
Submitted:
29 March 2024
Posted:
29 March 2024
You are already at the latest version
Abstract
Abstract: Colistin is a last-resort antimicrobial for treating multidrug-resistant Gram-negative bacteria. Phe-notypic colistin resistance is highly associated with plasmid-mediated mobile colistin resistance (mcr) genes. mcr-bearing Enterobacteriaceae have been detected in many countries, with the emergence of colistin-resistant pathogens a global concern. This study assessed the distribution of mcr-1, mcr-2, mcr-3, mcr-4, and mcr-5 genes with the phenotypic colistin resistance in isolates from diarrheal infants and children in Bangladesh. Bacteria were identified using the API-20E biochemical panel and 16s rDNA gene sequencing. Polymerase chain reac-tions detected mcr gene variants in the isolates. Their susceptibilities to colistin were determined by agar dilu-tion and E-test by minimal inhibitory concentration (MIC) measurements. Over 30.0% (69/225) of isolates showed colistin resistance by agar dilution assessment (MIC> 2.0 μg/mL). Overall, 15.5% of isolates carried mcr genes (7, mcr-1; 17, mcr-2; 13, mcr-3; and co-occurrence occurred in 2 isolates). Clinical breakout MIC val-ues (≥ 4 μg/mL) were associated with 91.3% of mcr-positive isolates. The mcr-positive pathogens include twenty Escherichia spp., five Shigella flexneri, five Citrobacter spp., two Klebsiella pneumoniae, and three Pseudo-monas parafulva. mcr-genes appeared to be significantly associated with phenotypic colistin resistance phe-nomena (p=0.000), with 100% colistin-resistant isolates showing MDR phenomena. Age and sex of patients showed no significant association with detected mcr variants. Overall, mcr-associated colistin-resistant bacte-ria have emerged in Bangladesh, which warrants further research to determine their spread and instigate ac-tivities to reduce resistance.
Keywords:
Mobile colistin-resistance
; mcr gene
; human-mcr
; diarrheal infant patients
; Bangladesh
; MDR
; antimicrobial stewardship programs
1. Introduction
In recent years, emerging antimicrobial resistance (AMR) has been recognized as a significant public health concern that transcends international boundaries [1]. In 2019, it was estimated there were 4.95 million deaths globally associated with bacterial AMR, including 1.27 million deaths directly attributable to bacterial AMR, with the highest mortality currently seen in South Asian and sub-Saharan African countries [2]. There is also considerable morbidity and costs associated with AMR [3,4,5,6]. If AMR is not adequately tackled, this could reduce gross domestic product per country by up to 3.8% [5,6]. As a result, AMR is increasingly seen as the next pandemic unless urgent measures are introduced to reverse the rising rate [7]. The overuse and misuse of antibiotics either prophylactically or therapeutically in either humans or animal husbandry generates selection pressure, increasing the development of AMR in pathogens and other diverse commensal microbial populations [8,9,10,11]. Whilst AMR is a universal phenomenon, the burden among low- and middle-income countries (LMICs) is appreciably higher due to economic, political, and environmental factors, including poor governance and infrastructures, as well as a limited number of national initiatives [12,13,14,15]. This is now changing with increasing recognition of the clinical and economic consequences of AMR. Global initiatives include the Global Action Plan (GAP) by the World Health Organization (WHO) to reduce AMR [1], as well as initiatives from the World Bank and OECD [6,16]. The GAP has been translated into the National Action Plans in Bangladesh, with countries at different stages of their implementation due to resource and other issues [17,18,19,20].
Other important global initiatives include dividing antibiotics into three different categories based on their resistance potential, which includes the ‘Access’, ‘Watch’, and ‘Reserve’ categories [21,22]. ‘Access’ antibiotics should typically be prescribed to treat commonly encountered infections with lower resistance rates and include penicillins. Those in the ‘Watch’ group, which includes azithromycin, quinolones, and a number of cephalosporins, should ideally only be prescribed in critical conditions as they have a greater chance of resistance development. Antibiotics in the ‘Reserve’ category, which include fifth-generation cephalosporins, some carbapenems, and linezolid, should only prescribed in multidrug resistance cases, with the aim of curbing rising AMR rates [21,23,24,25]. Reducing the extent of ‘Watch’ antibiotics being prescribed or dispensed can help to appreciably reduce the extent of multidrug-resistant organisms [26]. More recently, the WHO has launched the AWaRe book, giving prescribing guidance for approximately 35 clinical infections, including the choice of drug [24,27], dose, and duration for both children and adults, to reduce the extent of inappropriate antibiotic use. The target is that at least 60% of antibiotic use in given settings should be ‘Access’ antibiotics [23,24].
Colistin is also classified as a ‘Reserve’ antibiotic following the global increase in the prevalence of carbapenem-resistant Enterobacteriaceae, [22,28]. However, there are now concerns with its over use and resultant resistance development [29,30]. This includes in animals where there has been appreciable use of colistin as a growth promoter in recent years [30], with studies now showing that 96% or more of total worldwide use of colistin is still in poultry and pig farming [31,32]. Concerns with the overuse of colistin including both animals and humans, and resultant resistance development via zoonotic gene transfers, coupled with its importance in treating resistant gram-negative infections to reduce morbidity and mortality, has resulted in the World Health Organization and others classifying colistin as an antibiotic of very high importance for use in humans with its use reserved [21,33,34,35,36,37,38,39]. Alongside this, many countries now ban the use of colistin as a growth promoter in animal feeds and prophylactically to prevent bacterial infections [40,41,42]. Such measures have shown to be effective in reducing resistant strains [41,42,43]. This is important in Bangladesh, given extensive colistin-resistant Escherichia coli in broiler meat and chicken feces [44,45,46], exacerbating resistance among patients to colistin in Bangladesh [47,48,49,50]. Over-the-counter dispensing of antibiotics is also common in Bangladesh and a concern, especially when this involves ‘Watch’ and ‘Reserve’ antibiotics [51,52,53,54]
Overall, activities to enhance the appropriate use of colistin in both animals and humans are essential as colistin still remains the antibiotic of choice for multiple drug-resistant gram-negative bacterial infections (MDR-GNB). This includes carbapenem-resistant Acinetobacter baumannii (CRAB) as well as other pathogens resistant to the new antimicrobial agents [55,56,57]. However, the use of colistin as a last resort antibiotic is greatly threatened by the rise of plasmid-borne mobile colistin resistance gene [58,59,60], spreading rapidly via horizontal gene transfer [61]. Resistance to colistin is generated by the chromosomally mediated modification of lipopolysaccharide (LPS) [62]. Acquisition of colistin resistance by a novel plasmid-mediated gene, mcr-1, was first described in Enterobacteriales from both farm-animal products and humans [63]. Earlier studies have shown the genotypic linkage of the mobile colistin resistance genes, mcr-1, to the phenotypic colistin resistance [64,65]. Since then, the variants of mcr-carrying multiple species of Enterobacteriales have been detected in many countries from environments, animals, and humans [66,67,68,69]. Subsequently, more variants of transferable colistin resistance mcr gene (mcr-1 to mcr-9) have been described in Enterobacteriaceae [70,71].
In general, a substantial part of Enterobacteriales is normal flora; however, a few of those microbial pathogens can cause systemic bacteremia [72], community-acquired infections [73,74], and healthcare-associated infections (HAI) [75]. A recent outbreak with colistin-resistant pathogens in China ended a very high case-fatality rate in humans [76]. Consequently, the identification of the root cause, transmission, and trajectories of colistin-resistant infection is an increasing priority globally. We are aware that mcr-gene variants can be detected in the environment, animals, human fecal samples, and food products [77]. However, only a limited number of studies have also showed the dissemination of plasmids carrying these variants in infants with acute diarrhea [78]. This is important in Bangladesh, with diarrhea being a major cause of childhood mortality in the country, combined with the increasing prevalence of resistant genes in children exacerbated by the overuse of antibiotics [79,80,81].
Consequently, this study was designed to investigate different variants of the mcr gene and their association with colistin resistance among diarrheal pathogens in infants, children and adults in Bangladesh and the demographic factors associated with the identified mcr variants. The findings can be used to suggest future policies and initiatives where there are concerns.
2. Results
2.1. Study Patients
We collected a total of 179 diarrheal stool samples from infants, children, and adults in different locations in Bangladesh (Figure 1) throughout our study and isolated 228 distinct bacteria from 168 culture-positive diarrheal patients. The study patients comprised of 102 (57%) males and 77 (43%) females.
In eleven stool samples, no bacterial growth appeared. The majority of study patients were infants and children who needed hospitalization (admitted to Uttara Medical College, Dhaka), and the median and interquartile range (IQR) age was 1.17 (0.75-2.5) years. Additionally, 78.1% of patients were middle class, 21.1% were poor, and 0.8% were rich (Figure 2A). The duration of diarrhea among all patients ranged from 3- 40 days, and the mean duration (standard deviation) was 7.2± 4.78 days.
The ages of the patients were categorized into five groups, namely <1 year, 1-5 years, 6-10 years, 11-15, and >15 years. This revealed that disease occurrence was higher in the 1-5 years age group (48.0%). Figure 2B provides further information regarding the incidence of diarrhea associated with each age group. Among all the patients, 179 (100%) appeared with watery stools, 4 (2.2%) presented with abdominal cramps, 39 (21.8%) with vomiting, and 2 (1.10%) patients had blood in their stools (Figure 2C).
2.2. Identification of Diarrheal Pathogens and Their Phenotypic Colistin Susceptibility
Of the 228 isolates, 140 were categorized as Escherichia spp. (61.40%), 20 as Citrobacter spp. (8.77%), 18 as Klebsiella spp. (7.89%), 13 as Shigella flexneri (5.70%), 10 Enterobacter spp. (4.39%), and seven as Stenotrophomonas maltophi (3.07%). Figure 3 contains details of all the pathogens identified. One Proteus sp and two staphylococci were excluded from further study since colistin resistance is natural in Proteus and Staphylococcus species [82,83].
The agar dilution method determined the test bacteria as susceptible (S) when there is no growth at ≤2 μg/mL colistin sulfate concentrations and resistant (R) when growth appeared at >2 μg/mL. In addition, the disc diffusion method was used to evaluate the antibacterial potency of colistin sulfate (25 µg) in vitro for the 225 isolates, and bacteria were considered colistin-resistant (R) if ≤10 mm diameter zone of inhibition was recorded. Of the 225 isolates, the agar dilution test revealed that 69 (30.7%) isolates were resistant to colistin sulfate. However, the colistin disc diffusion method showed that 180 (80.7%) isolates were resistant. No significant relationship was observed between agar dilution and the disc diffusion method.
2.3. Prevalence of mcr Genes in Diarrheal Isolates
All 225 isolates were subjected to polymerase chain reaction (PCR) to find mcr-1 to mcr-5 genes. Three types of mcr genes (mcr-1, mcr-2, mcr-3) were detected in 35 isolates. These included 20 for Escherichia spp., five for Shigella flexneri, five for Citrobacter spp. and two Klebsiella pneumoniae and Enterobacter hormaechei, and one Pseudomonas parafulva. Bacteria identified from ten other genera did not appear with mcr genes (Table 1).
Of the 35 mcr positive isolates, co-occurrence was identified in 2 isolates, one contained mcr-1, mcr-2 and the other contained mcr-2, mcr-3. The harborage of mcr-1, mcr-2 and mcr-3 were identified as 3.1% (7 isolates), 7.6% (17 isolates), 5.8% (13 isolates) respectively. Combined, the presence of mcr variants was 15.56% (35/225).
2.4. Phenotypic-Genotypic Association
Of the 35 mcr positive isolates, the agar dilution test identified 32 (91.4%) resistant isolates and 3 (8.6%) sensitive isolates, which revealed high statistical significance of mcr variants gene associations (p= 0.000). Further separate analyses showed very significantly high statistical associations of mcr-1, mcr-2, and mcr-3 with phenotypic colistin resistance (p = .000, for all the three gene variants) (Table 2). All of the test isolates grew well on the control plate without colistin sulfate. As a susceptible control, Escherichia coli ATCC25922 strain with MIC of 2 µg/mL was employed.
The MIC was determined by E-test and agar dilution test separately using a range of ≤.5 µg/mL to >256 µg/mL. The median and IQR MIC for mcr-positive isolates was 32.0 (8.0-128) µg/mL (Table 3). One isolate (Escherichia coli) with co-carriage of mcr-1 and mcr-2 exhibited MIC values of 128 µg/mL. The other co-carrying bacterium (Escherichia coli) with mcr-2 and mcr-3 showed a MIC value of 8 µg/mL. The three mcr-positive bacteria that showed phenotypic susceptibilities to colistin-sulfate were Citrobacter portucalensis, Citrobacter freundii, and Escherichia coli (Table 3). Alongside, the median and IQR MIC for mcr-negative isolates was 1.0 (0.5-2.0) µg/mL. Whilst 20% of the mcr-negative isolates (38/190) exhibited resistance to colistin sulfate in agar dilution with the MIC range from 4 µg/mL to >128 µg/mL (Figure 4).
MIC analyses at each specific value between mcr-positive and mcr-negative isolates showed that most mcr-carrying isolates were identified with higher MIC levels of colistin sulfate ranging from >8 µg/mL to >128 µg/mL (Figure 4). At the same time, the vast majority of the mcr-negative isolates exhibited lower levels MIC, ranging from ≤.5 µg/mL to 2.0 µg/mL. A fraction of mcr-negative isolates demonstrated MIC values from 32 µg/mL to >128 µg/mL (Figure 4).
2.5. Multi-Drug Resistance and mcr-Carriage
All mcr-positive isolates (carrying at least one mcr gene) were examined for the presence of MDR. The disk diffusion test was conducted to identify whether the 35 mcr-positive isolates exhibited susceptibility to the other 17 different antibiotics from 8 different groups or not. Remarkably, 100% of mcr-positive organisms exhibited MDR outcomes (Table 4), and mcr-negative isolates revealed a comparatively lower frequency of MDR outcomes.
2.6. Demographic Factors Associated with the mcr-Carriage
More than one type of bacterial isolate was isolated and analyzed from each stool sample. Consequently, 130 bacteria were evaluated from 95 male stool samples and the other 95 from 73 female stool samples. Bacteria isolated from female stool samples carried more mcr genes (16.8%, 16/95) in comparison to male-origin isolates (14.6%, 19/130). In the five different age groups of the study population, almost uniform gene distributions were reported. Overall, the results showed that sex and age groups were not significantly associated with the presence of mcr-1, mcr-2, and mcr-3 (p= 0.445 to 0.781) (Table 5).
3. Discussion
We believe this is one of the first studies in Bangladesh to investigate colistin-resistant genes in human diarrheal pathogens among infants and children. Our data clearly showed the association of phenotypic colistin resistance and the mcr genes (mcr-1 to mcr-5), similar to other published studies on colistin resistant bacteria isolated from diarrhea [78,84,85]. This builds on earlier studies in Bangladesh including among children and adults [68,86].
Isolates harboring mcr genes have been detected with high MIC values, showcasing disparities between the agar dilution test and the disk diffusion method [87]. This variation can be attributed to the slow diffusion of colistin disks on agar medium for the complex and large molecular structure of colistin sulfate. In parallel, some mcr-negative isolates exhibited resistance to colistin. Several potential reasons may account for this phenomenon. Firstly, mutations in the mgrB gene [88], responsible for binding polymyxin antibiotics in the gram-negative cell wall, are particularly prevalent in Klebsiella pneumoniae [89,90]. Secondly, the absence of testing for other variants of mcr genes including mcr-6, mcr-7, mcr-8, mcr-9, and mcr-10, which could also attribute phenotypic colistin resistance [91,92]. Thirdly, some resistant bacteria may develop capsules, a polysaccharide coating on the outer surface of the cell wall [93,94,95]. Fourthly, overexpression of efflux pump systems could contribute to resistance development [96,97]. Fifthly, modulation in the bacterial cell surface, including alterations in the structure of lipopolysaccharides (LPS) of the cell membrane, might affect the binding of polymyxin antibiotics [98]. Additional investigations will be necessary to uncover potential molecular explanations for the observed differences between phenotypic and genotypic colistin resistance.
In this study, male children had a higher incidence rate of diarrhea than female children, similar to previous studies [99,100]. However, the reason for this difference is unclear. We also found that 57.14% of mcr-positive isolates were resistant to amoxicillin/clavulanic acid, and 57 to 71% of mcr-carrying isolates resistant to all generations of cephalosporins, although higher generation were more susceptible. This is a concern with clinicians now prescribing more carbapenems due to the decreased potency of cephalosporins, with meropenem showing more susceptibility among the 17 different antibiotics in the eight groups studied. However, recent studies showed carbapenem-resistant Enterobacteriaceae are now a global threat [101,102,103]. The MDR status of all mcr-positive isolates is positive, which is a threat to public health in Bangladesh and beyond, given the ensuing rise in untreatable infectious diseases [104,105,106].
This study underscores the clinical significance of establishing comprehensive surveillance systems for priority antibiotics such as colistin. In addition, we urge the Government of Bangladesh to ban the use of colistin as a growth promoter in animal feeds and prophylactically to prevent bacterial infections, similar to other countries [40,41,42]. This is because such measures have been shown to be effective in reducing resistant strains [41,42,43]. Alongside, this instigates antimicrobial stewardship programs (ASPs) in ambulatory care similar to other important antibiotics in Bangladesh where there are concerns about resistance development [107,108,109,110]. This includes ASPs among community pharmacists and drug sellers building on ASPs in other sectors in Bangladesh [52,111].
We are aware that there are several limitations with this study. Firstly, it was challenging to accurately estimate the real-world scenario since adults experiencing diarrheal problems could readily seek treatment at a healthcare center, whereas children relied on their parents’ decisions and assistance to access medical care. Secondly, the small sample size posed significant obstacles to conducting fully powered statistical analyses. However, our research spanned fifteen distinct districts across Bangladesh, and the outcomes are anticipated to be applicable if similar studies are conducted in other districts in the country. Thirdly, while this study examined mcr gene variants up to mcr-5, newer variants such as mcr-6, mcr-7, mcr-8, and mcr-9 were not investigated. Having said this, efforts were made to maintain internal validity by conducting independent trials when necessary. Despite these limitations, we believe our findings are robust, providing guidance to all key stakeholders in Bangladesh to enhance future sensitivity to colistin.
4. Materials and Methods
4.1. Study Design and Sampling
A prospective cross-sectional study was conducted between January 2020 and December 2020 among diarrheic children visiting the outpatient Department of Uttara Adhunik Medical College Hospital, Dhaka, Bangladesh. A total of 179 children and adults having acute diarrhea participated in this study prior to treatment with any prescribed antibiotics.
Acute diarrhea was defined as three or more liquid, loose, mucus, or bloody stools within 24 h, lasting no longer than 14 days. Fever was defined as a temperature of ≥37.5 °C. Demographic data was taken from each child, and informed consent was obtained from the parents or guardians before sample collection. All relevant demographic, clinical, and laboratory data were recorded and transferred to the questionnaire prepared for this study.
The ages of the children and patients were categorized into five groups, namely <1 year, 1-5 years, 6-10 years, 11-15, and >15 years, based on previous studies [112]. The income of parents was classified into rich, middle class, or poor, and as we are aware, this can make a difference [113].
A sterilized cotton swab was dipped in the mucus, purulent or bloody part of the freshly passed stool sample, placed immediately in CaryBlair Medium (Oxoid, Hampshire, UK), and transported to the laboratory for further analysis within six hours of collection.
4.2. Isolation and Identification of Bacteria
Collected samples were pre-enriched in buffered peptone water (Oxoid®, Hampshire, UK) at a dilution ratio of 1:10 and were incubated overnight at 37 ◦C. A loopful of each culture was streaked on MacConkey agar (Liofilchem Inc, Italy) and cysteine-, lactose-, and electrolyte-deficient (CLED) agar (Liofilchem Inc, Italy) and subsequently incubated at 37 ◦C for 24 h in aerobic condition simultaneously. MacConkey agar supports gram-negative diarrheal pathogens (Supplementary Figure S1A), while CLED agar aids in the growth of gram-negative bacteria and gram-positive cocci if present in diarrheal samples. Colony counts of 103 or 105 CFU/mL were considered for a cut-off value for a probable diarrheal sample [114].
Gram’s staining and biochemical tests were initially performed to identify growth-positive bacteria. A rapid biochemical-test kit API 20E (BioMe´rieux, Durham, NC), consisting of carbohydrate batteries and enzymatic substrates in a set of chromogenic panels, was used to verify the isolated identity (Supplementary Figure S1B) [115]. A part of the bacterial identity was confirmed by the polymerase chain reaction (PCR) amplification and sequencing of the 16S rDNA gene [116]. In total, 228 different isolates were generated from all the diarrheal samples. Three bacteria (one Proteus and two staphylococci) were excluded from the study for the next level analyses since colistin resistance is a natural phenomenon of the excluded isolates. The remaining 225 isolates were subjected to assessment of colistin susceptibility and mcr-1 to mcr-5 carriage. The isolates were preserved in 30% glycerol in Trypticase Soy Broth (TSB) at – 80°C until further use.
4.3. Phenotypic Colistin Susceptibility Testing
The phenotypic antibiogram profiles of diarrheal isolates against colistin were determined primarily using the Kirby–Bauer disk diffusion method according to the European Committee on Antimicrobial Susceptibility Testing (EUCAST) and Clinical & Laboratory Standards Institute (CLSI) guidelines (Supplementary Figure S1C) [117,118]. A 3-hour bacterial suspension in Mueller–Hinton broth was prepared to a concentration of McFarland 0.5 equivalent and then streaked on Mueller–Hinton agar (MHA, Oxoid, Basingstoke, UK) plates using a cotton swab to ensure consistent growth. The susceptibility of the isolate to the following discs of antibiotics (Oxoid, Basingstoke, UK) was evaluated: colistin (25 µg), Amoxycillin + Clavulanic acid (30 µg), Cefuroxime Sodium (30 µg), Cefixime (30 µg), Cefepime (30 µg), Imipenem (10 µg), Meropenem (10 µg), Nalidixic acid (10 µg), Ciprofloxacin (5 µg), Lomefloxacin (10 µg), levofloxacin (5 µg), Gentamicin (30 µg), Amikacin (30 µg), and Netilmicin (30 µg), Tobramycin (10 µg), Nitrofurantoin(300 µg), Trimethoprim-sulfamethoxazole (25 µg); by placing them on the bacterial lawns and incubating at 37°C overnight. A clear zone was developed around the disc for sensitive bacteria, and zone diameter was measured and evaluated to categorize Bacteria as susceptible (S), intermediate (I), and resistant (R) from the CLSI guideline charts for the appropriate antibiotics tested [118].
The isolates’ phenotypic antimicrobial susceptibilities were further tested by the agar dilution method [119]. The lowest concentration of colistin adequate to inhibit the visible growth of bacterial test isolates was determined by minimal inhibitory concentration (MIC) measurement by agar dilution method [119]. For agar dilution MIC, different concentrations of colistin-sulfate powder (Santa Cruz Biotechnology Inc, TX) from .50 µg/mL to 256.0 µg/mL in a two-fold dilution order were incorporated into MHA medium accordingly. One pure culture colony was inoculated into Mueller-Hinton broth to prepare each test inoculum and incubated for three hours at 37°C that develops a density of inoculum equivalent to 104 colony-forming units (CFU) per spot to drop on the MHA. The inoculum density was periodically compared to a 0.5 McFarland standard, equivalent to approximately 108 CFU/mL. The plates were incubated at 37°C in the air for 18-20 hours. Agar dilution MICs were performed in duplicates. The experiments were repeated when some single colonies or a thin haze growth was observed within the inoculated spot.
The epsilometer test (E-test) was performed parallel partly using a commercial strip containing a predefined gradient of colistin concentrations (Liofilchem Inc, Italy) to validate colistin MIC determination by the agar dilution method [87,120]. Concordant results were found in independent MIC assessment and E-test (Supplementary Figure S1D,E). E. coli ATCC 25922 strain was used as the quality control strain for disc diffusion and MIC testing. Besides, a control plate without colistin-sulfate was examined for the growth of both test and control isolates. Following EUCAST and CLSI guidelines, isolates were considered susceptible (S) when the MIC values exhibited ≤2 μg/mL and resistant (R) when MICs appeared >2 μg/mL [117]. Multidrug-resistant (MDR) isolates were described as those isolates that were found to be resistant to at least three different classes of antibiotics [121].
4.4. Detection of the Colistin Resistance mcr Genes
All 225 isolates were subjected to a single plex polymerase chain reaction (PCR) to detect the mcr-1 gene, yielding a 309 bp DNA band, using primers described elsewhere [63], and confirmed by sequencing. Amplicons were visualized under UV light after 1.2% agarose gel electrophoresis. The other four primer pairs to detect mcr-2, mcr-3, mcr-4, and mcr-5 gene amplicons were obtained from a recently published original study [122]. Multiplex polymerase chain reaction (PCR) was conducted to detect the mcr-1 to mcr-5 genes in the isolates. In brief, the modified protocol was as follows: prepared bacterial DNA (2.0 μL) was added to a 2× PCR premixture (15 μL, GeneON, Germany), and five pmol of each primer (1 μL), and deionized water was added to obtain a final volume of 30 μL. Reactions went through an initial denaturation at 94 °C for 15 min followed by 25 cycles of amplification (Applied Biosystems 2720 Thermal Cycler, Singapore), consisting of denaturation for 30 s at 94 °C, annealing for 90 s at 55 °C, and extension for 1 min at 72 °C, and a final 10 min elongation at 72 °C. Expected amplicons for mcr-1 (309 bp), mcr-2 (715 bp), mcr-3 (929 bp), mcr-4 (1116 bp), and mcr-5 (1644 bp) underwent electrophoresis through 1.2% agarose gel followed by staining with ethidium bromide and were visualized under UV light (Supplementary Figure S1E). Lastly, the obtained results were validated by separate single plex PCR analyses of the mcr genes.
4.5. Statistical Analysis
Using IBM SPSS statistics data editor (version 21) and GraphPad prism software (version 9.5), verified data were entered and then examined. The bivariate analysis did not include missing data. The mcr gene variations carried by diarrheal pathogens and their phenotypic traits were described using both descriptive and inferential statistical methods. Any associations between categorical data were examined using Pearson’s chi-square test, with the appropriate use of Yate’s continuity correction. Fisher’s Exact test results of a 2 x 2 contingency table were presented in place of the chi-square test results if the predicted frequency of the test cannot be assumed. Two-tailed p-values were computed, with a significance level of 0.05.
4.6. Ethics Statements
This study was authorized [No. UAMC/ERC/Recommend-62/2018, dated 09.07.2018] by the Ethics Review Committee of Uttara Adhunik Medical College. All research protocols complied with the Declaration of Helsinki regarding the use of human beings in research.
Each adult study participant provided written informed consent before the collection of their urine samples. For patients under the age of 18, parents or legal guardians were separately asked for written informed permission. Patient’s identities were anonymized.
5. Conclusions
Our findings indicate that multidrug-resistant pathogenic bacteria containing mcr genes are a major reservoir in the guts of young Bangladeshi children and adults. The mcr-1, mcr-2 and mcr-3 variants predominate in the Bangladeshi diarrheal bacteria over other variants, such as mcr-4 and mcr-5. We did not find any association between phenotypic colistin resistance and age, sex.
The advent of clinical MDR pathogens resistant to colistin intended as an antibiotic for last resort may spread diseases and illnesses that are subsequently incurable. The findings require immediate monitoring and action for both national and international antimicrobial stewardship. This includes limiting the use of colistin as a growth promoter agent in animal husbandry in Bangladesh, similar to other countries. This especially as AMR is increasingly being transferred from animals to people as a result of improper antibiotic use in animal feeding. There also needs to be increased patient education to address hygiene levels as well as seek measures to improve the availability of safe drinking water in the country. Alongside this, ASPs aim to help physicians, pharmacists, and drug sellers reduce the inappropriate prescribing and dispensing of colistin. We will continue to monitor the situation.
Supplementary Materials
The following supporting information on detailed procedures for bacterial isolation, antibiotic susceptibility testing, and mcr detection via Polymerase Chain Reaction (PCR), as outlined in Supplementary Figure S1.
Author Contributions
S.S and R.M.N.: Sample collection, Methodology, Investigation, Formal analysis, and Manuscript drafting; M.B.H: Data acquisition, Investigation, Data validation, Visualization; U.L.U and M.A.A.: Methodology, Data curation, Writing-Reviewing, Validation and Editing; A.S.M.M and M.R.K.K: Clinical and demographic data acquisition, Resources, Conceptualization, Visualization and Validation; SN: Project administration, Resources, Methodology, Validation, and Visualization; B.G.: Conceptualization, Methodology, Re-writing and Editing, Visualization; SI: Conceptualization, Supervision, Resources, Data curation and analysis, Writing-Reviewing, and Editing, Study coordination. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
This study was authorized [No. UAMC/ERC/Recommend-62/2018, dated 09.07.2018] by the Ethics Review Committee of Uttara Adhunik Medical College, Dhaka, Bangladesh.
Informed Consent Statement
Informed consent was obtained from parents before the start of sample collection.
Data Availability Statement
Data is contained within the article and available upon request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- WHO. GLOBAL ACTION PLAN ON ANTIMICROBIAL RESISTANCE. 2015. Available online: (accessed on https://apps.who.int/iris/bitstream/handle/10665/193736/9789241509763_eng.pdf?sequence=1.
- The burden of bacterial antimicrobial resistance in the WHO European region in 2019: a cross-country systematic analysis. Lancet Public Health 2022, 7, e897–e913. [CrossRef]
- Cassini, A.; Högberg, L.D.; Plachouras, D.; Quattrocchi, A.; Hoxha, A.; Simonsen, G.S.; Colomb-Cotinat, M.; Kretzschmar, M.E.; Devleesschauwer, B.; Cecchini, M.; et al. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: a population-level modelling analysis. Lancet Infect Dis 2019, 19, 56–66. [Google Scholar] [CrossRef]
- Dadgostar, P. Antimicrobial Resistance: Implications and Costs. Infect Drug Resist 2019, 12, 3903–3910. [Google Scholar] [CrossRef]
- Hofer, U. The cost of antimicrobial resistance. Nat Rev Microbiol 2019, 17, 3. [Google Scholar] [CrossRef] [PubMed]
- World Bank Group. Pulling Together to Beat Superbugs Knowledge and Implementation Gaps in Addressing Antimicrobial Resistance. 2019. . Available online: (accessed on.
- Gautam, A. Antimicrobial Resistance: The Next Probable Pandemic. JNMA J Nepal Med Assoc 2022, 60, 225–228. [Google Scholar] [CrossRef]
- Hossain, M.J.; Jabin, N.; Ahmmed, F.; Sultana, A.; Abdur Rahman, S.; Islam, M.R. Irrational use of antibiotics and factors associated with antibiotic resistance: findings from a cross-sectional study in Bangladesh. Health science reports 2023, 6, e1465. [Google Scholar] [CrossRef]
- Akram, F.; Imtiaz, M.; ul Haq, I. Emergent crisis of antibiotic resistance: A silent pandemic threat to 21st century. Microbial Pathogenesis 2023, 174, 105923. [Google Scholar] [CrossRef]
- Llor, C.; Bjerrum, L. Antimicrobial resistance: risk associated with antibiotic overuse and initiatives to reduce the problem. Ther Adv Drug Saf 2014, 5, 229–241. [Google Scholar] [CrossRef]
- Van, T.T.H.; Yidana, Z.; Smooker, P.M.; Coloe, P.J. Antibiotic use in food animals worldwide, with a focus on Africa: Pluses and minuses. Journal of Global Antimicrobial Resistance 2020, 20, 170–177. [Google Scholar] [CrossRef]
- Sulis, G.; Sayood, S.; Gandra, S. Antimicrobial resistance in low- and middle-income countries: current status and future directions. Expert Rev Anti Infect Ther 2022, 20, 147–160. [Google Scholar] [CrossRef]
- Iskandar, K.; Molinier, L.; Hallit, S.; Sartelli, M.; Hardcastle, T.C.; Haque, M.; Lugova, H.; Dhingra, S.; Sharma, P.; Islam, S.; et al. Surveillance of antimicrobial resistance in low- and middle-income countries: a scattered picture. Antimicrob Resist Infect Control 2021, 10, 63. [Google Scholar] [CrossRef] [PubMed]
- Collignon, P.; Athukorala, P.C.; Senanayake, S.; Khan, F. Antimicrobial resistance: the major contribution of poor governance and corruption to this growing problem. PLoS One 2015, 10, e0116746. [Google Scholar] [CrossRef]
- Ayukekbong, J.A.; Ntemgwa, M.; Atabe, A.N. The threat of antimicrobial resistance in developing countries: causes and control strategies. Antimicrob Resist Infect Control 2017, 6, 47. [Google Scholar] [CrossRef]
- Fernandes, M.R.; Ignacio, A.; Rodrigues, V.A.; Groppo, F.C.; Cardoso, A.L.; Avila-Campos, M.J.; Nakano, V. Alterations of Intestinal Microbiome by Antibiotic Therapy in Hospitalized Children. Microb Drug Resist 2017, 23, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Jouy, E.; Haenni, M.; Le Devendec, L.; Le Roux, A.; Chatre, P.; Madec, J.Y.; Kempf, I. Improvement in routine detection of colistin resistance in E. coli isolated in veterinary diagnostic laboratories. Journal of microbiological methods 2017, 132, 125–127. [Google Scholar] [CrossRef] [PubMed]
- Chua, A.Q.; Verma, M.; Hsu, L.Y.; Legido-Quigley, H. An analysis of national action plans on antimicrobial resistance in Southeast Asia using a governance framework approach. Lancet Reg Health West Pac 2021, 7, 100084. [Google Scholar] [CrossRef] [PubMed]
- Willemsen, A.; Reid, S.; Assefa, Y. A review of national action plans on antimicrobial resistance: strengths and weaknesses. Antimicrob Resist Infect Control 2022, 11, 90. [Google Scholar] [CrossRef]
- Godman, B.; Egwuenu, A.; Wesangula, E.; Schellack, N.; Kalungia, A.C.; Tiroyakgosi, C.; Kgatlwane, J.; Mwita, J.C.; Patrick, O.; Niba, L.L.; et al. Tackling antimicrobial resistance across sub-Saharan Africa: current challenges and implications for the future. Expert Opinion on Drug Safety 2022, 21, 1089–1111. [Google Scholar] [CrossRef]
- Sharland, M.; Pulcini, C.; Harbarth, S.; Zeng, M.; Gandra, S.; Mathur, S.; Magrini, N. Classifying antibiotics in the WHO Essential Medicines List for optimal use-be AWaRe. Lancet Infect Dis 2018, 18, 18–20. [Google Scholar] [CrossRef]
- Sharland, M.; Gandra, S.; Huttner, B.; Moja, L.; Pulcini, C.; Zeng, M.; Mendelson, M.; Cappello, B.; Cooke, G.; Magrini, N. Encouraging AWaRe-ness and discouraging inappropriate antibiotic use-the new 2019 Essential Medicines List becomes a global antibiotic stewardship tool. Lancet Infect Dis 2019, 19, 1278–1280. [Google Scholar] [CrossRef]
- Klein, E.Y.; Milkowska-Shibata, M.; Tseng, K.K.; Sharland, M.; Gandra, S.; Pulcini, C.; Laxminarayan, R. Assessment of WHO antibiotic consumption and access targets in 76 countries, 2000-15: an analysis of pharmaceutical sales data. Lancet Infect Dis 2021, 21, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Sharland, M.; Zanichelli, V.; Ombajo, L.A.; Bazira, J.; Cappello, B.; Chitatanga, R.; Chuki, P.; Gandra, S.; Getahun, H.; Harbarth, S.; et al. The WHO essential medicines list AWaRe book: from a list to a quality improvement system. Clin Microbiol Infect 2022, 28, 1533–1535. [Google Scholar] [CrossRef] [PubMed]
- Hsia, Y.; Lee, B.R.; Versporten, A.; Yang, Y.; Bielicki, J.; Jackson, C.; Newland, J.; Goossens, H.; Magrini, N.; Sharland, M. Use of the WHO Access, Watch, and Reserve classification to define patterns of hospital antibiotic use (AWaRe): an analysis of paediatric survey data from 56 countries. Lancet Glob Health 2019, 7, e861–e871. [Google Scholar] [CrossRef] [PubMed]
- Sulis, G.; Sayood, S.; Katukoori, S.; Bollam, N.; George, I.; Yaeger, L.H.; Chavez, M.A.; Tetteh, E.; Yarrabelli, S.; Pulcini, C.; et al. Exposure to World Health Organization’s AWaRe antibiotics and isolation of multidrug resistant bacteria: a systematic review and meta-analysis. Clin Microbiol Infect 2022, 28, 1193–1202. [Google Scholar] [CrossRef]
- Zanichelli, V.; Sharland, M.; Cappello, B.; Moja, L.; Getahun, H.; Pessoa-Silva, C.; Sati, H.; van Weezenbeek, C.; Balkhy, H.; Simão, M.; et al. The WHO AWaRe (Access, Watch, Reserve) antibiotic book and prevention of antimicrobial resistance. Bull World Health Organ 2023, 101, 290–296. [Google Scholar] [CrossRef]
- Lu, Q.; Li, G.-H.; Qu, Q.; Zhu, H.-H.; Luo, Y.; Yan, H.; Yuan, H.-Y.; Qu, J. Clinical efficacy of polymyxin B in patients infected with carbapenem-resistant organisms. Infection and drug resistance 2021, 1979–1988. [Google Scholar] [CrossRef] [PubMed]
- Van Boeckel, T.P.; Gandra, S.; Ashok, A.; Caudron, Q.; Grenfell, B.T.; Levin, S.A.; Laxminarayan, R. Global antibiotic consumption 2000 to 2010: an analysis of national pharmaceutical sales data. The Lancet infectious diseases 2014, 14, 742–750. [Google Scholar] [CrossRef]
- Zhang, S.; Abbas, M.; Rehman, M.U.; Wang, M.; Jia, R.; Chen, S.; Liu, M.; Zhu, D.; Zhao, X.; Gao, Q.; et al. Updates on the global dissemination of colistin-resistant Escherichia coli: An emerging threat to public health. Sci Total Environ 2021, 799, 149280. [Google Scholar] [CrossRef] [PubMed]
- Umair, M.; Hassan, B.; Farzana, R.; Ali, Q.; Sands, K.; Mathias, J.; Afegbua, S.; Haque, M.N.; Walsh, T.R.; Mohsin, M. International manufacturing and trade in colistin, its implications in colistin resistance and One Health global policies: a microbiological, economic, and anthropological study. Lancet Microbe 2023, 4, e264–e276. [Google Scholar] [CrossRef]
- Cuong, N.V.; Kiet, B.T.; Hien, V.B.; Truong, B.D.; Phu, D.H.; Thwaites, G.; Choisy, M.; Carrique-Mas, J. Antimicrobial use through consumption of medicated feeds in chicken flocks in the Mekong Delta of Vietnam: A three-year study before a ban on antimicrobial growth promoters. PLoS One 2021, 16, e0250082. [Google Scholar] [CrossRef]
- Andrade, F.F.; Silva, D.; Rodrigues, A.; Pina-Vaz, C. Colistin Update on Its Mechanism of Action and Resistance, Present and Future Challenges. Microorganisms 2020, 8. [Google Scholar] [CrossRef]
- WHO. Critically Important Antimicrobials for Human Medicine. 6th Revision 2018. Ranking of medically important antimicrobials for risk management of antimicrobial resistance due to non-human use. Available online: (accessed on.
- EMA. Categorisation of antibiotics used in animals promotes responsible use to protect public and animal health. 2020. Available online: (accessed on.
- Talat, A.; Miranda, C.; Poeta, P.; Khan, A.U. Farm to table: colistin resistance hitchhiking through food. Arch Microbiol 2023, 205, 167. [Google Scholar] [CrossRef]
- Shen, Y.; Zhang, R.; Schwarz, S.; Wu, C.; Shen, J.; Walsh, T.R.; Wang, Y. Farm animals and aquaculture: significant reservoirs of mobile colistin resistance genes. Environmental Microbiology 2020, 22, 2469–2484. [Google Scholar] [CrossRef]
- Javed, H.; Saleem, S.; Zafar, A.; Ghafoor, A.; Shahzad, A.B.; Ejaz, H.; Junaid, K.; Jahan, S. Emergence of plasmid-mediated mcr genes from Gram-negative bacteria at the human-animal interface. Gut Pathogens 2020, 12, 1–9. [Google Scholar] [CrossRef]
- Yin, Y.; Qiu, L.; Wang, G.; Guo, Z.; Wang, Z.; Qiu, J.; Li, R. Emergence and Transmission of Plasmid-Mediated Mobile Colistin Resistance Gene mcr-10 in Humans and Companion Animals. Microbiology Spectrum 2022, 10, e02097–02022. [Google Scholar] [CrossRef]
- Mendelson, M.; Brink, A.; Gouws, J.; Mbelle, N.; Naidoo, V.; Pople, T.; Schellack, N.; van Vuuren, M.; Rees, H. The One Health stewardship of colistin as an antibiotic of last resort for human health in South Africa. The Lancet. Infectious diseases 2018, 18, e288–e294. [Google Scholar] [CrossRef]
- Ribeiro, S.; Mourão, J.; Novais, Â.; Campos, J.; Peixe, L.; Antunes, P. From farm to fork: Colistin voluntary withdrawal in Portuguese farms reflected in decreasing occurrence of mcr-1-carrying Enterobacteriaceae from chicken meat. Environ Microbiol 2021, 23, 7563–7577. [Google Scholar] [CrossRef]
- Usui, M.; Nozawa, Y.; Fukuda, A.; Sato, T.; Yamada, M.; Makita, K.; Tamura, Y. Decreased colistin resistance and mcr-1 prevalence in pig-derived Escherichia coli in Japan after banning colistin as a feed additive. Journal of Global Antimicrobial Resistance 2021, 24, 383–386. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, C.; Zhang, R.; Chen, Y.; Shen, Y.; Hu, F.; Liu, D.; Lu, J.; Guo, Y.; Xia, X.; et al. Changes in colistin resistance and mcr-1 abundance in Escherichia coli of animal and human origins following the ban of colistin-positive additives in China: an epidemiological comparative study. Lancet Infect Dis 2020, 20, 1161–1171. [Google Scholar] [CrossRef]
- Nath, C.; Das, T.; Islam, M.S.; Hasib, F.M.Y.; Singha, S.; Dutta, A.; Barua, H.; Islam, M.Z. Colistin Resistance in Multidrug-Resistant Escherichia coli Isolated from Retail Broiler Meat in Bangladesh. Microb Drug Resist 2023, 29, 523–532. [Google Scholar] [CrossRef]
- Islam, S.; Urmi, U.L.; Rana, M.; Sultana, F.; Jahan, N.; Hossain, B.; Iqbal, S.; Hossain, M.M.; Mosaddek, A.S.M.; Nahar, S. High abundance of the colistin resistance gene mcr-1 in chicken gut-bacteria in Bangladesh. Sci Rep 2020, 10, 17292. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.B.; Alam, M.N.; Hasan, M.; Hossain, S.M.B.; Debnath, M.; Begum, R.; Samad, M.A.; Hoque, S.F.; Chowdhury, M.S.R.; Rahman, M.M.; et al. Molecular Detection of Colistin Resistance mcr-1 Gene in Multidrug-Resistant Escherichia coli Isolated from Chicken. Antibiotics (Basel) 2022, 11. [Google Scholar] [CrossRef]
- Sonia, S.J.; Uddin, K.H.; Shamsuzzaman, S.M. Prevalence of Colistin Resistance in Klebsiella pneumoniae Isolated from a Tertiary Care Hospital in Bangladesh and Molecular Characterization of Colistin Resistance Genes among Them by Polymerase Chain Reaction and Sequencing. Mymensingh Med J 2022, 31, 733–740. [Google Scholar]
- Ara, B.; Urmi, U.L.; Haque, T.A.; Nahar, S.; Rumnaz, A.; Ali, T.; Alam, M.S.; Mosaddek, A.S.M.; Rahman, N.A.A.; Haque, M.; et al. Detection of mobile colistin-resistance gene variants (mcr-1 and mcr-2) in urinary tract pathogens in Bangladesh: the last resort of infectious disease management colistin efficacy is under threat. Expert Rev Clin Pharmacol 2021, 14, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Dutta, A.; Islam, M.Z.; Barua, H.; Rana, E.A.; Jalal, M.S.; Dhar, P.K.; Das, A.; Das, T.; Sarma, S.M.; Biswas, S.K.; et al. Acquisition of Plasmid-Mediated Colistin Resistance Gene mcr-1 in Escherichia coli of Livestock Origin in Bangladesh. Microb Drug Resist 2020, 26, 1058–1062. [Google Scholar] [CrossRef] [PubMed]
- Kawser, Z.; Shamsuzzaman, S.M. Association of Virulence with Antimicrobial Resistance among Klebsiella pneumoniae Isolated from Hospital Settings in Bangladesh. Int J Appl Basic Med Res 2022, 12, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Rousham, E.K.; Nahar, P.; Uddin, M.R.; Islam, M.A.; Nizame, F.A.; Khisa, N.; Akter, S.M.S.; Munim, M.S.; Rahman, M.; Unicomb, L. Gender and urban-rural influences on antibiotic purchasing and prescription use in retail drug shops: a one health study. BMC Public Health 2023, 23, 229. [Google Scholar] [CrossRef] [PubMed]
- Unicomb, L.E.; Nizame, F.A.; Uddin, M.R.; Nahar, P.; Lucas, P.J.; Khisa, N.; Akter, S.M.S.; Islam, M.A.; Rahman, M.; Rousham, E.K. Motivating antibiotic stewardship in Bangladesh: identifying audiences and target behaviours using the behaviour change wheel. BMC Public Health 2021, 21, 968. [Google Scholar] [CrossRef]
- Orubu, E.S.F.; Samad, M.A.; Rahman, M.T.; Zaman, M.H.; Wirtz, V.J. Mapping the Antimicrobial Supply Chain in Bangladesh: A Scoping-Review-Based Ecological Assessment Approach. Glob Health Sci Pract 2021, 9, 532–547. [Google Scholar] [CrossRef]
- Islam, M.A.; Akhtar, Z.; Hassan, M.Z.; Chowdhury, S.; Rashid, M.M.; Aleem, M.A.; Ghosh, P.K.; Mah-E-Muneer, S.; Parveen, S.; Ahmmed, M.K.; et al. Pattern of Antibiotic Dispensing at Pharmacies According to the WHO Access, Watch, Reserve (AWaRe) Classification in Bangladesh. Antibiotics 2022, 11, 247. [Google Scholar] [CrossRef]
- Giacobbe, D.R.; Bassetti, M.; De Rosa, F.G.; Del Bono, V.; Grossi, P.A.; Menichetti, F.; Pea, F.; Rossolini, G.M.; Tumbarello, M.; Viale, P. Ceftolozane/tazobactam: place in therapy. Expert review of anti-infective therapy 2018, 16, 307–320. [Google Scholar] [CrossRef]
- Pogue, J.M.; Bonomo, R.A.; Kaye, K.S. Ceftazidime/avibactam, meropenem/vaborbactam, or both? Clinical and formulary considerations. Clinical Infectious Diseases 2019, 68, 519–524. [Google Scholar] [CrossRef]
- Van Duin, D.; Lok, J.J.; Earley, M.; Cober, E.; Richter, S.S.; Perez, F.; Salata, R.A.; Kalayjian, R.C.; Watkins, R.R.; Doi, Y. Colistin versus ceftazidime-avibactam in the treatment of infections due to carbapenem-resistant Enterobacteriaceae. Clinical Infectious Diseases 2018, 66, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-H.; Liu, Y.-Y.; Shen, Y.-B.; Yang, J.; Walsh, T.R.; Wang, Y.; Shen, J. Plasmid-mediated colistin-resistance genes: mcr. Trends in Microbiology 2023. [Google Scholar] [CrossRef] [PubMed]
- Phuadraksa, T.; Wichit, S.; Songtawee, N.; Tantimavanich, S.; Isarankura-Na-Ayudhya, C.; Yainoy, S. Emergence of plasmid-mediated colistin resistance mcr-3.5 gene in Citrobacter amalonaticus and Citrobacter sedlakii isolated from healthy individual in Thailand. Frontiers in Cellular and Infection Microbiology 2023, 12, 1917. [Google Scholar] [CrossRef]
- Zelendova, M.; Papagiannitsis, C.C.; Sismova, P.; Medvecky, M.; Pomorska, K.; Palkovicova, J.; Nesporova, K.; Jakubu, V.; Jamborova, I.; Zemlickova, H. Plasmid-mediated colistin resistance among human clinical Enterobacterales isolates: National surveillance in the Czech Republic. Frontiers in Microbiology 2023, 14, 1147846. [Google Scholar] [CrossRef]
- Wang, R.; van Dorp, L.; Shaw, L.P.; Bradley, P.; Wang, Q.; Wang, X.; Jin, L.; Zhang, Q.; Liu, Y.; Rieux, A.; et al. The global distribution and spread of the mobilized colistin resistance gene mcr-1. Nature communications 2018, 9, 1179. [Google Scholar] [CrossRef]
- Gogry, F.A.; Siddiqui, M.T.; Sultan, I.; Haq, Q.M. Current update on intrinsic and acquired colistin resistance mechanisms in bacteria. Frontiers in Medicine 2021, 1250. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. The Lancet. Infectious diseases 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Ćwiek, K.; Woźniak-Biel, A.; Karwańska, M.; Siedlecka, M.; Lammens, C.; Rebelo, A.R.; Hendriksen, R.S.; Kuczkowski, M.; Chmielewska-Władyka, M.; Wieliczko, A. Phenotypic and genotypic characterization of mcr-1-positive multidrug-resistant Escherichia coli ST93, ST117, ST156, ST10, and ST744 isolated from poultry in Poland. Brazilian Journal of Microbiology 2021, 52, 1597–1609. [Google Scholar] [CrossRef]
- Elias, R.; Spadar, A.; Phelan, J.; Melo-Cristino, J.; Lito, L.; Pinto, M.; Gonçalves, L.; Campino, S.; Clark, T.G.; Duarte, A. A phylogenomic approach for the analysis of colistin resistance-associated genes in Klebsiella pneumoniae, its mutational diversity and implications for phenotypic resistance. International Journal of Antimicrobial Agents 2022, 59, 106581. [Google Scholar] [CrossRef]
- Karim, M.R.; Zakaria, Z.; Hassan, L.; Faiz, N.M.; Ahmad, N.I. The occurrence and molecular detection of mcr-1 and mcr-5 genes in Enterobacteriaceae isolated from poultry and poultry meats in Malaysia. Frontiers in microbiology 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Rebelo, A.; MH, C.L.; Bortolaia, V.; Kjeldgaard, J.; Hendriksen, R. PCR for plasmid-mediated colistin resistance genes, mcr-1, mcr-2, mcr-3, mcr-4, mcr-5 and variants (multiplex), on DTU National Food Institute. 2018.
- Johura, F.-T.; Tasnim, J.; Barman, I.; Biswas, S.R.; Jubyda, F.T.; Sultana, M.; George, C.M.; Camilli, A.; Seed, K.D.; Ahmed, N. Colistin-resistant Escherichia coli carrying mcr-1 in food, water, hand rinse, and healthy human gut in Bangladesh. Gut pathogens 2020, 12, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Perdomo, A.; Webb, H.E.; Bugarel, M.; Friedman, C.R.; Francois Watkins, L.K.; Loneragan, G.H.; Calle, A. First Known Report of mcr-Harboring Enterobacteriaceae in the Dominican Republic. International Journal of Environmental Research and Public Health 2023, 20, 5123. [Google Scholar] [CrossRef]
- Ling, Z.; Yin, W.; Shen, Z.; Wang, Y.; Shen, J.; Walsh, T.R. Epidemiology of mobile colistin resistance genes mcr-1 to mcr-9. Journal of Antimicrobial Chemotherapy 2020, 75, 3087–3095. [Google Scholar] [CrossRef] [PubMed]
- Lemlem, M.; Aklilu, E.; Mohamed, M.; Kamaruzzaman, N.F.; Zakaria, Z.; Harun, A.; Devan, S.S.; Kamaruzaman, I.N.A.; Reduan, M.F.H.; Saravanan, M. Phenotypic and genotypic characterization of colistin-resistant Escherichia Coli with mcr-4, mcr-5, mcr-6, and mcr-9 genes from broiler chicken and farm environment. BMC microbiology 2023, 23, 392. [Google Scholar] [CrossRef] [PubMed]
- Tansarli, G.S.; Andreatos, N.; Pliakos, E.E.; Mylonakis, E. A systematic review and meta-analysis of antibiotic treatment duration for bacteremia due to Enterobacteriaceae. Antimicrobial Agents and Chemotherapy 2019, 63, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Yusuf, M.A.; Siddiqui, U.R.; Debnath, A.C.; Shil, R.C. Frequency and distribution of multidrug resistance enterobacteriaceae isolated from hospital and community acquired urinary tract infection patient attended at a tertiary level care hospital in Dhaka city. 2023.
- Villafuerte, D.; Aliberti, S.; Soni, N.J.; Faverio, P.; Marcos, P.J.; Wunderink, R.G.; Rodriguez, A.; Sibila, O.; Sanz, F.; Martin-Loeches, I. Prevalence and risk factors for Enterobacteriaceae in patients hospitalized with community-acquired pneumonia. Respirology 2020, 25, 543–551. [Google Scholar] [CrossRef]
- Choi, Y.K.; Byeon, E.J.; Park, J.J.; Lee, J.; Seo, Y.B. Antibiotic resistance patterns of Enterobacteriaceae isolated from patients with healthcare-associated infections. Infection & Chemotherapy 2021, 53, 355. [Google Scholar]
- Tian, G.-B.; Doi, Y.; Shen, J.; Walsh, T.R.; Wang, Y.; Zhang, R.; Huang, X. MCR-1-producing Klebsiella pneumoniae outbreak in China. The Lancet Infectious Diseases 2017, 17, 577. [Google Scholar] [CrossRef]
- Schwarz, S.; Johnson, A.P. Transferable resistance to colistin: a new but old threat. Journal of Antimicrobial Chemotherapy 2016, 71, 2066–2070. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Y.; Chen, M.; Hu, J.; Zhang, H.; Xiang, Y.; Yang, H.; Qiu, S.; Song, H. Plasmid-borne colistin resistance gene mcr-1 in a multidrug resistant Salmonella enterica serovar Typhimurium isolate from an infant with acute diarrhea in China. International Journal of Infectious Diseases 2021, 103, 13–18. [Google Scholar] [CrossRef]
- Billah, S.M.; Raihana, S.; Ali, N.B.; Iqbal, A.; Rahman, M.M.; Khan, A.N.S.; Karim, F.; Karim, M.A.; Hassan, A.; Jackson, B.; et al. Bangladesh: a success case in combating childhood diarrhoea. J Glob Health 2019, 9, 020803. [Google Scholar] [CrossRef]
- Sharif, N.; Nobel, N.U.; Sakib, N.; Liza, S.M.; Khan, S.T.; Billah, B.; Parvez, A.K.; Haque, A.; Talukder, A.A.; Dey, S.K. Molecular and Epidemiologic Analysis of Diarrheal Pathogens in Children With Acute Gastroenteritis in Bangladesh During 2014–2019. The Pediatric Infectious Disease Journal 2020, 39, 580–585. [Google Scholar] [CrossRef]
- Islam, M.R.; Nuzhat, S.; Fahim, S.M.; Palit, P.; Flannery, R.L.; Kyle, D.J.; Mahfuz, M.; Islam, M.M.; Sarker, S.A.; Ahmed, T. Antibiotic exposure among young infants suffering from diarrhoea in Bangladesh. J Paediatr Child Health 2021, 57, 395–402. [Google Scholar] [CrossRef]
- Khedher, M.B.; Baron, S.A.; Riziki, T.; Ruimy, R.; Raoult, D.; Diene, S.M.; Rolain, J.-M. Massive analysis of 64,628 bacterial genomes to decipher water reservoir and origin of mobile colistin resistance genes: is there another role for these enzymes? Scientific reports 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Aghapour, Z.; Gholizadeh, P.; Ganbarov, K.; Bialvaei, A.Z.; Mahmood, S.S.; Tanomand, A.; Yousefi, M.; Asgharzadeh, M.; Yousefi, B.; Kafil, H.S. Molecular mechanisms related to colistin resistance in Enterobacteriaceae. Infection and drug resistance 2019, 12, 965. [Google Scholar] [CrossRef] [PubMed]
- Gu, D.-x.; Huang, Y.-l.; Ma, J.-h.; Zhou, H.-w.; Fang, Y.; Cai, J.-c.; Hu, Y.-y.; Zhang, R. Detection of colistin resistance gene mcr-1 in hypervirulent Klebsiella pneumoniae and Escherichia coli isolates from an infant with diarrhea in China. Antimicrobial agents and chemotherapy 2016, 60, 5099–5100. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zhuang, Y.; Luo, J.; Xiao, Q.; Wu, Y.; Chen, Y.; Chen, M.; Zhang, X. Prevalence of colistin-resistant mcr-1-positive Escherichia coli isolated from children patients with diarrhoea in Shanghai, 2016–2021. Journal of Global Antimicrobial Resistance 2023, 34, 166–175. [Google Scholar] [CrossRef]
- Monira, S.; Shabnam, S.A.; Ali, S.; Sadique, A.; Johura, F.-T.; Rahman, K.Z.; Alam, N.H.; Watanabe, H.; Alam, M. Multi-drug resistant pathogenic bacteria in the gut of young children in Bangladesh. Gut pathogens 2017, 9, 1–8. [Google Scholar] [CrossRef]
- Maalej, S.M.; Meziou, M.R.; Rhimi, F.M.; Hammami, A. Comparison of disc diffusion, Etest and agar dilution for susceptibility testing of colistin against Enterobacteriaceae. Lett Appl Microbiol 2011, 53, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Shiraishi, T.; Hiyama, Y.; Honda, H.; Shinagawa, M.; Usui, M.; Kuronuma, K.; Masumori, N.; Takahashi, S.; Tamura, Y. Contribution of novel amino acid alterations in pmrA or pmrB to Colistin resistance in mcr-negative Escherichia coli clinical isolates, including major multidrug-resistant lineages O25b: H4-ST131-H 30Rx and Non-x. Antimicrobial agents and chemotherapy 2018, 62, e00864–00818. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Jayol, A.; Bontron, S.; Villegas, M.-V.; Ozdamar, M.; Türkoglu, S.; Nordmann, P. The mgrB gene as a key target for acquired resistance to colistin in Klebsiella pneumoniae. Journal of Antimicrobial Chemotherapy 2015, 70, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Cannatelli, A.; Giani, T.; D’Andrea, M.M.; Di Pilato, V.; Arena, F.; Conte, V.; Tryfinopoulou, K.; Vatopoulos, A.; Rossolini, G.M. MgrB inactivation is a common mechanism of colistin resistance in KPC-producing Klebsiella pneumoniae of clinical origin. Antimicrobial agents and chemotherapy 2014, 58, 5696–5703. [Google Scholar] [CrossRef] [PubMed]
- Borowiak, M.; Baumann, B.; Fischer, J.; Thomas, K.; Deneke, C.; Hammerl, J.A.; Szabo, I.; Malorny, B. Development of a novel mcr-6 to mcr-9 multiplex PCR and assessment of mcr-1 to mcr-9 occurrence in colistin-resistant Salmonella enterica isolates from environment, feed, animals and food (2011–2018) in Germany. Frontiers in microbiology 2020, 11, 80. [Google Scholar] [CrossRef]
- Hussein, N.H.; AL-Kadmy, I.; Taha, B.M.; Hussein, J.D. Mobilized colistin resistance (mcr) genes from 1 to 10: a comprehensive review. Molecular Biology Reports 2021, 48, 2897–2907. [Google Scholar] [CrossRef] [PubMed]
- Formosa, C.; Herold, M.; Vidaillac, C.; Duval, R.; Dague, E. Unravelling of a mechanism of resistance to colistin in Klebsiella pneumoniae using atomic force microscopy. Journal of Antimicrobial Chemotherapy 2015, 70, 2261–2270. [Google Scholar] [CrossRef] [PubMed]
- Gogry, F.A.; Siddiqui, M.T.; Sultan, I.; Haq, Q.M.R. Current update on intrinsic and acquired colistin resistance mechanisms in bacteria. Frontiers in medicine 2021, 8, 677720. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.; Vargas, M.; Regueiro, V.; Llompart, C.; Albert, í.S.; Bengoechea, J.A. Capsule polysaccharide mediates bacterial resistance to antimicrobial peptides. Infect. Immun, 2004; 72, 7107–7114. [Google Scholar]
- Bengoechea, J.A.; Skurnik, M. Temperature-regulated efflux pump/potassium antiporter system mediates resistance to cationic antimicrobial peptides in Yersinia. Molecular microbiology 2000, 37, 67–80. [Google Scholar] [CrossRef]
- Puja, H.; Bolard, A.; Noguès, A.; Plésiat, P.; Jeannot, K. The efflux pump MexXY/OprM contributes to the tolerance and acquired resistance of Pseudomonas aeruginosa to colistin. Antimicrobial agents and chemotherapy 2020, 64, e02033–02019. [Google Scholar] [CrossRef]
- NationRL, L. Colistin in the 21st century. Curr Opin Infect Dis 2009, 22, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Jarman, A.F.; Long, S.E.; Robertson, S.E.; Nasrin, S.; Alam, N.H.; McGregor, A.J.; Levine, A.C. Sex and gender differences in acute pediatric diarrhea: a secondary analysis of the Dhaka study. Journal of epidemiology and global health 2018, 8, 42. [Google Scholar] [CrossRef]
- Mero, W.; Jameel, A.Y.; Amidy, K.S.K. Microorganisms and viruses causing diarrhea in infants and primary school children and their relation with age and sex in Zakho city, Kurdistan Region, Iraq. 2015.
- Gupta, N.; Limbago, B.M.; Patel, J.B.; Kallen, A.J. Carbapenem-resistant Enterobacteriaceae: epidemiology and prevention. Clinical infectious diseases 2011, 53, 60–67. [Google Scholar] [CrossRef]
- Smith, H.Z.; Hollingshead, C.M.; Kendall, B. Carbapenem-Resistant Enterobacterales. In StatPearls; StatPearls Publishing Copyright © 2024, StatPearls Publishing LLC.: Treasure Island (FL), 2024.
- Hansen, G.T. Continuous Evolution: Perspective on the Epidemiology of Carbapenemase Resistance Among Enterobacterales and Other Gram-Negative Bacteria. Infect Dis Ther 2021, 10, 75–92. [Google Scholar] [CrossRef]
- Falagas, M.E.; Kasiakou, S.K.; Saravolatz, L.D. Colistin: the revival of polymyxins for the management of multidrug-resistant gram-negative bacterial infections. Clinical infectious diseases 2005, 40, 1333–1341. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.; Sharma, D.; Singh, A.; Sunita, K. Colistin Resistance and Management of Drug Resistant Infections. Can J Infect Dis Med Microbiol 2022, 2022, 4315030. [Google Scholar] [CrossRef]
- Davies, M.; Walsh, T.R. A colistin crisis in India. Lancet Infect Dis 2018, 18, 256–257. [Google Scholar] [CrossRef]
- Godman, B.; Egwuenu, A.; Haque, M.; Malande, O.O.; Schellack, N.; Kumar, S.; Saleem, Z.; Sneddon, J.; Hoxha, I.; Islam, S.; et al. Strategies to Improve Antimicrobial Utilization with a Special Focus on Developing Countries. Life (Basel) 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Haque, T.A.; Urmi, U.L.; Islam, A.B.M.M.K.; Ara, B.; Nahar, S.; Mosaddek, A.S.M.; Lugova, H.; Kumar, S.; Jahan, D.; Rahman, N.A.A.; Haque, M.; Islam, S.; Godman, B. Detection of qnr genes and gyrA mutation to quinolone phenotypic resistance of UTI pathogens in Bangladesh and the implications. Appl Pharm Sci, 2022; 12, 185–198. [Google Scholar]
- Shanta, A.S.; Islam, N.; Asad, M.A.; Akter, K.; Habib, M.B.; Hossain, M.J.; Nahar, S.; Godman, B.; Islam, S. Resistance and Co-resistance of Metallo-Beta-Lactamase Genes in Diarrheal and Urinary Tract Pathogens in Bangladesh. Preprints 2024. [Google Scholar] [CrossRef]
- Haque, M.; Godman, B. Potential strategies to improve antimicrobial utilisation in hospitals in Bangladesh building on experiences across developing countries. Bangladesh Journal of Medical Science 2021, 20, 469–477. [Google Scholar] [CrossRef]
- Harun, M.G.D.; Anwar, M.M.U.; Sumon, S.A.; Hassan, M.Z.; Mohona, T.M.; Rahman, A.; Abdullah, S.; Islam, M.S.; Kaydos-Daniels, S.C.; Styczynski, A.R. Rationale and guidance for strengthening infection prevention and control measures and antimicrobial stewardship programs in Bangladesh: a study protocol. BMC Health Serv Res 2022, 22, 1239. [Google Scholar] [CrossRef] [PubMed]
- Sears, C.L.; Islam, S.; Saha, A.; Arjumand, M.; Alam, N.H.; Faruque, A.; Salam, M.; Shin, J.; Hecht, D.; Weintraub, A. Association of enterotoxigenic Bacteroides fragilis infection with inflammatory diarrhea. Clinical Infectious Diseases 2008, 47, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.V.; Duflo, E. What is middle class about the middle classes around the world? Journal of economic perspectives 2008, 22, 3–28. [Google Scholar] [CrossRef] [PubMed]
- Okuma, T.; Nakamura, M.; Totake, H.; Fukunaga, Y. Microbial contamination of enteral feeding formulas and diarrhea. Nutrition 2000, 16, 719–722. [Google Scholar] [CrossRef] [PubMed]
- Djim-Adjim-Ngana, K.; Oumar, L.A.; Mbiakop, B.W.; Njifon, H.L.M.; Crucitti, T.; Nchiwan, E.N.; Yanou, N.N.; Deweerdt, L. Prevalence of extended-spectrum beta-lactamase-producing enterobacterial urinary infections and associated risk factors in small children of Garoua, Northern Cameroon. Pan African Medical Journal 2020, 36. [Google Scholar] [CrossRef] [PubMed]
- Van Der Zee, A.; Roorda, L.; Bosman, G.; Ossewaarde, J.M. Molecular diagnosis of urinary tract infections by semi-quantitative detection of uropathogens in a routine clinical hospital setting. PloS one 2016, 11, e0150755. [Google Scholar] [CrossRef]
- Satlin, M.J.; Lewis, J.S.; Weinstein, M.P.; Patel, J.; Humphries, R.M.; Kahlmeter, G.; Giske, C.G.; Turnidge, J. Clinical and Laboratory Standards Institute and European Committee on Antimicrobial Susceptibility Testing Position Statements on Polymyxin B and Colistin Clinical Breakpoints. Clinical Infectious Diseases 2020, 71, e523–e529. [Google Scholar] [CrossRef]
- M100, C. Performance Standards for Antimicrobial Susceptibility Testing. The Clinical and Laboratory Standards Institute (CLSI). 2018, 28th Volume.
- Wiegand, I.; Hilpert, K.; Hancock, R.E. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nature protocols 2008, 3, 163–175. [Google Scholar] [CrossRef]
- Behera, B.; Mathur, P.; Das, A.; Kapil, A.; Gupta, B.; Bhoi, S.; Farooque, K.; Sharma, V.; Misra, M.C. Evaluation of susceptibility testing methods for polymyxin. International journal of infectious diseases : IJID : official publication of the International Society for Infectious Diseases 2010, 14, e596–601. [Google Scholar] [CrossRef]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.; Giske, C.; Harbarth, S.; Hindler, J.; Kahlmeter, G.; Olsson-Liljequist, B. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clinical microbiology and infection 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Rebelo, A.R.; Bortolaia, V.; Kjeldgaard, J.S.; Pedersen, S.K.; Leekitcharoenphon, P.; Hansen, I.M.; Guerra, B.; Malorny, B.; Borowiak, M.; Hammerl, J.A.; et al. Multiplex PCR for detection of plasmid-mediated colistin resistance determinants, mcr-1, mcr-2, mcr-3, mcr-4 and mcr-5 for surveillance purposes. Euro Surveill 2018, 23. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Sampling Location. Different sites were marked, indicating the spatial distribution of diarrheal patients from whom samples were collected. The map displays sampling sites across various cities in Bangladesh.
Figure 1.
Sampling Location. Different sites were marked, indicating the spatial distribution of diarrheal patients from whom samples were collected. The map displays sampling sites across various cities in Bangladesh.
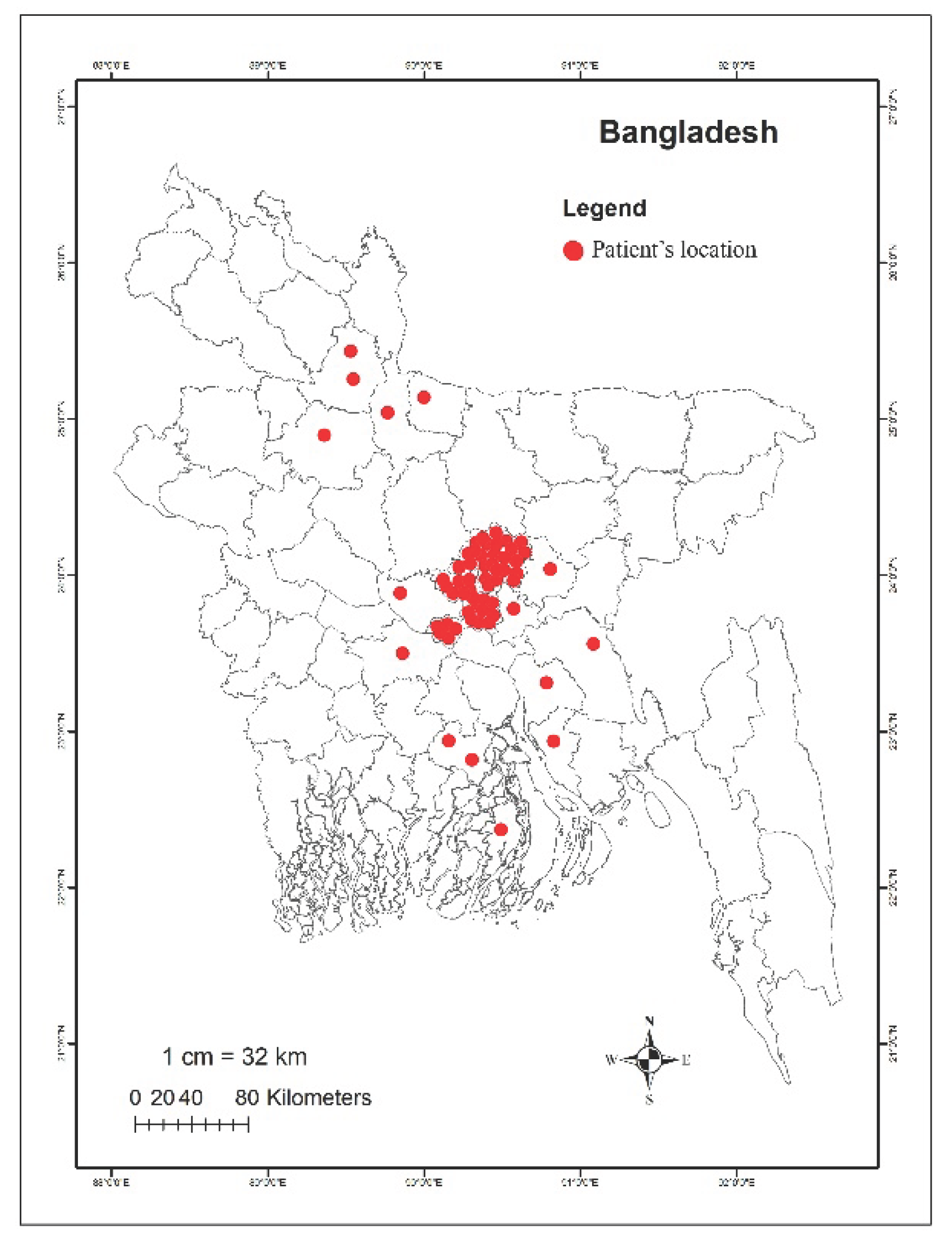
Figure 2.
Socioeconomic status of patients (A), as well as their age, sex (B), and clinical symptoms associated with diarrhea (C).
Figure 2.
Socioeconomic status of patients (A), as well as their age, sex (B), and clinical symptoms associated with diarrhea (C).
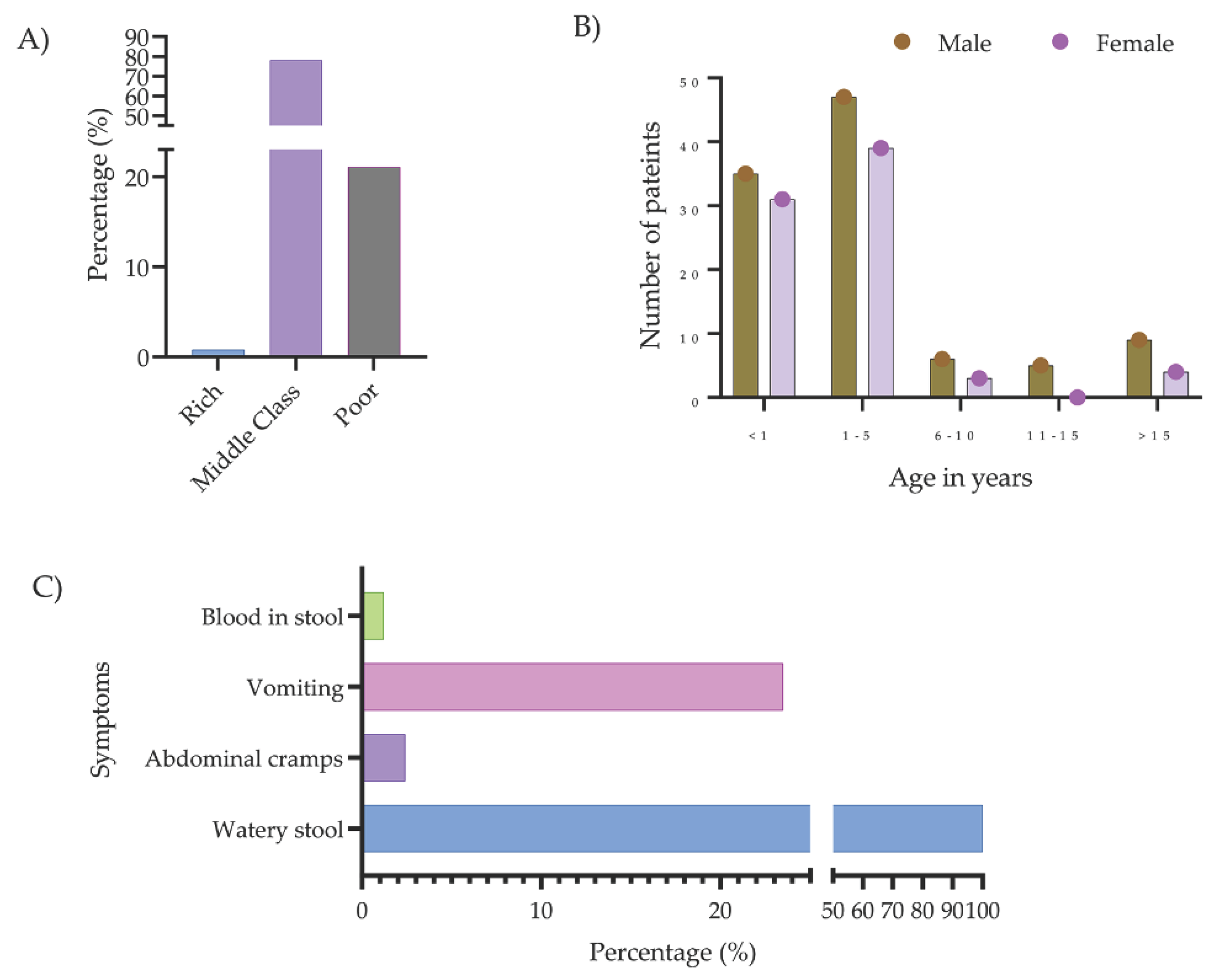
Figure 3.
Distribution of different bacteria identified from diarrheal patients. The pie chart illustrates the relative numbers of various bacteria identified among diarrheal patients. Each segment represents a specific bacterial species, with the size of each segment corresponding to the frequency of its occurrence in the sampled population.
Figure 3.
Distribution of different bacteria identified from diarrheal patients. The pie chart illustrates the relative numbers of various bacteria identified among diarrheal patients. Each segment represents a specific bacterial species, with the size of each segment corresponding to the frequency of its occurrence in the sampled population.
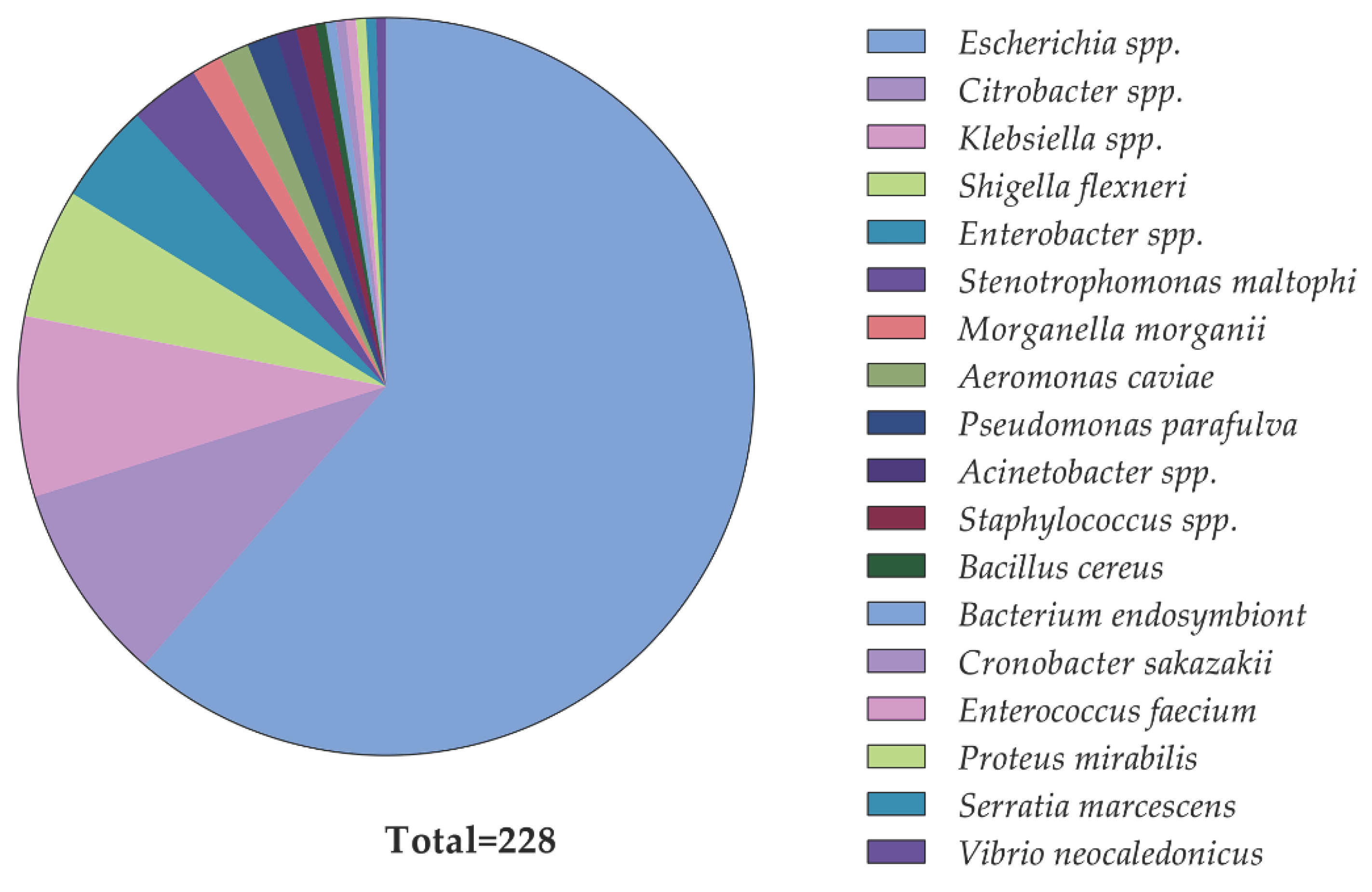
Figure 4.
Distribution of MIC levels of colistin sulfate among mcr-positive and mcr-negative isolates.
Figure 4.
Distribution of MIC levels of colistin sulfate among mcr-positive and mcr-negative isolates.
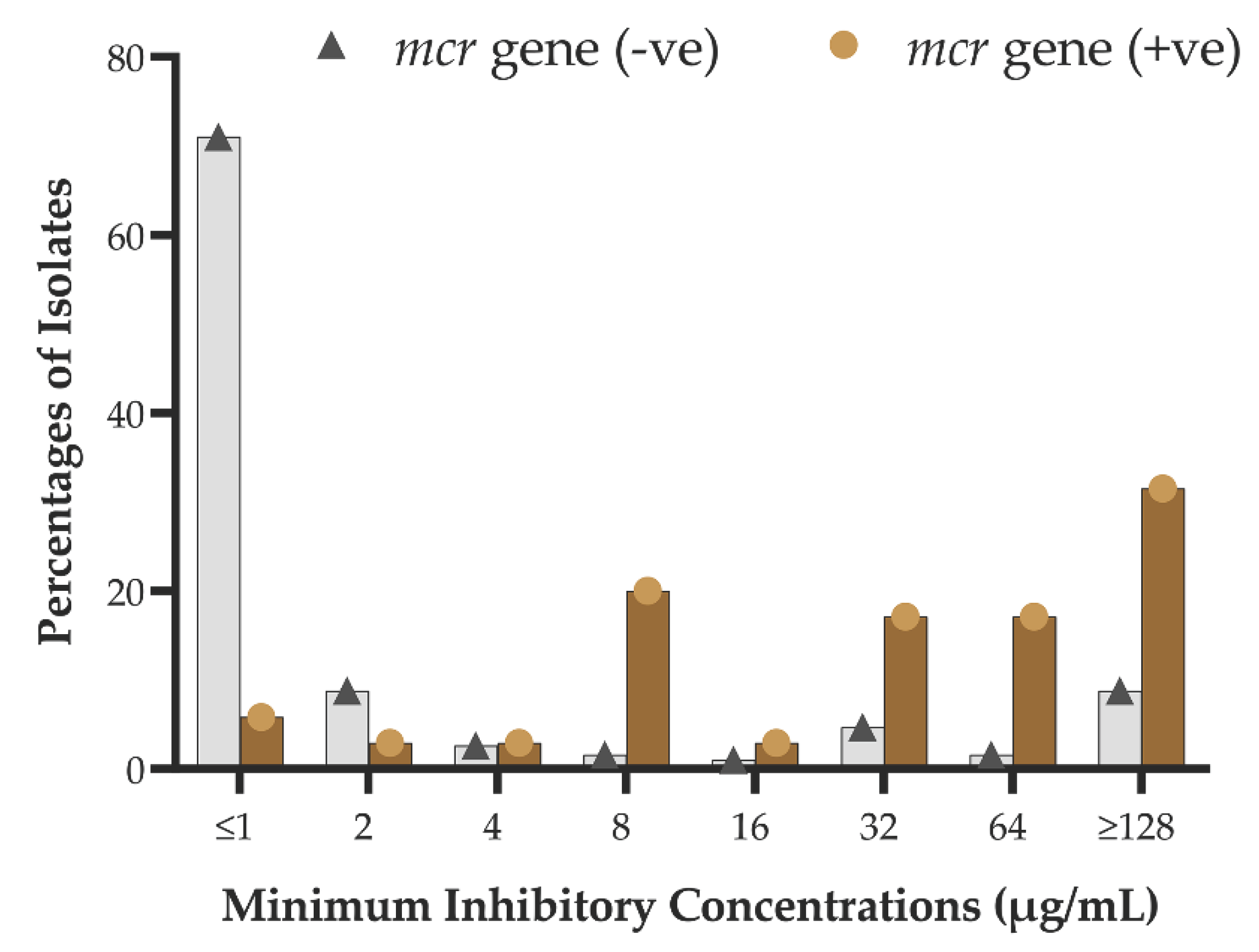

Table 1.
Identified diarrheal pathogens carrying mcr gene variantsa.
| Bacteria typeb | Number of isolates carrying mcr genes | Percentage of mcr-positive isolates | |
|---|---|---|---|
| Positive | Negative | ||
| Escherichia spp.c | 20 | 120 | 14.3 |
| Shigella flexneri | 5 | 8 | 38.5 |
| Citrobacter spp.d | 5 | 15 | 25.0 |
| Klebsiella pneumoniae | 2 | 16 | 11.1 |
| Enterobacter hormaechei | 2 | 8 | 20.0 |
| Pseudomonas parafulva | 1 | 2 | 33.3 |
| Aeromonas caviae. | 0 | 3 | - |
| Acinetobacter spp | 0 | 2 | - |
| Bacillus cereus | 0 | 1 | - |
| Bacterium endosymbiont | 0 | 1 | - |
| Morganella morganii | 0 | 3 | - |
| Serratia marcescens | 0 | 1 | - |
| Stenotrophomonas maltoph | 0 | 7 | - |
| Vibrio neocaledonicus | 0 | 1 | - |
| Cronobacter sakazakii | 0 | 1 | - |
| Enterococcus faecium | 0 | 1 | - |
| Total | 35 | 190 | |
Note : a12 mcr-1 and two mcr-2 genes were identified by polymerase chain reaction (PCR). bBacteria were primary identified based on their growth on selective culture media followed by biochemical tests. Further, identifications were confirmed by API 20E and 16s rDNA sequencing. cEscherichia spp. includes three Escherichia fergusonii and seventeen Escherichia coli. dCitrobacter spp. includes one Citrobacter europaeus, two Citrobacter portucalensis and two Citrobacter portucalensis
Table 2.
Phenotypic genotypic association of mcr genes and colistin resistance.
| Presence of mcr gene varients | Phenotypic susceptibility | P value | ||
|---|---|---|---|---|
| sensitive | resistance | |||
| mcr-1 | Positive (7) | 1 | 6 | .001 |
| Negative (218) | 155 | 63 | ||
| mcr-2 | Positive (17) | 3 | 14 | .000 |
| Negative (208) | 153 | 55 | ||
| mcr-3 | Positive (13) | 0 | 13 | .000 |
| Negative (212) | 156 | 56 | ||
| combined | Positive (35) | 3 | 32 | .000 |
| Negative (190) | 152 | 38 | ||
Table 3.
Diarrheal isolates with identified mcr-gene variants and associated minimum inhibitory concentration of colistin.
Table 3.
Diarrheal isolates with identified mcr-gene variants and associated minimum inhibitory concentration of colistin.
|
mcr-possitive isolate ID |
Identified bacteriaa |
Identified mcr gene varient | Phenotypic colistin susceptibility by MIC (µg/mL)b |
|---|---|---|---|
| PBD009 | Shigella flexneri | mcr-3 | 256 |
| PBD014 | Klebsiella pneumoniae | mcr-2 | 128 |
| PBD018 | Escherichia coli | mcr-3 | 8 |
| PBD021 | Escherichia coli | mcr-3 | 128 |
| PBD022 | Escherichia coli | mcr-3 | 32 |
| PBD027 | Citrobacter portucalensis | mcr-2 | .5 |
| PBD028 | Escherichia coli | mcr-3 | 256 |
| PBD033C2 | Pseudomonas parafulva | mcr-1 | 128 |
| PBD35 | Citrobacter portucalensis | mcr-2 | 8 |
| PBD35C1 | Citrobacter freundii | mcr-2 | 128 |
| PBD35C2 | Citrobacter freundii | mcr-2 | 1 |
| PBD039 | Escherichia fergusonii | mcr-2 | 256 |
| PBD040 | Citrobacter europaeus | mcr-2 | 64 |
| PBD043 | Klebsiella pneumoniae | mcr-2 | 16 |
| PBD058 | Escherichia coli | mcr-2, mcr-3 | 8 |
| PBD062 | Escherichia fergusonii | mcr-2 | 32 |
| PBD072 | Escheril chia fergusonii | mcr-2 | 64 |
| PBD077 | Escherichia coli | mcr-1 | 8 |
| PBD077C1 | Shigella flexneri | mcr-1 | 32 |
| PBD077C2 | Escherichia coli | mcr-1,mcr-2 | 128 |
| PBD080C2 | Escherichia coli | mcr-1 | 2 |
| PBD081C3 | Escherichia coli | mcr-2 | 32 |
| PBD081C4 | Enterobacter hormaechei | mcr-3 | 128 |
| PBD081C1 | Escherichia coli | mcr-2 | 32 |
| PBD082 | Shigella flexneri | mcr-1 | 128 |
| PBD083C1 | Shigella flexneri | mcr-3 | 32 |
| PBD083C2 | Escherichia coli | mcr-2 | 64 |
| PBD084C1 | Enterobacter hormaechei | mcr-2 | 128 |
| PBD84C2 | Escherichia coli | mcr-2 | 4 |
| PBD090 | Escherichia coli | mcr-3 | 64 |
| PBD096 | Escherichia coli | mcr-3 | 8 |
| PBD107 | Escherichia coli | mcr-1 | 8 |
| PBD114 | Escherichia coli | mcr-3 | 8 |
| PBD116 | Escherichia coli | mcr-3 | 64 |
| PBD117 | Shigella flexneri | mcr-3 | 64 |
Note: aBacteria were identified by rapid biochemical test kit API 20E system (BioMe ´rieux, Durham, NC) followed by16s rDNA sequencing. bThe minimum inhibitory concentration (MIC) measurement was conducted by agar dilution method following the EUCAST guidelines.
Table 4.
Multi-drug resistance (MDR) phenomena of mcr-positive isolates.
| List of antibiotics tested (n = 17, from eight drug-classes) | Phenotypic susceptibilities of mcr-positive diarrheal isolates | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Drug class | Antibiotic name | PBD009 | PBD014 | PBD018 | PBD021 | PBD022 | PBD027 | PBD028 | PBD033C2 | PBD035 | PBD035C1 | PBD035C2 | PBD039 | PBD040 | PBD043 | PBD058 | PBD062 | PBD072 | PBD077 |
| β-lactam with β-lactamase inhibitor | Amoxi-clava | R | R | R | R | R | R | I | R | R | R | S | R | I | R | I | S | S | S |
| Cephalosporins | Cefuroxime-G2 | R | R | R | R | I | I | I | R | R | R | R | R | R | R | S | R | R | S |
| Cefixime-G3 | R | R | R | R | I | R | R | R | R | R | R | R | R | R | S | R | R | S | |
| Cefepime-G4 | R | R | R | R | R | I | R | R | I | I | I | R | I | R | I | R | R | S | |
| Carbapenems | Imipenem | R | R | I | R | R | I | I | R | I | R | S | I | I | R | S | I | R | S |
| Meropenem | R | R | S | R | R | S | R | S | S | S | S | I | S | S | S | S | R | S | |
| Quinolone and fluoroquinolones | Nalidixic acid | I | S | R | I | I | I | I | R | I | R | R | R | I | R | R | R | R | R |
| Ciprofloxacin | R | S | R | S | S | S | R | R | I | R | R | R | R | S | I | R | I | R | |
| Levofloxacin | R | S | R | S | S | S | I | R | S | I | S | R | S | R | S | R | S | R | |
| Lomefloxacin | S | S | R | S | I | S | R | R | I | R | R | R | S | I | R | R | R | R | |
| Aminoglycosides | Gentamicin | S | S | I | S | R | S | I | S | S | S | S | S | S | R | S | R | R | R |
| Amikacin | R | S | I | S | S | S | R | S | S | I | I | R | I | R | S | I | I | I | |
| Netilmicin | S | S | I | I | S | S | R | S | S | S | I | I | R | S | R | S | R | I | |
| Tobramycin | S | S | S | S | R | S | R | I | S | S | S | R | S | R | S | R | R | S | |
| Polymyxins | Colistin | R | S | R | S | R | R | R | R | S | R | R | R | R | R | S | S | R | R |
| Nitrofuran | Nitrofurantoin | I | R | I | R | R | I | R | R | S | I | S | S | I | R | I | R | I | I |
| Trimethoprim | Trimethoprim-sulfamethoxazole | S | S | R | S | R | I | S | R | S | R | R | S | S | S | R | R | R | R |
| MDR statuse | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |
Note: R, Resistant; S, Sensitive; I, Intermediate; MDR, multidrug resistant. aAmoxicillin-clavulanic acid. bPBD033C2 is Pseudomonas spp. which is naturally resistant to Amoxicillin-clavulanic acid, therefore, AST results have been excluded from MDR calculation. cPBD081C4 and PBD084C1 are Enterobacter spp., which is naturally resistant to amoxicillin-clavulanic acid, therefore, AST results has been excluded from MDR calculation. ddifferent generations of cephalosporin. eMultidrug-resistant isolate is defined by center for disease control (CDC) as an isolate that is resistant to at least one antibiotic in three or more drug classes (https:// www.cdc.gov/narms/resources/glossary ). ‘+’, indicates MDR-positive; and ‘-‘, indicates MDR-negative.
Table 5.
Demographic factors associated with mcr genes (n = 225).
| Demography | Number (%) of different mcr-gene variants | p-value | ||
|---|---|---|---|---|
|
mcr-1 positive (n = 7) |
mcr-1 negative (n = 218) |
|||
| Gender | Male | 5 (3.8) | 125 (96.2) | 0.702* |
| Female | 2 (2.1) | 94 (97.9) | ||
| Age group (years) | <1 | 4 (4.6) | 83 (95.4) | 0.707* |
| 1-5 | 3 (3.3) | 88 (96.7) | ||
| 6-10 | 0 | 12 (100) | ||
| 11-15 | 0 | 5 (100) | ||
| >15 | 0 | 30 (100) | ||
|
mcr-2 positive (n = 17) |
mcr-2 negative (n = 208) |
|||
| Gender | Male | 8 (6.2) | 122 (93.8) | 0.445* |
| Female | 9 (9.5) | 87 (90.5) | ||
| Age group (years) | <1 | 8 (9.2) | 79 (90.8) | 0.480* |
| 1-5 | 4 (4.4) | 87 (95.6) | ||
| 6-10 | 1 (8.3) | 11 (91.7) | ||
| 11-15 | 0 | 5 (100) | ||
| >15 | 4 (13.3) | 26 (86.7) | ||
|
mcr-3 positive (n = 13) |
mcr-3 negative (n = 212) |
|||
| Gender | Male | 7 (5.4) | 123 (94.6) | 0.779* |
| Female | 6 (6.3) | 89 (93.7) | ||
| Age group (years) | <1 | 5 (5.7) | 82 (94.3) | 0.781* |
| 1-5 | 4 (4.4) | 87 (87) | ||
| 6-10 | 1 (8.3) | 11 (91.7) | ||
| 11-15 | 0 | 5 (100) | ||
| >15 | 3 (10.0) | 27 (90.0) | ||
|
mcr-1 to mcr-3 positive (n = 35) |
mcr-1 to mcr-3 negative (n = 190) |
|||
| Gender | Male | 19 (14.6) | 111 (85.4) | 0.711* |
| Female | 16 (16.8) | 79 (83.2) | ||
| Age group (years) | <1 | 15 (17.2) | 72 (82.8) | 0.503* |
| 1-5 | 11 (12.1) | 80 (87.9) | ||
| 6-10 | 2 (16.7) | 10 (83.3) | ||
| 11-15 | 0 | 5 (100) | ||
| >15 | 7 (23.3) | 23 (76.7) | ||
Note: %, row percentage; *Fisher’s Exact test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



