Submitted:
21 March 2024
Posted:
22 March 2024
You are already at the latest version
Abstract
Soil microorganisms plays an important role on plant development and the homogenization of soil microbiome is agri-environmental harmful. It is essential that agricultural practices are carried out taking soil microbiome preservation in consideration. Agroforestry systems are one of the most environmentally friendly agrosystems and its plant diversity influence directly the soil microbiome diversity. In addition, many technologies have been developed and used by applying inoculants in agriculture. However, most of the inoculants are developed with only one or a little number of microbial species to supply a specific plant nutritional demand. In this study we tested the efficacy of the microbial consortium (MC) obtained from compost and the cyanobacteria Arthrospira platensis (AP) comparing with the application of the vermicompost tea (VT) and bokashi (BK) in arugula, lettuce, beetroot, and carrot in an agroforestry system in two seasons. Soil changes on chemical properties and β-glucosidase and arylsulfatase were measured over time. The vegetables yield was measured and for arugula and lettuce the nutritional traits and plant root system were also evaluated. Greater plant yield, nutritional values, and plant root development was observed in the CM treated plants, AP and BK had similar results. Our results show that both CM and AP have potential to become a sustainable product for agricultural production
Keywords:
Inoculant
; soil biodiversity
; agroforestry system
1. Introduction
Agricultural practices have been intensified over the last decades causing harmful impacts on the environment [1]. However, it is known how much conventional agriculture contributes to global warming, due to the inadequate management of natural assets such as soil and water, associated with greenhouse gas emissions [2]. With the justification that it is necessary to increase food production due to population growth [3], there was no concern about the depletion of our natural resources, such as soil and water, fundamental for all types of life on the planet.
Agroforestry systems (AFS) are one of the most environmentally friendly agrosystems, where perennial woody plants (trees, shrubs, palm trees) are grown in association with herbaceous plants, agricultural and/or forage crops and/or with animals in the same area and can be planted all at once or in a time sequence, with high diversity of species and ecological interactions, favouring a more efficient recycling of nutrients, being able to restore soil health [4,5]. The trees, with deep roots, recapture and pump back leached nutrients, the biomass deposited in the soil by falling leaves, pruning branches and residues from annual crops improves the supply of nutrients and in addition to the root exudates favour the action of beneficial microorganisms in the soil [6,7].
Several microorganisms can interact with roots and promote plant growth [8,9]. These microorganisms have been studied for decades and biotechnological products have been developed using plant growth promoting microorganisms (PGPM) in agriculture as inoculants. This biotechnology is very important to the Brazilian agriculture [10]. In maize, for example, it is possible to replace in the country 25% of N chemical fertilizers by inoculation with Azospirillum brasilense, saving US$ 15 ha-1 and avoiding the emission of 236 kg of CO2e ha-1 [11]. In soybeans the replacement is by 100% using Bradyrhizobium spp [12]. There are researches with inoculation in horticulture, but not so far as it is in the extensive crops [12].
Mira et al. (2021) [13] isolated a microbial consortium from compost and inoculated in arugula, carrots, and radish obtaining great results on these three cultures. This microbial consortium differs from the conventional inoculants since it is inoculated a microbial community and not an isolated strain, it has potential to become a commercial inoculant, but more tests should be done. Another type of PGPM tested in horticulture are the cyanobacteria due to their ability to fix atmospheric nitrogen, solubilizing phosphorus and promote plant growth [14,15]. Also, they have potential to be applied as soils conditioners since they can improve soil structure and water retention [16], as well as the availability of nutrients [17,18].
In horticulture is common to use organic fertilizers to improve the productivity in a sustainable way. Vermicompost tea (worm tea) is a leachate obtained below the vermicomposting heap and has been used as organic fertilizer [19,20]. Bokashi is an organic fertilizer with a high content of organic matter and nutrients, high porosity and a high water retention capacity [21]. In addition, it contributes to soil fertility with different types of microorganisms such as fungi and bacteria, increasing life in the soil [22,23].
We tested the efficacy of the microbial consortium presented by Mira et al. (2021) [13] and the cyanobacteria Arthrospira platensis comparing with the application of the vermicompost tea and bokashi in arugula, lettuce, beetroot, and carrot in an agroforestry system implemented in an area that was used for construction debris deposit. Based on the literature and our previous pilot trials, we hypothesized that both the microbial consortium and A. platensis could show good results in vegetables development being potential for new inoculant development for horticulture.
2. Materials and Methods
2.1. Site Description and Area Preparation
The current study was conducted at the Social Organization Célio Lemos (Obra Social Célio Lemos) in São José dos Campos, State of São Paulo, Brazil. From April 2021 to August 2022, at the coordinates 23°11’21” S and 45°52’17” W, 596 m altitude. The study area presented a history of being used for deposing construction debris, also, one cinnamon and one eucalyptus trees were present there. The soil was classified as Dystrophic Red-Yellow Latosol according to the Brazilian classification system [24]. The climate is classified as Cwa (humid subtropical) according to Köppen-Geiger classification.
Firstly, the area was cleaned removing the rubble using a truck and cutting the cinnamon and eucalyptus trees. A 00-10 cm soil sample, composed by 15 soil subsamples was collected on May 27th, 2021, and sent to Ribersolo Laboratory in Ribeirão Preto, São Paulo State, Brazil, for determination of soil chemical and granulometric initial properties (Table 1). With a garden bedder and tilling the soil with a tiller, 10 garden beds spaced by 1 m, with 1 m x 16 m of size each, were built. No chemicals were applied in the soil. Using rakes, some rocks from the rubble remaining in the soil were removed.
A mix of seeds of green manure plants was sown by broadcast sowing on August 11th, 2021. The mix included: Avena strigosa (7.0 g/m2), Lupinus albus (6.5 g/ m2), Cajanus cajan (4.0 g/ m2), Crotalaria juncea (3.2 g/ m2), Helianthus annus (2.0 g/ m2), and Raphanus sativus (1.0 g/ m2). The seeds were mixed with garden soil (amount equivalent to 100 g/m2 of the total area). On October 22nd, 2021, the plants were cut, crushed, and placed on the garden beds for covering. Additionally, plant material from tree pruning carried out by the city hall was donated for the project and used for soil covering, the weight of the plant material laid in the soil was not measured, but it was sufficient to form a layer of approximately 5 cm of litter. This material was laid on the top of the beds and between them. The study can be divided in three stages: (1) area preparation and cover crop cultivation, (2) first experimental cycle, and (3) second experimental cycle.
2.2. Experimental Design
The experiment was conducted in a completely randomized blocks with five treatments: (C) Control; (CT) 0.5 L/ m2 of vermicompost tea diluted in the proportion 1:15; (Ap) 0.5 L/ m2 of Artrhospira platensis cultivated in CT; (MC) 0.5 L/ m2 of microbial consortium obtained from compost (1.2 x 109 cells/mL) [13]; (Bk) 1.0 kg/ m2 of Bokashi. Each block consisted of two rows of garden bed with 16 m2, each plot has 2 m of length spaced by 1 m length from each other, a space of 0.5 m of length in the ends of the garden beds was considered (Figure 1a). The vegetables were planted at the same time in a consortium including pepper, a single plant row spaced 1 m apart; lettuce, spaced by 0.3 m x 0.3 m; arugula, six plants spaced by 0.15 m between the pepper and lettuce row; carrot, seeds were applied in a row with 0.15 m from two rows of lettuce; beetroot, six plants spaced by 0.15 m between the lettuce row and pepper (Figure 1b). All plants were planted by seedlings, except the carrots which were planted by seeds. Around 5 days after the emergence, carrots were thinned to 6 plants per row. For the forestry component, two garden beds (one in the block 2 and other in the block 4) received some plant trees, Sphagneticola trilobata, Gliricidia sp, Musa paradisiaca, Ricinus communis, Diospyros kaki, Caesalpina pluviosa, Tabebuia roseo-alba, Mangirefa indica, Melia azedarach, Cajanus cajan and Citrus sp. It was conducted two planting and harvesting cycles. The first started on October 25th, 2021, and the second on May 5th, 2022.
2.3. Inoculum Preparation
The microbial consortium was the same that Mira et al. (2021) obtained from compost. A cryopreserved sample in Tryptic Soybean Broth (TSB) culture medium with 30% glycerol was reactivated by inoculating in 100 mL of TSB and maintaining at 30 °C for 72 h. Then, the reactivated culture was used to produce the inoculum for the experiment, 1 mL of the reactivated cells was transferred to several flasks containing 300 mL of TSB which were incubated at 30 °C for 72 h to get the amount necessary for the treatments. The cell concentration in the suspension was adjusted to get 1.2 x 109 cells/mL with the quantitative nephelometry test [25].
The Arthrospira platensis strain BMAKD7 was obtained from the “Aidar & Kutner Microorganism Bank of the Oceanographic Institute of the University of São Paulo”. The cyanobacteria were inoculated in 20 L of 1:5 diluted vermicompost tea in water without sterilization, also it was added 2.5 g/L of NaNO3 and 16 g/L of NaHCO3 to the medium. It was assembled a photobioreactor using an organizer box with 29 L of capacity, on the lid of the box an 18 W LED lamp was attached, it was connected to a timer programmed for 12 h of light time per day. Considering the variations in luminosity in the laboratory and the different depths at which the cells were positioned inside the box, a variation from 1570 to 2500 lux was measured without the presence of culture media and cells, which influence light absorption. The range of luminosity obtained is within good conditions established for cell growth [26]. The aeration was controlled using a pump and the system worked for 14 days at room temperature for the cyanobacterial growing. For the field application the cyanobacterial suspension was diluted in water in the proportion of 1:3 to give a 1:15 final dilution for the vermicompost tea.
It was utilized an artisanal Bokashi prepared by the local farmer at the experimental site for the experiments, it was used 45 kg of garden soil, 10 kg of rice straw, 4 kg of rice bran, 16 kg of manure, 1.4 kg of crushed coal, 800 g of yorin, 400 g of phosphite. The ingredients were mixed with 10 L of a mixture of 5 L of the E.M. suspension, 5 kg of sugar cane molasse and 100 L of water. E.M. were produced from samples of fungi isolated from an Atlantic Forest litter in the region of São José dos Campos, using cooked rice as bait. After 7 days, the colonies grown on cooked rice were selected by their colours, the dark ones were eliminated and the others were used as inoculum in a broth medium based on sugar cane molasses, milk and rock dust.
2.3. Data Collecting and Analysis
Randomly, taking care to preserve the plants roots systems it was harvested six plants from each plot for each specie, except for the pepper, which was harvested just the fruits. The arugula was harvested 28 days after planting (DAP), lettuce 45 DAP, beetroot 90 DAP, carrot 120 DAP, and pepper 120 DAP. The plants were transported to the laboratory, where it was made biometric measures. Firstly, the shoots and roots were separated, then it was measured the shoot and root fresh weigh for all the plants. For the lettuce and arugula, root volume was measured by immersion and displacement of water in a graduated cylinder. Total root length was assessed in fresh roots by the modified line-intersection method [27]. Root mean diameter (D) was calculated by the formula 2(V/Lπ)0.5 and root superficial area by the formula 2πRL + 2πR2, where V, L, and R represent the root system volume, total root length, and root mean radius (D/2), respectively [28]. The carrots root length was measured using a rule. Then, the shoots and roots were dried at 65 °C until getting constant mass. Shoot and roots dry weigh were measured. Due to budget limitations, just 3 repetitions (2, 3 and 4) of the arugula and lettuce in the first experiment were sent to bromatological analysis by the Ribersolo laboratory in Ribeirão Preto, São Paulo State. In the second experiment, bromatological analysis was not made.
2.3. Statistical Analysis
All data obtained were tested for the normality of variables and variance homogeneity, followed by an analysis of variance (ANOVA) at p < 0.05 and then means were compared by the Duncan test at p < 0.05. In cases where the variables did not show normality and/or variance homogeneity, data were transformed to √(x+1). These tests were performed using the ExpDes package of the R software (version 4.2.2, 2022). Data from biometric analysis were transformed into log(x+1) for clustering, the number of clusters were determined by the function “fviz_nbclust”, then the clusters representations were generated with “fviz_cluster” function, both from the “factoextra” package in R.
3. Results
In general, the first experiment showed more differences between the treatments than the second. In most cases, the vegetables were benefited by the Ap, MC, and Bk treatments, with a highlight to MC. In arugula, these treatments resulted in more shoot dry weigh in the first and second experiment (Table 2). Shoot dry weight in MC treated in the second experiment is the only of these three treatments mentioned that differs from VT. Furthermore, MC has more root dry weight and volume than the others in the first experiment. In the second experiment Ap, MC, and Bk did not differ from each other in relation to root dry weight and volume, but it differs from the control group. MC treatment also resulted in longer roots and in more root surface area in the first experiment. The treatments did not differ from each other in the root mean diameter. The Bk treated arugulas shown the lowest root specific length and the major root density values in the first experiment. The clustering plots based on the biometric analysis (Figure 2) shows two groups of treatments, the first comprises Ap, MC, and Bk, and the second comprises C and VT.
Means followed by different letters differ according to the Duncan test (p < 0.05).
In lettuce the treatments only shown differences between each other in the first experiment (Table 3). MC treated lettuces had more shoot and root dry weight and root volume than the others. Ap, MC, and Bk did not differ from each other in root system length, but it was longer than C and VT. There was no difference in root mean diameter between all the treatments. MC had the major root surface area, followed by Ap and Bk, and the C and VT. Only Ap and Bk differ from the control group in the root specific length. There were no differences between the treatments in root density. As it was for arugula, the treatments Ap, CM, and Bk in lettuce were like each other, being clustered separated from C and VT (Figure 3).
The bromatology analysis, performed in the first experiment, showed that, except for the Zn total content in the Ap treatment, Ap and Bk had always higher nutrient and protein content than C and VT. In relation to MC, these treatments were sometimes lower than it and sometimes similar to it in nutrient and protein content (Table 4). As it occurred on the arugulas, the bromatology analysis, performed in the first experiment, showed that, except for the Zn total content in the Ap treatment, Ap and Bk had always higher nutrient and protein content than C and VT. The MC treatment had, in general, higher nutrient content than the others, being like Ap and Bk in some of those (Table 4).
In the first experiment, beetroot treated with Ap, MC, and Bk resulted in more shoot fresh weight and root dry weigh comparing to the control group but did not differ from the VT group (Table 5). In the second experiment there was no differences between the treatments in beetroots. The treatments in carrots did not differ from each other in the first experiments but in the second VT treated carrots had more dry weight (Table 5).
4. Discussion
The maintenance of soil quality has a dependence of the soil microbial communities. Building a microbial functional diversity and redundancy is a key factor for a stable plant-soil system. This can be reached by adopting several sustainable practices such no tilling, soil covering, no or low inputs of chemicals, culture rotation, and microorganism application [29,30,31]. In the present study it was presented microbial treatments for using in agriculture. The microbial consortium has a biodiversity that came from the compost, bokashi has a microbial diversity which came from production process and the EM, and vermicompost tea has also microorganism diversity [32]. Although A. platensis is a single organism, it is known that cyanobacteria can stimulate soil microbiota diversity, moreover, it was used non sterilized vermicompost tea for the production, which is supposed to carry other beneficial microorganisms [32,33]. The initial hypothesis was that these microorganisms could improve the plants development and soil quality.
Plant growth promoting bacteria can induce improvements in morphological root traits by producing plant growth regulators and signaling molecules in the rhizosphere [34,35,36,37]. With a longer root system and more root surface area, the plant can scavenge more efficiently the soil nutrients, especially those with low motility in the soil [38,39]. Root tissue density is related with how the plant interact with the soil, even having impact in soil carbon dynamic. Specific root exudation rate (amount of root exudate per gram of dry root over time) is decreased with high root tissue density [40]. Root exudates represent a chemical interface for the communication between the plant and microorganisms, which will use it as energy source [41]. Also, high root tissue density is negatively correlated with root life span, it means a faster degradation of roots with lower tissue density in soil [42]. Regarding the plant physiology, low root tissue density may indicate less metabolic expenditure with the respiratory rate of the roots and faster shoot growth rate [43]. On other hand, plant can reduce root tissue density by the formation of air spaces in the root cortex when it has low nitrogen or phosphorus availability [2,44]. C and VT treatments had conditions that led to low general development comparing with the other treatments in the first experiment with arugula, one hypothesis is that these plants were in an environment with less plant growth promoting bacteria, reducing, for example, the N and P uptake. In the MC treatment, the low root tissue density was accompanied with a good plant development comparing with the other treatments, in this case root tissue density reflected not a worse plant development but a reduction on the necessity of high respiration rates by the roots. Although the treatments Ap, MC, and Bk did not show thinner roots than the control group on arugula and lettuce, in general, they had more root length, more root surface area, and less root specific length. These results may be attributed to the inoculated microorganisms and it leaded to a greater plant development and nutrient uptake when comparing to the control group.
Our results show that the A. platensis suspension in general improved the plants development and can also be used for agricultural systems improvements. There are corroborations in the literature with related benefits due to the inoculation of A. platensis in arugula, eggplant, lettuce, maize, and onion [45,46,47,48]. Micronutrient concentration increasing in plant parts has also been reported [49,50]. In the present work, A. platensis increased macro and micronutrients in arugula and lettuce, when compared with the control and VT groups, excepting for Zn in arugula, these results can be explained due to the roots development improving, which aids the plants to explore more efficiently the soil. The same rational thinking can be applied to the CM and Bk treatments.
The first trial with the microbial consortium used in this study was carried out by Mira et al. (2021) [13]. The authors isolated the microbial consortium from compost and applied in arugula, carrots, and radish. The MC improved significatively the fresh mass of the arugula, carrots, and radish in 2.5, 1.3, and 16 times, respectively. The increase in the arugula shoot dry weight in 2.98 and 1.94 times in the first and second experiment of the present study corroborates with it. On the other hand, while Mira et al. (2021) [13] related an increase in carrots biomass, the present results do not show that. Although the authors made just one application of the MC, it can be worthy to do another application in the middle of the growing period, since carrots takes a longer time to be harvested than arugula and lettuce. This approach of applying the microorganisms more than one time could also be adopted for the Ap treatment, Dias et al. (2016) [45] for example did four applications of A. platensis in an experiment with eggplant.
Bokashi, as an organic fertilizer, has a high content of organic matter and nutrients, in addition to the microorganisms [21]. The results with bokashi application show a general improvement on plan development when compared to the control group (Table 1, Table 2 and Table 3). Controversially, Mayer et al. (2010) [51] did not observe yield increase in a four-year experiment with potato, winter barley, alfalfa, and winter wheat, each one in a year as a rotation in temperate climate. The authors related some effects on soil traits but attributed it to the nutrient content in bokashi rather than the microorganisms, since the autoclaved bokashi treated group had similar results.
The use of bokashi through badly executed management can harm a crop. When the material is added to a substrate for seed germination or even when applied to the soil, the decomposition of the material continues and the negative impacts are caused by the decline in oxygen availability, N immobilization [52] or by the phytotoxicity generated by the presence of phytotoxic compounds [21]. Organic acids and ammonium are examples of these compounds and are generated when the raw material has a low C/N ratio [53]. Thus, it is recommended that biological tests be performed as quality control to assess the phytotoxicity of the product [21].
Therefore, adopting the inoculation of CM or A. platensis instead of Bokashi can be better for crop safety and yield, since the results shows better results to CM and similar results to Ap treatment comparing to Bokashi. However, for using CM in agriculture as a product, it is needed more research on formulation and scaling bioprocess, since using the TSB medium for growing the consortium is not economically worthy.
Author Contributions
Conceived and designed the experiments: TFR, MPMI, VM, CV, EE. Performed the experiments: TFR, MPMI, GB, VARM, EE. Analyzed the data: TFR, MPMI, EE. Contributed reagents/materials/analysis tools: CV, EE. Wrote the paper: TRF, EE. All authors read and approved the final manuscript.
Acknowledgments
T. F. Rodrigues acknowledges a PhD fellowship from CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior— Brasil—Finance Code 001) and M.P.M. Itkes Msc fellowship from Cargill. This project was financed by the Cargill Sustainability Notice (2019).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Muller, A.; Schader, C.; El-Hage Scialabba, N.; Brüggemann, J.; Isensee, A.; Erb, K.-H.; Smith, P.; Klocke, P.; Leiber, F.; Stolze, M.; et al. Strategies for feeding the world more sustainably with organic agriculture. Nat. Commun. 2017, 8, 1290. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P.; Ho, M.D.; Phosphorus, L. Rhizoeconomics: Carbon costs of phosphorus acquisition. Plant Soil 2005, 269, 45–56. [Google Scholar] [CrossRef]
- Beltran-Peña, A.A.; Rosa, L.; D’odorico, P. Global food self-sufficiency in the 21st century under sustainable intensification of agriculture. Environ. Res. Lett. 2020, 15, 095004. [Google Scholar] [CrossRef]
- Miccolis, A.; Peneireiro, F.M.; Vieira, D.L.M.; Marques, H.R.; Hoffmann, M.R.M. RESTORATION THROUGH AGROFORESTRY: OPTIONS FOR RECONCILING LIVELIHOODS WITH CONSERVATION IN THE CERRADO AND CAATINGA BIOMES IN BRAZIL. Exp. Agric. 2017, 55, 208–225. [Google Scholar] [CrossRef]
- Matos, P.S.; Cherubin, M.R.; Damian, J.M.; Rocha, F.I.; Pereira, M.G.; Zonta, E. Short-term effects of agroforestry systems on soil health in Southeastern Brazil. Agrofor. Syst. 2022, 96, 897–908. [Google Scholar] [CrossRef]
- Beule, L.; Vaupel, A.; Moran-Rodas, V.E. Abundance, Diversity, and Function of Soil Microorganisms in Temperate Alley-Cropping Agroforestry Systems: A Review. Microorganisms 2022, 10, 616. [Google Scholar] [CrossRef]
- Fahad, S.; Chavan, S.B.; Chichaghare, A.R.; Uthappa, A.R.; Kumar, M.; Kakade, V.; Pradhan, A.; Jinger, D.; Rawale, G.; Yadav, D.K.; et al. Agroforestry Systems for Soil Health Improvement and Maintenance. Sustainability 2022, 14, 14877. [Google Scholar] [CrossRef]
- de Souza, R.; Ambrosini, A.; Passaglia, L.M. Plant growth-promoting bacteria as inoculants in agricultural soils. Genet. Mol. Biol. 2015, 38, 401–419. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.J.d.S.; Dias-Filho, M.B.; Gurgel, E.S.C. Successful Plant Growth-Promoting Microbes: Inoculation Methods and Abiotic Factors. Front. Sustain. Food Syst. 2021, 5. [Google Scholar] [CrossRef]
- Bomfim, C.A.; Coelho, L.G.F.; Vale, H.M.M.D.; Mendes, I.d.C.; Megías, M.; Ollero, F.J.; Junior, F.B.d.R. Brief history of biofertilizers in Brazil: from conventional approaches to new biotechnological solutions. Braz. J. Microbiol. 2021, 52, 2215–2232. [Google Scholar] [CrossRef]
- Hungria, M.; Barbosa, J.Z.; Rondina, A.B.L.; Nogueira, M.A. Improving maize sustainability with partial replacement of N fertilizers by inoculation with Azospirillum brasilense. Agron. J. 2022, 114, 2969–2980. [Google Scholar] [CrossRef]
- Santos, M.S.; Nogueira, M.A.; Hungria, M. Microbial inoculants: reviewing the past, discussing the present and previewing an outstanding future for the use of beneficial bacteria in agriculture. AMB Express 2019, 9, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Mira, W.V.d.M.; Esposito, E.; Vilarraga, C.O.; Anatriello, E. APLICAÇÃO DE BIOINOCULANTES DE SOLO COMPOSTADO PARA PROMOÇÃO DO CRESCIMENTO DE RÚCULA (Eruca sativa), CENOURA (Daucus carota sativus) e RABANETE (Raphanus sativus). Rev. Bras. de Agroecol. 2021, 16, 117–122. [Google Scholar] [CrossRef]
- Kuraganti G, Edla S, Pallaval VB. Cyanobacteria as Biofertilizers: Current Research, Commercial Aspects, and Future Challenges, 2020, p. 259–78. [CrossRef]
- Zapata, D.; Arroyave, C.; Cardona, L.; Aristizábal, A.; Poschenrieder, C.; Llugany, M. Phytohormone production and morphology of Spirulina platensis grown in dairy wastewaters. Algal Res. 2021, 59, 102469. [Google Scholar] [CrossRef]
- Rossi, F.; Li, H.; Liu, Y.; De Philippis, R. Cyanobacterial inoculation (cyanobacterisation): Perspectives for the development of a standardized multifunctional technology for soil fertilization and desertification reversal. Earth-Science Rev. 2017, 171, 28–43. [Google Scholar] [CrossRef]
- Alobwede, E.; Leake, J.R.; Pandhal, J. Circular economy fertilization: Testing micro and macro algal species as soil improvers and nutrient sources for crop production in greenhouse and field conditions. Geoderma 2018, 334, 113–123. [Google Scholar] [CrossRef]
- Manjunath, M.; Kanchan, A.; Ranjan, K.; Venkatachalam, S.; Prasanna, R.; Ramakrishnan, B.; Hossain, F.; Nain, L.; Shivay, Y.S.; Rai, A.B.; et al. Beneficial cyanobacteria and eubacteria synergistically enhance bioavailability of soil nutrients and yield of okra. Heliyon 2016, 2, e00066. [Google Scholar] [CrossRef] [PubMed]
- Churilova, E.V.; Midmore, D.J. Vermiliquer (Vermicompost Leachate) as a Complete Liquid Fertilizer for Hydroponically-Grown Pak Choi (Brassica chinensis L.) in the Tropics. Horticulturae 2019, 5, 26. [Google Scholar] [CrossRef]
- Helaly, A.A.E.-H.; El-Dakak, R. Effect of Organic Liquid Vermicompost as a Substitute for Chemical Fertilizer on Morphological and Biochemical Characteristics in Lettuce. Assiut J. Agric. Sci. 2021, 52, 69–81. [Google Scholar] [CrossRef]
- Quiroz, M.; Céspedes, C. Bokashi as an Amendment and Source of Nitrogen in Sustainable Agricultural Systems: a Review. J. Soil Sci. Plant Nutr. 2019, 19, 237–248. [Google Scholar] [CrossRef]
- Restrepo J, Hensel J. El ABC de la agricultura orgánica, fosfitos y panes de piedra. Cali, Colombia,© Jairo Restrepo Rivera 2015.
- Wilmer J, Rodriguez W, Rosas G. CARACTERIZACIÓN FÍSICA Y QUÍMICA DE BOKASHI Y LOMBRICOMPOST Y SU EVALUACIÓN AGRONÓMICA EN PLANTAS DE MAÍZ 2019:2014.
- Santos HG, Jacomine PKT, Anjos LHC, Oliveira VA, Lumbreras JF, Coelho MR, et al. Brazilian Soil Classification System. 5th ed. Brasília: Embrapa; 2018.
- Morais, I.P.A.; Tóth, I.V.; Rangel, A.O.S.S. Turbidimetric and Nephelometric Flow Analysis: Concepts and Applications. Spectrosc. Lett. 2006, 39, 547–579. [Google Scholar] [CrossRef]
- Muliterno, A.; Mosele, P.C.; Costa, J.A.V.; Hemkemeier, M.; Bertolin, T.E.; Colla, L.M. Cultivo mixotrófico da microalga Spirulina platensis em batelada alimentada. 2005, 29, 1132–1138. [CrossRef]
- Tennant, D. A Test of a Modified Line Intersect Method of Estimating Root Length. J. Ecol. 1975, 63, 995. [Google Scholar] [CrossRef]
- Zangaro, W.; Rostirola, L.V.; de Souza, P.B.; Alves, R.d.A.; Lescano, L.E.A.M.; Rondina, A.B.L.; Nogueira, M.A.; Carrenho, R. Root colonization and spore abundance of arbuscular mycorrhizal fungi in distinct successional stages from an Atlantic rainforest biome in southern Brazil. Mycorrhiza 2012, 23, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Louca, S.; Polz, M.F.; Mazel, F.; Albright, M.B.; Huber, J.A.; O’Connor, M.I.; Ackermann, M.; Hahn, A.S.; Srivastava, D.S.; Crowe, S.A.; et al. Function and functional redundancy in microbial systems. Nat. Ecol. Evol. 2018, 2, 936–943. [Google Scholar] [CrossRef] [PubMed]
- Bertola, M.; Ferrarini, A.; Visioli, G. Improvement of Soil Microbial Diversity through Sustainable Agricultural Practices and Its Evaluation by -Omics Approaches: A Perspective for the Environment, Food Quality and Human Safety. Microorganisms 2021, 9, 1400. [Google Scholar] [CrossRef] [PubMed]
- Suman, J.; Rakshit, A.; Ogireddy, S.D.; Singh, S.; Gupta, C.; Chandrakala, J. Microbiome as a Key Player in Sustainable Agriculture and Human Health. Front. Soil Sci. 2022, 2. [Google Scholar] [CrossRef]
- Fritz, J.; Franke-Whittle, I.; Haindl, S.; Insam, H.; Braun, R. Microbiological community analysis of vermicompost tea and its influence on the growth of vegetables and cereals. Can. J. Microbiol. 2012, 58, 836–847. [Google Scholar] [CrossRef]
- Prasanna, R.; Kanchan, A.; Ramakrishnan, B.; Ranjan, K.; Venkatachalam, S.; Hossain, F.; Shivay, Y.S.; Krishnan, P.; Nain, L. Cyanobacteria-based bioinoculants influence growth and yields by modulating the microbial communities favourably in the rhizospheres of maize hybrids. Eur. J. Soil Biol. 2016, 75, 15–23. [Google Scholar] [CrossRef]
- Molina-Favero, C.; Creus, C.M.; Simontacchi, M.; Puntarulo, S.; Lamattina, L. Aerobic Nitric Oxide Production by Azospirillum brasilense Sp245 and Its Influence on Root Architecture in Tomato. Mol. Plant-Microbe Interactions® 2008, 21, 1001–1009. [Google Scholar] [CrossRef]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.-L.; Touraine, B.; Moënne-Loccoz, Y.; Muller, D.; Legendre, L.; Wisniewski-Dyé, F.; Prigent-Combaret, C. Plant growth-promoting rhizobacteria and root system functioning. Front. Plant Sci. 2013, 4, 356. [Google Scholar] [CrossRef]
- Cassán, F.; Vanderleyden, J.; Spaepen, S. Physiological and Agronomical Aspects of Phytohormone Production by Model Plant-Growth-Promoting Rhizobacteria (PGPR) Belonging to the Genus Azospirillum. J. Plant Growth Regul. 2013, 33, 440–459. [Google Scholar] [CrossRef]
- Fukami, J.; Cerezini, P.; Hungria, M. Azospirillum: benefits that go far beyond biological nitrogen fixation. AMB Express 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Hodge, A. The plastic plant: root responses to heterogeneous supplies of nutrients. New Phytol. 2004, 162, 9–24. [Google Scholar] [CrossRef]
- Comas, L.H.; Mueller, K.E.; Taylor, L.L.; Midford, P.E.; Callahan, H.S.; Beerling, D.J. Evolutionary Patterns and Biogeochemical Significance of Angiosperm Root Traits. Int. J. Plant Sci. 2012, 173, 584–595. [Google Scholar] [CrossRef]
- Williams, A.; Langridge, H.; Straathof, A.L.; Muhamadali, H.; Hollywood, K.A.; Goodacre, R.; de Vries, F.T. Root functional traits explain root exudation rate and composition across a range of grassland species. J. Ecol. 2021, 110, 21–33. [Google Scholar] [CrossRef]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant, Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef]
- Ryser, P. The Importance of Tissue Density for Growth and Life Span of Leaves and Roots: A Comparison of Five Ecologically Contrasting Grasses. Funct. Ecol. 1996, 10, 717. [Google Scholar] [CrossRef]
- Kramer-Walter, K.R.; Bellingham, P.J.; Millar, T.R.; Smissen, R.D.; Richardson, S.J.; Laughlin, D.C. Root traits are multidimensional: specific root length is independent from root tissue density and the plant economic spectrum. J. Ecol. 2016, 104, 1299–1310. [Google Scholar] [CrossRef]
- McPherson, D.C. CORTICAL AIR SPACES IN THE ROOTS OF ZEA MAYS L. New Phytol. 1939, 38, 190–202. [Google Scholar] [CrossRef]
- Dias, G.A.; Rocha, R.H.C.; Araújo, J.L.; Lima, J.F.; Guedes, W.A. Growth, yield, and postharvest quality in eggplant produced under different foliar fertilizer (Spirulina platensis) treatments. 2016, 37, 3893. [CrossRef]
- Dineshkumar, R.; Subramanian, J.; Arumugam, A.; Rasheeq, A.A.; Sampathkumar, P. Exploring the Microalgae Biofertilizer Effect on Onion Cultivation by Field Experiment. Waste Biomass- Valorization 2018, 11, 77–87. [Google Scholar] [CrossRef]
- Siringi, J.; Turoop, L.; Njonge, F. BIOSTIMULANT EFFECT OF SPIRULINA (ARTHROSPIRA PLATENSIS) ON LETTUCE (LACTUCA SATIVA) CULTIVATED UNDER AQUAPONIC SYSTEM. . 2022. [Google Scholar] [CrossRef]
- Wuang, S.C.; Khin, M.C.; Chua, P.Q.D.; Luo, Y.D. Use of Spirulina biomass produced from treatment of aquaculture wastewater as agricultural fertilizers. Algal Res. 2016, 15, 59–64. [Google Scholar] [CrossRef]
- Prasanna, R.; Bidyarani, N.; Babu, S.; Hossain, F.; Shivay, Y.S.; Nain, L. Cyanobacterial inoculation elicits plant defense response and enhanced Zn mobilization in maize hybrids. Cogent Food Agric. 2015, 1. [Google Scholar] [CrossRef]
- Rana, A.; Joshi, M.; Prasanna, R.; Shivay, Y.S.; Nain, L. Biofortification of wheat through inoculation of plant growth promoting rhizobacteria and cyanobacteria. Eur. J. Soil Biol. 2012, 50, 118–126. [Google Scholar] [CrossRef]
- Mayer, J.; Scheid, S.; Widmer, F.; Fließbach, A.; Oberholzer, H.-R. How effective are ‘Effective microorganisms® (EM)’? Results from a field study in temperate climate. Appl. Soil Ecol. 2010, 46, 230–239. [Google Scholar] [CrossRef]
- Bernal, M.; Alburquerque, J.; Moral, R. Composting of animal manures and chemical criteria for compost maturity assessment. A review. Bioresour. Technol. 2009, 100, 5444–5453. [Google Scholar] [CrossRef]
- Zucconi F, Monaco A, Forte M. Phytotoxins during the stabilization of organic matter, 1985.
Figure 1.
(a) Experimental design croquis and (b) vegetables arrangement in a plot. C = control; VT = vermicompost tea; Ap = Artrhospira platensis; MC = microbial consortium; Bk = bokashi. The highlighted lines in (a) indicate where the trees were planted.
Figure 1.
(a) Experimental design croquis and (b) vegetables arrangement in a plot. C = control; VT = vermicompost tea; Ap = Artrhospira platensis; MC = microbial consortium; Bk = bokashi. The highlighted lines in (a) indicate where the trees were planted.
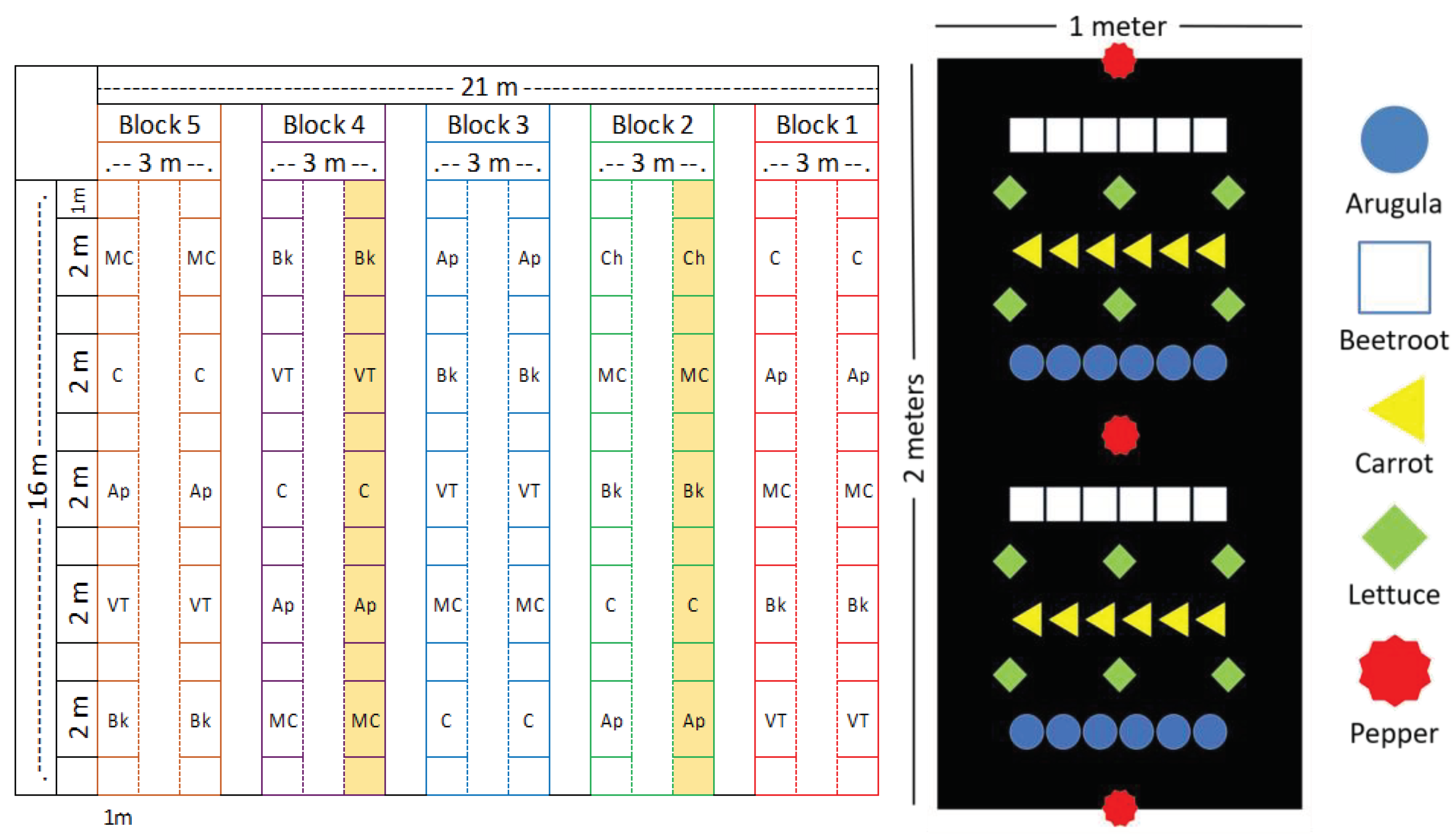
Figure 2.
Clustered treatments based on biometric data (Table 2 and Table 3) for the first experiment with arugula (a) and lettuce (b) and the second experiment with arugula (c) and lettuce (d). C = control; VT = vermicompost tea; Ap = Artrhospira platensis; MC = microbial consortium; Bk = Bokashi.
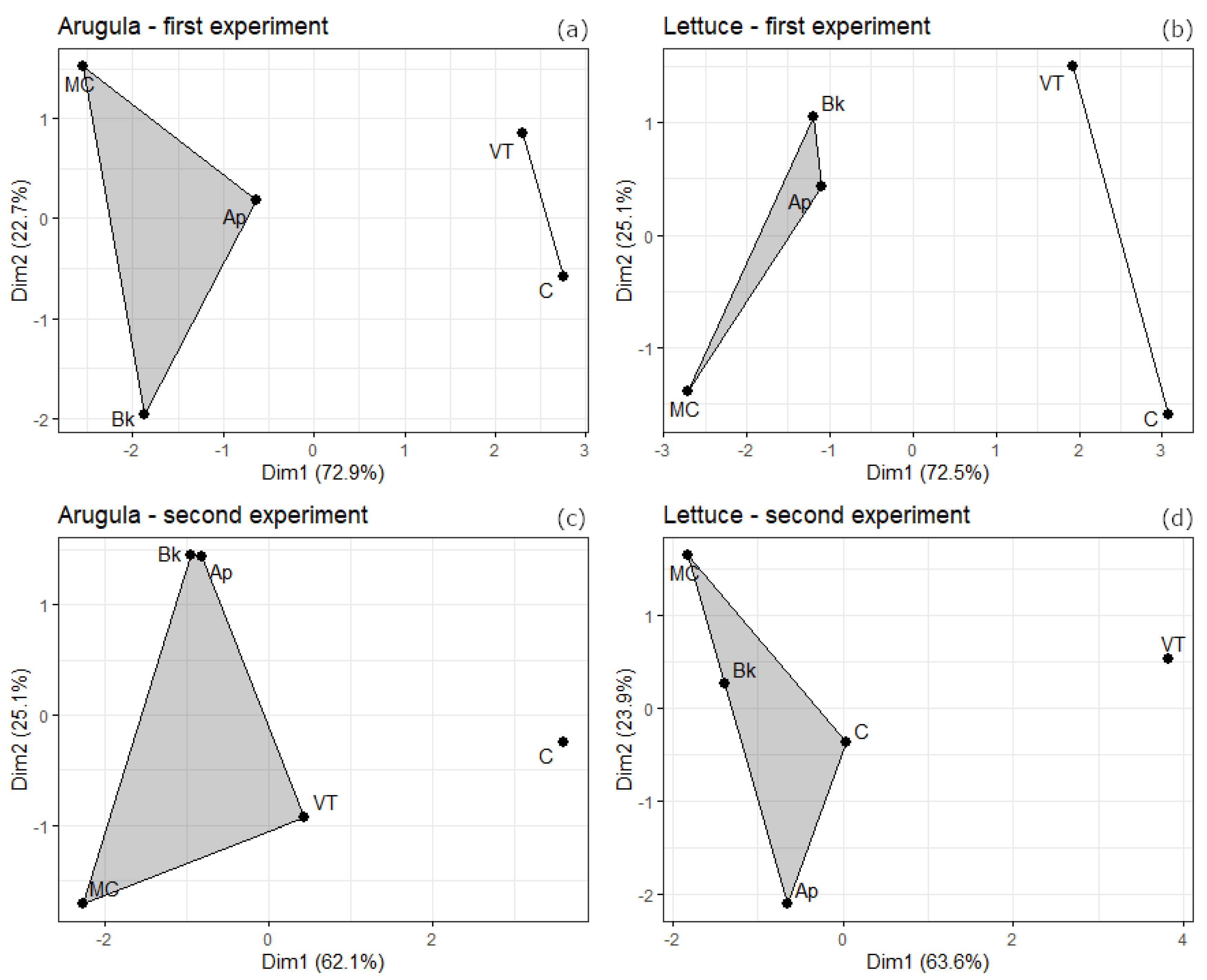

Table 1.
Soil chemical properties before starting the experiments.
| pH (CaCl2) | OM | P - res | K | Ca | Mg | H+Al | Al | SB | CEC | V | m | S | B | Cu | Fe | Mn | Zn | |||||
| g dm-3 | mg dm-3 | mmolc dm-3 | % | mg dm-3 | ||||||||||||||||||
| 6.5 | 17 | 26 | 3.1 | 52 | 9 | 14 | <0.7 | 64.1 | 78 | 82 | 1 | 10.0 | 0.22 | 2.50 | 27.0 | 1.90 | 6.30 | |||||
SB sum of bases; CEC cation exchange capacity.
Table 2.
Means of the variables analysed in the productivity of arugula with five repetitions in different treatments in AFS.
Table 2.
Means of the variables analysed in the productivity of arugula with five repetitions in different treatments in AFS.
| Shoot Dry Weight | Root Dry Weight | Root Volume |
Root System Length | Root Mean Diameter | Root Surface Area | Root Specific Length | Root Density | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| g | cm3 | cm | mm | cm2 | m/g | mg/ cm3 | ||||||||||||||||
| First Experiment | C | 1.33 | b | 0.10 | b | 1.33 | b | 190.66 | b | 0.94 | 56.38 | b | 22.68 | ab | 72.72 | bc | ||||||
| VT | 1.48 | b | 0.10 | b | 1.67 | b | 235.71 | b | 0.93 | 70.12 | b | 25.67 | a | 60.20 | c | |||||||
| Ap | 2.82 | ab | 0.21 | a | 2.21 | b | 293.84 | ab | 0.99 | 88.49 | b | 15.45 | bc | 94.57 | ab | |||||||
| MC | 3.97 | a | 0.26 | a | 3.68 | a | 348.13 | a | 1.15 | 125.96 | a | 13.70 | bc | 73.38 | bc | |||||||
| Bk | 3.42 | a | 0.26 | a | 2.41 | b | 209.03 | b | 1.25 | 78.18 | b | 8.58 | c | 105.97 | a | |||||||
| C.V. (%) | 44.55 | 39.94 | 37.20 | 28.49 | 18.51 | 26.75 | 38.95 | 23.87 | ||||||||||||||
| Second Experiment | C | 1.16 | c | 0.05 | b | 0.83 | b | 197.73 | b | 0.71 | 44.64 | b | 44.60 | 120.16 | ||||||||
| VT | 1.83 | bc | 0.08 | ab | 1.43 | ab | 467.49 | a | 0.69 | 90.57 | a | 46.40 | 107.79 | |||||||||
| Ap | 2.76 | ab | 0.11 | a | 1.63 | a | 375.04 | a | 0.76 | 87.13 | a | 35.33 | 144.55 | |||||||||
| MC | 3.15 | a | 0.13 | a | 2.10 | a | 459.90 | a | 0.63 | 101.37 | a | 58.40 | 142.25 | |||||||||
| Bk | 2.45 | ab | 0.12 | a | 1.73 | a | 406.99 | a | 0.80 | 98.27 | a | 37.69 | 133.00 | |||||||||
| C.V. (%) | 39.70 | 34.70 | 34.68 | 18.19 | 21.63 | 17.32 | 36.46 | 22.72 | ||||||||||||||
C = control; VT = vermicompost tea; Ap = Artrhospira platensis; MC = microbial consortium; Bk = bokashi; CV = coefficient of variation.
Table 3.
Means of the variables analysed in the productivity of lettuce with five repetitions in different treatments in AFS.
Table 3.
Means of the variables analysed in the productivity of lettuce with five repetitions in different treatments in AFS.
| Shoot Dry Weigh | Root Dry Weigh | Root Volume | Root System Length | Root Mean Diameter | Root Surface Area | Root Specific Length | Root Density | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| g | cm3 | cm | mm | cm2 | m/g | mg/ cm3 | |||||||||||||||||
| First Experiment | C | 3.56 | b | 0.48 | b | 4.80 | b | 556.70 | b | 1.08 | 181.81 | c | 11.62 | b | 494.21 | ||||||||
| VT | 3.32 | b | 0.44 | b | 4.67 | b | 662.35 | b | 0.94 | 193.49 | c | 15.81 | ab | 269.30 | |||||||||
| Ap | 5.58 | ab | 0.58 | b | 6.33 | b | 1,083.88 | a | 0.86 | 292.74 | b | 18.87 | a | 365.36 | |||||||||
| MC | 7.34 | a | 0.85 | a | 8.47 | a | 1,396.03 | a | 0.91 | 380.65 | a | 16.41 | ab | 408.67 | |||||||||
| Bk | 5.00 | b | 0.60 | b | 6.43 | b | 1,133.92 | a | 0.85 | 301.84 | b | 18.85 | a | 301.19 | |||||||||
| C.V. (%) | 33.02 | 23.78 | 21.14 | 30.86 | 15.49 | 21.16 | 22.67 | 6.54* | |||||||||||||||
| Second Experiment | C | 2.01 | 0.25 | 3.17 | 719.44 | 0.75 | a | 168.82 | 32.35 | a | 129.85 | ||||||||||||
| VT | 1.89 | 0.18 | 2.40 | 503.10 | 1.03 | b | 148.50 | 17.88 | b | 132.56 | |||||||||||||
| Ap | 1.84 | 0.23 | 3.20 | 854.84 | 0.68 | a | 184.74 | 38.24 | a | 122.55 | |||||||||||||
| MC | 2.79 | 0.30 | 3.27 | 743.27 | 0.66 | a | 182.80 | 44.21 | a | 138.58 | |||||||||||||
| Bk | 2.32 | 0.32 | 3.67 | 856.41 | 0.69 | a | 148.20 | 35.05 | a | 129.65 | |||||||||||||
| C.V. (%) | 40.39 | 36.69 | 31.42 | 26.93 | 19.19 | 22.3 | 30.26 | 11.33 | |||||||||||||||
C = control; VT = vermicompost tea; Ap = Artrhospira platensis; MC = microbial consortium; Bk = bokashi; CV = coefficient of variation. * Statistical analysis performed with values transformed into √(x + 1). Means followed by different letters differ according to the Duncan test (p < 0.05).
Table 4.
Total content of macronutrients, micronutrients, and proteins in arugula with different AFS treatments.
Table 4.
Total content of macronutrients, micronutrients, and proteins in arugula with different AFS treatments.
| N | P | K | Ca | Mg | S | B | Cu | Fe | Mn | Zn | Proteins | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mg plant-1 | µg plant-1 | mg plant-1 | |||||||||||||||||||||||||
| Arugula | C | 49.68 | c | 3.30 | b | 66.72 | c | 42.85 | d | 5.36 | d | 23.21 | c | 35.38 | d | 13.52 | c | 1672.35 | c | 35.42 | c | 150.74 | c | 310.54 | c | ||
| Ch | 50.94 | c | 4.26 | b | 74.74 | c | 49.94 | d | 5.89 | d | 24.37 | c | 42.74 | d | 15.21 | c | 955.67 | c | 39.98 | c | 171.83 | c | 318.36 | c | |||
| Ap | 111.92 | b | 9.83 | a | 146.33 | b | 94.73 | c | 10.87 | c | 38.61 | b | 78.53 | c | 30.15 | b | 3359.18 | b | 76.09 | b | 243.86 | bc | 699.55 | b | |||
| CM | 136.48 | a | 11.10 | a | 201.62 | a | 126.99 | a | 15.15 | a | 62.66 | a | 110.72 | a | 44.84 | a | 7202.48 | a | 112.17 | a | 467.73 | a | 853.05 | a | |||
| Bk | 115.45 | b | 11.87 | a | 184.31 | ab | 109.33 | b | 13.07 | b | 55.59 | a | 95.20 | b | 33.80 | b | 3416.98 | b | 104.66 | a | 355.55 | ab | 721.59 | b | |||
| C.V. (%) | 9.18 | 25.95 | 18.52 | 5.35 | 10.50 | 10.65 | 10.51 | 9.53 | 20.77* | 11.51 | 21.54 | 9.18 | |||||||||||||||
| Lettuce | C | 85.36 | b | 14.77 | c | 191.99 | c | 44.75 | b | 9.41 | c | 12.59 | b | 67.08 | c | 42.43 | b | 474.05 | b | 62.10 | b | 235.01 | b | 533.42 | b | ||
| Ch | 91.10 | b | 13.02 | c | 194.39 | c | 45.90 | b | 9.48 | c | 11.84 | b | 69.34 | c | 39.21 | b | 470.76 | b | 60.04 | b | 224.63 | b | 569.45 | b | |||
| Ap | 138.70 | a | 22.74 | b | 306.38 | b | 70.57 | a | 13.40 | b | 17.97 | a | 113.12 | b | 69.96 | a | 837.25 | a | 106.98 | a | 347.37 | a | 866.83 | a | |||
| CM | 154.48 | a | 28.04 | a | 400.93 | a | 88.94 | a | 18.60 | a | 22.61 | a | 140.88 | a | 75.61 | a | 1113.28 | a | 143.14 | a | 431.12 | a | 965.74 | a | |||
| Bk | 110.04 | b | 21.23 | b | 283.13 | b | 67.51 | a | 14.35 | b | 18.37 | a | 104.08 | b | 68.44 | a | 1089.10 | a | 97.59 | a | 318.74 | a | 687.77 | b | |||
| CV (%) | 9.82 | 15.00 | 9.98 | 17.67 | 17.62 | 14.39 | 7.11 | 15.47 | 18.15 | 11.17* | 17.11 | 9.82 | |||||||||||||||
C = control; VT = vermicompost tea; Ap = Artrhospira platensis; MC = microbial consortium; Bk = bokashi; CV = coefficient of variation. * Statistical analysis performed with values transformed into √(x + 1). Means followed by different letters differ according to the Duncan test (p < 0.05).
Table 5.
Means of the variables analysed in the productivity of beetroot and carrot with five repetitions in different treatments in AFS.
Table 5.
Means of the variables analysed in the productivity of beetroot and carrot with five repetitions in different treatments in AFS.
| Beetroot | Carrot | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Shoot Fresh Weight | Root Fresh Weight | Shoot Fresh Weight | Root Fresh Weight | Lengh | |||||||
| g | g | cm | |||||||||
| First Experiment | C | 4.30 | b | 1.34 | b | 34.07 | 53.22 | 16.62 | |||
| CT | 7.27 | ab | 3.51 | ab | 43.79 | 63.65 | 16.18 | ||||
| Ap | 9.21 | a | 5.70 | a | 52.90 | 68.60 | 16.69 | ||||
| MC | 17.43 | a | 6.71 | a | 39.20 | 61.95 | 16.50 | ||||
| Bk | 10.32 | a | 6.50 | a | 49.26 | 63.72 | 15.57 | ||||
| C.V. (%) | 47.35* | 26.38** | 25.24** | 34.5 | 10.55 | ||||||
| Second Experiment | C | 17.77 | 4.37 | 8.53 | 36.13 | ab | 14.43 | ||||
| CT | 25.77 | 6.91 | 13.63 | 57.56 | a | 14.93 | |||||
| Ap | 19.85 | 5.67 | 10.05 | 44.55 | ab | 14.28 | |||||
| MC | 21.45 | 6.98 | 13.22 | 57.53 | a | 15.25 | |||||
| Bk | 19.63 | 6.62 | 6.99 | 28.72 | b | 13.68 | |||||
| C.V. (%) | 28.24** | 23.83** | 37.84 | 35.46 | 17.46 | ||||||
C = control; VT = vermicompost tea; Ap = Artrhospira platensis; MC = microbial consortium; Bk = bokashi; CV = coefficient of variation. * Statistical analysis performed with values transformed into 1/(x + 1) ** Statistical analysis performed with values transformed into √(x + 1). Means followed by different letters differ according to the Duncan test (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



