
Submitted:
01 April 2024
Posted:
01 April 2024
You are already at the latest version
Abstract
This review aims to explore the intricate relationship between epigenetic mechanisms, stress, and affective disorders, focusing on how early life experiences and coping mechanisms contribute to susceptibility to mood disorders. Epigenetic factors play a crucial role in regulating gene expression without altering the DNA (deoxyribonucleic acid) sequence, and recent research has revealed associations between epigenetic changes and maladaptive responses to stress or psychiatric disorders. A scoping review of 33 studies employing the PRISMA-S (Preferred Reporting Items for Systematic Reviews and Meta-Analyses – Statement) guidelines investigates the role of stress-induced epigenetic mechanisms and coping strategies in affective disorders' occurrence, development, and progression. The analysis encompasses various stress factors, including childhood trauma, work-related stress, and dietary deficiencies, alongside epigenetic changes such as DNA methylation and altered gene expression. Findings indicate that specific stress-related genes frequently exhibit epigenetic changes associated with affective disorders. Moreover, the review examines coping mechanisms in patients with bipolar disorder and major depressive disorder, revealing mixed associations between coping strategies and symptom severity. While active coping is correlated with better outcomes, emotion-focused coping may exacerbate depressive or manic episodes. Overall, this review underscores the complex interplay between genetic predisposition, environmental stressors, coping mechanisms, and affective disorders. Understanding these interactions is essential for developing targeted interventions and personalized treatment strategies for individuals with mood disorders. However, further research is needed to elucidate specific genomic loci involved in affective disorders and the clinical implications of coping strategies in therapeutic settings.
Keywords:
epigenetic
; coping
; stress
; affective disorders
; DNA methylation
; childhood trauma
; stressor
; depressive disorder
; dietary deficiencies
1. Introduction
Epigenetic factors refer to functional changes in the genome without changes in the DNA sequence. Such modifications regulate gene expression and phenotype, for example, through mechanisms such as DNA methylation. Epigenetic differences may be a consequence of exposure to stress-related factors during critical periods of development and, therefore, contribute to susceptibility to certain psychiatric disorders. Recent studies have shown associations between specific epigenetic changes and the risk of maladaptive responses to stress or mental disorders [1].
Genes do not function as fixed patterns; their expression is regulated dynamically and often reversibly. Epigenetic molecular elements are cell chromatin, dynamic DNA, histone, and microRNA changes [2].
MicroRNAs are small RNA molecules that act as regulators of gene expression; they create a mechanism for modulating protein synthesis. Their aberrant expression contributes to the development of some diseases, with implications in epigenetic therapy. Unlike the genetic code, the epigenetic code is different and dynamic in all body cells; it responds to environmental changes and chemical, biological, and social aggression and targets psychopharmacological interventions [2].
Affective disorders tend to be both recurrent and progressive, in the sense that the episodes follow one another after shorter remission intervals or with an increased rate of cyclicity [3,4]. About 70% of patients who had the first episode of unipolar depression will later have multiple episodes, and almost all bipolar disorders are recurrent [5].
Numerous studies have shown that psychosocial factors can precipitate depression as well as mania [6,7]. However, most people who are exposed to stressful events do not develop a psychiatric disorder [8,9,10,11]. This begs the question: why do some people develop an emotional disorder about a stressful life event and others do not? The answer to this question is complex; it involves genetic predisposition, personality, previous experiences, family distress, adequate social support network and probably the individual’s response to stress [12].
P Preclinical animal studies have shown that exposure to stress is associated with changes in the epigenome (e.g., changes in the genes involved in the stress response), as well as the occurrence of depression-like behaviour; similarly, chronic exposure to unpredictable stress alters histone acetylation in the forebrain, with the appearance of anxiety-like behaviour and increased vulnerability to stress, as well as cognitive deficits. In the case of animals that were subjected to stress from the first days of life and separation from their mother, they showed depressive behaviours in adulthood [13].
Studies conducted on the families of patients with major depressive disorder or bipolar disorder have shown that there is a greater segregation of these affective disorders among family members [14,15]. The transmission rate to first-degree relatives of type I bipolar disorder is between 4-24%, type II bipolar disorder is 1-5%, and major depressive disorder is 4-24% [16]. Identical values were also described in schizophrenia and schizoaffective disorder.
Empirical data have highlighted the significant interindividual differences in response to stress and adversity. Some authors speak of the “invulnerable child”, referring to the positive interaction of events that determine the child’s resilience (invulnerability) to adversity [17]. A simple interpretation of such a phenomenon is the interaction between genetic and environmental factors that ultimately determine susceptibility. However, the need remains to translate the possible genetic influences of individual vulnerability into psychological mechanisms. This review aims to analyse clinical trials on coping mechanisms and their interaction with epigenetic mechanisms in patients with mood disorders. The effect of the interaction between the two mechanisms on these disorders’ onset, recurrence, and progression was mainly followed in the review.
2. Materials and Methods
According to PRISMA-S guidelines, 33 studies were reviewed in this article. These studies investigate both the role of epigenetic mechanisms induced by stressors in the occurrence/development and/or progression of affective disorders and the role of stress adaptation mechanisms in these disorders. Stress factors include childhood trauma, stress at work, dietary deficiencies, exposure to chemicals, and so on. Inclusion has not been limited to studies examining specific types of stressors.
Regarding epigenetic changes, they were defined to include any valid indicator of epigenetic changes (e.g., DNA methylation, DNA acetylation, altered transcription, direct changes in gene expression).
Inclusion criteria:
- studies in which a valid measurement of epigenetic changes that were associated with a significant stress factor (death of a loved one, emotional, physical, sexual abuse, etc.) was performed;
- studies that assessed whether the epigenetic change was correlated with a diagnosis of an affective disorder;
Exclusion criteria:
- subjects with somatic comorbidities,
- subjects under the age of 18 years old.
The studies were identified by searching the PubMed database between 2010-2023, using the following keywords: epigenetic mechanisms, stressors, major depressive disorder, suicidal ideation, bipolar disorder, mania, and coping results.
Figure 1.
Selection process in PRISMA flow diagram.
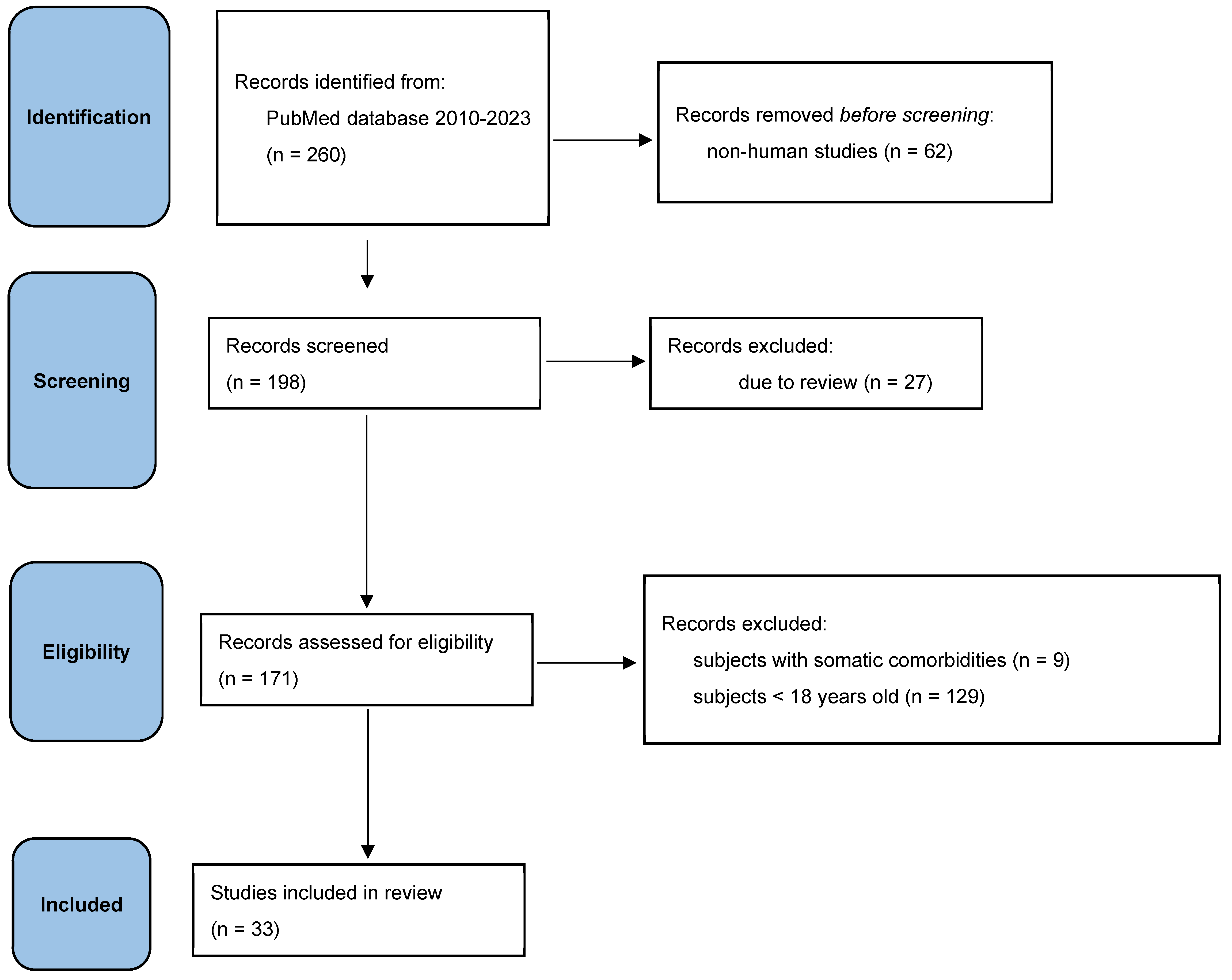
3. Results
Thirty-three articles were screened that followed the relationship between an epigenetic change associated with stress and the diagnosis of affective disorder, as well as the relationship between a stress factor, induced epigenetic changes and the evaluation of psychiatric symptoms (suicidal ideation, manic states, etc.). DNA methylation, in particular, was the change evaluated in these studies. Moreover, despite the broad meaning of the stressors, the identified studies assessed the adversity of early childhood experiences (abuse, abuse, early loss of a parent).
Epigenetic changes in the following stress-associated genes have been frequently linked to the diagnosis of affective disorder (Table 1):
- NR3C1 (human glucocorticoid receptor gene);
- SLC6A4 (serotonergic transporter gene);
- BDNF (brain-derived neurotrophic factor);
- FKBP5 (FK506 5 binding protein gene);
- SKA2 (kinetochore protein gene);
- OXTR (oxytocin receptor) and genes encoding oligodendrocytes.

Table 1.
Stress-related genes and their association with the presence of an affective disorder.
| GENE | STUDY | STRESS | AFFECTIVE DISORDER | QUESTIONNAIRE |
|---|---|---|---|---|
| NR3C1 | Bustamante et al. [18] | Positive association | Negative association | CTS (child trauma screen), CTQ (childhood trauma questionnaire) |
| Radtke et al. [19] | Didn’t report | Didn’t report | KERF 1 | |
| Perroud et al. [20] | Positive association | Didn’t report | CTQ | |
| De Assis Pinheiro et al. [21] | Negative association | Positive association | BDI-II (Beck Depression Inventory II) | |
| Comtois-Cabana et al. [22] | Positive association | Negative associaton | CTQ-SF (child trauma screen – short form), BDI-II | |
| FKPB 5 | Weder et al. [23] | Positive association | Negative association | CTQ |
| Tyrka et al. [24] | Negative association | Positive association | Interview | |
| Flasbeck&Brüne [25] | Negative association | Positive association | CTQ | |
| SLC6A4 | Sanwald et al. [26] | Positive association | Positive association | MADRS (Montgomery–ÅsbergDepression Rating Scale) |
| Swartz et al. [27] | Positive association | Positive association | Interview | |
| Booji et al. [28] | Positive association | Didn’t report | CTQ | |
| Lei et al. [29] | Positive association | Positive association | Non-standard 11 item scale | |
| Kang et al. [30] | Positive association | Didn’t report | Non-standard childhood adversity | |
| Alaasari et al. [31] | Negative association | Positive association | Karasek-Job Content Questionnaire | |
| Comtois-Cabana et al. [22] | Positive association | Positive association | CTQ-SF, BDI-II | |
| BDNF | Song et al. [32] | Positive association | Negative association | Non-standardself-reportquestionnaire |
| SKA2 | Weder et al. [23] | Didn’t report | Didn’t report | Report of parental abuse and neglect |
| Sadeh et al. [33] | Didn’t report | Positive association | PTSD (post-traumatic stress disorder) scale administered by clinicians | |
| Sadeh et al. [34] | Positive association | Didn’t report | PTSD scale administered by clinicians | |
| OXTR, LINGO3, POU3F1, ITGB1. | Smearman et al. [35] | Positive association | Didn’t report | CTQ |
| Lutz et al. [36] | Positive association | Negative association | CECA (Childhood Experience of Care and Abuse), Interview | |
| Ludwig et al. [37] | Didn’t report | Positive association | CTQ, HAM-D (Hamilton Depression Rating Scale) | |
| Kogan et al. [38] | Positive association | Positive association | ACE (Adverse Childhood Experiences) |
NR3C1 (human glucocorticoid receptor gene). Bustamante and co-workers [18] reported significantly higher scores on the scale of childhood abuse and trauma screening in the group of patients diagnosed with recurrent depressive disorder (n = 76) compared to the healthy control group (n = 76). Furthermore, methylation of the NR3C1 gene could be predicted by a history of childhood abuse or major depressive disorder. History of childhood abuse has been associated with increased methylation at the NR3C1 transcription factor binding site, leading to reduced NR3C1 gene expression; major depressive disorder has been associated with low methylation at the downstream locus, indicating that NR3C1 gene expression is unchanged.
Radtke et al. [19] analysed the association between NR3C1 methylation (at 41 CpG sites) and depressive symptoms in individuals with a history of childhood abuse (n = 46). No significant association was found between NR3C1 methylation and depressive symptoms. However, correlations have been found between the methylation of two CpG loci located in the NR3C1 gene promoter and the specific symptoms of depression.
Perroud et al. [20] described positive correlations between NR3C1 methylation and childhood abuse in patients with major depressive disorder (n = 99). The level of NR3C1 methylation was correlated with the form of abuse (physical abuse, sexual abuse, emotional neglect) and its severity.
De Assis Pinheiro et al. [21] showed that alcohol consumption, overweight, and high cortisol levels are related to NR3C1 non-methylation, while depression is related to its methylation (n = 386).
FKPB2 (FK506 5 binding protein gene). Weder et al. [23] compared children with a history of abuse (n = 94) and no history of abuse (n = 96), noting significant differences in NR3C1 methylation at six CpG sites in the promoter region. The researchers reported that methylation at two of these CpG sites was able to predict the onset of psychiatric symptoms. The study also looked at FKBP5 gene methylation in these groups and found positive correlations between gene methylation in abused children vs. the abused. Significant differences were found between the two groups regarding BDNF methylation.
On the other hand, Tyrka et al. [24] reported significantly lower NR3C1 methylation across the promoter region and at six other CpG sites in people with a history of childhood abuse and/or major depressive disorder, bipolar disorder or generalised anxiety disorder. This result contradicts previous studies, suggesting a much greater complexity in regulating the NR3C1 gene.
Also, Flasbeck&Brüne [25] demonstrated that FKBP5 was associated with anxiety and reduced empathy. Despite expectations, there was no discernible impact of childhood maltreatment on DNA methylation. Additionally, no methylation distinctions were evident between a clinical group and a non-clinical group concerning FKBP5. However, there was a slight discrepancy in NR3C1 methylation levels, although its biological significance is questionable.
SLC6A4 (serotonergic transporter gene). Sanwald et al. [26] concluded that SLC6A4 methylation was not related to depression severity, age at depression onset or SLEs in the entire group but positively related to depression severity in women (n = 95).
Swartz and colleagues [27] operationalized environmental stress, using adolescent socioeconomic status as a measurement method. The researchers found that poor socioeconomic status was associated with higher methylation of SLC6A4, which may lead to worsening of depressive symptoms. These findings suggest that adolescent stress may contribute to the severity of the disease through epigenetic changes in the SLC6A4 gene.
Booij et al. [28] compared individuals with a diagnosis of major depressive disorder (n = 33) and the healthy control group (n = 36), reporting insignificant differences in SLC6A4 methylation. However, the authors found positive correlations between the history of childhood abuse and SLC6A4 methylation.
In another study that operationalized environmental stress, Lei et al. [29] used the crime rate in the neighbourhood as a measure. These correlate positively with methylation of the SLC6A4 promoter, but only in individuals carrying the short allele gene. These findings suggest that the gene’s interaction with environmental factors may interact in a genotype-dependent manner.
Kang and colleagues [30] analysed the association between childhood adversity, the severity of depressive symptoms, and SLC6A4 methylation in patients with major depressive disorder (n = 108). Thus, higher methylation has been reported in patients with severe symptoms and a history of childhood abuse.
On the other hand, Alasaari et al. [31] reported significantly lower SLC6A4 promoter methylation in nurses (n = 24) compared to other areas with a low stress level (n = 25). These results contradict the results of previous studies, which support a positive association between stress and SLC6A4 methylation.
In 2023, Comtois-Cabana et al. [22] investigated the association between depressive symptoms and methylation levels of specific genes, including NR3C1 and SLC6A4. Adults with higher depressive symptoms exhibited higher methylation levels at two CpG sites across the NR3C1 promoter regions (n = 34) and lower methylation levels at three CpG sites across the SLC6A4 promoter region (n = 31). This study is the first to investigate the association between depressive symptoms and NR3C1 methylation levels in saliva samples of adults. The findings are consistent with some previous studies that also detected higher levels of NR3C1 methylation in blood samples of depressed adults compared to controls. While the majority of studies have found higher SLC6A4 methylation levels in association with depressive symptoms, some studies have reported conflicting results.
Song et al. [32] analyzed the correlations between BDNF methylation, scale scores for depressive symptoms, and work stress in the Japanese population (n = 774). Significantly lower BDNF methylation was reported in individuals with high questionnaire scores but considerably higher in individuals with high work stress levels.
Studies by Sadeh et al. [33,34] investigated the relationship between post-traumatic stress, depression and SKA2 gene methylation in a group of war veterans. In the first study, a positive association between PTSD symptoms and SKA2 methylation was reported, but no association with depressive symptoms (n = 145). On the other hand, in a subsequent study, Sadeh and colleagues reported an association between PTSD and SKA2 methylation, as well as an association between gene methylation and depressive symptoms (n = 466). The authors acknowledged that the discrepancy was due to chronic PTSD (i.e., the length of time since PTSD was diagnosed).
OXTR (oxytocin receptor). Smearman et al. [35] reported a positive association between a history of childhood abuse and OXTR methylation; however, the association was no longer valid after the correction for multiple comparisons.
Ludwig B. et al. [37] suggest a positive but nonsignificant association between the severity of depression symptoms and OXTR methylation. Also, the severity of emotional neglect in patients with affective disorders, but not childhood adverse experiences, was associated with OXTR methylation levels. On the other hand, Reiner et al. [39] found significantly lower exon 1 OXTR DNA methylation in depressed patients compared to healthy controls both before and after treatment. This suggests that lower methylation at CpG sites is associated with higher transcriptional activity of the OXTR gene, potentially leading to increased oxytocin receptor expression in brain areas implicated in depression.
Kogan et al. [38] demonstrate that contextual stressors, both in childhood and emerging adulthood, can increase defensive/hostile relational schemas. These schemas, in turn, are linked to substance abuse and depressive symptoms. Interestingly, the study also explores the moderating role of DNA methylation in the OXTR gene. When OXTR DNA methylation levels were high, the association between contextual stress and defensive/hostile relational schemas was exacerbated. Conversely, when OXTR DNA methylation levels were low, contextual stress did not significantly influence defensive/hostile schemas.
Regarding specific coping strategies, none of the studies thoroughly answered whether they are predictive of affective disorders. However, in most studies, emotion-focused coping is associated with the recurrence of depressive or manic episodes. These strategies could also be associated with a longer recovery time. Relatively few bipolar disorder studies have been identified, with findings mainly limited to major depressive disorder. In cross-sectional studies, a clear distinction between psychiatric symptoms and emotion-focused coping strategies is difficult to achieve, so the results of these studies can only partially address the goal (Table 2).
Table 2.
Coping mechanisms in patients with bipolar disorder and major depressive disorder.
| STUDY | NUMBER OF PATIENTS | GENDER | AGE (average) | QUESTIONNARE | RESULTS |
|---|---|---|---|---|---|
| Kasi et al. [40] | 162, MDD (major depressive disorder) and GAD (generalized anxiety disorder) | 74.4% M; 25.3% F | It does not specify | COPE (Coping Orientation to Problems Experienced Inventory) | In patients diagnosed with generalised anxiety disorder or major depressive disorder, “religion-oriented” was the most common coping mechanism identified. |
| Horwitz et al. [41] | 286, MDD | 41%M; 59% F | 18 | COPE, C-SSRS (Columbia Suicide Severity Rating Scale) | Active coping was correlated with lower C-SSRS sores at follow-up. |
| Fletcher et al. [42] | 379, BD (bipolar disorder) + MDD | 41% M 59%F | 39 | COPE, RPA (Responses to Positive Affect), CIPM (Coping Styles in Prodrome of Bipolar Mania), RSQ (Response Style Questionnaire),CERQ (Cognitive Emotion Regulation Questionnaire) | A number of differences were found between the group of patients with unipolar depression and the group with bipolar depression, the former being oriented towards active coping, focused on the problem. |
| Au CH et al. [43] | 115, BD | 37%M; 63%F | 47 | SCOS (Stigma Coping Orientation Scale) | It has been reported that low self-esteem is crucial to social functioning. Dysfunctional coping predominates among these patients. |
| Nitzburg et al. [44] | 92, BD | 48%M; 42%F | 45 | COPE | Dysfunctional coping is a predictive factor for many disabilities, while active coping is associated with resilience. Likewise, behavioral disengagement and guilt are predictors of disability. |
| Paans et al. [45] | 90, BD | 45%M; 55%F | 67 | UCL | The authors reported positive associations between better cognitive functioning and active coping. |
| Lin J et al. [7] | 310, MDD with suicidal risk | It does not specify | 30 | SCSQ (Simplified Coping Style Questionnaire) | Patients at risk of suicide had negative coping strategies and an inadequate social support network. |
| Kuiper et al. [45] | 89, MDD | It does not specify | 20 | COPE | Problem-centred coping has been shown to correlate with better functionality. Emotion-centred coping and dysfunctional coping have been associated with low resilience |
| Orzechowska et al. [46] | 80, MDD and BD | 48 women, 32 men | 49 | COPE | Unlike healthy people, depressed patients in stressful situations more often use strategies based on avoidance, denial and have more difficulty in finding positive aspects of stressful events. |
| Roohafza HR et al. [47] | 4685, MDD and GAD | It does not specify | 49 | COPE | The results show that positive interpretation and growth, active coping and a supportive social network are protective factors in major depressive disorder and generalised anxiety disorder. |
Longitudinal studies have consistently demonstrated the pivotal role of coping strategies in determining the trajectory of affective disorders. Research by Fletcher et al. [42] and Horwitz et al. [41], and Kasi et al. [40] highlight that adaptive coping strategies are associated with more extended remission periods and a reduced risk of recurrence. Conversely, passive coping is linked to an increased risk of recurrence and more severe symptoms of depression. Additionally, while individuals diagnosed with major depressive disorder may not significantly differ from their healthy counterparts in their response to stressful situations [46], problem-centred coping emerges as a good predictor of post-hospital symptoms [47].
4. Discussion
The review explores the complex interplay between epigenetic mechanisms, stress, coping strategies, and affective disorders. It synthesizes findings from 33 studies to elucidate the role of epigenetic changes induced by stressors in mood disorders’ occurrence, development, and progression, alongside the influence of coping mechanisms on these processes.
Epigenetic Mechanisms and Affective Disorders
The review highlights the significant associations between epigenetic changes in stress-related genes and the diagnosis of affective disorders. Specifically, genes such as NR3C1, SLC6A4, BDNF, FKBP5, SKA2, and OXTR exhibit alterations in DNA methylation patterns that are frequently linked to mood disorders. Notably, these changes often stem from early life experiences, including childhood trauma, parental abuse, or neglect. However, the relationship between specific epigenetic modifications and psychiatric symptoms is nuanced, with contradictory findings observed in some studies. For instance, while some studies report higher methylation levels associated with depressive symptoms, others find lower methylation levels [18,19,20].
Coping Mechanisms and Affective Disorders
The review discusses the role of coping strategies in modulating the severity and recurrence of affective disorders. Emotion-focused coping, characterized by strategies aimed at managing emotional distress, shows mixed associations with symptom severity. While some studies suggest that emotion-focused coping exacerbates depressive or manic episodes, others report no significant impact [40,41]. Conversely, active coping strategies, such as problem-solving and seeking social support, are correlated with better outcomes, including more extended remission periods and lower recurrence rates [42,44]. However, the distinction between adaptive and maladaptive coping strategies is complex, and longitudinal studies are needed to delineate their predictive value in affective disorders.
Individual Variability and Resilience
Empirical data highlight significant interindividual differences in response to stress and adversity [17]. While some individuals exhibit resilience to adverse experiences, others may develop affective disorders. The “invulnerable child” phenomenon suggests that positive interactions of events contribute to resilience, possibly through the interplay of genetic and environmental factors [12].
Clinical Implications and Future Directions
Understanding the intricate relationship between epigenetic mechanisms, stress, coping mechanisms, and affective disorders has important clinical implications [48,49,50]. Targeted interventions that address genetic predisposition and environmental stressors could lead to more effective treatment strategies. Longitudinal studies are needed to elucidate further the role of coping mechanisms in the onset, recurrence, and progression of affective disorders [41,42]. Additionally, future research should focus on identifying specific genomic loci involved in affective disorders and exploring the clinical implications of coping strategies in therapeutic settings [1].
The study has some limitations:
- Heterogeneity of study designs: The review encompasses studies with varying methodologies, including cross-sectional and longitudinal designs, which may limit the generalizability of findings.
- Sample characteristics: Studies included in the review involve diverse patient populations with variations in age, gender, and clinical characteristics, potentially confounding the interpretation of results.
- Measurement of epigenetic changes: The review primarily focuses on DNA methylation as a proxy for epigenetic alterations, overlooking other mechanisms such as histone modifications or microRNA regulation, which could also contribute to the pathophysiology of affective disorders.
- Causality and directionality: Most reviewed studies establish associations between epigenetic changes, stressors, coping mechanisms, and affective disorders, but causality and directionality remain unclear. Longitudinal studies are necessary to elucidate temporal relationships and causal pathways.
- Publication bias: The review may be subject to publication bias, as studies reporting statistically significant findings are more likely to be published, potentially skewing the evidence synthesis.
Further research should address the limitations mentioned above by employing longitudinal designs, integrating multi-omic approaches to explore comprehensive epigenetic mechanisms, and examining diverse coping strategies across different stages of affective disorders. Additionally, translational studies are warranted to translate research findings into personalized interventions and therapeutic approaches for individuals with mood disorders [1,12].
5. Conclusions
The review underscores the intricate interplay between epigenetic processes, stress responses, coping mechanisms, and affective disorders, providing valuable insights into the underlying mechanisms and avenues for future investigation and clinical intervention.
Despite challenges and limitations, understanding these interactions is critical for advancing our knowledge of mood disorders and developing more targeted and personalized treatment approaches.
Author Contributions
Conceptualization, B.C.B. and D.A.A.; methodology, E.S.B., D.L., and M.C.T.; formal analysis, D.L., V.E. and L.H.; investigation, A.T. and V.E.; data curation, V.N. and D.A.A.; writing—original draft preparation, B.C.B., E.S.B., and A.T.; writing—review and editing, B.C.B., E.S.B., D.L., M.C.T., V.N. and D.A.A.; supervision, E.S.B., L.H.,V.E. All authors have read and agreed to the published version of the manuscript.
Funding
We would like to acknowledge “Victor Babes” University of Medicine and Pharmacy Timisoara, Romania for their support in covering the costs of publication for this research paper.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schiele, M.A.; Gottschalk, M.G.; Domschke, K. The Applied Implications of Epigenetics in Anxiety, Affective and Stress-Related Disorders—A Review and Synthesis on Psychosocial Stress, Psychotherapy and Prevention. Clin. Psychol. Rev. 2020, 77, 101830. [Google Scholar] [CrossRef] [PubMed]
- Lacal, I.; Ventura, R. Epigenetic Inheritance: Concepts, Mechanisms and Perspectives. Front. Mol. Neurosci. 2018, 11, 292. [Google Scholar] [CrossRef] [PubMed]
- Knapik, A.; Krzystanek, E.; Szefler–Derela, J.; Siuda, J.; Rottermund, J.; Plinta, R.; Brzęk, A. Affective Disorder and Functional Status as Well as Selected Sociodemographic Characteristics in Patients with Multiple Sclerosis, Parkinson’s Disease and History of Stroke. Medicina 2020, 56, 117. [Google Scholar] [CrossRef]
- Post, R.M. Mechanisms of Illness Progression in the Recurrent Affective Disorders. Neurotox. Res. 2010, 18, 256–271. [Google Scholar] [CrossRef]
- Saunders, K.E.A.; Goodwin, G.M. The Course of Bipolar Disorder. Adv. Psychiatr. Treat. 2010, 16, 318–328. [Google Scholar] [CrossRef]
- Huang, Y.; Wongpakaran, T.; Wongpakaran, N.; Bhatarasakoon, P.; Pichayapan, P.; Worland, S. Depression and Its Associated Factors among Undergraduate Engineering Students: A Cross-Sectional Survey in Thailand. Healthcare 2023, 11, 2334. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Su, Y.; Lv, X.; Liu, Q.; Wang, G.; Wei, J.; Zhu, G.; Chen, Q.; Tian, H.; Zhang, K.; et al. Perceived Stressfulness Mediates the Effects of Subjective Social Support and Negative Coping Style on Suicide Risk in Chinese Patients with Major Depressive Disorder. J. Affect. Disord. 2020, 265, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.T.; Holmes, S.E.; Pietrzak, R.H.; Esterlis, I. Neurobiology of Chronic Stress-Related Psychiatric Disorders: Evidence from Molecular Imaging Studies. Chronic Stress 2017, 1, 247054701771091. [Google Scholar] [CrossRef]
- Enatescu, V.-R.; Bernad, E.; Gluhovschi, A.; Papava, I.; Romosan, R.; Palicsak, A.; Munteanu, R.; Craina, M.; Enatescu, I. Perinatal Characteristics and Mother’s Personality Profile Associated with Increased Likelihood of Postpartum Depression Occurrence in a Romanian Outpatient Sample. J Ment Health 2017, 26, 212–219. [Google Scholar] [CrossRef]
- Park, H.G.; Jeong, S.; Kwon, M. Factors Related to Depression According to the Degree of Loneliness in Adolescents with Severe Friend-Relationship Stress. Healthcare 2023, 11, 1354. [Google Scholar] [CrossRef]
- Răchită, A.; Strete, G.E.; Suciu, L.M.; Ghiga, D.V.; Sălcudean, A.; Mărginean, C. Psychological Stress Perceived by Pregnant Women in the Last Trimester of Pregnancy. Int. J. Environ. Res. Public. Health 2022, 19, 8315. [Google Scholar] [CrossRef] [PubMed]
- Christensen, M.V.; Kessing, L.V. Clinical Use of Coping in Affective Disorder, a Critical Review of the Literature. Clin. Pract. Epidemiol. Ment. Health 2005. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Pan, F.; Tang, Y.; Huang, J.H. Editorial: Early Life Stress-Induced Epigenetic Changes Involved in Mental Disorders. Front. Genet. 2021, 12, 684844. [Google Scholar] [CrossRef] [PubMed]
- Maher, B.S.; Marazita, M.L.; Zubenko, W.N.; Spiker, D.G.; Giles, D.E.; Kaplan, B.B.; Zubenko, G.S. Genetic Segregation Analysis of Recurrent, Early-onset Major Depression: Evidence for Single Major Locus Transmission. Am. J. Med. Genet. 2002, 114, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Zhao, Y.; Tan, Z.; Lai, J.; Chen, J.; Zhang, X.; Sun, J.; Chen, L.; Lu, K.; Cao, L.; et al. Differentiation between Bipolar Disorder and Major Depressive Disorder in Adolescents: From Clinical to Biological Biomarkers. Front. Hum. Neurosci. 2023, 17, 1192544. [Google Scholar] [CrossRef] [PubMed]
- Rowland, T.A.; Marwaha, S. Epidemiology and Risk Factors for Bipolar Disorder. Ther. Adv. Psychopharmacol. 2018, 8, 251–269. [Google Scholar] [CrossRef] [PubMed]
- Thorup, A.A.E.; Jepsen, J.R.; Ellersgaard, D.V.; Burton, B.K.; Christiani, C.J.; Hemager, N.; Skjærbæk, M.; Ranning, A.; Spang, K.S.; Gantriis, D.L.; et al. The Danish High Risk and Resilience Study—VIA 7—A Cohort Study of 520 7-Year-Old Children Born of Parents Diagnosed with Either Schizophrenia, Bipolar Disorder or Neither of These Two Mental Disorders. BMC Psychiatry 2015, 15, 233. [Google Scholar] [CrossRef]
- Bustamante, A.C.; Aiello, A.E.; Galea, S.; Ratanatharathorn, A.; Noronha, C.; Wildman, D.E.; Uddin, M. Glucocorticoid Receptor DNA Methylation, Childhood Maltreatment and Major Depression. J. Affect. Disord. 2016, 206, 181–188. [Google Scholar] [CrossRef]
- Radtke, K.M.; Schauer, M.; Gunter, H.M.; Ruf-Leuschner, M.; Sill, J.; Meyer, A.; Elbert, T. Epigenetic Modifications of the Glucocorticoid Receptor Gene Are Associated with the Vulnerability to Psychopathology in Childhood Maltreatment. Transl. Psychiatry 2015, 5, e571. [Google Scholar] [CrossRef]
- Perroud, N.; Paoloni-Giacobino, A.; Prada, P.; Olié, E.; Salzmann, A.; Nicastro, R.; Guillaume, S.; Mouthon, D.; Stouder, C.; Dieben, K.; et al. Increased Methylation of Glucocorticoid Receptor Gene (NR3C1) in Adults with a History of Childhood Maltreatment: A Link with the Severity and Type of Trauma. Transl. Psychiatry 2011, 1, e59. [Google Scholar] [CrossRef]
- De Assis Pinheiro, J.; Freitas, F.V.; Borçoi, A.R.; Mendes, S.O.; Conti, C.L.; Arpini, J.K.; Dos Santos Vieira, T.; De Souza, R.A.; Dos Santos, D.P.; Barbosa, W.M.; et al. Alcohol Consumption, Depression, Overweight and Cortisol Levels as Determining Factors for NR3C1 Gene Methylation. Sci. Rep. 2021, 11, 6768. [Google Scholar] [CrossRef]
- Comtois-Cabana, M.; Barr, E.; Provençal, N.; Ouellet-Morin, I. Association between Child Maltreatment and Depressive Symptoms in Emerging Adulthood: The Mediating and Moderating Roles of DNA Methylation. PLoS ONE 2023, 18, e0280203. [Google Scholar] [CrossRef] [PubMed]
- Weder, N.; Zhang, H.; Jensen, K.; Yang, B.Z.; Simen, A.; Jackowski, A.; Lipschitz, D.; Douglas-Palumberi, H.; Ge, M.; Perepletchikova, F.; et al. Child Abuse, Depression, and Methylation in Genes Involved With Stress, Neural Plasticity, and Brain Circuitry. J. Am. Acad. Child Adolesc. Psychiatry 2014, 53, 417–424. [Google Scholar] [CrossRef]
- Tyrka, A.R.; Parade, S.H.; Welch, E.S.; Ridout, K.K.; Price, L.H.; Marsit, C.; Philip, N.S.; Carpenter, L.L. Methylation of the Leukocyte Glucocorticoid Receptor Gene Promoter in Adults: Associations with Early Adversity and Depressive, Anxiety and Substance-Use Disorders. Transl. Psychiatry 2016, 6, e848. [Google Scholar] [CrossRef]
- Flasbeck, V.; Brüne, M. Association between Childhood Maltreatment, Psychopathology and DNA Methylation of Genes Involved in Stress Regulation: Evidence from a Study in Borderline Personality Disorder. PLoS ONE 2021, 16, e0248514. [Google Scholar] [CrossRef]
- Sanwald, S.; Widenhorn-Müller, K.; Schönfeldt-Lecuona, C.; GenEmo Research Group; Connemann, B.J.; Gahr, M.; Kammer, T.; Montag, C.; Kiefer, M. Factors Related to Age at Depression Onset: The Role of SLC6A4 Methylation, Sex, Exposure to Stressful Life Events and Personality in a Sample of Inpatients Suffering from Major Depression. BMC Psychiatry 2021, 21, 167. [Google Scholar] [CrossRef]
- Swartz, J.R.; Hariri, A.R.; Williamson, D.E. An Epigenetic Mechanism Links Socioeconomic Status to Changes in Depression-Related Brain Function in High-Risk Adolescents. Mol. Psychiatry 2017, 22, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Booij, L.; Szyf, M.; Carballedo, A.; Frey, E.-M.; Morris, D.; Dymov, S.; Vaisheva, F.; Ly, V.; Fahey, C.; Meaney, J.; et al. DNA Methylation of the Serotonin Transporter Gene in Peripheral Cells and Stress-Related Changes in Hippocampal Volume: A Study in Depressed Patients and Healthy Controls. PLoS ONE 2015, 10, e0119061. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.-K.; Beach, S.R.H.; Simons, R.L.; Philibert, R.A. Neighborhood Crime and Depressive Symptoms among African American Women: Genetic Moderation and Epigenetic Mediation of Effects. Soc. Sci. Med. 2015, 146, 120–128. [Google Scholar] [CrossRef]
- Kang, H.-J.; Kim, J.-M.; Stewart, R.; Kim, S.-Y.; Bae, K.-Y.; Kim, S.-W.; Shin, I.-S.; Shin, M.-G.; Yoon, J.-S. Association of SLC6A4 Methylation with Early Adversity, Characteristics and Outcomes in Depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 44, 23–28. [Google Scholar] [CrossRef]
- Alasaari, J.S.; Lagus, M.; Ollila, H.M.; Toivola, A.; Kivimäki, M.; Vahtera, J.; Kronholm, E.; Härmä, M.; Puttonen, S.; Paunio, T. Environmental Stress Affects DNA Methylation of a CpG Rich Promoter Region of Serotonin Transporter Gene in a Nurse Cohort. PLoS ONE 2012, 7, e45813. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Miyaki, K.; Suzuki, T.; Sasaki, Y.; Tsutsumi, A.; Kawakami, N.; Shimazu, A.; Takahashi, M.; Inoue, A.; Kan, C.; et al. Altered DNA Methylation Status of Human Brain Derived Neurotrophis Factor Gene Could Be Useful as Biomarker of Depression. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2014, 165, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Sadeh, N.; Spielberg, J.M.; Logue, M.W.; Wolf, E.J.; Smith, A.K.; Lusk, J.; Hayes, J.P.; Sperbeck, E.; Milberg, W.P.; McGlinchey, R.E.; et al. SKA2 Methylation Is Associated with Decreased Prefrontal Cortical Thickness and Greater PTSD Severity among Trauma-Exposed Veterans. Mol. Psychiatry 2016, 21, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Sadeh, N.; Wolf, E.J.; Logue, M.W.; Hayes, J.P.; Stone, A.; Griffin, L.M.; Schichman, S.A.; Miller, M.W. EPIGENETIC VARIATION AT SKA2 PREDICTS SUICIDE PHENOTYPES AND INTERNALIZING PSYCHOPATHOLOGY: 2015 Donald F Klein Investigator Award Finalist: Epigenetic Variation at SKA2 Predicts Suicide Phenotypes. Depress. Anxiety 2016, 33, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Smearman, E.L.; Almli, L.M.; Conneely, K.N.; Brody, G.H.; Sales, J.M.; Bradley, B.; Ressler, K.J.; Smith, A.K. Oxytocin Receptor Genetic and Epigenetic Variations: Association With Child Abuse and Adult Psychiatric Symptoms. Child Dev. 2016, 87, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Lutz, P.-E.; Tanti, A.; Gasecka, A.; Barnett-Burns, S.; Kim, J.J.; Zhou, Y.; Chen, G.G.; Wakid, M.; Shaw, M.; Almeida, D.; et al. Association of a History of Child Abuse With Impaired Myelination in the Anterior Cingulate Cortex: Convergent Epigenetic, Transcriptional, and Morphological Evidence. Am. J. Psychiatry 2017, 174, 1185–1194. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, B.; Carlberg, L.; Kienesberger, K.; Swoboda, P.; Swoboda, M.M.M.; Bernegger, A.; Koller, R.; Inaner, M.; Fuxjäger, M.; Zotter, M.; et al. Oxytocin Receptor Gene Methylation as a Molecular Marker for Severity of Depressive Symptoms in Affective Disorder Patients. BMC Psychiatry 2022, 22, 381. [Google Scholar] [CrossRef] [PubMed]
- Kogan, S.M.; Bae, D.; Cho, J.; Smith, A.K.; Nishitani, S. Pathways Linking Adverse Environments to Emerging Adults’ Substance Abuse and Depressive Symptoms: A Prospective Analysis of Rural African American Men. Dev. Psychopathol. 2021, 33, 1496–1506. [Google Scholar] [CrossRef] [PubMed]
- Reiner, I.C.; Gimpl, G.; Beutel, M.E.; Bakermans-Kranenburg, M.J.; Frieling, H. OXTR-Related Markers in Clinical Depression: A Longitudinal Case–Control Psychotherapy Study. J. Mol. Neurosci. 2022, 72, 695–707. [Google Scholar] [CrossRef]
- Kasi, P.M.; Naqvi, H.A.; Afghan, A.K.; Khawar, T.; Khan, F.H.; Khan, U.Z.; Khuwaja, U.B.; Kiani, J.; Khan, H.M. Coping Styles in Patients with Anxiety and Depression. ISRN Psychiatry 2012, 2012, 128672. [Google Scholar] [CrossRef]
- Horwitz, A.G.; Czyz, E.K.; Berona, J.; King, C.A. Prospective Associations of Coping Styles With Depression and Suicide Risk Among Psychiatric Emergency Patients. Behav. Ther. 2018, 49, 225–236. [Google Scholar] [CrossRef]
- Fletcher, K.; Parker, G.B.; Manicavasagar, V. Coping Profiles in Bipolar Disorder. Compr. Psychiatry 2013, 54, 1177–1184. [Google Scholar] [CrossRef] [PubMed]
- Au, C.-H.; Wong, C.S.-M.; Law, C.-W.; Wong, M.-C.; Chung, K.-F. Self-Stigma, Stigma Coping and Functioning in Remitted Bipolar Disorder. Gen. Hosp. Psychiatry 2019, 57, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Nitzburg, G.C.; Russo, M.; Cuesta-Diaz, A.; Ospina, L.; Shanahan, M.; Perez-Rodriguez, M.; McGrath, M.; Burdick, K.E. Coping Strategies and Real-World Functioning in Bipolar Disorder. J. Affect. Disord. 2016, 198, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Paans, N.P.G.; Dols, A.; Comijs, H.C.; Stek, M.L.; Schouws, S.N.T.M. Associations between Cognitive Functioning, Mood Symptoms and Coping Styles in Older Age Bipolar Disorder. J. Affect. Disord. 2018, 235, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Orzechowska, A.; Zajączkowska, M.; Talarowska, M.; Gałecki, P. Depression and Ways of Coping with Stress: A Preliminary Study. Med. Sci. Monit. 2013, 19, 1050–1056. [Google Scholar] [CrossRef]
- Roohafza, H.R.; Afshar, H.; Keshteli, A.H.; Mohammadi, N.; Feizi, A.; Taslimi, M.; Adibi, P. What’s the Role of Perceived Social Support and Coping Styles in Depression and Anxiety? J. Res. Med. Sci. 2014. [Google Scholar]
- Bagot, R.C.; Labonté, B.; Peña, C.J.; Nestler, E.J. Epigenetic Signaling in Psychiatric Disorders: Stress and Depression. Dialogues Clin. Neurosci. 2014, 16, 281–295. [Google Scholar] [CrossRef]
- Lee, J.S.; Jaini, P.A.; Papa, F. An Epigenetic Perspective on Lifestyle Medicine for Depression: Implications for Primary Care Practice. Am. J. Lifestyle Med. 2022, 16, 76–88. [Google Scholar] [CrossRef]
- Martins De Carvalho, L.; Chen, W.-Y.; Lasek, A.W. Epigenetic Mechanisms Underlying Stress-Induced Depression. In International Review of Neurobiology; Elsevier, 2021; Vol. 156, pp 87–126. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



