Submitted:
06 March 2024
Posted:
07 March 2024
You are already at the latest version
Abstract
Infections caused by KPC-producing K. pneumoniae continue to pose a significant clinical challenge due to emerging resistance to new antimicrobials. We investigated the association between two drugs whose roles have been repurposed against multidrug-resistant bacteria: fosfomycin and temocillin. Temocillin exhibits unusual stability against KPC enzymes, while fosfomycin acts as a potent “synergizer”. We conducted in vitro antimicrobial activity studies on 100 clinical isolates of KPC-producing K. pneumoniae using a combination of fosfomycin and temocillin. The results demonstrated synergistic activity in 91% of the isolates. Subsequently, we assessed the effect on Galleria mellonella larvae using 5 different KPC-Kp isolates. The addition of fosfomycin to temocillin increased larvae survival from 73 to 97% (+Δ 32%; isolate 1), from 93 to 100% (+Δ 7%; isolate 2), from 63 to 86% (+Δ 36%; isolate 3), from 63 to 90% (+Δ 42%; isolate 4), and from 93 to 97% (+Δ 4%; isolate 10), respectively. Among the temocillin-resistant KPC-producing K. pneumoniae isolates (24 isolates), the addition of fosfomycin reduced temocillin MIC values below the resistance breakpoint in all isolated except one. Temocillin combined with fosfomycin emerges as a promising combination against KPC-producing K. pneumoniae, warranting further clinical evaluation.
Keywords:
antibiotic
; combination
; fosfomycin
; Galleria mellonella
; in vivo
; KPC-producing Klebsiella pneumoniae
; resistance
; synergism
; temocillin
; therapy
1. Introduction
Infections caused by KPC-producing Klebsiella pneumoniae are associated with high mortality rates [1]. In Europe, from 2007 to 2015 the burden of carbapenem-resistant K. pneumoniae increased by 6.16 times both in terms of number of infections and number of deaths [2]. In patients with bloodstream infections (BSI) the resistance to carbapenems is associated with an excess of mortality [3]. It has been demonstrated that initiating the appropriate antibiotic therapy as early as possible for KPC-producing K. pneumoniae infections reduces mortality [4]. Currently, apart from colistin and eravacycline, we have four new beta-lactam drugs with activity against KPC-producing K. pneumoniae: cefiderocol, ceftazidime/avibactam, imipenem/relebactam and meropenem/vaborbactam. However, the microbial landscape is dynamic, with resistances emerging easily and increasingly reported [5]. According to the Infectious Diseases Society of America (IDSA) the preferred antibiotics for the treatment of infections outside of the urinary tract caused by carbapenem-resistant Enterobacterales (CRE) if KPC production is present are: meropenem-vaborbactam, followed by ceftazidime-avibactam, and then imipenem cilastatin-relebactam. Cefiderocol is listed as an alternative option [6]. The European Society of Clinical Microbiology and Infectious Diseases (ESCMID) guidelines recommend for patients with severe infections due to CRE meropenem-vaborbactam or ceftazidime-avibactam if active in vitro [7]. Combination therapies are not recommended by IDSA and ESCMID for CRE infections if susceptible to the new beta-lactams/beta-lactamase inhibitors. Moreover, antimicrobial stewardship principles advocate for the use of an antibiotic with the narrowest spectrum possible, at least for monomicrobial infections. In this context, temocillin, an old antibiotic, emerges as an interesting option. Temocillin is a semisynthetic derivative of ticarcillin, a carboxypenicillin commonly used in the ‘80-90 as an agent against Gram-negative bacteria [8]. Temocillin is used mainly for urinary tract infections (UTI), pneumonia, abdominal infections, and BSI. Generally, is well tolerated and it is associated with a low rate of C. difficile infections [8]. Temocillin exhibits unusual stability against extended-spectrum beta-lactamases (ESBL), AmpC and KPC enzymes [8], raising interest in drug repurposing. Indeed, at present, there are two ongoing clinical trials, whose aim is the comparison between temocillin and carbapenems in complicated UTI [9] and BSI [10] due to Gram-negative bacteria resistant to third generation cephalosporins. From an antimicrobial stewardship perspective, the inactivity of temocillin against metallo beta-lactamase and OXA-enzymes may be seen as an advantage, making it a strict anti-KPC agent. Fosfomycin is another old antibiotic with unique properties. It is a small molecule capable of synergizing with several other antibiotics, especially beta-lactams. It is a broad-spectrum antibiotic active against antimicrobial-resistant and multidrug-resistant (MDR) Gram-positive and Gram-negative bacteria including methicillin-resistant Staphylococcus aureus (MRSA), vancomycin-resistant enterococci (VRE), Enterobacterales, and Pseudomonas aeruginosa [11]. Moreover, both fosfomycin and temocillin have proven to be stable for at least 24 hours in elastomeric pumps, making them excellent options for outpatient parenteral antimicrobial therapy [12,13]. Temocillin offers the pharmacokinetic/pharmacodynamic advantages of a beta-lactam drug, while fosfomycin provides the advantages of a small molecule, with good tissue penetration and anti-biofilm activity. The combination of the two molecules theoretically helps in reducing bacterial minimum inhibitory concentration (MIC) against antibiotics without requiring a higher dose of the single drugs [14], potentially increasing bacterial killing and limiting drug-related toxicity. In the setting of MDR infections the number of in vitro and in vivo studies on different antimicrobial combination treatments are increasing [11,15]. To the best of our knowledge, currently there are only few studies on the synergistic activity in vitro and in vivo of temocillin and fosfomycin against Enterobacterales. Only one of these is focused on a KPC-producing E. coli infection in a murine model of peritonitis [16,17]. In light of this premise, we aimed to test the in vitro and in vivo potential of temocillin and fosfomycin combination against clinical isolates of KPC-producing K. pneumoniae.
2. Materials and Methods
Clinical bacterial isolates from various sample types (rectal swabs, urine, blood, bronchial aspirate) collected from January 2021 and August 2022 at the Microbiology Unit of the University Hospital of Trieste (Azienda Sanitaria Universitaria Giuliano Isontina) underwent screening for carbapenemase production using selective chromogenic agar medium (bioMérieux, Marcy-l’Etoile, France) as part of routine laboratory activities. Only one isolate per patient was studied. Species identification on bacterial isolates on selective medium was performed using MALDI-TOF mass spectrometry (bioMérieux, Marcy-l'Etoile, France), while antimicrobial susceptibility was assessed by the Vitek2 system (bioMérieux, Marcy-l'Etoile, France). The susceptibility profile of temocillin and fosfomycin was assessed using the Etest (bioMérieux, Marcy-l'Etoile, France). Isolates with meropenem MIC values greater than 0.125 mg/L underwent further investigated for carbapenemase production by PCR (Cepheid, Sunnyvale, California, United States) according to EUCAST criteria for detection of resistance mechanisms (https://www.eucast.org). Synergy tests between temocillin and fosfomycin were conducted for KPC-producing K. pneumoniae isolates. Although the recommended testing for fosfomycin is agar dilution, we used Etest also for fosfomycin since it allowed us to perform synergy testing Standard MIC of drugs A and B before creating the synergy. The "MTS Synergy Applicator System" was used for the synergy test. In particular, MIC Test Strip (MTS) of the antibiotic (A) was positioned on the MTS Synergy applicator platform. MTS (antibiotic A) was adjusted such that the MIC value of the first antibiotic (MICA) was placed at the intersection of the base. An MTS of the second antibiotic (B) was placed on the second base. The second STD (antibiotic B) was adjusted such that the MIC value of the second antibiotic (MICB) was positioned at the base intersection and intersects MTS-antibiotic A at its MIC value. Finally, the plates were incubated according to the standard MTS procedure for the specific microorganism [18]. EUCAST breakpoints were used to value susceptibility to temocillin and fosfomycin [19].
2.1. In Vivo Toxicity of Antibiotic Compounds
We have previously determined that fosfomycin is not toxic to Galleria mellonella larvae at doses of up to 512 mg/L [20]. In vivo toxicity of temocillin was determined according to the approach described previously for zidovudine and fosfomycin [21]. G. mellonella were purchased from LiveFoods UK Ltd (UK) and stored at 15°C prior to use. Only larvae weighing between 0.7 g and 1.3 g, showing no discoloration or injury were used in the assays. Groups of 10 larvae were injected into the proleg using a 26S gauge sterile syringe with either temocillin (48 mg/L) and separately, a combination of temocillin (48 mg/L) and fosfomycin (32 mg/L). These concentrations are equivalent to the highest MIC observed for each agent against K. pneumoniae isolates used in this study. Larvae mock-infected with phosphate buffered saline (PBS) were used as injection controls. Following inoculation, all larvae were incubated aerobically at 37°C for 72 hours and were assessed for mortality at 24 hour intervals. Larvae were classed as dead when an absence of movement in response to stimuli was observed.
2.2. In Vivo Challenge and Antibiotic Therapy
A single colony of the test K. pneumoniae isolates was harvested from a freshly streaked Luria-Bertani (LB) agar plate. The colony was then transferred to 10 ml of lysogeny broth and incubated at 37°C, with gentle agitation (150 rpm), under aerobic conditions for 16 hours. Following incubation, the cultures were serially diluted 10-fold in 1x PBS and enumerated on LB agar in triplicate. Groups of 10 G. mellonella larvae were placed in Petri Dishes, and challenged with 5000 CFU/ml of either K. pneumoniae isolate 1, 2, 3, 4, and 10-reference (Table 1). Additionally, a group of 10 larvae were mock-infected with 10 µl of sterile PBS. Within 15 minutes of infection, a second injection into the top right proleg was performed to administer fosfomycin, temocillin, a combination of both agents or PBS control. The concentration of each drug was equal to that used in combination with time-kill assays. Larvae were incubated aerobically at 37°C and scored for mortality at 0, 24, 48, and 72 hours post-infection. All assays were performed in triplicate and the data was plotted using GraphPad Prism 8.4.3 software (GraphPad Software Inc., San Diego, California, United States).
Results
2.3. Microbiological Findings
One-hundred non-repetitive K. pneumoniae clinical isolates were investigated. All isolates were recovered from different patients (n=100) and were KPC producers resistant to carbapenems (MICs >1 mg/L). Isolates were from rectal swabs (n=62), urine (n=29), blood culture (n=7), and bronchial aspirate (n=2). Isolates were recovered from patients hospitalized in the Medical Department (n=68), Surgical Department (n=9), Intensive Care Unit (n=22), and Emergency Department (n=1). In particular, among Medical Department patients, about 40% were from the Infectious Diseases Unit (n=27). Thirty-four isolates were resistant to temocillin (MICs >16 mg/L), while no resistant isolate was recovered for fosfomycin (MICs ≤ 32 mg/L). The fosfomycin-temocillin combination showed a prevalent synergistic activity against 91% of KPC-producing K. pneumoniae isolates, while additive activity was obtained against 8% of them. Only in one case indifferent activity was revealed. No antagonism was detected (Table 1). In particular, synergistic activity lowered temocillin MIC values under the resistance breakpoint (MICs ≤16 mg/L) for all isolates except one (99%), with a FIC index ranging from 0.21 to 1 considering all combinations (Table 1).
2.4. In Vivo Findings
Temocillin was determined to not be toxic to G. mellonella larvae, either alone or in combination with fosfomycin. A variable degree of synergism was observed between temocillin and fosfomycin in time-kill assays of K. pneumoniae. This synergism was recapitulated in vivo using the G. mellonella infection model, using select K. pneumoniae isolates. We observed clear increases in survival rates of larvae treated with combination therapy compared to either agent alone (Figure 1).
Larvae infected with K. pneumoniae isolate 1 and treated with a combination of temocillin and fosfomycin exhibited a survival rate of 96.6%, compared to 90% for fosfomycin alone, and 73.3% for temocillin alone (p=0.0252, n=30). In larvae infected with K. pneumoniae isolate 2, a combination of both antimicrobials resulted in a 100% survival rate, relative to a 93% survival rate for temocillin or fosfomycin monotherapy. K. pneumoniae isolate 3 elicited 90% mortality, however combination of temocillin at 24 and fosfomycin at 8 significantly improved survival rate relative to 86% relative to temocillin alone 63% (p=0.0768, n=30), or fosfomycin alone 53% (p=0.0205, n=30). Combination treatment of larvae infected with K. pneumoniae isolate 4 resulted in a survival rate of 90%, compared to a survival rate of 46% without antibiotic therapy, and 63% for temocillin alone (p=0.0344, n=30) and 73% for fosfomycin alone (p=0.1915, n=30). The survival rate of larvae infected with K. pneumoniae isolate 10 was 76.6%. Treatment of larvae with temocillin increased survival to 93.3%, and treatment with fosfomycin alone increased survival to 96.6%. Combination therapy with both of these agents resulted in a survival rate of 96.6% (Figure 1).
3. Discussion
KPC-producing isolates represent an important public health issue. Recently, emerging data on food-producing animals, and their products, as reservoirs for KPC-producing isolates are also being reported in different countries [22,23]. New beta-lactams/beta-lactamase inhibitors are now considered a first-line treatment option against KPC-producing isolates. Nevertheless, the widespread clinical use of ceftazidime/avibactam has forced CRE to mutate to adapt to the increasing antibiotic pressure. Several isolates carrying different KPC variants and resistant to ceftazidime/avibactam are emerging worldwide [5]. Resistance to ceftazidime/avibactam in Enterobacterales is commonly due to three different mechanisms: enzymatic alterations causing inactivation of the antibiotics; modification of the antibiotic target or expressions of an alternative target; and changes in cell permeability or expression of efflux pumps. Modification of β-lactamase hydrolytic properties due to specific mutations within class A carbapenemase is the most common mechanism related to ceftazidime/avibactam-resistance in Enterobacterales [24]. By March 2023, 145 blaKPC variants are registered in the National Center for Biotechnology Information (NCBI) database, all derived from mutations of blaKPC-2 or blaKPC-3. Overall, blaKPC-4 (n = 26), blaKPC-33 (n = 9), blaKPC-12 (n = 8), blaKPC-6 (n = 5), blaKPC-71 (n = 4), blaKPC-10 (n = 3), blaKPC-76 (n = 3), blaKPC-44 (n = 3), blaKPC-25 (n = 2), blaKPC-36 (n = 2), blaKPC-5 (n = 2), and blaKPC-90 (n = 2) were mutants from blaKPC-2. These KPC-2 variants were mainly identified from the US (n = 35), China (n = 20), and Italy (n = 3). In contrast, blaKPC-31 (n = 8), blaKPC-66 (n = 4), blaKPC-67 (n = 3), blaKPC-18 (n = 2), blaKPC-29 (n = 2), blaKPC-40 (n = 2), blaKPC-49 (n = 2), blaKPC-61 (n = 2), and blaKPC-70 (n = 2) were mutants from blaKPC-3. These blaKPC-3 variants were mainly reported from Italy (n = 15), the US (n = 8), and France (n = 1) [25]. Resistance to ceftazidime/avibactam represents a serious cause of concern, with cases of resistance especially reported from US, Greece and Italy. Moreover, an important rate of KPC producer strains worldwide (about 30%) shows a baseline resistance, mostly due to impermeability mechanisms. This is an important point, because isolates with baseline resistance to ceftazidime/avibactam could represent a reservoir of resistance that could be potentially enhanced under inappropriate ceftazidime/avibactambased treatment. These isolates commonly remain susceptible to meropenem/vaborbactam, imipenem/relebactam and cefiderocol, among the new beta-lactam options. Moreover, some isolates carrying specific KPC variants or increased enzyme expression in association with porin defects/loss showed resistance also to meropenem/vaborbactam and imipenem/relebactam [25,26]. Although resistance to meropenem/vaborbactam has been associated with decreased expression of ompK35 and ompK36 and concomitantly increased expression of blaKPC, MIC seems to be unaffected by an increase in expression of the blaKPC gene and efflux pump (acrB), or decreased expression of ompK35 alone [27]. Among Enterobacterales and other Gram-negative bacteria efflux pump systems, in particular AcrAB-TolC, are common resistance mechanisms against multiple antibiotic classes. Overexpression of acrAB in association with inactivated OmpK35 and OmpK36 porins increased the MIC of meropenem/vaborbactam [28]. Previous studies demonstrated that imipenem/relebactam resistance was associated with mutations resulting in a non-functional OmpK35 and OmpK36 porins in KPC-producing K. pneumoniae strains, or AmpC overexpression in combination with porins loss [29]. Similarly, resistance to cefiderocol, although still few, is increasingly being reported [30,31]. In this context, some old antibiotics such as temocillin and fosfomycin could represent beta-lactams/beta-lactamase inhibitors- and cefiderocol-sparing options, but also may provide pharmacokinetic/pharmacodynamic and biochemical advantages (e.g. central nervous system penetration, anti-biofilm activity, etc.) hence improving the antimicrobial armamentarium against KPC-producing isolates. Fosfomycin in combination with beta-lactams, represents a classical (and historical) synergistic option. Beta-lactam antibiotics target the bacterial cell wall penicillin-binding proteins, while fosfomycin inhibits the peptidoglycan precursor UDP N-acetylmuramic acid involved in peptidoglycan biosynthesis, hence providing a greater chance of synergistic/additive effects against Gram-negative isolates. Among beta-lactams, the best synergistic activity against KPC-producing K. pneumoniae isolates was found in ceftazidime/avibactam. Fosfomycin demonstrated synergistic activity against KPC-producing K. pneumoniae isolates with tigecycline, colistin, and aminoglycosides [11]. Moreover, recently, synergistic activity between fosfomycin and meropenem/vaborbactam [32,33], as well as between fosfomycin and cefiderocol [34], was also reported. In the present study we assessed the antimicrobial interaction among these drugs against KPC-producing K. pneumoniae clinical isolates, demonstrating a significant in vitro and in vivo synergistic activity. In particular, in vitro synergistic activity between temocillin and fosfomycin was demonstrated against 91% of KPC-producing K. pneumoniae isolates, lowering the temocillin MIC values under the resistance breakpoint in all cases. In the animal study, we tested the temocillin and fosfomycin combination on 5 different KPC-producing K. pneumoniae isolates. The addition of fosfomycin to temocillin increased larvae survival from 73 to 97% (+Δ 32%; isolate 1), from 93 to 100% (+Δ 7%; isolate 2), from 63 to 86% (+Δ 36%; isolate 3), from 63 to 90% (+Δ 42%; isolate 4), and from 93 to 97% (+Δ 4%; isolate 10). To our knowledge, another study evaluated in vitro and in vivo the synergism of fosfomycin and temocillin against Enterobacterales KPC-producers, in particular an E. coli KPC-3 producer strain. In vitro the addition of fosfomycin reduced temocillin MIC 16-fold for E. coli KPC-3 with a FICI of 0.562, showing an additive effect. This was eventually demonstrated in vivo, in murine models of peritonitis [16]. The combination of fosfomycin-temocillin was also found to be synergistic in 27% of K. pneumoniae strains in a observational study on Gram-negative invasive isolates of the UK. However, no KPC-producing Enterobacterales were reported [17].
Our study has limitations. First, it is monocentric. The epidemiology, especially for Gram-negative infections, could vary significantly among different areas within the same country. Thus, our study reflects only the local epidemiology. Secondly, we didn't include many invasive clinical isolates. Third, we didn't perform genotype analysis on our KPC-producing K. pneumoniae isolates; thus, we cannot provide any data regarding the presence of any KPC variants.
In conclusions, this study represents the first in vitro and in vivo description of temocillin-fosfomycin synergistic interaction against KPC-producing K. pneumoniae isolates. Temocillin combined with fosfomycin emerges as a promising combination against KPC-producing K. pneumoniae, warranting further clinical investigations.
Author Contributions
Conceptualization, S.D.B., V.C. and L.P.; methodology, V.C. and J.M.; validation, R.L.R. and M.B.; formal analysis, R.L.; investigation, V.C. and J.M.; resources, R.L.R.; data curation, V.C. and J.M.; writing—original draft preparation, S.D.B., V.Z., A.M. and L.P.; writing—review and editing, S.D.B. and A.M.; visualization, L.P.; supervision, R.L. and R.L.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
No procedures were undertaken using animals protected by the Animals in Scientific Procedures Act (1986).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Tumbarello, M.; Viale, P.; Viscoli, C.; Trecarichi, E.M.; Tumietto, F.; Marchese, A.; Spanu, T.; Ambretti, S.; Ginocchio, F.; Cristini, F.; et al. Predictors of Mortality in Bloodstream Infections Caused by Klebsiella Pneumoniae Carbapenemase-Producing K. Pneumoniae: Importance of Combination Therapy. Clin. Infect. Dis. 2012, 55, 943–950. [Google Scholar] [CrossRef]
- Cassini, A.; Högberg, L.D.; Plachouras, D.; Quattrocchi, A.; Hoxha, A.; Simonsen, G.S.; Colomb-Cotinat, M.; Kretzschmar, M.E.; Devleesschauwer, B.; Cecchini, M.; et al. Attributable Deaths and Disability-Adjusted Life-Years Caused by Infections with Antibiotic-Resistant Bacteria in the EU and the European Economic Area in 2015: A Population-Level Modelling Analysis. Lancet Infect. Dis. 2019, 19, 56–66. [Google Scholar] [CrossRef]
- Falcone, M.; Tiseo, G.; Carbonara, S.; Marino, A.; Di Caprio, G.; Carretta, A.; Mularoni, A.; Mariani, M.F.; Maraolo, A.E.; Scotto, R.; et al. Mortality Attributable to Bloodstream Infections Caused by Different Carbapenem-Resistant Gram-Negative Bacilli: Results From a Nationwide Study in Italy (ALARICO Network). Clin. Infect. Dis. 2023, 76, 2059–2069. [Google Scholar] [CrossRef]
- Falcone, M.; Bassetti, M.; Tiseo, G.; Giordano, C.; Nencini, E.; Russo, A.; Graziano, E.; Tagliaferri, E.; Leonildi, A.; Barnini, S.; et al. Time to Appropriate Antibiotic Therapy Is a Predictor of Outcome in Patients with Bloodstream Infection Caused by KPC-Producing Klebsiella Pneumoniae. Crit. Care 2020, 24, 29. [Google Scholar] [CrossRef]
- Di Bella, S.; Giacobbe, D.R.; Maraolo, A.E.; Viaggi, V.; Luzzati, R.; Bassetti, M.; Luzzaro, F.; Principe, L. Resistance to Ceftazidime/avibactam in Infections and Colonisations by KPC-Producing Enterobacterales: A Systematic Review of Observational Clinical Studies. J. Glob. Antimicrob. Resist. 2021, 25, 268–281.6. [Google Scholar] [CrossRef] [PubMed]
- Tamma, P.D.; Aitken, S.L.; Bonomo, R.A.; Mathers, A.J.; van Duin, D.; Clancy, C.J. Infectious Diseases Society of America 2023 Guidance on the Treatment of Antimicrobial Resistant Gram-Negative Infections. Clin. Infect. Dis. 2023. [Google Scholar] [CrossRef]
- Paul, M.; Carrara, E.; Retamar, P.; Tängdén, T.; Bitterman, R.; Bonomo, R.A.; de Waele, J.; Daikos, G.L.; Akova, M.; Harbarth, S.; et al. European Society of Clinical Microbiology and Infectious Diseases (ESCMID) Guidelines for the Treatment of Infections Caused by Multidrug-Resistant Gram-Negative Bacilli (endorsed by European Society of Intensive Care Medicine). Clin. Microbiol. Infect. 2022, 28, 521–547. [Google Scholar] [CrossRef]
- Lupia, T.; De Benedetto, I.; Stroffolini, G.; Di Bella, S.; Mornese Pinna, S.; Zerbato, V.; Rizzello, B.; Bosio, R.; Shbaklo, N.; Corcione, S.; et al. Temocillin: Applications in Antimicrobial Stewardship as a Potential Carbapenem-Sparing Antibiotic. Antibiotics (Basel) 2022, 11. [Google Scholar] [CrossRef]
- Temocillin Versus a Carbapenem as Initial Intravenous Treatment for ESBL Related Urinary Tract Infections (www.ClinicalTrials.gov, accessed 2nd march 2024).
- Marín-Candón, A.; Rosso-Fernández, C.M.; Bustos de Godoy, N.; López-Cerero, L.; Gutiérrez-Gutiérrez, B.; López-Cortés, L.E.; Barrera Pulido, L.; Borreguero Borreguero, I.; León, M.J.; Merino, V.; et al. Temocillin versus Meropenem for the Targeted Treatment of Bacteraemia due to Third-Generation Cephalosporin-Resistant (ASTARTÉ): Protocol for a Randomised, Pragmatic Trial. BMJ Open 2021, 11, e049481. [Google Scholar] [CrossRef] [PubMed]
- Antonello, R.M.; Principe, L.; Maraolo, A.E.; Viaggi, V.; Pol, R.; Fabbiani, M.; Montagnani, F.; Lovecchio, A.; Luzzati, R.; Di Bella, S. Fosfomycin as Partner Drug for Systemic Infection Management. A Systematic Review of Its Synergistic Properties from In Vitro and In Vivo Studies. Antibiotics (Basel) 2020, 9. [Google Scholar] [CrossRef]
- Carryn, S.; Couwenbergh, N.; Tulkens, P.M. Long-Term Stability of Temocillin in Elastomeric Pumps for Outpatient Antibiotic Therapy in Cystic Fibrosis Patients. J. Antimicrob. Chemother. 2010, 65, 2045–2046. [Google Scholar] [CrossRef] [PubMed]
- Manca, A.; Palermiti, A.; Mula, J.; Cusato, J.; Maiese, D.; Simiele, M.; De Nicolò, A.; D’Avolio, A. Stability Study of Fosfomycin in Elastomeric Pumps at 4 °C and 34 °C: Technical Bases for a Continuous Infusion Use for Outpatient Parenteral Antibiotic Therapy. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef]
- Antonello, R.M.; Di Bella, S.; Maraolo, A.E.; Luzzati, R. Fosfomycin in Continuous or Prolonged Infusion for Systemic Bacterial Infections: A Systematic Review of Its Dosing Regimen Proposal from in Vitro, in Vivo and Clinical Studies. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 1117–1126. [Google Scholar] [CrossRef] [PubMed]
- Tumbarello, M.; Raffaelli, F.; Giannella, M.; Mantengoli, E.; Mularoni, A.; Venditti, M.; De Rosa, F.G.; Sarmati, L.; Bassetti, M.; Brindicci, G.; et al. Ceftazidime-Avibactam Use for Klebsiella Pneumoniae Carbapenemase-Producing K. Pneumoniae Infections: A Retrospective Observational Multicenter Study. Clin. Infect. Dis. 2021, 73, 1664–1676. [Google Scholar] [CrossRef]
- Berleur, M.; Guérin, F.; Massias, L.; Chau, F.; Poujade, J.; Cattoir, V.; Fantin, B.; de Lastours, V. Activity of Fosfomycin Alone or Combined with Temocillin in Vitro and in a Murine Model of Peritonitis due to KPC-3- or OXA-48-Producing Escherichia Coli. J. Antimicrob. Chemother. 2018, 73, 3074–3080. [Google Scholar] [CrossRef] [PubMed]
- Suich, J.; Mawer, D.; van der Woude, M.; Wearmouth, D.; Burns, P.; Smeets, T.; Barlow, G. Evaluation of Activity of Fosfomycin, and Synergy in Combination, in Gram-Negative Bloodstream Infection Isolates in a UK Teaching Hospital. J. Med. Microbiol. 2022, 71. [Google Scholar] [CrossRef]
- Carretto, E.; Brovarone, F.; Gaibani, P.; Russello, G.; Farina, C. Multicentric evaluation of the reliability and reproducibility of synergy testing using the MIC strip test – synergy applications system (MTS-SAS™). ECCMID 2016. Poster 0802.
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint tables for interpretation of MICs and zone diameters. Version 14.0, 2024. http://www.eucast.org.
- Lagatolla, C.; Mehat, J.W.; La Ragione, R.M.; Luzzati, R.; Di Bella, S. In Vitro and In Vivo Studies of Oritavancin and Fosfomycin Synergism against Vancomycin-Resistant. Antibiotics (Basel) 2022, 11. [Google Scholar] [CrossRef]
- Antonello, R.M.; Di Bella, S.; Betts, J.; La Ragione, R.; Bressan, R.; Principe, L.; Morabito, S.; Gigliucci, F.; Tozzoli, R.; Busetti, M.; et al. Zidovudine in Synergistic Combination with Fosfomycin: An in Vitro and in Vivo Evaluation against Multidrug-Resistant Enterobacterales. Int. J. Antimicrob. Agents 2021, 58, 106362. [Google Scholar] [CrossRef]
- Valiatti, T.B.; Cayô, R.; Santos, F.F.; Bessa-Neto, F.O.; Brandão Silva, R.G.; Veiga, R.; de Nazaré Miranda Bahia, M.; Guerra, L.M.G.D.; Pignatari, A.C.C.; de Oliveira Souza, C.; Brasiliense, D.M.; Gales, A.C.; Guarani Network. Genomic Analysis of Klebsiella pneumoniae ST258 Strain Coproducing KPC-2 and CTX-M-14 Isolated from Poultry in the Brazilian Amazon Region. Antibiotics (Basel). 2022, 11, 1835. [Google Scholar] [CrossRef]
- Bonardi, S.; Cabassi, C.S.; Fiaccadori, E.; Cavirani, S.; Parisi, A.; Bacci, C.; Lamperti, L.; Rega, M.; Conter, M.; Marra, F.; Crippa, C.; Gambi, L.; Spadini, C.; Iannarelli, M.; Paladini, C.; Filippin, N.; Pasquali, F. Detection of carbapenemase- and ESBL-producing Klebsiella pneumoniae from bovine bulk milk and comparison with clinical human isolates in Italy. Int. J. Food Microbiol. 2023, 387, 110049. [Google Scholar] [CrossRef]
- Gaibani, P.; Giani, T.; Bovo, F.; Lombardo, D.; Amadesi, S.; Lazzarotto, T.; Coppi, M.; Rossolini, G.M.; Ambretti, S. Resistance to Ceftazidime/Avibactam, Meropenem/Vaborbactam and Imipenem/Relebactam in Gram-Negative MDR Bacilli: Molecular Mechanisms and Susceptibility Testing. Antibiotics (Basel). 2022, 11, 628. [Google Scholar] [CrossRef]
- Ding, L.; Shen, S.; Chen, J.; Tian, Z.; Shi, Q.; Han, R.; Guo, Y.; Hu, F. Klebsiella pneumoniae carbapenemase variants: the new threat to global public health. Clin. Microbiol. Rev. 2023, 36, e0000823. [Google Scholar] [CrossRef]
- Di Pilato, V.; Principe, L.; Andriani, L.; Aiezza, N.; Coppi, M.; Ricci, S.; Giani, T.; Luzzaro, F.; Rossolini, G.M. Deciphering Variable Resistance to Novel Carbapenem-Based β-Lactamase Inhibitor Combinations in a Multi-Clonal Outbreak Caused by Klebsiella Pneumoniae Carbapenemase (KPC)-Producing Klebsiella Pneumoniae Resistant to Ceftazidime/avibactam. Clin. Microbiol. Infect. 2023, 29, 537.e1–e537.e8. [Google Scholar] [CrossRef]
- Jorgensen, S.C.J.; Rybak, M.J. Meropenem and Vaborbactam: Stepping up the Battle against Carbapenem-resistant Enterobacteriales. Pharmacotherapy 2018, 38, 444–461. [Google Scholar] [CrossRef] [PubMed]
- Lomovskaya, O.; Sun, D.; Rubio-Aparicio, D.; Nelson, K.; Tsivkovski, R.; Griffith, D.C.; Dudley, M.N. Vaborbactam: Spectrum of β-lactamase inhibition and impact of resistance mechanisms on activity in Enterobacteriales. Antimicrob. Agents Chemother. 2017, 61, e01443-17. [Google Scholar] [CrossRef] [PubMed]
- Titov, I.; Wunderink, R.G.; Roquilly, A.; Gonzalez, D.R.; David-Wang, A.; Boucher, H.W.; Kaye, K.S.; Losada, M.C.; Du, J.; Tipping, R.; et al. A Randomized, Double-blind, Multicenter Trial Comparing Efficacy and Safety of Imipenem/Cilastatin/Relebactam Versus Piperacillin/Tazobactam in Adults with Hospital-acquired or Ventilator-associated Bacterial Pneumonia (RESTOREIMI 2 Study). Clin. Infect. Dis. 2020, 73, e4539–e4548. [Google Scholar] [CrossRef]
- Bovo, F.; Amadesi, S.; Palombo, M.; Lazzarotto, T.; Ambretti, S.; Gaibani, P. Clonal Dissemination of Resistant to Cefiderocol, Ceftazidime/avibactam, Meropenem/vaborbactam and Imipenem/relebactam Co-Producing KPC and OXA-181 Carbapenemase. JAC Antimicrob. Resist. 2023, 5, dlad099. [Google Scholar] [CrossRef]
- Bellinzona, G.; Merla, C.; Corbella, M.; Iskandar, E.N.; Seminari, E.; Di Matteo, A.; Gaiarsa, S.; Petazzoni, G.; Sassera, D.; Baldanti, F.; et al. Concomitant Resistance to Cefiderocol and Ceftazidime/Avibactam in Two Carbapenemase-Producing Isolates from Two Lung Transplant Patients. Microb. Drug Resist. 2024, 30, 21–26. [Google Scholar] [CrossRef]
- Oliva, A.; Curtolo, A.; Volpicelli, L.; Cogliati, D.F.; De Angelis, M.; Cairoli, S.; Dell’Utri, D.; Goffredo, B.M.; Raponi, G.; Venditti, M. Synergistic Meropenem/Vaborbactam Plus Fosfomycin Treatment of KPC Producing K. Pneumoniae Septic Thrombosis Unresponsive to Ceftazidime/Avibactam: From the Bench to the Bedside. Antibiotics (Basel, Switzerland) 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Boattini, M.; Bianco, G.; Iannaccone, M.; Bondi, A.; Cavallo, R.; Costa, C. Looking beyond Ceftazidime-Avibactam Resistance in KPC-Producing Klebsiella Pneumoniae: In Vitro Activity of the Novel Meropenem-Vaborbactam in Combination with the Old Fosfomycin. J. Chemother. 2021, 33. [Google Scholar] [CrossRef] [PubMed]
- Palombo, M.; Bovo, F.; Amadesi, S.; Gaibani, P. Synergistic Activity of Cefiderocol in Combination with Piperacillin-Tazobactam, Fosfomycin, Ampicillin-Sulbactam, Imipenem-Relebactam and Ceftazidime-Avibactam against Carbapenem-Resistant Gram-Negative Bacteria. Antibiotics (Basel, Switzerland) 2023, 12. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
In vivo combination therapy (fosfomycin + temocillin) – survival curves.
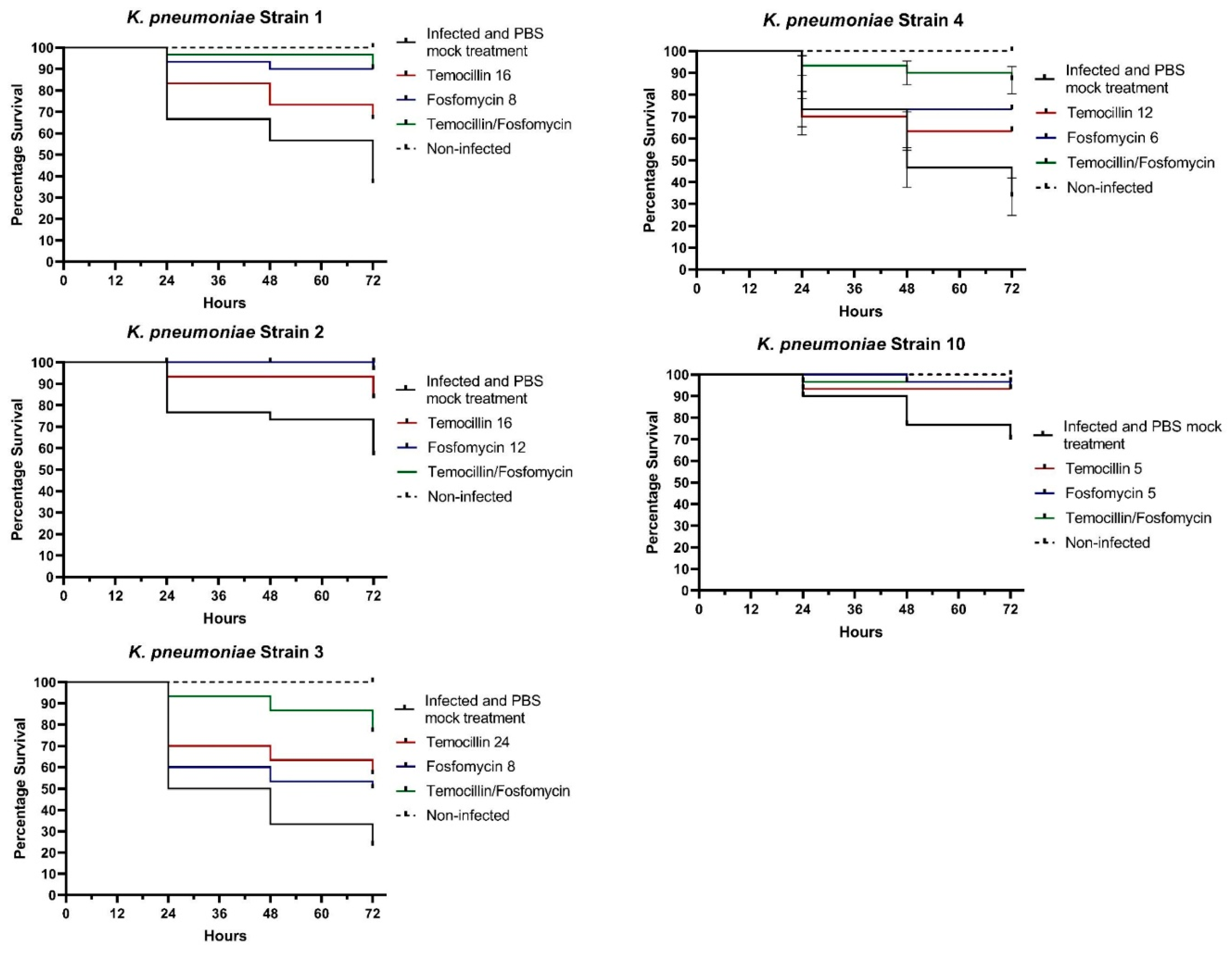

Table 1.
Fosfomycin + Temocillin: in vitro synergy testing.
| # | MIC FOS | MIC TEM | MIC FOS/TEM | MIC TEM/FOS | FICI | Interpretation |
|---|---|---|---|---|---|---|
| 1 | 8 | 16 | 1 | 2 | 0,25 | SYNERGISM |
| 2 | 12 | 16 | 3 | 4 | 0,5 | SYNERGISM |
| 3 | 8 | 24 | 3 | 6 | 0,62 | ADDITIVE |
| 4 | 6 | 12 | 1,5 | 2 | 0,41 | SYNERGISM |
| 5 | 4 | 8 | 1 | 2 | 0,5 | SYNERGISM |
| 6 | 8 | 12 | 2 | 3 | 0,5 | SYNERGISM |
| 7 | 6 | 8 | 1 | 2 | 0,41 | SYNERGISM |
| 8 | 12 | 16 | 1 | 3 | 0,27 | SYNERGISM |
| 9 | 12 | 24 | 3 | 6 | 0,5 | SYNERGISM |
| 10 | 4 | 6 | 0,75 | 1,5 | 0,43 | SYNERGISM |
| 11 | 6 | 12 | 1,5 | 2 | 0,41 | SYNERGISM |
| 12 | 16 | 24 | 3 | 8 | 0,52 | ADDITIVE |
| 13 | 12 | 24 | 3 | 4 | 0,41 | SYNERGISM |
| 14 | 4 | 8 | 0,75 | 2 | 0,43 | SYNERGISM |
| 15 | 12 | 16 | 2 | 3 | 0,35 | SYNERGISM |
| 16 | 24 | 32 | 18 | 16 | 1,25 | INDIFFERENT |
| 17 | 16 | 32 | 4 | 6 | 0,43 | SYNERGISM |
| 18 | 16 | 24 | 4 | 6 | 0,5 | SYNERGISM |
| 19 | 12 | 16 | 2 | 4 | 0,41 | SYNERGISM |
| 20 | 8 | 16 | 2 | 3 | 0,43 | SYNERGISM |
| 21 | 8 | 16 | 1 | 2 | 0,25 | SYNERGISM |
| 22 | 12 | 16 | 1,5 | 3 | 0,31 | SYNERGISM |
| 23 | 4 | 6 | 0,5 | 1,5 | 0,37 | SYNERGISM |
| 24 | 6 | 8 | 1 | 1,5 | 0,35 | SYNERGISM |
| 25 | 8 | 12 | 2 | 3 | 0,5 | SYNERGISM |
| 26 | 32 | 48 | 6 | 12 | 0,43 | SYNERGISM |
| 27 | 12 | 16 | 3 | 4 | 0,5 | SYNERGISM |
| 28 | 8 | 16 | 1,5 | 4 | 0,43 | SYNERGISM |
| 29 | 12 | 24 | 2 | 6 | 0,41 | SYNERGISM |
| 30 | 24 | 32 | 6 | 8 | 0,5 | SYNERGISM |
| 31 | 16 | 32 | 4 | 8 | 0,5 | SYNERGISM |
| 32 | 8 | 12 | 2 | 3 | 0,5 | SYNERGISM |
| 33 | 4 | 6 | 0,5 | 1 | 0,29 | SYNERGISM |
| 34 | 6 | 8 | 0,75 | 2 | 0,37 | SYNERGISM |
| 35 | 8 | 12 | 1 | 3 | 0,37 | SYNERGISM |
| 36 | 16 | 24 | 3 | 6 | 0,43 | SYNERGISM |
| 37 | 24 | 48 | 6 | 12 | 0,5 | SYNERGISM |
| 38 | 8 | 16 | 2 | 6 | 0,62 | ADDITIVE |
| 39 | 12 | 16 | 1 | 3 | 0,27 | SYNERGISM |
| 40 | 4 | 6 | 1 | 1,5 | 0,5 | SYNERGISM |
| 41 | 12 | 16 | 2 | 3 | 0,35 | SYNERGISM |
| 42 | 8 | 16 | 0,75 | 3 | 0,28 | SYNERGISM |
| 43 | 12 | 16 | 1 | 2 | 0,20 | SYNERGISM |
| 44 | 8 | 12 | 0,75 | 3 | 0,34 | SYNERGISM |
| 45 | 4 | 8 | 0,38 | 1 | 0,22 | SYNERGISM |
| 46 | 12 | 16 | 2 | 4 | 0,41 | SYNERGISM |
| 47 | 8 | 12 | 1 | 3 | 0,37 | SYNERGISM |
| 48 | 8 | 16 | 2 | 6 | 0,62 | ADDITIVE |
| 49 | 6 | 8 | 1 | 1,5 | 0,35 | SYNERGISM |
| 50 | 4 | 6 | 0,5 | 2 | 0,45 | SYNERGISM |
| 51 | 24 | 32 | 6 | 12 | 0,62 | ADDITIVE |
| 52 | 8 | 16 | 2 | 4 | 0,5 | SYNERGISM |
| 53 | 12 | 16 | 3 | 4 | 0,5 | SYNERGISM |
| 54 | 16 | 24 | 3 | 6 | 0,43 | SYNERGISM |
| 55 | 4 | 8 | 1 | 2 | 0,5 | SYNERGISM |
| 56 | 6 | 8 | 0,75 | 2 | 0,37 | SYNERGISM |
| 57 | 8 | 12 | 1 | 3 | 0,37 | SYNERGISM |
| 58 | 8 | 16 | 1 | 2 | 0,25 | SYNERGISM |
| 59 | 4 | 6 | 0,75 | 1,5 | 0,43 | SYNERGISM |
| 60 | 16 | 24 | 3 | 4 | 0,35 | SYNERGISM |
| 61 | 12 | 16 | 1,5 | 4 | 0,37 | SYNERGISM |
| 62 | 32 | 48 | 6 | 12 | 0,43 | SYNERGISM |
| 63 | 24 | 48 | 12 | 24 | 1 | ADDITIVE |
| 64 | 16 | 24 | 6 | 12 | 0,87 | ADDITIVE |
| 65 | 12 | 16 | 3 | 4 | 0,5 | SYNERGISM |
| 66 | 12 | 16 | 2 | 4 | 0,41 | SYNERGISM |
| 67 | 4 | 8 | 0,5 | 1 | 0,25 | SYNERGISM |
| 68 | 8 | 16 | 0,75 | 2 | 0,21 | SYNERGISM |
| 69 | 16 | 24 | 3 | 4 | 0,35 | SYNERGISM |
| 70 | 6 | 12 | 1,5 | 2 | 0,41 | SYNERGISM |
| 71 | 8 | 16 | 1,5 | 2 | 0,31 | SYNERGISM |
| 72 | 8 | 12 | 1 | 2 | 0,29 | SYNERGISM |
| 73 | 16 | 24 | 3 | 6 | 0,43 | SYNERGISM |
| 74 | 12 | 16 | 1,5 | 2 | 0,25 | SYNERGISM |
| 75 | 8 | 16 | 1 | 3 | 0,31 | SYNERGISM |
| 76 | 12 | 16 | 1 | 3 | 0,27 | SYNERGISM |
| 77 | 32 | 48 | 8 | 12 | 0,5 | SYNERGISM |
| 78 | 16 | 24 | 4 | 6 | 0,5 | SYNERGISM |
| 79 | 12 | 24 | 2 | 4 | 0,33 | SYNERGISM |
| 80 | 8 | 16 | 1 | 4 | 0,37 | SYNERGISM |
| 81 | 12 | 16 | 1,5 | 4 | 0,37 | SYNERGISM |
| 82 | 16 | 24 | 2 | 6 | 0,37 | SYNERGISM |
| 83 | 12 | 16 | 1 | 4 | 0,33 | SYNERGISM |
| 84 | 16 | 24 | 2 | 6 | 0,37 | SYNERGISM |
| 85 | 8 | 12 | 0,75 | 2 | 0,26 | SYNERGISM |
| 86 | 12 | 24 | 1,5 | 4 | 0,29 | SYNERGISM |
| 87 | 16 | 48 | 3 | 8 | 0,35 | SYNERGISM |
| 88 | 16 | 24 | 3 | 6 | 0,43 | SYNERGISM |
| 89 | 24 | 32 | 8 | 12 | 0,70 | ADDITIVE |
| 90 | 12 | 16 | 3 | 4 | 0,5 | SYNERGISM |
| 91 | 12 | 24 | 2 | 6 | 0,41 | SYNERGISM |
| 92 | 16 | 24 | 3 | 6 | 0,43 | SYNERGISM |
| 93 | 8 | 12 | 0,75 | 2 | 0,26 | SYNERGISM |
| 94 | 12 | 16 | 1,5 | 4 | 0,37 | SYNERGISM |
| 95 | 8 | 16 | 2 | 4 | 0,5 | SYNERGISM |
| 96 | 8 | 12 | 1,5 | 3 | 0,43 | SYNERGISM |
| 97 | 4 | 12 | 0,5 | 1,5 | 0,25 | SYNERGISM |
| 98 | 16 | 32 | 4 | 6 | 0,43 | SYNERGISM |
| 99 | 12 | 16 | 2 | 4 | 0,41 | SYNERGISM |
| 100 | 16 | 24 | 3 | 4 | 0,35 | SYNERGISM |
#: Isolate; FICI: FIC index; FOS: fosfomycin; MIC FOS/TEM: MIC of fosfomycin when combined with temocillin; MIC TEM/FOS: MIC of temocillin when combined with fosfomycin; TEM: temocillin.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



