM HSA and their description using the heterogeneous ligand model (Equation (2)) are shown in Figure 3. The respective averaged kinetic and equilibrium dissociation/association constants are represented in Table 2. The most pronounced ligand-binding induced changes in HSA affinity for Aβ40 in the presence of ethanol are observed for mefenamic acid with an increase in the KD values by a factor of 3-4. This effect is clearly seen in the scale of free energy changes accompanying the HSA-Aβ40 interaction (Figure 4). Similarly, the most marked changes in the presence of DMSO are observed for levothyroxine (3-fold increase in the KD1 value and 12-fold decrease in the KD2) and prednisone (decrease in the KD2 value by a factor of 13) (see Figure 4, Table 2).
Taken together, the SPR data demonstrate the ability of some representative members of the specific clusters (namely, I, III and IV) to affect HSA affinity for Aβ40. Meanwhile, one should expect various ligand- and cluster-specific effects, as exemplified by ibuprofen (cluster II [32]) and serotonin (cluster V [31]).
4. Discussion
The approved medications are a valuable source of the substances potentially suited for therapy of both common and rare diseases within the approach known as drug repurposing or repositioning [56]. For example, tetracyclines and some polyphenols are able to interfere with aggregation of several unrelated amyloidogenic proteins, such as α-synuclein (associated with Parkinson’s disease), amyloid polypeptide (type-2 diabetes), and transthyretin (senile systemic amyloidosis, familial amyloid polyneuropathy and cardiomyopathy) [57,58,59,60]. Similarly, some of the approved drugs have the potential to worsen certain disorders. In the present work, we systematically searched for the approved medications able to affect specificity to Aβ of HSA as a major natural depot for Aβ in the blood/CSF [18], with a focus on the HSA ligands associated with AD. In our previous works we identified several such HSA ligands [30,31,32], but here we used a general bioinformatic approach that allowed us to get a panel of 100 drug candidates (Table S1). They were grouped into five clusters according to the predicted location of their binding sites on HSA molecule, followed by ranking of the candidates within each cluster (Table S2). The massive involvement of polar and charged amino acid residues in the predicted binding sites of the ligands on HSA (Table S2) indicates potential dependence of the ligand-HSA interactions on pH and ionic strength of a solution.
The developed panel of 100 LMWLs of HSA represents a valuable source of the approved drugs with the potential to affect AD progression due to their ability to modulate HSA-Aβ interaction. To probe this suggestion, one representative member from each cluster was selected (Table 1) for experimental validation of their ability to affect Aβ40 binding to HSA, focusing on their outstanding clinical value: prednisone, warfarin, levothyroxine, propranolol are included in the 100 most commonly prescribed medications in USA according to the ClinCalc DrugStats Database (https://clincalc.com/DrugStats/Top300Drugs.aspx, according to data for 2021; accessed on 14 March 2024). The clinical data (Table 3) [61,62,63,64,65,66,67], as well as the results derived from animal models and in vitro studies [68,69,70], evidence an association of the selected drugs with AD. Meanwhile, the influence of the candidates on Aβ metabolism and HSA interaction with Aβ has not been reported to date.
The clusters III and IV overlap with the Aβ-binding site predicted for HSA [32] and confirmed in ref. [71] (the region between domains IB and IIIB), while the remaining clusters are more or less close to it. Therefore, the ligands belonging to these clusters may directly or allosterically affect HSA-Aβ40 interaction. This suggestion is supported by the data presented in Table 3: several members of the different clusters exert different effects on HSA affinity to Aβ. For instance, mefenamic acid (cluster III) favors dissociation of HSA-Aβ40 complex, while prednisone (cluster I) promotes this interaction. Furthermore, the effects for ligands within the same cluster may be opposite, as exemplified by prednisone and risperidone.
We have previously shown that ibuprofen [32] and serotonin [31] favor HSA interaction with Aβ (Table 3). The analogous effect is shown here for prednisone (Table 2), indicating its potential value in prevention of AD onset. Meanwhile, in a randomized, placebo-controlled multicenter trial low-dose prednisone did not show behavioral improvements compared with the placebo group [72]. At the same time, the use of high-doze intrathecal corticosteroids has been proposed as a promising approach to AD prevention [62]. On the contrary, the substances that prevent HSA interaction with Aβ, such as mefenamic acid and risperidone (Table 3), should be considered as potentially harmful with regard to stimulation of AD progression. Apparently, in vitro and clinical studies are needed to establish the relevance for AD in these cases. In any case, identification of the drugs that affect HSA-Aβ interaction is important for further studies of their impact on AD, regardless of the direction of the effect.
5. Conclusions
Our previous works have revealed the precious feature of the specific HSA ligands, such as ibuprofen [32] and serotonin [31], to improve HSA affinity for Aβ, which may contribute to prevention of AD. Here we extended search for the substances with a similar effect to all approved drugs. The careful filtering of the DrugBank according to several rational criteria, followed by selection of the candidates relevant to AD, gave rise to a panel of 100 top-ranked LMWLs. Although molecular docking studies enabled their classification depending on location of their binding sites on HSA molecule, the SPR data do not reveal clear regularities regarding the ability of the ligands belonging to different clusters to affect HSA-Aβ interaction. Thus, all 100 candidate ligands are of potential value in this respect.
Among the LMWLs studied in this work, prednisone has the most pronounced favorable effect on HSA affinity for monomeric Aβ, with a 13-fold decrease in the equilibrium dissociation constant. Meanwhile, HSA prevents Aβ fibrillation [20] via binding not only monomeric Aβ, but also its oligomeric forms and protofibrils [76,77,78], indicating the need to verify ability of the candidate ligands to favor the inhibition of Aβ fibrillation by HSA. The ability of LMWLs to modulate the HSA effects on the Ab fibril formation process has been previously shown for warfarin, palmitic acid, cholesterol, and ibuprofen [32,79]. Moreover, some of the HSA ligands may also inhibit Aβ fibrillation, as shown for serotonin [80]. Thus, search for the optimal therapeutic ligand involves extensive experimental studies and filtering of the candidates in accord with the abovementioned requirements. However, the present work provides a basis for these studies aimed at repurposing of the approved drugs for prevention and treatment of AD. Additional clinical trials should be performed for the ligands that prevent HSA-Aβ interaction (for instance, mefenamic acid and risperidone) to rule out the possibility of stimulation of Aβ depositions.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Figure S1: Scheme of the hierarchical classification of the AD-related low-molecular-weight HSA ligands according to the predicted location of their binding sites on HSA molecule; Table S1: List of the therapeutic low-molecular-weight HSA ligands associated with AD; Table S2: List of the AD-related low-molecular-weight HSA ligands, hierarchically classified according to the predicted location of their binding sites on HSA molecule.
Author Contributions
Conceptualization, E.I.D., E.A.L. and M.P.S.; methodology, E.A.L., E.I.D., M.P.S., A.V.M., V.A.R., A.A.V., M.E.P.; software, E.I.D., A.V.M.; validation, E.I.D., E.A.L. and M.P.S.; formal analysis, E.I.D., E.A.L., M.P.S., S.E.P., V.A.R.; investigation, E.I.D., E.A.L., V.A.R., M.P.S., E.L.N., A.A.V., A.A.N., M.E.P.; data curation, E.I.D., E.A.L., M.P.S.; writing—original draft preparation, E.I.D, E.A.L., M.P.S., E.L.N.; writing—review and editing, S.E.P.; visualization, A.V.M.; supervision, S.E.P.; project administration, E.A.L., S.E.P.; funding acquisition, E.A.L. All authors have read and agreed to the published version of the manuscript.
Funding
Funded by Russian Science Foundation, grant number 20-74-10072, https://rscf.ru/project/20-74-10072/. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article and supplementary materials.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Alvarez, B.; Carballal, S.; Turell, L.; Radi, R. Formation and Reactions of Sulfenic Acid in Human Serum Albumin. Methods Enzymol 2010, 473, 117–136. [Google Scholar] [CrossRef] [PubMed]
- Ezra, A.; Rabinovich-Nikitin, I.; Rabinovich-Toidman, P.; Solomon, B. Multifunctional Effects of Human Serum Albumin Toward Neuroprotection in Alzheimer Disease. In Neuroprotection in Alzheimer’s Disease; Elsevier, 2017; pp. 217–238. [Google Scholar]
- Ghuman, J.; Zunszain, P.A.; Petitpas, I.; Bhattacharya, A.A.; Otagiri, M.; Curry, S. Structural Basis of the Drug-Binding Specificity of Human Serum Albumin. J Mol Biol 2005, 353, 38–52. [Google Scholar] [CrossRef]
- Carter, D.C.; He, X.M. Structure of Human Serum Albumin. Science 1990, 249, 302–303. [Google Scholar] [CrossRef] [PubMed]
- He, X.M.; Carter, D.C. Atomic Structure and Chemistry of Human Serum Albumin. Nature 1992, 358, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Tayyab, S.; Feroz, S.R. Serum Albumin: Clinical Significance of Drug Binding and Development as Drug Delivery Vehicle. Adv Protein Chem Struct Biol 2021, 123, 193–218. [Google Scholar] [CrossRef]
- Zunszain, P.A.; Ghuman, J.; Komatsu, T.; Tsuchida, E.; Curry, S. Crystal Structural Analysis of Human Serum Albumin Complexed with Hemin and Fatty Acid. BMC Struct Biol 2003, 3, 6. [Google Scholar] [CrossRef] [PubMed]
- Jayaraj, A.; Schwanz, H.A.; Spencer, D.J.; Bhasin, S.; Hamilton, J.A.; Jayaram, B.; Goldman, A.L.; Krishna, M.; Krishnan, M.; Shah, A.; et al. Allosterically Coupled Multisite Binding of Testosterone to Human Serum Albumin. Endocrinology 2021, 162. [Google Scholar] [CrossRef]
- van der Vusse, G.J. Albumin as Fatty Acid Transporter. Drug Metab Pharmacokinet 2009, 24, 300–307. [Google Scholar] [CrossRef]
- Zaragoza, F. Non-Covalent Albumin Ligands in FDA-Approved Therapeutic Peptides and Proteins. J Med Chem 2023, 66, 3656–3663. [Google Scholar] [CrossRef]
- van Witteloostuijn, S.B.; Pedersen, S.L.; Jensen, K.J. Half-Life Extension of Biopharmaceuticals Using Chemical Methods: Alternatives to PEGylation. ChemMedChem 2016, 11, 2474–2495. [Google Scholar] [CrossRef]
- Bech, E.M.; Pedersen, S.L.; Jensen, K.J. Chemical Strategies for Half-Life Extension of Biopharmaceuticals: Lipidation and Its Alternatives. ACS Med Chem Lett 2018, 9, 577–580. [Google Scholar] [CrossRef]
- Biere, A.L.; Ostaszewski, B.; Stimson, E.R.; Hyman, B.T.; Maggio, J.E.; Selkoe, D.J. Amyloid β-Peptide Is Transported on Lipoproteins and Albumin in Human Plasma. Journal of Biological Chemistry 1996, 271, 32916–32922. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.M.; Kokjohn, T.A.; Kalback, W.; Luehrs, D.; Galasko, D.R.; Chevallier, N.; Koo, E.H.; Emmerling, M.R.; Roher, A.E. Amyloid-Beta Peptides Interact with Plasma Proteins and Erythrocytes: Implications for Their Quantitation in Plasma. Biochem Biophys Res Commun 2000, 268, 750–756. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Chen, W.-D.; Wang, Y.-D. β-Amyloid: The Key Peptide in the Pathogenesis of Alzheimer’s Disease. Front Pharmacol 2015, 6, 221. [Google Scholar] [CrossRef]
- Hampel, H.; Hardy, J.; Blennow, K.; Chen, C.; Perry, G.; Kim, S.H.; Villemagne, V.L.; Aisen, P.; Vendruscolo, M.; Iwatsubo, T.; et al. The Amyloid-β Pathway in Alzheimer’s Disease. Mol Psychiatry 2021, 26, 5481–5503. [Google Scholar] [CrossRef] [PubMed]
- Algamal, M.; Milojevic, J.; Jafari, N.; Zhang, W.; Melacini, G. Mapping the Interactions between the Alzheimer’s Aβ-Peptide and Human Serum Albumin beyond Domain Resolution. Biophys J 2013, 105, 1700–1709. [Google Scholar] [CrossRef] [PubMed]
- Menendez-Gonzalez, M.; Gasparovic, C. Albumin Exchange in Alzheimer’s Disease: Might CSF Be an Alternative Route to Plasma? Front Neurol 2019, 10, 1036. [Google Scholar] [CrossRef] [PubMed]
- Boada, M.; Ortiz, P.; Anaya, F.; Hernández, I.; Muñoz, J.; Núñez, L.; Olazarán, J.; Roca, I.; Cuberas, G.; Tárraga, L.; et al. Amyloid-Targeted Therapeutics in Alzheimer’s Disease: Use of Human Albumin in Plasma Exchange as a Novel Approach for Abeta Mobilization. Drug News Perspect 2009, 22, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Stanyon, H.F.; Viles, J.H. Human Serum Albumin Can Regulate Amyloid-β Peptide Fiber Growth in the Brain Interstitium: Implications for Alzheimer Disease. J Biol Chem 2012, 287, 28163–28168. [Google Scholar] [CrossRef]
- Costa, M.; Ortiz, A.M.; Jorquera, J.I. Therapeutic Albumin Binding to Remove Amyloid-β. J Alzheimers Dis 2012, 29, 159–170. [Google Scholar] [CrossRef]
- Boada, M.; López, O.; Núñez, L.; Szczepiorkowski, Z.M.; Torres, M.; Grifols, C.; Páez, A. Plasma Exchange for Alzheimer’s Disease Management by Albumin Replacement (AMBAR) Trial: Study Design and Progress. Alzheimers Dement (N Y) 2019, 5, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Boada, M.; Anaya, F.; Ortiz, P.; Olazarán, J.; Shua-Haim, J.R.; Obisesan, T.O.; Hernández, I.; Muñoz, J.; Buendia, M.; Alegret, M.; et al. Efficacy and Safety of Plasma Exchange with 5% Albumin to Modify Cerebrospinal Fluid and Plasma Amyloid-β Concentrations and Cognition Outcomes in Alzheimer’s Disease Patients: A Multicenter, Randomized, Controlled Clinical Trial. J Alzheimers Dis 2017, 56, 129–143. [Google Scholar] [CrossRef]
- Prajapati, K.D.; Sharma, S.S.; Roy, N. Current Perspectives on Potential Role of Albumin in Neuroprotection. Rev Neurosci 2011, 22, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.-M.; Byun, K.; Cho, K.; Kim, J.Y.; Yoo, J.S.; Kim, D.; Paek, S.H.; Kim, S.U.; Simpson, R.J.; Lee, B. Human Microglial Cells Synthesize Albumin in Brain. PLoS One 2008, 3, e2829. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, H.M.; Kozlowski, P.B. Evidence for Blood-Brain Barrier Changes in Senile Dementia of the Alzheimer Type (SDAT). Ann N Y Acad Sci 1982, 396, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Mokrzycki, M.H.; Kaplan, A.A. Therapeutic Plasma Exchange: Complications and Management. Am J Kidney Dis 1994, 23, 817–827. [Google Scholar] [CrossRef] [PubMed]
- Szczeklik, W.; Wawrzycka, K.; Włudarczyk, A.; Sega, A.; Nowak, I.; Seczyńska, B.; Fajfer, I.; Zając, K.; Królikowski, W.; Kózka, M. Complications in Patients Treated with Plasmapheresis in the Intensive Care Unit. Anaesthesiol Intensive Ther 2013, 45, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Sutton, D.M.; Nair, R.C.; Rock, G. Complications of Plasma Exchange. Transfusion (Paris) 1989, 29, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Litus, E.A.; Kazakov, A.S.; Sokolov, A.S.; Nemashkalova, E.L.; Galushko, E.I.; Dzhus, U.F.; Marchenkov, V.V.; Galzitskaya, O.V.; Permyakov, E.A.; Permyakov, S.E. The Binding of Monomeric Amyloid β Peptide to Serum Albumin Is Affected by Major Plasma Unsaturated Fatty Acids. Biochem Biophys Res Commun 2019, 510, 248–253. [Google Scholar] [CrossRef]
- Litus, E.A.; Kazakov, A.; Deryusheva, E.; Nemashkalova, E.; Shevelyova, M.; Nazipova, A.; Permyakova, M.; Raznikova, E.; Uversky, V.; Permyakov, S. Serotonin Promotes Serum Albumin Interaction with the Monomeric Amyloid β Peptide. Int J Mol Sci 2021, 22, 5896. [Google Scholar] [CrossRef]
- Litus, E.A.; Kazakov, A.S.; Deryusheva, E.I.; Nemashkalova, E.L.; Shevelyova, M.P.; Machulin, A. V; Nazipova, A.A.; Permyakova, M.E.; Uversky, V.N.; Permyakov, S.E. Ibuprofen Favors Binding of Amyloid-β Peptide to Its Depot, Serum Albumin. Int J Mol Sci 2022, 23, 6168. [Google Scholar] [CrossRef] [PubMed]
- Cunnane, S.C.; Schneider, J.A.; Tangney, C.; Tremblay-Mercier, J.; Fortier, M.; Bennett, D.A.; Morris, M.C. Plasma and Brain Fatty Acid Profiles in Mild Cognitive Impairment and Alzheimer’s Disease. Journal of Alzheimer’s Disease 2012, 29, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Vlad, S.C.; Miller, D.R.; Kowall, N.W.; Felson, D.T. Protective Effects of NSAIDs on the Development of Alzheimer Disease. Neurology 2008, 70, 1672–1677. [Google Scholar] [CrossRef] [PubMed]
- Cirrito, J.R.; Disabato, B.M.; Restivo, J.L.; Verges, D.K.; Goebel, W.D.; Sathyan, A.; Hayreh, D.; D’Angelo, G.; Benzinger, T.; Yoon, H.; et al. Serotonin Signaling Is Associated with Lower Amyloid-β Levels and Plaques in Transgenic Mice and Humans. Proc Natl Acad Sci U S A 2011, 108, 14968–14973. [Google Scholar] [CrossRef] [PubMed]
- Catanzariti, A.-M.; Soboleva, T.A.; Jans, D.A.; Board, P.G.; Baker, R.T. An Efficient System for High-Level Expression and Easy Purification of Authentic Recombinant Proteins. Protein Sci 2004, 13, 1331–1339. [Google Scholar] [CrossRef]
- Travis, J.; Pannell, R. Selective Removal of Albumin from Plasma by Affinity Chromatography. Clin Chim Acta 1973, 49, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Pace, C.N.; Vajdos, F.; Fee, L.; Grimsley, G.; Gray, T. How to Measure and Predict the Molar Absorption Coefficient of a Protein. Protein Sci 1995, 4, 2411–2423. [Google Scholar] [CrossRef] [PubMed]
- UniProt Consortium UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res 2023, 51, D523–D531. [CrossRef] [PubMed]
- Law, V.; Knox, C.; Djoumbou, Y.; Jewison, T.; Guo, A.C.; Liu, Y.; Maciejewski, A.; Arndt, D.; Wilson, M.; Neveu, V.; et al. DrugBank 4.0: Shedding New Light on Drug Metabolism. Nucleic Acids Res 2014, 42, D1091–7. [Google Scholar] [CrossRef]
- Wishart, D.S.; Knox, C.; Guo, A.C.; Cheng, D.; Shrivastava, S.; Tzur, D.; Gautam, B.; Hassanali, M. DrugBank: A Knowledgebase for Drugs, Drug Actions and Drug Targets. Nucleic Acids Res 2008, 36, D901–6. [Google Scholar] [CrossRef]
- Macielag, M.J. Chemical Properties of Antimicrobials and Their Uniqueness. In Antibiotic Discovery and Development; Springer US: Boston, MA, 2012; pp. 793–820. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J Comput Chem 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Battistuz, T.; Bhat, T.N.; Bluhm, W.F.; Bourne, P.E.; Burkhardt, K.; Feng, Z.; Gilliland, G.L.; Iype, L.; Jain, S.; et al. The Protein Data Bank. Acta Crystallogr D Biol Crystallogr 2002, 58, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Adasme, M.F.; Linnemann, K.L.; Bolz, S.N.; Kaiser, F.; Salentin, S.; Haupt, V.J.; Schroeder, M. PLIP 2021: Expanding the Scope of the Protein-Ligand Interaction Profiler to DNA and RNA. Nucleic Acids Res 2021, 49, W530–W534. [Google Scholar] [CrossRef] [PubMed]
- Giorgino, T. Computing and Visualizing Dynamic Time Warping Alignments in R : The Dtw Package. J Stat Softw 2009, 31. [Google Scholar] [CrossRef]
- Bhattacharya, A.A.; Grüne, T.; Curry, S. Crystallographic Analysis Reveals Common Modes of Binding of Medium and Long-Chain Fatty Acids to Human Serum Albumin. J Mol Biol 2000, 303, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, S.; Amisaki, T. Fatty Acid Binding to Serum Albumin: Molecular Simulation Approaches. Biochim Biophys Acta 2013, 1830, 5427–5434. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A.A.; Curry, S.; Franks, N.P. Binding of the General Anesthetics Propofol and Halothane to Human Serum Albumin. High Resolution Crystal Structures. J Biol Chem 2000, 275, 38731–38738. [Google Scholar] [CrossRef] [PubMed]
- Boudinot, F.D.; Jusko, W.J. Plasma Protein Binding Interaction of Prednisone and Prednisolone. J Steroid Biochem 1984, 21, 337–339. [Google Scholar] [CrossRef] [PubMed]
- Larsen, F.G.; Larsen, C.G.; Jakobsen, P.; Brodersen, R. Interaction of Warfarin with Human Serum Albumin. A Stoichiometric Description. Mol Pharmacol 1985, 27, 263–270. [Google Scholar]
- Rahim, S.; Oise Aubry, A.-F. Location of Binding Sites on Immobilized Human Serum Albumin for Some Nonsteroidal Anti-Inflammatory Drugs. J Pharm Sci 1995, 84, 949–952. [Google Scholar] [CrossRef]
- Zsila, F. Subdomain IB Is the Third Major Drug Binding Region of Human Serum Albumin: Toward the Three-Sites Model. Mol Pharm 2013, 10, 1668–1682. [Google Scholar] [CrossRef] [PubMed]
- Steiner, R.F.; Roth, J.; Robbins, J. The Binding of Thyroxine by Serum Albumin as Measured by Fluorescence Quenching. J Biol Chem 1966, 241, 560–567. [Google Scholar] [CrossRef]
- Housaindokht, M.R.; Rouhbakhsh Zaeri, Z.; Bahrololoom, M.; Chamani, J.; Bozorgmehr, M.R. Investigation of the Behavior of HSA upon Binding to Amlodipine and Propranolol: Spectroscopic and Molecular Modeling Approaches. Spectrochim Acta A Mol Biomol Spectrosc 2012, 85, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug Repurposing: Progress, Challenges and Recommendations. Nat Rev Drug Discov 2019, 18, 41–58. [Google Scholar] [CrossRef]
- Andrich, K.; Bieschke, J. The Effect of (-)-Epigallo-Catechin-(3)-Gallate on Amyloidogenic Proteins Suggests a Common Mechanism. Adv Exp Med Biol 2015, 863, 139–161. [Google Scholar] [CrossRef] [PubMed]
- Leri, M.; Nosi, D.; Natalello, A.; Porcari, R.; Ramazzotti, M.; Chiti, F.; Bellotti, V.; Doglia, S.M.; Stefani, M.; Bucciantini, M. The Polyphenol Oleuropein Aglycone Hinders the Growth of Toxic Transthyretin Amyloid Assemblies. J Nutr Biochem 2016, 30, 153–166. [Google Scholar] [CrossRef]
- Rahmani, M.; Negro Álvarez, S.E.; Hernández, E.B. The Potential Use of Tetracyclines in Neurodegenerative Diseases and the Role of Nano-Based Drug Delivery Systems. Eur J Pharm Sci 2022, 175, 106237. [Google Scholar] [CrossRef]
- Ferrié, L.; Figadère, B.; Rose, C.; Tomas-Grau, R.H.; Chehín, R.; Zabala, B.; Michel, P.P.; Raisman-Vozari, R.; Brunel, J.-M. C9-Functionalized Doxycycline Analogs as Drug Candidates to Prevent Pathological α-Synuclein Aggregation and Neuroinflammation in Parkinson’s Disease Degeneration. ChemMedChem 2024, e202300597. [Google Scholar] [CrossRef]
- Tokuda, T.; Oide, T.; Tamaoka, A.; Ishii, K.; Matsuno, S.; Ikeda, S. Prednisolone (30-60 Mg/Day) for Diseases Other than AD Decreases Amyloid Beta-Peptides in CSF. Neurology 2002, 58, 1415–1418. [Google Scholar] [CrossRef]
- Alisky, J.M. Intrathecal Corticosteroids Might Slow Alzheimer’s Disease Progression. Neuropsychiatr Dis Treat 2008, 4, 831–833. [Google Scholar] [CrossRef]
- Barber, M.; Tait, R.C.; Scott, J.; Rumley, A.; Lowe, G.D.O.; Stott, D.J. Dementia in Subjects with Atrial Fibrillation: Hemostatic Function and the Role of Anticoagulation. J Thromb Haemost 2004, 2, 1873–1878. [Google Scholar] [CrossRef] [PubMed]
- Grossmann, K. Direct Oral Anticoagulants (DOACs) for Therapeutic Targeting of Thrombin, a Key Mediator of Cerebrovascular and Neuronal Dysfunction in Alzheimer’s Disease. Biomedicines 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Johansson, P.; Almqvist, E.G.; Johansson, J.-O.; Mattsson, N.; Hansson, O.; Wallin, A.; Blennow, K.; Zetterberg, H.; Svensson, J. Reduced Cerebrospinal Fluid Level of Thyroxine in Patients with Alzheimer’s Disease. Psychoneuroendocrinology 2013, 38, 1058–1066. [Google Scholar] [CrossRef] [PubMed]
- Beaman, E.E.; Bonde, A.N.; Larsen, S.M.U.; Ozenne, B.; Lohela, T.J.; Nedergaard, M.; Gíslason, G.H.; Knudsen, G.M.; Holst, S.C. Blood-Brain Barrier Permeable β-Blockers Linked to Lower Risk of Alzheimer’s Disease in Hypertension. Brain 2023, 146, 1141–1151. [Google Scholar] [CrossRef] [PubMed]
- Melnikov, V.; Tiburcio-Jimenez, D.; Mendoza-Hernandez, M.A.; Delgado-Enciso, J.; De-Leon-Zaragoza, L.; Guzman-Esquivel, J.; Rodriguez-Sanchez, I.P.; Martinez-Fierro, M.L.; Lara-Esqueda, A.; Delgado-Enciso, O.G.; et al. Improve Cognitive Impairment Using Mefenamic Acid Non-Steroidal Anti-Inflammatory Therapy: Additional Beneficial Effect Found in a Controlled Clinical Trial for Prostate Cancer Therapy. Am J Transl Res 2021, 13, 4535–4543. [Google Scholar] [PubMed]
- Dobarro, M.; Gerenu, G.; Ramírez, M.J. Propranolol Reduces Cognitive Deficits, Amyloid and Tau Pathology in Alzheimer’s Transgenic Mice. Int J Neuropsychopharmacol 2013, 16, 2245–2257. [Google Scholar] [CrossRef]
- Dobarro, M.; Orejana, L.; Aguirre, N.; Ramírez, M.J. Propranolol Restores Cognitive Deficits and Improves Amyloid and Tau Pathologies in a Senescence-Accelerated Mouse Model. Neuropharmacology 2013, 64, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Joo, Y.; Kim, H.-S.; Woo, R.-S.; Park, C.H.; Shin, K.-Y.; Lee, J.-P.; Chang, K.-A.; Kim, S.; Suh, Y.-H. Mefenamic Acid Shows Neuroprotective Effects and Improves Cognitive Impairment in in Vitro and in Vivo Alzheimer’s Disease Models. Mol Pharmacol 2006, 69, 76–84. [Google Scholar] [CrossRef]
- Choi, T.S.; Lee, H.J.; Han, J.Y.; Lim, M.H.; Kim, H.I. Molecular Insights into Human Serum Albumin as a Receptor of Amyloid-β in the Extracellular Region. J Am Chem Soc 2017, 139, 15437–15445. [Google Scholar] [CrossRef]
- Aisen, P.S.; Davis, K.L.; Berg, J.D.; Schafer, K.; Campbell, K.; Thomas, R.G.; Weiner, M.F.; Farlow, M.R.; Sano, M.; Grundman, M.; et al. A Randomized Controlled Trial of Prednisone in Alzheimer’s Disease. Alzheimer’s Disease Cooperative Study. Neurology 2000, 54, 588–593. [Google Scholar] [CrossRef]
- Deryusheva, E.; Kazakov, A.; Nemashkalova, E.; Shevelyova, M.; Nazipova, A.; Permyakova, M.; Raznikova, E.; Permyakov, S.; Litus, E. Modulation of Human Serum Albumin Interaction with Amyloid b Peptide by Ibuprofen, Risperidone, Serotonin and Tryptophan. FEBS Open Bio 2022, 12, 230–230. [Google Scholar] [CrossRef]
- Brodaty, H.; Ames, D.; Snowdon, J.; Woodward, M.; Kirwan, J.; Clarnette, R.; Lee, E.; Greenspan, A. Risperidone for Psychosis of Alzheimer’s Disease and Mixed Dementia: Results of a Double-Blind, Placebo-Controlled Trial. Int J Geriatr Psychiatry 2005, 20, 1153–1157. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, H.S.; Kim, Y.H.; Kwon, M.J.; Kim, J.-H.; Min, C.Y.; Yoo, D.M.; Choi, H.G. The Association Between Thyroid Diseases and Alzheimer’s Disease in a National Health Screening Cohort in Korea. Front Endocrinol (Lausanne) 2022, 13, 815063. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Guo, C. Multipronged Regulatory Functions of Serum Albumin in Early Stages of Amyloid-β Aggregation. ACS Chem Neurosci 2021, 12, 2409–2420. [Google Scholar] [CrossRef] [PubMed]
- Milojevic, J.; Raditsis, A.; Melacini, G. Human Serum Albumin Inhibits Abeta Fibrillization through a “Monomer-Competitor” Mechanism. Biophys J 2009, 97, 2585–2594. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Cheng, F.; Xu, L.; Jia, L. HSA Targets Multiple Aβ42 Species and Inhibits the Seeding-Mediated Aggregation and Cytotoxicity of Aβ42 Aggregates. RSC Adv 2016, 6, 71165–71175. [Google Scholar] [CrossRef]
- Bode, D.C.; Stanyon, H.F.; Hirani, T.; Baker, M.D.; Nield, J.; Viles, J.H. Serum Albumin’s Protective Inhibition of Amyloid-β Fiber Formation Is Suppressed by Cholesterol, Fatty Acids and Warfarin. J Mol Biol 2018, 430, 919–934. [Google Scholar] [CrossRef]
- Hornedo-Ortega, R.; Da Costa, G.; Cerezo, A.B.; Troncoso, A.M.; Richard, T.; Garcia-Parrilla, M.C. In Vitro Effects of Serotonin, Melatonin, and Other Related Indole Compounds on Amyloid-β Kinetics and Neuroprotection. Mol Nutr Food Res 2018, 62. [Google Scholar] [CrossRef]
Figure 1.
The scheme of the bioinformatic search of the AD-related therapeutic HSA ligands, their filtering, structural analysis and ranking.
Figure 1.
The scheme of the bioinformatic search of the AD-related therapeutic HSA ligands, their filtering, structural analysis and ranking.
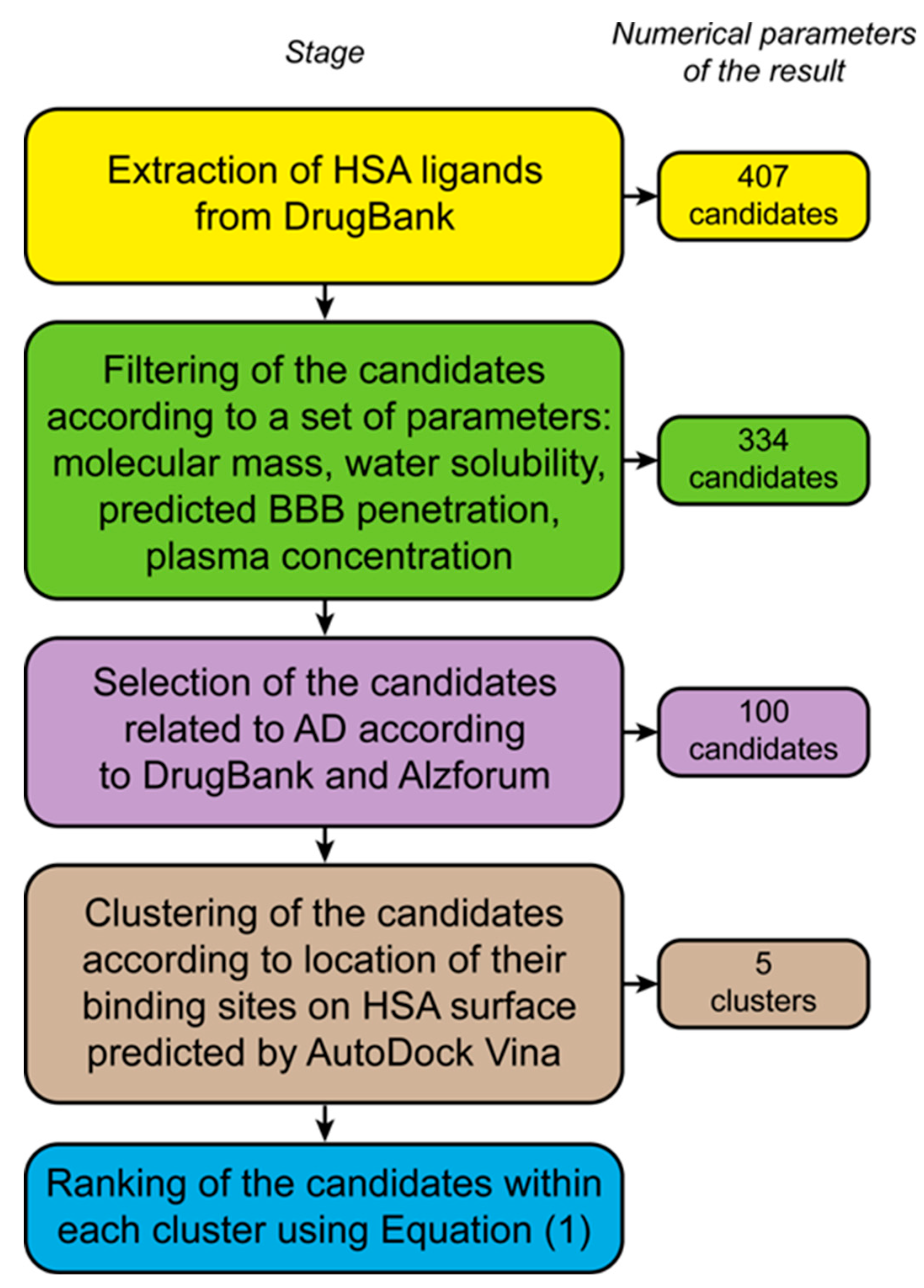
Figure 2.
Predicted location of the key HSA residues of the binding sites for the AD-related LMWLs clustered using the R language library ‘dtw’. The ligands of the individual clusters with highest R values (Equation (1)) are indicated (see Table S2 for the complete list), along with the R values (shown in the parentheses). The subdomains IA, IB, IIA, IIB, IIIA and IIIB are marked.
Figure 2.
Predicted location of the key HSA residues of the binding sites for the AD-related LMWLs clustered using the R language library ‘dtw’. The ligands of the individual clusters with highest R values (Equation (1)) are indicated (see Table S2 for the complete list), along with the R values (shown in the parentheses). The subdomains IA, IB, IIA, IIB, IIIA and IIIB are marked.
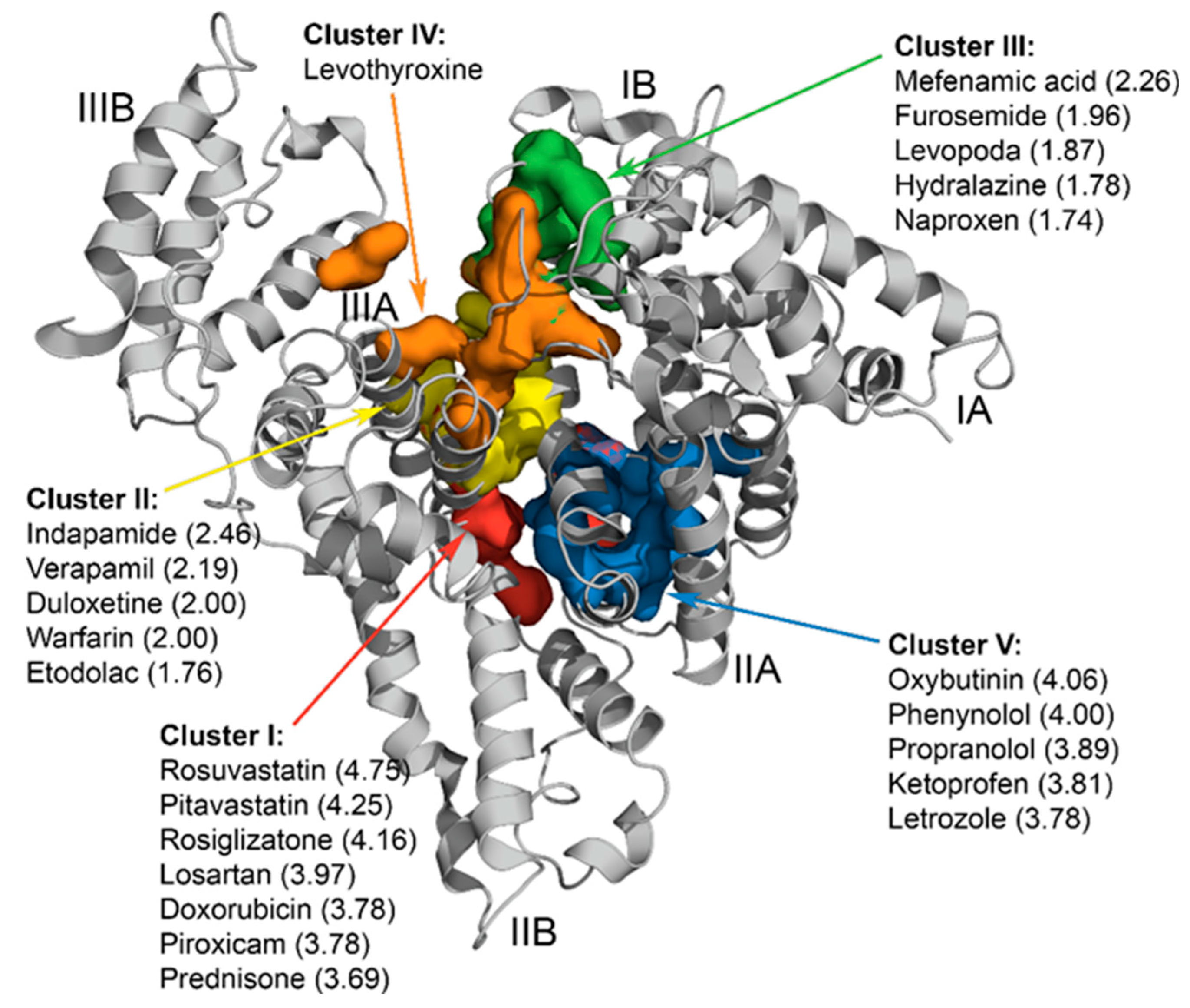
Figure 3.
The kinetics of HSA association (0-300 s) with the Aβ40 monomer immobilized on SPR chip’s surface by amine coupling, as well as the dissociation of their complex (300-2700 s), monitored by SPR spectroscopy in the presence (panels A-E) or absence (F) of the HSA ligands shown in Table 1: 2.5 mM prednisone (panel A), 1 mM warfarin (B), 250 μM mefenamic acid (C), 15 μM levothyroxine (D), 1 mM propranolol (E) (20 mM Tris-HCl, 150 mM NaCl, pH 7.4; 25 °C). The experimental curves are described within the heterogeneous ligand model (Equation (2)) (see Table 2 for the fitting parameters).
Figure 3.
The kinetics of HSA association (0-300 s) with the Aβ40 monomer immobilized on SPR chip’s surface by amine coupling, as well as the dissociation of their complex (300-2700 s), monitored by SPR spectroscopy in the presence (panels A-E) or absence (F) of the HSA ligands shown in Table 1: 2.5 mM prednisone (panel A), 1 mM warfarin (B), 250 μM mefenamic acid (C), 15 μM levothyroxine (D), 1 mM propranolol (E) (20 mM Tris-HCl, 150 mM NaCl, pH 7.4; 25 °C). The experimental curves are described within the heterogeneous ligand model (Equation (2)) (see Table 2 for the fitting parameters).
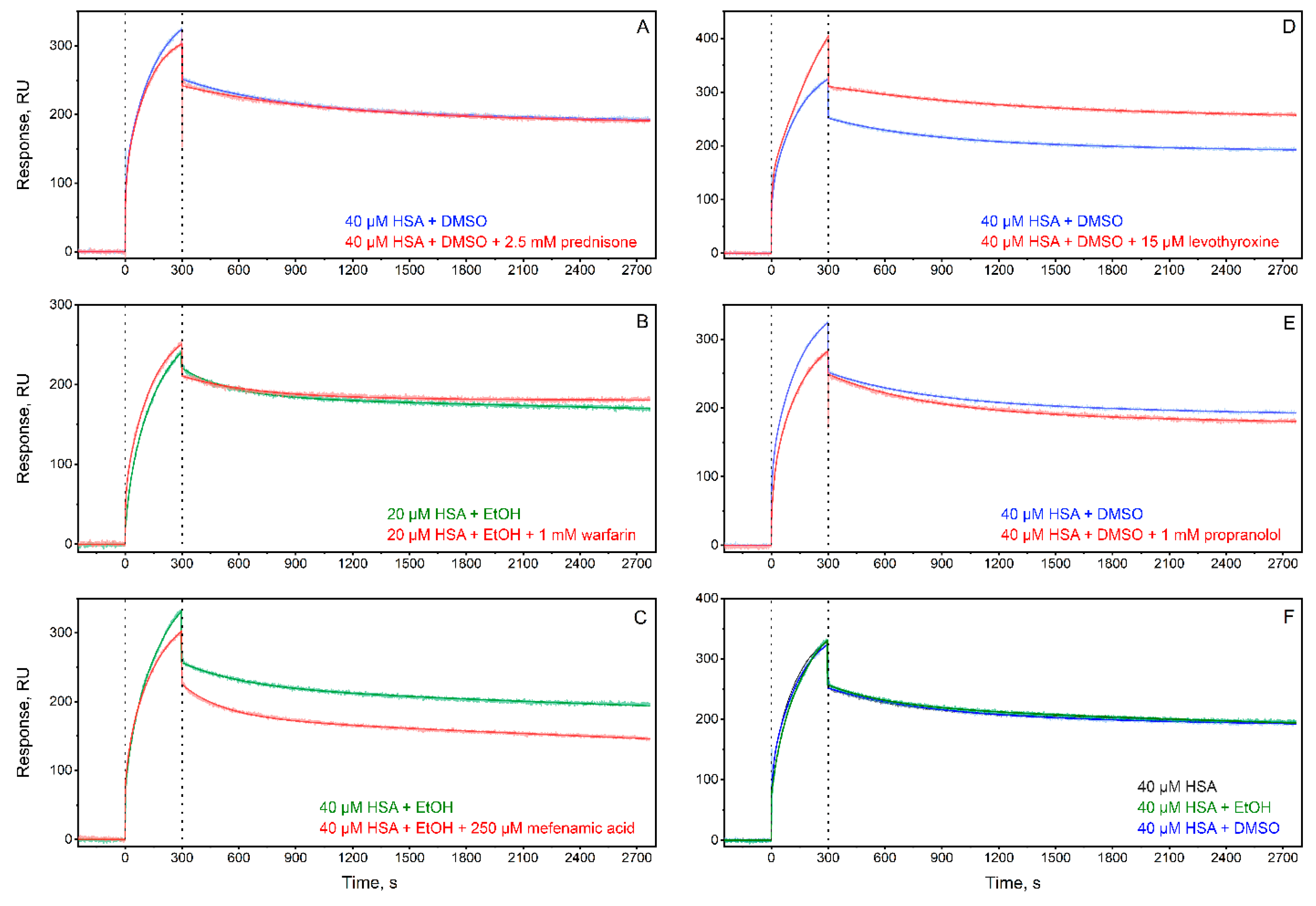
Figure 4.
The changes in the free energy changes accompanying HSA-Aβ40 interaction (δΔGi, i=1,2) induced by addition of the specific HSA ligands (see Table 1), calculated from the SPR data shown in Table 2 (ΔGi = – RT ln(55.3/KDi), i=1,2). Panel A corresponds to KD1, panel B ─ KD2.
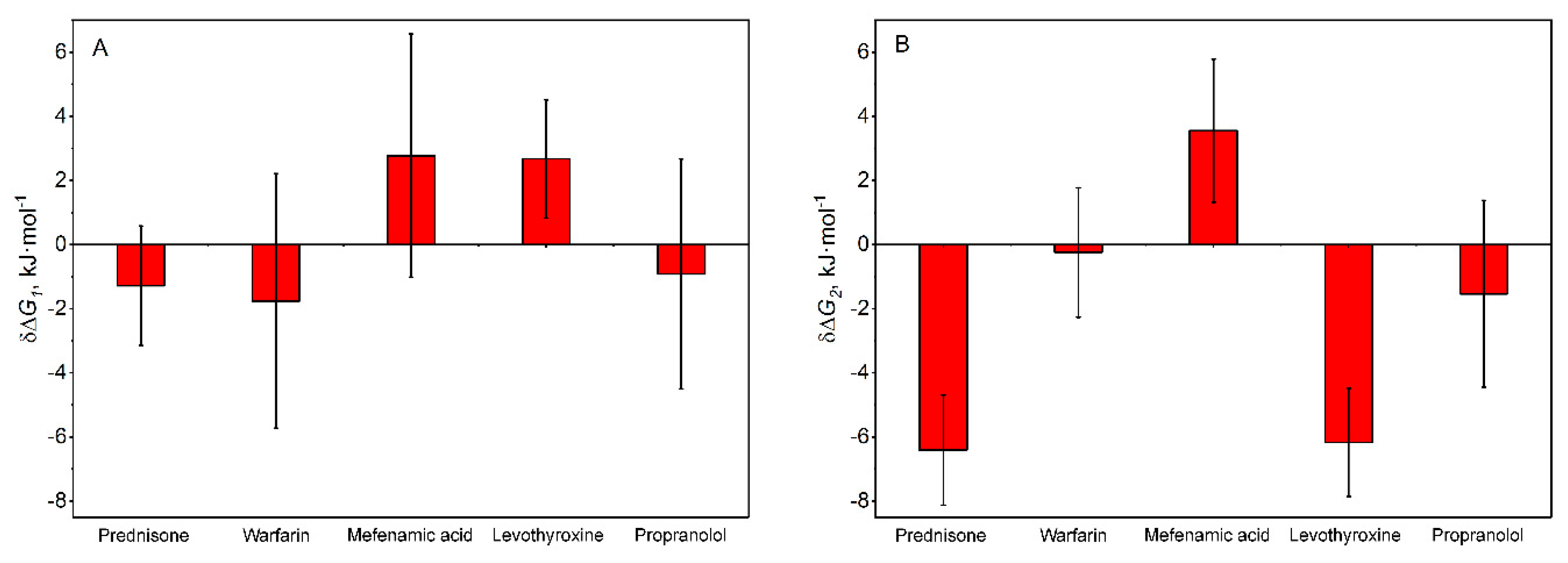

Table 1.
Information on the AD-associated LWMLs selected for experimental examination of their influence on HSA-Aβ interaction.
Table 1.
Information on the AD-associated LWMLs selected for experimental examination of their influence on HSA-Aβ interaction.
| Predicted cluster on HSA molecule | Drug | Discovery date | Drug class | Application area | Equilibrium association constant for the drug-HSA interaction | Calculated occupancy of HSA binding sites for 2.5-40 µM HSA | Drug concentration used for the SPR studies |
|---|---|---|---|---|---|---|---|
| I | Prednisone | 1950 | Corticosteroid | Transplantology; treatment of allergy, inflammation, infection, cancer, endocrine, autoimmune conditions | K = 103 M-1 [50] | 71% | 2.5 mM |
| II | Warfarin | 1945 | Antithrombotic agent | Thromboembolism treatment | K1 = 2×105 M-1 K2 = 5×104 M-1 [51] |
site1: 99-99.5%; site2: 98% | 1 mM |
| III | Mefenamic acid | 1961 | non-steroidal anti-inflammatory agent | Analgesia, treatment of inflammation и fever | K1=4×105 М-1 [52] K2 = 1×105 M-1 [53] |
95-96% | 250 μM |
| IV | Levothyroxine | 1914 | Thyroid hormone | treatment thyroid diseases including hypothyroidism and cancer | K = 105 M-1 (4 sites) [54] | 9-41% | 15 μM |
| V | Propranolol | Early 1960s | Beta blockers | Cardiology, including hypertension and myocardial infarction | K = 104 M-1 (2 sites) [55] | 90-91% | 1 mM |
Table 2.
Parameters of HSA-Aβ40 interaction in the presence/absence of the HSA ligands shown in Table 1 determined by SPR technique using the heterogeneous ligand model (Equation (2)).
Table 2.
Parameters of HSA-Aβ40 interaction in the presence/absence of the HSA ligands shown in Table 1 determined by SPR technique using the heterogeneous ligand model (Equation (2)).
| Ligand/Additive | ka1×102, M-1s-1 |
kd1×10-4, s-1 |
KD1×10-7, M |
ka2×102, M-1s-1 |
kd2×10-4, s-1 |
KD2×10-6, M |
|---|---|---|---|---|---|---|
| Without ligand + DMSO | 2.7±2.6 | 0.216±0.007 | 0.64±0.48 | 8.9±9.0 | 26±13 | 5.0±3.4 |
| Without ligand + ethanol | 5.6±4.7 | 0.27±0.20 | 1.2±1.6 | 13±8 | 34±11 | 3.2±1.5 |
| Prednisone + DMSO | 2.73±0.96 | 0.11±0.05 | 0.38±0.05 | 36±8 | 13.6±1.2 | 0.38±0.05 |
| Warfarin + ethanol | 7.9±3.7 | 0.50±0.39 | 0.57±0.43 | 24±28 | 32±11 | 2.9±1.9 |
| Mefenamic acid + ethanol | 3.3±1.6 | 0.97±0.56 | 3.6±2.1 | 3.6±3.4 | 31±12 | 13±10 |
| Levothyroxine + DMSO | 0.575±0.011 | 0.109±0.006 | 1.90±0.14 | 27.0±1.6 | 11.20±0.12 | 0.41±0.02 |
| Propranolol + DMSO | 22±31 | 0.204±0.010 | 0.45±0.55 | 25±34 | 18.8±0.2 | 2.7±2.6 |
Table 3.
Summary of the effect of the AD-related LMWLs of HSA on its interaction with Aβ and data on the role of these ligands in AD progression.
Table 3.
Summary of the effect of the AD-related LMWLs of HSA on its interaction with Aβ and data on the role of these ligands in AD progression.
| Cluster | HSA ligand | Effect of the ligand on HSA affinity for Aβ | Relevance for AD progression |
|---|---|---|---|
| I | prednisone | ↑ † | decline of AD biomarkers in non-AD patients after taking prednisone [61,62]; lack of effect in the treatment of AD patients [72] |
| risperidone | ↓ [73] | reduces psychosis and favors functioning in elderly patients with psychosis of AD and mixed dementia [74] | |
| II | ibuprofen | ↑ [32] | reduces the risk of AD progression [34] |
| III | mefenamic acid | ↓ † | not available |
| IV | levothyroxine | bidirectional effect † | hypothyroidism, thyroiditis and hyperthyroidism are more common among AD patients [75]; lowered levothyroxine level in cerebrospinal fluid of AD patients [65] |
| V | serotonin | ↑ [31] | modulates Aβ level in the central nervous system of AD patients [35] |
† See Table 2; ↑ - promote interaction; ↓ - prevent interaction.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



