Submitted:
25 March 2024
Posted:
26 March 2024
You are already at the latest version
Abstract
Cucurbitacin phytonematicides consistently suppress population densities of plant nematodes, but have no effect against bacteria, fungi and insects. This study investigates the mechanism through which cucurbitacin phytonematicides consistently exert nematicidal properties on nematodes. Post 72-h exposure of second-stage juveniles (J2) of Meloidogyne enterolobii to geometric concentration of Nemafric-BL phytonematicide, J2 total proteins were measured using TruspecCHNS Macro. Total proteins versus phytonematicide exhibited a negative quadratic relation (R2 = 0.97), with the minimum total proteins achieved at x = 4.9% phytonematicide. Beyond the minimum, total proteins increased gradually, depicting the existence of both isoprenylation (breakdown) and farnesylation (biosynthesis) of total proteins. In conclusion, cucurbitacins and nematode cuticles are both chemically structured, which provides the mechanism through which the test chemicals consistently decrease population densities of plant nematodes.
Keywords:
biosorption
; CARD
; cucurbitacins
; cuticles
; farneslation
; isoprenylation
; Meloidogyne enterolobii
; total protein
1. Introduction
Cucurbitacins are oxygenated triterpenoids with tetracyclic cucurbitane nucleus skeleton, namely, the 19-1→β-abeo-10α-lanost-5-ene [1,2]. Triterpenoids are biosynthesized in the endoplasmic reticulum and cytoplasm via the combination of two molecules of farnesyl diphosphate to form a C30 precursor, which is technically referred to as mevalonate/acetate pathway [3]. Cucurbitaceous plants that produce cucurbitacins include but are not limited to the genera Bryonia, cumis, Cucurbita, Luffa, Echinocystis, Lagenaria and Citrullus [1,2]. All cucurbitaceous plants originally biosynthesize two cucurbitacins, namely, cucurbitacin B and E, each with an acetyl function at C-25 [4]. All other identified cucurbitacins, at least 20, are produced from the two primary cucurbitacins through a wide range of chemical reactions, which include either hydrogenation by cucurbitacin Δ23–reductase, deacetylation by cucurbitacin acetylesterases, hydroxylation, dehydrogenation or isomerization [5,6,7]. Due to the dominance of any of these chemical reactions among and within the genera, cucurbitacin types may or may not be genus-specific. For example, within the genus Cucumis, C. myriocarpus and C. africanus produce cucurbitacin A (C32H46O9) and cucurbitacin B (C32H46O8), respectively, suggesting the existence of different chemical reactions. The identified cucurbitacins basically differ from one another from hydroxylation at C-2, C-3, C-19 and C-24, along with the existence of double chemical bonds between C-1 and C-2 or between C-23 and C-24, the acetylation of C-25 hydroxyl groups and the presence of a ketone function at C-3 [1,2]. These chemical arrangements, with the identified cucurbitacins still being viewed as work-in-progress, confer a wide assortment of novel biological activities to the test chemicals. Pharmacological studies had since shown that cucurbitacins have the potential to treat a wide variety of human and animal sicknesses [1,8,9]. In traditional medicine, plant organs with cucurbitacins had been used for the treatment of numerous human and animal ailments [10], including the deworming of canines [11].
In plant protection, cucurbitacin phytonematicides have been researched and developed from fermentation of wild cucumber (Cucumis myriocarpus Naude) and wild watermelon (Cucumis africanus L.), with cucurbitacins extracted from fruits using effective microorganisms [12]. In granular formulation (G) the products from C. myriocapus and C. africanus are available as Nemarioc-AG and Nemafric-BG phytonematicides, respectively. Similarly, in liquid formulation (L) the products are Nemarioc-AL and Nemafric-BL phytonematicides, with A and B representing the active ingredients. The two products have consistently suppressed population densities of r-nematode strategists such as the root-knot (Meloidogyne species) nematodes on various crops and the citrus nematode (Tylenchulus semipenetrans Cobb) on various rootstocks. Breakthroughs in phytonematicides were achieved through the Curve-fitting Allelochemical Response Dose (CARD) algorithm computer model, which was originated in Australia [13]. However, the model was perfected in our laboratory to embrace the indispensable concept of inference, thereby promoting normalization of data [14] as required in sample means to be the true reflection of the population means [15]. CARD computer model assisted us in avoiding toxicity concentrations on various crops, along with ensuring consistent results in suppression of nematode population densities [11,14].
During the improvement of CARD as a scientific tool, it was shown through morphometric adjustments that at the non-phytotoxic concentration of 2.62% phytonematicide, the K-strategist nematodes were somewhat having limited tolerance to the test phytonematicides [12,14,16]. However, the accumulative effects during the cropping cycle eventually have lethal effects on such nematodes. Notwithstanding the observed consistent lethal effects on nematodes, the products hardly have any deadly effect on bacteria and fungi [11,14] and insects such as mealybugs [17], leaf miners, aphids and mites (unpublished data). In contrast to the biocidal fumigant chemical nematicides, which have since been withdrawn from the agrochemical markets, the cucurbitacin phytonematicides have potent nematicidal properties, but are highly selective towards other pests. The primary difference between nematodes and the listed pests could mainly be on their body coverings, which had been used taxonomically to classify animals into cuticle-bearing and non-cuticle bearing animals, with reference to the Superphylum, Ecdysozoa [18].
In nematodes, one of the cuticle-bearing Ecdysozoa, the bodies and certain cavities are covered by the cuticle, which has distinct features [19]. The cuticle comprises an outer epicuticle layer that is lipid-rich and an inner subcuticular layers which are protein-rich (Figure 1). From the epicuticle to the interior is the (a) cortical layer with insoluble proteins – the cuticulins, then (b) the median layer with pillar-like proteins having interspaces filled with the gelatinous matrix, (c) the basal layer with distinct soluble proteins in the form of fibers that are dispersed as a dense gelatinous matrix (collagen) and then (e) the hypodermal layer. Lipids of the epicuticle are coated with a glycoprotein, technically called the surface coat, which plays a protective role to the epicuticle [19].
Lipids in the epicuticle enhance the mobility of nematodes in aqueous solutions [20]. During the molting process only the epicuticle and the median layers are shed off (Figure 1A), whereas the hypodermis is used to generate the new cuticle [20]. The newly molted juveniles exit the shed cuticular layers through the stomas.
The mechanism through which the cucurbitacin phytonematicide are nematicidal to plant nematodes is not documented. Exposure of Meloidogyne species to both fermented crude extracts and pure cucurbitacins resulted in paralysis, which was irreversible [21]. By definition, paralysis is the loss of the ability to move and feel in all or part of the body and mostly involves some damaging effects on the nervous system. In their review of the mechanisms through which systemic nematostatic chemicals such as organophosphates, carbamates and oxycarbamates affected nematodes, Van Gundy and McKenry [22] found no empirical evidence that the test chemicals had any effect on the nervous system. Cucurbitacins have distinct functional groups that attack lipids and proteins in various entities [23]. Since the epicuticle and the subcuticular layers of nematodes are lipid-rich and protein-rich, respectively, the arrangement could possibly enhance the interaction with the functional groups of cucurbitacins. As illustrated in Figure 1B, the nematode cuticle consists of the lipid-containing epicuticle to the exterior of the body, with five interiorly arranged sublayers of proteins. Since the protein-containing sublayers are relatively large when compared with the lipid-containing epicuticle, the former is an ideal entity for investigating the mechanism through which cucurbitacins consistently suppress nematode population densities. The objective of this study was, therefore, to determine whether increasing concentration of Nemafric-BL phytonematicides would affect the total proteins in M. enterolobii.
2. Materials and Methods
2.1. Preparation of Juveniles
Ten egg masses of M. enterolobii were picked from roots of kenaf (Hibiscus cannabinus L.) using a toothpick stick into water in a Petri dish and torn open using two toothpick sticks. Eggs were incubated for three days at 28ºC and hatched second-stage juveniles (J2) were separated from eggs using the modified Baerman method [24]. Approximately 7000 J2 were used to inoculate 100 seedlings of nematode susceptible tomato cv. ′Floradade‵, each in 20-cm-diameter plastic pot filled with 2700 ml growing mixture comprising pasteurised (300°C for 3 h) river sand and Hygromix (Hygrotech, Tshwane, South Africa) at 3:1 (v/v) ratio. At 95 days shoots were severed and discarded, with roots removed from the growing mixture and slightly shaken in a bucket half-filled with tapwater to release the adhering particles without dislodging the egg masses. Approximately 5000 egg masses were picked, eggs released and incubated for the eventual extraction of J2 as described above.
2.2. Treatment and Experimental Design
Approximately 250 000 J2 per Petri dish were subjected to 0, 2, 4, 8, 18, 32 and 64% Nemafric-BL phytonematicide, arranged in a completely randomised design, with three replicates, inside the growth chamber 25°C. The study was repeated three times on weekly basis.
2.3. Data Collection
At 72 h after exposure, total proteins were measured using TruSpecCHNS Macro (Leo, St. Joseph, MI, USA) as described previously [25].
2.4. Data Analysis
Prior to analysis, phytonematicide concentration (x-axis) data were expressed as exponentials (20, 21, 22, 23, 24, 25 and 26) % and transformed using log2x = x (1) to homogenise the variance [26]. Interval interactions using analysis of variance were assessed using Statistix 10.0 software. Since interval interactions were not significant at 5% level of probability [15], data were pooled (n = 63). Mean total proteins versus the transformed data (0, 1, 2, 3, 4, 5 and 6%) were subjected to CARD algorithm computer model [11,13,21].
3. Results
Total proteins versus cucurbitacins exhibited negative quadratic relations, with the model explained by the quadratic equation Y = b2x2 + b1x + c, along with an overall association (R2) of 97% (Figure 2). Using the relation x = –b1/2b2 from the quadratic equation [15,21], the minimum occurred at x = 4.9% Nemafric-BL phytonematicide, which translates to 31.36% untransformed concentration of Nemafric-BL phytonematicide. Relative to the untreated control, at the minimum Nemafric-BL had decreased total proteins by 82.29%. After the minimum of 2.53% total protein, the latter started to increase gradually to 2.54%, 3.13% and 4.7% at 5, 6 and 7% Nemafric-BL phytonematicide, respectively (Figure 2). The gradual increases in total proteins following the minimum, the model suggested that the subsequent biosynthesis of proteins increased the total proteins. Relative to the minimum, at 7% Nemafric-BL phytonematicide had increased total proteins by 85.77%.
4. Discussion
The observed results suggested that three processes, namely, sorption of cucurbitacins, breakdown of proteins and then biosynthesis of proteins, occurred. Sorption of cucurbitacins could be ascribed to the existence of the functional groups in cucurbitacins that target lipids and then proteins [23]. This sorption does not occur in entities devoid of the features observed in cuticles of nematodes. In J2 bodies sorption was followed by the breakdown of total proteins towards the detectable minimum, which was then followed by the biosynthesis of total proteins. The observation had the biological density-dependent growth (DGG) patterns, which describe various biological responses to the bioactive phytochemicals [11,13,21].
The observed process in our study could be compared to processes which occur during bioremediation. In the definition of bioremediation, there is first the concept of sorption, where the chemical compounds are removed from the environment [27]. Cucurbitacins are lipophilic [23], which therefore confers them the status of being hydrophobic [27]. The two properties improve the loading capabilities of cucurbitacins in the development of cucurbitacin drugs [27]. Lipids in the epicuticles have the capability of attracting cucurbitacin molecules from aqueous solutions and therefore their natural ability of being lipophilic [23]. This attraction provides sufficient reason as to why nematodes are highly sensitive to cucurbitacin phytonematicides in crude and purified forms [21,28,29,30,31]. Explained differently, this provide evidence for the first time why cucurbitacin phytonematicides had been highly consistent in suppressing nematode population densities under different environments and crops [14]. This attraction of cucurbitacins by the epicuticles, removes the active ingredients of cucurbitacin phytonematicides from the environment, which agrees with the description of bioremediation processes [32]. Once cucurbitacins are removed from the environment by the epicuticles of nematodes, they can, due to the hydrophobic properties of cucurbitacins, further be attracted to proteins in protein-rich sublayers of cuticles, thereby resulting in the breakdown of proteins, which is technically referred to as isoprenylation reactions [33].
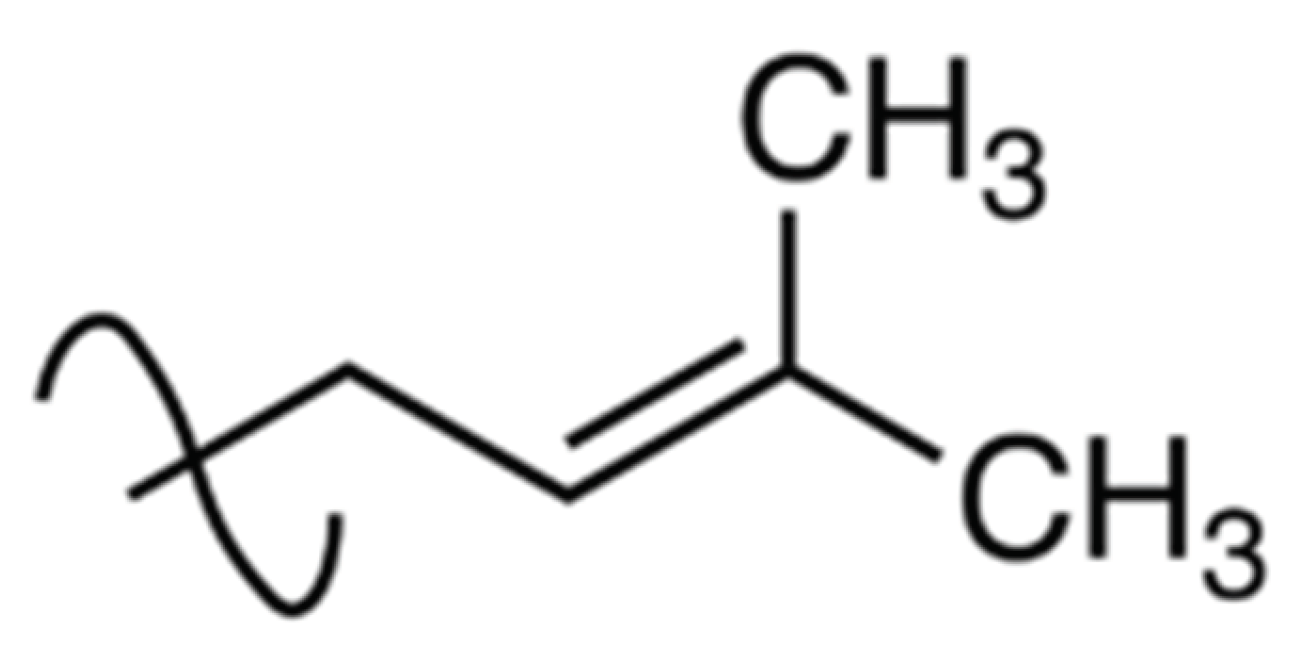
Isoprenylation involves the addition of hydrophobic molecules such as cucurbitacins through the prenyl groups (3-methylbut-2-en-1-yl) to the proteins [33,34,35]. In cucurbitacins the prenyl groups are located at C-9 [36] and are important for protein-to-protein binding through the highly specialized prenyl-binding domains (Figure 3). Generally, prenylation involves the transfer of either a farnesyl or a geranylgeranyl moiety to C-terminal cysteine(s) of the target protein, using one of the three enzymes, namely, farnesyl transferase, Caax protease and geranylgeranyl transferase I [35]. Obviously, the prenyl groups, also referred to as the isoprenyl groups (Figure 3), have one hydrogen atom more than
The isoprenes.
Importantly, farnesyl transferase is one of the enzymes required in the biosynthesis of cucurbitacins through the methyl-D-ereythritol 4-phosphate (MEP)/1-deoxy-D-xylulose 5-phosphate (MEP/DOXP) biosynthesis pathway [1 [35,36,37,38]. Although isoprenylation (breakdown) and farnesylation (biosynthesis) are two opposed reactions, they may occur separately or concurrently [35]. Farnesylation is a post-translational modification of proteins in which the isoprenyl groups are added to a cysteine residue [33,34,35]. This is an important process to mediate protein-to protein interactions, thereby increasing total proteins through the occurance of the enabling protein-to-protein membrane interactions.
Both isoprenylation and farnesylation as observed in our study supported the principle of density-dependent growth (DDG) patterns, which are key in biological reactions when entities are subjected to increasing concentration of bioactive phytonematicides such as cucurbitacins [11,14,21]. Notwithstanding the observed effects on nematode population densities, cucurbitacin phytonematicides have no deleterious effects on bacteria, fungi and mycoplasms [11] and insects such as mealybug [17], leaf miner and aphids as well as mites (unpublished data), which can be enumerated as entities that lack features that are in nematode cuticles. Nematodes are not the only cuticle-bearing animals. In the broadest sense, there are cuticle and non-cuticle bearing animals. In the cuticle-bearing animals, nematodes are grouped alongside ecdysones.
5. Conclusions
In the current study, cucurbitacin-nematode interactions resulted in isoprenylation (breakdown) of total proteins to the minimum, after which farneslation (biosynthesis) of proteins resulted in the gradual increase in the total proteins. At the minimum, which is equivalent to 100% inhibition, the test nematode is dead and although farneslation reactions, occur, the latter could not revive the affected nematode and thus, the irreversible effects of the test products on nematodes. In conclusion, isoprenylation of total proteins confer the mechanism through which cucurbitacin phytonematicides confer consistent nematicidal attributes on nematodes, thereby providing some empirically-based evidence on the reason the test products are highly selective and effective in suppressing nematode population densities.
Supplementary Materials
All data were included in the manuscript.
Author Contributions
Conceptualization, PWM and KMP; methodology, PWM and ARN.; software, PWM and ARN; validation, PWM and KMP; formal analysis, PWM; investigation, KMP; resources, PWM; data curation, KMP; writing—original draft preparation, PWM; writing, review and editing, PWM; visualization, ARN; project administration, KMP; funding acquisition, PWM. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Research Foundation and the Department of Agriculture, Land Reform and Rural Development, grant number 136124 and the APC was funded by the University of Limpopo.
Data Availability Statement
No new data were created.
Acknowledgments
The Agricultural Research Council of South Africa for the use of TruSpecCHNS Macro to collect total proteins.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Chen, J.C.; Chiu, M.H.; Nie, R.L.; Cordell, G.A.; Qiu, X.S. The cucurbitacins and cucurbitane glycosides: Structures and biological activities. Nat. Prod. Rep. 2005, 22, 386–399. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Kuo, T.C.; Yang, M.; Chien, T.; Chu, M.; Huang, L.; et al. Identification of cucurbitacins and assembly of a draft genome for Aquilaria agallocha. BMC Genom. 2014, 15, 578. [Google Scholar] [CrossRef] [PubMed]
- Noushahi, H.A.; Khan, A.H.; Noushahi, U.F.; Hussain, M.; Javed, T.; Zafar, M.; et al. Biosynthetic pathways of triterpenoids and strategies to improve their biosynthetic efficiency. Plant Growth Regul. 2022, 97, 437–454. [Google Scholar] [CrossRef] [PubMed]
- Gry, J.; Søborg, I.; Andersson, H.C. Cucurbitacins in plant food. Ekspressen Tryk and Kopicenter: Copenhagen, Denmark, 2006.
- Dirr, H.; Schabort, J.C.; Weitz, C. Cucurbitacin 23 reductase from fruit of Cucurbita maxima var. Green Hubbard. Biochem. J. 1986, 33, 649–653. [Google Scholar] [CrossRef] [PubMed]
- Schabort, J.C.; Teijema, H.L. The role of cucurbitacin Δ23 reductase in the breakdown pathway of toxic bitter principles in Cucurbita maxima. Phytochemistry. 1968, 7, 2107–2110. [Google Scholar] [CrossRef]
- Zhou, Y.; Ma, Y.; Zeng, J.; Duan, L.; Xue, X.; Wang, H.; et al. Convergence and divergence of cucurbitacin biosynthesis and regulation in Cucurbitaceae. Nat. Plants. 2016, 2, 16183–16202. [Google Scholar] [CrossRef] [PubMed]
- Abbas SVincourt, J.; Habib, L.; Netter, P.; Greige-Gerges, H.; Magdalou, J. The cucurbitacins E, D and I: Investigation of their cytotoxicity towards human chondrosarcoma SW 1353 cell line and their biotransformation in man liver. Toxicol. Lett. 2016, 216, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Mirr, S.A.; Mukherjee, S.; Makar, S.; Pal, G. Cucurbitacins a vibrant triterpenoid: A review on its anticancer property. PharmaTut. 2019, 7, 43–54. [Google Scholar]
- Mphahlele, R.R.; Mashela, P.W.; Pofu, K.M. Post-harvest fruit decay-inducing pathogen in medicinally important Cucumis species indigenous to South Africa. Afrn. J. Agric. Res. 2012, 6, 3786–3791. [Google Scholar] [CrossRef]
- Mashela, P.W.; De Waele, D.; Dube, Z.; KHosa, M.C.; Pofu, K.M.; Tefu, G.; et al. Alternative nematode management strategies. In Nematology in South Africa: A review from the 21st Century, 1st ed.; Fourie, H., et al., Eds.; Springer Nature: Berlin, Germany, 2017; pp. 151–181. [Google Scholar]
- Mashela, P.W.; De Waele, D.; Pofu, K.M. Use of indigenous Cucumis technologies as alternative to synthetic nematicides in management of root-knot nematodes in low-input agricultural farming systems: A review. Sci. Res. Essays. 2011, 33, 6762–6768. [Google Scholar]
- Mashela, P.W.; Pofu, K.M. Cucumis oilseed cake: Nematicidal attributes and management of associated challenges. In Oilseed cake for nematode management, 1st ed.; Amad, F., Pandey, R., Eds.; CRC Press: Boca Raton, FL, 2023; pp. 15–38. [Google Scholar]
- Liu, D.L.; Johnson, I.R.; Lovett, J.V. Mathematical modelling of allelopathey. III: a model for curve-fitting allelochemical dose responses. Nonlin. Biol., Toxicol. Med. 2003, 1, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Gomez, K.A.; Gomez, A.A. Statistical procedures for agricultural research. Wily: New York, USA. 1984.
- Matlala, M.C. Response of morphometrics to cucurbitacin-containing phytonematicides in Scutellonema brachyurus. MSc Dissert., University of Limpopo, Sovenga. South Africa. 2023.
- Mokhoelele, F. Nemarioc-AL, Nemafric-BL and momorfly phytoinsecticides with and without Dicerocaryum senecioides wettener in the management of mealybug on Mimusops zeyheri. Masters Dissertation, . University of Limpopo, Sovenga, South Africa. 2018. [Google Scholar]
- Aguinaldo, A.M.; Tubeville, J.M.; Linford, L.S.; Rivera, M.C.; Garey, J.R.; Raff, R.A.; Lake, J.A. Evidence for a clade of nematodes, arthropods and other moulting animals. Nature. 1997, 29, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.N.; Moens, M. Plant nematology. Wallingford, Oxfordshire, UK. 2013.
- Schultz, R.D.; Bennett, E.E.; Ellis, E.A.; Gumienny, T.L. Regulation of extracellular matrix organization by BMP signaling in Caenorhabditis elegans. PLoS ONE 2014, 9, e101929. [Google Scholar] [CrossRef] [PubMed]
- Dube, Z.P. Nemarioc-AL and Nemafric-BL phytonematicides: Bioactivities in Meloidogyne incognita, tomato crop, soil type and organic matter. PhD thesis, University of Limpopo, Sovenga, South Africa. 2016.
- Van Gundy, S.D.; McKenry, M.V. Action of nematicides; Horsfall, J.G. , Cowling, E.B., Eds.; Plant disease I Academic Press: New York, 1975; pp. 263–280. [Google Scholar]
- Van Wyk, B.E.; Wink, M. Medicinal plants of the world; Timber Press, Inc.: Portland, Oregon, USA, 2004. [Google Scholar]
- Marais, M.; Swart, A.; Fourie, H.; Berry, S.D.; Knoetze, R.; Malan, A.P. Techniques and procedures. In Nematology in South Africa: A view from the 21st century; Fourie, H., Spaull, V., Jones, R., Daneel, M., De Waele, D., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 73–117. [Google Scholar]
- Mashela, P.W.; Shokoohi, E. Morphometric and total protein responses in Meloidogyne incognita second-stage juveniles to Nemafric-BL phytonematicide. Sci. Rep. 2021, 11, 1135–1147. [Google Scholar] [CrossRef] [PubMed]
- Causton, D.R. Contemporary biology: A biologist’s mathematics. Bedford Square, London, 1977.
- Patel, S.K.; Lavasanifar, A.; Choi, P. Roles of non-polar and polar intermolecular interactions in the improvement of the drug loading capacity of PEO-b-PLC with hydrophobic cucurbitacin drugs. Biomacromolecules. 2009, 10, 2584–2591. [Google Scholar] [CrossRef] [PubMed]
- Dube, Z.P.; Mashela, P.W. Response of Meloidogyne incognita egress and overall sensitivity active ingredients of Nemarioc-AL and Nemafric-BL phytonematicides, Transylv. Rev. 2016, 7, 954–960. [Google Scholar]
- Dube, Z.P.; Mashela, P.W. Confirmation of bioactivities of active ingredients of Nemarioc-AL and Nemafric-BL phytonematicides. Acta Agric. Scand. Sect. B-Soil and Plant Sci. 2017, 67, 571–575. [Google Scholar] [CrossRef]
- Dube, Z.P.; Mashela, P.W. Effects of cucurbitacin A on mobility of Meloidogyne incognita second-stage juveniles. Res. Crops. 2018, 19, 504–508. [Google Scholar]
- Dube, Z.P.; Mashela, P.W.; De Waele, D. Sensitivity of Meloidogyne incognita second-stage juvenile hatch, motility and viability to pure cucurbitacins and cucurbitacin phytonematicides. S. Afrn. J. Plant and Soil. 2019, 36, 29–32. [Google Scholar] [CrossRef]
- Jørgensen, K.S. In Situ bioremediation. Adv. Appl. Microbiol. 2007, 61, 285–305. [Google Scholar]
- Casey, P.J.; Seabra, M.C. Protein prenyltransferases. J. Biol. Chem. 1996, 27, 5289–5292. [Google Scholar] [CrossRef] [PubMed]
- Marshall, C.J. Protein prenylation: A mediator of protein-protein interactions. Science. 1993, 259, 1865–1866. [Google Scholar] [CrossRef] [PubMed]
- Novelli, G.; D’Apice, M.R. Protein farneslation and disease. J. Inher. Metab. Dis. 2012, 35, 917–926. [Google Scholar] [CrossRef]
- Wanke, M.; Skorupinska-Tudek, K.; Swiezewska, E. Isoprenoid biosynthesis via 1-deoxy-D-xylulose 5-phosphate/2-C-methyl-D-erythritol 4-phosphate (DOXP/MEP) pathway. Acta Biochem. Pol. 2001, 48, 663–672. [Google Scholar] [CrossRef]
- Dinan, L.; Harmatha, J.; Volodin, V.; Lafont, R. Phytoecdysteroids: Diversity, biosynthesis and distribution. In Ecdysomes: Structures and functions, 1st ed.; Smagghe, G., Ed.; 2009; pp. 3–45.
- Lichtenthaler, H.K. The Non-mevalonate DOXP/MEP (Deoxyxylulose 5-Phosphate/Methylerythritol 4-Phosphate) Pathway of Chloroplast Isoprenoid and Pigment Biosynthesis. In The Chloroplast. Advances in Photosynthesis and Respiration; Rebeiz, C.A., et al., Eds.; Springer: Dordrecht, 2010; Volume 31, pp. 95–118. [Google Scholar]
Figure 1.
Nematode cuticle during molting (A) and lipid-rich epicuticle and protein-rich sublayers (B) from transmission electron microscopy photograph of Caenorhabditis elegans (Improved by Dr E. Shokoohi).
Figure 1.
Nematode cuticle during molting (A) and lipid-rich epicuticle and protein-rich sublayers (B) from transmission electron microscopy photograph of Caenorhabditis elegans (Improved by Dr E. Shokoohi).
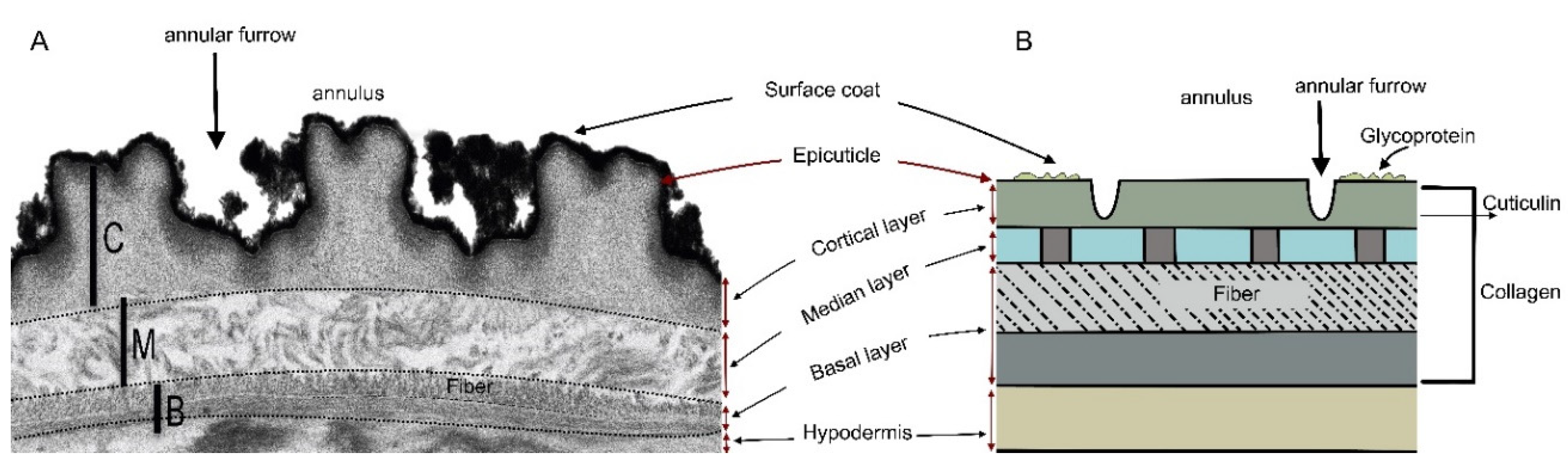
Figure 2.
Isoprenylation and farnesylation in Meloidogyne enterolobii induced by Nemafric-BL phytonematicide.
Figure 2.
Isoprenylation and farnesylation in Meloidogyne enterolobii induced by Nemafric-BL phytonematicide.
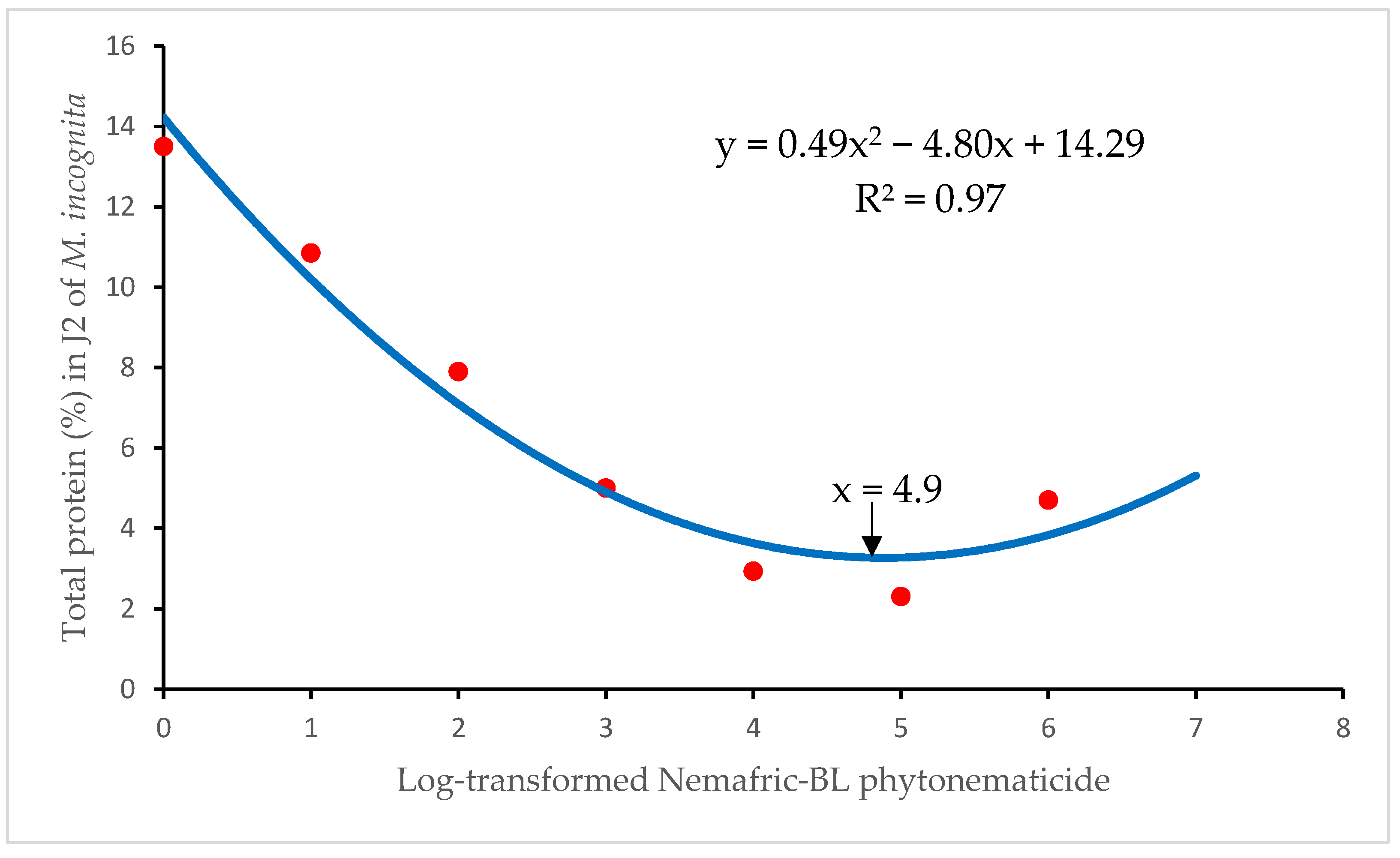
Figure 3.
Structure of the prenyl group.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



