
Submitted:
19 March 2024
Posted:
19 March 2024
You are already at the latest version
Abstract
The pivotal role of the basolateral amygdala (BLA) in the emotional modulation of hippocampal plasticity and memory consolidation is well established. Specifically, multiple studies have demonstrated that the activation of the noradrenergic (NA) system within BLA governs these modulatory effects. However, most of the current evidence has been obtained by direct infusion of synthetic NA or beta-adrenergic agonists. In the present study, we aimed to investigate the effect of endogenous NA release in the BLA, induced by a natural aversive stimulus (coyote urine), on memory consolidation for a neutral hippocampal-dependent task. We combined a weak version of the object location task (OLT) with mild predator odor exposure (POE) for our experiments. To investigate the role of the BLA in memory modulation, a subset of the animals (Wistar rats) were treated with the non-selective beta-blocker propranolol at the end of the behavioral procedures. Hippocampal tissue was collected either 90 minutes after drug infusion or 24 hours after the OLT test. The samples were used to determine the levels of phosphorylated CREB (pCREB) and activity-regulated cytoskeleton-associated protein (Arc) - two molecular markers of experience-dependent changes in neuronal activity. The result suggests that POE is a behavioral paradigm suitable for studying the interaction between BLA and the hippocampus in memory prioritization and selectivity.
Keywords:
basolateral amygdala
; noradrenaline
; memory consolidation
; object location task
; predator odor exposure
; pCREB
; Arc
1. Introduction
Episodic memory is the ability to store information about events that have transpired in the surrounding environment [1,2]. By enabling animals to make memory-based predictions, avoid potential threats and find the most efficient ways to obtain vital resources, this unique cognitive faculty supports adaptability to the ever-changing environmental conditions [2]. Episodic memories requires the binding of information about the spatial/environmental context (“where”), the objects/landmarks encountered within the environment (“what”), the temporal order of appearance of the stimuli within the environment (“when”) and in most occasions, the presence of familiar and/or unknown conspecifics (“who”) [1,2]. The formation of associations between the different aspects of an event is highly-dependent on the hippocampus, specifically the dorsal section of the structure [1,2,3,4]. Newly formed memories are labile at first and their long-term storage requires the synthesis of specific plasticity-related proteins (PRPs) [5]. The protein-dependent process that governs the transition of a memory from its short-term form into its stable long-term one is collectively known as cellular (or initial) memory consolidation [5]. Importantly, hippocampal glutamatergic transmission, while indispensable for memory formation, could not alone trigger the expression of PRPs that characterizes initial memory consolidation. The catecholaminergic neurotransmitters, noradrenaline (NA) and dopamine (DA), as well as the glucocorticoid hormone cortisol (corticosterone in laboratory rodents) are essential for the long-term storage of memories [6,7,8]. Emotional salience and environmental novelty are two of the main conditions capable of evoking the physiological state required for the prioritization of information in the hippocampus [9,10,11]. During emotionally arousing and/or distinctly novel situations the pontine nucleus locus coeruleus rearranges the firing patterns of its neurons to increase the secretion of NA and DA in dorsal hippocampus [12,13,14]. Thus, events lacking emotional relevance or other salient characteristics (like novelty) may not activate catecholaminergic release in the dorsal hippocampus to a degree that will allow their long-term stabilization and availability for later retrieval [11].
Nevertheless, neutral stimuli encountered close to a salient event may also be significant since they could be used as predictors for particular aversive or rewarding outcomes in the future [11]. The behavioral equivalent of the “Synaptic tagging and capture hypothesis” [15], the “Behavioral tagging hypothesis”, is a learning and memory model that offers reliable evidence of how emotional or distinctly novel experiences could modulate the consolidation of closely occurring low-arousing or neutral events [16]. According to this hypothesis, the encoding of a weak event (memory about neutral or low-emotional behavioral tasks) closely before or after the encoding of a second, strong event (emotionally arousing behavioral task), will induce structural and functional modifications in sets of synapses belonging to the cellular assembly that constitutes the trace of the memory (behavioral tag) [11,16,17]. Subsequently, the elevated levels of catecholamines (or other neuromodulators like corticosterone) triggered by the strong event would activate the expression of PRPs needed to stabilize the induced alterations in the synapses and to consolidate the memory for the weak event. So far, the validity of the Behavioral tagging hypothesis has been confirmed by a plethora of studies combining weak protocols of various behavioral tasks (e.g. contextual fear conditioning, inhibitory avoidance, spatial object recognition, novel object recognition) with novel environment exposure (a session of open field (OF) exploration) [18,19,20,21]. Pairing exposure to OF with weak inhibitory avoidance protocol (wIA), Moncada and colleagues (2011) [20] demonstrated the importance of NA for the stabilization of the behavioral tag after memory encoding. By infusing beta adrenoreceptor antagonist (propranolol) in the dentate gyrus (DG), the researchers were able to disrupt the beneficial effect of OF on the consolidation of the memory for the wIA task. Moreover, the same study was able to establish that blocking the catecholaminergic transmission did not interfere with the formation of short-term IA memory, thus highlighting the role of NA in PRPs provision. A different set of studies established acute stress as another emotional state capable of modulating the consolidation of otherwise forgetful experiences [22,23]. The study by da Cunha et al. (2022) [23] provided evidence that mild stressful experiences like placing Wistar rats on an elevated platform (EP) one hour before learning is sufficient to induce a long-term memory for wIA. The observed effects were dependent on the activation of the glucocorticoid receptors (GRs), since the infusion of the GRs antagonist mifepristone in the CA1 field of the dorsal hippocampus abolished the storage of the memory for wIA. Another significant finding reported by the research team was that when EP exposure was combined with strong IA (sIA), the formation of a long-term memory for the behavioral task was impaired. This raises the possibility that intensity of the emotional states established by two events determines the dynamics of their interaction on the molecular level - two temporarily proximal salient events may compete for PRPs, leading to insufficient consolidation and fragmented memory.
Beside the hippocampus, emotionally charged or stressful events activate the amygdala - a heterogenous group of interconnected nuclei located in the temporal lobe and involved in the regulation of emotional behaviors [24]. The amygdala is necessary for the encoding emotional valence, emotional learning and regulation of the stress responses [24,25,26,27]. Substantial evidence has been provided that during emotional events one particular subregion of the structure - the basolateral complex of the amygdala (BLA) - incorporates the arousal signals originating from inside the brain (greater NA release from the LC) and from the peripheral hormone systems (heighten corticosterone level) to foster a positive modulatory effect on the processes required for the initial consolidation of memories [9,28,29,30,31,32]. Therefore, the BLA could also influence the prioritization of information in the hippocampus [33]. Since the BLA does not project directly to the dorsal hippocampus, the effect it exhibits on memory processes is reliant on its known anatomical connections with the entorhinal cortex, a cortical area that functions as a gateway and provides the hippocampus with sensory information originating from other specialized circuits [34,35]. Most studies that have established the modulatory role of the BLA on memory consolidation combined behavioral training of the experimental animals with pharmacological manipulations, mainly direct post-encoding infusions of different pharmacological agents [9]. However, evidence has also been provided by experiments using electrophysiological and optogenetic stimulation of the BLA, either during training or immediately after [36,37,38]. Importantly, post-training stimulation of the BLA affects the consolidation of various behavioral tasks regardless of the emotional valence of the training [38,39,40,41,42,43,44]. These findings allow for comparison with the protocols used by the behavioral tagging studies, since the stimulation of the BLA may mimic the effects on memory consolidation produced by a strong modulatory event. On the contrary, it has been reported that the memory modulation capacities of BLA stimulation depend on the levels of emotional arousal induced by the learning event [39,42]. For example, Roozendaal et al. (2006) [39] provided evidence that system injection of corticosterone immediately after the training trial of object recognition task (ORT) improved the performance of the animals during the test session 24h later. This effect was not observed in the animals that were substantially habituated to the training context before the beginning of the experiments, showing that habituation may reduce novelty-induced emotional arousal in the brain. Moreover, the same study reveals that infusion of propranolol in the BLA immediately after training blocks the corticosterone enhancement of memory consolidation, which demonstrates the role of BLA for the integration of arousal signals and the need for NA activation of the structure for the emotional modulation of memory formation. In a similar study, McReynolds et al. (2010) [40] have provided further evidence about the interaction between corticosterone and NA in the BLA. They observed that a systemic injection of corticosterone immediately after an IA training session increased the levels of NA in the BLA after 15 minutes, which was estimated through in vivo microdialysis. Furthermore, the administration of corticosterone elevated the synaptic levels of the activity-regulated cytoskeletal-associated protein (Arc) and enhanced memory consolidation for the IA task. However, when propranolol was infused in the BLA immediately after IA training and corticosterone administration, both the effect on memory and Arc expression were attenuated. Arc is an immediate early gene, the product of which is highly important for the stabilization of the synaptic modifications inducted by learning [45,46,47]. Several other publications have demonstrated that intra-BLA administration of the beta-adrenergic receptor agonist clenbuterol enhances memories for different tasks and increases the expression of Arc in the dorsal hippocampus [41,42,44]. In a series of experiments, McReynolds and colleagues (2014) [42] revealed that post-encoding infusion of clenbuterol in the BLA of rats had a positive effect on the consolidation of memories for both IA and ORT tasks. This was accompanied by an increase in the synaptic expression of Arc. Remarkably, the effects of clenbuterol on memory performance and Arc expression in the ORT experiment were noticeable only in the animals that were not subjected to habituation prior to the training. This once again illustrates that a certain degree of arousal must be induced during training in order for BLA to exhibit its modulatory function. Experiments examining BLA-induced augmentation of memory consolidation in the hippocampus like the ones described here have led to the proposal of the “Emotional tagging hypothesis” [29]. This hypothetical account is concerned with the role of the various aspects of an emotional event - intensity, controllability, duration - in facilitating of the interaction between the BLA on one side and memory-related plasticity in the hippocampus on the other. Despite the great number of studies examining the emotional modulation of memory consolidation, a thorough behavioral protocol to study interactions mentioned above, like those implemented by the behavioral tagging studies, is still lacking.
In light of the research reviewed above, our present experimental work intends to establish a novel behavioral protocol that could provide further evidence for the role of BLA in memory modulation by taking advantage of the natural changes in homeostatic conditions induced by an emotionally charged event. That is to say, we aimed to demonstrate that endogenous activation of the BLA by a stimulus with innate emotional valence promotes the stabilization of the behavioral tag set by a previous weak event. To do this, we decided to couple OLT with predatory odor exposure (POE) in a novel cage two minutes after the training trial. The presence of predatory cues has been proven to evoke neuronal responses in the BLA [48]. For the purpose of our experiments we used urine from the predatory species Canis latrans (coyote). Urine from predatory species is a highly aversive stimulus for rodents as it contains kairomones - natural chemical substances that serve as ecological signals informing the animal of the presence of potential predatory threat [49,50]. Previous studies have used coyote urine as unconditioned stimuli in hippocampus-dependent contextual fear conditioning paradigm [51,52]. Recent publication has also provided a detailed analysis of the repertoire of defensive behaviors of rats exposed to coyote urine [53]. Considering all these factors, we hypothesized that POE may be a potent modulatory event capable of inducing strong effect on the memory consolidation for a weaker behavioral task. Moreover, we predicted that memory enhancement prompt by POE may be dependent on NA transmission in BLA. To test these ideas, our group conducted two separate experiments. In Experiment #1, rats were assigned to three groups depending on the behavioral procedures and pharmacological treatment. Animals from the control group received a bilateral infusion of the vehicle in the BLA upon the compilation of the sample phase of the OLT. The other two groups were subjected to OLT training and exposed to coyote urine, followed by infusion of either vehicle or propranolol in the BLA. Rats from all the groups were sacrificed 90 minutes following pharmacological treatment and both of their hippocampi were extracted to examine the outcomes of BLA manipulation on experience-dependent plasticity. Experiment #2 followed the same protocol, but to establish possible retroactive enhancement of memory induced by POE, the animals from the three groups were submitted to a test 24 hours later. As in the first experiment, animals were sacrificed 90 minutes after the test and both hippocampi were collected to assess the retrieval-related changes. The selected tissue samples were used to assess the levels of the phosphorylated form of cAMP response element-binding protein (pCREB) and the expression of Arc through ELISA and western blotting (WB), respectively. We selected these molecular markers based on existing evidence for their experience-dependent activation and role in different memory processes [45,54,55,56].
2. Results
The experimental procedure is shown in Scheme 1. Details concerning the steps in Experiment #1 and Experiment #2 are shown in Section 4.
2.1. Experiment #1
2.1.1. Post-Training Propranolol Infusion Suppresses the Effect of the Exposure to a Natural Aversive Stimulus (Coyote Urine) on Hippocampal pCREB Expression
To investigate the role of endogenous NA released from the BLA as a result of POE on the expression of plasticity marker (pCREB) in the hippocampus, the rats were trained in an OLT, which involved the exploration of two identical stimulus objects located in different positions in the sample and the test sessions, respectively. Immediately after the sample session, each rat from the two stressed groups was exposed to a stressful stimulus (coyote urine) for 5 minutes. The matched control group (C + veh) was placed under the same conditions except for the delivery of the Petri dish. Immediately thereafter, the rats were infused with a vehicle into the BLA (C + veh) and (POE + veh) or propanonol (POE + prop). At the 90 min after the sample test, the hippocampus was isolated from all groups. One-way ANOVA revealed a significant difference between groups [F2,20 = 3.809, p = 0. 0418] for the expression of activated CREB (pCREB) in the hippocampus at the sample phase. The post-hoc test showed that pCREB expression in the hippocampus was significantly increased in the stress + veh group compared to the C + veh group (p = 0.0011) (Figure 1). At the same time, post-training infusion of propranolol after training suppressed the expression of this transcription factor in the hippocampus (p = 0.0013 compared to the POE + veh group).
2.1.2. The Arc Expression in the Hippocampus Showed a Tendency for Elevation as a Sequence of Post-Training Exposure to a Natural Aversive Stimulus (Coyote Urine)
To ascertain whether the Arc protein expression in the hippocampus is involved in the effect of endogenous NA released from the BLA on the consolidation process, the three groups (C + veh; POE + veh and POE + prop) were decapitated 90 min after the sample test. The Arc expression was explored with the Western blot (WB) test. A tendency for increased Arc expression in the hippocampus was detected in the stress + veh group compared to the C + veh group in homogenates tested by WB test (Figure 2).
2.2. Experiment # 2
2.2.1. Acute Post-Training Exposure to a Natural Aversive Stimulus (Coyote Urine) Did Not Affect the Performance of Rats on the Object Location Task
The procedure for infusing vehicle/propranolol into the BLA of rats in Experiment #2 was the same as in Experiment #1. Twenty-four hours after training, each rat from the three groups was re-exposed to two identical objects placed in a different location (Figure 3). One-way ANOVA analysis revealed no significant difference between groups [F2,25 = 1.402, p > 0.05]. Although there was a tendency for impaired retention in the stress + prop group, the results suggest a lack of memory-enhancing effect of acute stress-induced release of NA from the BLA.
2.2.2. Acute Post-Training Exposure to a Natural Aversive Stimulus (Coyote Urine) and Propranolol Infusion Did Not Change the pCREB Expression in the Hippocampus in the Retention Phase
No significant difference was found between the three groups after the test session for the pCREB expression in the hippocampus [F2,24 = 0.824, p > 0.05] (Figure 4), suggesting a lack of effect of endogenous NA released from the BLA after acute stress on this marker of plasticity.
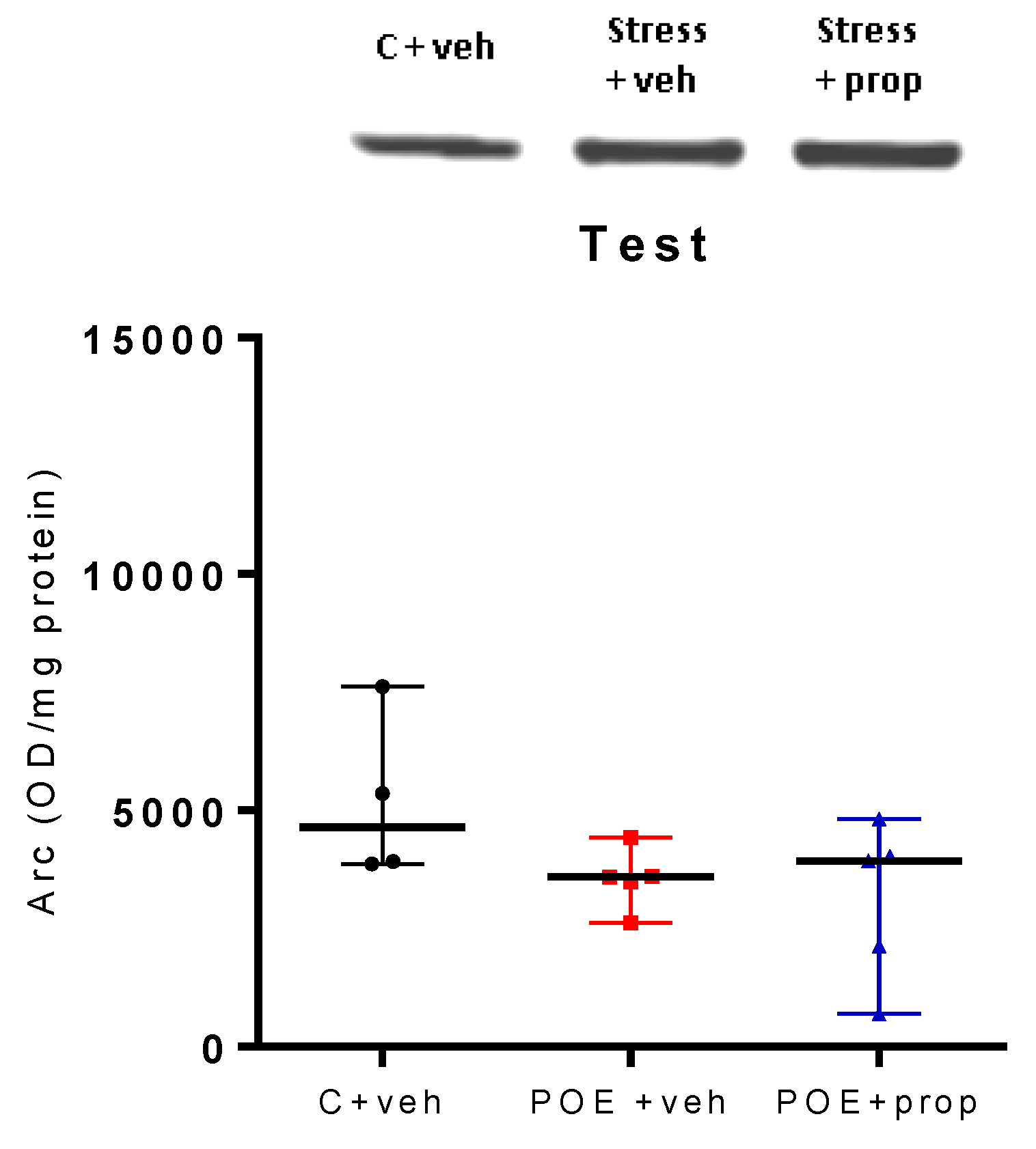
2.2.3. Acute Post-Training Exposure to Predator Odor and Propranolol Infusion Did Not Change the Arc Expression in the Hippocampus in the Retention Phase
There was no significant difference between the three groups (C + veh; POE + veh and stress + prop) in Arc expression in the hippocampus, measured by the WB test, in the retention phase (Figure 5).
3. Discussion
The experiments presented here were designed to demonstrate that NA activation of the BLA during emotionally arousing situations is critical for any retrograde enhancement effect on memory consolidation. As our protocol relied on exposing animals to a stimulus with an innate negative valence to induce emotional arousal rather than a pharmacological intervention, we aimed to provide direct evidence for the emotional aspects of the behavioral tagging. Our study is conceptually close to those by da Cunha and colleagues (2019) [22] and da Cunha and collaborators (2021) [23]. Specifically, the design of the 2019 study [22] paired weak OLT (4 minutes sample session) with EP session 1 hour after the initial training and discovered that the induced state of acute stress was able to save the memory for OLT from forgetting. The observed effects were dependent on elevated levels of corticosterone in the dorsal hippocampus, since infusion of antagonists for both mineralocorticoid (MRs) and glucocorticoid receptors (GRs) in the CA1 field disrupted the effects induced by the stressful experience. Furthermore, they showed that when EP was coupled with the strong version of OLT (10 minutes long training trial), the formation of a memory for the task was impaired, highlighting once again the observation that two temporarily proximal strong events may compete for PRPs. Although this was not the focus of their experiment, the results obtained by the researchers regarding the contribution of hippocampal MRs and GRs to memory enhancement could be interpreted from the perspective of a BLA-dependent process. As mentioned above, both the studies by Roozendaal et al. (2006) [39] and McReynolds at al. (2010) [40] have the demonstrated that the beneficial effect of a systemic corticosterone injection on memory could be prevented by infusion of propranolol in the BLA. These observations further motivated us to investigate how endogenous NA release in BLA during emotionally arousing situations influences the dynamics of memory formation in the hippocampus.
The main finding of our study is that the animals that were exposed to the predator odor and infused with vehicle immediately immediately after the end of the OLT sample session showed a significant upregulation in their pCREB levels and a trend toward increased Arc expression in the hippocampus compared to those infused with propranolol. Most importantly, this effect was mediated by the BLA. This is evident by the results obtained from our first experiment, where the animals that were infused with vehicle immediately after completion of the sample session of the OLT and the exposure to the predatory odor showed notably higher availability of both markers compared to those infused with propranolol. Furthermore, the substantial elevation in pCREB levels and the tendency for higher Arc expression in the rats from the POE/veh group compared to the control group, highlights the influence of the emotional event on hippocampal plasticity. A previous study by Kabitzke, Silva and Wiedenmayer (2011) [57] reported that using cat odor as an unconditioned stimulus in a contextual fear conditioning paradigm significantly increased pCREB levels in both dorsal and ventral hippocampus of juvenile rats. However, when an intraperitoneal injection of propranolol was administered after the exposure to the predatory cue, this experience-dependent effect was not observed and the animals showed no conditioned response when they were re-exposed to the training context 24 hours later, suggesting that the antagonist may have blocked the activation of CREB by NA and hindered the cellular mechanism of memory consolidation. Finally, post-training systemic injection of the non-selective beta-adrenergic blocker nadolol, which cannot cross the blood-brain barrier, had no effects on pCREB levels or the studied behavioral responses, suggesting that the effect on memory consolidation was facilitated by the endogenous release of NA in the brain. Our findings regarding pCREB extend the evidence described above, revealing that the effects of predatory cues on hippocampus-dependent memory and memory-induced behavioral responses are also influenced by the activation of the noradrenergic neurotransmission in the BLA. Importantly, in the experiment of Kabitzke, Silva and Wiedenmayer (2011) [57], the hippocampal tissue was collected 60 minutes after odor exposure. In our study, both hippocampi were extracted 90 minutes after the completion of all experimental procedures. This time point was selected because we wanted to establish the possible existence of a prolonged window for CREB activation. Since CREB is one of the main transcription factors involved in memory consolidation [55,56], a prolonged activation of CREB can be interpreted as an indication for increased transcriptional activity of the genes responsible for PRPs. With regard to Arc, our results showed a tendency for increased expression of the protein in the animals from the POE/veh group. This further emphasizes the role of BLA in modulating memory-related plasticity in the hippocampus during emotionally charged events. The studies that have provided evidence for BLA-modulated Arc expression have used synaptoneurosome preparation to determine synaptic levels of the protein in the dorsal hippocampus [40,42,44]. In addition, these studies have collected the samples at different timepoints and have shown that the peak concentration may be approximately 1 hour after training [40,44]. In our experiment we homogenized the entire hippocampus. This, together with the time point of tissue collection, may have limited our ability to obtain more detailed results. Nevertheless, we still were able to demonstrate the presence of an emotional arousal-dependent effect on Arc expression regulated by BLA.
Unfortunately, the second experiment was unable to provide firm evidence for a POE-induced modulatory influence on memory consolidation. This is evident by the lack of pronounced effect in the performance of the POE/veh group during the test session of the memory task. In line with the collected behavioral data, Arc expression in the test groups also showed no significant recognition-induced differences. Regarding the pCREB levels of the test groups, the results confirm the notion established by Moncada and Viola (2006) [58] that a familiar spatial environment can reduce the phosphorylation of the protein. As this is a pilot study, the behavioral protocol we used may have had some drawbacks that must be carefully evaluated and eliminated in the future. These drawbacks may partially explain the lack of behavioral effect observed in Experiment #2. Considering the behavioral procedures, animals in both experiments were habituated to the experimental context for 10 minutes, one day before the sample session of OLT and POE. This was done in order to limit the presence of possible anxiety-related and neophobic behaviors during the training. However, the studies by Roozendaal et al. (2006) [39] and McReynolds et al. [42] reported that habituation may neutralize the influence of post-encoding BLA activation on memory consolidation. In their paper, McReynolds et al. (2014) [42] suggest that there may be a requirement for parallel arousal-induced activation of both the BLA and the efferent memory-related structures for the modulatory effect to occur and promote greater synaptic expression of Arc. Thus, by familiarizing the animals with the experimental context, the habituation session of the current experiments may have limited the modulatory influences of the BLA by reducing novelty-induced activation of the dorsal hippocampus. Notably, in a study by Ballarini and colleagues (2009) [18] - that coupled weak spatial object recognition task with OF - animals were subjected to two habituation sessions, but the presence of an enhancing effect on memory consolidation was still observed. This suggests that the modulatory events characterized either by environmental novelty or the presence of emotionally arousing stimuli may promote memory consolidation through the activation of different neural circuits. Confirmation or rejection of this notion requires more behavioral tagging studies that include emotionally valenced stimuli in their modulatory events. Another factor that may have influenced the lack of noticeable modulatory effect on memory consolidation in our second experiment is the temporal window between the weak and the modulatory events. The rats underwent POE only two minutes after the end of the OLT sample session. This interval was chosen, because we wanted to design a more naturalistic behavioral paradigm. In natural environmental conditions emotional stimuli may appear surprisingly and influence the consolidation of the memories for the most proximal neutral stimuli or contextual features that could be used to optimize future memory-based predictions. Most behavioral tagging experiments that reported positive modulation on memory consolidation have exposed the animals to the strong event at different time points - ranging from 15 minutes to 1 hour before or after training [18,20,22]. Importantly, da Cunha and colleagues (2022) [23] found that acute stress (EP) experienced immediately after wIA did not impair the formation of memory for the task and facilitated its long-term stabilization. However, given the different nature of the emotionally arousing events selected in ours and theirs (POE vs. EP), currently we cannot rule out the possibility that the animals in our experiment may have formed a strong association between the two events that could have influenced their performance during the test session of OLT. When re-exposed to the experimental context the following day, animals may have expected to encounter the aversive stimuli once again, which may explain why we did not observe the memory-related behaviors we expected. In a recent review by Sazma, Shields and Yonelinas (2019) [59], which examined the effects of post-encoding stress in animal and human studies, the authors have expressed similar concerns about the possible negative effect of acute stress on subsequent memory retrieval. Considering the fact that the POE was only performed for 4 minutes, we doubt that the intensity of our modulatory event could have negatively impacted the consolidation of the memory for OLT. Detailed studies on how different physiological parameters of the emotional response to different predatory odors might influence the cellular and molecular basis of memory processes are still lacking. Nevertheless, a recent report by Migliaro and colleagues (2019) [60] provides some insights into the observed behavioral changes depending on the duration of the exposure. Using various tests to assess anxiety-like and depressive-like behavior 24 hours after exposure of Wistar rats to bobcat urine, they found that animals exposed for 10 or 20 minutes showed clear change in their behavioral profile, while those exposed to the predatory cue for 3 minutes showed no profound alterations. Taking all these different possibilities into account, we must emphasize that the results of our first experiment on pCREB and Arc expression directly argues against the likelihood of our modulatory event being too mild to produce experience-dependent neural activity in the hippocampus.
4. Materials and Methods
4.1. Animals
Young adult male Wistar rats purchased from the vivarium of the Institute of Neurobiology, BAS. Upon arrival, they were acclimated for one week prior to surgery. The rats were maintained under standard environmental conditions (22-23°C, 12/12 h light/dark cycle with light on at 7 am), in groups of 3-4 in Plexiglas cages, with access to laboratory chow and tap water ad libitum.
The experiments were performed in accordance with the Council Directive 2010/63/EC on animal experiments and approved by the Bulgarian Food Safety Agency (research project: N◦349).
4.2. Experimental Design
The experiments were performed in two separate protocols. Rats were randomly (blindly) allocated to experiment 1 (pCREB and Arc expression in the sampling phase) and experiment 2 (retention test, pCREB and Arc expression). Rats in one of three different conditions (n = 6-8) were as follows Group I (C + veh), Group II (stress + veh) and Group III (stress + prop). Rats were anaesthetized with ketamine (80 mg/kg, i.p.) and xylazine (20 mg/kg, s.c.) and implanted bilaterally by stereotaxic surgery with two cannulas according to the atlas of Paxinos and Watson [61] at the following coordinates (AP = - 2.5 L = ± 4.8; H = 6.5). One week after recovery, animals from the three groups were habituated for 5 min in the open field used for the memory task. Twenty-four hours later, the rats were trained in the OLT (a probe phase). Three minutes after training, each rat from Group II and Group III was exposed to a predator odour (coyote urine) for 5 minutes. Rats were injected with vehicle (groups I and II) or propranolol (0.75 µg) using a 30-gauge needle extended 2 mm beyond the cannula. The injection needle was connected to a 5 ml Hamilton syringe via a silicone tube. A volume of 0.5 µl vehicle (saline) or propranolol (0.5 µl/60 s) was infused. The injection needle was left in place for a further 60 seconds, after which the rat was returned to its home cage.
4.3. Object Localization Test
The object location test was conducted in an OF box (50 cm × 50 cm × 50 cm) with grey walls. The procedure consisted of three separate trials: familiarization, probe (training) and retention. A video camera placed above the box was used in each trial and the video recordings were analyzed offline. During habituation, each rat was placed in the OF without an object for 3 minutes. Twenty-four hours later, the sample trial was performed and each tested rat was exposed to two identical plastic objects (A1 and A2) for 3 min, positioned as shown in (Figure 3). The retention trial was performed 24 h later and the tested subject was placed in the same OF box where one of the objects remained in the same position as in the sample trial (A1) and the other object (A2) was placed in a novel position (Nov) for 3 min. The exploration time (sec), defined as the nose approaching the object in the test trial, was scored. The discrimination index (DI) for each object was calculated as follows D = Nov/A1 + Nov. The OF box and objects were thoroughly cleaned with 70% alcohol (v/v) after each trial. Animals were sacrificed by guillotine, and left and right hippocampal tissue was isolated, immediately frozen in liquid nitrogen and stored in a refrigerator at -20°C.
4.4. Enzyme-Linked Immunosorbent Assay (ELISA) Test
Hippocampal samples were weighed and and homogenized with HEPES buffer (20 mM HEPES; 1 mM EGTA; 210 mM mannitol; 70 mM sucrose; pH 7.2) and protease inhibitor cocktail (100mM PMSF, 100mM NaF, 35mM EDTA). The homogenates were centrifuged at 10 000 x g for 5 min at 4 °C. The supernatants were used for the determination of total protein by the Bradford method. Thereafter, two ELISA tests were performed to determine the protein expression of ARC and pCREB. ELISA assay was performed according to the manufacturer’s instructions for the specific kits (ELISA kit for Activity Regulated Cytoskeleton Associated Protein (ARC), SEJ620Ra; Rat phospho-cAMP response element binding protein (p-CREB), SL1344Ra).
4.5. Western Blot Test
Homogenized hippocampal tissue samples were mixed 1:1 with glycerol loading buffer and separated by electrophoresis on 12% SDS gels. Proteins were transferred to nitrocellulose membranes by semi-dry transfer. Non-specific binding was blocked by incubating overnight with 5% (w/v) BSA in TBS. The membranes were then incubated for two hours at 37°C with anti-ARC primary antibodies (Affinity Biosciences) at a dilution of 1:500 in TBS+5%BSA buffer. The membranes were washed with TTBS and incubated with HRP-conjugated goat anti-rabbit IgG (Elabscience, E-AB-1003) for one hour at room temperature. After washing, the bands were detected using chemiluminescent dye (Abcam ab134640). To determine the relative molecular weight of the proteins, a weight electrophoresis calibration kit (Elabscience) was run on the same slab gels. The membranes were dried and scanned.
Statistical Analysis
Experimental data were presented as mean ± S.E.M. The parametric statistical test was used in case of normality data distribution (one-way ANOVA followed by Tukey’s test) while Kruskal–Wallis on ranks followed by the Mann–Whitney U test was applied for non-parametric data. The significant level was set at p < 0.05.
5. Conclusions
Taking into account the result of Experiment #1, we are confident that with more detailed adjustments POE could become an even more suitable strategy for investigating the complex neurobiological systems behind emotional modulation of memory processes in laboratory rodents. In the future, it will be interesting to couple POE with other weak behavior protocols that have been extensively used to study hippocampal-dependent memory processes and behavioral tagging (contextual fear conditioning, inhibitory avoidance). In addition, the use of methods such as immunohistochemistry and in situ hybridization will allow us to better understand the interaction between POE and the learning task it modulates at the structural level. By activating the neural circuits involved in threat detection and defensive behaviors, predatory odors induce heightened emotional arousal. Studies have shown that the BLA is one of the main structures responsible for regulating the unconditioned response towards predatory odors. However, to our knowledge, no previous experiment has attempted to examine possible modulatory influences of the emotional state induced by predatory cues on the consolidation of memories for events. Thus, here we attempted to bridge two distinct and independently developing research lines. The Behavioral tagging hypothesis offers exciting opportunities to study the effects of different internal states and environmental conditions on memory formation. The result of such studies may provide valuable insights into the search for novel therapeutic strategies targeting the pathological alterations of episodic memory associated with Alzheimer’s disease and affective disorders. Emotional arousal is a strong modulator of hippocampal plasticity and investigating how its effects on memory are regulated by the noradrenergic neurotransmission in the BLA will provide a more in-depth understanding of the mechanism for information prioritization.
Author Contributions
Conceptualization, B.P. and J.T.; methodology, B.P,P.I.,D.K., L.K.,P.R.J,T.; formal analysis, B.P.,P.I.,J.T.; investigation, B.P.,P.I.,D.K.; resources, D.A.; data curation, B.P.; writing—original draft preparation, B.P.,J.T; writing—review and editing, B.P.,J.T.; project administration, D.A.; funding acquisition, D.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Bulgarian National Science Fund (Grant No. KП-06-Н61/12). The studies was supported by the Bulgarian Ministry of Education and Science within the framework of the National Recovery and Resilience Plan of Bulgaria, Component “Innovative Bulgaria”, project No. BG-RRP-2.004-0006-C02 “Development of research and innovation at Trakia University in service of health and sustainable well-being”. .
Institutional Review Board Statement
The study was conducted in accordance with the Decla-ration of Helsinki, and the experimental design was prepared in full accordance with the Europe-an Communities Council Directive 2010/63/ E.U. and approved by the Bulgarian Food Safety Agency (research project: #349).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sugar, J.; Moser, M. Episodic Memory: Neuronal Codes for What, Where, and When. Hippocampus 2019, 29, 1190–1205. [Google Scholar] [CrossRef] [PubMed]
- Allen, T. A.; Fortin, N. J. The Evolution of Episodic Memory. Proc. Natl. Acad. Sci. U.S.A. 2013, 110 (Suppl. S2), 10379–10386. [Google Scholar] [CrossRef] [PubMed]
- Fanselow, M. S.; Dong, H.-W. Are the Dorsal and Ventral Hippocampus Functionally Distinct Structures? Neuron 2010, 65, 7–19. [Google Scholar] [CrossRef]
- Eichenbaum, H. Hippocampus. Neuron 2004, 44, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Okuda, K.; Højgaard, K.; Privitera, L.; Bayraktar, G.; Takeuchi, T. Initial Memory Consolidation and the Synaptic Tagging and Capture Hypothesis. Eur J of Neuroscience 2021, 54, 6826–6849. [Google Scholar] [CrossRef] [PubMed]
- Lisman, J.; Grace, A. A.; Duzel, E. A neoHebbian Framework for Episodic Memory; Role of Dopamine-Dependent Late LTP. Trends in Neurosciences 2011, 34, 536–547. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P. V.; Connor, S. A. Noradrenergic Regulation of Hippocampus-Dependent Memory. CNSAMC 2019, 19, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Finsterwald, C.; Alberini, C. M. Stress and Glucocorticoid Receptor-Dependent Mechanisms in Long-Term Memory: From Adaptive Responses to Psychopathologies. Neurobiology of Learning and Memory 2014, 112, 17–29. [Google Scholar] [CrossRef]
- LaLumiere, R. T.; McGaugh, J. L.; McIntyre, C. K. Emotional Modulation of Learning and Memory: Pharmacological Implica-tions. Pharmacol Rev 2017, 69, 236–255. [Google Scholar] [CrossRef]
- Fernández, G.; Morris, R. G. M. Memory, Novelty and Prior Knowledge. Trends in Neurosciences 2018, 41, 654–659. [Google Scholar] [CrossRef]
- Dunsmoor, J. E.; Murty, V. P.; Clewett, D.; Phelps, E. A.; Davachi, L. Tag and Capture: How Salient Experiences Target and Rescue Nearby Events in Memory. Trends in Cognitive Sciences 2022, 26, 782–795. [Google Scholar] [CrossRef]
- Dahl, M. J.; Kulesza, A.; Werkle-Bergner, M.; Mather, M. Declining Locus Coeruleus–Dopaminergic and Noradrenergic Modulation of Long-Term Memory in Aging and Alzheimer’s Disease. Neuroscience & Biobehavioral Reviews 2023, 153, 105358. [Google Scholar] [CrossRef]
- Duszkiewicz, A. J.; McNamara, C. G.; Takeuchi, T.; Genzel, L. Novelty and Dopaminergic Modulation of Memory Persistence: A Tale of Two Systems. Trends in Neurosciences 2019, 42, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Hansen, N. The Longevity of Hippocampus-Dependent Memory Is Orchestrated by the Locus Coeruleus-Noradrenergic System. Neural Plasticity 2017, 2017, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Redondo, R. L.; Morris, R. G. M. Making Memories Last: The Synaptic Tagging and Capture Hypothesis. Nat Rev Neurosci 2011, 12, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Moncada, D.; Ballarini, F.; Viola, H. Behavioral Tagging: A Translation of the Synaptic Tagging and Capture Hypothesis. Neural Plasticity 2015, 2015, 1–21. [Google Scholar] [CrossRef]
- Vishnoi, S.; Raisuddin, S.; Parvez, S. Behavioral Tagging: A Novel Model for Studying Long-Term Memory. Neuroscience & Biobehavioral Reviews 2016, 68, 361–369. [Google Scholar] [CrossRef]
- Ballarini, F.; Moncada, D.; Martinez, M. C.; Alen, N.; Viola, H. Behavioral Tagging Is a General Mechanism of Long-Term Memory Formation. Proc. Natl. Acad. Sci. U.S.A. 2009, 106, 14599–14604. [Google Scholar] [CrossRef]
- Vishnoi, S.; Raisuddin, S.; Parvez, S. Behavioral Tagging: Role of Neurotransmitter Receptor Systems in Novel Object Recognition Long-Term Memory. ACS Omega 2022, 7, 11587–11595. [Google Scholar] [CrossRef]
- Moncada, D.; Ballarini, F.; Martinez, M. C.; Frey, J. U.; Viola, H. Identification of Transmitter Systems and Learning Tag Molecules Involved in Behavioral Tagging during Memory Formation. Proc. Natl. Acad. Sci. U.S.A. 2011, 108, 12931–12936. [Google Scholar] [CrossRef]
- Lima, K. R.; Da Rosa, A. C. D. S.; Picua, S. S.; E Silva, S. S.; Soares, N. M.; Mello-Carpes, P. B. Novelty Promotes Recognition Memory Persistence by D1 Dopamine Receptor and Protein Kinase A Signalling in Rat Hippocampus. Eur J of Neuroscience 2022, 55, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Lopes Da Cunha, P.; Villar, M. E.; Ballarini, F.; Tintorelli, R.; Ana María Viola, H. Spatial Object Recognition Memory Formation under Acute Stress. Hippocampus 2019, 29, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Lopes Da Cunha, P.; Tintorelli, R.; Correa, J.; Budriesi, P.; Viola, H. Behavioral Tagging as a Mechanism for Aversive-memory Formation under Acute Stress. Eur J of Neuroscience 2022, 55, 2651–2665. [Google Scholar] [CrossRef] [PubMed]
- Janak, P. H.; Tye, K. M. From Circuits to Behaviour in the Amygdala. Nature 2015, 517, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.-H.; Zhang, J.-Y.; Holmes, A.; Pan, B.-X. Amygdala Circuit Substrates for Stress Adaptation and Adversity. Biological Psychiatry 2021, 89, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Brockett, A. T.; Vázquez, D.; Roesch, M. R. Prediction Errors and Valence: From Single Units to Multidimensional Encoding in the Amygdala. Behavioural Brain Research 2021, 404, 113176. [Google Scholar] [CrossRef]
- Pignatelli, M.; Beyeler, A. Valence Coding in Amygdala Circuits. Current Opinion in Behavioral Sciences 2019, 26, 97–106. [Google Scholar] [CrossRef]
- Roesler, R.; Parent, M. B.; LaLumiere, R. T.; McIntyre, C. K. Amygdala-Hippocampal Interactions in Synaptic Plasticity and Memory Formation. Neurobiology of Learning and Memory 2021, 184, 107490. [Google Scholar] [CrossRef]
- Bergado, J. A.; Lucas, M.; Richter-Levin, G. Emotional Tagging—A Simple Hypothesis in a Complex Reality. Progress in Neurobiology 2011, 94, 64–76. [Google Scholar] [CrossRef]
- Hermans, E. J.; Battaglia, F. P.; Atsak, P.; De Voogd, L. D.; Fernández, G.; Roozendaal, B. How the Amygdala Affects Emotional Memory by Altering Brain Network Properties. Neurobiology of Learning and Memory 2014, 112, 2–16. [Google Scholar] [CrossRef]
- Roozendaal, B.; McEwen, B. S.; Chattarji, S. Stress, Memory and the Amygdala. Nat Rev Neurosci 2009, 10, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Paré, D.; Headley, D. B. The Amygdala Mediates the Facilitating Influence of Emotions on Memory through Multiple Interacting Mechanisms. Neurobiology of Stress 2023, 24, 100529. [Google Scholar] [CrossRef] [PubMed]
- Manns, J. R.; Bass, D. I. The Amygdala and Prioritization of Declarative Memories. Curr Dir Psychol Sci 2016, 25, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Roesler, R.; McGaugh, J. L. The Entorhinal Cortex as a Gateway for Amygdala Influences on Memory Consolidation. Neuroscience 2022, 497, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Paz, R.; Pare, D. Physiological Basis for Emotional Modulation of Memory Circuits by the Amygdala. Current Opinion in Neurobiology 2013, 23, 381–386. [Google Scholar] [CrossRef]
- Bass, D. I.; Nizam, Z. G.; Partain, K. N.; Wang, A.; Manns, J. R. Amygdala-Mediated Enhancement of Memory for Specific Events Depends on the Hippocampus. Neurobiology of Learning and Memory 2014, 107, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Bass, D. I.; Partain, K. N.; Manns, J. R. Event-Specific Enhancement of Memory via Brief Electrical Stimulation to the Basolateral Complex of the Amygdala in Rats. Behavioral Neuroscience 2012, 126, 204–208. [Google Scholar] [CrossRef]
- Huff, M. L.; Miller, R. L.; Deisseroth, K.; Moorman, D. E.; LaLumiere, R. T. Posttraining Optogenetic Manipulations of Basolateral Amygdala Activity Modulate Consolidation of Inhibitory Avoidance Memory in Rats. Proc. Natl. Acad. Sci. U.S.A. 2013, 110, 3597–3602. [Google Scholar] [CrossRef]
- Roozendaal, B.; Okuda, S.; Van Der Zee, E. A.; McGaugh, J. L. Glucocorticoid Enhancement of Memory Requires Arousal-Induced Noradrenergic Activation in the Basolateral Amygdala. Proc. Natl. Acad. Sci. U.S.A. 2006, 103, 6741–6746. [Google Scholar] [CrossRef]
- McReynolds, J. R.; Donowho, K.; Abdi, A.; McGaugh, J. L.; Roozendaal, B.; McIntyre, C. K. Memory-Enhancing Corticosterone Treatment Increases Amygdala Norepinephrine and Arc Protein Expression in Hippocampal Synaptic Fractions. Neurobiology of Learning and Memory 2010, 93, 312–321. [Google Scholar] [CrossRef]
- McIntyre, C. K.; Miyashita, T.; Setlow, B.; Marjon, K. D.; Steward, O.; Guzowski, J. F.; McGaugh, J. L. Memory-Influencing Intra-Basolateral Amygdala Drug Infusions Modulate Expression of Arc Protein in the Hippocampus. Proc. Natl. Acad. Sci. U.S.A. 2005, 102, 10718–10723. [Google Scholar] [CrossRef]
- McReynolds, J. R.; Anderson, K. M.; Donowho, K. M.; McIntyre, C. K. Noradrenergic Actions in the Basolateral Complex of the Amygdala Modulate Arc Expression in Hippocampal Synapses and Consolidation of Aversive and Non-Aversive Memory. Neurobiology of Learning and Memory 2014, 115, 49–57. [Google Scholar] [CrossRef]
- Barsegyan, A.; McGaugh, J. L.; Roozendaal, B. Noradrenergic Activation of the Basolateral Amygdala Modulates the Consolidation of Object-in-Context Recognition Memory. Front. Behav. Neurosci. 2014, 8. [Google Scholar] [CrossRef] [PubMed]
- McReynolds, J. R.; Carreira, M. B.; McIntyre, C. K. Post-Training Intra-Basolateral Complex of the Amygdala Infusions of Clenbuterol Enhance Memory for Conditioned Place Preference and Increase ARC Protein Expression in Dorsal Hippocampal Synaptic Fractions. Neurobiology of Learning and Memory 2021, 185, 107539. [Google Scholar] [CrossRef]
- Korb, E.; Finkbeiner, S. Arc in Synaptic Plasticity: From Gene to Behavior. Trends in Neurosciences 2011, 34, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Nikolaienko, O.; Patil, S.; Eriksen, M. S.; Bramham, C. R. Arc Protein: A Flexible Hub for Synaptic Plasticity and Cognition. Seminars in Cell & Developmental Biology 2018, 77, 33–42. [Google Scholar] [CrossRef]
- Bramham, C. R.; Alme, M. N.; Bittins, M.; Kuipers, S. D.; Nair, R. R.; Pai, B.; Panja, D.; Schubert, M.; Soule, J.; Tiron, A.; Wibrand, K. The Arc of Synaptic Memory. Exp Brain Res 2010, 200, 125–140. [Google Scholar] [CrossRef]
- Mitra, R. Neuronal Plasticity in the Amygdala Following Predator Stress Exposure. Front. Behav. Neurosci. 2019, 13, 25. [Google Scholar] [CrossRef]
- Ferrero, D. M.; Lemon, J. K.; Fluegge, D.; Pashkovski, S. L.; Korzan, W. J.; Datta, S. R.; Spehr, M.; Fendt, M.; Liberles, S. D. Detection and Avoidance of a Carnivore Odor by Prey. Proc. Natl. Acad. Sci. U.S.A. 2011, 108, 11235–11240. [Google Scholar] [CrossRef]
- Fortes-Marco, L.; Lanuza, E.; Martinez-Garcia, F. Of Pheromones and Kairomones: What Receptors Mediate Innate Emotional Responses? The Anatomical Record 2013, 296, 1346–1363. [Google Scholar] [CrossRef]
- Wang, M. E.; Fraize, N. P.; Yin, L.; Yuan, R. K.; Petsagourakis, D.; Wann, E. G.; Muzzio, I. A. Differential Roles of the Dorsal and Ventral Hippocampus in Predator Odor Contextual Fear Conditioning. Hippocampus 2013, 23, 451–466. [Google Scholar] [CrossRef] [PubMed]
- Yuan, R. K.; Hebert, J. C.; Thomas, A. S.; Wann, E. G.; Muzzio, I. A. HDAC I Inhibition in the Dorsal and Ventral Hippocampus Differentially Modulates Predator-Odor Fear Learning and Generalization. Front. Neurosci. 2015, 9. [Google Scholar] [CrossRef] [PubMed]
- Maestas-Olguin, C. R.; Parish, M. M.; Pentkowski, N. S. Coyote Urine, but Not 2-Phenylethylamine, Induces a Complete Profile of Unconditioned Anti-Predator Defensive Behaviors. Physiology & Behavior 2021, 229, 113210. [Google Scholar] [CrossRef]
- Morin, J.-P.; Guzmán-Ramos, K.; Bermudez-Rattoni, F. New Insights on Retrieval-Induced and Ongoing Memory Consolidation: Lessons from Arc. Neural Plasticity 2015, 2015, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Alberini, C. M.; Kandel, E. R. The Regulation of Transcription in Memory Consolidation. Cold Spring Harb Perspect Biol 2015, 7, a021741. [Google Scholar] [CrossRef] [PubMed]
- Alberini, C. M. Transcription Factors in Long-Term Memory and Synaptic Plasticity. Physiological Reviews 2009, 89, 121–145. [Google Scholar] [CrossRef] [PubMed]
- Kabitzke, P. A.; Silva, L.; Wiedenmayer, C. Norepinephrine Mediates Contextual Fear Learning and Hippocampal pCREB in Juvenile Rats Exposed to Predator Odor. Neurobiology of Learning and Memory 2011, 96, 166–172. [Google Scholar] [CrossRef]
- Moncada, D.; Viola, H. Phosphorylation State of CREB in the Rat Hippocampus: A Molecular Switch between Spatial Novelty and Spatial Familiarity? Neurobiology of Learning and Memory 2006, 86, 9–18. [Google Scholar] [CrossRef]
- Sazma, M. A.; Shields, G. S.; Yonelinas, A. P. The Effects of Post-Encoding Stress and Glucocorticoids on Episodic Memory in Humans and Rodents. Brain and Cognition 2019, 133, 12–23. [Google Scholar] [CrossRef]
- Migliaro, M. Effects on Behavior by Different Exposure Durations of Predator Scent Stress. aip 2020, 10, 17–26. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Academic Press: San Diego, CA, USA, 2007. [Google Scholar]
Scheme 1.
Experimental timeline.
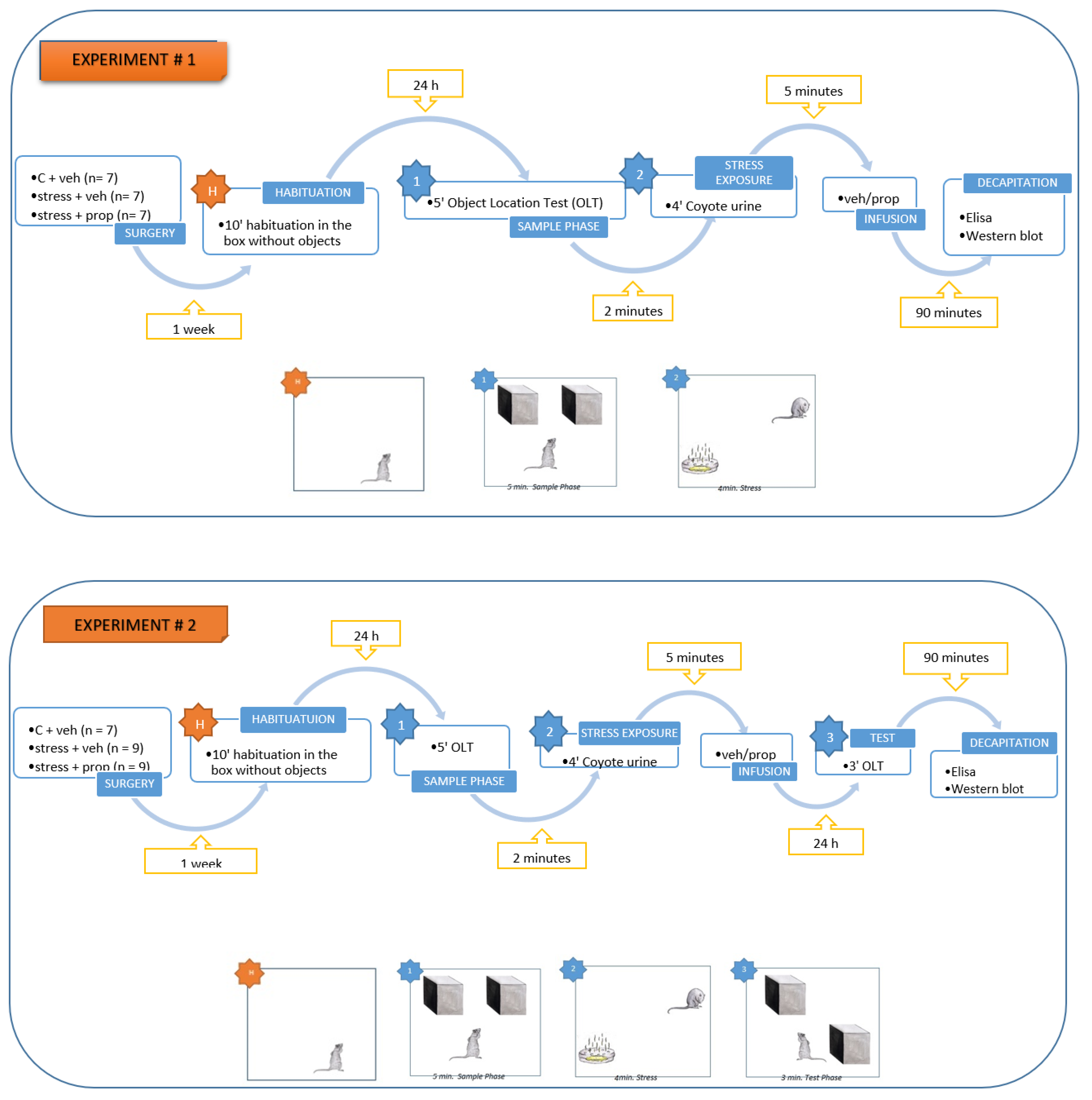
Figure 1.
Hippocampal pCREB levels (pg/mg protein), measured by ELISA test, was significantly increased after a predator odor exposure (POE) immediately after the sample phase of the object location task and infusion of vehicle in the basolateral amygdala (BLA). **p = 0.001, POE + veh group vs. C + veh group; **p = 0.0013, POE + prop group compared to the POE + veh group. n = 5-7.
Figure 1.
Hippocampal pCREB levels (pg/mg protein), measured by ELISA test, was significantly increased after a predator odor exposure (POE) immediately after the sample phase of the object location task and infusion of vehicle in the basolateral amygdala (BLA). **p = 0.001, POE + veh group vs. C + veh group; **p = 0.0013, POE + prop group compared to the POE + veh group. n = 5-7.
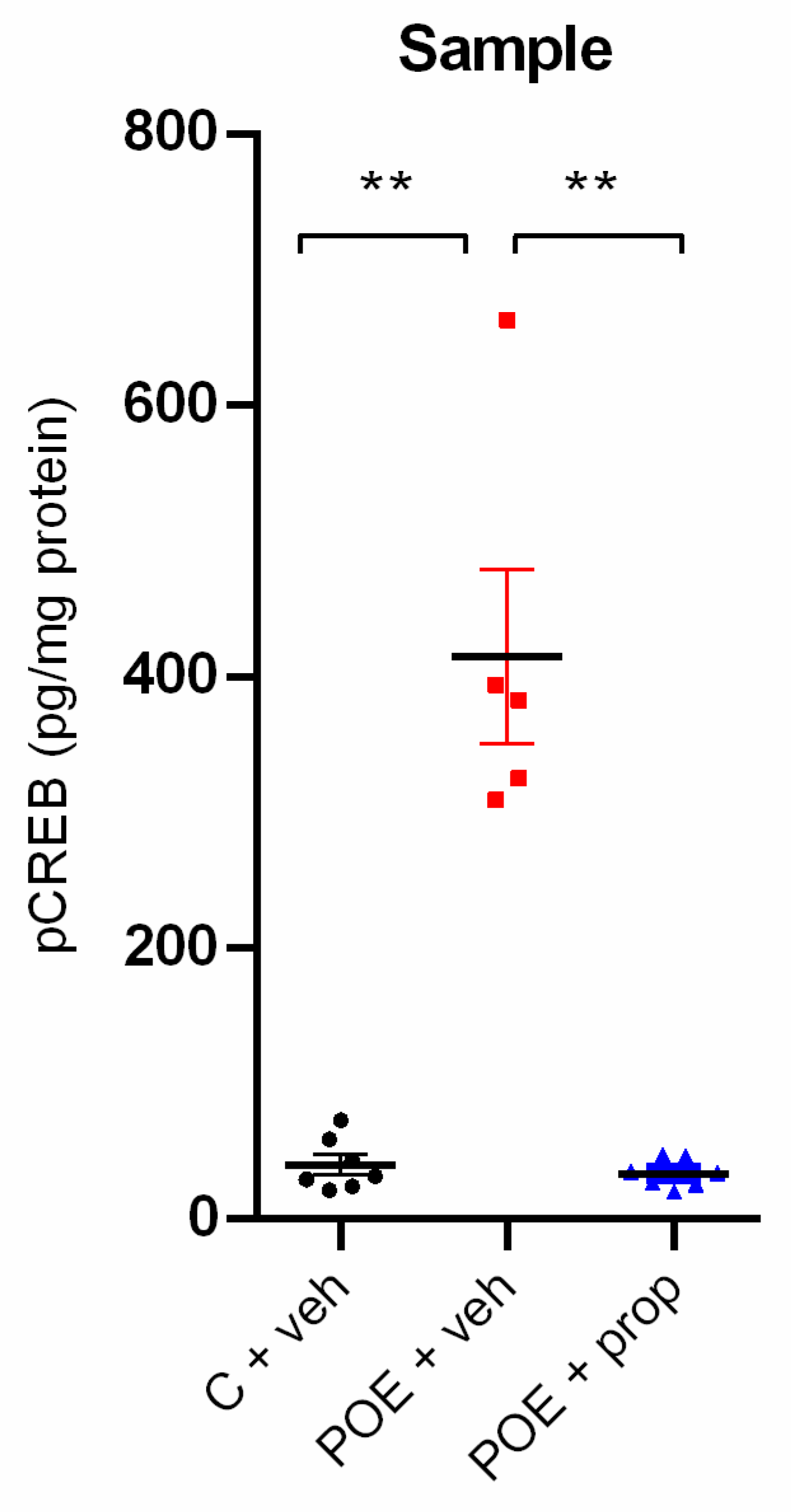
Figure 2.
Hippocampal Arc expression (OD/mg protein), measured by Western blot test, showed a tendency for increase after POE immediately after the sample phase of the object location task and vehicle infusion in the basolateral amygdala (BLA). n = 6.
Figure 2.
Hippocampal Arc expression (OD/mg protein), measured by Western blot test, showed a tendency for increase after POE immediately after the sample phase of the object location task and vehicle infusion in the basolateral amygdala (BLA). n = 6.
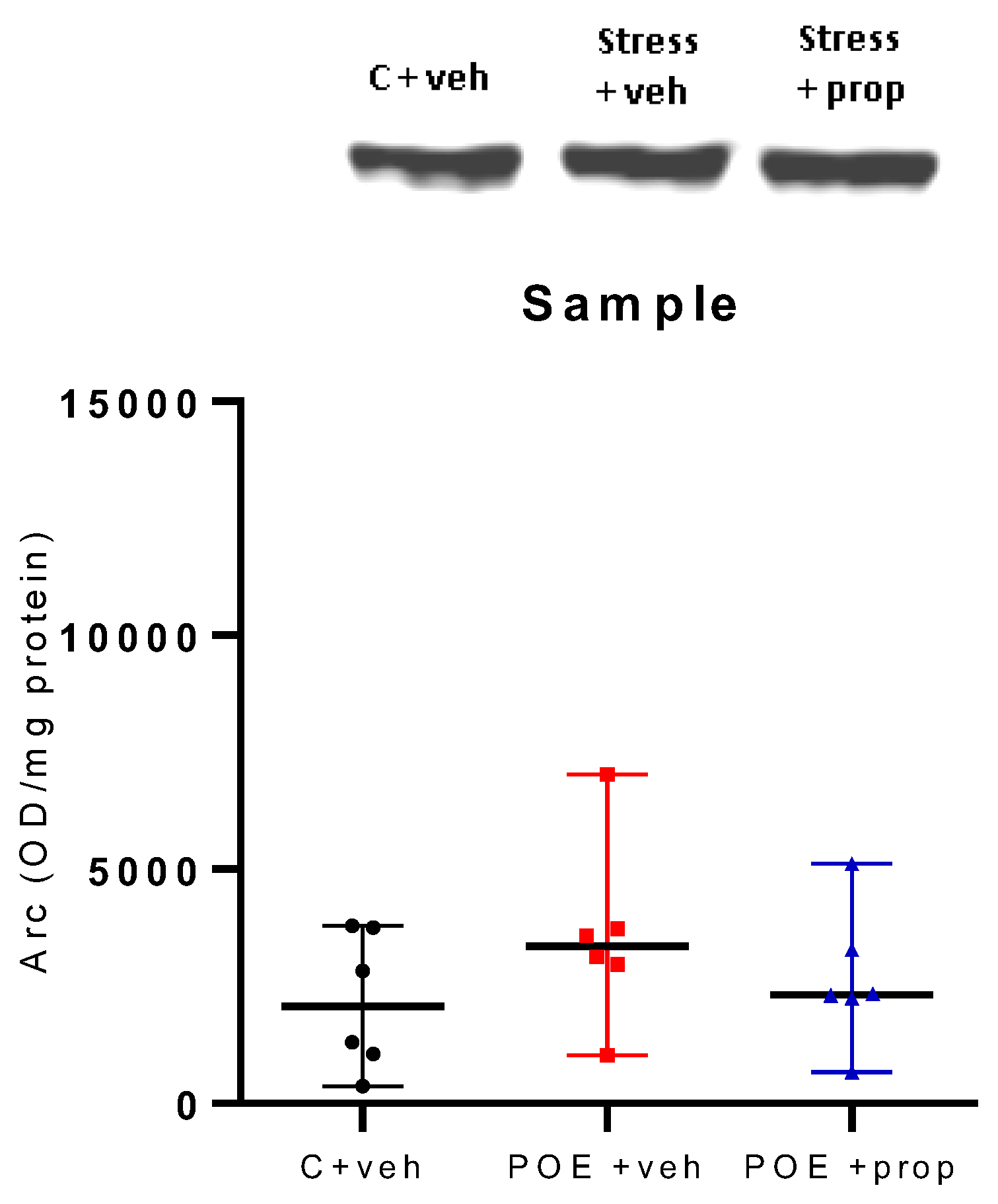
Figure 3.
No significant difference was found between the three groups tested, c + veh, POE + veh and POE + prop, in the object location task. n = 7-9.
Figure 3.
No significant difference was found between the three groups tested, c + veh, POE + veh and POE + prop, in the object location task. n = 7-9.

Figure 4.
Hippocampal pCREB levels (pg/mg protein), measured by ELISA test, were not significantly different between the three groups in the test phase. n = 6-9.
Figure 4.
Hippocampal pCREB levels (pg/mg protein), measured by ELISA test, were not significantly different between the three groups in the test phase. n = 6-9.
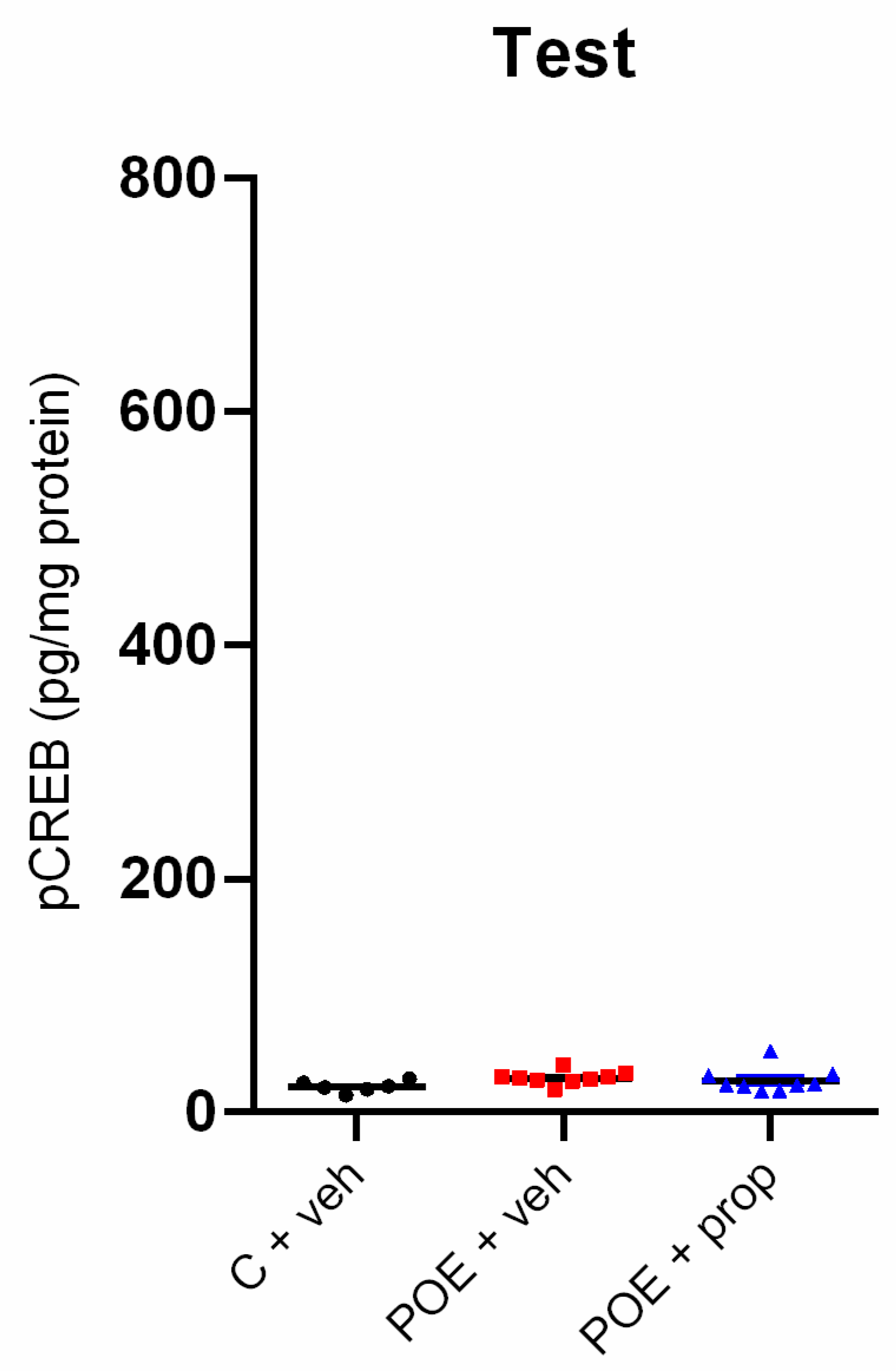
Figure 5.
Hippocampal Arc expression (OD/mg protein), measured by Western blot test, was not different between the three groups, C + veh, POE + veh and POE + prop. n = 6.
Figure 5.
Hippocampal Arc expression (OD/mg protein), measured by Western blot test, was not different between the three groups, C + veh, POE + veh and POE + prop. n = 6.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



