Submitted:
26 February 2024
Posted:
12 March 2024
You are already at the latest version
Abstract
Background: Bacteria can survive antibiotic exposure by mechanisms like tolerance without a shift in antimicrobial susceptibility. In this study, we investigated the presence of tolerance to three antimicrobials, ceftriaxone (CRO), azithromycin (AZM), and ciprofloxacin (CIP), in clinical isolates and WHO reference panel of N. gonorrhoeae and explored the prevalence between urogenital and anorectal infection sites. Methods: We examined 66 N. gonorrhoeae clinical isolates, evenly divided between anorectal and urogenital infection sites, using a modified tolerance disk (TD test) for three antibiotics. The isolates underwent a three-step incubation process wherein the isolates were exposed to an antibiotic disc for 20 hours of incubation (Step I), followed by the replacement of the antibiotic disc with a nutrient disc for overnight incubation (Step II) and additional overnight incubation with extra nutrients (Step III). Results: Four isolates were lost due to contamination. Of 62 clinical isolates, 7 exhibited tolerance to AZM, with 6 from anorectal and 1 from urogenital sites (p = 0.0496). Tolerance to CRO was observed in 11 isolates, which was significantly associated with the infection site, with 9 from the anorectum, and 2 from the urogenital site being tolerant (p = 0.02). Tolerance to CIP was found in 11 isolates, but was not significantly associated with the infection site. Tolerance was also detected in 8 (K, M, N, O, P, U, V, W) out of the 14 WHO (World Health Organization) reference strains, with varying patterns of tolerance to CRO (n=8), CIP (n=2) and AZM (n=1). Conclusion: This study identified CRO, AZM, and CIP tolerance in clinical Neisseria gonorrhoeae isolates. Significant variations in tolerance proportions between anorectal and urogenital infection sites, particularly for CRO, were observed. Additionally, tolerance was detected in WHO reference strains.
Keywords:
Neisseria gonorrhoeae
; tolerance
; antimicrobial resistance
; anorectal infection
; urogenital infection
; azithromycin
; ceftriaxone
; ciprofloxacin
Impact Statement
Tolerance, defined as the ability of a bacteria to survive transient antibiotic exposure without exhibiting a rise in the minimal inhibitory concentration, is a growing concern in high-priority pathogens, such as Neisseria gonorrhoeae, that causes gonorrhoea, a sexually transmitted infection. This study confirmed the findings from a previous study using a larger sample size and three antimicrobials (ciprofloxacin, azithromycin, and ceftriaxone) to detect tolerance in clinical and WHO reference isolates of Neisseria gonorrhoeae. Furthermore, tolerance to these antibiotics varied significantly between anorectal and urogenital infection sites, with ceftriaxone tolerance particularly prominent in anorectal isolates.
Introduction
Tolerance is defined as the ability of bacteria to survive transient exposure to high bactericidal concentrations of antibiotics by slowing their metabolism through an extension of the lag phase without a change in minimum inhibitory concentration (MIC) [1,2,3]. Antibiotic tolerance has been shown to play an important role in the emergence of antimicrobial resistance (AMR) [2,4]. Notably, in E. coli, tolerance has been shown to accelerate the development of AMR and has been implicated in treatment failure [5]. Recent studies have validated the use of the cheap and easy-to-perform Tolerance Disk (TD) test for detecting tolerance in clinical isolates of bacteria such as Staphylococcus aureus and Escherichia coli [6,7].
Gonorrhoea, the second-most-common bacterial sexually transmitted infection (STI), is caused by the bacteria Neisseria gonorrhoeae [8,9,10,11]. It has developed resistance to multiple classes of antibiotics, including ceftriaxone (CRO), the recommended treatment for gonorrhoea. Due to the increase in antimicrobial resistance (AMR), there is a real possibility Neisseria gonorrhoeae may become untreatable [12,13,14]. Previously, we demonstrated that tolerance to ceftriaxone (CRO) could be induced in Neisseria gonorrhoeae by intermittent exposure to high-concentration of CRO followed by the detection of tolerance in N. gonorrhoeae using the modified TD- test [15]. Furthermore, using the modified TD-test we detected CRO tolerance in clinical isolates of N. gonorrhoeae [15]. In addition, our study found that N. gonorrhoeae isolates from the anorectum were more likely to be CRO tolerant than isolates from urogenital sites. However, the limited number of samples assessed was too small to warrant assessing if the difference was statistically significant [15]. Moreover, the study only evaluated tolerance to a single antimicrobial [15]. Previous studies have found important pheno- and genotypic differences between N. gonorrhoeae isolated from different anatomical sites [16,17].
These findings led to the current study, where we aimed to assess if tolerance to three antimicrobials, azithromycin (AZM), ceftriaxone (CRO), and ciprofloxacin (CIP), could be detected in clinical isolates and WHO reference panel of N. gonorrhoeae using the previously established modified TD-test. In addition, we aimed to assess if the prevalence of tolerance to each antimicrobial differed between urogenital and anorectal infection sites.
Materials and methods
Bacterial strains
- (i)
- WHO reference panel
Fourteen gonococcal WHO reference strains were used in the study (F, G, K, L, M, N, O, P, U, V, W, X, Y, Z ) [18].
- (ii)
- Clinical isolates
N. gonorrhoeae clinical isolates (n = 66) from the Belgian National Reference Centre of STIs (2023) were selected for the study. The isolates were equally divided between anorectal (n=33) and urogenital (n=33) infection sites. The isolates were selected at random by an independent researcher instructed to randomly select 33 anorectal and 33 urogenital isolates received by the Belgian National Reference Centre of STIs in 2023.
MICs of three antimicrobials (AZM, CRO, and CIP) were determined using the E-test method, following the manufacturer's instructions (BioMérieux, France) on Gonococcal (GC) agar plates [3.6% BD DifcoTM GC Medium Base (Fisher Scientific) supplemented with 1% IsoVitalex (BD)] (Supplementary Table S1). The isolates were revived from storage at -80°C in 20% glycerol skim milk. They were subcultured and incubated twice on a BDTM Columbia Agar with 5% Sheep Blood at 5.5% CO2 atmosphere and 36°C.
Tolerance Detection
The modified TD-test was carried out according to Balduck et al., 2022. [15]The test comprises a three-step incubation process. In brief, the direct colony suspension method was used to prepare inoculum [19], wherein a small quantity of inoculum from an overnight subculture on chocolate agar was suspended in phosphate buffer saline (PBS). The turbidity of the suspension was adjusted to 0.5-1.0 to McFarland standard and inoculated on BDTM GC agar plates. Three antibiotics disks were placed on each inoculated plate: AZM 0.75 µg, CRO 0.064 µg CIP 0.032 µg. Disks were created by soaking 6 mm blank disks (Merck Life Science, Darmstadt, Germany) in 25 µL of the required antibiotic; the antibiotic concentration was calculated so that it would fall below the MIC. After approximately 20 to 24 hours of incubation at 36 °C and 6.0% CO2 (Step I TD test), the antibiotic disks were removed and replaced by 6 mm blank disks containing 25 µl gonococcal broth (distilled water supplemented with 15 g/L bacto protease peptone no 3, 1 g/L soluble starch, 4 g/L K2HPO4 (174.18 g/mol), 1 g/L KH2PO4 (136.08 g/mol), 5 g/L NaCl (58.44 g/mol) and 1% BD BBLTM IsoVitaleX). The plates were again incubated overnight (Step II TD test). This was followed by adding 10 µl of GC medium onto the same nutrient disk. The plates were incubated for another night (Step III TD test) (Figure 1).
All plates were analyzed for tolerance by three independent researchers and were photographed. The TD-test was duplicated and triplicated for the WHO reference and clinical N. gonorrhoeae isolates, respectively. If tolerant colonies were found, they were harvested and grown on BDTM GC Chocolate Agar for an incubation period of 48 hours and stored in 20% glycerol skim milk at −80 °C.
Antimicrobial Susceptibility Testing
Following the TD tests, the MIC of tolerant colonies was determined using the E-test method (range AZM: 0.016 µg/mL – 256 µg/mL CRO: 0.016 µg/mL – 256 µg/mL, CIP: 0.002 – 32 µg/mL) (BioMérieux, France), according to the manufacturer's instructions on GC agar plates [20].
Statistical Analysis
Fisher exact test implemented in XLSTAT (https://www.xlstat.com/enersion 28.0; IBM Corporation, Armonk, NY, USA) was used to determine the association between the prevalence of tolerance/non-tolerance isolates and infection sites (anorectal, urogenital) for each antibiotic (AZM, CRO, CIP). Three replicates per clinical isolate were available. If any replicates displayed tolerant colonies, the isolate was categorized as 'tolerant'. If only one or two out of the three replicates exhibited tolerant colonies, they were classified as 'heterotolerant'. Two sensitivity analyses were performed, where tolerance was defined as the detection of tolerance in 2 or 3 of the triplicate TD-tests. A p-value lower than 0.05 was considered statistically significant.
Results
The following isolates were lost during the TD test due to contamination: one replicate from the WHO reference panel (WHO V) and four clinical isolates. Among the clinical isolates, one urogenital and three anorectal isolates were lost, resulting in 30 anorectal and 32 urogenital clinical isolates. The pictures of the TD-tests are available on request.
Detection of Tolerance and Heterotolerance in WHO Reference Strains
Out of the 14 isolates, tolerance to AZM, CIP and CRO was detected in one (WHO Z), two (WHO P and WHO U) and eight WHO isolates (K, M, N, O, P, U, V, and W), respectively (Supplementary Table S1, Table 1).
The TD tests were performed in duplicate, which resulted the identification of heterotolerance in one of the 14 isolates. Specifically, tolerance was identified in one of two replicates for CIP and CRO in WHO-U (Table 1).
Differences in Azithromycin and Ceftriaxone Tolerance Across Infection Sites
Eleven (17%) of the 62 clinical N. gonorrhoeae isolates exhibited tolerance to AZM. Among the 30 anorectal clinical isolates, 10 (33%) showed tolerance to AZM, whereas among the 32 urogenital clinical isolates, one (3%) exhibited AZM tolerance (p = 0.002, Table 2).
The prevalence of tolerance to CRO in clinical isolates of N. gonorrhoeae was 13 (20%) out of 62. Tolerance to CRO was higher in N. gonorrhoeae clinical isolates from anorectal infection sites than urogenital infection sites. Ten (33%) out of 30 anorectal clinical isolates and three (9%) out of 32 urogenital clinical isolates showed tolerance to CRO (p = 0.029, Table 2).
No Difference in Tolerance to Ciprofloxacin between the Infection Sites
No statistically significant difference was found for the rate of tolerance to CIP based on the infection site of clinical N. gonorrhoeae isolates. Tolerance to CIP was detected in 14 (22.5%) of the 62 clinical N. gonorrhoeae isolates. Five (17%) and nine (28%) of the 30 anorectal and 32 urogenital clinical isolates showed tolerance to CIP, respectively (p = 0.367, Table 2, Supplementary Table S2.
Heterotolerance and Variability in Azithromycin, Ciprofloxacin and Ceftriaxone Tolerance among Clinical N. gonorrhoeae Isolates
Heterolerance to AZM, CIP and CRO were detected in 7 (11.2%), 10 (16.1%) and 5 (8%) clinical isolates out of the 62 N. gonorrhoeae isolates, respectively (Table 2). Out of the 30 anorectal isolates, six (20%), three (10%), and four (13%) isolates showed heterotolerance to AZM, CIP and CRO, respectively. Of the 32 urogenital isolates, one (3%), seven (22%) and one (3%) showed heterotolerance to AZM, CIP and CRO, respectively (Table 2).
No Decreased Susceptibility to Azithromycin, Ceftriaxone, and Ciprofloxacin in Tolerant Colonies
The detected tolerant colonies did not have an increase in MIC compared to the baseline isolates (data was shown). The sensitivity analysis, where we defined isolates as tolerant only if two (sensitivity analysis 1) or three (sensitivity analysis 2) of the three replicates were tolerant did not substantively change the results (Supplementary Table S3).
Discussion
We previously established that tolerance to CRO could be detected in N. gonorrhoeae clinical isolates, but this was limited to a small sample size [15]. This study confirmed the previous finding using a larger sample size, 14 of the WHO reference strains and using three antibiotics to detect tolerance. In the clinical N. gonorrhoeae isolates, we found that the prevalence of tolerance to AZM and CRO (but not CIP) was higher in anorectal clinical isolates than in urogenital clinical isolates.
Previous studies have found tolerance in clinical isolates of different bacterial species, such as methicillin-resistant Staphylococcus aureus (MRSA) blood infections, Pseudomonas aeruginosa infections in cystic fibrosis patients, and Enterococcus faecium infection in a leukaemia patient [21,22,23]. Lazarovits et al. (2022) described how tolerance to multiple antibiotics, including ampicillin, CRO, and ertapenem, was detected via the TD test in E. coli isolates of patients with bloodstream infections. Importantly, they found that the detection of tolerance in E. coli was associated with an increased risk of reinfection [24].
There are a number of possible explanations for the higher prevalence of AZM and CRO tolerance in the anorectal than the urethral infections. Urethral infections are typically symptomatic and of short duration, whereas the vast majority of anorectal infections are asymptomatic and persist for months [25]. These differences are, in turn, related to factors such as differences in the microbiome and immune response in these locales (Figure 2). The rectal microbiome is considerably more diverse and abundant than the urethral microbiome [26,27]. A large number of bacterial species have been found to interact with N. gonorrhoeae. A number of bacterial species, such as numerous Enterobacteriales spp. that are prevalent in the anorectum inhibit the growth of N. gonorrhoeae through the production of substances such as bacteriocins [28,29,30]. Various streptococcal species have been noted to exhibit a similar effect [31]. It is possible that differences in these inhibitory effects between the urethra and anorectum may explain the higher prevalence of tolerance in the anorectum. Differences in the host immune responses between the urethra and anorectum may also play a role (Figure 2a,b). The abundance of bacteria in the rectum is partially enabled by a downregulation of the host immune system at this site. For example, the toll-like receptors on the apical surface of the rectal epithelium are strongly down-regulated [32]. The downregulated immune system in the rectum may favour the emergence of bacterial tolerance. A further possibility is that the longer duration of colonization in the anorectum than the urethra means that anorectal infections are more likely to be exposed to bystander selection from antibiotics used for other indications that, in turn, select for tolerance [33].
Our findings are commensurate with those of studies that have found that the anatomical site of infection selects for specific pheno- and genotypic traits in N. gonorrhoeae. One study, for example, found that the cervix selected for loss of function mutations in the mtrCDE and farAB efflux pumps, which were, in turn, associated with increased susceptibility to various antimicrobials[16]. Another study has found that anorectal gonococcal infections exhibited a higher expression of mtrCDE than urethral infections [17].
Clinical cure rates for urethral and anorectal infections are typically high for most recommended treatments. This is less in the case of pharyngeal infections, where the cure rate for agents such as aminoglycosides and zoliflodacin is lower than for other sites [34]. Whilst poor drug penetration into the oropharynx likely plays an important role in this regard, it may be worthwhile testing the hypothesis that tolerance contributes to this poor cure rate. Whilst we did not find that tolerance increased the probability of AMR emerging, studies in other species have found that tolerance plays a crucial role in the emergence of AMR [5,35]. Future studies will be required to test this hypothesis more extensively in N. gonorrhoeae.
It is possible that tolerance, just like resistance, could be underpinned by stochastic pheno- and genotypic variations. These variations could explain our finding of heterotolerance. Only one of four published studies using TD-tests reported conducting the test in replicate [6,7,24,36]. This one study reported conducting the TD-tests in duplicate but did not report if there was any discordance in the TD-test between replicates [36]. Our study was thus the first to report heterotolerance. It is worth noting that using the replicates in the TD-test we identified heterotolerance to both CIP and CRO in one WHO reference isolate (WHO-U).
The limitations of this study include the use of only TD tests to detect tolerance; other techniques, such as MDK99 killing curves, could have provided useful complementary information. In addition, no genotyping or transcriptomics was performed on the obtained tolerant colonies, as this was beyond the scope of the current study. However, we have recently performed omics on tolerant colonies that will create a better understanding of the mutations associated with CRO tolerance in N. gonorrhoeae [37]. Although we tested all the clinical isolates in triplicate, we did not rerun the TD-tests on a separate occasion to assess the reproducibility of our findings. Finally, we do not have an explanation for why there was no difference in the prevalence of ciprofloxacin tolerance between anatomical sites.
Nonetheless, this is the first in vitro study to detect tolerance to AZM, CIP and CRO in clinical isolates of N. gonorrhoeae. The study established a difference in the rate of tolerance to AZM and CRO based on the infection site. Moreover, we used a large sample size (the biggest to date), performed the experiment in triplicates, and performed the investigation blinded to infection sites. Furthermore, our discovery of heterotolerance, where not all replicates of the isolates showed tolerance, presents a novel dimension to explore.
Funding
The study was funded by SOFI 2021 grant—‘PReventing the Emergence of untreatable STIs via radical Prevention’ (PRESTIP)
Conflicts of Interest
All the authors declare that they have no conflicts of interest.
References
- Brauner A, Fridman O, Gefen O, Balaban NQ. Distinguishing between resistance, tolerance and persistence to antibiotic treatment. Nature reviews. Microbiology 2016, 14, 320–330.
- Sulaiman, J.E.; Lam, H. Evolution of Bacterial Tolerance Under Antibiotic Treatment and Its Implications on the Development of Resistance. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Fridman, O.; Goldberg, A.; Ronin, I.; Shoresh, N.; Balaban, N.Q. Optimization of lag time underlies antibiotic tolerance in evolved bacterial populations. Nature 2014, 513, 418–421. [Google Scholar] [CrossRef] [PubMed]
- Handwerger, S.; Tomasz, A. Antibiotic Tolerance Among Clinical Isolates of Bacteria. Clin. Infect. Dis. 1985, 7, 368–386. [Google Scholar] [CrossRef]
- Levin-Reisman, I.; Ronin, I.; Gefen, O.; Braniss, I.; Shoresh, N.; Balaban, N.Q. Antibiotic tolerance facilitates the evolution of resistance. https://www.science.org (2017). [CrossRef]
- Gefen, O.; Chekol, B.; Strahilevitz, J.; Balaban, N.Q. TDtest: easy detection of bacterial tolerance and persistence in clinical isolates by a modified disk-diffusion assay. Sci. Rep. 2017, 7, srep41284. [Google Scholar] [CrossRef]
- Kotková, H.; Cabrnochová, M.; Lichá, I.; Tkadlec, J.; Fila, L.; Bartošová, J.; Melter, O. Evaluation of TD test for analysis of persistence or tolerance in clinical isolates of Staphylococcus aureus. J. Microbiol. Methods 2019, 167, 105705. [Google Scholar] [CrossRef]
- Hill, S.A.; Masters, T.L.; Wachter, J. Gonorrhea – an evolving disease of the new millennium. Microb. Cell 2016, 3, 371–389. [Google Scholar] [CrossRef] [PubMed]
- Piszczek J, St. Jean R, Khaliq Y. Gonorrhea: Treatment update for an increasingly resistant organism. Can Pharm J / Rev des Pharm du Canada 2015, 148, 82–89. [Google Scholar]
- Unemo M, Seifert HS, Hook EW, Hawkes S, Ndowa F, et al. Gonorrhoea. Nat Rev Dis Prim 2019, 5, 79.
- world Health Organization, Prudden HJ. WHO preferred product characteristics for gonococcal vaccines. 2020.
- World Health Organisation. WHO guidelines for the treatment of Neisseria gonorrhoeae. https://apps.who.int/iris/bitstream/handle/10665/246114/9789241549691-eng.pdf?sequence=1 (2016, accessed 23 May 2023).
- Ohnishi M, Saika T, Hoshina S, Iwasaku K, Nakayama S, et al. Ceftriaxone-resistant Neisseria gonorrhoeae, Japan. Emerging infectious diseases 2011, 17, 148–149.
- Unemo, M.; Golparian, D.; Nicholas, R.; Ohnishi, M.; Gallay, A.; Sednaoui, P. High-Level Cefixime- and Ceftriaxone-Resistant Neisseria gonorrhoeae in France: Novel penA Mosaic Allele in a Successful International Clone Causes Treatment Failure. Antimicrob. Agents Chemother. 2012, 56, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Balduck, M.; Laumen, J.G.E.; Abdellati, S.; De Baetselier, I.; de Block, T.; Manoharan-Basil, S.S.; Kenyon, C. Tolerance to Ceftriaxone in Neisseria gonorrhoeae: Rapid Induction in WHO P Reference Strain and Detection in Clinical Isolates. Antibiotics 2022, 11, 1480. [Google Scholar] [CrossRef]
- Ma, K.C.; Mortimer, T.D.; Hicks, A.L.; Wheeler, N.E.; Sánchez-Busó, L.; Golparian, D.; Taiaroa, G.; Rubin, D.H.F.; Wang, Y.; Williamson, D.A.; et al. Adaptation to the cervical environment is associated with increased antibiotic susceptibility in Neisseria gonorrhoeae. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- A Morse, S.; Lysko, P.G.; McFarland, L.; Knapp, J.S.; Sandstrom, E.; Critchlow, C.; Holmes, K.K. Gonococcal strains from homosexual men have outer membranes with reduced permeability to hydrophobic molecules. Infect. Immun. 1982, 37, 432–438. [Google Scholar] [CrossRef]
- Unemo M, Golparian D, Sánchez-Busó L, Grad Y, Jacobsson S, et al. The novel 2016 WHO Neisseria gonorrhoeae reference strains for global quality assurance of laboratory investigations: phenotypic, genetic and reference genome characterization. J Antimicrob Chemother 2016, 71, 3096–3108.
- Mcdermott PF, White DG, Zhao S, Simjee S, Walker RD. Antimicrobial Susceptibility Testing. Preharvest Postharvest Food Saf Contemp Issues Futur Dir 2008, 25.
- BioMérieux. ETEST - Trusted Leader in MIC Gradient Strip Technology. https://www.biomerieux-usa.com/sites/subsidiary_us/files/prn_056750_rev_03.a_etest_brochure_final_art_2.pdf.
- Liu, J.; Gefen, O.; Ronin, I.; Bar-Meir, M.; Balaban, N.Q. Effect of tolerance on the evolution of antibiotic resistance under drug combinations. Science 2020, 367, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Mulcahy, L.R.; Burns, J.L.; Lory, S.; Lewis, K. Emergence of Pseudomonas aeruginosa Strains Producing High Levels of Persister Cells in Patients with Cystic Fibrosis. J. Bacteriol. 2010, 192, 6191–6199. [Google Scholar] [CrossRef] [PubMed]
- Honsa, E.S.; Cooper, V.S.; Mhaissen, M.N.; Frank, M.; Shaker, J.; Iverson, A.; Rubnitz, J.; Hayden, R.T.; Lee, R.E.; Rock, C.O.; et al. RelA Mutant Enterococcus faecium with Multiantibiotic Tolerance Arising in an Immunocompromised Host. mBio 2017, 8, e02124–16. [Google Scholar] [CrossRef]
- Lazarovits, G.; Gefen, O.; Cahanian, N.; Adler, K.; Fluss, R.; Levin-Reisman, I.; Ronin, I.; Motro, Y.; Moran-Gilad, J.; Balaban, N.Q.; et al. Prevalence of Antibiotic Tolerance and Risk for Reinfection Among Escherichia coli Bloodstream Isolates: A Prospective Cohort Study. Clin. Infect. Dis. 2022, 75, 1706–1713. [Google Scholar] [CrossRef]
- Chow, E.P.F.; Camilleri, S.; Ward, C.; Huffam, S.; Chen, M.Y.; Bradshaw, C.S.; Fairley, C.K. Duration of gonorrhoea and chlamydia infection at the pharynx and rectum among men who have sex with men: a systematic review. Sex. Heal. 2016, 13, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Dekaboruah, E.; Suryavanshi, M.V.; Chettri, D.; Verma, A.K. Human microbiome: an academic update on human body site specific surveillance and its possible role. Arch. Microbiol. 2020, 202, 2147–2167. [Google Scholar] [CrossRef]
- Galiwango, R.M.; Park, D.E.; Huibner, S.; Onos, A.; Aziz, M.; Roach, K.; Anok, A.; Nnamutete, J.; Isabirye, Y.; Wasswa, J.B.; et al. Immune milieu and microbiome of the distal urethra in Ugandan men: impact of penile circumcision and implications for HIV susceptibility. Microbiome 2022, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Akomoneh, E.A.; Laumen, J.G.E.; Abdellati, S.; Van Dijck, C.; Vanbaelen, T.; Britto, X.B.; Manoharan-Basil, S.S.; Kenyon, C. The Discovery of Oropharyngeal Microbiota with Inhibitory Activity against Pathogenic Neisseria gonorrhoeae and Neisseria meningitidis: An In Vitro Study of Clinical Isolates. Microorganisms 2022, 10, 2497. [Google Scholar] [CrossRef] [PubMed]
- Baquero F, Moreno F. The microcins. FEMS Microbiol Lett 1984, 23, 117–124.
- Simpson, D.M.; Davis, C.P. Properties of a Gonococcal Inhibitor Produced by Escherichia coli. J. Gen. Microbiol. 1979, 115, 471–477. [Google Scholar] [CrossRef] [PubMed]
- McBride, M.E.; Duncan, W.C.; Knox, J.M. Bacterial interference of Neisseria gonorrhoeae by alpha-haemolytic streptococci. Sex. Transm. Infect. 1980, 56, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.C.-H.; Wang, J.-T.; Wei, S.-C.; Ni, Y.-H. Host-microbial interactions and regulation of intestinal epithelial barrier function: From physiology to pathology. World J. Gastrointest. Pathophysiol. 2012, 3, 27–43. [Google Scholar] [CrossRef]
- Tedijanto, C.; Olesen, S.W.; Grad, Y.H.; Lipsitch, M. Estimating the proportion of bystander selection for antibiotic resistance among potentially pathogenic bacterial flora. Proc. Natl. Acad. Sci. 2018, 115, 201810840–E11995. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.Y.S.; Hatzis, C.L.; Lau, A.; A Williamson, D.; Chow, E.P.F.; Fairley, C.K.; Hocking, J.S. Treatment efficacy for pharyngeal Neisseria gonorrhoeae: a systematic review and meta-analysis of randomized controlled trials. J. Antimicrob. Chemother. 2020, 75, 3109–3119. [Google Scholar] [CrossRef] [PubMed]
- Levin-Reisman, I.; Brauner, A.; Ronin, I.; Balaban, N.Q. Epistasis between antibiotic tolerance, persistence, and resistance mutations. Proc. Natl. Acad. Sci. 2019, 116, 14734–14739. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Ma, K.; Wan, I.; Willcox, M.D. Ciprofloxacin resistance and tolerance of Pseudomonas aeruginosa ocular isolates. Contact Lens Anterior Eye 2023, 46, 101819. [Google Scholar] [CrossRef]
- Manoharan-Basil S, Balduck M, Laumen J, Kenyon C. Transcriptomic profiling of ceftriaxone-tolerant phenotypes of Neisseria gonorrhoeae WHO P reference strain. ECCMID 2023, Copenhagen; 2023.
Figure 1.
TD test of clinical N. gonorrhoeae isolates (n = 66) with 0.75 µg AZM, 0.064 µg CRO, and 0.032 µg CIP 6 mm disks, performed in triplicate. The arrow indicates the presence of tolerant colonies after step III of the TD test.
Figure 1.
TD test of clinical N. gonorrhoeae isolates (n = 66) with 0.75 µg AZM, 0.064 µg CRO, and 0.032 µg CIP 6 mm disks, performed in triplicate. The arrow indicates the presence of tolerant colonies after step III of the TD test.
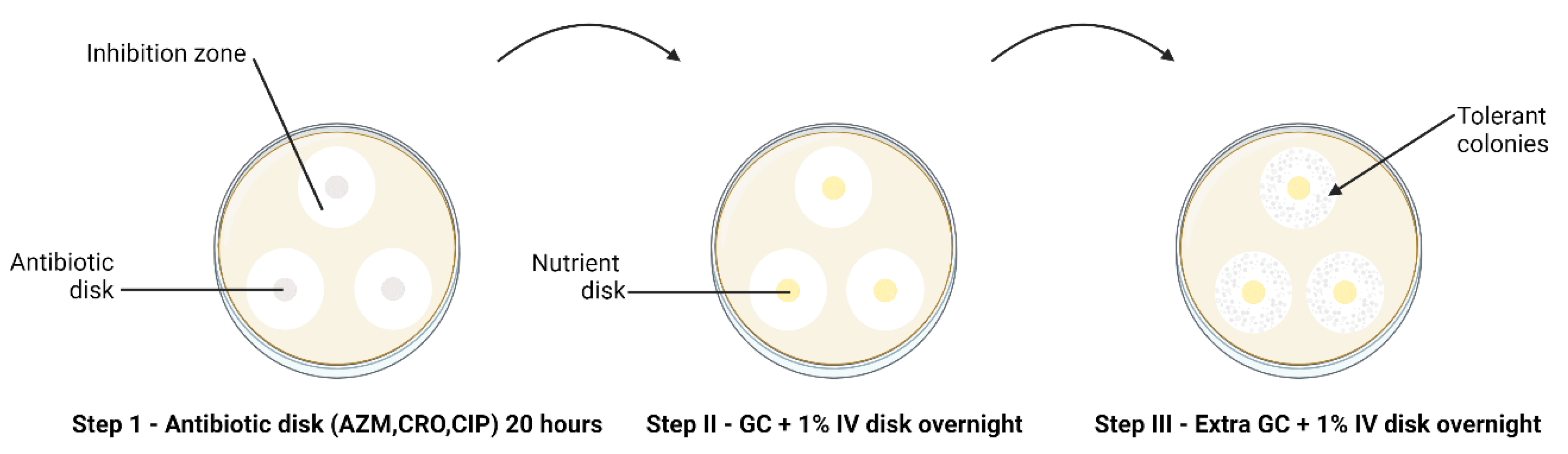
Figure 2.
(a) Relative paucity of commensal microbes and related pronounced immune response in the mucosa of the urogenital tract result in a decreased probability of tolerance developing in Neisseria gonorrhoeae (b) In contract the abundance of microbes in the GIT lumen is associated with immune tolerance. In this setting, incoming Neisseria gonorrhoeae is more likely to acquire tolerance (tolerance indicated by N. gonorrhoeae diplococci changing from red to green; DC – dendritic cell, IL – interleukin, PMN – polymorph neutrophil, TLR- toll like receptors, PAMPs – Pathogen associated molecular pattern, Figure produced in BioRender.com).
Figure 2.
(a) Relative paucity of commensal microbes and related pronounced immune response in the mucosa of the urogenital tract result in a decreased probability of tolerance developing in Neisseria gonorrhoeae (b) In contract the abundance of microbes in the GIT lumen is associated with immune tolerance. In this setting, incoming Neisseria gonorrhoeae is more likely to acquire tolerance (tolerance indicated by N. gonorrhoeae diplococci changing from red to green; DC – dendritic cell, IL – interleukin, PMN – polymorph neutrophil, TLR- toll like receptors, PAMPs – Pathogen associated molecular pattern, Figure produced in BioRender.com).
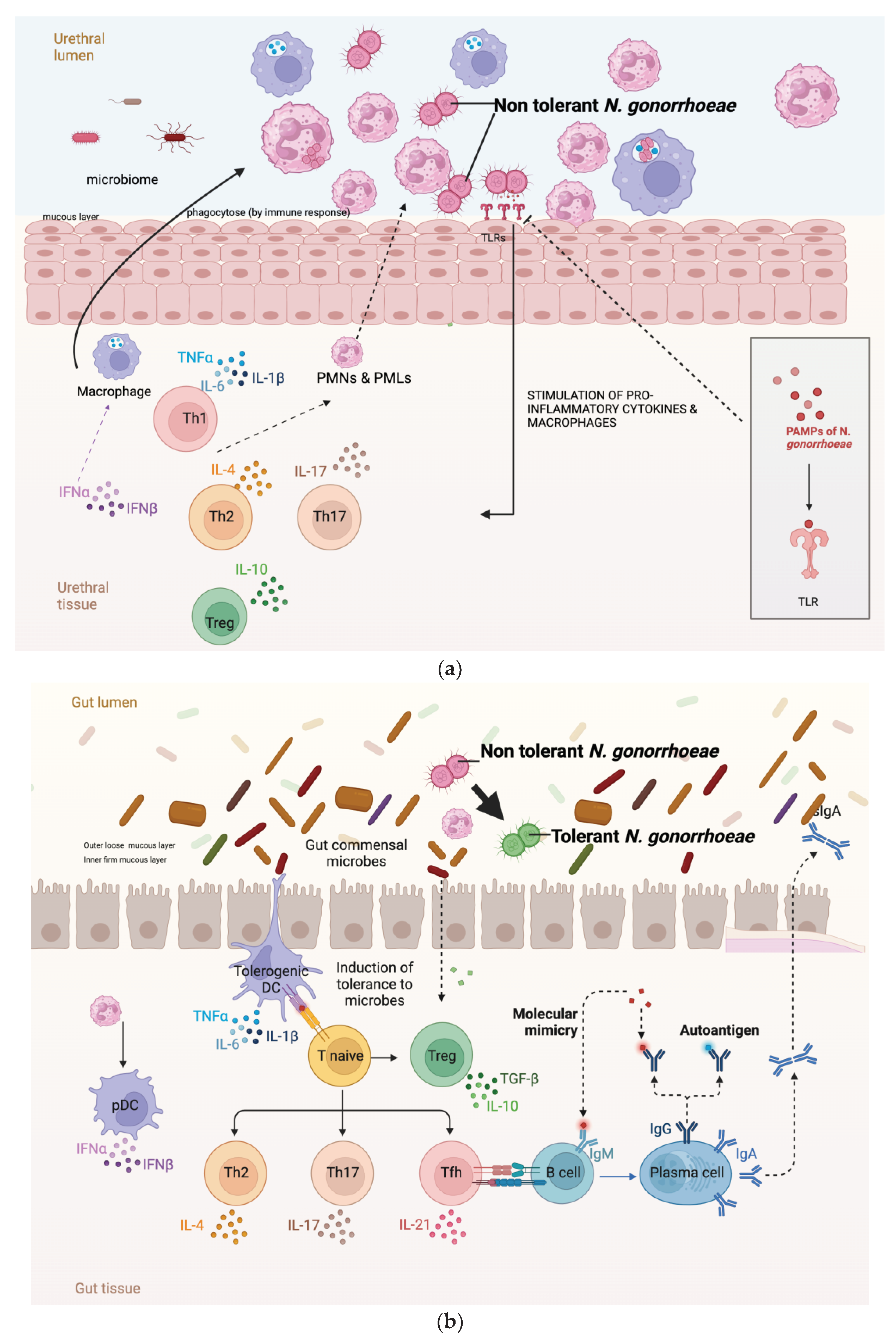

Table 1.
Azithromycin, ciprofloxacin and ceftriaxone tolerance and heterotolerance emergence following a TD test on 14 WHO N. gonorrhoeae reference strains.
Table 1.
Azithromycin, ciprofloxacin and ceftriaxone tolerance and heterotolerance emergence following a TD test on 14 WHO N. gonorrhoeae reference strains.
| Antibiotics | Tolerance | Heterotolerance | ||
| Number | WHO isolates | Number | WHO isolates | |
| Azithromycin | 1 | Z | 0 | |
| Ciprofloxacin | 2 | P, U | 1 | U |
| Ceftriaxone | 8 | K, M, N, O, P, U, V, W | 0 | |
Table 2.
Azithromycin, ciprofloxacin and ceftriaxone tolerance and heterotolerance emergence following a TD test on anorectal (n = 30) and urogenital (n = 32) N. gonorrhoeae clinical isolates. α - 0.05.
Table 2.
Azithromycin, ciprofloxacin and ceftriaxone tolerance and heterotolerance emergence following a TD test on anorectal (n = 30) and urogenital (n = 32) N. gonorrhoeae clinical isolates. α - 0.05.
| Antibiotics |
Tolerance | Heterotolerance | ||||
| Yes | No | Proportion | Yes | No | Proportion | |
| Azithromycin | ||||||
| Anorectal | 10 | 20 | 33% | 6 | 24 | 20% |
| Urogenital | 1 | 31 | 3% | 1 | 31 | 3% |
| p value | 0.002 | p value | 0.050 | |||
| Ciprofloxacin | ||||||
| Anorectal | 5 | 25 | 17% | 3 | 27 | 10% |
| Urogenital | 9 | 23 | 28% | 7 | 25 | 22% |
| p value | 0.367 | p value | 0.304 | |||
| Ceftriaxone | ||||||
| Anorectal | 10 | 20 | 33% | 4 | 26 | 13% |
| Urogenital | 3 | 29 | 9% | 1 | 31 | 3% |
| p value | 0.029 | p value | 0.189 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



