Submitted:
04 April 2024
Posted:
05 April 2024
You are already at the latest version
Abstract
Poplars (Populus spp.) are of significant ecological and economic importance. Long-term breeding efforts were aimed mainly at obtaining fast-growing and productive plants and less considered resistance to pests. This study aimed to identify the patterns of susceptibility to Saperda carcharias (Linnaeus, 1758) (Coleoptera: Cerambycidae) among Populus hybrids and pure species, considering crossing combinations and some other traits. In our research, 35 clones of poplar species and hybrids of the Ukrainian and foreign selection from the Aigeiros, Tacamahaca, and Leucoides sections were tested in 2021 and 2023 in the plantation created in 2014 in Eastern Ukraine. S. carcharias preferred infesting clones with earlier foliage development, larger diameters, and height increments. Specifically, hybrids with American maternal lineage showed the highest susceptibility, whereas those with European maternal lineage or Asian origin displayed lower infestation levels or resistance, respectively. This variability suggests a complex relationship between genetic background and pest resistance, underlining the need for a deeper understanding of the factors influencing susceptibility to S. carcharias. Selecting native species clones or creating mixed clone plantations could enhance the resilience of poplar plantations to pest threats.
Keywords:
Populus spp.
; hybrids
; clones
; Saperda carcharias
; phenology
; height increment.
1. Introduction
Poplar trees (Populus spp.) play a crucial role ecologically and economically due to their rapid growth and versatility in various applications, including timber production, bioenergy, and environmental remediation [1,2,3,4,5]. However, the Short Rotation Woody Crops (SRWC) yields are often reduced by a range of interacting environmental stressors, including frost [6], drought [27], salinity [8], pathogens [9], and phytophagous insects [10]. With climate change and increasing ecological pressures, understanding the complex interplay between insect herbivory resistance and the tolerance mechanisms to these abiotic stressors has become paramount. Dozens of species of phytophagous insects feed and develop in all organs of poplar causing various effects on tree viability, growth rate, and wood quality [9,11,12]. It depends on insect biological traits, population density, plant susceptibility, and environmental conditions [13,14,15,16,17].
Breeding efforts have historically focused on obtaining fast-growing and productive plants [18,19,20,21,22,23,24]. However, these plants had various responses to stress induced by abiotic factors of the environment and activities of living organisms within the environment. Particularly, as susceptibility to insect damage varies widely among clones, the limited genetic diversity of poplar in large SRWC areas may heighten the risk of pest-related losses or abiotic disasters [10].
Most studies of insect resistance in poplar hybrids and clones have focused on defoliators [27] because the foliage damage is visible and easy to quantify, and these insects can be controlled by spraying with an insecticide [9,11,28]. At the same time, phloem-boring or xylophagous insects at SRWC can, at high population densities, weaken trees by maturation feeding on leaves, gnaw the galleries of different diameters and at different depths in trunks, branches, and roots, actively or passively vector the pathogens, or create conditions for the penetration of pathogens through entrance holes and promote their development by weakening the trees [14,15,29,30,31,32]. Some insects of this group have relatively low prevalence and harmfulness in forests but increase the population density at a single-species plantation, particularly Cryptorhynchus lapathi (Linnaeus, 1758) (Coleoptera: Curculionidae: Cryptorhynchinae), Saperda carcharias (Linnaeus, 1758) (Coleoptera: Cerambycidae), Cossus cossus (Linnaeus, 1758) (Lepidoptera: Cossidae), Paranthrene tabaniformis (Rottemburg, 1775) (Lepidoptera: Sesiidae) [9,11,12], and Anoplophora glabripennis Motschulsky, 1853 [32]. In eastern Ukraine, ten of 72 investigated xylophagous poplar species, were rated as the most dangerous. However, most of them are highly harmful in the case of inhabiting over 60 % of trees [14,15].
Large poplar longhorn beetle S. carcharias is one of them because it inhabits only living, healthy trees regardless of age [33,34], damages foliage and the bark of new growth at maturation feeding [35,36], and promotes the pathogen invasion [37] causing physiological harm [14]. The larva excavates a long and wide gallery deep into the wood [33,38] causing technical harm. The timber of damaged trees may be used for paper, pulp, or match production but not for saw timber, furniture, and veneering [39]. The life cycle of S. carcharias lasts 2 to 4 years, according to climatic conditions [40,41,42]. Even within the same tree, larvae develop in the upper part of the trunk for 2 years, and in the base of the trunk for 3–4 years [35,36]. The larvae pupate starting in May, depending on the instars of the hibernating individuals, and due to the heterogeneous temperature regime both within the plantation and in one tree, the emergence of adults and the colonization of new trees continues until September [40,41,42].
Since detecting the xylophagous insects in the early stages of tree colonization requires more effort than in the case of defoliators, and control is expensive and little effective [11], the best way to reduce negative consequences for plantation production is to increase the diversity of clones and hybrids [9].
Fritz et al. [43] have suggested that clone resistance against herbivores is affected by hybridization in four different ways: the same resistance of hybrids and parents, intermediate resistance of hybrids compared to the parents, hybrid susceptibility, and hybrid resistance is close to either more susceptible or more resistance parent. Responses of different insect species may vary widely to the same hybrid host. This indicates diverse genetic effects of interspecific hybridization on resistance.
The resistance of parental forms as a result of long-term coevolution may be one of the important factors of resistance to pests. For example, Manchurian ash (Fraxinus mandshurica Rupr.) coevolved with emerald ash borer, Agrilus planipennis Fairmaire, 1888 (Coleoptera: Buprestidae) and is more resistant than North American or European ash species [44]. Usually, clones with the same parents have similar resistance. However, the progeny of P. trichocarpa is generally more resistant to caterpillar-like damage [17]. Research has shown that cultivar ‘Robusta’ (Populus deltoides × P. nigra), obtained about 1910, is more susceptible to some insects than other P. deltoides × P. nigra crosses of more recent origin. At the same time, P. deltoides × P. nigra hybrids were damaged more intensively than P. trichocarpa (Torr. & Gray) clones [17].
In Ukraine, numerous studies have been carried out with hybrids P. deltoides, P. × euramericana, P. trichocarpa, P. laurifolia, P. lasiocarpa primarily focusing on growth [1,20,45,46,47,48,49,50,51] and biology [21,52]. However, modern studies on the resistance of poplar hybrids to insects are only starting in eastern Ukraine.
This article aims to identify patterns of susceptibility or resistance to Saperda carcharias (Linnaeus, 1758) (Coleoptera: Cerambycidae) infestation among Populus hybrids and pure species, focusing on the influence of their seasonal development, stem diameter, height increment, and crossing combinations.
2. Materials and Methods
2.1. Study Region and Tested Poplar Clones
The experimental site is located on the territory of the State Enterprise “Kharkiv Forest Research Station”, 15 km from Kharkiv, in the Kharkiv region, Ukraine (50°05′01″ N, 36°18′25″ W, 156 m a.s.l.) (Figure 1). This area falls under the Forest-Steppe zone as per comprehensive forestry zoning [53]. Climate data obtained from ClimateCharts.net (Zepner et al., 2020) indicates an average annual temperature of +8.8°C and an average annual rainfall of 535.2 mm for the period 2001-2020 [54]. The climate is classified as temperate continental (Dfb) according to the Köppen classification system [55].
Researches were carried out in the collection of poplar (Populus sp.) and willow (Salix sp.) clones. The plot is flat, with a slight slope to the north. On the northern and western sides, it is bordered by deciduous forests, which include mainly Quercus robur L., Acer platanoides L., Tilia cordata Mill., and Ulmus laevis Pall. The rows of plants on the plantation are arranged from south to north. The distance between rows is 2 m and between plants in a row 2 m.
The cuttings, rooted the previous summer in an open field nursery and dormant for the winter, were planted in the spring of 2014. The plot encompasses 35 clones of poplar, representing species and hybrids selected from the Aigeiros, Tacamahaca, and Leucoides sections (Table 1).
In 2021, part of the trees on the plantation were cut to assess the biomass obtained from individual clones. In 2022, access to the plantation was impossible due to military operations. In 2023, it was discovered that the crowns of the pruned plants had been restored.
2.2. Weather Data Analysis
To compare the weather conditions of individual years, the following indicators were evaluated: average air temperature for a year and vegetation period (period with air temperature over 10 °C); the sum of precipitation for a year, and vegetation period; the dates of stable transition of temperature over 5 and 10 °C, and Hydrothermal index (HTI).
The dates of stable transition of temperature over 5 and 10 °C were evaluated according to a method by V. Meshkova [56]. G. T. Selyaninov hydrothermal index (HTI) was calculated as
where ΣP is precipitation for a period with mean monthly air temperature over 10 °C, mm; Σt is the sum of daily air temperature for the same period, °C [57].
2.3. Field Data
Surveys of the distribution of phytophagous insects on plantations and assessment of the condition of clones were carried out in 2019–2021 and 2023. When examining each plant, the stem diameter at the lower part was measured with a caliper. Plant condition (healthy, dead) and the presence of pest infestation were recorded (Figure 2, Table S1).
Particular attention was paid to identifying S. carcharias. Its presence at the site was evidenced by characteristic frass, and foliage damage as a result of maturation feeding of the adults (Figure 3a,b).
Galleries of this longhorn beetle inside the stem could be seen after cutting the plant (Figure 3cd).
Poplar infestation by S. carcharias was calculated as the ratio of infested and all plants of each clone expressed as a percentage. The error of this indicator was calculated considering the number of plants in a given clone, and the comparison of infestation for different clones was assessed using the criterion Z [58]. Mean monthly air temperature and precipitation for 1990–2020 for Kharkiv were taken from the ClimateCharts.net web platform [54], and respective weather indicators for the years of field research were taken from the website [59].
The rate of poplar foliage development was assessed on 10 plants of each clone by the scoring scheme: 1—dormant buds completely enveloped by the scales; 2—bud swelling with scales slightly diverging showing a narrow yellow margin; the presence of one or more droplets of balsam; 3—bud sprouting, with tips of the small leaves emerging out of the scales; 4—buds completely opened with leaves still clustered together; scales still present; 5—leaves diverging with their blades still rolled up; scales may be present or absent; 6—leaves completely unfolded (but smaller in size than mature ones); lengthening of the axis of the shoot evident; scales absent [60].
The average score on the day of recording was calculated. Considering the preliminary assessment of the seasonal development of the clones, the arithmetic mean score of each clone, obtained from the assessment on April 20, was used for comparison. The clones with foliage in phase ≤ 3 were considered as late, 3.1–4—medial, and 4.1–5 early.
The date of completing the growing season was assessed on 10 plants of each clone on October 15 by the proportion of fallen leaves. The clones with a percentage of foliage fall ≤ 60% were considered as late, 61–90 %—medial, and >90 %—early.
The stem diameter and height increment of these trees was also measured on October 15. The clones with stem diameter ≤ 6 cm were considered as small, 6.1–7 cm—medial, and >7 cm—large. The clones with height increment ≤ 60% were considered as low, 61–90 %—medial and >90 %—high.
3. Results
3.1. Weather Conditions
In all years of the study, the annual air temperature in the plantation area exceeded long-term data: from 2.1°C (by 23.7%) in 2019 to 0.4°C (by 4.6%) in 2021 (Table 2). However, the temperature of the growing season exceeded long-term data to a lesser extent: from 1.8°C (by 10.1%) in 2019 to 0.3°C (by 1.9%) in 2021. The date of stable temperature transition over 5°C was earlier by 10 days in 2019, 20 days in 2020, and 13 days in 2023. In 2021, this date was 4 days later than long-term data. However, the date of stable temperature transition over 10°C was earlier than long-term data only in 2019 and 2023 (by 7 and 6 days, respectively), and in 2020 and 2021 it occurred 8 and 7 days later, respectively.
The annual precipitation in 2019–2021 was lower than long-term data (in 2019—by 180.1 mm, or by 34.5%). However, in 2023 it exceeded long-term data by 171.7 mm (32.9%). Precipitation for the growing season was almost twice less than the long-term values. In 2020, it was almost the same as the long-term values, and in 2023 it exceeded them by 23.3%. The hydrothermal index in 2019 was almost 2 times less than the long-term value, in 2020 it was almost equal to the long-term value, and in 2023 it even exceeded it (Table 2).
3.2. Prevalence of S. carcharias Depending on Clone Phenology, Size, and Management
A survey in 2020 showed that out of 696 trees inspected, 72 (10.3%) were infested by S. carcharias, 2 (0.3%) by Saperda populnea (Linnaeus, 1758), 4 (0.58%) by Sesia apiformis (Clerck, 1759) (Lepidoptera: Sesiidae), and 4 (0.58%) died of unknown causes. In 2021, S. carcharias has already infested 12 % of trees. After cutting in 2021 and restoring the crowns, in the survey of 2023, uncut trees amounted to 32.5%, trees after cutting with restored crowns—57.6%, and dead trees—10%. At the same time, S. carcharias was found in all three dead trees, and among cut and uncut trees—18.8% and 26.3%, respectively (Figure 4).
Infestation of uncut and cut groups of trees by S. carcharias was higher in 2023 compared with 2020, however, it was significantly greater in uncut trees (see Figure 4).
Infestation by S. carcharias was higher in 2023 compared with 2020 for most clones (Figure 5). In both years of assessment, the clone ‘Ivantiivska’ was the most infested by S. carcharias. In three more clones (‘Kytaiska × pyramidalna’, ‘Karolinska 162’, and ‘Volosystoplidna’) over 50 % of trees were infested.
Four clones (‘Perspektyvna’, ‘Sakrau79’, ‘Rosijska’, and ‘Addita’) were infested only in 2023. 12 clones were not infested in both years, particularly, ‘Deltopodibna’, Gradizka’, ‘Gulliver’, ‘Kanadska×balsamichna’, ‘Lubenska’, ‘Mobilna’, ‘Nocturne’, ‘Novoberlinska-7’, ‘Robusta 16’, ‘Rohanska’, ‘Sakrau45-51’, ‘Slava Ukrayiny’. To reveal the possible causes of various infestation levels of clones, some of their traits were analyzed.
Distribution by the dates of spring foliage development for poplar clones with S. carcharias presence and absence differed significantly (χ2 = 17.4; χ2 0.05 = 6.0; χ2 0.01 = 9.2). The clones with earlier foliage development were more susceptible to infestation (Figure 6a). At the same time, the dates of the fall have no significant impact on S. carcharias presence (χ2 = 4.0; χ2 0.05 = 6.0) (Figure 6b).
The average diameter of non-infested trees was 5.9 ± 0.06 cm, and of trees infested with S. carcharias 6.6 ± 0.19 cm, but in both groups of trees, there were the plants with a diameter of 1 to 12 cm. S. carcharias preferred colonizing clones with larger diameters (χ2 = 8.6; χ2 0.05 = 6.0) (Figure 7a) and height increment (χ2 = 41.9; χ2 0.05 = 6.0; χ2 0.01 = 9.2) (Figure 7b).
3.3. Prevalence of S. carcharias Depending on Clone Origin and Crossing Combination
S. carcharias infested clones of all tested poplar sections and their combinations, except T × L (Figure 8).
However, within each of the four susceptible combinations of sections, there were hybrids with high or low resistance to infestation by this pest (Figure 9). Among the hybrids from section Aigeiros, those with a maternal plant of American origin (P. deltoides) exhibited significantly higher susceptibility to infestation by S. carcharias compared with hybrids with a maternal plant of European origin (P. nigra) (Figure 9a).
Maternal plants of crossing combination A × L were also of American origin, and paternal (P. lasiocarpa) were of Asian origin. Although the S. carcharias infestation of the presented hybrids differed almost twice, the differences were not statistically confirmed (Figure 9b). In the A × L hybrid group, the clones with a maternal plant of Asian origin (P. suaveolens, P. simonii) were considerably more heavily infested compared with clones of European maternal lineage. However, the hybrid P. deltoides × P. balsamifera, descending from two American parents, showed no infestation (Figure 9c). P. trichocarpa plants were the most infested (44.2 %) in the Tacamahaca section. Only 3.4% of plants having an American maternal and Asian paternal (P. trichocarpa × P. laurifolia) lineage were infested by S. carcharias, and the third member of this group having Asian origin (P. simonii f. fastigiata) was not infested at all (Figure 9d).
4. Discussion
In our research, 35 clones of poplar species and hybrids of the Ukrainian and foreign selection from the Aigeiros, Tacamahaca, and Leucoides sections were tested in 2021 and 2023 for sustainability or resistance to infestation by S. carcharias in the plantation created in 2014 in Eastern Ukraine.
Infestation by S. carcharias was higher in 2023 compared with 2020 for most clones (Figure 5). In both years of assessment, the clone ‘Ivantiivska’ was the most infested by S. carcharias. Three more clones (‘Kytaiska × pyramidalna’, ‘Karolinska 162’, and ‘Volosystoplidna’) had over 50 % infestation.
12 clones were not infested in both years, particularly, ‘Deltopodibna’, Gradizka’, ‘Gulliver’, ‘Kanadska × balsamichna’, ‘Lubenska’, ‘Mobilna’, ‘Nocturne’, ‘Novoberlinska-7’, ‘Robusta 16’, ‘Rohanska’, ‘Sakrau45-51’, ‘Slava Ukrayiny’. To reveal the possible causes of various infestation levels of clones, some of their traits were analyzed.
In all years of the study, the annual air temperature in the plantation area exceeded long-term data, but the dates of the stable temperature transition over 10°C occurred earlier than long-term data in 2019 and 2023 by 7 and 6 days, respectively, and occurred 8 and 7 days later it in 2020 and 2021 (see Table 2). Such variations could affect the attractiveness, suitability, and susceptibility of poplars to S. carcharias and larvae development inside the trunk. This warrants further investigation into their role in pest infestation dynamics.
In our research, the precipitation and hydrothermal index in 2019–2021 were inferior to long-term data, and in 2023 exceeded them (see Table 2). Lack of precipitation usually promotes phytophagous insect development while its increase promotes tree resistance [56]. Since the development length of S. carcharias larvae in the poplar trunk varies depending on locality and year, this aspect is also advisable for further study in different clones.
Infestation of uncut and cut trees by S. carcharias was higher in 2023 compared with 2020, however, it was significantly greater in uncut trees (see Figure 4). This may be because after cutting a part of the trunk was left with insufficient height for the galleries of S. carcharias. However, the plants were cut at a height of about 40 cm, and the length of the galleries in the dissected plants from which the adults emerged did not exceed 17 cm (our unpublished data). It is possible also that wood moisture decreased after cutting, which was unfavorable for larvae development.
In our research, the clones with earlier foliage development were more susceptible to infestation (see Figure 6a), while the dates of the fall had no significant impact on S. carcharias presence (Figure 6b). However, in Canada, resistant to Cryptorhynchus lapathi (L.) (Coleoptera: Curculionidae) poplar clones were flushed approximately three weeks earlier than the susceptible clones [63]. The conflicting findings may be because C. lapathi larvae mine in the bark while developing through three instars, and then move into the xylem, while S. carcharias larva immediately gnaws a gallery to the sapwood. The choice of plants with earlier leaf development may be explained by the availability of foliage for S. carcharias maturation feeding, which then becomes able to mate and lay eggs on nearby trees.
The dates of the fall have no significant impact on clone infestation by S. carcharias because it needs foliage only for maturation feeding after adult emergence, which goes on from May to August [35,36].
In our research, S. carcharias preferred colonizing clones with larger diameters (Figure 7a) and height increments (Figure 7b). The first may be because the larva cannot complete its development in young plants with insufficient wood volume. The second phenomenon may be because the plants with greater height increments usually have greater radial increments [53] and sufficient wood volume for larvae development [39].
S. carcharias showed a wide capacity to infest clones across all presented poplar sections and their crossing combinations, except the Tacamahaca and Leucoides cross. Within each crossing combination, there was a discernible variation in susceptibility levels. This variation underscores the complex nature of hybrid resistance, suggesting that neither clone origin nor crossing combination alone can predict susceptibility to S. carcharias.
The studies have provided mixed results regarding the susceptibility of Populus hybrids compared to pure species in the face of pest infestations. In Sweden, Christersson [18] has studied that hybrids P. trichocarpa and P. deltoides were more prone to pest attacks, unlike their pure P. trichocarpa counterparts which were largely spared. In Canada, Kalischuk et al. [64] reported that poplar hybrids suffered severe infestations by poplar bud gall mites, and no trees were severely infested in areas where pure poplars grew.
Contrary to these findings, in Latvia, the study by Zeps et al. [37], no significant differences in susceptibility of rot and poplar borer were found between hybrid and European aspen. In Finland, Välimäki and Heliövaara’s [38] research indicates no significant difference in poplar borer larval galleries between aspens and hybrid aspens. A study by Moore and Wilson [65] in Michigan found no discernible preference by S. inornata for hybrid over pure Populus species, despite attacking a substantial proportion of the population. This lack of preference challenges the notion that Populus hybrids are inherently more vulnerable to pests.
Among the hybrids from section Aigeiros, those with a maternal plant of American origin (P. deltoides) exhibited higher susceptibility to infestation. Conversely, hybrids with a maternal plant of European origin (P. nigra) showed significantly lower infestation levels. This pattern suggests a pivotal role of maternal lineage in determining hybrid susceptibility to pest attacks.
For the crossing combinations of Aigeiros and Leucoides, P. deltoides × P. lasiocarpa showed a significantly high infestation rate, while P. × canadensis cv. ‘Regenerata’ × P. lasiocarpa had a lower infestation rate, suggesting variability in pest resistance even within specific crossing combinations.
In the Aigeiros and Tacamahaca hybrid group, clones with a maternal plant of Asian origin (P. suaveolens, P. simonii) were more heavily infested, while those with European maternal lineage were less affected. Interestingly, the hybrid P. deltoides × P. balsamifera, descending from two American parents, showed no infestation, highlighting the complexity of genetic influences on pest resistance. The experience of Hannon et al. [29] suggests that hybrids with P. deltoides × P. nigra parentage are more resistant to stem borers than P. trichocarpa × P. deltoides or P. trichocarpa × P. nigra parentage.
Among the representatives of section Tacamahaca, P. trichocarpa was significantly more susceptible to S. carcharias infestation. However, a genome-wide association study (GWAS) conducted by Sepúlveda et al. [31] on P. trichocarpa provenances revealed three SNP markers significantly associated with resistance to S. calcarata. This discovery indicates that P. trichocarpa employs a sophisticated machinery of genetic expression and metabolite production to fend off S. calcarata attacks in different conditions. Another representative with an American maternal and Asian paternal (P. trichocarpa × P. laurifolia) lineage exhibited low infestation rates. The third representative, of Asian origin (P. simonii f. fastigiata), was not infested, suggesting a potential genetic basis for resistance.
When sorting non-infested clones by parent origin, it was observed that European and American species were more prevalent by plant count, whereas European-American and European-Asian hybrids dominated by clone count. This distribution suggests a nuanced relationship between genetic background and pest resistance, warranting further exploration into the specific genetic or environmental factors that confer resilience against S. carcharias infestation. Hybrids with European maternal lineage generally exhibited lower susceptibility to pest infestations, suggesting a robust genetic basis for resistance within these lines. Hybrids derived from American maternal lineage tend to be more vulnerable to pests, necessitating careful consideration in breeding programs aiming for pest-resistant poplar cultivars. The presence of Asian maternal lineage or complex crossing combinations involving Asian lineage can result in higher susceptibility.
The findings underscore the importance of considering maternal lineage and specific hybrid genetic makeup when developing poplar breeding strategies to enhance resistance to pest infestations. Further research into the genetic and environmental interactions influencing these susceptibility patterns is crucial for advancing pest-resistant poplar cultivation.
The exploration of alternative strategies, such as the utilization of selected clones of native species or the creation of mixed clone plantations, emerges as a viable path toward the sustainable development of poplar plantations resilient to the dynamic threats of pests and pathogens. Our results corroborate the alternative strategy proposed by Biselli et al. [9] about creating mixed clone plantations, which emerge as viable paths toward the sustainable development of poplar plantations resilient to the dynamic threats of pests and pathogens.
Insect herbivory resistance, a key trait in poplar trees, influences their ability to withstand and recover from damage caused by herbivorous insects. This resistance trait is critical for maintaining tree health and productivity. On the other hand, the ability of hybrid poplars to tolerate environmental stresses like frost, drought, and salinity can significantly affect their survival and growth in diverse ecosystems. These abiotic stressors are becoming more pronounced due to changing climate patterns and land-use practices, making it imperative to investigate how insect herbivory resistance may interact with the trees’ abilities to tolerate such stressors. Further research into the genetic and environmental interactions influencing these susceptibility patterns is crucial for advancing pest-resistant poplar cultivation.
5. Conclusions
Among 35 clones of poplar species and hybrids of the Ukrainian and foreign selection from the Aigeiros, Tacamahaca, and Leucoides sections in the plantation created in 2014 in Eastern Ukraine, S. carcharias preferred infesting clones with earlier foliage development, larger diameters, and height increments. Infestation by S. carcharias was greater in 2023 compared with 2020 for most clones, however, 12 clones from 35 tested were not infested in both years. S. carcharias preferred colonizing clones with earlier foliage development, larger diameters, and height increments. American origin showing higher susceptibility, European origin demonstrating lower susceptibility, and Asian origin or mixed lineages presenting variable resistance to S. carcharias infestation. The findings underscore the importance of considering maternal lineage and specific hybrid genetic makeup when developing poplar breeding strategies to enhance resistance to pest infestations. Further research into the genetic and environmental interactions influencing these susceptibility patterns is crucial for advancing pest-resistant poplar cultivation. The utilization of selected clones of native species or the creation of mixed clone plantations emerges as a viable path toward the sustainable development of poplar plantations resilient to the dynamic threats of pests.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Database of tested clones’ health condition.
Author Contributions
Author Contributions: Conceptualization, V.M.; methodology, V.M., N.V.; software, O.B., N.V.; validation, O.B. and N.V.; formal analysis, V.M., N.V., Y.S.; investigation, V.M., N.V., Y.S., K.Zh., O.Z.; data curation, K.Zh., O.Z. and Y.S.; writing—original draft preparation, V.M., N.V.; writing—review and editing, V.M., N.V., Y.S; visualization, O.B., N.V.; supervision, V.M.. All authors have read and agreed to the published version of the manuscript.
Funding
The paper was prepared by the authors in the framework of a research plan of URIFFM (grant 0120U101891), which was supported by the State Forest Resources Agency of Ukraine
Data Availability Statement
Data are available upon email request to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kutsokon, N.K.; Khudolieieva, L.V.; Los, S.A.; Vysotska, N.Y.; Torosova, L.O.; Tkach, V.P; Nesterenko, O.G; Rashydov, N.M. Evaluation of growth characteristics of one-year poplar and willow clones in short-rotation plantation in Kharkiv region. Studia Biologica 2018, 12, 55–64. [Google Scholar] [CrossRef]
- Niemczyk, M.; Hu, Y.; Thomas, B.R. Selection of poplar genotypes for adapting to climate change. Forests 2019, 10, 1041. [Google Scholar] [CrossRef]
- Oliveira, N.; Pérez-Cruzado, C.; Cañellas, I.; Rodríguez-Soalleiro, R.; Sixto, H. Poplar short rotation coppice plantations under Mediterranean conditions: The case of Spain. Forests 2020, 11, 1352. [Google Scholar] [CrossRef]
- Schiberna, E.; Borovics, A.; Benke, A. Economic modelling of poplar short rotation coppice plantations in Hungary. Forests 2021, 12, 623. [Google Scholar] [CrossRef]
- Fürtner, D.; Perdomo Echenique, E. A.; Hörtenhuber, S. J.; Schwarzbauer, P.; Hesser, F. Beyond Monetary Cost-Benefit Analyses: Combining Economic, Environmental and Social Analyses of Short Rotation Coppice Poplar Production in Slovakia. Forests 2022, 13, 349. [Google Scholar] [CrossRef]
- Čakšs, R.; Zeltinš, P.; Cakša, L.; Zeps, M.; Jansons, A. The effects ¯ of frost cracks and large poplar borer damage on stem rot in hybrid aspen (Populus tremula L. × Populus tremuloides Michx.) clones. Forests 2022, 13, 593. [Google Scholar] [CrossRef]
- Rosso, L.; Cantamessa, S.; Bergante, S.; Biselli, C.; Fricano, A.; Chiarabaglio, P. M. . & Carra, A. Responses to drought stress in poplar: what do we know and what can we learn? Life 2023, 13, 533. [Google Scholar]
- Zhou, X.; Dong, Y.; Zhang, Q.; Xiao, D.; Yang, M.; Wang, J. Expression of multiple exogenous insect resistance and salt tolerance genes in Populus nigra L. Frontiers in Plant Science 2020, 11, 546447. [Google Scholar] [CrossRef]
- Biselli, C.; Vietto, L.; Rosso, L.; Cattivelli, L.; Nervo, G.; Fricano, A. Advanced breeding for biotic stress resistance in poplar. Plants 2022, 11, 2032. [Google Scholar] [CrossRef]
- Nordman, E.E.; Robison, D.J. , Abrahamson, L.P., Volk, T.A. Relative resistance of willow and poplar biomass production clones across a continuum of herbivorous insect specialization: Univariate and multivariate approaches. Forest Ecology and Management, 2005; 217, 307–318. [Google Scholar]
- Isebrands, J.G.; Richardson, J. Insect and Other Pests of Poplars and Willows. Poplars and Willows: Trees for Society and the Environment. Isebrands, J.G.; Richardson, J. FAO, 2014; 459–526. [Google Scholar]
- De Tillesse, V.; Nef, L.; Charles, J.; Hopkin, A.; Augustin, S. Damaging poplar insects. FAO, Rome. 2007.
- Mattson, W.; Lawrence, R.; Haack, R.; Herms, D.; Charles, P. Defensive strategies of woody plants against different insect feeding guilds in relation to plant ecological strategies and intimacy of association with insects. In Mechanism of woody plant defenses against insects, search for pattern; Mattson, W., Levieux, J., Bernard-Dagan, J. (eds). Springer, Berlin, 1988: p 416.
- Skrylnyk, Y.Y.; Zhupinska, K.Y.; Koshelyaeva, Y.V.; Meshkova, V.L. Physiological harmfulness of xylophagous insects in poplar and aspen stands in the Left-Bank Forest-Steppe. Forestry & Forest melioration 2023, 142, 147–157. [Google Scholar]
- Skrylnyk, Yu.Ye.; Zhupinska, K.Yu.; Koshelyaeva, Ya.V.; Meshkova, V.L. Xylophagous insects (Insecta: Coleoptera, Hymenoptera, Lepidoptera) of Populus sp. (Malpighiales: Salicaceae) in the eastern regions of Ukraine. The Kharkov Entomological Society Gazette 2023, 31, 24–30. [Google Scholar] [CrossRef]
- Timchenko, G.A.; Treml’, A.G. Pests of poplars in the eastern part of Ukraine and Crimea. Entomol. review 1963, 42, 793–810. [Google Scholar]
- Tomescu, R.; Nef, L. Leaf-eating insect damage on different poplar clones and sites. Annals of Forest Science 2007, 64, 99–108. [Google Scholar] [CrossRef]
- Christersson, L. Biomass production of intensively grown poplars in the southernmost part of Sweden: observations of characters, traits and growth potential. Biomass Bioenergy 2006, 30, 497–508. [Google Scholar] [CrossRef]
- Chiarabaglio, P. M.; Deidda, A.; Bergante, S.; Castro, G.; Facciotto, G.; Giorcelli, A. . & Carbonaro, C. Life Cycle Assessment (LCA): new poplar clones allow an environmentally sustainable cultivation. Ann. Silvic. Res. 2020, 45, 72–82. [Google Scholar]
- Fuchylo, Y.D.; Sbytna, M.V.; Hayda, Y.I.; Kozatska, N.Y. Growth and productivity of hybrid poplar plantations in conditions of western forest-steppe of Ukraine. Scientific Bulletin of UNFU 2017, 27, 43–47. [Google Scholar] [CrossRef]
- Fuchylo, Ya; Maurer, V. ; Sbytna, M.; Odarchenko, I.; Fuchylo, D. Features of woody biomass and planting-stock of poplar in “stump” type of plantation management. Scientific works of the Forestry Academy of Sciences of Ukraine 2016, 14, 134–140. [Google Scholar]
- Kang, M.S.; Pham, H.N. Simultaneous selection for high-yielding and stable crop genotypes. Agron. J. 1991, 83, 161–165. [Google Scholar] [CrossRef]
- Neyko, I.S.; Kolchanova, O.V. Adaptability and features of poplar varieties growth in conditions of the Podillya region. Scientific bulletin of UNFU 2018, 28, 53–56. [Google Scholar] [CrossRef]
- Niemczyk, M.; Kaliszewski, A.; Jewiarz, M.; Wróbel, M.; Mudryk, K. Productivity and biomass characteristics of selected poplar (Populus spp.) cultivars under the climatic conditions of northern Poland. Biomass and Bioenergy 2018, 111, 46–51. [Google Scholar] [CrossRef]
- Caldbeck, E.S.; McNabb, Jr.H.S.; Hart, E.R. Poplar clonal preferences of the cottonwood leaf beetle. Journal of Economic Entomology 1978, 71, 518–520. [Google Scholar] [CrossRef]
- Castellani, E.; Freccero, V.; Lapietra, G. Proposal of a scale of differentiation of poplar leaf buds useful for antiparasitic interventions. Plant Biosystem 1967, 101, 355–360. [Google Scholar]
- Yousuf, P.; Razzak, S.; Parvaiz, S.; Rather, Y.A.; Lone, R. Role of plant phenolics in the resistance mechanism of plants against insects. In Plant Phenolics in Biotic Stress Management (pp. 191-215). Singapore: Springer Nature Singapore, 2024.
- Meshkova, V.L. (Ed.) Methodical guidelines for a survey, assessment and forecasting of the spread of forest pests and diseases for the lowland part of Ukraine; Planeta-Print: Kharkiv, Ukraine, 2020; 90p, ISBN 978-617-7897-00-1. [Google Scholar]
- Hannon, E.R.; Kittelson, N.T.; Eaton, J.A.; Brown, J.J. Screening hybrid poplar clones for susceptibility to Cryptorhynchus lapathi (Coleoptera: Curculionidae). Journal of economic entomology 2008, 101, 199–205. [Google Scholar] [CrossRef]
- Marquina, J.L.; Marlats, R.; Núñez Cresto, M. Cloning susceptibility of poplar (Populus sp. ) when attacked by Platypus mutatus in Buenos Aires, Argentinaю Bosque (Valdivia) 2006, 27, 92–97. [Google Scholar]
- Sepúlveda, S.L.; Neale, D.B.; Holliday, J.A.; Famula, R.; Fiehn, O.; Stanton, B.J.; Guerra, F.P. GWAS on the attack by aspen borer Saperda calcarata on black cottonwood trees reveals a response mechanism involving secondary metabolism and independence of tree architecture. Forests 2023, 14, 1129. [Google Scholar] [CrossRef]
- Zhou, Y.; Ge, X.; Zou, Y.; Guo, S.; Wang, T.; Zong, S. Prediction of the potential global distribution of the Asian longhorned beetle Anoplophora glabripennis (Coleoptera: Cerambycidae) under climate change. Agricultural and Forest Entomology 2021, 23, 557–568. [Google Scholar] [CrossRef]
- Palm, T. The wood borers and bark beetles of southern and central Swedish deciduous trees. Opuscula Entomol, 1959; 15–16, 334–335. [Google Scholar]
- Latzel, M.J.; Gruppe, A.; Lemme, H. Analysis of larval galleries of the large poplar longhorn beetle Saperda carcharias L. 1758 (Coleoptera, Cerambycidae). Mitteilungen der Deutschen Gesellschaft für allgemeine und angewandte Entomologie 2018, 21, 83–87. [Google Scholar]
- Ritchie, W. The structure, bionomics, and economic importance of Saperda carcharias L., the large poplar longhorn. Rev Appl Entomol, 1921; A IX, 171–172. [Google Scholar]
- Šrot, M. Bionomics of poplar borer (Saperda carcharias L. ). Reports of the Forest Research Institutes of Czechoslovakia 1962, 25, 85–114. [Google Scholar]
- Zeps, M.; Senhofa, S.; Zadina, M.; Neimane, U.; Jansons, A. Stem damages caused by heart rot and large poplar borer on hybrid and European aspen. For. Stud. 2017, 66, 21–26. [Google Scholar] [CrossRef]
- Välimäki, S.; Heliövaara, K. Hybrid aspen is not preferred by the large poplar borer (Saperda carcharias). Arthropod-Plant Interactions 2007, 1, 205–211. [Google Scholar] [CrossRef]
- Isebrands, J.G.; Richardson, J. (Eds.) . Poplars and willows: trees for society and the environment. CABI. 2014.
- Schneiderowa, J. Economical meaning of the large poplar borer (Saperda carcharias L.—Cerambycidae, Coleoptera). Prace IBL 1961, 234, 3–98. [Google Scholar]
- Petitpierre, E. Saperda carcharias (L., 1758) nueva especie para la fauna balear (Coleoptera: Cerambycidae). Revista gaditana de Entomología 2018, 9, 191–192. [Google Scholar]
- Nuorteva, M.; Patomäki, J.; Saari, L. Large poplar longhorn, Saperda carcharias (L.), as food for white-backed woodpecker, Dendrocopos leucotos (Bechst.). Silva Fennica 1981, 15, 208–221. [Google Scholar] [CrossRef]
- Fritz, R.S.; Moulia, C.; Newcombe, G. Resistance of hybrid plants and animals to herbivores, pathogens, and parasites. Annu Rev Ecol Syst 1999, 30, 565–591. [Google Scholar] [CrossRef]
- Villari, C.; Herms, D.A.; Whitehill, J.G.; Cipollini, D.; Bonello, P. Progress and gaps in understanding mechanisms of ash tree resistance to emerald ash borer, a model for wood-boring insects that kill angiosperms. New Phytol. 2016, 209, 63–79. [Google Scholar] [CrossRef]
- Patlai, I.M.; Rudenko, V.N. Selection of fast-growing forest trees in Ukraine. Forestry and Forest Melioration 1990, 81, 3–7. [Google Scholar]
- Starova, N.V. Selection of Salicaceae. Moscow: Forest Industry, 1980, p. 1–208.
- Los, S.A.; Tereshchenko, L.I.; Gayda, Yu.І.; Ustimenko, P.М. State of forest genetic resources in Ukraine. Kharkiv: PLANETA-PRINT, 2014. 138 p. ISBN 978-617-7229-06-2.
- Torosova, L.A.; Vysotska, N.Y.; Los, S.A.; Orlovska, T.V.; Zolotyh, I.V. Studies of morphological characters for representatives of Populus genus. Forestry and Forest Melioration 2015, 126, 148–157. [Google Scholar]
- Kutsokon, N.; Rakhmetov, D.; Rakhmetova, S.; Khudolieieva, L.; Rashydov, N. Nursery screening of poplar and willow clones for biofuel application in Ukraine. iForest-Biogeosciences and Forestry 2022, 15, 401. [Google Scholar] [CrossRef]
- Vysotska, N.Y. Current state and prospects of the poplar genetic resources conservation in Ukraine. Scientific Works of the Forestry Academy of Sciences of Ukraine 2017, 15, 38–44. [Google Scholar] [CrossRef]
- Vysotska, N.; Torosova, L. Collection of species and varieties of willows and poplars in the Ukrainian Research Institute of Forestry and Forest Melioration named after G. M. Vysotsky. In: Conservation of Forest Genetic Resources: Proceedings of the 5th International Conference, 39. Homel, Belarus: NAS of Belarus, Scientific and Practical Center of the NAS of Belarus. 2017 p.38–40 http://www.forinst.basnet.by/2017/files/2017-InForest-conference_proceedings.pdf.
- Krupej, N.S. Inheritance of crown pyramidality in pyramidal poplar hybrids. Forestry & Forest Melioration 1970, 23, 71–79. [Google Scholar]
- Nazarenko, V.V.; Pasternak, V.P. Patterns of formation of forest types of Forest-steppe of the Kharkiv region. Kharkiv, KhNAU, 2016, 190 p.
- Zepner, L. , Karrasch, P., Wiemann, F., Bernard, L.,: ClimateCharts. net—an interactive climate analysis web platform. International Journal of Digital Earth, 2021, 14, 338–356. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Meshkova, V.L. Seasonal development of foliage browsing insects; Novoe Slovo: Kharkov, Ukraine, 2009; pp. 1–396. ISBN 978-966-2046-69-4. [Google Scholar]
- Selyaninov, G. T. Methodology of Agricultural Climate Characteristics. World Agro-climatic reference book. L.-M. 1937; 5–29. [Google Scholar]
- Atramentova, L.A.; Utevskaya, O.M. Statistical Methods in Biology; Likhtar: Gorlovka, Ukraine, 2008; 248p, ISBN 978-966-2129-26-7. [Google Scholar]
- Meteofor: weather in Kharkiv. Available online: https://meteofor.com.ua/ru/weather-kharkiv-14410 (accessed on 2 March 2024).
- Turok, J.; Lefèvre, F.; Cagelli, L. ; De Vries, SMG (compilers). Populus nigra Network. Report of the second meeting, 10–12 September 1995, Casale Monferrato, Italy. Rome, Italy: International PlantGenetic Resources Institute, 1996, 26 pp.
- Python Software Foundation, Python Language Reference, Available at http://www.www.python.org. (accessed on 2 March 2024).
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Broberg, C.L.; Borden, J.H. Hybrid poplar clones with Populus maximowiczii parentage demonstrate postoviposition antibiosis to Cryptorhynchus lapathi (Coleoptera: Curculionidae). Journal of Economic Entomology 2005, 98, 2254–2259. [Google Scholar] [CrossRef]
- Kalischuk, A.R.; Gom, L.A.; Floate, K.D.; Rood, S.B. Intersectional cottonwood hybrids are particularly susceptible to the poplar bud gall mite. Can J Bot 1997, 75, 1349–1355. [Google Scholar] [CrossRef]
- Moore, L.M.; Wilson, L.F. Impact of the poplar-gall saperda, Saperda inornata (Coleoptera: Cerambycidae) on a hybrid Populus plantation in Michigan. Great Lakes Entomol 1986, 19, 163–167. [Google Scholar] [CrossRef]
Figure 1.
Location of study plot in the map of Kharkiv region (Ukraine).
Figure 2.
Location of alive, dead, cut, and infested by S. carcharias plants at the experimental plot in 2020 (before cutting) and in 2023 (after cutting).
Figure 2.
Location of alive, dead, cut, and infested by S. carcharias plants at the experimental plot in 2020 (before cutting) and in 2023 (after cutting).
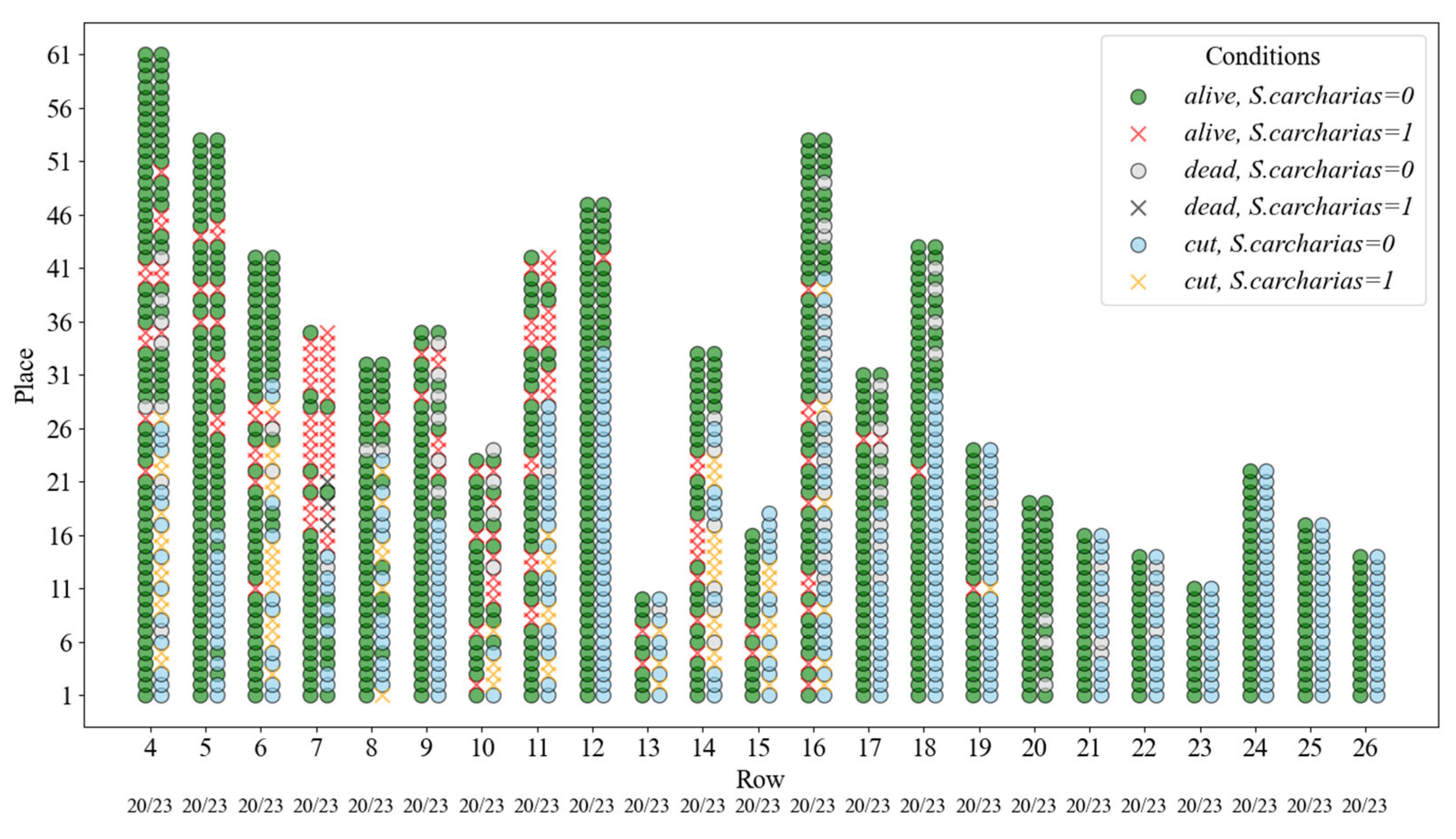
Figure 3.
Signs of S. carcharias in plantation: (a) frass at the base of the trunk, chips, and streaks are visible; (b) maturation feeding of an adult; (c) a gallery inside the stem; (d) cross-section in the lower part of infested poplar stem.
Figure 3.
Signs of S. carcharias in plantation: (a) frass at the base of the trunk, chips, and streaks are visible; (b) maturation feeding of an adult; (c) a gallery inside the stem; (d) cross-section in the lower part of infested poplar stem.

Figure 4.
Proportion of poplar trees, infested by S. carcharias in 2021 (before cutting) and 2023 (after cutting). The infestation of groups of trees with the same letters in parentheses has no significant difference at p=0.05.
Figure 4.
Proportion of poplar trees, infested by S. carcharias in 2021 (before cutting) and 2023 (after cutting). The infestation of groups of trees with the same letters in parentheses has no significant difference at p=0.05.
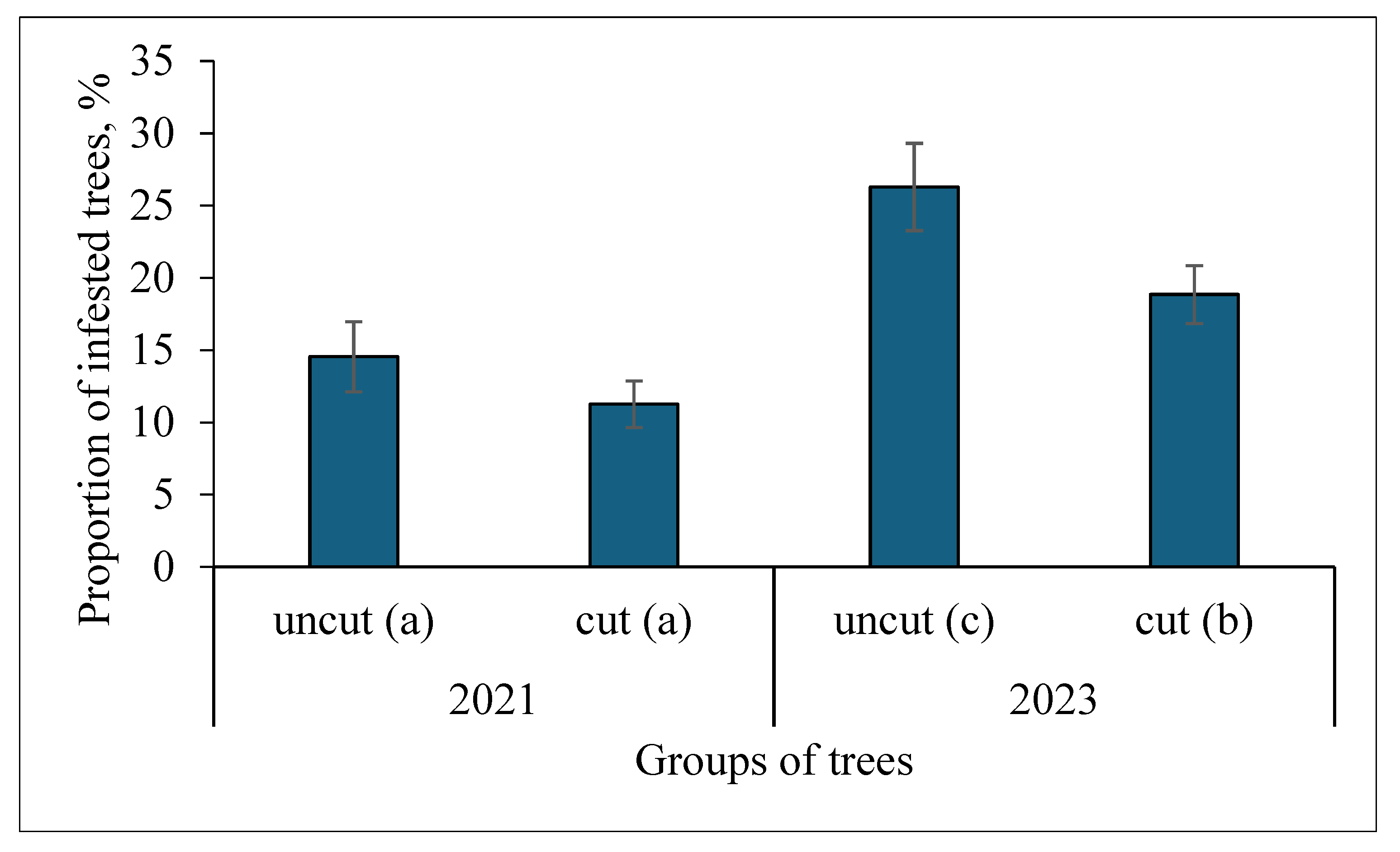
Figure 5.
The proportion of trees infested by S. carcharias in poplar clones in 2020 and 2023 (non-infested clones are not shown). Full clone names are presented in Table 1.
Figure 5.
The proportion of trees infested by S. carcharias in poplar clones in 2020 and 2023 (non-infested clones are not shown). Full clone names are presented in Table 1.
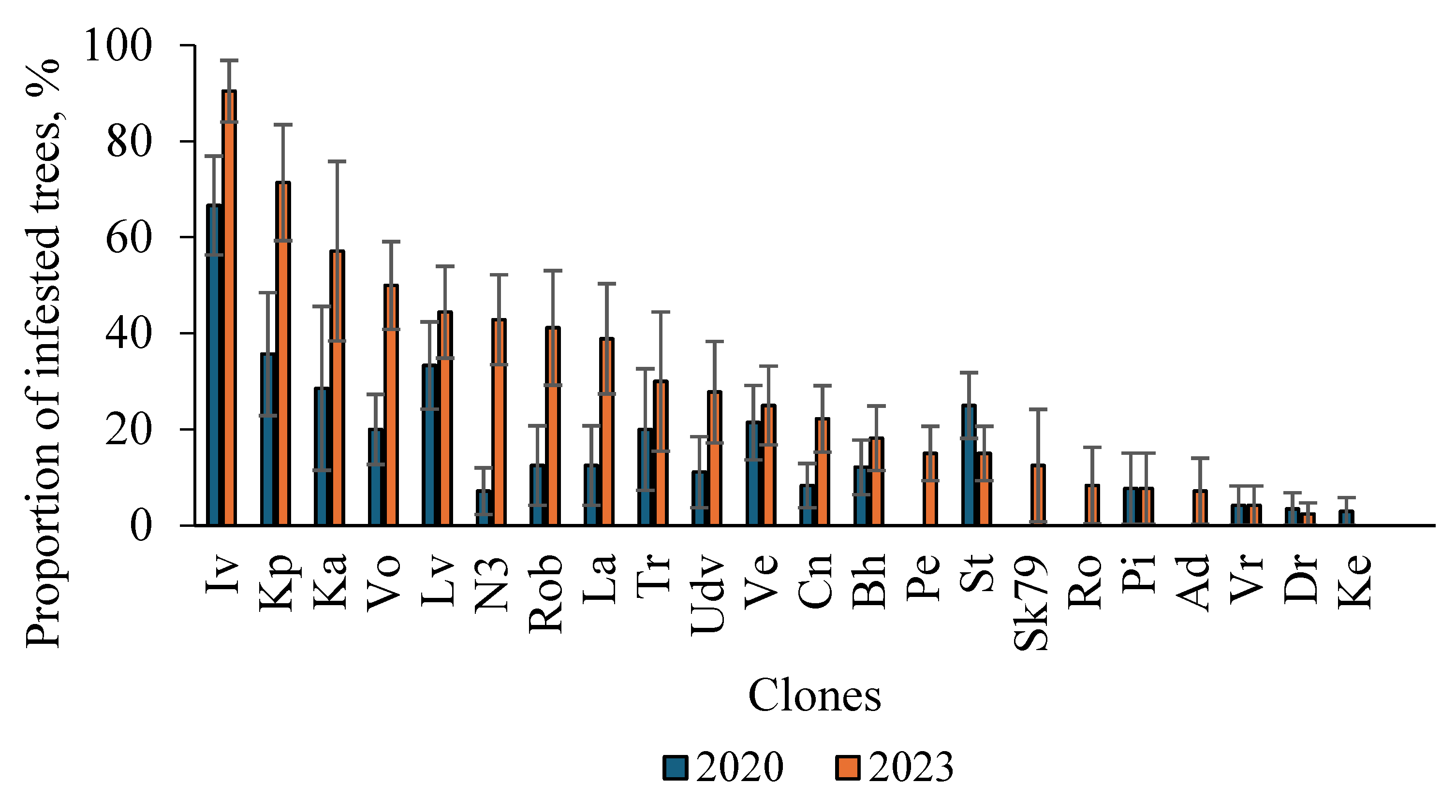
Figure 6.
Distribution of poplar clones, infested by S. carcharias (present) and non-infested (absent) (a) depending on the score of foliage development on April 20: phase ≤ 3—late; 3.1–4—medial; 4.1–5 early; (phase 3—bud sprouting, with tips of the small leaves emerging out of the scales; phase 4—buds completely opened with leaves still clustered together; scales still present; phase 5—leaves diverging with their blades still rolled up; scales may be present or absent;) (b) depending on the percentage of foliage fall on October 15: fall ≤ 60%—late; 61–90 %—medial; >90 %—early.
Figure 6.
Distribution of poplar clones, infested by S. carcharias (present) and non-infested (absent) (a) depending on the score of foliage development on April 20: phase ≤ 3—late; 3.1–4—medial; 4.1–5 early; (phase 3—bud sprouting, with tips of the small leaves emerging out of the scales; phase 4—buds completely opened with leaves still clustered together; scales still present; phase 5—leaves diverging with their blades still rolled up; scales may be present or absent;) (b) depending on the percentage of foliage fall on October 15: fall ≤ 60%—late; 61–90 %—medial; >90 %—early.
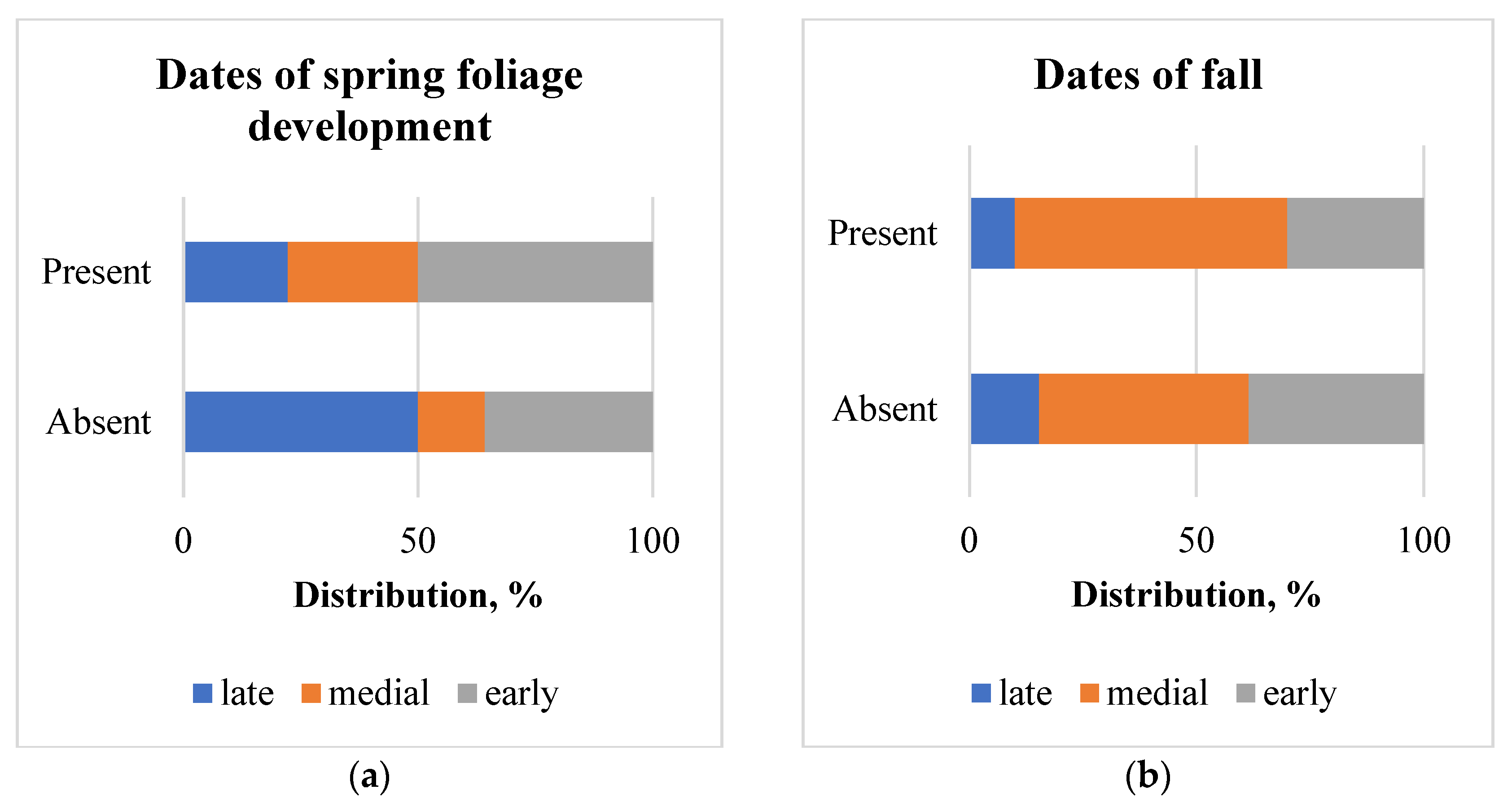
Figure 7.
Distribution of poplar clones, infested by S. carcharias (present) and non-infested (absent) (a) depending on poplar stem diameter: small ≤ 6 cm; medial—6.1–7 cm; large—>7 cm; (b) depending on height increment: low ≤ 60%; 61–90 %—medial; >90 %—high.
Figure 7.
Distribution of poplar clones, infested by S. carcharias (present) and non-infested (absent) (a) depending on poplar stem diameter: small ≤ 6 cm; medial—6.1–7 cm; large—>7 cm; (b) depending on height increment: low ≤ 60%; 61–90 %—medial; >90 %—high.
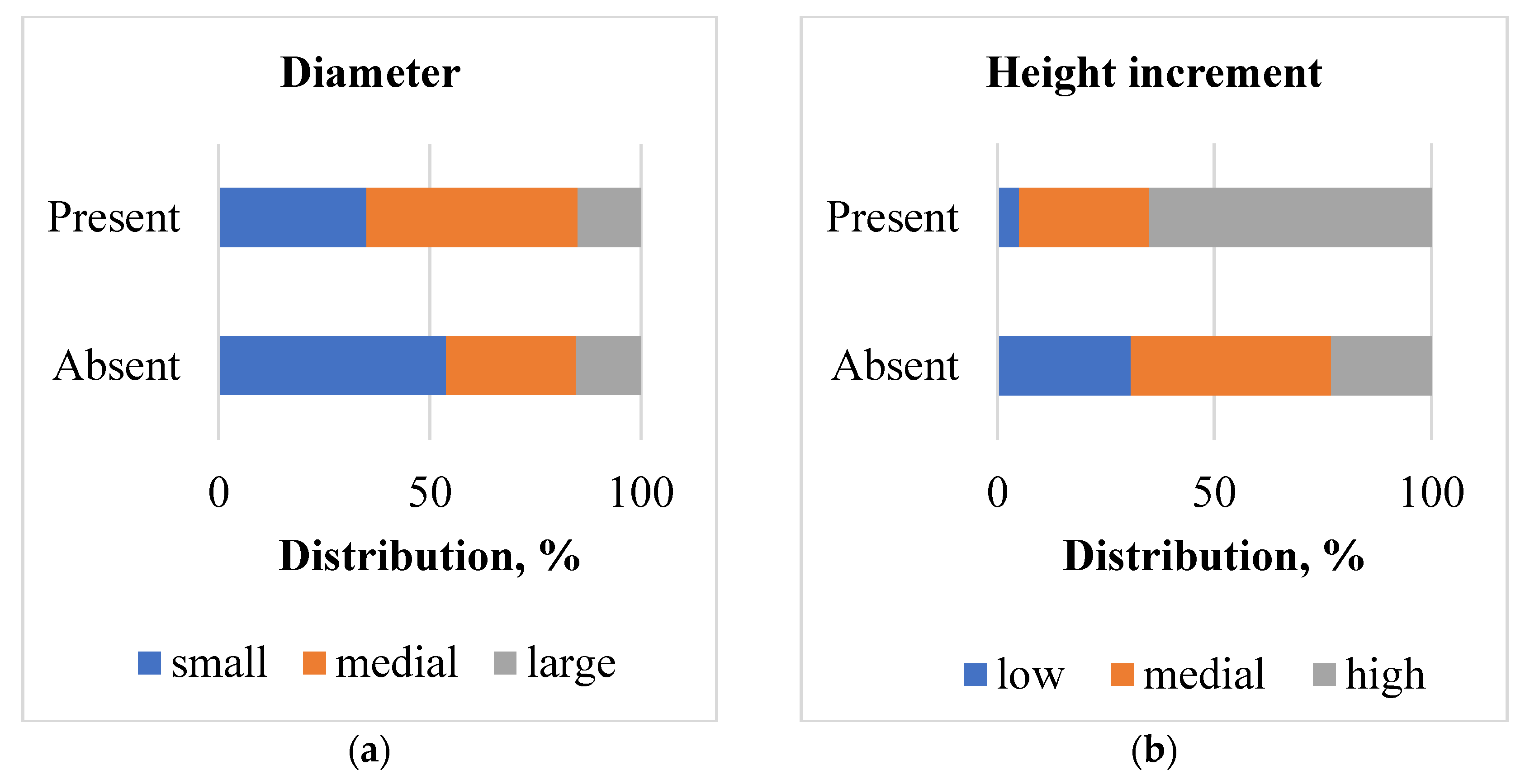
Figure 8.
Proportion of poplar clones, infested by S. carcharias, depending on the group of sections. The columns with the same letters have no significant difference at p=0.05. Sections: A—Aigeiros, T—Tacamahaca, L—Leucoides.
Figure 8.
Proportion of poplar clones, infested by S. carcharias, depending on the group of sections. The columns with the same letters have no significant difference at p=0.05. Sections: A—Aigeiros, T—Tacamahaca, L—Leucoides.
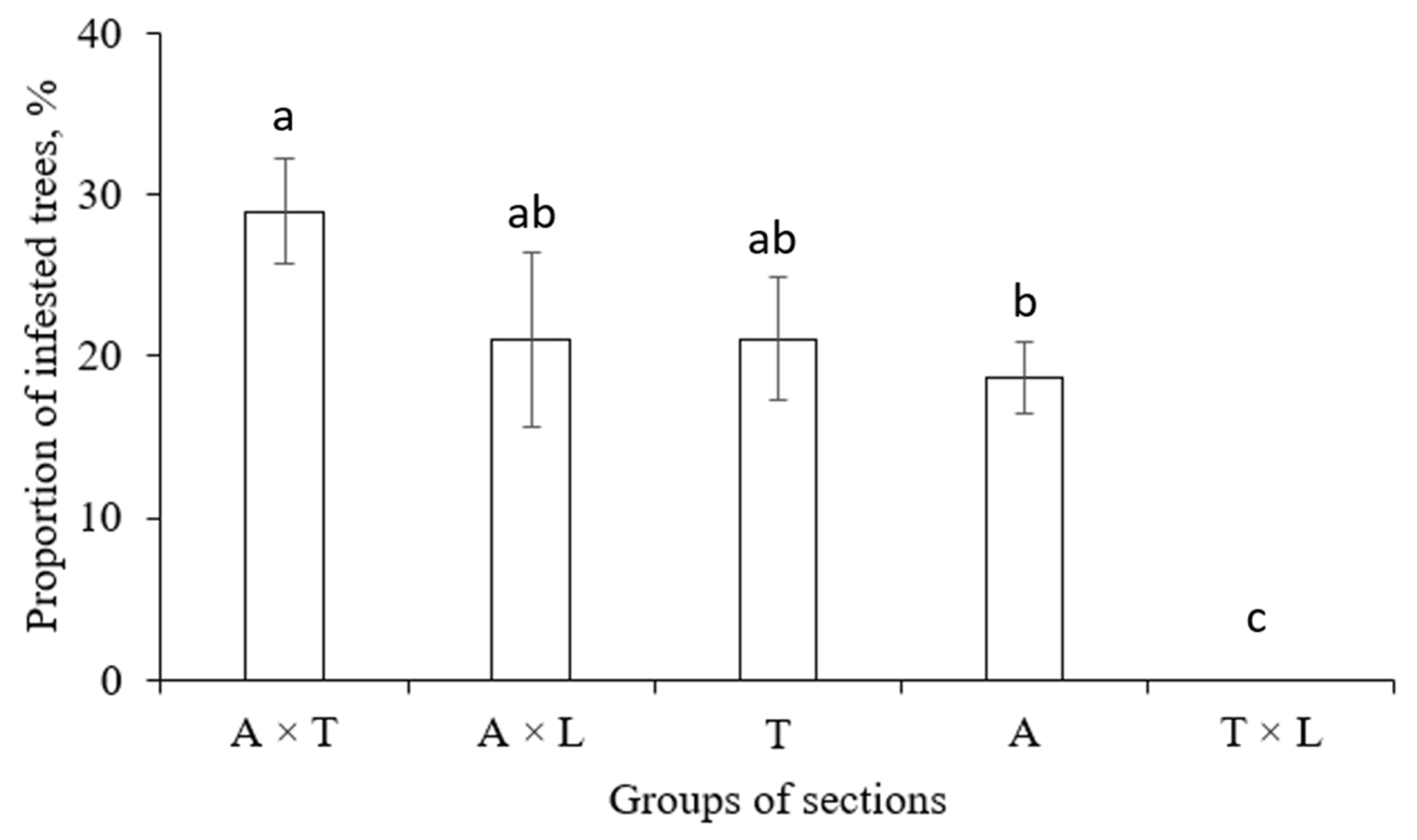
Figure 9.
The proportion of poplar clones, infested by S. carcharias, depending on crossing combinations in the groups of sections: (a)—Section A; (b)—A × L; (c)—A × T; (d)—Section T. The columns with the same letters in parentheses have no significant difference at p=0.05. The abbreviations of crossing combinations are mentioned in Table 1.
Figure 9.
The proportion of poplar clones, infested by S. carcharias, depending on crossing combinations in the groups of sections: (a)—Section A; (b)—A × L; (c)—A × T; (d)—Section T. The columns with the same letters in parentheses have no significant difference at p=0.05. The abbreviations of crossing combinations are mentioned in Table 1.
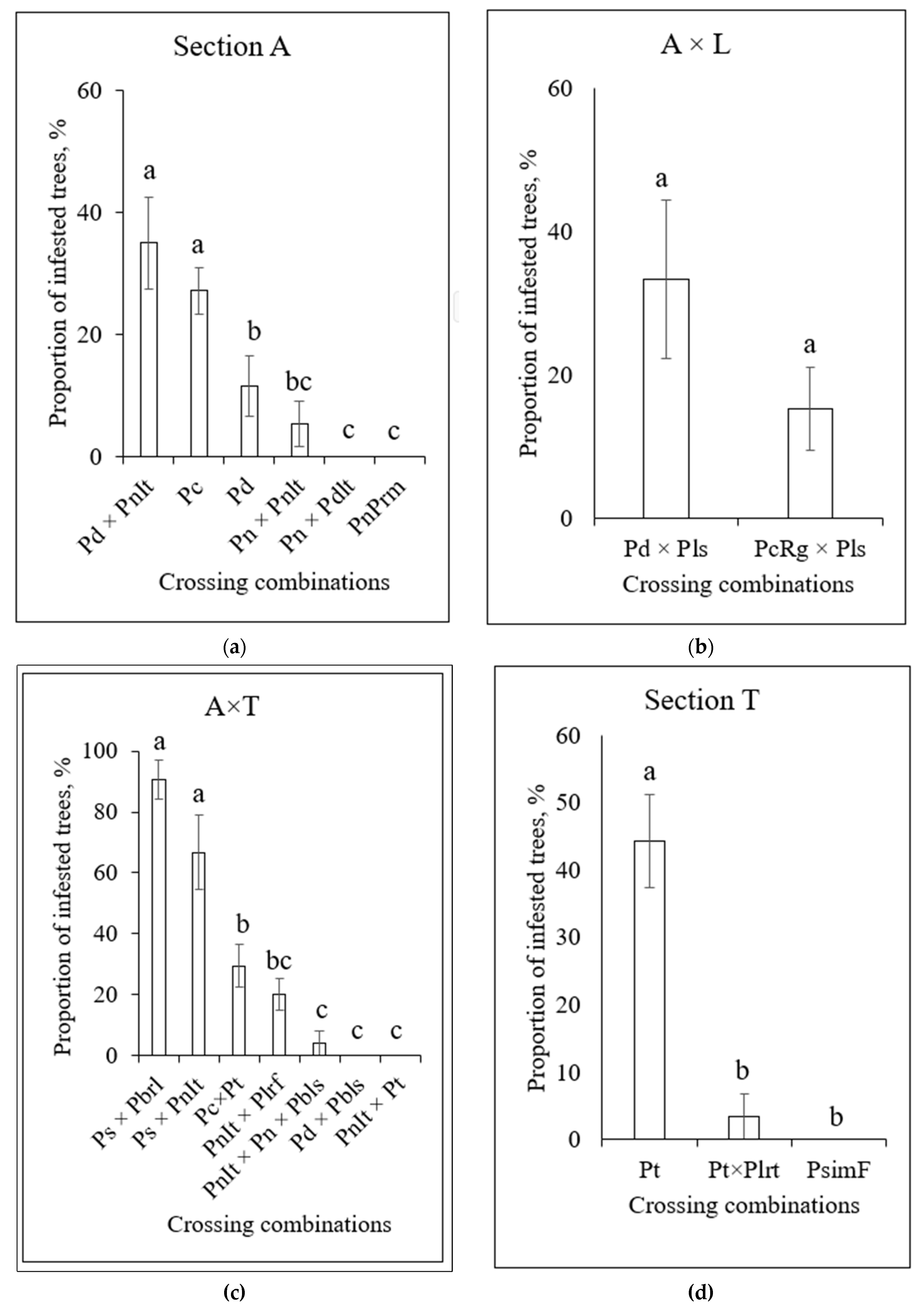

Table 1.
Tested poplar clones.
| Section abbr. | Hybrids / Crossing combination | Clone name | Ref. |
|---|---|---|---|
| A | P. × canadensis (Pc) | ‘Bachelieri’ (Bh), ‘Sakrau79’ (Sk79), ‘Robusta’ (Rob), ‘Tronco’ (Tr), ‘Constanta’ (Cn), ‘Veryla’ (Ve), ‘Sakrau45-51’ (Sk45), ‘Brabantika’ (Br) | [39,45,48] |
| A | P. deltoides (Pd) | ‘Gulliver’ (Gu), ‘Deltopodibna’ (De), ‘Karolinska 162’ (Ka), | [45,48] |
| A | P. deltoides × P. nigra cv. ‘Italica’ (Pd × PnIt) | ‘Strilopodibna’ (St), | [46] |
| A | P. nigra × P. deltoides (Pn × Pd) | ‘Gradizka’ (Gr), ‘Keliberdynska’ (Ke), ‘Robusta 16’ (Rob16) | [39,45] |
| A | P. nigra × P. nigra cv. ‘Italica’ (Pn × PnIt) | ‘Rosijska’ (Ro), ‘Pioner’ (Pi), ‘Addita’ (Ad) | [39,48] |
| A | P. nigra cv. ‘Pyramidalis’ (PnPrm) | ‘Slava Ukrayiny’ (Sl) | [45] |
| T | P. trichocarpa (Pt) | ‘Volosystoplidna’ (Vo), ‘Lada’ (La) | [48] |
| T | P. trichocarpa × P. laurifolia (Pt × Plrf) | ‘Druzhba’ (Dr) | [48] |
| T | P. simonii f. fastigiata (PsimF) | ‘Rohanska’ (Rh) | [48] |
| A × L | P. × canadensis cv. ‘Regenerata’ × P. lasiocarpa (PcRg × Pls) | ‘Perspektyvna’ (Pe) | [45] |
| A × L | P. deltoides × P. lasiocarpa (Pd × Pls) | ‘Udyvytelnaya’ (Udv) | [45] |
| A × T | P. × canadensis × P. trichocarpa (Pc ×Pt) | ‘Lvivska’ (Lv), ‘Mobilna’ (Mo) | [45] |
| A × T | P. deltoides × P. balsamifera (Pd × Pbls) | ‘Kanadska×balsamichna’ (KB) | [48] |
| A × T | P. nigra cv. ‘Italica’ × P. laurifolia (PnIt × Plrf) | ‘Novoberlinska-7’ (N7), ‘Novoberlinska-3’ (N3) | [45,48] |
| A × T | P. nigra cv. ‘Italica’ × P. trichocarpa (PnIt × Pt) | ‘Lubenska’ (Lu) | [45] |
| A × T | (P. nigra cv. ‘Italica’ х P. nigra) × P. balsamifera (PnIt × Pn × Pbls) | ‘Versia’ (Vr) | [45] |
| A × T | P. suaveolens ×P. × berolinensis (Psv × Pbrl) | ‘Ivantiivska’ (Iv) | [46,48] |
| A × T | P. simonii × P. nigra cv. ‘Italica’ (Psim × PnIt) | ‘Kytaiska × pyramidalna’ (Kp) | [46] |
| T × L | P. trichocarpa × P. lasiocarpa (Pt × Pls) | ‘Nocturne’ (No) | [46,48] |
Note: A—Aigeiros, T—Tacamahaca, L—Leucoides.
Table 2.
Climatic indicators for 1990–2020 and the years of field research for Kharkiv meteorological station.
Table 2.
Climatic indicators for 1990–2020 and the years of field research for Kharkiv meteorological station.
| Climatic indicators * | 1990–2020 | 2019 | 2020 | 2021 | 2023 |
|---|---|---|---|---|---|
| Air temperature, T°C | |||||
| – for year | 8.8 | 10.8 | 10.4 | 9.2 | 10.3 |
| – for vegetative period | 17.4 | 19.2 | 17.9 | 17.8 | 18.2 |
| Date of stable transition of temperature | |||||
| – over 5°C | 29/03 | 19/03 | 9/03 | 2/04 | 16/03 |
| – over 10°C | 16/04 | 9/04 | 24/04 | 23/04 | 10/04 |
| Precipitation, mm | |||||
| – for year | 535.2 | 342.3 | 494.6 | 399.0 | 694.1 |
| – for the vegetative period | 287.2 | 159.4 | 285.1 | 217.8 | 354.2 |
| Hydrothermal index** | 0.90 | 0.45 | 0.87 | 0.67 | 1.06 |
*Note: data for 2022 are not full because of military actions; **HTI=10*R/Σt where R is precipitation in millimeters for the period with temperatures above 10°, Σt is the sum of temperatures (°C) for the same time [57].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



