Submitted:
02 April 2024
Posted:
04 April 2024
You are already at the latest version
Abstract
Gliomas are the most common primary brain tumors in adults. Despite multidisciplinary treatment approaches, the survival rates for patients with malignant glioma have only improved marginally, and few prognostic biomarkers have been identified. Peroxisome proliferator-activated receptor γ (PPARγ) coactivator-1α (PGC-1α) is a crucial regulator of cancer metabolism, playing a vital role in cancer cell adaptation to fluctuating energy demands. In this study, the clinicopathological roles of PGC-1α in gliomas were evaluated. Employing immunohistochemistry, cell culture, siRNA transfection, cell viability assays, western blot analyses, and in vitro and in vivo invasion and migration assays, we explored the functions of PGC-1α in glioma progression. High PGC-1α expression was significantly associated with an advanced pathological stage in patients with glioma and with poorer overall survival. The downregulation of PGC-1α inhibited glioma cell proliferation, invasion, and migration and altered the expression of oncogenic markers. These results conclusively demonstrated that PGC-1α plays a critical role in maintaining the malignant phenotype of glioma cells and indicated that targeting PGC-1α could be an effective strategy to curb glioma progression and improve patient survival outcomes.
Keywords:
glioma
; PGC-1α
; prognosis
1. Introduction
Gliomas are the most common primary brain tumors in adults [1] and have a poor prognosis [2,3,4]. Although multidisciplinary treatment strategies have been applied, the survival of patients with malignant glioma has improved only slightly due to the high invasive ability and resistance to conventional treatments [5,6]. Few valuable molecules have been validated and widely applied as prognostic indicators in clinical practice, and a better understanidng of the biological and molecular factors involved in gliomas is necessary to for biomarker development [7].
Metabolic reprogramming is considered a hallmark of cancer [8]. Glycolysis, mitochondrial respiration, glutaminolysis, and fatty acid metabolism are important processes in cancer development [9,10,11,12,13]; they provide cancer cells with an adaptable metabolic feature and improve cancer cell survival under stress [14,15,16]. Mitochondrial biogenesis is critically regulated by the peroxisome proliferator-activated receptor γ (PPARγ) coactivator (PGC) family, which consists of PPARγ coactivator-1 alpha (PGC-1α) (PPARGC1A), PGC-1β (PPARGC1B), and PRC (PPRC1) [17]. Among numerous regulators or mediators of cancer metabolism, PGC1α is emerging as an essential controller of multiple metabolic pathways [18,19]. PGC1α functions as a necessary adaptor for cells to maintain a metabolic balance under harsh situations. It has protective effects against chronic diseases, such as skeletal muscle atrophy, heart failure, neurodegeneration, obesity, diabetes, and hepatic steatosis, some of which are predisposing factors for cancer [18,19,20]. There is recent evidence that PGC1α expression is closely associated with cancer progression. The exceptional ability of PGC1α to manipulate cellular metabolism enables cancer cells to thrive under a constantly fluctuating energy status and highlights the importance of PGC1α in effective cancer therapy [21]. PGC1α promotes carcinogenesis in chemical-induced colon and liver carcinoma mouse models [22]. The ectopic expression of PGC1α has been observed in several cancer types [23,24,25,26], and PGC1α is regulated by several oncogenes and signaling pathways [27,28,29]. Hence, the role of PGC1α in glioma as not been determined. This study aimed to validate the clinical and pathological roles of PGC1α in glioma.
2. Materials and Methods
2.1. Patients
Patients with glioma at the Neurosurgery Department of Chung Ho Hospital, Kaohsiung Medical University (KMUHIRB-E(I)-20190150) were included. Patients diagnosed based on biopsies alone or with incomplete medical records, no follow-up visits, low-quality pathological results, or poor immunohistochemical staining were excluded. A total of 68 patients were finally included in this study.
2.2. Immunohistochemistry Staining
For every case, 3 µm sections were cut from formalin-fixed, paraffin-embedded tissue blocks. These sections were deparaffinized, rehydrated, and autoclaved at 121°C for 10 min in Target Retrieval solution, pH 6.0 (DAKO, Glostrup, Denmark; S2369) to retrieve antigens. After 20 min at room temperature, 3% hydrogen peroxide was applied to the section for 5 min for blocking endogenous peroxidase at room temperature. After washing twice with Tris buffer, the sections were incubated with a 1:200 dilution of PGC1α for 1 h at room temperature. After washing twice with Tris buffer, the sections were incubated with the secondary antibody conjugated with horseradish peroxidase for 30 min at room temperature. Finally, the slides were incubated in 3,3-diaminobenzidine (Dako; K5007) for 5 min, followed by Mayer’s hematoxylin counterstaining for 90 s and mounting with Malinol. For the IHC scoring system, samples were classified into two types based on staining intensity: low expression and high expression. The proportion of positive tumor cells was scored as follows: 0 (no positive tumor cells), 1 (<10%), 2 (10–50%), and 3 (>50%). The intensity of staining was determined as: 0 (no staining), 1 (weak staining), 2 (moderate staining), and 3 (strong staining). The staining index (SI) was calculated as the product of the intensity and percentage of positive tumor cells, resulting in scores of 0, 1, 2, 3, 4, 6, and 9. A cut off value of 4 for the total score was established; scores of >4 indicated high PGC1α expression, and scores of <3 indicated low PGC1α expression.
2.3. Cell Culture
Cell lines utilized in this study were sourced from the American Type Culture Collection Cell Line Bank and maintained under conditions of 5% CO2 at a temperature of 37°C. The GBM8401, GBM8901, and DBTRG-05MG cell lines were propagated in RPMI medium enriched with 10% fetal bovine serum (FBS). U87-MG and SVGp12 cell lines were grown in minimum essential medium supplemented with 10% FBS. Additionally, the G5T/VGH, Hs683, and A172 cell lines were cultivated in Dulbecco’s modified Eagle’s medium (DMEM) with 10% FBS. The M059K cell line was cultured in a 1:1 mixture of DMEM and F12 medium with 10% FBS. The cell lines, including GBM8401, GBM8901, U87-MG, G5T/VGH, DBTRG-05MG, M059K, Hs683, and A172, were derived from tissues of patients with glioblastoma (GBM), while the SVG cell line, used as a normal control, originated from healthy brain tissue.
2.4. Transfection
PGC1α siRNA in glioma cells was achieved using DharmaFECT Transfection Reagent (Dharmacon, Lafayette, CO, USA). The sequence of human PGC1α siRNA (Sigma, St. Louis, MO, USA) included siRNA#1: GAGAAUUCAUGGAGCAAUA, siRNA#2: GAAGAGCGCCGUGUGAUUU, siRNA#3: ACACUCAGCUAAGUUAUAA, and siRNA#4: GCAGGUAACAUGUUCCCUA. Fo transfection, 5 μM PGC1α siRNA was used. Following transfection with siRNA, cells were cultured for 3 days before use. PGC1α protein expression was detected by western blot analyses.
2.5. Cell Viability
Cells were added to DMEM containing 10% serum and placed in a 24-well plate with approximately 1 × 104 cells per 0.5 ml in each well. These cells were incubated under 5% CO2, saturated humidity, and 37 °C for 24 h. Cells were counted using an MTT assay after co-culture with 5 μM PGC1α siRNA for 24, 48, and 72 h.
2.6. Western Blotting
All samples were lysed in 200 μl lysis buffer. Then, 50 μg of protein for every sample was loaded for separation by SDS-PAGE at 50 V for 4 h. The protein was transferred from the gel to a PVDF membrane. After 1 h in blocking buffer, the membranes were incubated with primary antibodies [PGC1α (1:500; Biorbyt, Ely, UK; orb13647), β-actin (1:20000; Sigma; A5441), N-cadherin (1:500; Proteintech, Rosemont, IL, USA; 22018-1-AP), E-cadherin (1:500; Proteintech, Rosemont, IL, USA; 20874-1-AP), cyclin D1 (1:500; Proteintech; 60186-1-lg), VEGF (1:500; ABGENT, San Diego, CA, USA; P15692)] for 2 h at room temperature and secondary antibodies [Goat anti-Rabbit (1:2000; Millipore, Billerica, MA, USA; AP132P) and Goat anti-Mouse (1:2000; Millipore; AP124P)] for 90 min. ECL solution (Western Lightning; 205-14621) was used to detect specific bands with MINICHEMI (Thermo).
2.7. Invasion Assay In Vitro
Cell invasion was evaluated using a Transwell assay (CORNING Inc., Corning, NY, USA; COR3452) in vitro. Cells were seeded at 1 × 104 per insert and the lower chamber of the Transwell was filled with 2 ml of medium with nonsense siRNA (si-non) or PGC1α siRNA. After incubation for 24 h, cells remaining on the upper surface of the Transwell membrane were removed using a cotton swab. Cells that had passed through the Transwell to the bottom of the insert were fixed, stained, photographed, and quantified by counting in six random high-powered fields.
2.8. Migration Assay In Vitro
Cell invasion was evaluated using a wound healing assay (ibidi; 80209). Briefly, 24-well plates were coated in Matrigel and cells were cultured at 37°C for 12 h. Then, 70 μl of cells at 1 × 106/ml was incubated with siRNA for 24 h. Images were obtained at 0, 8, 12, and 24 h.
2.9. Animal Model
GBM8401 cells, engineered to express fluorescence (1 × 105 cells in 5 μl), were intracranially implanted into the striatum of immunodeficient mice (LASCO Laboratory Animal Center, Taipei, Taiwan). The animal protocol received approval from the Kaohsiung Medical University Committee of Institutional Animal Research (IACUC 108285). The mice were kept under constant environmental conditions with a temperature maintained at 24°C and subjected to regular light/dark cycles (12 h each), with unrestricted access to a standard diet. For this experiment, the control group received injections of unmodified GBM8401 cells (n = 12), and the experimental group was treated with GBM8401 cells in which PGC-1α had been knocked down using shRNA targeting PGC-1α (n = 12). The mice were anesthetized using isoflurane, and fluorescence detection was conducted using the Xenogen IVIS® Spectrum Noninvasive Quantitative Molecular Imaging System (J&H; IVIS Lumina LT 2D) at intervals of 7, 14, and 21 days following the cell injections.
2.10. Data Analysis
SPSS 26.0 (IBM, Armonk, NY, USA) was used for statistical analyses. Chi-square tests were performed to evaluate correlations between PGC1α protein expression and clinicopathological parameters. The survival rate was analyzed by the Kaplan–Meier method with the log-rank test. Multivariate Cox regression analyses were used to verify the independent effect of each variable. The western blotting results were analyzed using Lane 1D. A P-value of <0.05 was considered statistically significant.
3. Results
3.1. Associations of PGC-α Expression with Clinicopathological Characteristics in Patients with Intracranial Glioma
Ninety-five patients from Kaohsiung Medical University Hospital diagnosed with intracranial gliomas were included. Clinicopathological characteristics, such as age, sex, World Health Organization (WHO) pathological classification, tumor size, receipt of chemotherapy and radiotherapy, and Karnofsky performance score (KPS) (a quality-of-life index), were evaluated in relation to PGC-1α expression levels. Among patients under 60 years of age, 30.5% showed low PGC-1α expression and 44.2% exhibited high expression. Among patients over 60 years of age, 7.4% had low PGC-1α expression and 17.9% had high expression. The age-related difference in PGC-1α expression was not statistically significant (P = 0.342). Regarding sex, 21.1% of male patients had low PGC-1α expression and 33.7% had high expression. For female patients, 16.8% had low and 28.4% had high PGC-1α expression, with no significant difference in expression based on sex (P = 1.000). The WHO grade showed a significant correlation with PGC-1α expression. In particular, 15.8% of patients with grade II glioma had low expression and 8.4% had high expression, while in grades III/IV gliomas, 22.1% had low expression and 53.7% had high expression (P = 0.003). No significant differences were found in PGC-1α expression with respect to tumor size, radiotherapy, chemotherapy, or KPS score (Table 1). In summary, there was a significant association between PGC-1α expression and pathological staging in patients with glioma, where higher PGC-1α expression was correlated with higher tumor grades.
3.2. Prognostic Significance of PGC-1α Expression in Glioma
As shown in Figure 1, a Kaplan–Meier analysis and subsequent log-rank test confirmed that PGC-1α expression is correlated with survival in astrocytoma; in particular, a high level of PGC-1α expression was significantly correlated with poor overall survival. In univariate analyses, the impact of each factor on the prognosis of patients with glioma was assessed. Patient outcomes were not significantly correated with age (relative risk = 0.648, 95% confidence interval (CI), 0.311–1.351, P = 0.247), sex (relative risk = 0.892, 95% CI, 0.473–1.682, P = 0.724), tumor size (relative risk = 1.114, 95% CI, 0.572–2.171, P = 0.751), chemotherapy (relative risk = 1.542, 95% CI, 0.776 to 3.065, P = 0.217), or KPS (relative risk = 1.167, 95% CI, 0.586 to 2.324, P = 0.660). The WHO grade was identified as a significant prognostic factor (relative risk = 0.382, 95% CI, 0.171–0.851, P = 0.019) (Table 2). Additionally, radiotherapy had a relative risk of 1.845 with a 95% CI of 0.965–3.526 (P = 0.064) (Table 2), indicating a trend towards significance, suggesting that it has an impact on outcomes.
PGC-1α expression was closely related to prognosis (relative risk = 0.177, 95% CI, 0.075–0.417, P < 0.001), highlighting its potential as a critical prognostic factor in glioma. In a multivariate analysis, PGC-1α expression remained significant (relative risk = 0.197, 95% CI, 0.080–0.482, P < 0.001). However, the WHO grade, while showing a reduced relative risk of 0.546 and a 95% CI of 0.223–1.335, was not a significant factor in this analysis (P = 0.185) (Table 2). Therefore, PGC-1α was identified as an independent biomarker for prognosis in glioma.
3.3. Elevated PGC-1α Protein Expression in GBM Cells Relative to Normal Cells
To elucidate the difference in PGC-1α expression between normal glial cells and GBM, real-time PCR was conducted (Figure 2). PGC-1α mRNA levels were evaluated across various glioma cell lines, including GBM8401, U87-MG, G5T/VGH, DBTRG-05MG, M059K, and A172, and compared with levels in normal primary human astrocytes (SVGp12). PGC-1α expression levels in the glioma cell lines GBM8401, U87-MG, G5T/VGH, DBTRG-05MG, M059K, and A172 were significantly higher than those in SVGp12, suggesting that PGC-1α in integral in the pathogenesis of glioma. In comparison with levels in SVGp12, the average increases in PGC-1α expression in various glioma cell lines were as follows: 5.4-fold in GBM8401 cells, 1.7- to 2-fold in U87-MG cells at different time points, and approximately 4.6-fold in A172 cells. G5T/VGH cells displayed a moderate 1.5-fold elevation, DBTRG-05MG cells exhibited an approximately 2.5-fold increase, and M059K cells had an average increase of about 3-fold over levels in SVGp12. Given the high PGC-1α protein levels in GBM8401 and A172 cells, they were chosen for further analyses of the impact of PGC-1α downregulation on glioma progression. In particular, real-time PCR and western blot analyses were utilized to quantify the impact of PGC-1α siRNA silencing (Figure 3). In GBM8401 cells, there was a notable reduction in PGC-1α mRNA expression to an average of approximately 0.23 (relative to levels in wild-type cells) after siRNA treatment. Western blotting further demonstrated a significant decrease in PGC-1α protein levels (i.e., 0.35) following siRNA application, indicating effective knockdown. For A172 cells, real-time PCR revealed a decrease in PGC-1α mRNA levels to an average of 0.38 with siRNA treatment. Western blotting results echoed this finding, with PGC-1α protein levels diminishing to an average of 0.20 in comparison with levels in the control. These findings underscore the efficiency of siRNA targeting PGC-1α in downregulating its expression at both the mRNA and protein levels.
3.4. PGC-1α Knockdown Inhibited GBM Cell Proliferation
To evaluate the effect of PGC-1α on GBM cell survival, we compared cell viability in GBM8401 and A172 cell lines with or without the siRNA-mediated knockdown of PGC-1α. Viability was measured at 24, 48, and 72 h after transfection. As shown in Figure 4, in GBM8401 cells, there was a discernible decrease in cell viability to an average of 58% at 48 h post-transfection with si-PGC-1α. This reduction became more pronounced at 72 h, with an average viability of about 27%. In the A172 cell line, viability also decreased after si-PGC-1α transfection, averaging 42% at 48 h and approximately 27% at 72 h. These results highlight the significant impact of PGC-1α knockdown on the viability of GBM cell lines, affirming the potential value of PGC-1α as a target for therapeutic strategies. These data indicate that that suppression of PGC-1α can markedly hinder the survival of GBM cells.
3.5. PGC-1α Knockdown Inhibited GBM Cell Invasion and Migration
To analyze cell migration, a wound healing assay was employed with GBM8401 and A172 cells to observe the effects of PGC-1α suppression (Figure 5). PGC-1α was silenced using siRNA, and the migration percentage was recorded at 0, 8, 12, and 24 h post-transfection. For GBM8401 cells, the average migration percentages in the control group were approximately 1.9% at 0 h, escalating to about 32% at 8 h, 44% at 12 h, and approximately 78% at 24 h. In the si-PGC-1α group, the cells exhibited a significant impairment in migration, with average percentages of approximately 1.8% at 0 h, 16% at 8 h, 28% at 12 h, and 46% at 24 h post-transfection. Similarly, A172 cells demonstrated an average initial migration percentage of approximately 1.4%, with increases to approximately 19% at 8 h, 33% at 12 h, and 49% at 24 h in the control group. Post-transfection with si-PGC-1α, these cells showed a reduction in migration to averages of approximately 1.4% at 0 h, 7% at 8 h, 17% at 12 h, and 29% at 24 h.
Cell invasion capabilities for GBM8401 and A172 cell lines were evaluated to determine the effect of PGC-1α silencing on GBM invasiveness using a Matrigel invasion assay (Figure 6). Following transfection with siRNAs targeting PGC-1α, both cell lines exhibited a significant decrease in invasion rates compared with those in the control and negative control groups. In the GBM8401 cell line, the average percentage of invasion for the control group was approximately 71%, while si-PGC-1α transfection led to a significant reduction, with an average invasion rate of approximately 35%. For the A172 cell line, the average invasion rate in the control group was approximately 51%, and si-PGC-1α treatment resulted in a marked decrease to an average of approximately 11%. These results suggest that PGC-1α contrnibutes to the invasive capacity of GBM cells, with its knockdown by siRNA significantly inhibiting invasion. This indicates that PGC-1α could be a promising target for therapeutic strategies aimed at mitigating GBM invasiveness.
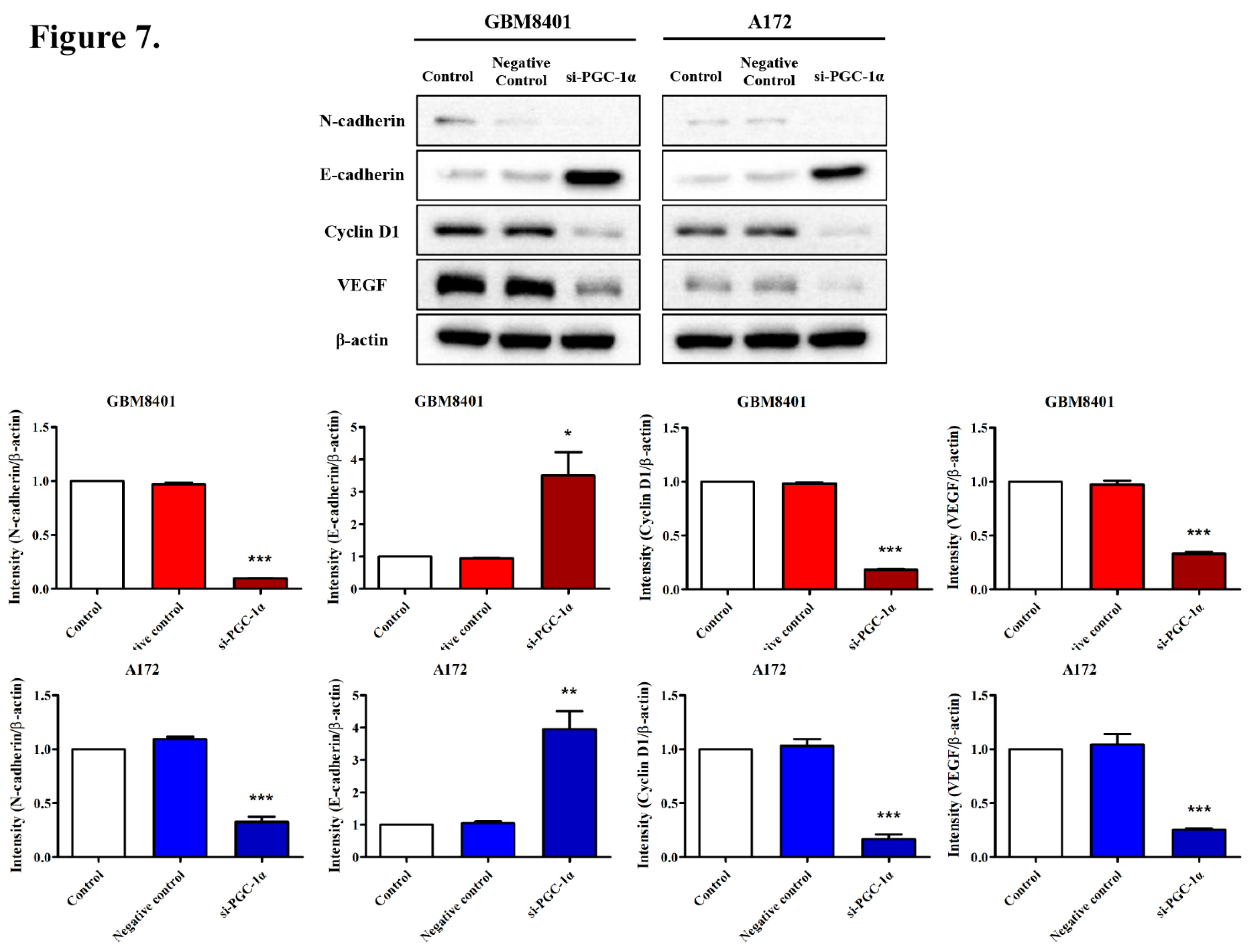
3.6. Modulation of Carcinogenic Marker Expression by PGC-1α Knockdown in Glioma Cells
The effects of PGC-1α on key molecular markers associated with glioma progression were assessed in GBM8401 and A172 cell lines using western blot analyses. PGC-1α silencing through siRNA led to observable changes in the expression levels of several critical proteins. In GBM8401 cells, siRNA-mediated knockdown of PGC-1α resulted in marked reductions in the expression of N-cadherin (0.10-fold), Cyclin D1 (0.20-fold), and VEGF (0.33-fold), while E-cadherin expression showed an increase (2.85-fold) over levels in the control and negative control groups. For A172 cells, following PGC-1α knockdown, similar trends were observed. There were significant decreases in the expression of N-cadherin, Cyclin D1, and VEGF, with verage fold-changes of approximately 0.33, 0.17, and 0.26, respectively. E-cadherin expression increased with an average fold change of approximately 4.28. These patterns suggest that PGC-1α has a substantial role in the regulation of molecules that are pivotal for glioma cell invasiveness and proliferation, providing insight into how PGC-1α suppression could alter the behavior of glioma cells and serve as a therapeutic target.
Figure 7.
Protein Expression Analysis of N-cadherin, E-cadherin, Cyclin D1, and VEGF Following PGC-1α Silencing in GBM Cells. Western blot analysis of N-cadherin, E-cadherin, Cyclin D1, and VEGF following PGC-1α siRNA transfection in GBM cells. Protein expression levels were assessed in GBM8401 and A172 cells for the control, negative control, and si-PGC-1α-transfected groups. The bar graphs below the blots quantify the relative expression normalized to β-actin. **P < 0.01 and ***P < 0.001 compared with the control group.
Figure 7.
Protein Expression Analysis of N-cadherin, E-cadherin, Cyclin D1, and VEGF Following PGC-1α Silencing in GBM Cells. Western blot analysis of N-cadherin, E-cadherin, Cyclin D1, and VEGF following PGC-1α siRNA transfection in GBM cells. Protein expression levels were assessed in GBM8401 and A172 cells for the control, negative control, and si-PGC-1α-transfected groups. The bar graphs below the blots quantify the relative expression normalized to β-actin. **P < 0.01 and ***P < 0.001 compared with the control group.
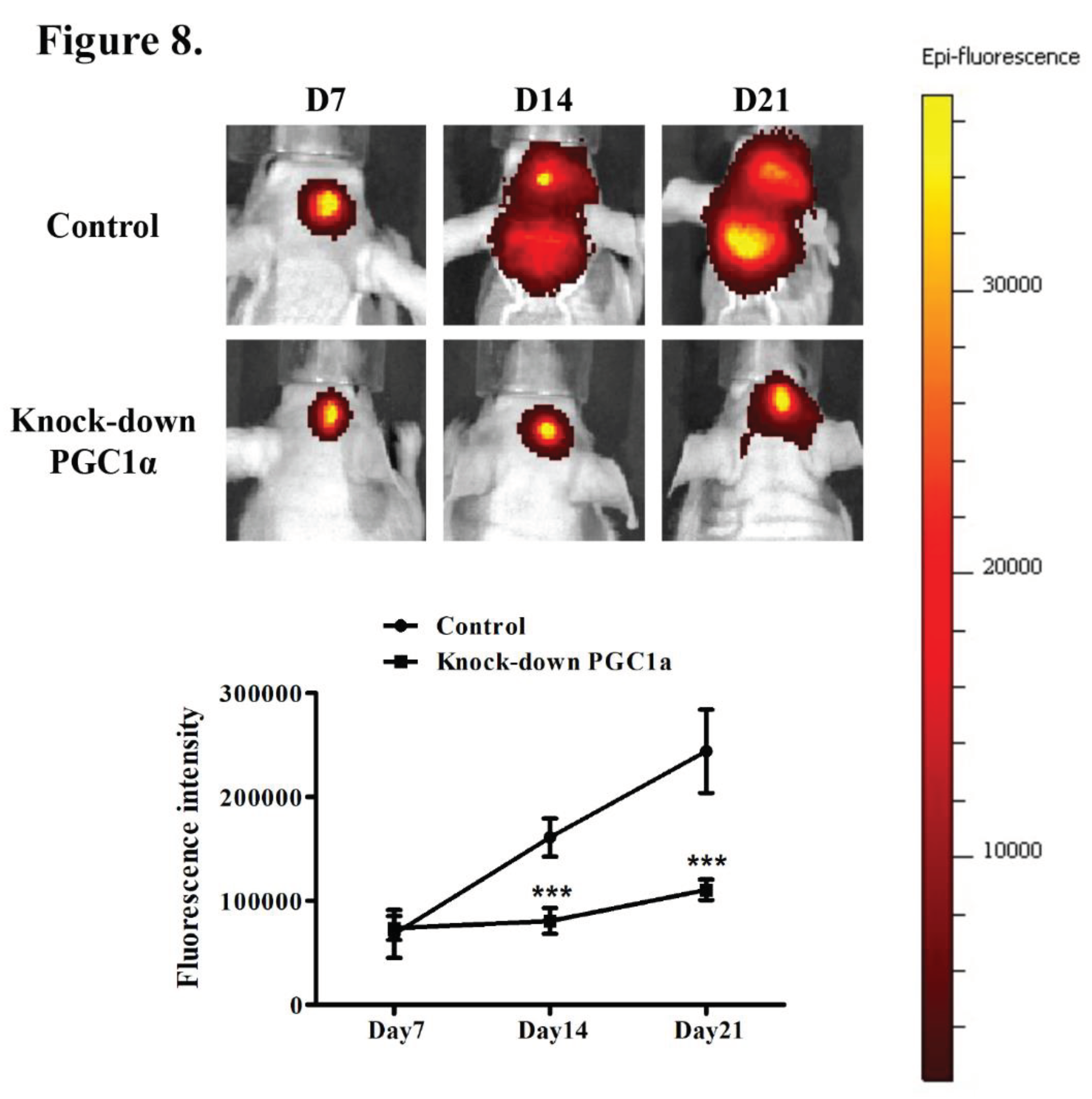
3.7. Effects of PGC-1α Knockdown on Tumor Growth and Survival in a Murine Glioblastoma Model
The effect of PGC-1α knockdown on tumor dynamics was monitored in vivo using a fluorescence-based tumor growth assay with a murine model. Mice were implanted with GBM cells expressing normal levels of PGC-1α (control group) or with PGC-1α knockdown. Fluorescence intensity measurements at days 7, 14, and 21 post-implantation revealed that tumor growth was markedly lower in the PGC-1α knockdown group than in the control group. This significant reduction in fluorescence intensity in the PGC-1α knockdown group was correlated with decreases in tumor activity and progression, implying that PGC-1α is integral to tumor growth within this biological system. These data further suggest that PGC-1α is a therapeutic target for inhibiting tumor growth and extending survival in GBM.
Figure 8.
In Vivo Tumor Growth Following PGC-1alpha Knockdown. Effects of PGC-1α knockdown on tumor progression in GBM cells in vivo. After intracranial implantation of GBM8401 cells with fluorescence in the striatum, (A) the comparative epi-fluorescence intensity was evaluated. (B) progression over time post-implantation in the PGC-1α knockdown and control groups. ***P < 0.001 compared with the control group on Day 14 and Day 21.
Figure 8.
In Vivo Tumor Growth Following PGC-1alpha Knockdown. Effects of PGC-1α knockdown on tumor progression in GBM cells in vivo. After intracranial implantation of GBM8401 cells with fluorescence in the striatum, (A) the comparative epi-fluorescence intensity was evaluated. (B) progression over time post-implantation in the PGC-1α knockdown and control groups. ***P < 0.001 compared with the control group on Day 14 and Day 21.
4. Discussion
PGC-1α, a transcriptional coactivator that orchestrates mitochondrial biogenesis and metabolic flux, has emerged as a pivotal factor in the metabolic reprogramming of cancer cells—a key adaptation that supports their rapid growth and survival under diverse conditions [17,30]. In several types of carcinomas, including breast, colon, and ovarian cancers, PGC-1α is frequently downregulated, and its downregulation is associated with a loss of metabolic flexibility and an increased reliance on glycolysis for energy production, even in the presence of oxygen, a phenomenon known as the Warburg effect [31,32,33]. The decreased expression in these contexts suggests that of PGC-1α is involved in maintaining metabolic checks that prevent cancer cell proliferation, thereby underscoring the importance of its tumor-suppressive functions [34]. Conversely, the upregulation of PGC-1α in certain cancers illustrates the adaptability of cancer cells in exploiting physiological mechanisms for their survival and expansion, contributing to enhanced mitochondrial biogenesis and metabolic efficiency, which provide cancer cells with the energy and biosynthetic precursors necessary for rapid growth [35,36]. Cancer cells undergo a fundamental shift in their metabolism to support increased demands for energy and biosynthetic materials, with PGC-1α standing at the crossroads of this metabolic reprogramming, regulating key pathways involved in energy production and the synthesis of macromolecules [37]. The roles of PGC-1α in mitochondrial biogenesis and function are a double-edged sword in the context of cancer [38]. On one hand, the ability of PGC-1α to enhance mitochondrial efficiency can support the energetic and biosynthetic needs of rapidly proliferating cancer cells [39]. On the other hand, the role of PGC-1α in oxidative metabolism can lead to the increased production of reactive oxygen species (ROS), which can damage cancer cells and potentially limit their growth [30,40]. The Warburg effect, characterized by increased glycolysis in the presence of oxygen, is a hallmark of many cancers [41]. While PGC-1α is traditionally associated with oxidative metabolism, its influence on cancer metabolism extends to glycolytic pathways as well [33,42]. Through its regulatory network, PGC-1α can impact the expression of enzymes involved in glycolysis, thereby affecting the metabolic strategy of cancer cells [17]. In our study, PGC-1α expression was significantly correlated with the pathological stage of gliomas, with higher expression in advanced tumors. While age, gender, tumor size, and treatments (e.g., chemotherapy and radiotherapy) did not significantly affect PGC-1α levels, the WHO grade was a notable exception, with support for its prognostic value. Kaplan–Meier and log-rank analyses further established PGC-1α as a crucial prognostic marker, with high expression levels associated with poorer overall survival in patients with glioma, suggesting that it is an independent biomarker for glioma prognosis.
PGC-1α is also closely related to oncogenic processes within astrocytic cells [43]. The elevation of PGC-1α and mitochondrial transcription factor A (TFAM) expression in astrocytoma tissues is a potential adaptive mechanism where cancer cells exploit mitochondrial biogenesis and enhanced energy production to support their rapid growth and survival in the hostile tumor microenvironment [44]. This scenario is further complicated by the positive correlation between PGC-1α levels and the activity of AMP-activated protein kinase (AMPK) and its phosphorylated form, suggesting a sophisticated regulatory network that promotes the invasive growth of astrocytomas [45]. In our result, knocking down PGC-1α in GBM cells led to significant reductions in their proliferation, migration, and invasion as well as altered expression levels of key carcinogenic markers, pointing to the protein's critical role in tumor growth and aggressiveness. In vivo experiments further demonstrated that PGC-1α knockdown drastically curbs tumor growth in a murine model, further indicating that PGC-1α is a viable target for GBM therapy. These findings collectively emphasize the importance of PGC-1α in maintaining the malignant phenotype of GBM cells and suggest that targeting PGC-1α could be an effective approach to inhibit GBM progression and improve patient outcomes.
5. Conclusions
PGC-1α plays a crucial role in GBM progression, influencing cell proliferation, invasion, migration, and the expression of oncogenic markers. The knockdown of PGC-1α in GBM cell lines, such as GBM8401 and A172, resulted in marked reductions in cell viability and migratory and invasive capabilities as well as altered expression levels of key proteins associated with cancer progression. These in vitro findings were substantiated by in vivo evidence showing reduced tumor growth in a murine model with PGC-1α knockdown, highlighting the significance of the protein in tumor development and maintenance. Collectively, the results support the important role of PGC-1α in GBM and suggest that targeting PGC-1α could serve as an effective therapeutic strategy to impede GBM progression and improve outcomes. This research underscores the potential of PGC-1α not only as a prognostic biomarker but also as a promising target for the development of novel GBM treatments.
Author Contributions
Conceptualization, J.-H. L.; methodology, T.-T.T.; software, C.-H.C.; validation, Y.-W.C. and C.-J.L.; formal analysis, J.-H. L.; investigation, Y.-W.C.; resources, C.-Y. C.; data curation, A.-S. L.; writing—original draft preparation, Y.-W.C.; writing—review and editing, A.-L.K. and A.-S. L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from Kaohsiung Medical University Chung-Ho Memorial Hospital (KMUH110-0M28 and KMUH111-1M21).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board of Kaohsiung Medical University Hospital (KMUHIRB-20190150) for studies involving humans. The study was approved by the Committee of Institutional Animal Research of Kaohsiung Medical University (IACUC 107078).
Informed Consent Statement
Patient consent was waived due to the fact that all data from patients were from a biobank and delinked.
Data Availability Statement
No new data were created or analysed in this study. Data sharing is not applicable to this article.
Acknowledgments
The authors thank the Center for Laboratory Animals in Kaohsiung Medical University for the animal care and for IVIS Lumina imaging.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Davis, F.G.; McCarthy, B.J. Current epidemiological trends and surveillance issues in brain tumors. Expert Rev. Anticancer Ther. 2001, 1, 395–401. [Google Scholar] [CrossRef]
- Ohgaki, H.; Kleihues, P. Epidemiology and etiology of gliomas. Acta Neuropathol. 2005, 109, 93–108. [Google Scholar] [CrossRef]
- Fuller GN, Scheithauer BW. The 2007 Revised World Health Organization (WHO) Classification of Tumours of the Central Nervous System: newly codified entities. Brain Pathol. 2007, 17, 304–307. [Google Scholar] [CrossRef] [PubMed]
- Wen, P.Y.; Kesari, S. Malignant gliomas in adults. N. Engl. J. Med. 2008, 359, 492–507. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Ducray, F.; Idbaih, A.; Wang, X.W.; Cheneau, C.; Labussiere, M.; Sanson, M. Predictive and prognostic factors for gliomas. Expert Rev. Anticancer Ther. 2011, 11, 781–789. [Google Scholar] [CrossRef]
- Tsai, H.P.; Tsai, T.H.; Hsieh, Y.J.; Chen, Y.T.; Lee, C.L.; Tsai, Y.C.; et al. Overexpression of Fli-1 in astrocytoma is associated with poor prognosis. Oncotarget. 2017, 8, 29174–29186. [Google Scholar] [CrossRef]
- Boroughs, L.K.; DeBerardinis, R.J. Metabolic pathways promoting cancer cell survival and growth. Nat. Cell Biol. 2015, 17, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Ward, P.S.; Thompson, C.B. Metabolic reprogramming: a cancer hallmark even warburg did not anticipate. Cancer Cell. 2012, 21, 297–308. [Google Scholar] [CrossRef]
- Currie, E.; Schulze, A.; Zechner, R.; Walther, T.C.; Farese, R.V., Jr. Cellular fatty acid metabolism and cancer. Cell Metab. 2013, 18, 153–161. [Google Scholar] [CrossRef]
- Hensley, C.T.; Wasti, A.T.; DeBerardinis, R.J. Glutamine and cancer: cell biology, physiology, and clinical opportunities. J. Clin. Invest. 2013, 123, 3678–3684. [Google Scholar] [CrossRef] [PubMed]
- Locasale, J.W. Serine, glycine and one-carbon units: cancer metabolism in full circle. Nat. Rev. Cancer. 2013, 13, 572–583. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, S.E.; Chandel, N.S. Targeting mitochondria metabolism for cancer therapy. Nat. Chem. Biol. 2015, 11, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Bobrovnikova-Marjon, E.; Hurov, J.B. Targeting metabolic changes in cancer: novel therapeutic approaches. Annu. Rev. Med. 2014, 65, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Seyfried, T.N.; Flores, R.E.; Poff, A.M.; D'Agostino, D.P. Cancer as a metabolic disease: implications for novel therapeutics. Carcinogenesis. 2014, 35, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Staubert, C.; Bhuiyan, H.; Lindahl, A.; Broom, O.J.; Zhu, Y.; Islam, S.; et al. Rewired metabolism in drug-resistant leukemia cells: a metabolic switch hallmarked by reduced dependence on exogenous glutamine. J. Biol. Chem. 2015, 290, 8348–8359. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Widlund, H.R.; Puigserver, P. PGC-1 coactivators: Shepherding the mitochondrial biogenesis of tumors. Trends Cancer. 2016, 2, 619–631. [Google Scholar] [CrossRef]
- Girnun, G.D. The diverse role of the PPARgamma coactivator 1 family of transcriptional coactivators in cancer. Semin. Cell Dev. Biol. 2012, 23, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.W.; Yao, Z.; Vicencio, J.M.; Karkucinska-Wieckowska, A.; Szabadkai, G. PGC-1 family coactivators and cell fate: roles in cancer, neurodegeneration, cardiovascular disease and retrograde mitochondria-nucleus signalling. Mitochondrion. 2012, 12, 86–99. [Google Scholar] [CrossRef]
- Austin, S.; St-Pierre, J. PGC1alpha and mitochondrial metabolism--emerging concepts and relevance in ageing and neurodegenerative disorders. J. Cell. Sci. 2012, 125, 4963–4971. [Google Scholar] [CrossRef]
- Wolf, D.A. Is reliance on mitochondrial respiration a "chink in the armor" of therapy-resistant cancer? Cancer Cell. 2014, 26, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Bhalla, K.; Hwang, B.J.; Dewi, R.E.; Ou, L.; Twaddel, W.; Fang, H.B; et al. PGC1alpha promotes tumor growth by inducing gene expression programs supporting lipogenesis. Cancer Res. 2011, 71, 6888–6898. [Google Scholar] [CrossRef] [PubMed]
- Shiota, M.; Yokomizo, A.; Tada, Y.; Inokuchi, J.; Tatsugami, K.; Kuroiwa, K.; et al. Peroxisome proliferator-activated receptor gamma coactivator-1alpha interacts with the androgen receptor (AR) and promotes prostate cancer cell growth by activating the AR. Mol Endocrinol. 2010, 24, 114–127. [Google Scholar] [CrossRef]
- McGuirk, S.; Gravel, S.P.; Deblois, G.; Papadopoli, D.J.; Faubert, B.; Wegner, A.; et al. PGC-1alpha supports glutamine metabolism in breast cancer. Cancer Metab. 2013, 1, 22. [Google Scholar] [CrossRef]
- Vazquez, F.; Lim, J.H.; Chim, H.; Bhalla, K.; Girnun, G.; Pierce, K.; et al. PGC1alpha expression defines a subset of human melanoma tumors with increased mitochondrial capacity and resistance to oxidative stress. Cancer Cell. 2013, 23, 287–301. [Google Scholar] [CrossRef]
- Taguchi, A.; Delgado, O.; Celiktas, M.; Katayama, H.; Wang, H.; Gazdar, A.F; et al. Proteomic signatures associated with p53 mutational status in lung adenocarcinoma. Proteomics. 2014, 14, 2750–2759. [Google Scholar] [CrossRef]
- Haq, R.; Shoag, J.; Andreu-Perez, P.; Yokoyama, S.; Edelman, H.; Rowe, G.C.; et al. Oncogenic BRAF regulates oxidative metabolism via PGC1alpha and MITF. Cancer Cell. 2013, 23, 302–315. [Google Scholar] [CrossRef]
- LaGory, E.L.; Wu, C.; Taniguchi, C.M.; Ding, C.C.; Chi, J.T.; von Eyben, R.; et al. Suppression of PGC-1alpha is critical for reprogramming oxidative metabolism in renal cell carcinoma. Cell Rep. 2015, 12, 116–127. [Google Scholar] [CrossRef]
- Sancho, P.; Burgos-Ramos, E.; Tavera, A.; Bou Kheir, T.; Jagust, P.; Schoenhals, M.; et al. MYC/PGC-1alpha balance determines the metabolic phenotype and plasticity of pancreatic cancer stem cells. Cell Metab. 2015, 22, 590–605. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.; Luo, X.; Xiao, L.; Tang, M.; Bode, A.M.; Dong, Z.; et al. The role of PGC1alpha in cancer metabolism and its therapeutic implications. Mol. Cancer Ther. 2016, 15, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Han, C.Y.; Patten, DA.; Richardson, R.B.; Harper, M.E.; Tsang, B.K. Tumor metabolism regulating chemosensitivity in ovarian cancer. Genes Cancer. 2018, 9, 155–175. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, N.; Das, G.M. Metabolic reprogramming in breast cancer and its therapeutic implications. Cells. 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Cassim, S.; Vucetic, M.; Zdralevic, M.; Pouyssegur, J. Warburg and beyond: the power of mitochondrial metabolism to collaborate or replace fermentative glycolysis in cancer. Cancers. 2020, 12, 1119. [Google Scholar] [CrossRef]
- Villena, J.A. New insights into PGC-1 coactivators: redefining their role in the regulation of mitochondrial function and beyond. Febs J. 2015, 282, 647–672. [Google Scholar] [CrossRef] [PubMed]
- LeBleu, V.S.; O'Connell, J.T.; Herrera, K.N.G.; Wikman, H.; Pantel, K.; Haigis, M.C.; et al. PGC-1α mediates mitochondrial biogenesis and oxidative phosphorylation in cancer cells to promote metastasis. Nat. Cell. Biol. 2014, 16, 1125–1125. [Google Scholar] [CrossRef]
- Mastropasqua, F.; Girolimetti, G.; Shoshan, M. PGC1α: friend or foe in cancer? Genes-Basel, 2018; 9. [Google Scholar]
- Hipolito, A.; Martins, F.; Mendes, C.; Lopes-Coelho, F.; Serpa, J. Molecular and metabolic reprogramming: pulling the strings toward tumor metastasis. Front. Oncol. 2021, 11, 656851. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Chen, S.; Hu, X. Peroxisome proliferator-activated receptor gamma coactivator-1 alpha: a double-edged sword in prostate cancer. Curr. Cancer Drug Targets. 2022, 22, 541–559. [Google Scholar] [CrossRef]
- LeBleu, V.S.; O'Connell, J.T.; Gonzalez Herrera, K.N.; Wikman, H.; Pantel, K.; Haigis, M.C.; et al. PGC-1alpha mediates mitochondrial biogenesis and oxidative phosphorylation in cancer cells to promote metastasis. Nat. Cell Biol. 2014, 16, 992–1003. [Google Scholar] [CrossRef]
- Rius-Perez, S.; Torres-Cuevas, I.; Millan, I.; Ortega, A.L.; Perez, S. PGC-1alpha, inflammation, and oxidative stress: an integrative view in metabolism. Oxid. Med. Cell Longev. 2020, 2020, 1452696. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, L.; Supuran, C.T.; Alfarouk, K.O. The Warburg effect and the hallmarks of cancer. Anticancer Agents Med. Chem. 2017, 17, 164–170. [Google Scholar] [CrossRef]
- Liang, H.; Ward, W.F. PGC-1alpha: a key regulator of energy metabolism. Adv. Physiol. Educ. 2006, 30, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.N.; Zhang, M.Q.; Yu, F.L.; Han, B.; Bao, M.Y.; Li, X.; et al. Peroxisom proliferator-activated receptor-γ coactivator-1α in neurodegenerative disorders: A promising therapeutic target. Biochem. Pharmacol. 2023, 215. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lou, L.; Li, D.; Wang, Y.; Jia, X.; Hao, X.; et al. Expression, clinicopathological significance, and prognostic potential of AMPK, p-AMPK, PGC-1α, and TFAM in astrocytomas. J. Neuropath. Exp. Neur. 2023, 83, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Bruns, I.; Sauer, B.; Burger, M.C.; Eriksson, J.; Hofmann, U.; Braun, Y.; et al. Disruption of peroxisome proliferator-activated receptor coactivator (PGC)-1 reverts key features of the neoplastic phenotype of glioma cells. J. Biol. Chem. 2019, 294, 3037–3050. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Correlation Between PGC-1α Expression Levels and Patient Survival in GBM. (A) Immunohistochemical staining for PGC-1α expression in glioma. Representative immunohistochemical staining results for gliomas with high (left) and low (right) PGC-1α expression. (B) Kaplan–Meier survival analysis correlating PGC-1α expression levels with patient survival outcomes, assessed by the log-rank test. There was a significant difference in prognosis between the high and low expression groups.
Figure 1.
Correlation Between PGC-1α Expression Levels and Patient Survival in GBM. (A) Immunohistochemical staining for PGC-1α expression in glioma. Representative immunohistochemical staining results for gliomas with high (left) and low (right) PGC-1α expression. (B) Kaplan–Meier survival analysis correlating PGC-1α expression levels with patient survival outcomes, assessed by the log-rank test. There was a significant difference in prognosis between the high and low expression groups.
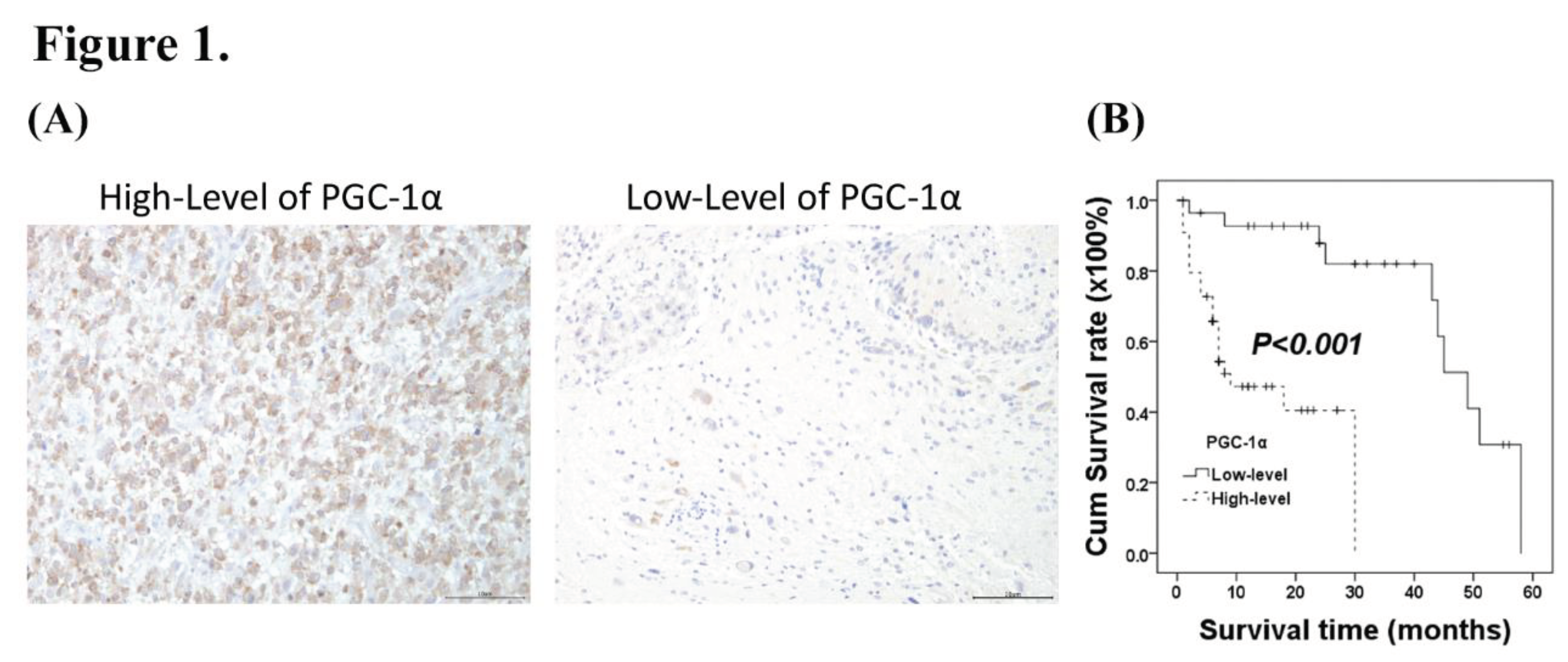
Figure 2.
Relative PGC-1α mRNA Expression Between Glial and GBM Cell Lines. Comparison of PGC-1α mRNA expression between normal glial and GBM cell lines. Bar graph represents the relative expression levels of PGC-1α mRNA in normal glial cells (SVGp12) and various GBM cell lines (GBM8401, U87-MG, A172, G5T/VGH, DBTRG-05MG, and M059K), as measured by real-time PCR. Expression levels are normalized against the SVGp12 glial cell line, which served as the control. A statistically significant increase in expression was observed in the GBM8401 and A172 cell lines (*P < 0.05, **P < 0.01) when compared with levels in SVGp12.
Figure 2.
Relative PGC-1α mRNA Expression Between Glial and GBM Cell Lines. Comparison of PGC-1α mRNA expression between normal glial and GBM cell lines. Bar graph represents the relative expression levels of PGC-1α mRNA in normal glial cells (SVGp12) and various GBM cell lines (GBM8401, U87-MG, A172, G5T/VGH, DBTRG-05MG, and M059K), as measured by real-time PCR. Expression levels are normalized against the SVGp12 glial cell line, which served as the control. A statistically significant increase in expression was observed in the GBM8401 and A172 cell lines (*P < 0.05, **P < 0.01) when compared with levels in SVGp12.
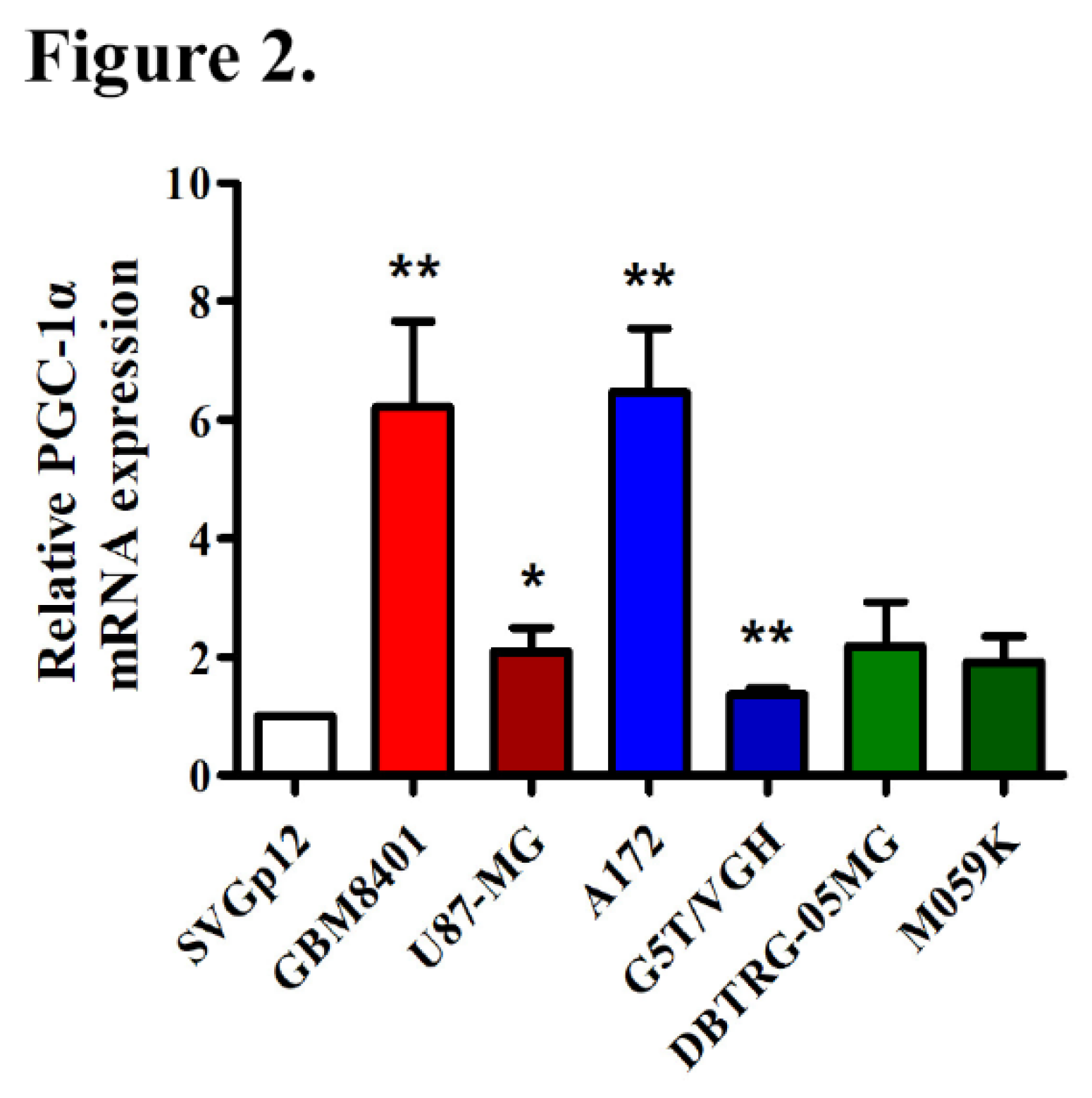
Figure 3.
Effects of PGC-1α Knockdown on mRNA and Protein Expression in GBM Cells. (A) Relative PGC-1α mRNA levels in GBM8401 and A172 cells. The control group was untreated cells. The negative control group was cells treated with non-targeting siRNA. The si-PGC-1α group included cells with transfected with PGC-1α siRNA. mRNA levels were significantly lower following PGC-1α knockdown than the control group for both GBM8401 and A172 (***P < 0.001, **P < 0.01). (B) Western blot images and quantitative analysis of PGC-1α protein expression. The protein bands for PGC-1α and β-actin are shown for GBM8401 and A172 cells. Quantification relative to β-actin demonstrates significantly lower levels of PGC-1α in the si-PGC-1α group than in the control group for both GBM8401 and A172 (***P < 0.001, **P < 0.01).
Figure 3.
Effects of PGC-1α Knockdown on mRNA and Protein Expression in GBM Cells. (A) Relative PGC-1α mRNA levels in GBM8401 and A172 cells. The control group was untreated cells. The negative control group was cells treated with non-targeting siRNA. The si-PGC-1α group included cells with transfected with PGC-1α siRNA. mRNA levels were significantly lower following PGC-1α knockdown than the control group for both GBM8401 and A172 (***P < 0.001, **P < 0.01). (B) Western blot images and quantitative analysis of PGC-1α protein expression. The protein bands for PGC-1α and β-actin are shown for GBM8401 and A172 cells. Quantification relative to β-actin demonstrates significantly lower levels of PGC-1α in the si-PGC-1α group than in the control group for both GBM8401 and A172 (***P < 0.001, **P < 0.01).
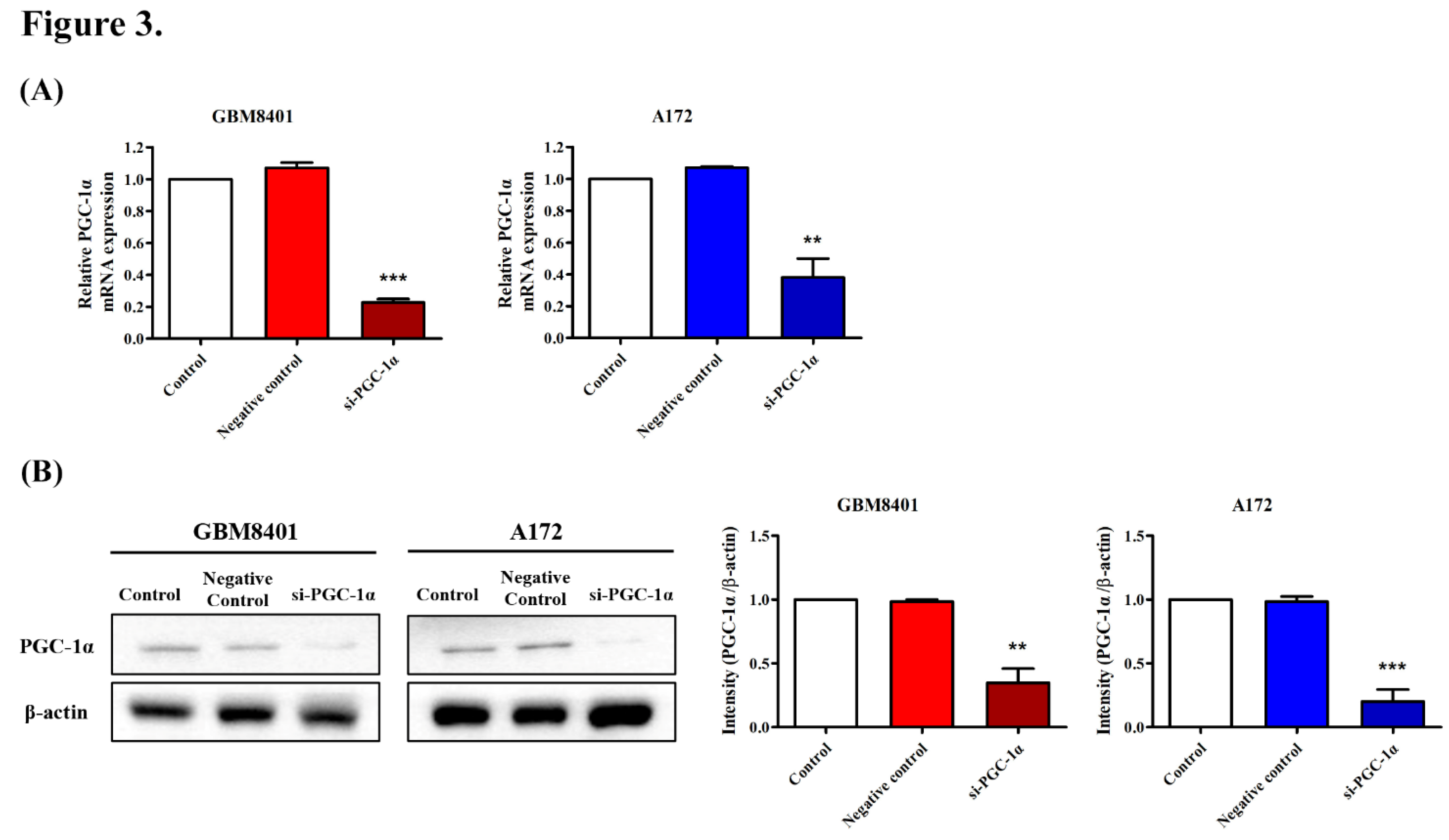
Figure 4.
Impact of PGC-1α Knockdown on GBM Cell Viability. Viability following PGC-1α siRNA transfection in GBM cells. Cell viability is shown for the control, negative control, and si-PGC-1α-transfected groups for GBM8401 and A172 cells at 1, 2, and 3 days post-transfection. The results indicate a significant decrease in cell viability in the si-PGC-1α groups over time (*P < 0.05, **P < 0.01, and ***P < 0.001).
Figure 4.
Impact of PGC-1α Knockdown on GBM Cell Viability. Viability following PGC-1α siRNA transfection in GBM cells. Cell viability is shown for the control, negative control, and si-PGC-1α-transfected groups for GBM8401 and A172 cells at 1, 2, and 3 days post-transfection. The results indicate a significant decrease in cell viability in the si-PGC-1α groups over time (*P < 0.05, **P < 0.01, and ***P < 0.001).
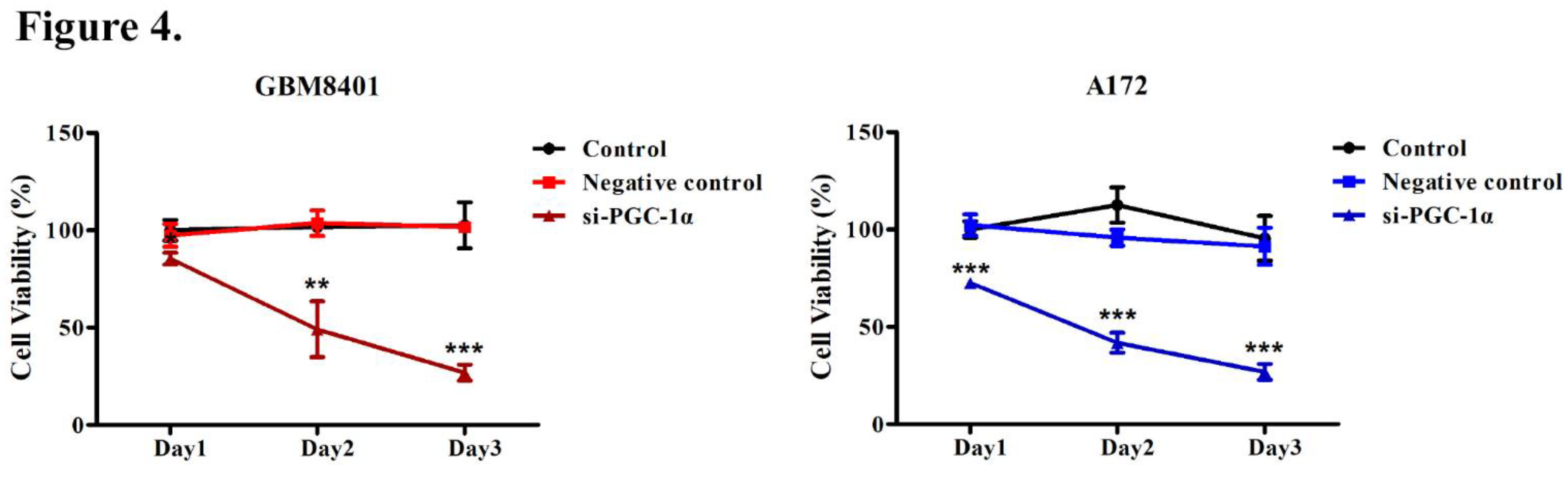
Figure 5.
Migration Assay of GBM Cells Following PGC-1α Knockdown. A wound healing assay post-PGC-1α siRNA transfection in GBM cells. The assay was conducted on GBM8401 and A172 cells divided into three groups: control, negative control, and si-PGC-1α. Time-lapse images were captured at 0, 8, 12, and 24 h post-transfection to assess the migration rate. Bar graphs quantify the percentage of wound closure, with significant inhibitory effects observed in the si-PGC-1α group compared with estimates in controls at later time points. Statistical significance was observed at 12 and 24 h post-transfection (*P < 0.05, **P < 0.01, ***P < 0.001).
Figure 5.
Migration Assay of GBM Cells Following PGC-1α Knockdown. A wound healing assay post-PGC-1α siRNA transfection in GBM cells. The assay was conducted on GBM8401 and A172 cells divided into three groups: control, negative control, and si-PGC-1α. Time-lapse images were captured at 0, 8, 12, and 24 h post-transfection to assess the migration rate. Bar graphs quantify the percentage of wound closure, with significant inhibitory effects observed in the si-PGC-1α group compared with estimates in controls at later time points. Statistical significance was observed at 12 and 24 h post-transfection (*P < 0.05, **P < 0.01, ***P < 0.001).
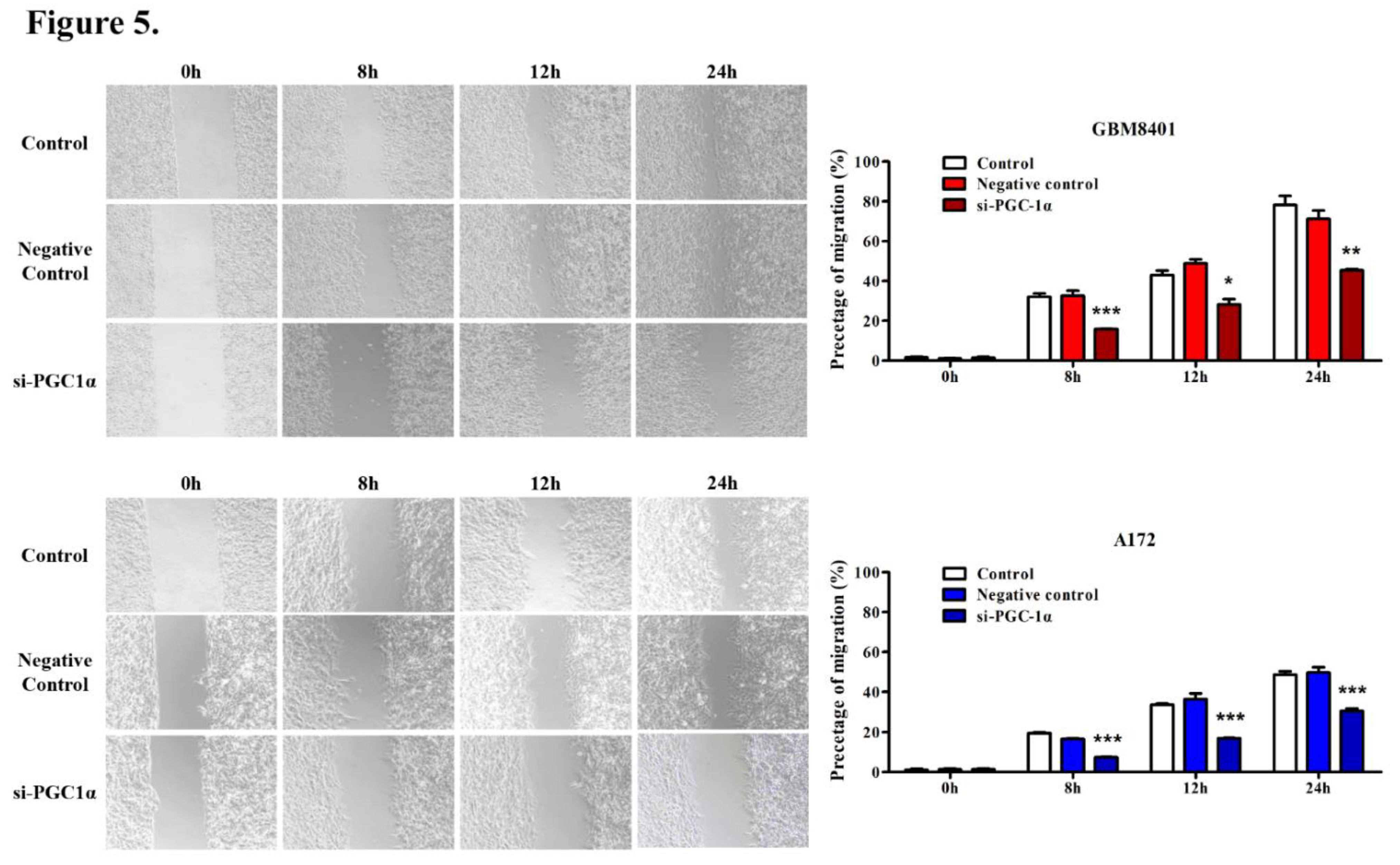
Figure 6.
Invasion Assay of GBM Cells Following PGC-1α Knockdown. Transwell invasion assay following PGC-1α siRNA transfection in GBM cells. Displayed are images of GBM8401 and A172 cells from the control group, negative control group, and si-PGC-1alpha group. The bar graphs to the right show the percentage of invasion, revealing a significant reduction in cell invasion in the si-PGC-1α group compared with the control group in GBM8401 and A172 cells (***P < 0.001).
Figure 6.
Invasion Assay of GBM Cells Following PGC-1α Knockdown. Transwell invasion assay following PGC-1α siRNA transfection in GBM cells. Displayed are images of GBM8401 and A172 cells from the control group, negative control group, and si-PGC-1alpha group. The bar graphs to the right show the percentage of invasion, revealing a significant reduction in cell invasion in the si-PGC-1α group compared with the control group in GBM8401 and A172 cells (***P < 0.001).
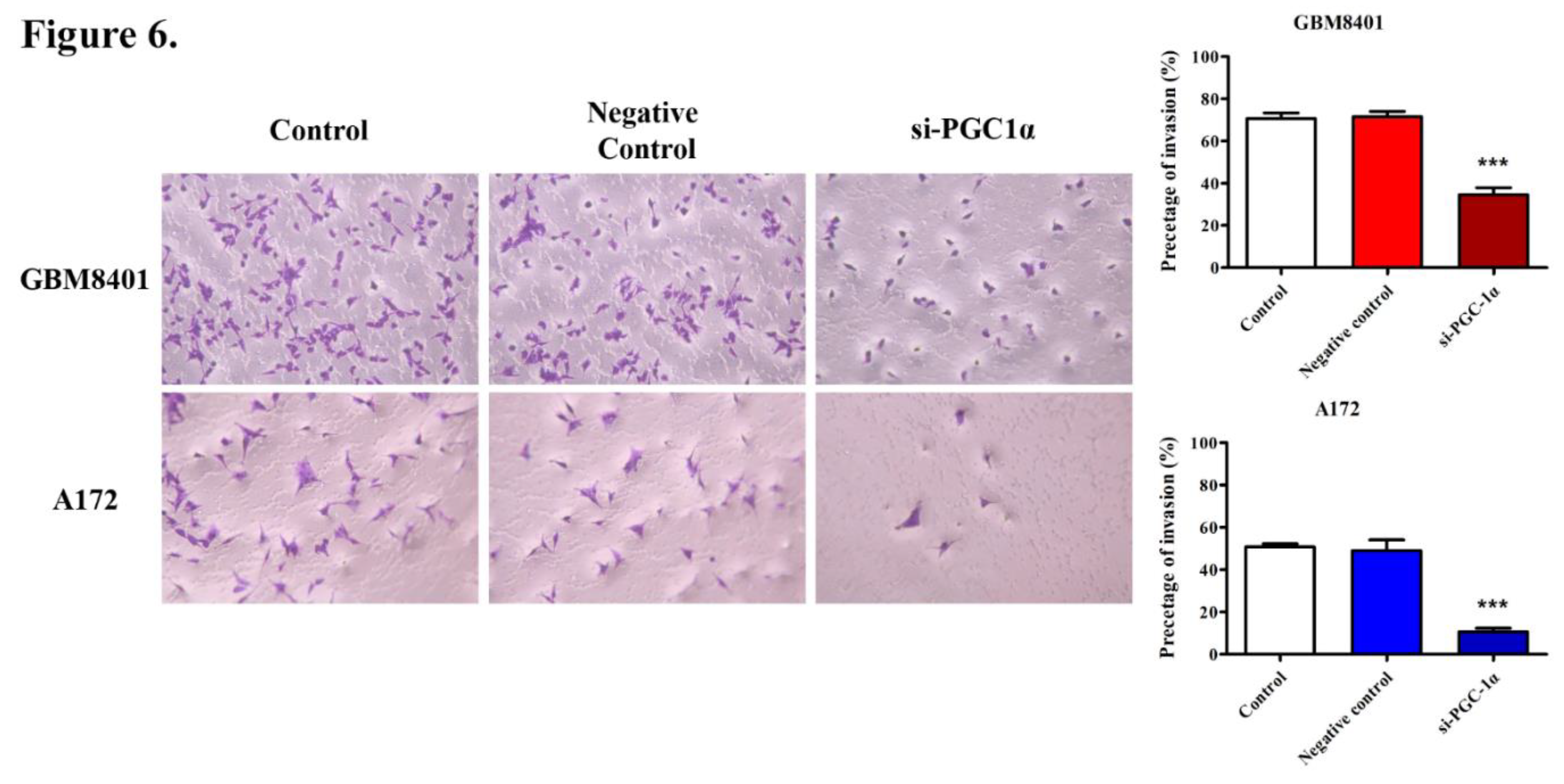

Table 1.
Correlation of PGC-1α expression with clinicopathologic parameters in patients with glioma.
Table 1.
Correlation of PGC-1α expression with clinicopathologic parameters in patients with glioma.
| No. of patients | PGC-1α expression (n, %) | P-value | ||
|---|---|---|---|---|
| Low | High | |||
| Age | 0.342 | |||
| <60 | 71 | 29 (30.5%) | 42 (44.2%) | |
| >60 | 24 | 7 (7.4%) | 17 (17.9%) | |
| Gender | 1.000 | |||
| Male | 52 | 20 (21.1%) | 32 (33.7%) | |
| Female | 43 | 16 (16.8%) | 27 (28.4%) | |
| WHO Grade | 0.003** | |||
| II | 23 | 15 (15.8%) | 8 (8.4%) | |
| III/IV | 72 | 21 (22.1%) | 51 (53.7%) | |
| Tumor size | 1 | |||
| ≤3cm | 59 | 22 (23.2%) | 37 (38.9%) | |
| >3cm | 36 | 14 (14.7%) | 22 (23.2%) | |
| Radiotherapy | 1.000 | |||
| No | 54 | 20 (21.1%) | 34 (38.9%) | |
| Yes | 41 | 16 (16.8%) | 25 (26.3%) | |
| Chemotherapy | 0.052 | |||
| No | 59 | 27 (28.4%) | 32 (33.7%) | |
| Yes | 36 | 9 (9.5%) | 27 (28.4%) | |
| KPS | 0.643 | |||
| ≤70 | 67 | 24 (25.3%) | 43 (45.3%) | |
| >70 | 28 | 12 (12.6%) | 16 (16.8%) | |
**Statistical significance (P < 0.01). WHO, World Health Organization; KPS, Karnofsky performance score.
Table 2.
Univariate and multivariate Cox regression analyses of prognostic parameters in patients with glioma.
Table 2.
Univariate and multivariate Cox regression analyses of prognostic parameters in patients with glioma.
| Univariate analysis | Multivariate analysis | |||||
|---|---|---|---|---|---|---|
| Relative risk | 95% CI | P | Relative risk | 95% CI | P | |
| Age | 0.648 | 0.311-1.351 | 0.247 | |||
| Gender | 0.892 | 0.473-1.682 | 0.724 | |||
| WHO grade | 0.382 | 0.171-0.851 | 0.019 | 0.546 | 0.223-1.335 | 0.185 |
| Tumor size | 1.114 | 0.572-2.171 | 0.751 | |||
| Radiotherapy | 1.845 | 0.965-3.526 | 0.064 | |||
| Chemotherapy | 1.542 | 0.776-3.065 | 0.217 | |||
| KPS | 1.167 | 0.586-2.324 | 0.660 | |||
| PGC-1α expression | 0.177 | 0.075-0.417 | <0.001 | 0.197 | 0.080-0.482 | <0.001 |
*Statistical significance (P < 0.05). CI, confidence interval; WHO, World Health Organization; KPS, Karnofsky performance score.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



