
Submitted:
03 April 2024
Posted:
04 April 2024
You are already at the latest version
Abstract
Neurodegenerative diseases that include Alzheimer's disease (AD), amyotrophic lateral sclerosis (ALS), Parkinson's disease (PD), Huntington's disease (HD), and multiple sclerosis (MS), arise due to numerous causes like protein accumulation and autoimmunity, characterized by neurologic depletion which lead to incapacity in normal physiological function such as thinking and movement in these patients. Glial cells perform a critical role in protective neuronal function; in the case of neuroinflammation, glial cell dysfunction can promote the development of neurodegenerative diseases. MiRNA participates in gene regulation and plays a vital role in many biological processes in the body; in the central nervous system (CNS), it can play an essential part in neural maturation and differentiation. In neurodegenerative diseases, miRNA dysregulation occurs, enhancing the development of these diseases. In this review, we discuss neurodegenerative diseases (Alzheimer's disease (AD), Parkinson's disease (PD), amyotrophic lateral sclerosis (ALS), and multiple sclerosis (MS)) and how miRNA is preserved as a diagnostic biomarker or therapeutic agent in these disorders. Finally, we highlighted miRNA as therapy.
Keywords:
Microglia cell
; neurodegenerative disease
; miRNA
; protein accumulation
Introduction
Neurodegenerative disease is a collective neurological disorder characterized by neuronal degradation that ultimately leads to cognitive and movement disability, includes Alzheimer's disease (AD), amyotrophic lateral sclerosis (ALS), Parkinson's disease (PD), Huntington's disease (HD) and multiple sclerosis (MS) [1,2]. According to the World Health Organization, this disorder could become the second deadliest disease after cancer in the next fifteen years [3]. nervous system is nearly non- regenerative ,thus ; the development of these diseases are permanent[4].
Glial cells, which include microglia, astrocyte, and oligodendrocyte, play a significant role in protective neuronal function and regulation[5]. In neurodegenerative disease, Progression of the neuroinflammatory process occurs predominantly through microglia and astrocyte activation [6].
Neuroinflammation plays an active role in the pathogenesis of different neurodegenerative diseases [7], and microglia activation is a key factor in neuroinflamation[8]. microglia resident macrophage in brain[9] can act as either M1(pro-inflammatory) or M2 (anti-inflammatory) [10]. In the case of neuron impairment, this leads to microglia activation and the release of inflammatory cytokines to improve neuronal damage [11]. However, a neurodegenerative disorder characterized by persistent chronic inflammation will further activate microglia, which indicates more damage to neurons [9], through secreting various inflammatory and cytokine mediators [12]. (Figure 1). CX3CR1 receptor is found on the surface of different cells, including microglial cells [13], that interact with CX3CL1,[14] that found on neurons within globus pallidus, thalamus, striatum, [15], CX3CR1/CX3CL1 which acts as in different biological phenomena in CNS through regulating interaction between microglia, neurons, and immune cell, and participates in different cases of neuropathologies [14].
Astroceyt is one of the glial cells that are abundant in CNS and participate in metabolic supplement for neurons and participate in adjacent cell protection through liberating important chemical messengers such as growth factors, glutamine, lactic acid, and elimination of K+ and increased glutamate[16]. The metabolic exchange between astrocytes, microglia, and neurons mediates chronic inflammation and oxidative stress, leading to neurodegenerative disease progression[16,17].
The essential purpose of oligodendrocytes is myelin generation, and they provide support for neurons and prevent cell death via stimulating myelin recovery[18,19]. Chronic demyelination, such as in (MS) leads to myelin loss due to different pathological mechanisms such as loss of function of oligodendrocyte and mitochondrial dysfunction, oxidative stress enhancement, and at last lead to neuronal cell death. [20].
Early diagnosis of neurodegenerative disease can be helpful in disease treatment and elimination of its possible consequences. Thus one of the biomarkers reliable for this purpose is miRNA [21]
MicroRNAs (miRNAs) which play a significant role in neurodegenerative disease through different physiological process, belongs to non-coding RNAs (ncRNA)[3], and one of the characteristics of ncRNA do not encode specific protein[22]. NcRNA that is present typically in (CNS) that participates in the development and pathogenesis of this system, can be classified as small ncRNA that includes transfer RNA (tRNA), microRNAs (miRNAs), short small interference RNAs(siRNAs), piwi-interacting RNAs (pi-RNA), and long non-coding RNAs that include natural antisense transcript (NATs) and long intragenic RNAs non-coding RNAs (linc RNAs)[23],
MicroRNA with lengths of 21-25 nucleotides plays a critical role in the performance of many diverse biological functions in the cells, such as cell development, stem cell differentiation, oxidative stress[24], and tumor genesis [25]. Several miRNAs circulate in the central nervous system and play an essential role in neural maturation, differentiation, and development [26,27].
miRNA are synthesis from double–strand primary miRNA (pri-miRNA) that is synthesized via RNA polymerase II [28] and undergo more processing in the nucleus to generate precursor miRNA( pre-miRNA) hairpins by Drosha ( type III ribonuclease ) and DiGeorge critical region 8 (DGCR8) (RNA binding protein) [2930]. Then pre-miRNA exists in the cytoplasm through the nuclear pore complex in compensation with exportin-5 (XPO5) and GTPase RanGTP to protect it from the degradation process that occurs through nuclease enzyme[31,32]. In the cytoplasm, Dicer ( type III ribonuclease) leads to cleavage of pre-miRNA to form mature miRNA; this mature miRNA is loaded into Argonaute protein (Ago) into the RNA-induced silencing complex ( RISC), then when binding to the target region on mRNA (3'UTR) lead to suppression this region.[33].in case of any defect occurs in these processes, it can participate in the development of neurodegenerative diseases [35], Figure 2 (c).
In neurodegenerative disease, miRNAs play a significant role in neuronal malfunction through an increased buildup of protein and peptides in the pathogenic form [35]. The dicer enzyme in the nucleus is essential in miRNA synthesis [36]. When knocked out of specific areas in the brain, this leads to removing the dicer enzyme, and specific miRNA will affect and not be made, thus leading to the manifestation of neurodegenerative disorders[37].
This study focuses on the role of miRNAs in participation as diagnostic or prognosis in AD, PD, MS, and ALS or as therapy; Finally, we discuss types of therapeutic miRNA overall.
Role of miRNAs in neurodegenerative diseases.
Many mechanisms can facilitate damaging neurons and play a role in the development of different neurodegenerative diseases (Figure 3). In this section, we discuss AD, PD, MS and ALS and how can preserve miRNA in the neurodegenerative diseases as pathological biomarkers, diagnosis or therapy, Table 1.
Alzheimer's Disease (AD)
Alzheimer's disease (AD) is the most common prevalent neurodegenerative disease affecting aged people[61], and development as mental decline lead to memory problem and difficulty in language and social communication [62,63].
AD occurs due to the accumulation of (amyloid β-peptide )Aβ plaques extracellular in the post-mortem, and neurofibrillary tangles (NFTs) that( composed from abnormal hyperphosphorylated tau-protein) as intracellular [64]. Thus, these triggers lead to changing occurs in synapses and synaptic plasticity in different parts of the brain, such as the neocortex, hippocampus, and limbic system, and lead to cognitive impairment in these patients with AD [65,66,67].
Many other proteins can participate in the pathological of AD, such as cytoplasmic inclusion of TDP-43 accumulation in different parts of the brain hippocampus, amygdala, frontal neocortex, and entorhinal cortex/ inferior temporal cortex[68] and nuclear protein impairments hnRNP A1 and A2/B1 due to neurotoxin –mediated cholinergic impairments, could be lead to defective in processing of miRNA that contribute to pathogenesis and disease development [34,69].
Many investigators informed the role of miRNA in different diseases, such as AD [70], that showed miR-133b could be a promising biomarker for AD patients. Others published that miR-29,miR-181, and miR-9 play a significant role in immune response and inflammation in AD.[71]
There was noticed when overexpression of miR-455-3P in patients with AD this play as a pathological role in AD and can be used as a peripheral biomarker[72]. When Mir-125b overexpression in the brain frontal cortex of AD patients, this accelerated hyperphosphorylation of tau protein brain mice led to memory and learning skills impairments [73] and reported that miR-28-3P, miR-125b, and miR-9 might consider as prospective indicators for AD[74]. As explained previously, Aβ plaques accumulation in the brain, they have found numbers of miRNAs participate in the regulation of Aβ levels, as mir-15 plays a significant role in the regulation of BCL2 [75], while miR-34 downregulates BCL2 translation. BCL2 was associated with AD[76] when increased in a level reduced cognitive impairment progression and Aβ plaques in the AD of mouse model [77]. Mir 106 can be targeted as therapy in AD due to its ability to inhibit phosphorylation of tau protein through inhibition of Aβ1-42 at Tyr18[78]; therefore, when directing this pathway that participated in the pathogenesis of AD, through this miRNA can resort to the function the cell of the brain.
miRNAs can play an essential role in BACE-1 regulation[79] that act as β-secrets 1 that proteolysis of amyloid precursor protein (APP) to liberated Aβ peptide [80], through increasing or decreasing levels of BACE-1 [79], one of these miRNAs, is mir 149 was noted decreased in expression in serum of AD patients and can prospective diagnostic biomarker for AD, in case of overexpression could suppress Aβ accumulation through targeting BACE-1 in AD model cells and improving neuronal viability and can target as therapy for this disease [81].
Parkinson's Disease (PD)
It is a disease that belongs to neurodegenerative disorders, affecting people with age over 65 years [82]. Arise from losing of dopamine neurons in the brain; this dopamine can act as a neurotransmitter, and due to this depletion, leads to movement slowing and reduced balance [83], furthermore was noted formation of inclusions of Lewy and Lewy neuritis bodies in the cytoplasm of a neuronal cell that mainly involved synuclein alpha protein (α-syn) [84]. Less than 20% account for a genetic mutation that participates in PD development that includes Parkin (PARK2), Parkinsonism-associated deglaze (DJ-1), α-synuclein ( SNCA), and leucine-rich repeat kinase 2 ( LRRK2)[85].
PD can advance as pathological and nonpathological progression as non–motor indications can be noted, such as constipation, sleep disorders, and depression[86].
Due to miRNAs play an essential role in biological cellular functions [87]; therefore, they can act as a biomarker in the identification of PD; one of these miRNAs is mir 132; in one study that showed overexpression of miR-132 in plasma samples in males with PD than females in compression to control, and established a potential role of mir 132 in the regulation of Nurr1 protein that participate in dopamine regulation[49]. Other miRNAs contributing to the rule of α-syn are miR-223 and miR-153, noted down-regulated in expression in serum, brain, and saliva of the PD mice model and thus can be considered a diagnostic biomarker in this disease [88]. Other published that miR-153 Increased in level from the extracted tissue of mouse brain and serum [89,90], and was noted a negative correlation between miR-153 and nuclear factor-E2-related 2 (Nrf2) that participate in varied antioxidant genes transcription[91] when inhibited of miR-153 in MPP+ induced PD model, induced Nrf2 signalling pathway and preserve neuron from oxidative stress and can promote new strategy for therapy for PD [90].
Another indicated an increased level of mir 29a and mir 29c in the serum of females than males infected with PD; however, mir 29 significantly down-regulated in PD in comper healthy control, anyway needed further study to be indicated as a biomarker for PD [92].
Mir 375 upregulated in the human spinal motor neurons development and assisted spinal motor neurogenesis [93]; one study demonstrated that mir-375 overexpression improved dopaminergic neurons and inflammation by inhibition of specific protein-1 (SP-1)[94]; this protein act as a transcriptional factor, and when the elevated level in the brain lead to increased neuronal death [95].
miR-216a can inhibit the expression of Bax protein [96]. Bax protein belongs to the Bcl-2 gen family[97]; it acts in case of elevated level as prompter apoptosis and leads to cell death, and when decreased level, indicates inhibition of apoptosis [98]; thus, mir 216a and can potentially target in PD through regulation of Bax protein [96].
Multiple Sclerosis (MS)
Multiple sclerosis (MS) is an autoimmune inflammatory disease characterized by demyelination of the myelin sheath of nerves in the spinal cord and brain; it is a disease of young adult onset[99]. Affecting females more than males [100]. Demyelination formation in white matter occurs due to increased infiltration of T-cells and the release of the cytokine in CNS, which will activate macrophage; this leads B-cells to transform into plasma cells and secreted autoantibodies which attack the myelin sheaths that surround the nerve fibres [101,102].
Many miRNAs have been identified in the different samples of MS patients; one of these, miR-922 and miR-181C in CSF [103], that's associated with transformation from clinically isolated syndrome (CIS) to relapsing-remitting MS (RRMS) characterized by deterioration that initiated via infiltration autoreactive immune cell in CNS [104], and the same association was noted in serum for miR-922 [103]. Mir-181c can control neuroinflammation by reducing microglial activation [105]and diminishing proinflammatory cytokines expressed through microglia [106]. Furthermore, it can indicate miR-181c as a biomarker in an increased inflammatory condition [107].
Mir-150 showed alteration in level in both plasma and CSF after treatment and was used as a biomarker for this disease [108]; mir-155 acts as a proinflammatory activator and plays an essential role in autoimmune diseases [109,110], such as MS pathogenesis; therefore, can be interpreted as one of the miRNAs that use in diagnosis in MS disease [111] Overexpression of miR-21, miR-146a/b in CSF of patients with MS with active lesions can give used as valuable markers in MS patients [112].
Another indicated that Mir-223 participated in remyelination and activation of M2 phagocyte, and in the case of knout mice, led to remyelination impairment [113]. Another study discussed when over-expression of mir-125a-3p contributed to the impairment of oligodendrocyte precursor cell (OPC) maturation, whereas inhibited expression stimulates this cell development [114]; thus, mir-125a-3p overexpression adds to the MS progression, which involvement repair impairment of demyelinated lesions [115].
ALS
Amyotrophic lateral sclerosis (ALS) is a neurodegenerative disease of adult onset that is characterized by muscle paralysis that occurs due to advanced loss of upper and lower motor neurons ,and finally respiratory failure and death through the first five years of diagnosis[116], which can be either familial (fALS ) that associated with high genetic adversity[117] or sporadic ( sALS) that both environmental and genetic factors were involved in development [118].
The emergence of biomarkers for diagnosing ALS is needed; many studies showed the possible detection of miRNAs can help rapidly identify and observe ALS disease [119].
Many studies noted these miRNAs widely disrupted in skeletal muscle and brain and several biofluids in patients with ALS [120,121], such as mir 206, mir 124, mir 181, mir 155 [122,123,124,125]. One study showed that miR-27a-3p in the serum of ALS patients downregulated compared to controls and could indicate miR-27a-3p can serve as a diagnostic biomarker for ALS [126].
Many miRNAs participated in the pathogenesis of ALS; one of these was the downregulation of mir 218-2 due to processing defecting by dicer, which could affect neuronal robustness and, as a result, could be possible targeting therapy in diseases of motor neurons [127]. Fused in sarcoma (FUS) and Transactive response DNA binding protein-43 (TDP-43) that acts as a nuclear protein that participates in RNAs splicing and transcription initiation[128], in nucleus and cytoplasm through Drosha and Dicer, respectively [129], that was noted TDP-43 widely expressed in ALS cases[130]. At the same time, FUS is less common[131]. So, disturbance in these proteins associated with ALS can lead to dysregulation in the miRNA processing mechanism [132], which is one of the possible pathological involved in ALS disease.
A systematic review specified that serum miR-133b,miR-206, and miR-338-3p are prospective ALS markers[133], and miR-206 and miR-133, known as myomiRNA, are identified in skeletal muscle [134]. According to Pegoraro et al.'s study, mir-133 decreased in level in sALS patients, while miR-206 overexpression in muscle specimens in chromosome 9 open reading frame 72 (C9orf72) and superoxide dismutase 1(SOD1) mutations in ALS patients [135]. Mir1- act as muscle differential and Mir 133 act as muscle proliferation[135,136] , while mir-206 decelerates the progression of ALS disease via adding NM synapses regeneration[137]
mir 155 suggested a therapeutic element for treating ALS disease that acts as a proinflammatory activator when inhibited in ALS mice, improving disease progression [138]. Mir-146a, when down-regulated, can lead to malfunction of astrocytes and microglia and could contribute to the degeneration of motor neurons(MN); while upregulation provides a protective effect in ALS patients [139], it also leads to a level of NFL proteins decreased, and this maintenance of neuronal morphology[140].
miRNAs as Therapy
One of the difficulties that prevent transfusion of miRNA is the blood-brain barrier (BBB) that prevents the miRNA from acting appropriately in brain tissues [33]; many promising transporting methods can improve this issue, such as using small extravascular vesicles (sEVs) [141], which can be administrated through intravenous injection [142], stereotactic injection, or nasally administrated,[143,144,145]. Another method of transportation is either a viral vector such as a recombinant adeno-associated virus or adeno-associated virus [146,147], or through a non-viral vector such as gold nanoparticles[148], and many others met such as hydrogel, Carbone-based nanoparticles like nano-diamonds, and smart mesoporous silica nanoparticles[149,150,151], that facilities loading of miRNA in these system to specific site , Figure 2(a).
Several strategies have been developed to use miRNA as a therapeutic agent, such as mimic miRNA, by using synthetic double-strand miRNA, one as a guide and another as a passenger that is linked to another molecule, such as cholesterol, to enhance cellular uptake, lead to restoring of specific miRNA to an average level, that absent or downregulated during disease development and can improvement their specific target protein[152,153], or use (anti-miRNA) miRNA inhibitors, such as using synthetic oligonucleotides that are chemically modified, to target specific miRNA that is characterized in overexpression through targeting some pathway that involved in the miRNA biogenesis, at last, provided functions effectiveness, [154,155,156].
Masking miRNA works to prevent inhibition mRNA from affecting endogenous miRNA [153,157], consisting of single-strand antisense oligonucleotides that interact with the binding site of miRNA in the 3'UTR of the target mRNA[158].
Another type was miRNA sponges that contain multiple binding sites to targeted miRNA that is separated by a few nucleotides, which is typically either viral vector or plasmid and introduced to gen through insertion into 3'UTR of selective gen that is driven by RNA polymerase II promoter [159], Figure 2(b).
There are some limitations of using miRNA as therapy, such as extracellular blockage in case of phagocytosis, degradation by nucleases enzyme, complement opsonization, or through intracellular blocking like non-specific site targeting ineffective cellular taking; regardless of this limitation, miRNA is still ideal targeting as a therapy due to safety, simplicity, effectiveness and easy contributed [160,161] and took multiple advantageous to the improvement of neurodegenerative disease consequences.
Conclusion and Future Perspective
Recently increased evidence of neurodegenerative disease progression, which can affect aged people and young adults. Therefore, these diseases are a tremendous burden on the health organizations of each country, consequently needing emerging tools and biomarkers facilities in the early diagnosis and treatment to decrease the problem issues of these disorders. One of the promising molecules that add to the early diagnosis of this disease and identify possible pathological mechanisms that lead to the development of those disorders through increased or decreased expression, even if it can be used as therapy through many strategies that developed, is miRNAs. Despite related problems to blood-brain briar (BBB) that prevent the miRNA from acting appropriately in brain tissues, many promising transporting methods can improve this problem, such as viral vectors and non-viral vectors [33,162]. So we need increasing investigation of brain-enriched miRNA due to plays a protective or inflammatory role, and by identifying novel pathways that participate in developing this neurodegenerative disease that can assist to considered miRNA-based therapy designed.
Abbreviations---111
AD, Alzheimer`s disease; ALS, amyotrophic lateral sclerosis; PD, Parkinson's disease; HD, Huntington's disease; MS, multiple sclerosis; CX3CL1,fractalkine; miRNAs, MicroRNAs; nc-RNA, non-coding RNAs; pri-miRNA,primary miRNA; pre-miRNA, precursor miRNA; DGCR8, DiGeorge critical region 8;Ago,Argonaute;RISC,RNA-induced silencing complex; NF-KB, nuclear factor-kappa B; FUS, Fused in sarcoma;TDP-43,Transactive response DNA binding protein; C9orf72,chromosome 9 open reading frame 72; SOD1,superoxide dismutase 1; MN, motor neurons; MPP+, 1- mythel-4-phenylpyridinium; DA, dopamine; Aβ, amyloid β-peptide; NFTs, neurofibrillary tangles; CIS, clinically isolated syndrome; RRMS, relapsing remitting multiple sclerosis ; PPMS, primary progressive multiple sclerosis; sEVs , small extravascular vesicle; PGC-1α, peroxisome proliferator-activated receptor-γ coactivator; ROS, reactive oxygen species; ECF, extracellular fluid ; HC, healthy control, ROS; reactive oxygen species.
Author's contribution
This review was theorized by Jalil Tavakol-Afshari. Literature search was performed and was written by Zahraa Alkhazaali-Ali. The manuscript was critically reviewed and edited by Jalil Tavakol-Afshari, Sajad Sahab-Negah, and Amir Reza Boroumand. All authors read and approved the final manuscript.
Acknowledgements
We would like to thank all individuals participate in literature review.
Consent to participate
Not applicable
Ethical Approval
Not applicable
Consent to Publish
Not applicable.
Competing Interests
The authors declare no competing interests.
Data availability
Not applicable
Funding
The authors declare that no funds were received during the preparation of this manuscript.
References
- Gitler, A.D.; Dhillon, P.; Shorter, J. Neurodegenerative disease: models, mechanisms, and a new hope. Dis. Model. Mech. 2017, 10, 499–502. [Google Scholar] [CrossRef] [PubMed]
- De Felice, B.; Mondola, P.; Sasso, A.; Orefice, G.; Bresciamorra, V.; Vacca, G.; Biffali, E.; Borra, M.; Pannone, R. Small non-coding RNA signature in multiple sclerosis patients after treatment with interferon-β. BMC Med Genom. 2014, 7, 1–9. [Google Scholar] [CrossRef]
- Li, S.; Lei, Z.; Sun, T. The role of microRNAs in neurodegenerative diseases: a review. Cell Biol. Toxicol. 2022, 39, 53–83. [Google Scholar] [CrossRef]
- Heemels M-T Neurodegenerative diseases. Nature 2016, 539, 179–180. [CrossRef] [PubMed]
- Ettle, B.; Schlachetzki, J.C.M.; Winkler, J. Oligodendroglia and myelin in neurodegenerative diseases: more than just bystanders? Mol Neurobiol 2016, 53, 3046–3062. [Google Scholar] [CrossRef]
- Heneka, M.T.; Kummer, M.P.; Latz, E. Innate immune activation in neurodegenerative disease. Nat. Rev. Immunol. 2014, 14, 463–477. [Google Scholar] [CrossRef]
- Marogianni, C.; Sokratous, M.; Dardiotis, E.; Hadjigeorgiou, G.M.; Bogdanos, D.; Xiromerisiou, G. Neurodegeneration and Inflammation—An Interesting Interplay in Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 8421. [Google Scholar] [CrossRef]
- Tang, Y.; Le, W. Differential Roles of M1 and M2 Microglia in Neurodegenerative Diseases. Mol. Neurobiol. 2016, 53, 1181–1194. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Dong, X.; Zheng, D.; Nao, J. MiR-124 and the Underlying Therapeutic Promise of Neurodegenerative Disorders. Front. Pharmacol. 2020, 10, 1555. [Google Scholar] [CrossRef]
- Essandoh, K.; Li, Y.; Huo, J. ; Fan G-C MiRNA-mediated macrophage polarization and its potential role in the regulation of inflammatory response. Shock 2016, 46, 122. [Google Scholar] [CrossRef]
- Colonna, M.; Butovsky, O. Microglia Function in the Central Nervous System During Health and Neurodegeneration. Annu. Rev. Immunol. 2017, 35, 441–468. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y. ; Holtzman DM Interplay between innate immunity and Alzheimer disease: APOE and TREM2 in the spotlight. Nat Rev Immunol 2018, 18, 759–772. [Google Scholar] [CrossRef] [PubMed]
- Chamera, K.; Trojan, E.; Szuster-Głuszczak, M.; Basta-Kaim, A. The Potential Role of Dysfunctions in Neuron-Microglia Communication in the Pathogenesis of Brain Disorders. Curr. Neuropharmacol. 2020, 18, 408–430. [Google Scholar] [CrossRef] [PubMed]
- Poniatowski. A.; Wojdasiewicz, P.; Krawczyk, M.; Szukiewicz, D.; Gasik, R.; Kubaszewski,.; Kurkowska-Jastrzębska, I. Analysis of the Role of CX3CL1 (Fractalkine) and Its Receptor CX3CR1 in Traumatic Brain and Spinal Cord Injury: Insight into Recent Advances in Actions of Neurochemokine Agents. Mol. Neurobiol. 2016, 54, 2167–2188. [Google Scholar] [CrossRef]
- Tarozzo, G.; Bortolazzi, S.; Crochemore, C.; Chen, S.; Lira, A.; Abrams, J.; Beltramo, M. Fractalkine protein localization and gene expression in mouse brain. J. Neurosci. Res. 2003, 73, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yuan, Z.; Yang, S.; Zhu, Y.; Xue, M.; Zhang, J.; Leng, L. Brain Energy Metabolism: Astrocytes in Neurodegenerative Diseases. CNS Neurosci. Ther. 2022, 29, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Tcw, J.; Qian, L.; Pipalia, N.H.; Chao, M.J.; Liang, S.A.; Shi, Y.; Jain, B.R.; Bertelsen, S.E.; Kapoor, M.; Marcora, E.; et al. Cholesterol and matrisome pathways dysregulated in astrocytes and microglia. Cell 2022, 185, 2213–2233. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, F.; Huang, N.; et al. Oligodendrocytes and myelin: Active players in neurodegenerative brains? Dev Neurobiol 2022, 82, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Rezzonico, M.G.; Friedman, B.A.; Huntley, M.H.; Meilandt, W.J.; Pandey, S.; Chen, Y.-J.J.; Easton, A.; Modrusan, Z.; Hansen, D.V.; et al. TREM2-independent oligodendrocyte, astrocyte, and T cell responses to tau and amyloid pathology in mouse models of Alzheimer disease. Cell Rep. 2021, 37, 110158. [Google Scholar] [CrossRef]
- Correale, J.; Marrodan, M. ; Ysrraelit MC Mechanisms of neurodegeneration and axonal dysfunction in progressive multiple sclerosis. Biomedicines 2019, 7, 14. [Google Scholar] [CrossRef]
- V. V.; I.T.; Vasu, M.M.; A Poovathinal, S.; Anitha, A. miRNAs as biomarkers of neurodegenerative disorders. Biomarkers Med. 2017, 11, 151–167. [Google Scholar] [CrossRef]
- Szelągowski, A.; Kozakiewicz, M. A Glance at Biogenesis and Functionality of MicroRNAs and Their Role in the Neuropathogenesis of Parkinson’s Disease. Oxidative Med. Cell. Longev. 2023, 2023, 1–18. [Google Scholar] [CrossRef]
- Rege, S.D.; Geetha, T.; Pondugula, S.R.; Zizza, C.A.; Wernette, C.M.; Babu, J.R. Noncoding RNAs in Neurodegenerative Diseases. ISRN Neurol. 2013, 2013, 1–5. [Google Scholar] [CrossRef]
- Cabezas, R.; Baez-Jurado, E.; Hidalgo-Lanussa, O.; Echeverria, V.; Ashrad, G.M.; Sahebkar, A.; Barreto, G.E. Growth Factors and Neuroglobin in Astrocyte Protection Against Neurodegeneration and Oxidative Stress. Mol. Neurobiol. 2019, 56, 2339–2351. [Google Scholar] [CrossRef]
- Jovanovic, M. ; Hengartner MO miRNAs and apoptosis: RNAs to die for. Oncogene 2006, 25, 6176–6187. [Google Scholar] [CrossRef]
- Sharma, S. ; Lu H-C microRNAs in neurodegeneration: current findings and potential impacts. 2018. [Google Scholar]
- Cho, K.H.T.; Xu, B.; Blenkiron, C.; Fraser, M. Emerging Roles of miRNAs in Brain Development and Perinatal Brain Injury. Front. Physiol. 2019, 10, 227. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.-H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef]
- Lee, Y.; Ahn, C.; Han, J.; et al. The nuclear RNase III Drosha initiates microRNA processing. Nature 2003, 425, 415–419. [Google Scholar] [CrossRef]
- Gregory, R.I.; Yan, K.-P.; Amuthan, G.; Chendrimada, T.; Doratotaj, B.; Cooch, N.; Shiekhattar, R. The Microprocessor complex mediates the genesis of microRNAs. Nature 2004, 432, 235–240. [Google Scholar] [CrossRef]
- Okada, C.; Yamashita, E.; Lee, S.J.; et al. A high-resolution structure of the pre-microRNA nuclear export machinery. Science 2009, 326, 1275–1279. [Google Scholar] [CrossRef]
- Zeng, Y. ; Cullen BR Structural requirements for pre-microRNA binding and nuclear export by Exportin 5. Nucleic Acids Res 2004, 32, 4776–4785. [Google Scholar] [CrossRef]
- Roy, B.; Lee, E.; Li, T.; Rampersaud, M. Role of miRNAs in Neurodegeneration: From Disease Cause to Tools of Biomarker Discovery and Therapeutics. Genes 2022, 13, 425. [Google Scholar] [CrossRef]
- Weng, Y.-T.; Chang, Y.-M.; Chern, Y. The Impact of Dysregulated microRNA Biogenesis Machinery and microRNA Sorting on Neurodegenerative Diseases. Int. J. Mol. Sci. 2023, 24, 3443. [Google Scholar] [CrossRef]
- Junn, E. ; Mouradian MM MicroRNAs in neurodegenerative diseases and their therapeutic potential. Pharmacol Ther 2012, 133, 142–150. [Google Scholar] [CrossRef]
- Song M-S, Rossi JJ Molecular mechanisms of Dicer: endonuclease and enzymatic activity. Biochem J 2017, 474, 1603–1618. [CrossRef]
- Chmielarz, P.; Konovalova, J.; Najam, S.S.; Alter, H.; Piepponen, P.; Erfle, H.; Sonntag, K.-C.; Schütz, G.; Vinnikov, I.; Domanskyi, A. Dicer and microRNAs protect adult dopamine neurons. Cell Death Dis. 2017, 8, e2813–e2813. [Google Scholar] [CrossRef]
- Saeidi, N.; Goudarzvand, H.; Mohammadi, H.; Mardi, A.; Ghoreishizadeh, S.; Shomali, N.; Goudarzvand, M. Dysregulation of miR-193a serves as a potential contributor to MS pathogenesis via affecting RhoA and Rock1. Mult. Scler. Relat. Disord. 2022, 69, 104468. [Google Scholar] [CrossRef]
- Martín, M.M.-S.; Gómez, I.; Quiroga-Varela, A.; Río, M.G.-D.; Cedeño, R.R.; Álvarez, G.; Buxó, M.; Miguela, A.; Villar, L.M.; Castillo-Villalba, J.; et al. miRNA Signature in CSF From Patients With Primary Progressive Multiple Sclerosis. Neurol. - Neuroimmunol. Neuroinflammation 2023, 10. [Google Scholar] [CrossRef]
- Saeidi, N.N.; Dabiri, A.; Mansouri, R.; Moomivand, A.; Goudarzvand, M. miRNAs as Valuable Diagnostic Biomarkers in Patients with Multiple Sclerosis. J. Biomed. Res. Environ. Sci. 2023, 4, 773–778. [Google Scholar] [CrossRef]
- Abolghasemi, M.; Ashrafi, S.A.; Asadi, M.; et al. 2022; -10.
- Unlu, H.T.; Saridas, F.; Taskapilioglu, O.; Cecener, G.; Egeli, U.; Turan, O.F.; Tunca, B.; Zarifoglu, M. Investigation of miR-146a Expression Profiles in Fecal Samples of Patients With Multiple Sclerosis for Early Diagnosis and Treatment. Neurol. Sci. Neurophysiol. 2023, 40, 81–87. [Google Scholar] [CrossRef]
- Citterio, L.A.; Mancuso, R.; Agostini, S.; Meloni, M.; Clerici, M. Serum and Exosomal miR-7-1-5p and miR-223-3p as Possible Biomarkers for Parkinson’s Disease. Biomolecules 2023, 13, 865. [Google Scholar] [CrossRef]
- Cao, X.-Y.; Lu, J.-M.; Zhao, Z.-Q.; Li, M.-C.; Lu, T.; An, X.-S.; Xue, L.-J. MicroRNA biomarkers of Parkinson’s disease in serum exosome-like microvesicles. Neurosci. Lett. 2017, 644, 94–99. [Google Scholar] [CrossRef]
- Dos Santos, M.C.T.; Barreto-Sanz, M.A.; Correia, B.R.S.; et al. miRNA-based signatures in cerebrospinal fluid as potential diagnostic tools for early stage Parkinson’s disease. Oncotarget 2018, 9, 17455. [Google Scholar] [CrossRef]
- Zago, E.; Dal Molin, A.; Dimitri, G.M.; et al. Early downregulation of hsa-miR-144-3p in serum from drug-naïve Parkinson’s disease patients. Sci Rep 2022, 12, 1330. [Google Scholar] [CrossRef]
- Chen, L.; Yang, J.; Lü, J.; et al. Identification of aberrant circulating mi RNA s in Parkinson’s disease plasma samples. Brain Behav 2018, 8, e00941. [Google Scholar] [CrossRef]
- Kim, J.; Fiesel, F.C.; Belmonte, K.C.; Hudec, R.; Wang, W.-X.; Kim, C.; Nelson, P.T.; Springer, W.; Kim, J. miR-27a and miR-27b regulate autophagic clearance of damaged mitochondria by targeting PTEN-induced putative kinase 1 (PINK1). Mol. Neurodegener. 2016, 11, 1–16. [Google Scholar] [CrossRef]
- Yang, Z.; Li, T.; Li, S.; Wei, M.; Qi, H.; Shen, B.; Chang, R.C.-C.; Le, W.; Piao, F. Altered Expression Levels of MicroRNA-132 and Nurr1 in Peripheral Blood of Parkinson’s Disease: Potential Disease Biomarkers. ACS Chem. Neurosci. 2019, 10, 2243–2249. [Google Scholar] [CrossRef]
- Baumert, B.; Sobuś, A.; Gołąb-Janowska, M.; Ulańczyk, Z.; Paczkowska, E.; Łuczkowska, K.; Zawiślak, A.; Milczarek, S.; Osękowska, B.; Meller, A.; et al. Local and Systemic Humoral Response to Autologous Lineage-Negative Cells Intrathecal Administration in ALS Patients. Int. J. Mol. Sci. 2020, 21, 1070. [Google Scholar] [CrossRef]
- Russell, A.P.; Wada, S.; Vergani, L.; Hock, M.B.; Lamon, S.; Léger, B.; Ushida, T.; Cartoni, R.; Wadley, G.D.; Hespel, P.; et al. Disruption of skeletal muscle mitochondrial network genes and miRNAs in amyotrophic lateral sclerosis. Neurobiol. Dis. 2013, 49, 107–117. [Google Scholar] [CrossRef]
- Kong, Y.; Li, S.; Zhang, M.; Xu, W.; Chen, Q.; Zheng, L.; Liu, P.; Zou, W. Acupuncture Ameliorates Neuronal Cell Death, Inflammation, and Ferroptosis and Downregulated miR-23a-3p After Intracerebral Hemorrhage in Rats. J. Mol. Neurosci. 2021, 71, 1863–1875. [Google Scholar] [CrossRef]
- Takahashi, I.; Hama, Y.; Matsushima, M.; Hirotani, M.; Kano, T.; Hohzen, H.; Yabe, I.; Utsumi, J.; Sasaki, H. Identification of plasma microRNAs as a biomarker of sporadic Amyotrophic Lateral Sclerosis. Mol. Brain 2015, 8, 1–9. [Google Scholar] [CrossRef]
- Alexandrov, P.N.; Dua, P.; Hill, J.M.; Bhattacharjee, S.; Zhao, Y.; Lukiw, W.J. microRNA (miRNA) speciation in Alzheimer’s disease (AD) cerebrospinal fluid (CSF) and extracellular fluid (ECF). Int. J. Biochem. Mol. Biol. 2012, 3, 365–373. [Google Scholar]
- Lukiw, W.J. ; Pogue AI Vesicular transport of encapsulated microRNA between glial and neuronal cells. Int J Mol Sci 2020, 21, 5078. [Google Scholar] [CrossRef]
- Kumar, S.; Reddy, A.P.; Yin, X. ; Reddy PH Novel MicroRNA-455-3p and its protective effects against abnormal APP processing and amyloid beta toxicity in Alzheimer’s disease. Biochim Biophys Acta (BBA)-Molecular Basis Dis 2019, 1865, 2428–2440. [Google Scholar] [CrossRef]
- Kumar, S. ; Reddy PH A new discovery of MicroRNA-455-3p in Alzheimer’s disease. J Alzheimer’s Dis 2019, 72, S117–S130. [Google Scholar] [CrossRef]
- Zhao, Y.; Pogue, A.I. ; Lukiw WJ MicroRNA (miRNA) signaling in the human CNS in sporadic Alzheimer’s disease (AD)-novel and unique pathological features. Int J Mol Sci 2015, 16, 30105–30116. [Google Scholar] [CrossRef]
- Fan, W.; Liang, C.; Ou, M.; Zou, T.; Sun, F.; Zhou, H.; Cui, L. MicroRNA-146a Is a Wide-Reaching Neuroinflammatory Regulator and Potential Treatment Target in Neurological Diseases. Front. Mol. Neurosci 2020, 13, 90. [Google Scholar] [CrossRef]
- Zhao, Y.; Bhattacharjee, S.; Jones, B.M.; et al. Regulation of neurotropic signaling by the inducible, NF-kB-sensitive miRNA-125b in Alzheimer’s disease (AD) and in primary human neuronal-glial (HNG) cells. Mol Neurobiol 2014, 50, 97–106. [Google Scholar] [CrossRef]
- Andersen, E.; Casteigne, B.; Chapman, W.D.; et al. Diagnostic biomarkers in Alzheimer’s disease. Biomarkers in Neuropsychiatry 2021, 5, 100041. [Google Scholar] [CrossRef]
- McKhann, G.; Drachman, D.; Folstein, M.; Katzman, R.; Price, D.; Stadlan, E.M. Clinical diagnosis of Alzheimer's disease: Report of the NINCDS—ADRDA Work Group under the auspices of Department of Health and Human Services Task Force on Alzheimer's Disease. Neurology 2011, 77, 333–333. [Google Scholar] [CrossRef]
- Mayeux, R.; Stern, Y. Epidemiology of Alzheimer disease. Cold Spring Harb Perspect Med 2012, 2, a006239. [Google Scholar] [CrossRef]
- Hyman, B.T.; Phelps, C.H.; Beach, T.G.; Bigio, E.H.; Cairns, N.J.; Carrillo, M.C.; Dickson, D.W.; Duyckaerts, C.; Frosch, M.P.; Masliah, E.; et al. National Institute on Aging–Alzheimer's Association guidelines for the neuropathologic assessment of Alzheimer's disease. Alzheimer's Dement. 2012, 8, 1–13. [Google Scholar] [CrossRef]
- Masliah, E.; Crews, L.; Hansen, L. Synaptic remodeling during aging and in Alzheimer's disease. J. Alzheimer's Dis. 2006, 9, 91–99. [Google Scholar] [CrossRef]
- Pennanen, C.; Kivipelto, M.; Tuomainen, S.; Hartikainen, P.; Hänninen, T.; Laakso, M.P.; Hallikainen, M.; Vanhanen, M.; Nissinen, A.; Helkala, E.-L.; et al. Hippocampus and entorhinal cortex in mild cognitive impairment and early AD. Neurobiol. Aging 2004, 25, 303–310. [Google Scholar] [CrossRef]
- DeKosky, S.T.; Scheff, S.W. ; Styren SD Structural correlates of cognition in dementia: quantification and assessment of synapse change. Neurodegeneration 1996, 5, 417–421. [Google Scholar] [CrossRef]
- Katsumata, Y.; Fardo, D.W.; Kukull, W.A. ; Nelson PT Dichotomous scoring of TDP-43 proteinopathy from specific brain regions in 27 academic research centers: associations with Alzheimer’s disease and cerebrovascular disease pathologies. Acta Neuropathol Commun 2018, 6, 1–11. [Google Scholar] [CrossRef]
- Berson, A.; Barbash, S.; Shaltiel, G.; Goll, Y.; Hanin, G.; Greenberg, D.S.; Ketzef, M.; Becker, A.J.; Friedman, A.; Soreq, H. Cholinergic-associated loss of hnRNP-A/B in Alzheimer's disease impairs cortical splicing and cognitive function in mice. EMBO Mol. Med. 2012, 4, 730–742. [Google Scholar] [CrossRef]
- Yang, Q.; Zhao, Q.; Yin, Y. miR-133b is a potential diagnostic biomarker for Alzheimer's disease and has a neuroprotective role. Exp. Ther. Med. 2019, 18, 2711–2718. [Google Scholar] [CrossRef]
- Dorval, V.; Nelson, P.T. ; Hébert SS Circulating microRNAs in Alzheimer’s disease: the search for novel biomarkers. Front Mol Neurosci 2013, 6, 24. [Google Scholar]
- Kumar, S.; Vijayan, M. ; Reddy PH MicroRNA-455-3p as a potential peripheral biomarker for Alzheimer’s disease. Hum Mol Genet 2017, 26, 3808–3822. [Google Scholar] [CrossRef]
- Banzhaf-Strathmann, J.; Benito, E.; May, S.; Arzberger, T.; Tahirovic, S.; Kretzschmar, H.; Fischer, A.; Edbauer, D. MicroRNA-125b induces tau hyperphosphorylation and cognitive deficits in Alzheimer's disease. EMBO J. 2014, 33, 1667–1680. [Google Scholar] [CrossRef]
- Hong, H.; Li, Y.; Su, B. Identification of Circulating miR-125b as a Potential Biomarker of Alzheimer’s Disease in APP/PS1 Transgenic Mouse. J. Alzheimer's Dis. 2017, 59, 1449–1458. [Google Scholar] [CrossRef]
- Pan, Y.; Liu, R.; Terpstra, E.; Wang, Y.; Qiao, F.; Wang, J.; Tong, Y.; Pan, B. Dysregulation and Diagnostic Potential of microRNA in Alzheimer’s Disease. J. Alzheimer's Dis. 2015, 49, 1–12. [Google Scholar] [CrossRef]
- Callens, M.; Kraskovskaya, N.; Derevtsova, K.; Annaert, W.; Bultynck, G.; Bezprozvanny, I.; Vervliet, T. The role of Bcl-2 proteins in modulating neuronal Ca2+ signaling in health and in Alzheimer's disease. Biochim. et Biophys. Acta (BBA) - Mol. Cell Res. 2021, 1868, 118997–118997. [Google Scholar] [CrossRef]
- Rohn, T.T.; Vyas, V.; Hernandez-Estrada, T.; Nichol, K.E.; Christie, L.-A.; Head, E. Lack of Pathology in a Triple Transgenic Mouse Model of Alzheimer's Disease after Overexpression of the Anti-Apoptotic Protein Bcl-2. J. Neurosci. 2008, 28, 3051–3059. [Google Scholar] [CrossRef]
- Liu, W.; Zhao, J.; Lu, G. miR-106b inhibits tau phosphorylation at Tyr18 by targeting Fyn in a model of Alzheimer's disease. Biochem. Biophys. Res. Commun. 2016, 478, 852–857. [Google Scholar] [CrossRef]
- Di Meco, A.; Pratico, D. MicroRNAs as therapeutic targets for Alzheimer’s disease. J Alzheimer’s Dis 2016, 53, 367–372. [Google Scholar] [CrossRef]
- Vassar, R. Bace 1: The β-secretase enzyme in alzheimer’s disease. J Mol Neurosci 2004, 23, 105–113. [Google Scholar] [CrossRef]
- Du, W.; Lei, C.; Dong, Y. MicroRNA-149 is downregulated in Alzheimer’s disease and inhibits β-amyloid accumulation and ameliorates neuronal viability through targeting BACE1. Genet. Mol. Biol. 2021, 44, e20200064. [Google Scholar] [CrossRef]
- Kouli, A.; Torsney, K.M. ; Kuan W-L Parkinson’s disease: etiology, neuropathology, and pathogenesis. Exon Publ 2018, 3–26. [Google Scholar]
- Jankovic, J. Parkinson's disease: clinical features and diagnosis. J. Neurol. Neurosurg. Psychiatry 2008, 79, 368–376. [Google Scholar] [CrossRef]
- Narhi, L.; Wood, S.J.; Steavenson, S.; Jiang, Y.; Wu, G.M.; Anafi, D.; Kaufman, S.A.; Martin, F.; Sitney, K.; Denis, P.; et al. Both Familial Parkinson's Disease Mutations Accelerate α-Synuclein Aggregation. J. Biol. Chem. 1999, 274, 9843–9846. [Google Scholar] [CrossRef]
- Alcalay, R.N.; Caccappolo, E.; Mejia-Santana, H.; et al. Frequency of known mutations in early-onset Parkinson disease: implication for genetic counseling: the consortium on risk for early onset Parkinson disease study. Arch Neurol 2010, 67, 1116–1122. [Google Scholar] [CrossRef]
- Chaudhuri, K.R.; Healy, D.G. ; Schapira AH V Non-motor symptoms of Parkinson’s disease: diagnosis and management. Lancet Neurol 2006, 5, 235–245. [Google Scholar] [CrossRef]
- Goh, S.Y.; Chao, Y.X.; Dheen, S.T.; Tan, E.-K.; Tay, S.S.-W. Role of MicroRNAs in Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 5649. [Google Scholar] [CrossRef]
- Cressatti, M.; Juwara, L.; Galindez, J.M.; et al. Salivary microR-153 and microR-223 levels as potential diagnostic biomarkers of idiopathic Parkinson’s disease. Mov Disord 2020, 35, 468–477. [Google Scholar] [CrossRef]
- Gui, Y.; Liu, H.; Zhang, L.; Lv, W.; Hu, X. Altered microRNA profiles in cerebrospinal fluid exosome in Parkinson disease and Alzheimer disease. Oncotarget 2015, 6, 37043–37053. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, S.; Qi, W.; et al. Overexpression of miR-153 promotes oxidative stress in MPP+-induced PD model by negatively regulating the Nrf2/HO-1 signaling pathway. Int J Clin Exp Pathol 2018, 11, 4179. [Google Scholar]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. et Biophys. Acta (BBA) - Mol. Cell Res. 2018, 1865, 721–733. [Google Scholar] [CrossRef]
- Bai, X.; Tang, Y.; Yu, M.; Wu, L.; Liu, F.; Ni, J.; Wang, Z.; Wang, J.; Fei, J.; Wang, W.; et al. Downregulation of blood serum microRNA 29 family in patients with Parkinson’s disease. Sci. Rep. 2017, 7, 1–7. [Google Scholar] [CrossRef]
- Bhinge, A.; Namboori, S.C.; Bithell, A.; Soldati, C.; Buckley, N.J.; Stanton, L.W. MiR-375 is Essential for Human Spinal Motor Neuron Development and May Be Involved in Motor Neuron Degeneration. STEM CELLS 2015, 34, 124–134. [Google Scholar] [CrossRef]
- Cai L-J, Tu, L. ; Li, T.; et al. Up-regulation of microRNA-375 ameliorates the damage of dopaminergic neurons, reduces oxidative stress and inflammation in Parkinson’s disease by inhibiting SP1. Aging 2020, 12, 672.
- Citron, B.A.; Dennis, J.S.; Zeitlin, R.S.; Echeverria, V. Transcription factor Sp1 dysregulation in Alzheimer's disease. J. Neurosci. Res. 2008, 86, 2499–2504. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, M.; Wei, M.; Wang, A.; Deng, Y.; Cao, H. MicroRNA-216a inhibits neuronal apoptosis in a cellular Parkinson’s disease model by targeting Bax. Metab. Brain Dis. 2020, 35, 627–635. [Google Scholar] [CrossRef]
- Reed JC Proapoptotic multidomain Bcl-2/Bax-family proteins: mechanisms, physiological roles, and therapeutic opportunities. Cell death Differ 2006, 13, 1378–1386. [CrossRef]
- De Falco, M.; De Luca, L.; Acanfora, F.; Cavallotti, I.; Cottone, G.; Laforgia, V.; De Luca, B.; Baldi, A.; De Luca, A. Alteration of the Bcl-2:Bax ratio in the placenta as pregnancy proceeds. Histochem. J. 2001, 33, 421–425. [Google Scholar] [CrossRef]
- Cantoni, C.; Ghezzi, L.; Choi, J.; Cross, A.H.; Piccio, L. Targeting miR-223 enhances myeloid-derived suppressor cell suppressive activities in multiple sclerosis patients. Mult. Scler. Relat. Disord. 2023, 76, 104839–104839. [Google Scholar] [CrossRef]
- Orton, S.-M.; Herrera, B.M.; Yee, I.M.; Valdar, W.; Ramagopalan, S.V.; Sadovnick, A.D.; Ebers, G.C.; Canadian Collaborative Study Group. Sex ratio of multiple sclerosis in Canada: a longitudinal study. Lancet Neurol. 2006, 5, 932–936. [Google Scholar] [CrossRef]
- Maciak, K.; Dziedzic, A.; Saluk, J. Remyelination in multiple sclerosis from the miRNA perspective. Front. Mol. Neurosci. 2023, 16, 1199313. [Google Scholar] [CrossRef]
- Engelhardt, B. ; Ransohoff RM The ins and outs of T-lymphocyte trafficking to the CNS: anatomical sites and molecular mechanisms. Trends Immunol 2005, 26, 485–495. [Google Scholar] [CrossRef]
- Ahlbrecht, J.; Martino, F.; Pul, R.; Skripuletz, T.; Sühs, K.-W.; Schauerte, C.; Yildiz. ; Trebst, C.; Tasto, L.; Thum, S.; et al. Deregulation of microRNA-181c in cerebrospinal fluid of patients with clinically isolated syndrome is associated with early conversion to relapsing–remitting multiple sclerosis. Mult. Scler. J. 2016, 22, 1202–1214. [Google Scholar] [CrossRef]
- Harris, V.K.; Tuddenham, J.F. ; Sadiq SA Biomarkers of multiple sclerosis: current findings. Degener Neurol Neuromuscul Dis 2017, 19–29. [Google Scholar]
- Ma, Q.; Zhao, H.; Tao, Z.; Wang, R.; Liu, P.; Han, Z.; Ma, S.; Luo, Y.; Jia, J. MicroRNA-181c Exacerbates Brain Injury in Acute Ischemic Stroke. Aging Dis. 2016, 7, 705–714. [Google Scholar] [CrossRef]
- Zhang, L. ; Dong L-Y, Li Y-J, et al. The microRNA miR-181c controls microglia-mediated neuronal apoptosis by suppressing tumor necrosis factor. J Neuroinflammation 2012, 9, 1–12. [Google Scholar]
- Kramer, S.; Haghikia, A.; Bang, C.; et al. Elevated levels of miR-181c and miR-633 in the CSF of patients with MS: A validation study. e: Neurol Neuroinflammation 2019, 6, 2019. [Google Scholar]
- Bergman, P.; Piket, E.; Khademi, M.; James, T.; Brundin, L.; Olsson, T.; Piehl, F.; Jagodic, M. Circulating miR-150 in CSF is a novel candidate biomarker for multiple sclerosis. Neurol. - Neuroimmunol. Neuroinflammation 2016, 3, e219–e219. [Google Scholar] [CrossRef]
- Moore, C.S.; Rao, V.T.S.; Durafourt, B.A.; et al. miR-155 as a multiple sclerosis–relevant regulator of myeloid cell polarization. Ann Neurol 2013, 74, 709–720. [Google Scholar] [CrossRef]
- Leng, R.-X.; Pan, H.-F.; Qin, W.-Z.; Chen, G.-M.; Ye, D.-Q. Role of microRNA-155 in autoimmunity. Cytokine Growth Factor Rev. 2011, 22, 141–147. [Google Scholar] [CrossRef]
- Maciak, K.; Dziedzic, A.; Miller, E.; Saluk-Bijak, J. miR-155 as an Important Regulator of Multiple Sclerosis Pathogenesis. A Review. Int. J. Mol. Sci. 2021, 22, 4332. [Google Scholar] [CrossRef]
- Martín, M.M.-S.; Reverter, G.; Robles-Cedeño, R.; Buxò, M.; Ortega, F.J.; Gómez, I.; Tomàs-Roig, J.; Celarain, N.; Villar, L.M.; Perkal, H.; et al. Analysis of miRNA signatures in CSF identifies upregulation of miR-21 and miR-146a/b in patients with multiple sclerosis and active lesions. J. Neuroinflammation 2019, 16, 1–10. [Google Scholar] [CrossRef]
- Galloway, D.A.; Blandford, S.N.; Berry, T.; Williams, J.B.; Stefanelli, M.; Ploughman, M.; Moore, C.S. miR-223 promotes regenerative myeloid cell phenotype and function in the demyelinated central nervous system. Glia 2018, 67, 857–869. [Google Scholar] [CrossRef]
- Marangon, D.; Boda, E.; Parolisi, R.; Negri, C.; Giorgi, C.; Montarolo, F.; Perga, S.; Bertolotto, A.; Buffo, A.; Abbracchio, M.P.; et al. In vivo silencing of miR-125a-3p promotes myelin repair in models of white matter demyelination. Glia 2020, 68, 2001–2014. [Google Scholar] [CrossRef]
- Lecca, D.; Marangon, D.; Coppolino, G.T.; Méndez, A.M.; Finardi, A.; Costa, G.D.; Martinelli, V.; Furlan, R.; Abbracchio, M.P. MiR-125a-3p timely inhibits oligodendroglial maturation and is pathologically up-regulated in human multiple sclerosis. Sci. Rep. 2016, 6, srep34503. [Google Scholar] [CrossRef]
- Brusati, A.; Ratti, A.; Pensato, V.; Peverelli, S.; Gentilini, D.; Bella, E.D.; Sorce, M.N.; Meneri, M.; Gagliardi, D.; Corti, S.; et al. Analysis of miRNA rare variants in amyotrophic lateral sclerosis and in silico prediction of their biological effects. Front. Genet. 2022, 13, 1055313. [Google Scholar] [CrossRef]
- Hardiman, O.; Al-Chalabi, A.; Chio, A.; et al. Amyotrophic lateral sclerosis. Nat Rev Dis Prim 2017, 3, 1–19. [Google Scholar] [CrossRef]
- Goutman, A.S.; Hardiman, O.; Al-Chalabi, A.; Chió, A.; Savelieff, M.G.; Kiernan, M.C.; Feldman, E.L. Emerging insights into the complex genetics and pathophysiology of amyotrophic lateral sclerosis. Lancet Neurol. 2022, 21, 465–479. [Google Scholar] [CrossRef]
- Joilin, G.; Leigh, P.N.; Newbury, S.F.; Hafezparast, M. An Overview of MicroRNAs as Biomarkers of, A.L.S. Front. Neurol.
- Rinchetti 2019, P.; Rizzuti, M.; Faravelli, I.; Corti, S. MicroRNA metabolism and dysregulation in amyotrophic lateral sclerosis. Mol Neurobiol 2018, 55, 2617–2630. [Google Scholar] [CrossRef]
- Alvia, M.; Aytan, N.; Spencer, K.R.; Foster, Z.W.; Rauf, N.A.; Guilderson, L.; Robey, I.; Averill, J.G.; Walker, S.E.; Alvarez, V.E.; et al. MicroRNA Alterations in Chronic Traumatic Encephalopathy and Amyotrophic Lateral Sclerosis. Front. Neurosci. 2022, 16, 855096. [Google Scholar] [CrossRef]
- Toivonen, J.M.; Manzano, R.; Oliván, S.; Zaragoza, P.; García-Redondo, A.; Osta, R. MicroRNA-206: A Potential Circulating Biomarker Candidate for Amyotrophic Lateral Sclerosis. PLOS ONE 2014, 9, e89065. [Google Scholar] [CrossRef]
- Vaz, A.R.; Vizinha, D.; Morais, H.; Colaço, A.R.; Loch-Neckel, G.; Barbosa, M.; Brites, D. Overexpression of miR-124 in Motor Neurons Plays a Key Role in ALS Pathological Processes. Int. J. Mol. Sci. 2021, 22, 6128. [Google Scholar] [CrossRef]
- Magen, I.; Yacovzada, N.S.; Yanowski, E.; Coenen-Stass, A.; Grosskreutz, J.; Lu, C.-H.; Greensmith, L.; Malaspina, A.; Fratta, P.; Hornstein, E. Circulating miR-181 is a prognostic biomarker for amyotrophic lateral sclerosis. Nat. Neurosci. 2021, 24, 1534–1541. [Google Scholar] [CrossRef]
- Cunha, C.; Santos, C.; Gomes, C.; Fernandes, A.; Correia, A.M.; Sebastião, A.M.; Vaz, A.R.; Brites, D. Downregulated Glia Interplay and Increased miRNA-155 as Promising Markers to Track ALS at an Early Stage. Mol. Neurobiol. 2017, 55, 4207–4224. [Google Scholar] [CrossRef]
- Xu, Q.; Zhao, Y.; Zhou, X.; Luan, J.; Cui, Y.; Han, J. Comparison of the extraction and determination of serum exosome and miRNA in serum and the detection of miR-27a-3p in serum exosome of ALS patients. Intractable Rare Dis. Res. 2018, 7, 13–18. [Google Scholar] [CrossRef]
- Reichenstein, I.; Eitan, C.; Diaz-Garcia, S.; Haim, G.; Magen, I.; Siany, A.; Hoye, M.L.; Rivkin, N.; Olender, T.; Toth, B.; et al. Human genetics and neuropathology suggest a link between miR-218 and amyotrophic lateral sclerosis pathophysiology. Sci. Transl. Med. 2019, 11. [Google Scholar] [CrossRef]
- Yang, L.; Embree, L.J. ; Hickstein DD TLS-ERG leukemia fusion protein inhibits RNA splicing mediated by serine-arginine proteins. Mol Cell Biol 2000, 20, 3345–3354. [Google Scholar] [CrossRef]
- Kawahara, Y.; Mieda-Sato, A. TDP-43 promotes microRNA biogenesis as a component of the Drosha and Dicer complexes. Proc. Natl. Acad. Sci. 2012, 109, 3347–3352. [Google Scholar] [CrossRef]
- Lagier-Tourenne, C.; Polymenidou, M. ; Cleveland DW TDP-43 and FUS/TLS: emerging roles in RNA processing and neurodegeneration. Hum Mol Genet 2010, 19, R46–R64. [Google Scholar] [CrossRef]
- Ling S-C, Polymenidou, M. ; Cleveland DW Converging mechanisms in ALS and FTD: disrupted RNA and protein homeostasis. Neuron 2013, 79, 416–438.
- Koike, Y.; Onodera, O. Implications of miRNAs dysregulation in amyotrophic lateral sclerosis: Challenging for clinical applications. Front. Neurosci. 2023, 17, 1131758. [Google Scholar] [CrossRef]
- Liu, H.; Lan, S.; Shi, X.-J.; Fan, F.-C.; Liu, Q.-S.; Cong, L.; Cheng, Y. Systematic review and meta-analysis on microRNAs in amyotrophic lateral sclerosis. Brain Res. Bull. 2023, 194, 82–89. [Google Scholar] [CrossRef]
- Di Pietro, L.; Lattanzi, W.; Bernardini, C. Skeletal Muscle MicroRNAs as Key Players in the Pathogenesis of Amyotrophic Lateral Sclerosis. Int. J. Mol. Sci. 2018, 19, 1534. [Google Scholar] [CrossRef]
- Pegoraro, V.; Marozzo, R.; Angelini, C. MicroRNAs and HDAC4 protein expression in the skeletal muscle of ALS patients. Clin. Neuropathol. 2020, 39, 105–114. [Google Scholar] [CrossRef]
- Nie M, Deng Z-L, Liu J, Wang D-Z (2015) Noncoding RNAs, emerging regulators of skeletal muscle development and diseases. Biomed Res Int 2015.
- Williams, A.H.; Valdez, G.; Moresi, V.; Qi, X.; McAnally, J.; Elliott, J.L.; Bassel-Duby, R.; Sanes, J.R.; Olson, E.N. MicroRNA-206 Delays ALS Progression and Promotes Regeneration of Neuromuscular Synapses in Mice. Science 2009, 326, 1549–1554. [Google Scholar] [CrossRef]
- Butovsky, O.; Jedrychowski, M.P.; Cialic, R.; Krasemann, S.; Murugaiyan, G.; Fanek, Z.; Greco, D.J.; Wu, P.M.; Doykan, C.E.; Kiner, O.; et al. Targeting miR-155 restores abnormal microglia and attenuates disease in SOD1 mice. Ann. Neurol. 2014, 77, 75–99. [Google Scholar] [CrossRef]
- Gomes, C.; Sequeira, C.; Likhite, S.; Dennys, C.N.; Kolb, S.J.; Shaw, P.J.; Vaz, A.R.; Kaspar, B.K.; Meyer, K.; Brites, D. Neurotoxic Astrocytes Directly Converted from Sporadic and Familial ALS Patient Fibroblasts Reveal Signature Diversities and miR-146a Theragnostic Potential in Specific Subtypes. Cells 2022, 11, 1186. [Google Scholar] [CrossRef]
- Campos-Melo, D.; Droppelmann, C.A.; He, Z.; Volkening, K.; Strong, M.J. Altered microRNA expression profile in amyotrophic lateral sclerosis: a role in the regulation of NFL mRNA levels. Mol. Brain 2013, 6, 26. [Google Scholar] [CrossRef]
- He, M.; Zhang, H.-N.; Tang, Z.-C.; Gao, S.-G. Diagnostic and Therapeutic Potential of Exosomal MicroRNAs for Neurodegenerative Diseases. Neural Plast. 2021, 2021, 1–13. [Google Scholar] [CrossRef]
- Akbari, A.; Jabbari, N.; Sharifi, R.; Ahmadi, M.; Vahhabi, A.; Seyedzadeh, S.J.; Nawaz, M.; Szafert, S.; Mahmoodi, M.; Jabbari, E.; et al. Free and hydrogel encapsulated exosome-based therapies in regenerative medicine. Life Sci. 2020, 249, 117447. [Google Scholar] [CrossRef]
- Liu, X.; Yang, Y.; Li, Y.; Niu, X.; Zhao, B.; Wang, Y.; Bao, C.; Xie, Z.; Lin, Q.; Zhu, L. Integration of stem cell-derived exosomes with in situ hydrogel glue as a promising tissue patch for articular cartilage regeneration. Nanoscale 2017, 9, 4430–4438. [Google Scholar] [CrossRef]
- Kim, H.J.; Seo, S.W.; Chang, J.W.; Lee, J.I.; Kim, C.H.; Chin, J.; Choi, S.J.; Kwon, H.; Yun, H.J.; Lee, J.M.; et al. Stereotactic brain injection of human umbilical cord blood mesenchymal stem cells in patients with Alzheimer's disease dementia: A phase 1 clinical trial. Alzheimer's Dementia: Transl. Res. Clin. Interv. 2015, 1, 95–102. [Google Scholar] [CrossRef]
- Erdő, F.; Bors, L.A.; Farkas, D.; Bajza, Á.; Gizurarson, S. Evaluation of intranasal delivery route of drug administration for brain targeting. Brain Res. Bull. 2018, 143, 155–170. [Google Scholar] [CrossRef]
- Beutler, A.S.; Reinhardt, M. AAV for pain: steps towards clinical translation. Gene Ther. 2009, 16, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Kaplitt, M.G.; Feigin, A.; Tang, C.; Fitzsimons, H.L.; Mattis, P.; A Lawlor, P.; Bland, R.J.; Young, D.; Strybing, K.; Eidelberg, D.; et al. Safety and tolerability of gene therapy with an adeno-associated virus (AAV) borne GAD gene for Parkinson's disease: an open label, phase I trial. Lancet 2007, 369, 2097–2105. [Google Scholar] [CrossRef] [PubMed]
- Ekin, A.; Karatas, O.F.; Culha, M.; Ozen, M. Designing a gold nanoparticle-based nanocarrier for microRNA transfection into the prostate and breast cancer cells. J. Gene Med. 2014, 16, 331–335. [Google Scholar] [CrossRef]
- Conde, J.; Oliva, N.; Atilano, M.; Song, H.S.; Artzi, N. Self-assembled RNA-triple-helix hydrogel scaffold for microRNA modulation in the tumour microenvironment. Nat. Mater. 2015, 15, 353–363. [Google Scholar] [CrossRef]
- Zhao, J.; Lai, H.; Lu, H.; Barner-Kowollik, C.; Stenzel, M.H.; Xiao, P. Fructose-Coated Nanodiamonds: Promising Platforms for Treatment of Human Breast Cancer. Biomacromolecules 2016, 17, 2946–2955. [Google Scholar] [CrossRef]
- Yu, C.; Qian, L.; Uttamchandani, M.; Li, L.; Yao, S.Q. Single-Vehicular Delivery of Antagomir and Small Molecules to Inhibit miR-122 Function in Hepatocellular Carcinoma Cells by using “Smart” Mesoporous Silica Nanoparticles. Angew. Chem. 2015, 127, 10720–10724. [Google Scholar] [CrossRef]
- Austin TM First microRNA mimic enters clinic. Nat Biotechnol 2013, 31, 577.
- Van Rooij, E.; Kauppinen, S. Development of micro RNA therapeutics is coming of age. EMBO Mol Med 2014, 6, 851–864. [Google Scholar] [CrossRef]
- Rupaimoole, R. ; Slack FJ MicroRNA therapeutics: towards a new era for the management of cancer and other diseases. Nat Rev Drug Discov 2017, 16, 203–222. [Google Scholar] [CrossRef]
- Walayat, A.; Yang, M.; Xiao, D. Therapeutic Implication of miRNA in Human Disease. In Antisense Therapy; Sharad, S., Kapur, S., Eds.; IntechOpen: London, UK, 2019; ISBN 978-1-78984-532-7. [Google Scholar]
- Lennox, K.A. ; Behlke MA Chemical modification and design of anti-miRNA oligonucleotides. Gene Ther 2011, 18, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Li, Z. ; Rana TM Therapeutic targeting of microRNAs: current status and future challenges. Nat Rev Drug Discov 2014, 13, 622–638. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z. The principles of MiRNA-masking antisense oligonucleotides technology. MicroRNA Cancer Methods Protoc 2011, 43–49. [Google Scholar]
- Ebert, M.S. ; Sharp PA MicroRNA sponges: progress and possibilities. Rna 2010, 16, 2043–2050. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhao, H.; Tan, Z.; et al. Bottleneck limitations for microRNA-based therapeutics from bench to the bedside. Die Pharm Int J Pharm Sci 2015, 70, 147–154. [Google Scholar]
- Gavrilov, K. ; Saltzman WM Therapeutic siRNA: principles, challenges, and strategies. Yale J Biol Med 2012, 85, 187. [Google Scholar]
- Yang, N. An overview of viral and nonviral delivery systems for microRNA. Int. J. Pharm. Investig. 2015, 5, 179–81. [Google Scholar] [CrossRef]
Figure 1.
Microglia and principle activation in the brain. Microglia normally present in a resting state that contains toll-like receptor (TLR) and CX3CR1, and due to activation such as in case of ageing, pathogen, and depression of dead cell lead to activation of microglia through TLR and CX3CR1; to become either (M1) that secreted proinflammatory cytokine-like tumor necrotic factor-α (TNF-α), interleukin-6 (IL-6) , reactive oxygen species (ROS) , or (M2) that secreted anti-inflammatory cytokine that have protect effect for example (IL-4, IL-10). In case of persist activation of microglia, this enhancing the persistent secretion of the pro-inflammatory cytokine, increased T-cell activation, which in turn activation of microglia that will affect astrocyte activation and loss of oligodendrocyte function; all this contributes to the neuronal death. This Fig was created by using (Adobe Illustrator 2020).
Figure 1.
Microglia and principle activation in the brain. Microglia normally present in a resting state that contains toll-like receptor (TLR) and CX3CR1, and due to activation such as in case of ageing, pathogen, and depression of dead cell lead to activation of microglia through TLR and CX3CR1; to become either (M1) that secreted proinflammatory cytokine-like tumor necrotic factor-α (TNF-α), interleukin-6 (IL-6) , reactive oxygen species (ROS) , or (M2) that secreted anti-inflammatory cytokine that have protect effect for example (IL-4, IL-10). In case of persist activation of microglia, this enhancing the persistent secretion of the pro-inflammatory cytokine, increased T-cell activation, which in turn activation of microglia that will affect astrocyte activation and loss of oligodendrocyte function; all this contributes to the neuronal death. This Fig was created by using (Adobe Illustrator 2020).
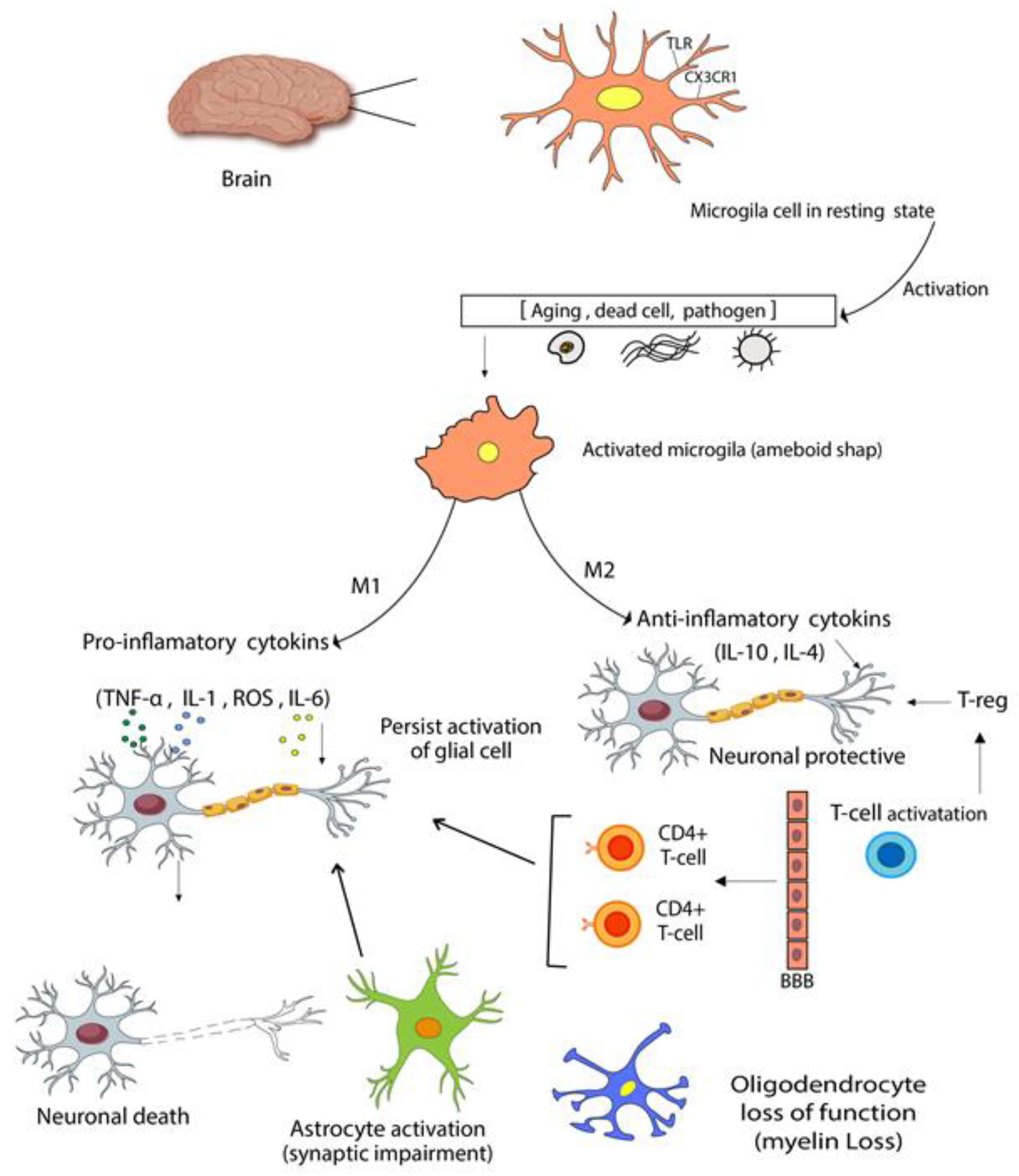
Figure 2.
Schematic diagram of miRNA's transportation system, therapeutic, and machinery synthesis. (A) Showed transportation system, miRNA can be transported to cells inside the virus-like particles, lipid nanoparticles and exosomes. (B) Therapeutics miRNA can be a synthetic double or single strand that targets miRNA syntheses in the nucleus or cytoplasm, like (mimic miRNA, anti-miRNA, masking miRNA). (C) Synthesis of miRNA starts in the nucleus, pri-miRNA transcript from RNA polymerase II; after that, pre-miRNA formed through the processing of pri-miRNA by dorsha and DGCR8, pre-miRNA exported into the cytoplasm through nuclear pore in combination with XPO5, RanGTP, dicer lead to form mature miRNA in the cytoplasm, that mature miRNA loaded to (AGO), the unwanted stranded degraded, the functional strand loaded to RISC and lastly targeted complementary region 3' UTR on mRNA, which prompts mRNA degradation or block translation, this Fig. created by using (Adobe Illustrator 2020).
Figure 2.
Schematic diagram of miRNA's transportation system, therapeutic, and machinery synthesis. (A) Showed transportation system, miRNA can be transported to cells inside the virus-like particles, lipid nanoparticles and exosomes. (B) Therapeutics miRNA can be a synthetic double or single strand that targets miRNA syntheses in the nucleus or cytoplasm, like (mimic miRNA, anti-miRNA, masking miRNA). (C) Synthesis of miRNA starts in the nucleus, pri-miRNA transcript from RNA polymerase II; after that, pre-miRNA formed through the processing of pri-miRNA by dorsha and DGCR8, pre-miRNA exported into the cytoplasm through nuclear pore in combination with XPO5, RanGTP, dicer lead to form mature miRNA in the cytoplasm, that mature miRNA loaded to (AGO), the unwanted stranded degraded, the functional strand loaded to RISC and lastly targeted complementary region 3' UTR on mRNA, which prompts mRNA degradation or block translation, this Fig. created by using (Adobe Illustrator 2020).

Figure 3.
Schematics overview of normal and damaging neurons. (A); showed normal constituents of neuron form cell body with usually present (nucleus, mitochondria,) to axon and how is generally warped in the myelin sheath, the axonal terminal that consists of normal function of telodendria and synaptic terminals, which ads in the neurotransmitter. (B) showed damaged neuron that occurs due to different mechanisms such as mitochondrial dysfunction, DNA damage, accumulation of different protein such as (TDP-43, Lewy body, Aβ plaques ) in the cell body and around the neuronal cell, axonal transport loss such as (neurofilaments deposition, demyelination) which boost losing of the axonal terminal, all of these mechanisms participate in the loss of normal function of neurons, this Fig. created by using (Adobe Illustrator 2020).
Figure 3.
Schematics overview of normal and damaging neurons. (A); showed normal constituents of neuron form cell body with usually present (nucleus, mitochondria,) to axon and how is generally warped in the myelin sheath, the axonal terminal that consists of normal function of telodendria and synaptic terminals, which ads in the neurotransmitter. (B) showed damaged neuron that occurs due to different mechanisms such as mitochondrial dysfunction, DNA damage, accumulation of different protein such as (TDP-43, Lewy body, Aβ plaques ) in the cell body and around the neuronal cell, axonal transport loss such as (neurofilaments deposition, demyelination) which boost losing of the axonal terminal, all of these mechanisms participate in the loss of normal function of neurons, this Fig. created by using (Adobe Illustrator 2020).
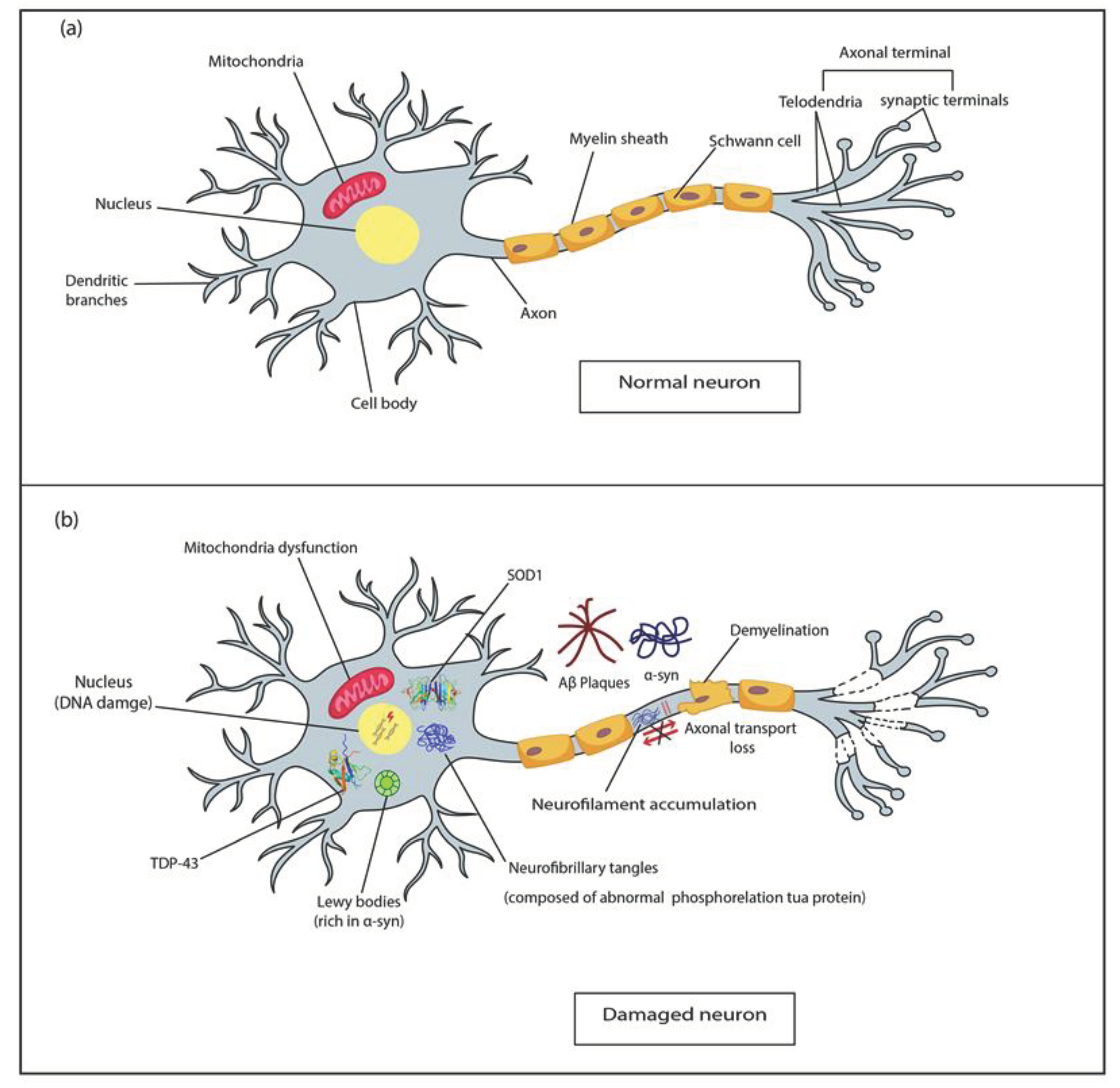

Table 1.
Some miRNA and their potential use in neurodegenerative diseases.
| Type of Disease | Type of miRNA | Expression level | indication | reference |
| MS | Mir-193a | decreased | Prognostic, diagnostic marker, and act as therapeutic | [38] |
| MS | Let7b-5p, mir-143-3p | Decreased in CSF | Promising miRNA candidate to discriminate PPMS | [39] |
| MS | Mir-155a, mir-146a | Increased in level (serum) | Act as diagnostic, prognostic and therapeutic | [40] |
| Mir-34a, mir-143a,mir-373a | Decreased in level (serum) | |||
| MS | Mir-10,mir-21,mir-124 | Decreased (blood) | in the pathogenesis of MS | [41] |
| MS | MIR-146a | Decreased in whole blood and feces of RRMS in compression with CIS, decreased in female. | In fecal as Diagnostic, prognostic biomarker | [42] |
| PD | Mir -7-1-5p, mir-223-3p | Increased in serum and serum isolated exosome | play a role in inflammation in PD, a Potential biomarker to discriminate PD from HC | [43] |
| PD | MIR-24, MIR-195 | Increased serum-EVs |
Act as an active biomarker in the diagnosis of PD |
[44] |
| MIR-19b | Decreased Serum -EVs | |||
| PD | MIR-22-3P | Decreased |
As a diagnostic parameter in the early stage of PD |
[45] |
| 22-3P, MIR-10b-5p,mir-151a-3p | increased(CSF) | |||
| PD | Has-mir-144-3p | Decreased inserum | Play a role in the progression of the disease and as an Early marker of PD. | [46] |
| PD | Mir-27-a, | Increased plasma | Can act in early diagnosis of PD, Use of mir-27-a, mir 27-b as a potential therapeutic target |
[47,48] |
| Mir-142-3p, mir-222,let-7a,let-7f | Decreased (plasma) | |||
| PD | Mir-132 | Increased( in males than in females) in prephrial blood | Biomarker for PD as diagnostic and disease progression | [49] |
| ALS | miR-16-5p | increased in plasma, CSF | Neuroprotective role in ALS after administration of intrathecal linage negative cell, | [50] |
| mir-206 | Increased plasma |
|||
| ALS | miR-23a | Increase in skeletal muscle in ALS patients and mouse model | Therapeutic inhibition of mir-23a may be a strategy to rescue peroxisome proliferator-activated receptor-γ coactivator (PGC-1α )activity and ameliorate skeletal muscle mitochondrial function in ALS, and down-regulation of miR-23a-3p has been proven to alleviate neuronal cell death and ROS | [51,52] |
| ALS | Hsa-mir-4649-5p |
Increased (plasma) |
Acts as diagnostic markers | [53] |
| Has-mir-4299 | Decreased (plasma) | |||
| AD | MIR-155,mir-146a, mir-125b ,mir-9,mir-34a | Upregulation Brain tissue of AD, ECF and CSF |
Biomarker for AD | [54,55] |
| AD | Mir-455-3p | Increased levels in blood serum, CSF post-mortem brain tissues, AD fibroblasts, AD β-lymphocytes, AD cell lines, transgenic AD (TgAD) mouse models and AD CSF | Biomarker and therapeutic | [56,57] |
| AD | Mir146a-5p | Upregulated in AD brain, neocortex and hippocampus |
Act as Pathogenesis and therapeutic biomarker | [58,59] |
| AD | Mir-125b | Upregulated in AD brain tissue | Pathological role can be used as treating therapy by using anti-mir 125b, anti-NF-kB | [60] |
Table 1 Abbreviations: AD, Alzheimer`s disease; ALS, amyotrophic lateral sclerosis; PD, Parkinson's disease; MS, multiple sclerosis, PPMS; primary progressive multiple sclerosis; CIS, clinically isolated syndrome; ROS; reactive oxygen species , NF-KB, nuclear factor-kappa B; EVs , extravascular vesicle; HC, healthy control; CSF, cerebrospinal fluid. .
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



