
Submitted:
03 April 2024
Posted:
03 April 2024
You are already at the latest version
Abstract
The rest-breaking agent, hydrogen cyanamide (HC), can substitute insufficient chill unit accumulation in Vitis vinifera and induce uniform bud-break; however, due to its toxicity it is being banned. In South Africa, red seedless grapes, including V. vinifera Crimson Seedless (CS), are the largest table grape export group; therefore, replacing HC in V. vinifera CS is crucial. This study aimed to confirm the molecular triggers induced by HC and assess the bud-break-enhancing abilities of commercial plant biostimulants. Forced bud-break assay experiments using V. vinifera CS single-node cuttings and a small-scale field trial were performed. Results demonstrated that increased chill unit accumulation (CUA), reduced HC efficacy. Bud-break started between 10 and 20 days after treatment, irrespective of final CUA. The small-scale field trial found that HC3 % resulted in a slightly delayed bud-break rate, whilst biostimulants were similar to the negative control. The treatment of dormant grapevine compound buds with nitric oxide (NO), hydrogen peroxide (H2O2), and hypoxia trigger dormancy-release to a certain extent, supporting the molecular models proposed for HC action. NO, H2O2, and hypoxia, in combination with PBs, may potentially replace HC; however, this needs to be confirmed in future experiments.
Keywords:
Vitis vinifera
; Crimson Seedless
; bud break
; plant biostimulant
; hypoxia
1. Introduction
Woody perennial crops survive cold winter months by suspending visible growth. This phenomenon is called dormancy [1]. When crops are dormant, compound buds are covered by bud scales and their vascular systems are physically separated from the rest of the vine [2]. This protects reproductive tissues, which were formed during the previous growth season. Only upon environmental cues (e.g., warming and increasing daylight hours) do the vascular systems of the buds re-connect to the cane for growth and differentiation to begin [2]. This is known as bud-break, defined as the day the first green tissue becomes visible underneath the bud scales [1]. Rapid, uniform bud-break is vital for optimal fruit set and quality, which is directly affected by the number of chill units (CU) accumulated during endodormancy.
Vitis vinifera generally requires a minimum of 200 hours of exposure to temperatures between 0 and 10°C to achieve a high bud-break percentage, defined as a bud-break of 80% or above [3]. Furthermore, to achieve high and uniform bud-break percentages across vineyards, a minimum exposure time of 400 h at 3°C is necessary [3]. However, slight variations in cold accumulation requirements may occur due to crop and cultivar differences [4]. CU models provide climate predictions and trends that affect the development of grapevine compound buds and can thus be used to determine optimal farming strategies.
Commonly used CU models include the Dokoozlian model, the Richardson (Utah) model, the Infruitec model (daily positive Utah model), and the dynamic model [4,5,6,7,8]. The Dokoozlian model calculates the ratio between exposure to cold temperatures (hours < 7°C) and cold-negating temperatures (hours > 20°C)[4]. The Richardson and Infruitec models are very similar regarding unit accumulation by allocating units per hour and suggesting that full units are accumulated at temperatures between 2.5 and 9.1°C [8]. The dynamic model is more complex, taking multiple factors into account, including sequences of both cold and hot temperatures during winter, and determines how much chill was reversed by the latter [6,7,8]. Furthermore, it is more accurate than the other models [8,9]. Nevertheless, all these models are used by South African fruit growers [4,8]. The choice of which model to use depends on the preferences of individual farmers and field experts.
South Africa has five table grape production regions: the Northern Provinces, Orange River, Olifants River, Berg River and Hex River (Fig. 3.1A: SATI 2018). The Northern Provinces, Orange River and Olifants River accumulate insufficient CU to achieve high percentages of bud-break (Figure S1a) [4,10]. Whilst the Berg River and Hex River regions accumulate enough CU for a high percentage bud-break, they do not always accumulate enough CU for uniform bud-break. The rest-breaking agent Dormex®, of which the active ingredient is 49% (w/v) hydrogen cyanamide (HC), can substitute insufficient chill unit accumulation (CUA) in crops and induce uniform bud-break. Dormex® is typically applied at a concentration of 3% or 5% (v/v) about four weeks before the expected onset of natural bud-break.
Regardless of cold accumulation in South Africa, farmers apply HC to guarantee high bud-break percentages and rapid and uniform dormancy release in their vineyards [4]. The need for the use of HC by fruit growers is predicted to increase with the effects of climate change and a general warming of growing regions [11]. HC is toxic, and its use has either already been banned or is predicted to be banned in several countries [12]. This is especially concerning as markets are sensitive, emphasizing the importance of looking for environmentally friendly production practices for grape products. The five largest export markets for South African table grapes are the European Union (54%), United Kingdom (22%), Canada (6%), Middle East (5%), and Southeast Asia (5%) [13]. Within each of these markets, excluding Southeast Asia, red seedless grapes are the largest table grape export group, including V. vinifera Crimson Seedless (CS) (Figure S1b) [13]. V. vinifera CS is a late-season red seedless table grape cultivar and is the most widely planted table grape cultivar in South Africa, with 3798 and 3660 hectares planted in 2021 and 2022, respectively [13]. Therefore, finding a replacement for HC in V. vinifera CS should be a priority for the South African agricultural sector.
A significant amount of research to date has focused on the molecular mode of action of HC on grapevine compound buds [1,14,15,16,17,18,19,20,21,22,23]. Recently, two models of the molecular mode of action of HC and bud-break have been reported [24,25]. Briefly, HC inhibits catalase (CAT), which creates a hypoxic environment in the buds, subsequently inducing fermentative metabolism, which is associated with an increase in nitric oxide (NO) and reactive oxygen species (ROS) [26]. The subsequent ROS and NO species signalling triggers the expression of transcription factors that upregulate genes related to growth and differentiation [24,25]. Additionally, phytohormone profiles change in the buds during dormancy release. For instance, abscisic acid (ABA) is well known to maintain bud dormancy, whilst ethylene is associated with dormancy release [25]. Furthermore, in grapevine buds, the exogenous application of ethylene has both been found to enhance dormancy release and act as a key signalling molecule within the antioxidant defence system [17,22].
The proposed molecular mechanisms of HC have assisted in developing less toxic commercial alternatives, such as plant biostimulants (PBs), which have been sold amongst agrochemical products over the last few years. Indeed, several PBs have shown a potential to enhance bud-break, compared to no treatment, in apple, grapevine, pear, sweet cherry, blackberry, peach, and kiwi [27,28,29,30,31,32,33,34]. However, very few reports, especially in the grapevine, exist of PBs that enhanced bud-break in a similar or superior manner to that of HC. Nevertheless, the success of PBs reported in bud-break studies may be attributed to their ability to cause major oxidative stress in plants whilst providing nutrients necessary for cellular and tissue recovery [35]. It is typical for such treatments to be amino acid and nitrite salt-based and to be applied in combination with an adjuvant or oil-based substance as well as additional supplementation of NO and calcium containing biochemical agents [35]. Interestingly, all these elements have been reported in the molecular models of HC action during bud-break [24,25,26].
Oxidative stress induced by PB-based treatments is likely attributed to the formation of respiratory stress and NO formation. Nitrites and nitrates form enzymatically via the activation of nitrogen nitrite and nitrate reductases under hypoxic conditions [30]. These conditions can be artificially created by commercial rest-breaking oils or oil-based adjuvants. Respiratory stress induced by rest-breaking oils leads to ROS accumulation, such as hydrogen peroxide (H2O2), a molecular trigger for various processes involved in dormancy release [26]. Calcium is an essential nutrient for plant growth, which enters plant cells in its divalent cation form, calcium ion (Ca2+), but can be toxic when present in excessive amounts [36]. ROS, such as H2O2, produced by NADPH oxidase, activate Ca2+ channels, which causes cell expansion and growth [37].
NO is a type of ROS used and produced by plants and has several important regulatory functions. These include abiotic and biotic stress responses and are involved in various physiological processes, including dormancy release [38]. NO is a well-known trigger of dormancy release in seeds and bulbs [39,40]. Additionally, its accumulation in grapevine buds has also been well-documented after the exogenous application of HC [24,25,30]. Indeed, NO can inhibit cytochrome oxidase by competing with oxygen and reversibly inhibiting CAT [41,42]. Whilst the success of dormancy-breaking agrochemicals is attributed to their ability to induce oxidative stress, uncontrolled increases in ROS accumulation can damage cellular components [43].
Agrochemicals with high amino acid contents may protect against uncontrolled ROS levels. Indeed, changes in amino acid profile are associated with plants under major oxidative stress, and the exogenous application of several amino acids is known to improve antioxidant defence responses [44]. Protein degradation, which occurs because of oxidative stress, releases amino acids, which the plant uses to synthesise various protective metabolites. For instance, proline, arginine, glutamine, asparagine, and gamma-aminobutyric acid (GABA) are used to produce osmolytes, secondary metabolites, or are stored as organic nitrogen. Interestingly, GABA has been found to reduce H2O2 during grapevine bud-break by activating VvCAT2 while repressing VvCAT1, both of which are isogenes of CAT [22]. Furthermore, aromatic amino acids, such as phenylalanine and tyrosine, are direct precursors for pathways related to hormone, polyphenol, and cell wall biosynthesis [44]. Similarly, branch-chain amino acids, such as isoleucine and valine, are used to produce cyanogenic glycosides, glucosinolates and acyl sugars. Finally, sulfur-containing amino acids, such as methionine and cysteine, play key roles in sulfur metabolism, which shifts during oxidative stress. This certainly emphasizes the importance of including amino acids as a key ingredient in treatments which induce bud-break.
A common method for studying dormancy in grapevine is single node cuttings (SNCs) under forced conditions. This approach is useful, as buds display behavioural similarities to those in the vineyard [17,22,45]. Another benefit of such an approach is that it enables the evaluation of the depth of dormancy via eliminating para- and ecodormant factors. Considering this, the current study aimed first to confirm the various molecular triggers induced by HC in dormant grapevine buds [24,25]. This was achieved via the external application of mineral oil, H2O2, and ethylene and two slow-releasing NO donors: diethylamine NONOate sodium salt hydrate (NO2) and S-nitroso glutathione. The second aim was to assess the bud-break enhancing abilities of four commercially available PBs: a riboflavin derivative (7,8 dimethylalloxazine), a citrus-based plant extract, an L-glutamic acid-based agrochemical, and a nitrogen and amino acid-based agrochemical. These aims were achieved via the execution of a series of forced bud-break assay experiments using V. vinifera CS SNCs over two years (with material from two growth seasons) and performing a small-scale field trial, which was conducted in the second year.
2. Materials and Methods
2.1. Plant Material
All assays were conducted using V. vinifera CS plant material from the 9-year-old CS block (Ramsey rootstock) at Windmeul Farm, Hoekstra Fruit, in Paarl, Western Cape, South Africa. Each row in the CS vineyard block consists of twenty-one line posts, with four vines between posts. The open gable trellis design consists of six catch wires, of which the bottom three are typically used for binding the one-year-old canes after winter pruning (Figure 1a).
2.2. Chill Unit Recordings
Local weather data was acquired from the Stratus 200 4G weather station (Hortec, Somerset West, Western Cape, South Africa, https://www.hortec.co.za/), which is located on Môrewag farm, Paarl (approx. 3.6 km from the experimental site). The CUs in this study were expressed as Infruitec Units (IUs) as this is the measure commonly used by local farmers and industry experts [8].
2.3. Forced Bud-Break Assays
2.3.1. Preparation of Plant Material, Experimental Design & Data Collection
Dormant ten node canes in E-L stage 1 [46], were collected from three middle rows of the CS block during May, June and July (2021 and 2022), before the accumulation of the final total CU required for each experiment (Figure 1a). Canes were separated into SNCs. These SNCs were soaked in contact fungicide (Captab 500 g/L), drained for a few minutes, and placed into heavy-duty plastic bags, then sealed.
The SNCs were stored at 4 °C in darkness until the pre-determined total number of IUs was reached. Finally, the exposed buds were dipped in the various respective treatments. To prevent dehydration of the SNCs in between treatment steps, such as the separation of canes into SNCs and treatment application, they were placed upright in trays filled with tap water (Figure 1b). Following treatment, SNCs were positioned in styrofoam supports and floated on water in plastic trays at a bud-break forcing temperature of 22°C, with a 16/8 h light/dark photoperiod and light intensity of 35 µmol/m2/s (F36W/GRO, GRO-LUX T8, Sylvania, Germany, https://www.sylvania-lighting.com/), (Figure 1c) [4]. Temperature was monitored with Tinytag TGP-4500 loggers (RS Components, Midrand, South Africa, https://za.rs-online.com/web/), (Figure S1). The location of individual SNCs in the supports was determined using randomized block experimental designs. The water in the trays was replaced every 14 days.
All laboratory equipment used for treatment preparation was thoroughly rinsed with distilled water (dH2O) before use. The SNCs were monitored for bud-break daily, with the bud-break date being defined as the date at which green tissue became visible underneath the bud scales (Figure 1d) [1]. More specifically, this is defined as E-L stage 3 and E-L stage 4 [46].
2.3.2 Candidate Treatments & Application Concentrations
During the 2021 and 2022 winter seasons, a series of forced bud-break assays were performed, which varied in the final accumulated CU. In each experiment, dH2O served as the negative control and 3% Dormex® as the positive control (referred to as dH2O and HC 3%, respectively, in all experiments).
In 2021, several biochemical agents and two PBs (BC204 and lumichrome) were tested individually (Table 1). In 2022, two agrochemical-based PBs were tested at the manufacturers’ recommended concentrations and lower concentrations, along with a mixture treatment of all promising biochemical agents and PBs included in the 2021 assays (Table 2). At the request of the supply companies, all commercial biostimulant and bud-break product treatments were assigned code names.
2.4. Small-Scale Field Trial Dormancy-Release Assay
2.4.1. Experimental Site and Design
A small-scale field trial was conducted to test the two PBs included in the glasshouse assays of 2022. This trial was executed during September and October 2022 in two middle rows of the CS block. Treatments were applied in replicates of eight groups of two plants between individual sets of line posts, using a randomised block design. The two middle vines within each post were the experimental targets. Four canes facing opposite directions of the vine, bound to the bottom wire of the gable trellis system, were selected as experimental target canes. The first 10 buds of each cane were clearly marked for analysis using white acrylic paint, starting from the head of the vine (Figure 2a). To prevent contamination by commercial spray applications, three guard rows on either side and at least two guard line posts on the terminal ends of each row were included in the experimental design (Figure 2b).
2.4.2. Treatments Evaluated and Dosage Concentrations
Treatments evaluated during the small-scale field trial include SNH & PBXH (Table 2). The positive control was 3% Dormex®, while the negative control was the tap water used to make up the treatments on the farm (which was referred to as dH2O regardless). With the assistance of Curativo (Pty) Ltd (Wellington, South Africa), treatments were applied at a spray volume of 611 L/ha using STHIL SR 430 backpack mist blowers (Stuttgart, Germany, https://www.stihl.com/default.aspx) within the first three days after winter pruning had taken place (approx. two weeks before expected natural bud-break date). Target compound buds were monitored daily for dormancy release, using E-L stage 4 as a reference [46].
2.5. Data Handling
2.6. Statistical Analysis
Using the R package drc, a four-parameter logistic growth curve was fitted to determine the final bud-break percentage, rate, and EC50 (Equation 2) [52], and a piecewise linear regression model was fitted to determine the onset (Equation 3) [30].
Listed below are the four parameters which were used to assess bud-break datasets of the different treatments, along with their definitions according to Equations 2 and 3 (Figure S3):
- Onset: The number of days until the bud-break date was reached. Larger positive values indicate delayed time until onset was reached.
- Bud-break rate: The slope between 0 % bud-break and final bud-break % indicates bud-break rate between the first and last buds that have broken. The slope parameter (or more generally termed “hillslope”) indicates the steepness of decrease. Thus, the larger the slope value, the steeper the decrease. For an increasing function (such as in the current study), the opposite is true: the larger the negative slope is, the steeper the increase, and the faster the bud-break rate.
- Half maximal effective concentration (EC50): Number of days to reach 50 % bud-break after treatment. This is also a parameter indicating speed. Larger positive values indicate delayed time until EC50 was reached.
- Final bud-break percentage: The upper value of the cumulative bud-break percentage growth curve (plateau). Larger positive values indicate a higher final bud-break percentage.
An unadjusted post hoc t-test, with a 5% confidence interval (p<0.05), was used to determine significant differences between growth curve parameters [53].
3. Results
3.1. Chill Units in the VINEYARD
CU recordings, which were measured as IUs in the vineyard by the Môrewag Stratus weather station, were used for all dormancy-release assays in the current study, including glasshouse forced bud-break trials performed in the laboratory and the small-scale field trial (Figure S4). In 2021, 57.5 CU were accumulated by the end of May and reached a maximum of 795.5 CU by the end of October. In 2022, a total of 63.5 CU was accumulated by the end of May, and maximum of 662 CU were reached by the end of October. Furthermore, during each month after May, a higher final monthly number of CU were accumulated in 2021 than in 2022. In both years, CU started to accumulate in May, reaching final values of 57.5 (2021) and 63.5 (2022) respectively. By the end of July in both years, enough CU (479.5 in 2021, & 368 in 2022) had accumulated for rapid, uniform dormancy release to take place in the V. vinifera CS block [3,54,55].
3.2. Forced bud-Break Assays
3.2.1. Evaluation of Plant Biostimulants (BC204 and Lumichrome), and Individual Biochemical Agents
During June 2021, the effects of the following treatments were evaluated on bud-break in V. vinifera CS SNCs after 100, 200 and 400 CUA: HC 3%, dH2O, BC204 0.05%, Lum 5 nM, H2O2 1%, Eth 1000 ppm, BB 3%, SNO 10 μM, and NO2 10 μM (Table 1). Each treatment consisted of 30 biological replicates. All samples were collected on 31 May 2021, prepared, and left at 4 °C until the desired final total CU had accumulated. At the time of sampling, 57.5 natural CU had already accumulated in the vineyard (Figure S4).
Cumulative plots of bud-break % across the three experiments showed that with increased CUA, the effect of Dormex® (HC 3%) was reduced in efficacy compared to both the negative control and other treatments (Figure 3). This indicates that there was less need for Dormex® with increased CUA. An interesting observation was that the days until bud-break was first observed lengthened with increasing CU values. After 100, 200 and 400 CUA, bud-break was first observed after about 9 (Figure 3a), 16 (Figure 3b) and 22 (Figure 3c) days respectively. These longer periods before bud-break onset with more CUA may be due to increased exposure to cold stress of SNCs, which had accumulated more CU during storage for artificial cold accumulation. Nevertheless, clear trends were observed amongst the experiments after careful evaluation of bud-break onset, rate, EC50 and final cumulative % datasets.
After 100 and 200 CUA, HC 3%-treated buds took 9.86 and 16.12 days to reach onset, respectively, significantly less than the other treatments (p=0) (Table 3). In contrast, after 400 CUA, HC 3%-treated buds had an onset of 25.4 days, similar to the other treatments. The negative control, dH2O, yielded onset values of 20.07, 22.17 and 27.08 days after 100, 200 and 400 CUA, respectively. Two treatments, H2O2 1% and BB 3%, resulted in significantly less days for bud-break onset to be reached than dH2O in all three experiments (p=0). Furthermore, the onset after BB 3% treatment was earlier than that of H2O2 1% in each experiment. The onset of H2O2 1%-treated SNCs was 18.8, 21.07 and 24.48 days; for BB 3%, they were 16.48, 19.16 and 25.17 days after 100, 200 and 400 CUA, respectively.
Bud-break rates of SNCs treated with dH2O were -0.37, -0,46 and -0,31, and for HC 3%, -0.48, -0.48 and -0.71 after 100, 200 and 400 CUA, respectively (the larger negative the slope is, the steeper the increase of bud-break rate), (Table 3). The HC 3% treatment rates were significantly higher than the negative control after 100 and 400 CUA (p=0). Additionally, compared to the negative control, buds treated with NO2 10 μM consistently had significantly higher rates of -0.43, -0.54 and 0.39, respectively, in the order of increased CUA (p=0 to 0.02). In addition to this, the rate of NO2 10 μM after 200 CUA was significantly higher than that of HC 3% (p=0). Some interesting trends were also observed in the EC50 results.
The EC50 values of HC 3%-treated SNCs after 100, 200 and 400 CUA were 15.51, 21.32, and 25.86 respectively (Table 3). Across all experiments, these were significantly the lowest EC50 values (p=0). The EC50s of the negative control SNCs were 26.82, 27.99 and 35.39, in the order of increased CUA. Two treatments, H2O2 1% and BB3 %, had significantly lower EC50 values than dH2O in all three experiments (p=0 to 0.02), resulting in less days to reach 50% bud-break. These precise onset values after 100, 200 and 400 CUA were 24.77, 27.16, and 34.92 for H2O2 1 %, and 23.79, 25.82 and 33.52 for BB 3%. Bud-break EC50 was reached earlier following BB 3% treatment compared to H2O2 1% in each case (p=0). One treatment, Eth 1000 ppm, resulted in consistent delayed bud-break results compared to all other treatments.
The days it took SNCs treated with Eth 1000 ppm to reach onset were 20.93, 24.49 and 30.81 days after 100, 200 and 400 CUA, respectively (Table 3). Furthermore, SNCs treated with Eth 1000 ppm took 28.03, 31.71 and 36.50 days to reach EC50 after 100, 200 and 400 CUA respectively. These onsets and EC50 values were significantly higher than those of the negative and positive controls (p=0 to 0.02), excluding the onset of HC 3 % after 400 CUA, in which case the values were similar. Treatments did not result in major differences in the final bud-break %; however, there were still some significant differences seen in the datasets:
HC 3%-treated SNCs reached 95.95, 96.40 and 95.65 final bud-break %, whilst dH2O -treated SNCs reached 95.81, 97.48 and 100 final bud-break % after 100, 200 and 400 CUA respectively. In comparing the differences between these results, the higher % reached by dH2O-treated SNCs after 400 CUA was the only significant difference (p=0). Furthermore, there were only two treatments, SNO 10 μM and NO2 10 μM, which resulted in significantly higher bud-break percentages than dH2O in two of the three experiments (p=0 to 0.01). These experiments included SNCs with 100 and 200 CUA. The final bud-break percentages of SNO 10 μM and NO2 10 μM treated SNCs were 97.77 and 99.34 after 100 CUA and 99.38 and 99.27 after 200 CUA, respectively. Nevertheless, all treatments across all three experiments resulted in final bud-break percentages above 90%.
3.2.2. Evaluation of Plant Biostimulants SN and PBX, as Well as Biochemical Agents Combined with Lumichrome and BC204
In 2022, the effects of the following treatments were evaluated on bud-break in V. vinifera CS SNCs after 100, 200 and 250 CU: HC 3 %, dH2O, MIX2, SNH, SNL, PBXH, and PBXL. This selection of treatments included two commercially available PBs, SN and PBX, and a combination of treatments tested in the previous year, which was named MIX2. The CU chosen for each experiment was based on assay optimisation from last year. Each treatment consisted of 45 biological replicates, and each experiment was performed in duplicate (data for second replicates not shown).
Fresh samples were collected for each experiment between mid-June and mid-July 2022 at approximately the final desired CU (Table S1). This procedure was implemented to avoid potential stress that may be introduced by long periods of exposure to artificial CUA. The results were that SNCs with higher CUA had more time to mature on the vines before sampling. Visualisations of cumulative bud break over time revealed several general trends for further consideration.
Bud-break across treatments generally started between 10 and 20 days after treatment, irrespective of final CUA (Figure 4). Additionally, the range of onset values became more condensed with increased CUA. The earliest onset date recorded in each experiment was approx. 10 days. This shows that stress introduced by artificial CUA was successfully avoided. After 100 CUA, HC 3% generally resulted in advanced bud-break, compared to other treatments; however, it increasingly caused phytoinhibition compared to the negative control, dH2O, after 200 and 250 CUA. This is especially evident in the final bud-break percentage, which was close to 70 after 250 CUA. Most treatments generally resulted in cumulative bud-break curves with trends similar to that of the negative control across the three experiments. However, after 100 CUA, there were two exceptions, MIX2, and SNH, which had resulted in bud-break trends more similar to that of HC 3%. It was also noteworthy that SNH continued to produce bud-break curves similar in shape to that of HC 3% after 200 and 250 CUA. This included some phytoinhibitory results after treatment with SNH with higher CUA, particularly lower final bud-break %, which was around 80% after 250 CUA.
The negative control, dH2O, resulted in 18.20, 15.48 and 13.77 days after 100, 200 and 250 CUA until bud-break onset was reached (Table 4). After HC 3% treatment, bud-break was generally initiated early across experiments, compared to the negative control and other treatments. For instance, the onset of HC 3% treated SNCs after 100 CUA was 9.69 days, which was significantly lower than all other treatments (p=0). Compared to dH2O, the onset of HC 3% treated SNCs was significantly lower after 200 CUA (p=0), yet similar after 250 CUA, which were 10.35 and 13.72 days respectively. Compared to HC 3%, one treatment, SNH, resulted in equal or reduced days to onset in two of the three experiments. Firstly, SNH resulted in a similar onset of 9.81 days after 200 CU and a significantly lower onset after 250 CUA, which was 10.14 days (p=0). The latter also had the lowest onset compared to all treatments following 250 CUA.
Considering the bud-break rate, HC 3%-treated SNCs had significantly the slowest rates, which were -0.32, -0.29 and -0.23 after 100, 200 and 250 CUA, respectively (Table 4). The bud-break rates of the negative control, which were -0.52, -0.79 and -0.59 after 100, 200 and 250 CUA, were the most rapid in each experiment, with one exception. This exception was recorded after 100 CUA, during which MIX2 treatment resulted in significantly the highest rate of -0.62 compared to all treatments (p=0). The EC50 values of each experiment provided further information on the effect of treatment on the rate of bud-break between treatments:
Generally, HC 3%-treated buds took longer to reach EC50 with increased CUA and were eventually surpassed by both the negative control and all treatments after 250 CUA. For instance, after 100 CUA, the EC50 value recorded after HC 3% treatment of 17.02 was significantly the least days (p=0), and that of dH2O, which was 22.88, was significantly the most days (p=0) (Table 4). After 200 CUA, the EC50s of HC 3% and dH2O treated buds were 18.71 and 18.57 days, respectively, which were similar. Furthermore, after 250 CUA, HC 3%-treated SNCs took the most days, which was 24.96, to reach EC50 (p=0). The 17.99 days it took buds treated with the negative control in this experiment was average amongst treatments. Notably, after 200 and 250 CUA, SNH-treated SNCs significantly displayed the lowest EC50s, which were 14.05 and 15.67, respectively (p=0). After 100 CUA, this treatment resulted in an EC50 of 18.29, which was the second least, outcompeted only by HC % (p=0).
The final bud-break % of all the treatments across experiments resulted in values above 90%, except for HC 3% and SNH after 250 CUA (Table 4). The final bud-break percentage for HC 3% in this experiment was 70.71%, which was significantly the lowest (p=0). This was followed by the 81.03% recorded after SNH treatment, which was significantly lower than the negative control and other experimental treatments (p=0).
3.3.3. Small-Scale Field Trial: Evaluation of Plant Biostimulants SN and PBX
A small-scale field trial was started on the 6th of September 2022, during which the effect of dH2O, HC 3%, SNH and PBXH was evaluated. At the time of application, 627 CU had already accumulated in the V. vinifera CS block, which reached a maximum of 662 CU during the experiment (Figure 5). The cumulative bud-break plots show that HC 3% had resulted in a slightly delayed bud-break rate, whilst PBXH and SNH displayed results similar to that of the negative control.
Buds treated with the negative control took 6.53 days until bud-break was initiated, very similar to the 6.07 days it took PBXH buds to reach onset (Table 5). The treatment that significantly resulted in the least days to reach onset was SNH, which was achieved in 5.07 days (p=0). Furthermore, the onset of buds treated with HC 3% was 8.38, which was significantly the longest time recorded for this parameter (p=0). Nevertheless, in addition to resulting in the most days until onset, HC 3% treatment also resulted in the most days until EC50 was reached, which was 15.61 days. This is significantly longer than the 13.09, 12.62 and 12.74 days it took dH2O and PBXH to reach EC50, respectively (p=0). However, HC 3% reached a final bud-break % of 85.26, which was significantly the highest among treatments (p=0). The 80.20 and 82.28 final bud-break percentages reached by PBXH and SNH, respectively, were significantly higher than that of the negative control, dH2O (p=0).
4. Discussion
4.1. Forced Bud-Break Assays
4.1.1. Evaluation of Plant Biostimulants BC204, Lumichrome, and Candidate Biochemical Agents
The forced bud-break assays, which took place in 2021, evaluated the effects of two PBs, which were BC204 and lumichrome, as well as a selection of biochemical agents. The treatments included HC 3%, dH2O, BC204 0.05%, Lum 5 nM, H2O2 1%, Eth 1000 ppm, BB 3%, SNO 10 μM, and NO2 10 μM. Treatments were evaluated after insufficient CUA to reach a high percentage bud-break (100 CU), sufficient CUA for a high percentage bud-break (200 CU), and sufficient CUA to result in both a high percentage and uniform bud-break (400 CU) [3,4,54,55]. All samples were collected on the same day and were thus all at the same level of maturity across experiments. Overall, the results of the 2021 forced bud-break assays demonstrated the relationship between HC and CUA and provided insight into possible mixture compositions that could lead to new formulations or alternative treatment strategies.
Overall, HC 3% advanced bud-break onset the most effectively, however, this became less distinct with higher CUA (Figure 3). Similarly, bud-break rate was accelerated strongly by HC 3% treatment, which was only outcompeted by 10 μM NO2 (NO2 10 μM) after 200 CUA (Table 3). Interestingly, NO2 10 μM was also the only treatment that resulted in higher bud-break rates than the negative control in all three experiments (Table 3). What was also noteworthy considering NO treatments, was that both NO donors in the current study, including NO2 10 μM and 10 μM S-nitrosoglutathione (SNO 10 μM), resulted in enhanced final cumulative bud-break percentages in two of the three experiments (Table 3). Such results are in line with literature which has linked NO donors to bud dormancy-release, as well as its involvement in the HC-induced cascade [19,24,26,30,56,57].
In the current study, NO likely triggered hormone response in a similar fashion to HC [24,25,26]. For instance, it has been found that the exogenous application of NO donors, 8% (w/v) potassium nitrate (KNO3) and 6% (w/v) calcium nitrate (Ca [NO3]2), increased gibberellic acid and indoleacetic acid levels and a decreased that of ABA in dormant ‘Anna’ apple tree buds [58]. It was also reported that both donors hastened and improved bud-break percentage compared to the negative control; however, these results were not superior to those of Dormex®. In agreement, it was shown that in ‘Fuji’ apple tree shoots, the bud-break enhancing abilities of 0.1-2 M KNO3, and 0.05-1M sodium nitrite (NaNO2), applied in combination with 3% (v/v) mineral oil were also not superior to that of 3% (v/v) Dormex® [30]. However, it was reported that the enzymatic generation of NO via a combination of NaNO2 and ascorbic acid was, in some cases, equally effective to Dormex®. The product of this reaction directly generates NO and nitrites and does not rely on nitrite and/or nitrate reductases to produce NO. Indeed, this is a similar concept to using NO donors that release NO in a controlled manner once dissolved in water, as was done in the present study, and supports the bud-break-enhancing abilities of NO in current results. Besides NO donors, which have shown potential regarding rate and final bud-break percentage, two other treatments, 1% H2O2 (H2O2 1%), and 3% BUDBREAK® mineral oil (BB 3%), showed potential in enhancing onset and EC50 values.
Despite HC 3% treatment reducing the number of days it took to reach bud-break onset and EC50 the most in each experiment, which was not surprising, H2O2 1% and BB 3% produced results superior to the negative control (Table 3). ROS (H2O2) and hypoxia, which can be created artificially with mineral oils such as BB3 %, are clearly associated with dormancy-release in grapevine buds [24,25,26]. It was notable that BB 3% had produced slightly better results than 1% H2O2. Indeed, mineral and vegetable oils have been reported to enhance bud-break properties in woody perennial crops, such as grapevine and pear; however, they generally have not produced results superior to HC treatment [47,59,60]. This is in harmony with current results for BB 3%.
Compared to no treatment in ‘Carmen’ grapevine, 2% (v/v) vegetable oil and 2 % (v/v) mineral oil have been found to increase bud-break percentage, both independently and in combination with one another, which agrees with current results [47]. Additionally, it was shown that the combination of the two oils enhanced the bud-break percentage in a similar manner to 3% (v/v) HC, which may be considered for future optimisation purposes. Furthermore, 2% (v/v) mineral oil was reported to lower CAT activity within 24 h after treatment, compared to the negative control. These results are supported by another study in Asian ‘Hosui’ pear trees, in which 4% mineral oil also inhibited CAT activity [60]. This may explain the lower onset and EC50 values displayed by grapevine buds treated with BB 3% in current results since the inhibition of CAT is known to occur under hypoxic conditions, which induces bud-break [24].
Hydrogen peroxide is a key signalling compound in the physiological regulation of natural dormancy release [26,61]. In agreement with current results, it has been shown that H2O2 may partially promote bud-break in perennial crops such as pear, grapevine, and walnut [22,61,62,63,64]. For example, in V. vinifera Thompson Seedless, 1% (v/v) H2O2 enhanced bud-break percentage compared to the negative control; however, it was not as effective as 2.5% (w/v) HC [63]. Furthermore, it was suggested that after application, H2O2 was rapidly degraded by CAT and did not result in activation of grape dormancy-breaking related protein kinase, which is known to be upregulated by HC [1,63]. In support of this, it was reported that the application of 1 % (v/v) H2O2 to V. vinifera, Thompson Seedless buds that were in the stage of endo dormancy-release triggered genes encoding antioxidant enzymes, such as CAT [22].
The degradation by CAT may explain the inability of H2O2 treatment to produce results equal to that of HC in the current study. Additionally, it could explain the slightly inferior results compared to BB 3%, which likely activated anaerobic respiratory pathways that led to H2O2 accumulation and subsequent signalling [26]. It is known that H2O2 is a negative regulator of dormancy, which can even negatively affect bud-break and shoot growth when applied in too high concentrations or when buds have matured past the stage of endodormancy [61]. However, this may not have been the case in current results as 1% (v/v) H2O2 has been tested for bud-break by other research studies in grapevine, which reported similar trends in results [22,63].
Ethylene treatment using 0.206% v/v Ethephon®, an ethylene supplement (Eth 1000 ppm), resulted in prolonged bud-break compared to HC. Eth 1000 ppm-treated SNCs consistently took the greatest number of days to reach both bud-break onset and EC50 (Table 3). Indeed, it has been reported that the exogenous application of 500 and 1000 ppm ethephon, which is similar to the treatment concentration applied in the present study, prolonged dormancy-release in ‘Muscat of Alexandria’ grapevine plants grown in a greenhouse [65]. However, such results contrast reports that 100 ppm ethylene enhanced the bud-break of V. vinifera ‘Perlette’ cuttings [17]. In addition to this, another study on V. vinifera Thompson Seedless found that the exogenous application of 10 mg ethephon, dissolved in 10 mL 100 mM phosphate buffer, upregulated genes involved in oxidative stress responses, which is associated with bud-break [22]. Thus, the ability of ethylene supplementation to enhance bud-break traits may be subjected to optimal application concentrations.
4.1.2. Evaluation of Plant Biostimulants SN and PBX, as Well as Biochemical Agents Combined with Lumichrome and BC204
Forced bud-break assays, which took place in 2022, evaluated the effects of two controls, which were HC 3% and dH2O, two commercial PBs, which were SN and PB, and MIX2, which was a combination treatment consisting of treatments tested in the previous year, after 100, 200 and 250 CUA (Table 2). All treatments resulted in high final bud-break percentages, except for HC 3% and SNH after 250 CUA (Figure 4) [3,4,54,55]. The fact that HC 3% had displayed increased phytoinhibition at higher CUA was unexpected as this did not occur in the forced bud-break assays of 2021, during which buds had accumulated up to 400 CU, which is much higher than the maximum of 250 CUA in these results (Figure 3). A possible explanation for this phenomenon is that the SNCs for all experiments in the previous year were collected on the 31st of May, which at the time had accumulated only 14.3 to 50% of final CUA. In contrast, SNCs in current experiments were collected individually for each experiment on slightly later dates, 10th of June to 12th of July, and had accumulated 80 to 95% of the final CUA. Buds in the later experiments were thus allowed a longer time to age on the vine.
The reasoning behind the strategy of individual sampling per experiment was to bypass any stress introduced during artificial cold accumulation in 2022. Seemingly, this was successful, with no overall delay in onset between experiments in 2022, as seen in the previous year (Figure 4). However, the introduction of variation in bud maturity should be considered in future studies. Indeed, the optimal timing of application of forced bud-break agents such as HC is vital, and early application may cause uneven bud-break. In contrast, late application risks bud damage due to phytoinhibition or other stress factors [66]. Considering this, it is most likely that after 250 CUA, the buds were in a metabolic state unsuitable for HC treatment.
One of the most noteworthy outcomes was that after 250 CUA, SNH treatment had negatively affected final bud-break %, in a similar manner, albeit to a lesser extent compared to HC 3% (Figure 4). Cumulative bud-break percentage plots of SNCs treated with SNH were also quite similar in shape to those after HC 3% treatment. These results suggest that SNH may have induced similar physiological changes within dormant buds to HC, whilst being less phytoinhibitory than HC, or simultaneously providing nutrients, which aids in stress response and cell recovery. Indeed, chemical and physical methods other than HC that are known to induce bud-break, including heat shock, sodium azide, and hypoxia, have been found to simulate the pathways induced by HC in grapevine [17,22,24,58]. Besides the bud-break percentage, the onset and EC50 values of HC 3% and SNH were superior to other treatments and the negative control (Table 4). Furthermore, SNH treatment generally accelerated the onset and EC50.
SNH consists of two synergistically-acting substances, including 2% (v/v) SBB-01 and 20 % (v/v) NDY-01 (code names were supplied by manufacturer/supplier). Both SBB-01 and NDY-01 are rich in various forms of nitrogen. Additionally, SBB-01 is an amino acid-based biostimulant, and NDY-01 contains water-soluble calcium (CaO), which could enhance Ca signaling similarly to HC [24]. Indeed, the most common agrochemicals evaluated for dormancy release are amino acid and nitrite salt-based [35]. The performance of such agrochemicals has been tested in crops such as kiwi, apple, sweet cherry, grapevine, blackberry, and peach [23,28,31,32,33,34,67,68]. Generally, treatments showed enhancing effects on bud-break and, in some cases, resulted in similar or superior results compared to HC, which are aligned with current results for SNH. For instance, in ‘Gala’ and ‘Fuji’ apple trees, it was demonstrated that Erger®, Synchron® and Vorax® in combination with mineral oil or calcium nitrate, can all effectively enhance axillary and terminal bud-break percentages [23]. Overall results varied compared to HC treatment, with Synchron® performing better, Erger® similarly, and Vorax® worse than HC. In another study, Erger® application with calcium nitrate was tested in the ‘Fiano’ grapevine cultivar, during which 3, 5 and 7% (v/v) Erger® enhanced bud-break compared to the control, but not compared to 4% (v/v) Dormex® [33]. Nevertheless, one treatment, which was MIX2, in the current study showed the potential to enhance the bud-break rate.
Bud-break rates of the negative controls were not better than HC 3%, nor any other treatment across experiments, except MIX2 after 100 CU. This treatment produced a notable result. MIX2 treatment probably introduced hypoxia, as well as ROS and NO signalling. This may have been due to the inclusion of BB 3%, H2O2 1%, NO2 10 μM and SNO 10 μM in the treatment. The relevance of these individual treatments to dormancy release has previously been explored. However, two PBs were also included in MIX2, which were 5 nM lumichrome and 0.05% (v/v) BC204.
All the components of MIX2 were evaluated independently during the 2021 forced bud-break assays, all of which had shown some potential in enhancing bud-break, except for the two PBs. Nevertheless, a synergistic interaction between two or more components may have been at play, which ultimately enhanced bud-break rate and/or stress tolerance in the SNCs. In addition to activating anaerobic metabolism, mineral oil prevents evaporation and ensures the penetration of treatments into plant tissue [51]. The mineral oil likely improved the uptake of the other ingredients in the treatment. Collectively, interference with aerobic respiration, as well as NO and ROS signalling, are likely to enhance the bud-break rate in a similar fashion to the way HC works on dormant buds [24,25,26].
The two PBs may have promoted growth in the presence of oxidative stress introduced by the other ingredients of MIX2. Their enhanced performance in the current treatment may have been explained by their application and BB3 %, which may have improved treatment uptake. BC204, which is currently being used for colour enhancement on various crops, including CS table grapes, has been found to increase crop yield, fruit quality, plant health and stress response [50]. For example, 0.01% (v/v) BC204 enhanced Arabidopsis thaliana growth, even in the presence of osmotic stress introduced by 100 mM NaCl. An RNA-seq study showed that in A. thaliana shoot tissue, BC204 differentially affected genes related to dormancy release, such as cell wall synthesis, carbon metabolism, transcription factors, calcium regulation and phytohormones [50]. Furthermore, 5 nM lumichrome increased photosynthesis and enhanced plant growth via turgor-driven cell expansion (upregulation of XTH9 & XPA4) in A. thaliana [49]. However, these present data indicate that BC204 and lumichrome do not seem to contribute to dormancy release in grapevine buds. However, they may be found to enhance the growth and general health of the shoots that emerge post-bud-break upon further evaluations.
4.2. Small-Scale Field Trial: Evaluation of Plant Biostimulants SN and PBX
In 2022, a small-scale field trial was conducted, during which the effects of HC 3 %, dH2O, SNH, and PBXH on bud-break were evaluated. Treatments were applied approx. Two weeks before predicted bud-break, in accordance with commercial guidelines and pruning schedules based on grower advice. The Berg River region, where the trial was conducted, may simply not have been the optimal location to conduct bud-break trials, considering high numbers of CU (627 CUA) that had accumulated by the time that the trial was initiated (Figure 5). This is well above the required CU for high and even bud-break, which eliminated the need for artificial dormancy-release [3,4,54,55].
However, using HC locally in the Paarl area is implemented as a routine treatment amongst most growers. Nonetheless, HC 3% treated buds took the longest to reach EC50, suggesting that HC may negatively affect bud-break uniformity when applied to buds already receiving sufficient CU (Table 5)[66]. However, HC 3% achieved the highest final bud-break % among all treatments. The two PBs displayed qualities similar to that of water. However, future more expansive controlled field trials are necessary to obtain more robust and accurate datasets.
4.3. On the Development of Novel Treatments: Summary Model of Key Components
Alternative dormancy-release treatments typically consist of the following three ingredients, which should be considered in the development of future treatments (Figure 6):
- A source of hypoxia, such as an oil-based adjuvant or mineral/vegetable oil
- Inclusion of additional supplementation of nitrites and/or nitrates such as potassium nitrite (KNO2) or KNO3
- Amino acids
Together, the abovementioned components should ideally induce dormancy-release and aid in cell recovery in the following ways. Oxidative stress induced by the candidate alternative PB-based agrochemical treatments is likely attributed lead to the formation of respiratory stress and NO from nitrites and nitrates enzymatically via the activation of nitrogen nitrite and nitrite reductases under hypoxic conditions. NO is a type of ROS, which both inhibits cytochrome oxidase and has been shown to inhibit CAT in a reversible manner, which further contributes to the spike in ROS [41,42]. Hypoxic conditions are artificially created by commercial rest breaking oils or oil-based adjuvants. Additionally respiratory stress, which is induced by rest breaking oils leads to ROS accumulation, such as H2O2, which is a natural trigger for various processes involved in dormancy-release [26]. Amino acids aid in stress response and recovery:
Changes in amino acid profiles are associated in plants which are under major oxidative stress, and the exogenous application of several amino acids are known to improve antioxidant defence [44]. Protein degradation, which occurs because of oxidative stress, results in the release of amino acids, which are used by the plant to biosynthesise various protective metabolites such as osmolytes, secondary metabolites, structural components needed for cell wall repair, hormones, polyphenols, cyanogenic glycosides, glucosinolates, and acyl sugars.
5. Conclusions
Rising global winter temperatures due to climate change are an increasing threat to agricultural practices globally, continuously introducing abiotic stress at a pace too rapid to allow for evolution and adaption [69]. Average winter temperatures have increased annually and are predicted to continue rising all over the world, including grape-producing regions in South Africa [8,11,70,71]. This is in accordance with the lower CUA accumulated in 2022 than in 2021 in the experimental site, which was in the Berg River table grape-producing region of South Africa (Figure S4). Despite sufficient CUA in this region, Dormex® is typically used as a preventative measure. Its use may soon become unavoidable, which is already the case for most of the remaining South African table grape-producing regions, including the Berg-, Olifants- and Orange River regions, and the Northern Regions. This emphasizes the importance of research results aimed at discovering an alternative product, such as those obtained in the current study.
The results of the current study highlighted several key elements which should be included in future candidate HC replacement treatments. The results obtained during the 2021 forced bud-break assays indicated that treatment of dormant grapevine compound buds with NO, H2O2, and hypoxia do indeed trigger dormancy-release to a certain extent, supporting the molecular models proposed for HC action [24,25]. Furthermore, during the 2022 forced bud-break assays, it was shown that NO, H2O2, and hypoxia, in combination with PBs, may potentially replace HC, provided the correct application strategy is used. However, this needs to be confirmed in future experiments. The relevance of amino acid and nitrogen-salt-based agrochemicals that induce oxidative stress was also confirmed by the results recorded after SNH treatment. These products might be best applied with an adjuvant and/or dormancy oil to enhance uptake and performance in future studies [51,59,72]. Finally, in 2022, the complexity of field trials compared to controlled glasshouse conditions and the need for optimal timing of HC application were emphasized and require further exploration.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Information from the South African Table Grape Industry; Figure S2: Example of growth room temperatures during glasshouse assays; Figure S3: Visual representation and definitions of four parameters used to assess bud-break efficacy; Figure S4: Infruitec units accumulated in the V. vinifera CS block on Windmeul farm. Table S1: Summary of experiment dates and CU information for experiments evaluating the effects of HC 3%, MIX2, SNH, SNL, PBXH, PBXL, and dH2O, in in V. vinifera Crimson Seedless (CS) buds after, 100, 200 and 250 CUA.
Author Contributions
Conceptualization, P.N.H., T.N.K. and J.P.M.; performing all experiments, N.C.V.; writing—original draft preparation, N.C.V.; writing—review and editing, P.N.H., E.A. and J.P.M.; supervision, P.N.H., E.A., and J.P.M.; funding acquisition, T.N.K. and J.P.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially funded to T.N.K. (and J.P.M.) by the Winfield United (Villa Crop) Chair in Plant Health at Stellenbosch University, and Stellenbosch University. The APC was funded by Stellenbosch University. The funders were not involved in the design of the study; in the collection, analysis and interpretation of data; in the writing of the paper; or in the decision to submit the article for publication.
Data Availability Statement
Data are available upon request from the corresponding author.
Acknowledgements
Melané Vivier (South African Grape and Wine Research Institute and Department of Viticulture and Oenology at Stellenbosch University) is thanked for the provision of the glasshouse facilities. Martin Kidd of the Centre of Statistical Consultation (Stellenbosch University) is thanked for assistance with statistical analyses. Hoekstra Fruit Farms in Paarl is thanked for the use of their vineyards. Curativo is thanked for assistance with the small-scale trial.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Or, E.; Vilozny, I.; Eyal, Y.; Ogrodovitch, A. The Transduction of the Signal for Grape Bud Dormancy Breaking Induced by Hydrogen Cyanamide May Involve the SNF-like Protein Kinase GDBRPK. Plant Mol Biol 2000, 43, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Forney, C.F.; Bondada, B. Renewal of Vascular Connections between Grapevine Buds and Canes during Bud Break. Scientia Horticulturae 2018, 233, 331–338. [Google Scholar] [CrossRef]
- Dokoozlian, N.K. Chilling Temperature and Duration Interact on the Budbreak of `Perlette’ Grapevine Cuttings. HortScience 1999, 34, 1–3. [Google Scholar] [CrossRef]
- Avenant, E.; Avenant, J.H. Chill Unit Accumulation and Necessity of Rest Breaking Agents in South African Table Grape Production Regions. BIO Web of Conferences 2014, 3, 01017. [Google Scholar] [CrossRef]
- Richardson, E.A.; Seeley, S.D.; Walker, D.R. A Model for Estimating the Completion of Rest for ‘Redhaven’ and ‘Elberta’ Peach Trees1. HortScience 1974, 9, 331–332. [Google Scholar] [CrossRef]
- Fishman, S.; Erez, A.; Couvillon, G.A. The Temperature Dependence of Dormancy Breaking in Plants: Computer Simulation of Processes Studied under Controlled Temperatures. Journal of Theoretical Biology 1987, 126, 309–321. [Google Scholar] [CrossRef]
- Fishman, S.; Erez, A.; Couvillon, G.A. The Temperature Dependence of Dormancy Breaking in Plants: Mathematical Analysis of a Two-Step Model Involving a Cooperative Transition. Journal of Theoretical Biology 1987, 124, 473–483. [Google Scholar] [CrossRef]
- Ogundeji, A.A.; Jordaan, H. A Simulation Study on the Effect of Climate Change on Crop Water Use and Chill Unit Accumulation. South African Journal of Science 2017, 113, 7–7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Taylor, C. The Dynamic Model Provides the Best Description of the Chill Process on ‘Sirora’ Pistachio Trees in Australia. HortScience 2011, 46, 420–425. [Google Scholar] [CrossRef]
- SATI Production Overview Statistical Booklet; South African Table Grape Industry, 2018;
- Tharaga, P.C. Impacts of Climate Change on Accumulated Chill Units at Selected Fruit Production Sites in South Africa. 2014.
- Sheshadri, S.H.; Sudhir, U.; Kumar, S.; Kempegowda, P. DORMEX®-Hydrogen Cyanamide Poisoning. Journal of Emergencies, Trauma, and Shock 2011, 4, 435. [Google Scholar] [CrossRef] [PubMed]
- SATI Statistics of Table Grapes in South Africa; South African Table Grape Industry, 2022;
- Nir, G.; Shulman, Y.; Fanberstein, L.; Lavee, S. Changes in the Activity of Catalase (EC 1.11.1.6) in Relation to the Dormancy of Grapevine (Vitis Vinifera L.) Buds 1 2. Plant Physiology 1986, 81, 1140–1142. [Google Scholar] [CrossRef] [PubMed]
- Petri, J.L.; Stuker, H. Effect of Mineral Oil and Hydrogen Cyanamide Concentrations on Apple Dormancy, Cv. Gala. Acta Hortic. 1995, 161–168. [Google Scholar] [CrossRef]
- Pérez, F.J.; Lira, W. Possible Role of Catalase in Post-Dormancy Bud Break in Grapevines. Journal of Plant Physiology 2005, 3, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Ophir, R.; Pang, X.; Halaly, T.; Venkateswari, J.; Lavee, S.; Galbraith, D.; Or, E. Gene-Expression Profiling of Grape Bud Response to Two Alternative Dormancy-Release Stimuli Expose Possible Links between Impaired Mitochondrial Activity, Hypoxia, Ethylene-ABA Interplay and Cell Enlargement. Plant Mol Biol 2009, 71, 403. [Google Scholar] [CrossRef] [PubMed]
- Pérez, F.J.; Vergara, R.; Or, E. On the Mechanism of Dormancy Release in Grapevine Buds: A Comparative Study between Hydrogen Cyanamide and Sodium Azide. Plant Growth Regul 2009, 59, 145–152. [Google Scholar] [CrossRef]
- Petri, J.L.; Berenhauser Leite, G.; Putti, G.L. Apple Tree Budbreak Promoters in Mild Winter Conditions. Acta Hortic. 2008, 291–296. [Google Scholar] [CrossRef]
- Pérez, F.J.; Kühn, N.; Vergara, R. Expression Analysis of Phytochromes A, B and Floral Integrator Genes during the Entry and Exit of Grapevine-Buds from Endodormancy. Journal of Plant Physiology 2011, 168, 1659–1666. [Google Scholar] [CrossRef] [PubMed]
- Vergara, R.; Rubio, S.; Pérez, F.J. Hypoxia and Hydrogen Cyanamide Induce Bud-Break and up-Regulate Hypoxic Responsive Genes (HRG) and VvFT in Grapevine-Buds. Plant Mol Biol 2012, 79, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Vergara, R.; Parada, F.; Rubio, S.; Pérez, F.J. Hypoxia Induces H2O2 Production and Activates Antioxidant Defence System in Grapevine Buds through Mediation of H2O2 and Ethylene. J Exp Bot 2012, 63, 4123–4131. [Google Scholar] [CrossRef] [PubMed]
- Petri, J.L.; Leite, G.B.; Couto, M.; Gabardo, G.C.; Haverroth, F.J. Chemical Induction of Budbreak: New Generation Products to Replace Hydrogen Cyanamide. Acta Hortic. 2014, 159–166. [Google Scholar] [CrossRef]
- Sudawan, B.; Chang, C.-S.; Chao, H.; Ku, M.S.B.; Yen, Y. Hydrogen Cyanamide Breaks Grapevine Bud Dormancy in the Summer through Transient Activation of Gene Expression and Accumulation of Reactive Oxygen and Nitrogen Species. BMC Plant Biology 2016, 16, 202. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Huang, X.; Shen, Y.; Shen, T.; Zhang, H.; Lin, L.; Wang, J.; Deng, Q.; Lyu, X.; Xia, H. Hydrogen Cyanamide Induces Grape Bud Endodormancy Release through Carbohydrate Metabolism and Plant Hormone Signaling. BMC Genomics 2019, 20, 1034. [Google Scholar] [CrossRef] [PubMed]
- Beauvieux, R.; Wenden, B.; Dirlewanger, E. Bud Dormancy in Perennial Fruit Tree Species: A Pivotal Role for Oxidative Cues. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Erez, A.; Yablowitz, Z.; Aronovitz, A.; Hadar, A. Dormancy Breaking Chemicals; Efficiency with Reduced Phytotoxicity. Acta Hortic. 2008, 105–112. [Google Scholar] [CrossRef]
- Segantini, D.M.; Leonel, S.; Ripardo, A.K. da S.; Tecchio, M.A.; Souza, M.E. de Breaking Dormancy of “Tupy” Blackberry in Subtropical Conditions. American Journal of Plant Sciences 2015, 6, 1760–1767. [Google Scholar] [CrossRef]
- Petri, J.L.; Leite, G.B.; Couto, M.; Francescatto, P. A New Product to Induce Apple Bud Break and Flowering – Syncron ®. Acta Hortic. 2016, 103–110. [Google Scholar] [CrossRef]
- Allderman, L.; Steyn, W.J.; Louw, E.D. Alternative Rest Breaking Agents and Vigour Enhancers Tested on “Fuji” Apple Shoots under Mild Conditions. In Proceedings of the Acta Horticulturae; International Society for Horticultural Science (ISHS), Leuven, Belgium, September 30 2022; pp. 145–152. [Google Scholar]
- Ardiles, M.; Ayala, M. An Alternative Dormancy-Breaking Agent to Hydrogen Cyanamide for Sweet Cherry (Prunus Avium L.) under Low Chilling Accumulation Conditions in the Central Valley of Chile. In Proceedings of the Acta Horticulturae, International Society for Horticultural Science (ISHS), Leuven, Belgium, June 1 2017; pp. 423–430. [Google Scholar]
- Tavares, D.F.; Rodrigues, S.I.A.; Oliveira, C.M.M.S. Biostimulants to Promote Budbreak in Kiwifruit (Actinidia Chinensis Var. Deliciosa ‘Hayward’). Acta Hortic. 2018, 367–372. [Google Scholar] [CrossRef]
- Fowler, J.G.; Margoti, G.; Biasi, L.A. Sprouting, Phenology, and Maturation of the Italian Grapevine ‘Fiano’ in Campo Largo, PR, Brazil. Semina: Ciências Agrárias 2020, 41, 83–94. [Google Scholar] [CrossRef]
- Ferreira, R.B.; Leonel, S.; Souza, J.M.A.; Silva, M. de S.; Ferraz, R.A.; Martins, R.C.; Silva, M.S.C. da Peaches Phenology and Production Submitted to Foliar Nitrogen Fertilizer and Calcium Nitrate. Bioscience Journal 2019, 35, 752–762. [Google Scholar] [CrossRef]
- Guillamón, J.G.; Dicenta, F.; Sánchez-Pérez, R. Advancing Endodormancy Release in Temperate Fruit Trees Using Agrochemical Treatments. Front. Plant Sci. 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Broadley, M.R. Calcium in Plants. Annals of Botany 2003, 92, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Foreman, J.; Demidchik, V.; Bothwell, J.H.F.; Mylona, P.; Miedema, H.; Torres, M.A.; Linstead, P.; Costa, S.; Brownlee, C.; Jones, J.D.G.; et al. Reactive Oxygen Species Produced by NADPH Oxidase Regulate Plant Cell Growth. Nature 2003, 422, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Wendehenne, D.; Hancock, J.T. New Frontiers in Nitric Oxide Biology in Plant. Plant Science 2011, 181, 507–508. [Google Scholar] [CrossRef] [PubMed]
- Šírová, J.; Sedlářová, M.; Piterková, J.; Luhová, L.; Petřivalský, M. The Role of Nitric Oxide in the Germination of Plant Seeds and Pollen. Plant Science 2011, 181, 560–572. [Google Scholar] [CrossRef]
- Niu, L.; Li, B.; Liao, W.; Zhu, Y.; Wang, M.; Jin, X.; Xu, Q. Effect of Nitric Oxide on Dormancy Release in Bulbs of Oriental Lily (Lilium Orientalis) ‘Siberia. ’ The Journal of Horticultural Science and Biotechnology 2015, 90, 594–598. [Google Scholar] [CrossRef]
- Brown, G.C. Reversible Binding and Inhibition of Catalase by Nitric Oxide. European Journal of Biochemistry 1995, 232, 188–191. [Google Scholar] [CrossRef]
- Brown, G.C.; Borutaite, V. Nitric Oxide Inhibition of Mitochondrial Respiration and Its Role in Cell Death. Free Radical Biology and Medicine 2002, 33, 1440–1450. [Google Scholar] [CrossRef] [PubMed]
- Asada, K. Production and Scavenging of Reactive Oxygen Species in Chloroplasts and Their Functions. Plant Physiology 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Savchenko, T.; Tikhonov, K. Oxidative Stress-Induced Alteration of Plant Central Metabolism. Life 2021, 11, 304. [Google Scholar] [CrossRef] [PubMed]
- Or, E.; Vilozny, I.; Fennell, A.; Eyal, Y.; Ogrodovitch, A. Dormancy in Grape Buds: Isolation and Characterization of Catalase cDNA and Analysis of Its Expression Following Chemical Induction of Bud Dormancy Release. Plant Science 2002, 162, 121–130. [Google Scholar] [CrossRef]
- Dry, P.R.; Coombe, B.G.; Anderson, C.J. Viticulture. Volume 1, Resources; 2nd ed., repr. with alterations.; Winetitles: Ashford, S. Aust., 2005; ISBN 978-0-9756850-0-6.
- Botelho, R.V.; Sato, A.J.; Maia, A.J.; Marchi, T.; Oliari, I.C.R.; Rombolà, A.D. Mineral and Vegetable Oils as Effective Dormancy Release Agents for Sustainable Viticulture in a Sub-Tropical Region. The Journal of Horticultural Science and Biotechnology 2016, 91, 370–377. [Google Scholar] [CrossRef]
- Egbichi, I.; Keyster, M.; Ludidi, N. Effect of Exogenous Application of Nitric Oxide on Salt Stress Responses of Soybean. South African Journal of Botany 2014, 90, 131–136. [Google Scholar] [CrossRef]
- Pholo, M.; Coetzee, B.; Maree, H.J.; Young, P.R.; Lloyd, J.R.; Kossmann, J.; Hills, P.N. Cell Division and Turgor Mediate Enhanced Plant Growth in Arabidopsis Plants Treated with the Bacterial Signalling Molecule Lumichrome. Planta 2018, 248, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Loubser, J.; Hills, P.N. Analysis of the Molecular and Physiological Effects Following Treatment with BC204 in Arabidopsis Thaliana and Solanum Lycopersicum. PhD thesis, Stellenbosch University: South Africa, 2020.
- Sagredo, K.X.; Theron, K.I.; Cook, N.C. Effect of Mineral Oil and Hydrogen Cyanamide Concentration on Dormancy Breaking in ‘Golden Delicious’ Apple Trees. South African Journal of Plant and Soil 2005, 22, 251–256. [Google Scholar] [CrossRef]
- Dubois, J.-M.; Ouanounou, G.; Rouzaire-Dubois, B. The Boltzmann Equation in Molecular Biology. Progress in Biophysics and Molecular Biology 2009, 99, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Vetter, T.R.; Mascha, E.J. Unadjusted Bivariate Two-Group Comparisons: When Simpler Is Better. Anesthesia & Analgesia 2018, 126, 338. [Google Scholar] [CrossRef]
- Dokoozlian, N.K.; Williams, L.E.; Neja, R.A. Chilling Exposure and Hydrogen Cyanamide Interact in Breaking Dormancy of Grape Buds. HortScience 1995, 30, 1244–1247. [Google Scholar] [CrossRef]
- Dokoozlian, N.K. Quantifying the Chilling Status of Grapevines and the Response to Dormancy-Breaking Chemicals in the Coachella Valley of California.; HortScience, 1998; Vol. 33, p. 510.
- Hernández, G.; Craig, R.L. Effects of Alternatives to Hydrogen Cyanamide on Commercial Kiwifruit Production. Acta Hortic. 2011, 357–363. [Google Scholar] [CrossRef]
- Pereira, N.; Oliveira, C.M.; Mota, M.; Sousa, R.M. Evaluation of Five Dormancy Breaking Agents to Induce Synchronized Flowering in “Rocha” Pear. Acta Hortic. 2011, 423–428. [Google Scholar] [CrossRef]
- Seif El-Yazal, M.A.; Seif El-Yazal, S.A.; Rady, M.M. Exogenous Dormancy-Breaking Substances Positively Change Endogenous Phytohormones and Amino Acids during Dormancy Release in ‘Anna’ Apple Trees. Plant Growth Regul 2014, 72, 211–220. [Google Scholar] [CrossRef]
- Makarem, M.M.; Taha, N.M.; Shakweer, N.H. Studies the Effect of Alternative Dormancy Breaking Agents on “Le-Cont” Pear Cultivar. Journal of Plant Production 2016, 7, 943–950. [Google Scholar] [CrossRef]
- Botelho, R.V.; Biasi, L.A.; Maia, A.J.; Nedilha, L.C.B.M.; Viencz, T. Dormancy Release in Asian ‘Hosui’ Pear Trees with the Use of Vegetable and Mineral Oils. Acta Hortic. 2021, 317–324. [Google Scholar] [CrossRef]
- Pérez, F.J.; Noriega, X.; Rubio, S. Hydrogen Peroxide Increases during Endodormancy and Decreases during Budbreak in Grapevine (Vitis Vinifera L.) Buds. Antioxidants 2021, 10, 873. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, H.; Sugiura, T.; Sugiura, H. Effect of Hydrogen Peroxide on Breaking Endodormancy in Flower Buds of Japanese Pear (Pyrus Pyrifolia Nakai). Journal of the Japanese Society for Horticultural Science 2005, 74, 255–257. [Google Scholar] [CrossRef]
- Pérez, F.J.; Vergara, R.; Rubio, S. H2O2 Is Involved in the Dormancy-Breaking Effect of Hydrogen Cyanamide in Grapevine Buds. Plant Growth Regul 2008, 55, 149–155. [Google Scholar] [CrossRef]
- Gholizadeh, J.; Sadeghipour, H.R.; Abdolzadeh, A.; Hemmati, K.; Hassani, D.; Vahdati, K. Redox Rather than Carbohydrate Metabolism Differentiates Endodormant Lateral Buds in Walnut Cultivars with Contrasting Chilling Requirements. Scientia Horticulturae 2017, 225, 29–37. [Google Scholar] [CrossRef]
- Iwasaki, K. Effects of Bud Scale Removal, Calcium Cyanamide, GA3, and Ethephon on Bud Break of ‘Muscat of Alexandria’ Grape (Vitis Vinifera L.). Journal of the Japanese Society for Horticultural Science 1980, 48, 395–398. [Google Scholar] [CrossRef]
- Or, E.; Nir, G.; Vilozny, I. Timing of Hydrogen Cyanamide Application to Grapevine Buds. VITIS - Journal of Grapevine Research 1999, 38, 1–1. [Google Scholar] [CrossRef]
- Hawerroth, F.J.; Petri, J.L.; Leite, G.B. Erger and Calcium Nitrate Concentration for Budbreak Induction in Apple Trees. Acta Hortic. 2010, 239–244. [Google Scholar] [CrossRef]
- Rodrigo, J.; Espada, J.L.; Bernad, D.; Martin, E.; Herrero, M. Effects of Syncron ® and Nitroactive ® on Flowering and Ripening in Sweet Cherry. Acta Hortic. 2017, 389–394. [Google Scholar] [CrossRef]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of Climate Change on Crops Adaptation and Strategies to Tackle Its Outcome: A Review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; Pérez-López, D.; Sánchez, E.; Centeno, A.; Gómara, I.; Dosio, A.; Ruiz-Ramos, M. Chilling Accumulation in Fruit Trees in Spain under Climate Change. Natural Hazards and Earth System Sciences 2019, 19, 1087–1103. [Google Scholar] [CrossRef]
- Theron, H.; Hunter, J.J. Mitigation and Adaptation Practices to the Impact of Climate Change on Wine Grape Production, with Special Reference to the South African Context. South African Journal of Enology and Viticulture 2022, 43, 10–25. [Google Scholar] [CrossRef]
- Hernández, G.; Craig, R.L. Effects of Alternatives to Hydrogen Cyanamide on Commercial ‘Hayward’ Kiwifruit Production. Acta Hortic. 2016, 131–138. [Google Scholar] [CrossRef]
Figure 1.
Photos taken during glasshouse forced bud-break assays of V. vinifera Crimson Seedless (CS) plant material: (a) Gable trellis system of CS vineyard block on Windmeul Farm, Hoekstra Fruit, Paarl. (b) Single node cuttings (SNCs) placed upright in trays filled with tap water prior to treatment applications. (c) SNCs that are positioned in styrofoam supports and floated on water in plastic trays at 22°C, with a 16/8 h light/dark photoperiod. (d) Green tissue and leaf tips visible underneath the bud scales, specifically E-L stage 3, and E-L stage 4 (arrow pointing towards the greening bud)[1,46].
Figure 1.
Photos taken during glasshouse forced bud-break assays of V. vinifera Crimson Seedless (CS) plant material: (a) Gable trellis system of CS vineyard block on Windmeul Farm, Hoekstra Fruit, Paarl. (b) Single node cuttings (SNCs) placed upright in trays filled with tap water prior to treatment applications. (c) SNCs that are positioned in styrofoam supports and floated on water in plastic trays at 22°C, with a 16/8 h light/dark photoperiod. (d) Green tissue and leaf tips visible underneath the bud scales, specifically E-L stage 3, and E-L stage 4 (arrow pointing towards the greening bud)[1,46].

Figure 2.
Small scale field trial experimental design: (a) V. vinifera CS target cane for small scale field trial, bound to the bottom wire of the gable trellis system. Ten target compound buds were clearly marked for analysis using white acrylic paint, starting from the head of the vine. (b) Experimental rows of V. vinifera CS block of Windmeul farm, Paarl. Two plant biostimulants, SNH and PBXH were evaluated against 3% Dormex® (HC 3%), the positive control, and dH2O, the negative control in the months of September and October 2022. One biological repeat of each replicate consists of the two middle vines of the four planted between line posts. Four target canes were selected per target vine, of which the first ten target compound buds from the base of the vine were marked.
Figure 2.
Small scale field trial experimental design: (a) V. vinifera CS target cane for small scale field trial, bound to the bottom wire of the gable trellis system. Ten target compound buds were clearly marked for analysis using white acrylic paint, starting from the head of the vine. (b) Experimental rows of V. vinifera CS block of Windmeul farm, Paarl. Two plant biostimulants, SNH and PBXH were evaluated against 3% Dormex® (HC 3%), the positive control, and dH2O, the negative control in the months of September and October 2022. One biological repeat of each replicate consists of the two middle vines of the four planted between line posts. Four target canes were selected per target vine, of which the first ten target compound buds from the base of the vine were marked.
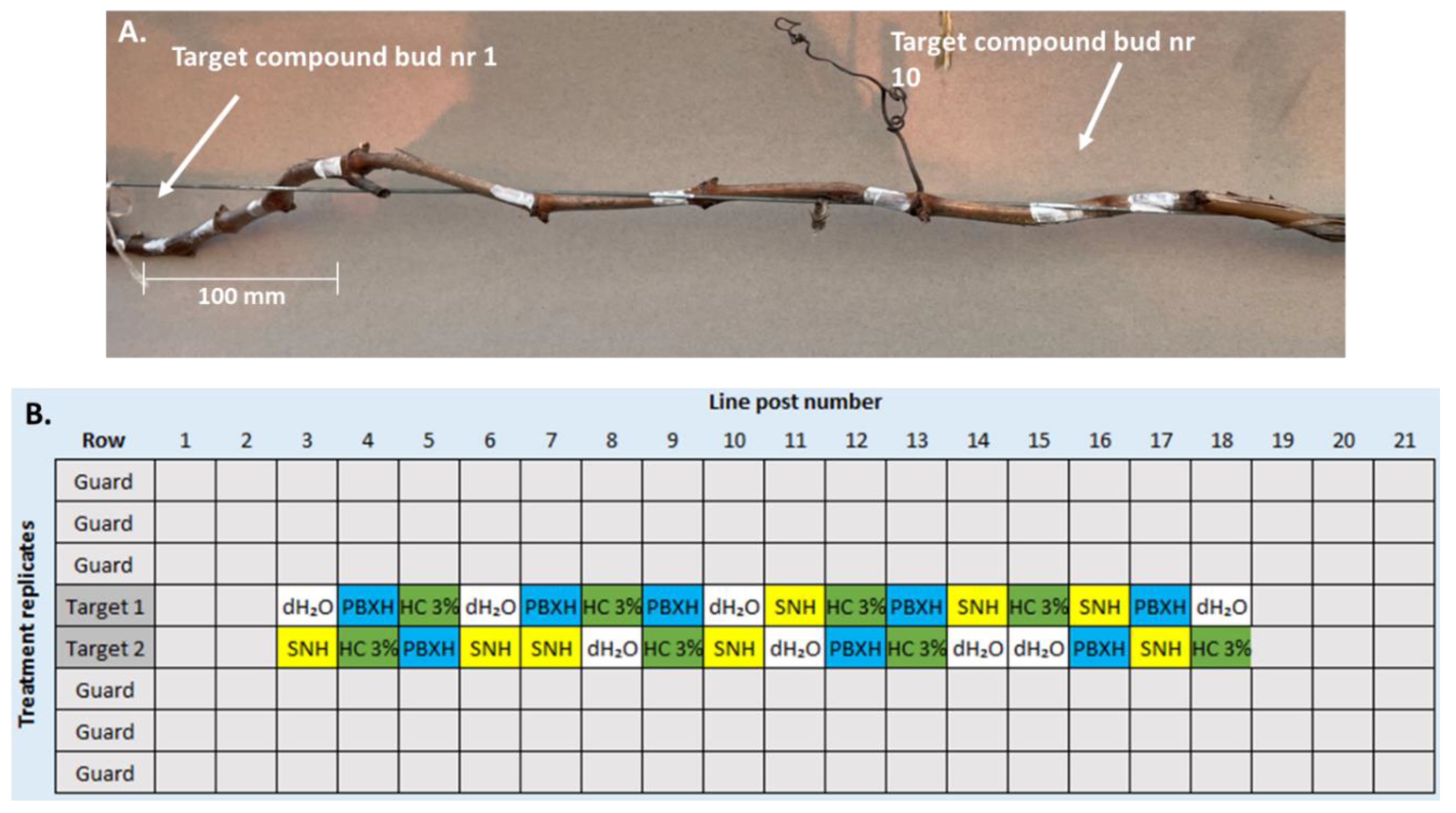
Figure 3.
Effects of HC 3%, dH2O, BC204 0.05%, Lum 5 nM, H2O2 1%, Eth 1000 ppm, BB 3%, SNO 10 μM, & NO2 10 μM on the bud-break of V. vinifera CS SNCs, after (a) 100, (b) 200, and (c) 400 CUA (30 biological replicates/treatment).
Figure 3.
Effects of HC 3%, dH2O, BC204 0.05%, Lum 5 nM, H2O2 1%, Eth 1000 ppm, BB 3%, SNO 10 μM, & NO2 10 μM on the bud-break of V. vinifera CS SNCs, after (a) 100, (b) 200, and (c) 400 CUA (30 biological replicates/treatment).
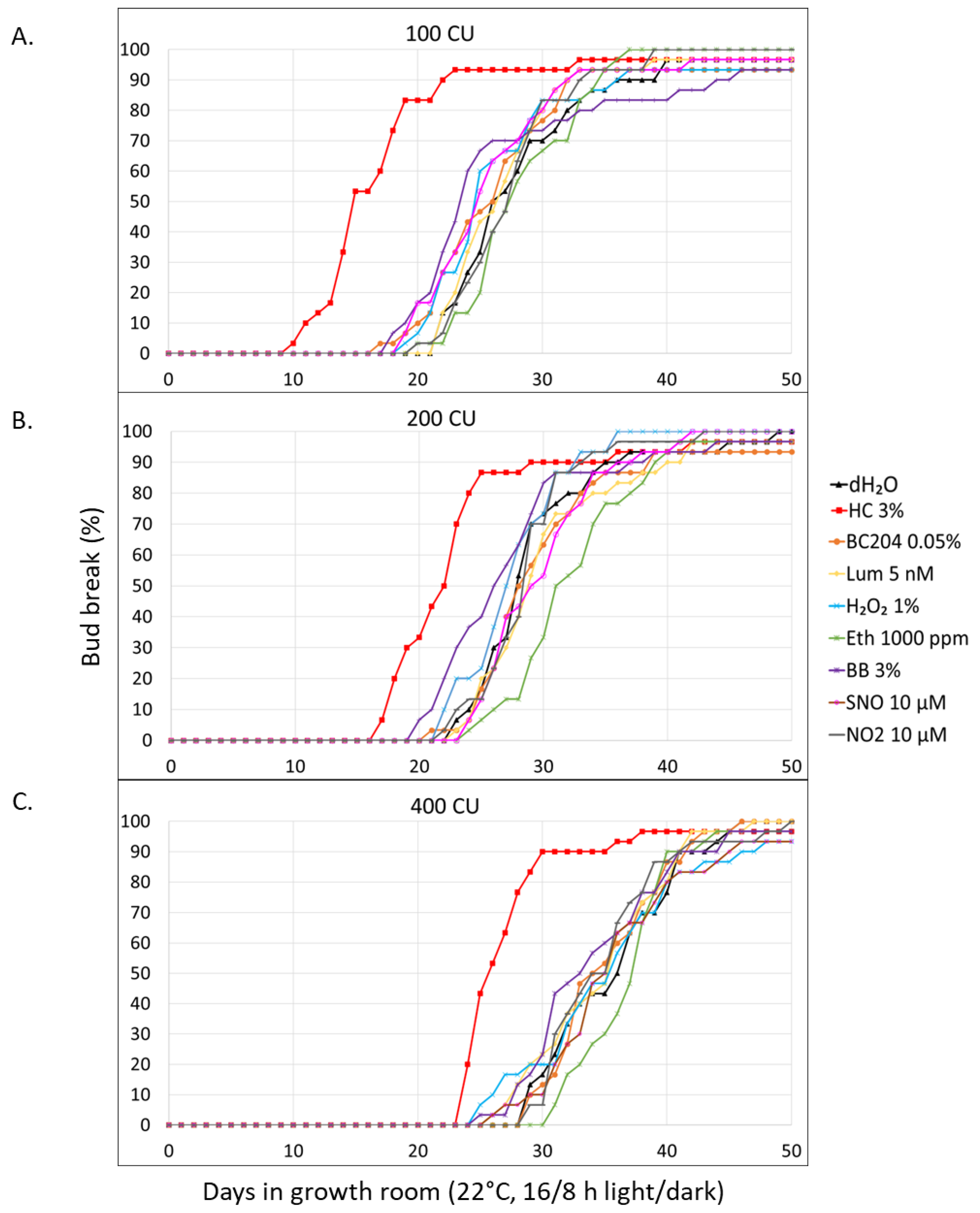
Figure 4.
Effects of HC 3%, dH2O, SNH, SNL, PBXH, PBXL & MIX2 on the bud-break of V. vinifera CS SNCs, after (a) 100, (b) 200, and (c) 250 CUA (45 biological replicates/treatment).
Figure 4.
Effects of HC 3%, dH2O, SNH, SNL, PBXH, PBXL & MIX2 on the bud-break of V. vinifera CS SNCs, after (a) 100, (b) 200, and (c) 250 CUA (45 biological replicates/treatment).
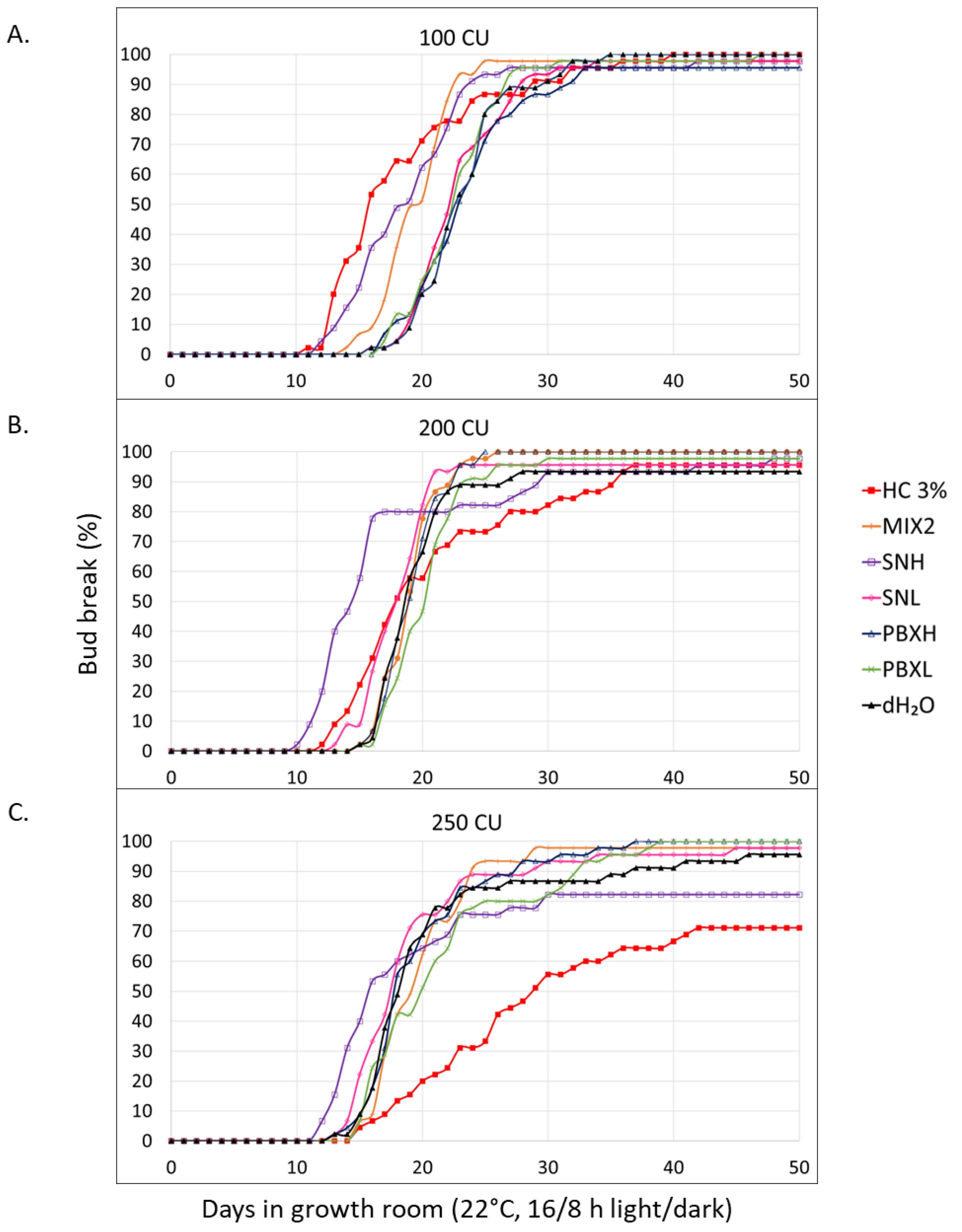
Figure 5.
Effects of HC 3%, dH2O, SNH, & PBXH on the bud-break of V. vinifera CS canes during the small-scale field trial.
Figure 5.
Effects of HC 3%, dH2O, SNH, & PBXH on the bud-break of V. vinifera CS canes during the small-scale field trial.
Figure 6.
The components typically present in alternative dormancy-release treatments, which are agrochemical and biochemical agent based. Nitric oxide donors, and hypoxia create oxidative stress, which act as central signaling mechanisms for dormancy-release. Amino acid supplementation is used by the plant to produce compounds which aid in protection against uncontrolled reactive oxygen accumulation and recovery.
Figure 6.
The components typically present in alternative dormancy-release treatments, which are agrochemical and biochemical agent based. Nitric oxide donors, and hypoxia create oxidative stress, which act as central signaling mechanisms for dormancy-release. Amino acid supplementation is used by the plant to produce compounds which aid in protection against uncontrolled reactive oxygen accumulation and recovery.
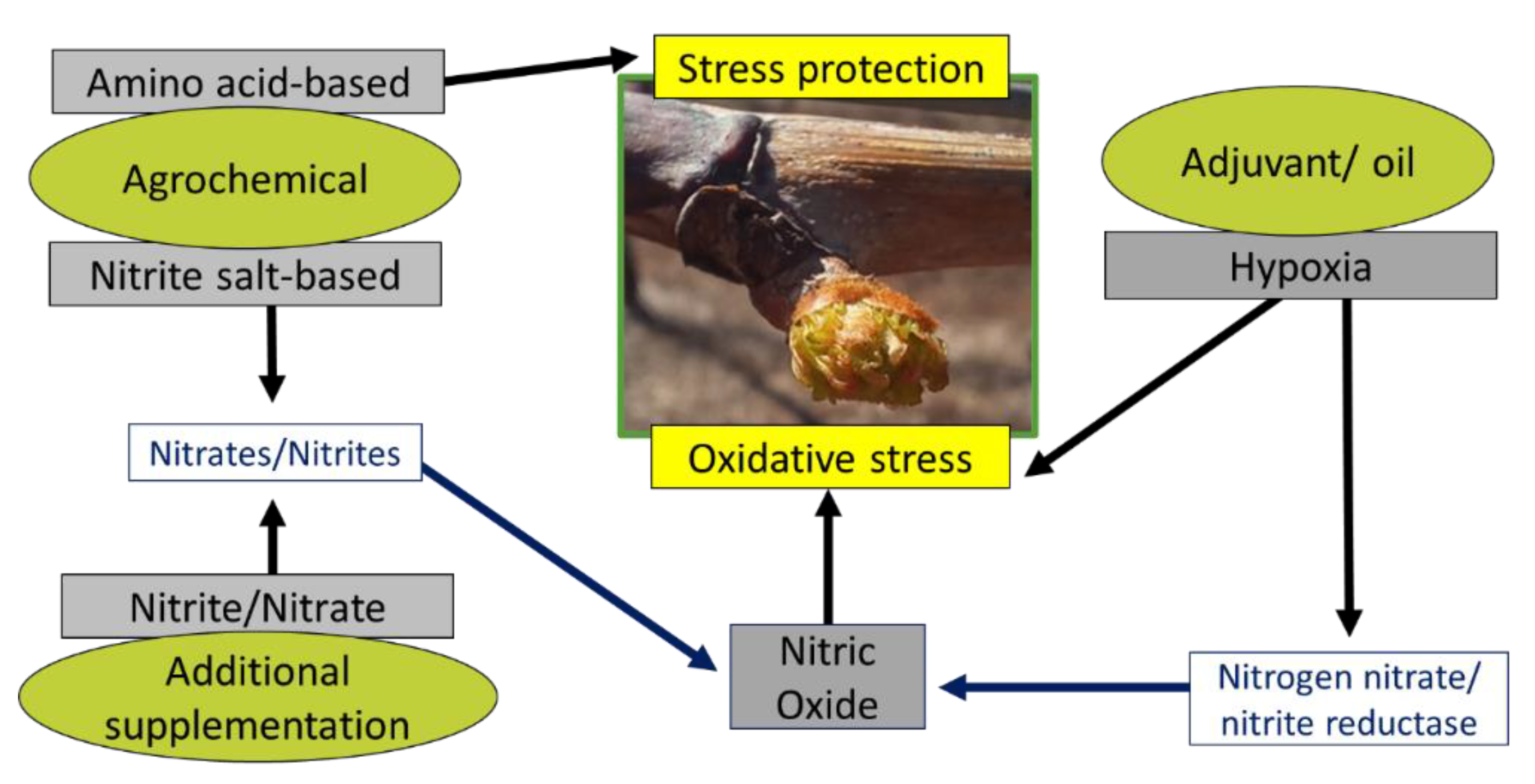

Table 1.
Biochemical agents and plant biostimulants (PBs) tested on V. vinifera CS grapevine SNCs in 2021 during forced bud-break assays. Each treatment was assigned a code name.
Table 1.
Biochemical agents and plant biostimulants (PBs) tested on V. vinifera CS grapevine SNCs in 2021 during forced bud-break assays. Each treatment was assigned a code name.
| Treatment | Relevance to dormancy-release molecular models | Application concentration |
Treatment code name |
|---|---|---|---|
| Dormex® | Positive control | 3% v/v [47] | HC 3% |
| Distilled water | Negative control | NA | dH2O |
| H2O2 | ROS | 1% v/v [22] | H2O2 1% |
| S-Nitrosoglutathione | NO donor | 10 μM [48] | SNO 10 μM |
| Diethylamine NONOate sodium salt hydrate | NO donor | 10 μM [48] | NO2 10 μM |
| Lumichrome | Riboflavin derivative | 5 nM [49] | Lum 5 nM |
| BC204 (Commercial PB) | Citrus-based plant extract | 0.05% v/v [50] | BC204 0.05% |
| BUDBREAK® mineral oil | Hypoxia | 3% v/v [51] | BB 3% |
| Ethephon | Ethylene supplement | 0.206% v/v [22] |
Table 2.
Biochemical agents and PBs tested on V. vinifera CS grapevine SNCs in 2021 during forced bud-break assays. Each treatment was assigned a code name.
Table 2.
Biochemical agents and PBs tested on V. vinifera CS grapevine SNCs in 2021 during forced bud-break assays. Each treatment was assigned a code name.
| Treatment | Relevance to dormancy-release molecular models |
Application concentration | Treatment code name |
|---|---|---|---|
| Dormex® | Positive control | 3% v/v | HC 3% |
| Distilled water | Negative control | NA | dH2O |
| SN (Commercial PB) | Amino acids formulations (SBB-01), synergistic effect provided by a calcium supplement (NDY-01) | 2% v/v SBB-01 and 20% v/v NDY-01 (recommended by manufacturer) | SNH |
| SN | 0.2% v/v SBB-01 and 2% v/v NDY-01 | SNL | |
| PBX (Commercial PB) | Amino acid formulations which alter metabolism in plants | 1.5% v/v (recommended by manufacturer) | PBXH |
| PBX | 0.15% v/v | PBXL | |
| Combination of selected biochemical agents (refer to Table 1) | Refer to Table 1 | 1% v/v hydrogen peroxide, 10 μM S-nitrosoglutathione, 10 μM diethylamine NONOate sodium salt hydrate, 5nM lumichrome, 0.05% v/v BC204 and 3% v/v BUDBREAK® mineral oil | MIX2 |
Table 3.
Respective coefficient estimates measured after HC 3%, dH2O, BC204 0.05%, Lum 5 nM, H2O2 1%, Eth 1000 ppm, BB 3%, SNO 10 μM, & NO2 10 μM application on V. vinifera CS SNCs after 100, 200, and 400 CUA (30 biological replicates/treatment). Treatments have been assigned letters which indicate significant differences at 5% (p<0.05) amongst the coefficient estimates. These letters only apply to individual experiments and parameters.
Table 3.
Respective coefficient estimates measured after HC 3%, dH2O, BC204 0.05%, Lum 5 nM, H2O2 1%, Eth 1000 ppm, BB 3%, SNO 10 μM, & NO2 10 μM application on V. vinifera CS SNCs after 100, 200, and 400 CUA (30 biological replicates/treatment). Treatments have been assigned letters which indicate significant differences at 5% (p<0.05) amongst the coefficient estimates. These letters only apply to individual experiments and parameters.
| Coefficient estimate | ||||
|---|---|---|---|---|
| Bud-break parameter: definition | Treatment | 100 | 200 | 400 |
| Onset: Number of days to first bud-break | HC 3% | 9.86f | 16.12f | 25.41abcd |
| dH2O | 20.07a | 22.17b | 27.08b | |
| BC204 0.05% | 18.08c | 22.49bc | 27.58b | |
| Lum 5 nM | 20.47ab | 22.34bc | 25.91c | |
| H2O2 1% | 18.80d | 21.07d | 24.48d | |
| Eth 1000 ppm | 20.93b | 24.49a | 30.81a | |
| BB 3% | 16.48e | 19.16e | 25.17cd | |
| SNO 10 μM | 17.81c | 22.74bc | 27.36b | |
| NO2 10 μM | 20.83b | 22.86c | 27.85b | |
| Rate: Slope between onset and final bud-break % | HC 3% | -0.48a | -0.48ab | -0.71a |
| dH2O | -0.37b | -0.46ac | -0.31bc | |
| BC204 0.05% | -0.34b | -0.42dc | -0.36de | |
| Lum 5 nM | -0.45a | -0.39d | -0.30bc | |
| H2O2 1% | -0.42a | -0.45ac | -0.25f | |
| Eth 1000 ppm | -0.37b | -0.38d | -0.49g | |
| BB 3% | -0.34b | -0.38d | -0.30b | |
| SNO 10 μM | -0.36b | -0.38d | -0.33dc | |
| NO2 10 μM | -0.43a | -0.54b | -0.39e | |
| EC50: Number of days to 50 % of final bud-break %. | HC 3% | 15.51g | 21.32h | 25.86f |
| dH2O | 26.82b | 27.99e | 35.39b | |
| BC204 0.05% | 25.43d | 28.37c | 34.80c | |
| Lum 5 nM | 26.07c | 28.82d | 34.60c | |
| H2O2 1% | 24.77e | 27.16f | 34.92c | |
| Eth 1000 ppm | 28.03a | 31.71a | 36.50a | |
| BB 3% | 23.79f | 25.82g | 33.52e | |
| SNO 10 μM | 24.98e | 29.25b | 34.93c | |
| NO2 10 μM | 26.96b | 28.16ce | 34.26d | |
| Final percentage: Upper limit of cumulative bud-break % curve | HC 3% | 95.95c | 96.40bc | 95.65e |
| dH2O | 95.81c | 97.48b | 100a | |
| BC204 0.05% | 96.85bc | 94.23e | 99.99ab | |
| Lum 5 nM | 96.35c | 95.27ec | 100a | |
| H2O2 1% | 92.61d | 99.94a | 98.19cd | |
| Eth 1000 ppm | 100a | 96.69bd | 97.10d | |
| BB 3% | 91.87d | 95.43ecd | 98.14cd | |
| SNO 10 μM | 97.77b | 99.38a | 95.55e | |
| NO2 10 μM | 99.34a | 99.27a | 98.72cb | |
Table 4.
Respective coefficient estimates measured after HC 3%, dH2O, SNH, SNL, PBXH, PBXL & MIX2 application on V. vinifera CS SNCs after 100, 200 and 250 CUA (45 biological replicates/treatment). Treatments have been assigned letters which indicate significant differences amongst the coefficient estimates. These letters only apply to individual experiments and parameters.
Table 4.
Respective coefficient estimates measured after HC 3%, dH2O, SNH, SNL, PBXH, PBXL & MIX2 application on V. vinifera CS SNCs after 100, 200 and 250 CUA (45 biological replicates/treatment). Treatments have been assigned letters which indicate significant differences amongst the coefficient estimates. These letters only apply to individual experiments and parameters.
| Coefficient estimate | ||||
|---|---|---|---|---|
| Bud-break parameter: definition | Treatment | 100 | 200 | 250 |
| Onset: Number of days to first bud-break | HC 3% | 9.69f | 10.35c | 13.72ab |
| dH2O | 18.20a | 15.48a | 13.77ac | |
| MIX2 | 15.32d | 15.47a | 14.14a | |
| SNH | 11.67e | 9.81c | 10.14e | |
| SNL | 17.33b | 13.86b | 12.60bd | |
| PBXH | 16.76c | 15.50a | 13.20bc | |
| PBXL | 17.32b | 15.88a | 12.34d | |
| Rate: Slope between onset and final bud-break % | HC 3% | -0.32a | -0.29a | -0.23a |
| dH2O | -0.52d | -0.79b | -0.59c | |
| MIX2 | -0.62b | -0.83b | -0.52bc | |
| SNH | -0.39c | -0.57c | -0.44b | |
| SNL | -0.50d | -0.72bd | -0.52bc | |
| PBXH | -0.44e | -0.77b | -0.47b | |
| PBXL | -0.52d | -0.65cd | -0.31d | |
| EC50: Number of days to 50 % of final bud-break %. | HC 3% | 17.02e | 18.71bc | 24.96a |
| dH2O | 22.88b | 18.57c | 17.99f | |
| MIX2 | 19.31c | 18.77bc | 19.09c | |
| SNH | 18.29d | 14.05e | 15.67g | |
| SNL | 22.35a | 17.64d | 17.46e | |
| PBXH | 22.83b | 18.89b | 18.57d | |
| PBXL | 22.40a | 19.81a | 20.18b | |
| Final percentage: Upper limit of cumulative bud-break % curve | HC 3% | 98.15b | 93.25d | 70.71e |
| dH2O | 99.46a | 92.90d | 91.57c | |
| MIX2 | 97.95b | 99.85a | 97.42a | |
| SNH | 97.29b | 92.65d | 81.03d | |
| SNL | 97.43b | 95.76c | 95.24b | |
| PBXH | 95.31c | 99.89a | 98.11a | |
| PBXL | 99.18a | 97.56b | 97.66a | |
Table 5.
Respective coefficient estimates measured during V. vinifera CS dormancy-release small scale field trial after HC 3 %, dH2O, SNH and PBXH treatment. Treatments have been assigned letters which indicate significant differences amongst the coefficient estimates. These letters only apply to individual experiments and parameters.
Table 5.
Respective coefficient estimates measured during V. vinifera CS dormancy-release small scale field trial after HC 3 %, dH2O, SNH and PBXH treatment. Treatments have been assigned letters which indicate significant differences amongst the coefficient estimates. These letters only apply to individual experiments and parameters.
| Bud-break parameter: definition | Treatment | Coefficient estimate |
|---|---|---|
| Onset: Number of days to first bud-break | HC 3% | 8.38a |
| dH2O | 6.53b | |
| PBXH | 6.07bc | |
| SNH | 5.70c | |
| Rate: Slope between onset and final bud-break % | HC 3% | -0.36a |
| dH2O | -0.38a | |
| PBXH | -0.39a | |
| SNH | -0.38a | |
| EC50: Number of days to 50 % of final bud-break %. | HC 3% | 15.61a |
| dH2O | 13.09b | |
| PBXH | 12.62c | |
| SNH | 12.74c | |
| Final percentage: Upper limit of cumulative bud-break % curve | HC 3% | 85.26a |
| dH2O | 78.69d | |
| PBXH | 80.20c | |
| SNH | 82.28b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



