
Submitted:
05 March 2024
Posted:
05 March 2024
You are already at the latest version
Abstract
This study addresses the urgent need for sustainable protein sources in aquaculture due to the depletion of marine resources and rising costs. Animal protein sources, particularly poultry by-product meal (PBM) and insect exuviae meal, are investigated as viable alternatives to fishmeal (FM). Research confirms the successful replacement of FM with a combination of PBM and insect exuviae meal (up to 50%) in the diet of European seabass without compromising growth, feed conversion, gut health and liver fat content. In particular, growth performance was robust with both PBM formulations, with the 25% PBM diet showing better results. Histological examinations showed good gut and liver health, contradicting the concerns of previous studies. The study emphasizes the importance of holistic analyzes that go beyond growth parameters and include histomorphological investigations. The results show that PBM in combination with insect/exuviae meal is well tolerated by seabass, which is consistent with literature reports of mitigating negative effects on gut health. A detailed analysis of the microbiota revealed a decrease in the Firmicutes/Proteobacteria ratio due to an increase in potentially pathogenic bacteria. However, the formulation containing insect exuviae partially counteracted this effect by preserving the beneficial Lactobacillus and promoting the synthesis of short-chain fatty acids (SCFA), particularly butyrate. Chitin-rich components from insect exuviae were associated with improved gut health, which was supported by increased production of SCFA, which are known for their anti-inflammatory properties. The study concludes that a combination of PBM and insect/exuviae meal can replace up to 50% of FM in the diet of seabass, supporting sustainable aquaculture practices. Despite some changes in the microbiota, the negative effects are mitigated by the addition of insect exuviae, highlighting their potential as a prebiotic to increase fish productivity and contribute to a circular economy in aquaculture.
Keywords:
insect meal
; exuviae
; poultry meal
; seabass
; gut microbiota
1. Introduction
Fishmeal (FM) is undoubtedly an excellent source of protein in aquafeeds due to its essential amino acid (EAA) profile, palatability, and other remarkable properties [1]. However, in recent years, we have observed a steady decline in the inclusion levels of FM in aquaculture feeds due to limited supply, increasing price, and ethical concerns [1,2]. Currently, FM is widely regarded as no longer a sustainable and essential component of fish feed.
Therefore, alternative, and more sustainable protein sources are needed for the development and promotion of sustainable aquaculture. Since the beginning of this century, considerable research efforts have been made to find alternative proteins, focusing mainly on plant-based ingredients such as soybean meal, corn gluten meal and rapeseed meal. Plant proteins are used extensively in feed formulations and will continue to be important raw materials in aquafeeds, although they contain several antinutritional factors (ANFs), complex indigestible carbohydrates, and low EAAs, which can have negative side effects on feed intake, digestion and absorption of nutrients, and fish health [3].
A real alternative to vegetable proteins as FM substitutes are animal by-products, which have similar properties to FM in terms of AA content, digestibility, palatability, and lack of ANFs. Since June 1, 2013, the European Commission has approved processed animal proteins from non-ruminants in aquafeed [4]. Before this date, their use was prohibited due to restrictive regulations to prevent the spread of bovine spongiform encephalopathy [5]. Among the by-products of land animals, poultry by-product meal (PBM), which consists of ground, rendered, and cleaned parts of the carcasses of slaughtered poultry including legs, necks, intestines, and undeveloped eggs, is the most economical and widely used component of aquafeed [6]. PBM is a high quality palatable and digestible protein source due to its content of EAA (except lysine and methionine), fatty acids, vitamins, and minerals [7].
To date, the suitability of PBM inclusion has been reported for several marine species [8]. PBM has successfully partially or completely replaced FM in the diet of a number of marine fish species, including gilthead seabream (Sparus aurata) [9,10,11,12], black seabass (Centropristis striata) [13], and red seabream (Pagrus major) [14]. In particular, in gilthead seabream, up to 100% of FM replacement (corresponding to 38% of inclusion) was achieved with lysine and methionine supplementation [12,15].
Optimal growth performance has even been obtained in salmonids, including rainbow trout (Oncorhynchus mykiss) and Atlantic salmon (Salmo salar), fed feeds with high PBM content [16,17,18,19,20]. However, it should be noted that differences in maximum dietary PBM inclusion rate are highly dependent on fish species, PBM quality, and overall feed formulation. Although some studies suggest that PBM can completely replace conventional protein sources, most studies recommend partial replacement of FM to maintain the nutritional balance, palatability, and digestibility of the feed [8,9,10,11,17,21,22].
Moreover, mixtures of alternative protein sources are usually preferred over a single protein source to replace FM in fish feed, especially in FM-free formulations. For example, mixtures of poultry and insect meal are being investigated as alternatives to FM in aquafeeds. Like PBM, insect meal is a promising alternative to FM in aquafeed as it has several advantages. Insects have a high protein (34-74% DM) and lipid (10-30% DM) content, balanced EAA, and a good content of vitamins (B12) and minerals (iron and zinc); they also contain several bioactive compounds such as chitin, fatty acids, and antimicrobial peptides [23].
The use of insects in animal feed is strictly regulated by the European Union (EU). Regulation no 2001/999 (Annex IV), amended by Regulation 2017/893 (Annex X), authorises the use of seven insect species in feed for aquaculture, namely the common housefly (Musca domestica), the black soldier fly (Hermetia illucens), lesser mealworm (Alphitobius diaperinus), mealworm (Tenebrio molitor), house cricket (Acheta domesticus), field cricket (Gryllus assimilis), and banded cricket (Gryllodes sigillatus). As of November 2021 (Regulation 2021/1925), the list was extended to eight authorized species with the inclusion of the silkworm (Bombyx mori). Among them, the black soldier fly and the mealworm are the most commonly used insect species in aquafeed [24,25].
Recent studies have shown that an effective replacement of FM in aquafeed can be achieved by combining black soldier fly (BSF) larval meal with PBM. In rainbow trout fed a plant-based diet without FM, a high proportion of plant protein-rich components in the diet with both BSF and PBM resulted in better growth and gut/liver health outcomes than BSF and PBM alone [20]. Similarly, BSF and PBM successfully replaced plant proteins in a diet without FM in gilthead seabream, improving nutrient uptake and promoting gut health [26].
In addition, the combination of BSF and PBM in particular restored the gut microbiota of fish negatively affected by a plant-based diet by improving the richness of bacterial species and the abundance of beneficial bacteria [27,28].
The prebiotic effect of chitin, the indigestible polysaccharide that makes up the exoskeleton of insects, and insect meal containing chitin has been widely demonstrated in fish [24,29]. The addition of insect meal usually increases the amount of lactic acid bacteria (LAB) and the genus Bacillus, which are commonly used as probiotics in aquaculture. Finally, the gut microbiota of fish is known to be very sensitive to dietary manipulations [30], and since proteins are the most important nutrient, they have a major impact on the composition of the gut microbiota.
Accordingly, in the present study we tested the effect of partial replacement of FM by PBM alone or in combination with insect meal or exuviae meal, a chitin source from insect exoskeletons, in European seabass. A multidisciplinary approach was used in which growth performance, gut and liver morphology, gut microbiota and gut volatile short chain fatty acid production, were evaluated.
2. Materials and Methods
2.1. Ethics Approval
The handling of animals and all procedures complied with the European Union Council Directive 2010/63 on the use of experimental animals. The protocol was approved by the Ethics Committee for Animal Welfare and Use of Animals of the University of Insubria and by the Italian Ministry of Health (No. 285/2020-PR).
2.2. Diets
Three isoproteic and isolipidic diets were formulated: a control diet (A) containing 20% FM, and two test diets in which 50% of the FM was replaced by 20% (B) or 25% (C) of PBM. In addition, 5% insect meal (Mutatec, France) and 0.5% exuviae meal were added to feed B and C, respectively. The complete formulation and the proximate composition of the three feed types produced by “Leocata Mangimi” in Modica (Ragusa), Italy, are listed in Table 1 and Table 2.
2.3. Feeding Trial and Sampling
The feeding experiment was carried out in a recirculating aquaculture system (RAS) at the Department of Biotechnology and Life Sciences of the University of Insubria (Varese, Italy). Two hundred and ten juvenile European seabass (D. labrax) (mean initial body weight 52 ± 0.8 g), purchased from Società Agricola CIVITA ITTICA S.r.l. (Verona, Italy), were randomly distributed in six circular fiberglass tanks with a capacity of 700 l (35 fish/tank). The pH, temperature, and dissolved oxygen (DO) of the water were strictly controlled throughout the experiment. Water temperature was 20 ± 1.5 °C, salinity was 22 g l−1, pH was 7.5-8.0; total ammonia nitrogen was ≤ 0.1 mg l−1, and DO saturation was maintained above 85%. After a one-week acclimatization period, the fish were fed once daily (6 days per week) with three experimental diets (2 tanks/feed) for 15 weeks. The experimental groups fed diets A, B, or C were named CTRL, PM20, and PM25, respectively. The fish were individually weighed and measured in length at the beginning of the feeding trial (t0) and then after 4 weeks (t1), 10 weeks (t2), and at the end of 15 weeks (t3). The feeding rate was adjusted according to biomass and ranged between 1.5-2.5% of body weight. The weight of ingested and uneaten feed was recorded daily for each tank. Mortality was also monitored. These data were used as the basis for calculating feed conversion ratio (FCR = dry feed intake/wet weight gain) and specific growth rate [SGR (%/day) = 100 x [ln (final body weight) - ln (initial body weight)]/day], for each dietary fish group.
At the end of the 16-week experiment, 4 fish per tank (8 fish/diet) were euthanized with an overdose of tricaine methanesulfonate (MS-222, 400 mg l-1, Sigma-Aldrich). Liver, proximal, and distal intestine were removed from each fish and immediately fixed in a 10% solution of neutral buffered formalin (NBF) for histological analysis. Samples were taken from an additional four fish per tank (8 fish/diet) and the entire intestine (excluding pyloric ceca) was aseptically removed from each fish. The digesta and intestinal mucosa were collected and mixed together in a sterile tube with 800 μl Xpedition™ Lysis/Stabilization Solution (Zymo Research, Irvine, CA, USA) and stored at 4°C until metabarcoding analysis [31]. For the quantification of volatile fatty acids, 18 additional faecal samples were collected from each experimental group (9 fish/tank), pooled in a centrifuge tube (3 pools/tank), and stored at -80°C until analysis.
2.4. Histological Analysis
NBF-fixed liver and intestinal samples were embedded in paraffin and cut into 5μm sections using a microtome (Leica RM2245). To observe the tissue structure, the slides were stained with hematoxylin and eosin (H&E) and examined under a light microscope (Zeiss Axiophot microscope) using a CMOS Discovery C30 digital camera. The acquired images were processed using the Fiji software (open-source Java-based image processing programme). For intestinal morphology, villus height (ViH), villus width (ViW), lamina propria width (LPW), and submucosal layer thickness (SMT) were measured according to Escaffre et al. [32]. For the liver, a semi-quantitative assessment approach based on a grading score (1 = not observed/low, 2 = moderate, and 3 = severe) was used [33,34,35]. In particular, hepatocyte vacuolization (HV), nuclear displacement (ND), cellular hypertrophy (CH), and irregular nuclear shapes (NS) were considered as histological features in the liver.
2.5. Analysis of Short Chain Fatty Acids in Faecal Samples
The qualitative and quantitative determination of SCFAs (acetate, propionate, iso-butyrate, and butyrate) was performed according to the modified extraction method of Chlebicz-Wójcik and Śliżewska [36]. The detailed protocol of SCFA extraction was described by Rimoldi et al. [37]. The extracts were analysed using an HPLC-UV-VIS system (Shimadzu, Milan, Italy) equipped with two LC -20AD pumps, a CBM-Alite controller, a DGU-20A5 degasser, and an automatic injector. Samples were separated on a 150 mm × 4.6 mm I.D., 2.7 µm particle Ascentis Express 90A C18 column (Merck KGaA, Darmstadt, Germany). The flow rate of the mobile phase (0.005 M H2SO4) was 0.6 mL/min, and the injection volume was 10 µL. The column temperature was maintained at 60°C and the UV-VIS absorbance was measured at a wavelength of 210 nm. The raw data were processed using the LC software (Shimadzu, Milan, Italy). The peaks were identified based on the retention time of the certified standards. Quantitative analyses were performed using the calibration curve method in a range of different concentrations. The results were expressed in mmol/l.
2.6. Bacterial DNA Extraction, Multiplex-Amplicon Library Preparation, and Sequencing
Bacterial DNA was extracted from 300 mg of intestinal material (faeces + mucosa) and 200 mg of each feed in triplicate. For DNA extraction, the DNeasy® PowerSoil® Pro kit (Qiagen, Italy) was used according to the manufacturer’s instructions, with an additional mechanical lysis step using a TissueLyser II (Qiagen, Italy) [38]. The 16S V4 library was prepared by the next generation sequencing (NGS) service GALSEQ srl (Milan, Italy) and sequenced on the NovaSeq 6000 System – Illumina, using a paired-end 2 × 150 bp sequencing strategy and a cluster density of 300K/sample. A detailed description of library preparation and sequencing has already been reported [18,39].
2.7. NGS Raw Data Analysis
The QIIME 2TM pipeline (v. 2020.2) was used to process raw amplicon data [40]. Taxonomy was assigned to amplicon sequence variants (ASVs) down to genus level using the SILVA database (https://www.arb-silva.de/). The entire data preparation workflow has been previously described in detail [41]. It included pre-processing steps, taxonomy classification, and calculation of alpha and beta diversity based on weighted and unweighted UniFrac distances [42,43].
2.8. Predicted Functional Pathway Analysis of Gut Microbiome
PICRUSt (Phylogenetic Investigation of Communities by Reconstruction of Unobserved States) was used to assess the functional potential of microbial communities [44]. The predicted function of the metagenome was based on the analysis of KEGG pathways. The extended error bar graphs showing the significantly different abundance of KEGG pathways between control and PM20 and PM25 groups were generated from the PICRUSt output files using the Statistical Analysis of Metagenomic Profiles (STAMP) software package and two-sided Welch’s t-test [45].
2.9. Statistics
All data were tested for normality and homoscedasticity using the Shapiro–Wilk test and the Levene’s test, respectively. For statistical comparisons, the two-way analysis of variance or the Kruskal-Wallis test was used if the data were not normally distributed. Significant differences in beta diversity were tested using permutational multivariate analysis of variance (PERMANOVA). All tests were performed using the Past4 v. 4.02 software with significance set at p < 0.05 [46]. Differences in the abundance of bacterial taxa between samples were tested with the two-sided Welch’s t-test using the STAMP software package.
3. Results
3.1. Growth Performance
The data on growth performance and feed conversion are shown in Table 3. During the entire duration of the feeding trial (88 days), the mortality rate was less than 2%. All fish grew efficiently and doubled their weight. However, a significantly lower final body weight (FBW) was observed in fish fed diet PM20 with respect to the controls. There were no significant differences between the diet groups in specific growth rate (SGR) and feed conversion ratio (FCR).
3.2. Gut and Liver Morphology
A detailed examination of liver and intestinal histopathology was performed to evaluate the effects of PM in the diet of European seabass (as described in Table 4 and Table 5). Standard histological analysis of proximal and distal intestinal cross-sections of the PM20 and PM25 fish groups showed a well-organized and preserved tissue structure with no obvious signs of damage or inflammation compared to specimens from control animals (Figure 1, panels A–F). Morphometric analysis of the intestinal cross-sections also showed no morphological changes in response to diet in the structure of the mucosa and submucosa layers, the thickening of the lamina propria, and the height and width of the villi in both the distal and proximal intestines (Table 4).
Histological sections of the liver are shown in Figure 2 (panels A-F). The liver tissue samples yielded a total score of six for the control group, five for the PM20 and 5.67 for the PM25 group (Table 5). These values define a normal or slightly altered morphology. Accordingly, all samples showed moderate lipid infiltration, which did not affect the size and shape of the hepatocytes. Most of the nuclei were in a peripheral position, but did not alter the shape and size of the hepatocytes and thus the physiological morphology of the organ. No signs of hepatocyte ballooning, vacuolar degeneration, capillary hyperemia, or vasodilatation were observed in the different experimental groups.
3.3. Volatile SCFAs in Faecal Samples
The SCFAs in the feces of fish fed three experimental diets were quantified by HPLC. The concentrations of selected SCFAs (acetate, propionate, and butyrate) found in our samples are shown in Table 6. The highest amount of propionate (7.41 mmol/L) and butyrate (3.03 mmol/L) was found in the gut of fish of group PM25, fed with insect exuviae. In contrast, fish of the PM20 group fed with diet B had a higher concentration of acetate in their feces compared to the other two feeding groups.
3.4. Sequencing Efficiency
Of the bacterial DNA extracted from the gut samples, only twenty were efficiently amplified to obtain a V4 amplicon library, seven from fish groups CTRL and PM25 and six from PM20. In contrast, all nine DNA samples from the feeds were amplified correctly and yielded the expected amplicon size. All twenty-nine samples were successfully sequenced and yielded 2,393,031 high quality reads, of which 2,021,977 were from gut samples and 371,054 from feed pellets (Supplementary data file S1). Good’s coverage values were above 99% for all samples, indicating that the sequencing depth of all datasets in this study was sufficient to reveal the bacterial communities in the gut and feed. Based on the rarefaction curves, the sequencing depth for the calculation of alpha diversity indices was set at 55,556 reads.
All raw data (Fastq) were submitted to the public database of European Nucleotide Archive (EBI ENA) under the access code PRJEB70800.
3.5. Feed Microbial Profiles
The microbiota profile of three feeds was outlined at the level of phylum, class, order, family, and genus. Considering only the most representative taxa (relative abundance ≥ 1%), the microbial community of the feeds was composed of 4 phyla (Figure 3A), 5 classes, 17 orders, 59 families (Figure 3B), and 73 genera. The result of alpha diversity analysis performed on the feed microbiota data showed the highest species richness (observed OTUs) and biodiversity (Shannon’s index) in feed A, and a lower Chao 1 index value in feed C compared to B (Table 7).
Pairwise comparisons of microbial communities from different diets using the two-sided Welch’s t-test revealed that 37 and 16 genera differed significantly between control A and diets B and C, respectively. Most of these genera belonged to the phylum Firmicutes and were more abundant in the experimental feed pellets (Table 8 and Table 9). In contrast, the relative abundances of 52 bacterial genera were significantly different between two experimental feed B and C (Table 10). Twenty-two of them were assigned to the Firmicutes phylum. In particular, the probiotic genera Bacillus and Lactobacillus were associated with diet C, which contained 25% PBM and exuviae meal in its formulation.
3.6. Gut Microbiota Profiles
The total intestinal microbiota of our samples comprised 5 phyla (Figure 4A), 5 classes, 13 orders, 46 families (Figure 4B), and 49 genera.
Analysis of the alpha diversity of the intestinal samples (Table 11) revealed significant differences only for the Shannon biodiversity index, which was lower than in fish fed diet B than in controls (CTRL= 6.20 ± 1.06; PM20= 4.69 ± 0.65; PM25= 5.18 ± 1.23).
Analysis of beta diversity of gut microbial communities based on phylogenetic UniFrac distances revealed both qualitative (unweighted UniFrac) and quantitative (weighted UniFrac) differences as a function of diet. As shown in the PCoA diagrams in Figure 5A and B, the gut samples formed separate groups from the feed samples. In particular, the PCoA diagram based on the weighted UniFrac distances showed that the gut samples of fish fed the experimental diets PM20 and PM25 and the control samples were clustered separately (Figure 5A). The results of the PCoA analysis were statistically validated by both the ANOSIM test and the PERMANOVA test (Table 12).
3.7. Modulating Effect of Diet on the Composition of Gut Microbiota
To determine any differences in the gut microbiota, pairwise comparisons were made between the feeding groups. The relative abundances of bacterial genera were compared using Welch’s t-test and the results are shown in Table 13 and Table 14. Firmicutes and Proteobacteria are the dominant phyla in all experimental groups (Figure 4A). These two phyla were significantly affected by the replacement of FM with PBM. In particular, a decrease in Firmicutes (8-10%) combined with an increase in the proportion of Proteobacteria (80-83%) was observed in fish fed with experimental diets B and C compared to controls (Firmicutes: 76%; Proteobacteria: 18%). At the genus level, a decrease in Lactobacillus was observed in the gut of PM20 (Table 13), but not in the PM25 group (Table 14). Compared to CTRL, both PM20 and PM25 fish had a lower relative abundance of the genera Clostridium sensu stricto, belonging to the Clostridiaceae family, and Cetobacterium (Table 13 and Table 14). In addition, the experimental diets B and C had a bactericidal effect on the genus Staphylococcus. In contrast, the PM20 and PM25 samples showed a marked increase in the genera Vibrio and Photobacterium, both of which belong to the Vibrionaceae family (Table 13 and Table 14). When comparing PM20 and PM25 fish, their gut microbiota differed only in the genus Paenibacillus, which was more abundant in the gut of PM20 fish (5%) than in the gut of PM25 fish (0.06%).
3.8. Predictive Functional Analysis of Gut Microbiota Communities
The PICRUSt predictions of the functional composition of the gut microbiota based on the KEGG database showed large differences in the abundance of KEGG metabolic pathways between the control and treatment groups. In particular, 20 and 19 metabolic pathways differed between the CTRL and PM20 and PM25 samples, respectively (Figure 6 and Figure 7). The gut microbiota of fish fed PBM diets showed increased abundance of metabolic pathways mainly involved in biofilm formation, peptidoglycan and lipopolysaccharide biosynthesis, structural proteins, bacterial secretion system and unsaturated fatty acids biosynthesis. In contrast, cell growth, nucleotide metabolism and Staphylococcus aureus infections decreased in the PM20 and PM25 groups. No significant difference was found between the functional profile of PM20 and PM25.
4. Discussion
The drive to find alternative and more sustainable protein sources to replace FM stems from the need to alleviate the pressure on global marine resource depletion and also to contain the cost of aquaculture production, as protein is the most expensive nutrient in fish diets, accounting for 30-50% of the total cost.
Various plant protein sources have traditionally been used to partially or completely replace FM in aquafeeds. However, nowadays animal protein sources are considered the best alternative to FM due to their higher protein and lipid content, better amino acid profile, and palatability [47,48].
The results of this study seem to go in this direction, confirming that PBM together with insect or exuviae meal successfully meet the requirements for adequate growth in seabass. This result confirms what has already been reported in the literature, according to which the optimal substitution rate of FM by PBM is between 25% and 50% for most marine carnivorous animals [8]. Indeed, both formulations containing PBM were well accepted and at the end of the feeding trial, the fish doubled their weight, regardless of the diet.
However, only minor differences were found between the control group and the experimental feeding groups PM20 and PM25. The growth performance determined from the final weight showed that the fish group fed with the 20% PBM diet performed worse than the group fed with the control diet, resulting in a lower final weight compared to the control, but did not differ from the PM25 group. In fact, in marine fish, such as seabream (Sparus aurata) and red seabream (Pagrus major), there is evidence that even the complete replacement of FM with PBM had no negative effect on the growth parameters and productivity of the fish [10,14,15].
The inclusion of PBM in the diet of freshwater fish has been shown to be successful as well. In particular, the present study is consistent with our previous results in rainbow trout. The growth data showed that trout fed a diet rich in PBM (55-70%) grew as well as fish fed a control diet rich in FM (37.3%) and free of PBM [18].
The formulation of optimal diets for farmed fish species requires the application of different types of analyzes to verify their effects on the health status of the specimens, which cannot be limited to growth performance and feed efficiency alone. Histomorphological examination is a good biomarker for the assessment of fish welfare, especially in gut and liver histomorphology [49]. No potential changes related to intestinal inflammation or hepatic lipid accumulation in response to FM replacement were detected in the histological examinations, suggesting that PBM in combination with insect and exuviae meal was well tolerated by seabass. In agreement with our results, Pleić and colleagues [28] showed that partial replacement of plant protein diet with insect and PBM could even mitigate the negative effects of plant proteins on the proximal and distal gut of seabass by significantly improving all gut morphometric parameters, the degree of vacuolization and cellular infiltration. The health and integrity of the intestinal epithelium are critical to nutrient absorption and fish health, as damage to the gut can lead to immune dysfunction, reduced disease resistance, loss of appetite and slow growth.
In addition to the gut, the liver is also considered a valuable marker for nutritional pathologies, as it plays a key role in energy metabolism and storage, as well as in immune defense and detoxification [50,51]. As for the present study, the histological results indicate a favorable liver health status, with a moderate accumulation of lipids in hepatocytes in all fish, regardless of diet. In contrast, Donadelli et al. [52] recently observed a significant accumulation of hepatocyte lipids associated with marked histological changes indicative of an incipient steatotic state in seabass fed a FM-free diet in which 40% of the plant protein was replaced with insect or poultry by-product. These changes appeared to be slightly attenuated when insect meal and PBM were combined to partially replace plant proteins in the diet. The data available in the literature indicate species-specific differences in the responses to alternative protein-rich feed components. For example, the inclusion of Hermetia illucens and PBM in diets containing no FM resulted in improved gut and liver health in gilthead seabream and rainbow trout [20,26,52].
Interestingly, a higher amount of SCFAs was found in the fecal samples of fish fed experimental diets containing insect-derived raw material compared to controls. Insect meal and especially exuviae are considered valuable sources of chitin. Chitin is an insoluble dietary fiber consisting of β-1,4-poly-N-acetyl-D-glucosamine and can be used as a substrate for bacterial fermentations leading to the production of acetate, propionate, and butyrate as the main end products with positive effects on gut health [24,53]. Indeed, chitin and its deacetylated derivative chitosan are bioactive compounds which can positively influence the health and performance of farmed fish. In the present study, the highest amount of propionate and butyrate was found in the gut of seabass fed with exuviae meal. Accordingly, trout fed with pupal exuviae meal had the highest content of SCFAs, especially butyrate, in their feces [29]. Butyrate is well known to have anti-inflammatory properties and to promote fish intestinal health, barrier function, and mucosal immunity in fish. Therefore, its increased production in the fish gut should be considered a desirable effect [54,55,56].
As a prebiotic, chitin may also increase the biodiversity of the gut microbiota by promoting the proliferation of beneficial chitin-degrading bacteria, such as Bacillus and Paenibacillus, which have recently been isolated from the gastrointestinal tract of European seabass fed chitin-enriched diets [57]. It is known that replacing FM with insect meal affects the biodiversity and number of gut bacteria. The partial replacement of FM with at least 10% insect meal had an important effect in modulating the transient gut microbial communities by increasing both butyrate-producing bacteria and beneficial lactic acid bacteria [37,58,59,60,61]. Similarly, the ingestion of H. illucens exuviae meal led to an enrichment of the gut microbiota with the families Bacillaceae, Staphylococcaceae, Paenibacillaceae, and Brevibacteriaceae in seabass and rainbow trout [29,58]. Unlike previous studies, the proportion of insect or exuviae meal utilized in the present study was not sufficient to promote the proliferation of beneficial bacteria. In fact, the gut microbiota of PM20 and PM25 seabass was characterized by an increase in the Proteobacteria/Fimicutes ratio compared to the FM diet-fed controls. A similar increase in the Proteobacteria fraction, mainly represented by Gammaproteobacteria, was previously reported in trout fed diets containing a high proportion of alternative terrestrial animal proteins (>50%), mainly represented by PBM [18]. Accordingly, the trout gut microbiota was characterized by a high abundance of bacterial genera belonging to the class of Gammaproteobacteria, such as Vibrio, Pasteurella, and Proteus. Also, at the genus level, PM20 and PM25 fish showed an enrichment of Vibrio and Photobacterium genera in their gut, which are normally considered potential pathogens for fish. Similarly, we found an increase of the genus Photobacterium in the gut microbiota of trout fed a diet containing 20% of head shrimp meal, another chitin-rich ingredient [29]. However, it is also true that this genus includes several chitinase-producing bacterial species [62,63]. In contrast, it was recently reported that seabass fed a plant-based diet supplemented with a combination of 10% H. illucens meal and 30% PBM showed increased abundance of the phylum Firmicutes, particularly the beneficial genera Lactobacillus and Bacillus in their gut [28].
In relation to the present study, the genus Lactobacillus decreased significantly in the gut of PM20 seabass, but not in PM25 compared to controls. In contrast to the exuviae meal, the amount of insect meal was not sufficient to attenuate the negative effects of PBM on the abundance of lactic acid bacteria. Similarly, in gilthead seabream, a 10% content of H. illucens meal was not sufficient to increase the abundance of lactic acid bacteria in the gut [37,59].
The comparison of microbial profiles in the gut of PM20 and PM25 fish showed a positive association of the genus Paenibacillus with insect meal. This result confirmed what was previously reported in rainbow trout and gilthead seabream in response to dietary H. illucens meal ingestion [37,39,59]. In contrast, Rangel et al. [58] found that consumption of diets enriched with H. illucens exuviae, but not insect meal, led to an increase in the genus Paenibacillus in the gut of European seabass. This genus is of interest because it is considered a good probiotic candidate. In fact, Paenibacillus shares several characteristics with members of the genus Bacillus. It can produce antimicrobial and volatile organic compounds and degrade non-starch polysaccharides, and last, but not least, the bacteria of this genus have chitinolytic activity [57,58,64,65,66]. The presence of chitinolytic bacteria is particularly important when feeding insect-derived ingredients to fish, as bacterial chitinases help to improve the digestibility of the feed [53]. In rainbow trout, on the other hand, the addition of 1.6% pupal exuviae meal of H. illucens to a diet containing 20% FM resulted in an enrichment of various bacteria genera, such as Corynebacterium, Bacillus, Facklamia, and Brevibacterium. These divergent results suggest that the relationship between dietary components and the gut microbiota are complex and species-specific.
The predictive functional analysis PICRUSt showed large differences between the controls and the PBM-fed experimental groups. Compared to controls, fish fed PBM diets were associated with biofilm formation, peptidoglycan and lipopolysaccharide biosynthesis, structural proteins, bacterial secretion system, and unsaturated fatty acid biosynthesis. Much more interesting was the reduction of signaling pathways related to Staphylococcus aureus infection in PM20 and PM25 fish. This result is consistent with the lower amount of the genus Staphyloccoccus observed in these samples.
5. Conclusions
Overall, this work has shown that in practical feeds for European seabass, formulated to current industry standards, FM can sometimes be replaced by up to 50% by a combination of PBM and insect or exuviae meal without compromising growth performance, feed conversion, gut health, and liver lipid content. The only negative effect of PBM was a decrease in the Firmicutes/Proteobacteria ratio at the gut microbiota level due to an increase in potentially pathogenic bacteria belonging to the Gammaproteobacteria class, such as Vibrio and Photobacterium. However, of the two experimental formulations, the diet containing insect exuviae meal partially mitigated this negative disadvantage by preserving the amount of the beneficial Lactobacillus genus comparable to controls fed FM diet and promoting the synthesis of SCFAs, especially butyrate. These data confirm our previous findings that insect exuviae are a valid prebiotic candidate to contribute to more sustainable aquaculture practices by increasing fish productivity through efficient utilization of organic waste thus contributing to a circular economy system.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, supplementary DATA file S1.
Author Contributions
All authors contributed to the study conception and design. Material preparation, data collection and analysis were performed by S.R., A.D, R.A., I.H., V.K, G.T, and B.C.. The first draft of the manuscript was written by S.R., G.T., and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Funding
Research partly supported by the 1. project “BIO = C = O,” PON-MISE I&C 2014–2020 FESR, ASSE I—Innovazione e Azione, Azione 1.1.3. D.M. 05/03/2018, project codex N. F/200078/01–03/X45, CUP B41B20000280005; 2. I-FISH. Protocollo nr: 414352—del 07/12/2023—AOO_IAI—AOO_Incentivi Fondo per la Crescita Sostenibile—Accordi per l’innovazione di cui al D.M. 31 Dicembre 2021 e D.D. 14 Novembre 2022. Impresa: VRM S.R.L. (C.F. 02779360243)—Capofila.
Data Availability Statement
All raw sequencing data were submitted to the European Nucleotide Archive (EBI ENA) public database under accession code PRJEB70800.We encourage all authors of articles published in MDPI journals to share their research data.
Acknowledgments
The authors thank Dr Micaela Antonini for assistance with the animal care and feeding. IH and VK are PhD students in the “Dottorato in Scienze della Vita e Biotecnologie” at the “Università degli Studi dell’Insubria” in Varese, Italy.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Tacon, A.G.J.; Metian, M. Fishing for aquaculture: non-food use of small pelagic forage fish-a global perspective. Rev. Fish. Sci. 2009, 17, 305–317. [CrossRef]
- Oliva-Teles, A.; Enes, P.; Peres, H. Replacing fishmeal and fish oil in industrial aquafeeds for carnivorous fish. Feed Feed. Pract. Aquac. 2015, 203–233. [CrossRef]
- Daniel, N. A review on replacing fish meal in aqua feeds using plant protein sources. Int. J. Fish. Aquat. Stud. 2018, 6, 164–179.
- European Commission Commission Regulation (EU) No 56/2013 of 16 January 2013 Amending Annexes I and IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council Laying down Rules for the Prevention, Control and Eradication of Certain Transmissible Spongifo. Off. J. Eur. Union 2013, 21, 3–16.
- European Commission Commission Regulation (EC) No 999/2001 of the European Parliament and of the Council of 22 May 2001 Laying down Rules for the Prevention, Control and Eradication of Certain Transmissible Spongiform Encephalopathies. Off. J. Eur. Union 2001, L147.
- Yu, Y. Replacement of fish meal with poultry by-product meal and hydrolyzed feather meal in feeds for finfish. In Alternative Protein Sources in Aquaculture Diets; CRC Press, 2008; pp. 51–93.
- Cheng, Z.; Hardy, R.W. Apparent digestibility coefficients of nutrients and nutritional value of poultry by-product meals for rainbow trout Oncorhynchus mykiss measured in vivo using settlement. J. World Aquac. Soc. 2002, 33, 458–465. [CrossRef]
- Galkanda-Arachchige, H.S.C.; Wilson, A.E.; Davis, D.A. Success of fishmeal replacement through poultry by-product meal in aquaculture feed formulations: a meta-analysis. Rev. Aquac. 2019. [CrossRef]
- Fontinha, F.; Magalhães, R.; Moutinho, S.; Santos, R.; Campos, P.; Serra, C.R.; Aires, T.; Oliva-Teles, A.; Peres, H. Effect of dietary poultry meal and oil on growth, digestive capacity, and gut microbiota of gilthead seabream (Sparus aurata) juveniles. Aquaculture 2021, 530, 735879. [CrossRef]
- Karapanagiotidis, I.T.; Psofakis, P.; Mente, E.; Malandrakis, E.; Golomazou, E. Effect of fishmeal replacement by poultry by-product meal on growth performance, proximate composition, digestive enzyme activity, haematological parameters and gene expression of gilthead seabream (Sparus aurata). Aquac. Nutr. 2019, 25, 3–14. [CrossRef]
- Davies, S.J.; Laporte, J.; Gouveia, A.; Salim, H.S.; Woodgate, S.M.; Hassaan, M.S.; El-Haroun, E.R. Validation of processed animal proteins (mono-PAPS) in experimental diets for juvenile gilthead sea bream (Sparus aurata L.) as primary fish meal replacers within a European perspective. Aquac. Nutr. 2019, 25, 225–238. [CrossRef]
- Psofakis, P.; Meziti, A.; Berillis, P.; Mente, E.; Kormas, K.A.; Karapanagiotidis, I.T. Effects of dietary fishmeal replacement by poultry by-product meal and hydrolyzed feather meal on liver and intestinal histomorphology and on intestinal microbiota of gilthead seabream (Sparus aurata). Appl. Sci. 2021, 11. [CrossRef]
- Dawson, M.R.; Alam, M.S.; Watanabe, W.O.; Carroll, P.M.; Seaton, P.J. Evaluation of poultry by-product meal as an alternative to fish meal in the diet of juvenile black sea bass reared in a recirculating aquaculture system. N. Am. J. Aquac. 2018, 80, 74–87. [CrossRef]
- Takakuwa, F.; Murashita, K.; Noguchi, Y.; Inui, T.; Watanabe, K.; Sugiyama, S.; Yamada, S.; Biswas, A.; Tanaka, H. Effects of long-term feeding of fishmeal-free diet on growth parameters, bile acid status, and bile acid-related gene expression of yearling red sea bream Pagrus major (Temminck & Schlegel, 1843). Aquaculture 2023, 570, 739444. [CrossRef]
- Sabbagh, M.; Schiavone, R.; Brizzi, G.; Sicuro, B.; Zilli, L.; Vilella, S. Poultry by-product meal as an alternative to fish meal in the juvenile gilthead seabream (Sparus aurata) diet. Aquaculture 2019, 511, 734220. [CrossRef]
- Burr, G.S.; Wolters, W.R.; Barrows, F.T.; Hardy, R.W. Replacing fishmeal with blends of alternative proteins on growth performance of rainbow trout (Oncorhynchus mykiss), and early or late stage juvenile Atlantic salmon (Salmo salar). Aquaculture 2012, 334–337, 110–116. [CrossRef]
- Barreto-Curiel, F.; Parés-Sierra, G.; Correa-Reyes, G.; Durazo-Beltrán, E.; Viana, M.T. Total and partial fishmeal substitution by poultry by-product meal (petfood grade) and enrichment with acid fish silage in aquafeeds for juveniles of rainbow trout Oncorhynchus mykiss. Lat. Am. J. Aquat. Res. 2016, 44, 327–335. [CrossRef]
- Rimoldi, S.; Terova, G.; Ascione, C.; Giannico, R.; Brambilla, F. Next generation sequencing for gut microbiome characterization in rainbow trout (Oncorhynchus mykiss) fed animal by-product meals as an alternative to fishmeal protein sources. PLoS One 2018, 13, 1–29. [CrossRef]
- Hatlen, B.; Jakobsen, J. V.; Crampton, V.; Alm, M.; Langmyhr, E.; Espe, M.; Hevrøy, E.M.; Torstensen, B.E.; Liland, N.; Waagbø, R. Growth, Feed utilization and endocrine responses in Atlantic salmon (Salmo salar) fed diets added poultry by-product meal and blood meal in combination with poultry oil. Aquac. Nutr. 2015, 21, 714–725. [CrossRef]
- Randazzo, B.; Zarantoniello, M.; Gioacchini, G.; Cardinaletti, G.; Belloni, A.; Giorgini, E.; Faccenda, F.; Cerri, R.; Tibaldi, E.; Olivotto, I. Physiological response of rainbow trout (Oncorhynchus mykiss) to graded levels of hermetia illucens or poultry by-product meals as single or combined substitute ingredients to dietary plant proteins. Aquaculture 2021, 538, 736550. [CrossRef]
- Wang, X.; Luo, H.; Zheng, Y.; Wang, D.; Wang, Y.; Zhang, W.; Chen, Z.; Chen, X.; Shao, J. Effects of poultry by-product meal replacing fish meal on growth performance, feed utilization, intestinal morphology and microbiota communities in juvenile large yellow croaker (Larimichthys crocea). Aquac. Reports 2023, 30. [CrossRef]
- Yu, H.; Li, M.; Yu, L.; Ma, X.; Wang, S.; Yuan, Z.; Li, L. Partial replacement of fishmeal with poultry by-product meal in diets for coho salmon (Oncorhynchus kisutch) post-smolts. Animals 2023, 13, 1–14. [CrossRef]
- Gasco, L.; Biancarosa, I.; Liland, N.S. From Waste to Feed: A review of recent knowledge on insects as producers of protein and fat for animal feeds. Curr. Opin. Green Sustain. Chem. 2020, 23, 67–79. [CrossRef]
- Hasan, I.; Rimoldi, S.; Saroglia, G.; Terova, G. Sustainable fish feeds with insects and probiotics positively affect freshwater and marine fish gut microbiota. Anim. 2023, 13, 1633. [CrossRef]
- Maulu, S.; Langi, S.; Hasimuna, O.J.; Missinhoun, D.; Munganga, B.P.; Hampuwo, B.M.; Gabriel, N.N.; Elsabagh, M.; Van Doan, H.; Abdul Kari, Z.; et al. Recent advances in the utilization of insects as an ingredient in aquafeeds: a review. Anim. Nutr. 2022, 11, 334–349. [CrossRef]
- Randazzo, B.; Zarantoniello, M.; Cardinaletti, G.; Cerri, R.; Giorgini, E.; Belloni, A.; Contò, M.; Tibaldi, E.; Olivotto, I. Hermetia illucens and poultry by-product meals as alternatives to plant protein sources in gilthead seabream (Sparus aurata) diet: a multidisciplinary study on fish gut status. Animals 2021, 11, 1–22. [CrossRef]
- Gaudioso, G.; Marzorati, G.; Faccenda, F.; Weil, T.; Lunelli, F.; Cardinaletti, G.; Marino, G.; Olivotto, I.; Parisi, G.; Tibaldi, E.; et al. Processed animal proteins from insect and poultry by-products in a fish meal-free diet for rainbow trout: impact on intestinal microbiota and inflammatory markers. Int. J. Mol. Sci. 2021, 22. [CrossRef]
- Pleić, I.L.; Bušelić, I.; Messina, M.; Hrabar, J.; Žuvić, L.; Talijančić, I.; Žužul, I.; Pavelin, T.; Anđelić, I.; Pleadin, J.; et al. A plant-based diet supplemented with hermetia illucens alone or in combination with poultry by-product meal: one step closer to sustainable aquafeeds for European seabass. J. Anim. Sci. Biotechnol. 2022, 13, 1–22. [CrossRef]
- Rimoldi, S.; Ceccotti, C.; Brambilla, F.; Faccenda, F.; Antonini, M.; Terova, G. Potential of shrimp waste meal and insect exuviae as sustainable sources of chitin for fish feeds. Aquaculture 2023, 567, 739256. [CrossRef]
- Ringø, E.; Zhou, Z.; Vecino, J.L.G.; Wadsworth, S.; Romero, J.; Krogdahl; Olsen, R.E.; Dimitroglou, A.; Foey, A.; Davies, S.; et al. Effect of dietary components on the gut microbiota of aquatic animals. A never-ending story? Aquac. Nutr. 2016, 22, 219–282. [CrossRef]
- Rimoldi, S.; Gini, E.; Koch, J.F.A.; Iannini, F.; Brambilla, F.; Terova, G. Effects of hydrolyzed fish protein and autolyzed yeast as substitutes of fishmeal in the gilthead sea bream (Sparus aurata) diet, on fish intestinal microbiome. BMC Vet. Res. 2020 161 2020, 16, 1–13. [CrossRef]
- Escaffre, A.M.; Kaushik, S.; Mambrini, M. Morphometric evaluation of changes in the digestive tract of rainbow trout (Oncorhynchus mykiss) due to fish meal replacement with soy protein concentrate. Aquaculture 2007, 273, 127–138. [CrossRef]
- Caballero, M.J.; Izquierdo, M.S.; Kjørsvik, E.; Fernández, A.J.; Rosenlund, G. Histological alterations in the liver of sea bream, Sparus aurata L., caused by short- or long-term feeding with vegetable oils. recovery of normal morphology after feeding fish oil as the sole lipid source. J. Fish Dis. 2004, 27, 531–541. [CrossRef]
- Rodrigues, S.; Antunes, S.C.; Nunes, B.; Correia, A.T. Histological alterations in gills and liver of rainbow trout (Oncorhynchus mykiss) after exposure to the antibiotic oxytetracycline. Environ. Toxicol. Pharmacol. 2017, 53, 164–176. [CrossRef]
- Camargo, M.M.P.; Martinez, C.B.R. Histopathology of Gills, Kidney and liver of a neotropical fish caged in an urban stream. Neotrop. Ichthyol. 2007, 5, 327–336. [CrossRef]
- Chlebicz-Wójcik, A.; Śliżewska, K. The effect of recently developed synbiotic preparations on dominant fecal microbiota and organic acids concentrations in feces of piglets from nursing to fattening. Anim. 2020, 10, 1–18. [CrossRef]
- Rimoldi, S.; Di Rosa, A.R.; Oteri, M.; Chiofalo, B.; Hasan, I.; Saroglia, M.; Terova, G. The impact of diets containing Hermetia illucens meal on the growth, intestinal health, and microbiota of gilthead seabream (Sparus aurata). Fish Physiol. Biochem. 2024. [CrossRef]
- Rimoldi, S.; Gini, E.; Iannini, F.; Gasco, L.; Terova, G. The effects of dietary insect meal from Hermetia illucens prepupae on autochthonous gut microbiota of rainbow trout (Oncorhynchus mykiss). Animals 2019, 9, 143. [CrossRef]
- Rimoldi, S.; Antonini, M.; Gasco, L.; Moroni, F.; Terova, G. Intestinal microbial communities of rainbow trout (Oncorhynchus mykiss) may be improved by feeding a Hermetia illucens meal/low-fishmeal diet. Fish Physiol. Biochem. 2021, 47, 365–380. [CrossRef]
- Bolyen, E., Rideout, J.R., Dillon, M.R. et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [CrossRef]
- Torrecillas, S.; Rimoldi, S.; Montero, D.; Serradell, A.; Acosta, F.; Fontanillas, R.; Allal, F.; Haffray, P.; Bajek, A.; Terova, G. Genotype x Nutrition interactions in European sea bass (Dicentrarchus labrax): effects on gut health and intestinal microbiota. Aquaculture 2023, 574, 739639. [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [CrossRef]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and qualitative beta diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [CrossRef]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16s rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [CrossRef]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. Genome analysis STAMP: Statistical Analysis of Taxonomic and Functional Profiles. 2014, 30, 3123–3124. [CrossRef]
- Hammer, D.A.T.; Ryan, P.D.; Hammer, Ø.; Harper, D.A.T. Past: Paleontological Statistics Software Package for education and data analysis. Palaeontol. Electron. 2001, 4, 178.
- Macusi, E.D.; Cayacay, M.A.; Borazon, E.Q.; Sales, A.C.; Habib, A.; Fadli, N.; Santos, M.D. Protein fishmeal replacement in aquaculture: a systematic review and implications on growth and adoption viability. Sustain. 2023, 15. [CrossRef]
- Aragão, C.; Gonçalves, A.T.; Costas, B.; Azeredo, R.; Xavier, M.J.; Engrola, S. Alternative proteins for fish diets: implications beyond growth. Animals 2022, 12. [CrossRef]
- De Marco, G.; Cappello, T.; Maisano, M. Histomorphological changes in fish gut in response to prebiotics and probiotics treatment to improve their health status: a review. Animals 2023, 13. [CrossRef]
- Bruslé, J.; Gonzàlez i Anadon, G. The structure and function of fish liver. Fish Morphol. 2017, 77–93. [CrossRef]
- Roberts, R.J. Nutritional pathology. In Fish Nutrition; Academic Press: London, UK, 2022; pp. 823–855 ISBN 978-0-12-819587-1.
- Donadelli, V.; Di Marco, P.; Mandich, A.; Finoia, M.G.; Cardinaletti, G.; Petochi, T.; Longobardi, A.; Tibaldi, E.; Marino, G. Effects of dietary plant protein replacement with insect and poultry by-product meals on the liver health and serum metabolites of sea bream (Sparus aurata) and sea bass (Dicentrarchus labrax). Animals 2024, 14. [CrossRef]
- Hasan, I.; Gai, F.; Cirrincione, S.; Rimoldi, S.; Saroglia, G.; Terova, G. Chitinase and insect meal in aquaculture nutrition: a comprehensive overview of the latest achievements. Fishes 2023, 8, 1–16. [CrossRef]
- Terova, G.; Díaz, N.; Rimoldi, S.; Ceccotti, C.; Gliozheni, E.; Piferrer, F. effects of sodium butyrate treatment on histone modifications and the expression of genes related to epigenetic regulatory mechanisms and immune response in european sea bass (Dicentrarchus labrax) fed a plant-based diet. PLoS One 2016, 11, 1–20. [CrossRef]
- Tran, N.T.; Liang, H.; Li, J.; Deng, T.; Zhang, M.; Li, S. Health benefits of butyrate and its producing bacterium, Clostridium butyricum, on aquatic animals. Fish Shellfish Immunol. Reports 2023, 4, 100088. [CrossRef]
- Tran, N.T.; Li, Z.; Wang, S.; Zheng, H.; Aweya, J.J.; Wen, X.; Li, S. Progress and perspectives of short-chain fatty acids in aquaculture. Rev. Aquac. 2020, 12, 283–298. [CrossRef]
- Rangel, F.; Santos, R.A.; Monteiro, M.; Lavrador, A.S.; Gasco, L.; Gai, F.; Oliva-Teles, A.; Enes, P.; Serra, C.R. Isolation of chitinolytic bacteria from European sea bass gut microbiota fed diets with distinct insect meals. Biology (Basel). 2022, 11. [CrossRef]
- Rangel, F.; Enes, P.; Gasco, L.; Gai, F.; Hausmann, B.; Berry, D.; Oliva-Teles, A.; Serra, C.R.; Pereira, F.C. Differential modulation of the European sea bass gut microbiota by distinct insect meals. Front. Microbiol. 2022, 13, 1249. [CrossRef]
- Busti, S.; Bonaldo, A.; Candela, M.; Scicchitano, D.; Trapella, G.; Brambilla, F.; Guidou, C.; Trespeuch, C.; Sirri, F.; Dondi, F.; et al. Hermetia illucens larvae meal as an alternative protein source in practical diets for gilthead sea bream (Sparus aurata): a study on growth, plasma biochemistry and gut microbiota. Aquaculture 2024, 578. [CrossRef]
- Terova, G.; Rimoldi, S.; Ascione, C.; Gini, E.; Ceccotti, C.; Gasco, L. rainbow trout (Oncorhynchus mykiss) gut microbiota is modulated by insect meal from Hermetia illucens prepupae in the diet. Rev. Fish Biol. Fish. 2019, 29, 465–486. [CrossRef]
- Rimoldi, S.; Antonini, M.; Gasco, L.; Moroni, F.; Terova, G. Intestinal Microbial Communities of rainbow trout (Oncorhynchus mykiss) may be improved by feeding a Hermetia illucens meal/low-fishmeal diet. Fish Physiol. Biochem. 2021, 47, 365–380. [CrossRef]
- Paulsen, S.S.; Andersen, B.; Gram, L.; MacHado, H. Biological potential of chitinolytic marine bacteria. Mar. Drugs 2016, 14. [CrossRef]
- Swiontek Brzezinska, M.; Jankiewicz, U.; Burkowska, A.; Walczak, M. Chitinolytic Microorganisms and their possible application in environmental protection. Curr. Microbiol. 2014, 68, 71. [CrossRef]
- Gupta, A.; Gupta, P.; Dhawan, A. Paenibacillus polymyxa as a water additive improved immune response of Cyprinus carpio and disease resistance against Aeromonas hydrophila. Aquac. Reports 2016, 4, 86–92. [CrossRef]
- Chen, S.W.; Liu, C.H.; Hu, S.Y. Dietary administration of probiotic Paenibacillus ehimensis npust1 with bacteriocin-like activity improves growth performance and immunity against Aeromonas hydrophila and Streptococcus iniae in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2019, 84, 695–703. [CrossRef]
- Amoah, K.; Huang, Q. cheng; Dong, X. hui; Tan, B. ping; Zhang, S.; Chi, S. yan; Yang, Q. hui; Liu, H. yu; Yang, Y. zhi Paenibacillus polymyxa improves the growth, immune and antioxidant activity, intestinal health, and disease resistance in Litopenaeus vannamei challenged with Vibrio parahaemolyticus. Aquaculture 2020, 518, 734563. [CrossRef]
Figure 1.
Standard hematoxylin-eosin (H&E) histochemical analysis of proximal (panels A, C, E), and distal (panels B, D, F) seabass intestine. 5 µm cross sections were obtained from the proximal and distal paraffin-embedded intestine of CTRL (panels A and B), PM20 (panels C and D), and PM25 (panels E and F) fish. M, mucosa; SM, submucosa; CM, circular muscle layer; LM, longitudinal muscle layer. Scale bar = 1000 µm.
Figure 1.
Standard hematoxylin-eosin (H&E) histochemical analysis of proximal (panels A, C, E), and distal (panels B, D, F) seabass intestine. 5 µm cross sections were obtained from the proximal and distal paraffin-embedded intestine of CTRL (panels A and B), PM20 (panels C and D), and PM25 (panels E and F) fish. M, mucosa; SM, submucosa; CM, circular muscle layer; LM, longitudinal muscle layer. Scale bar = 1000 µm.
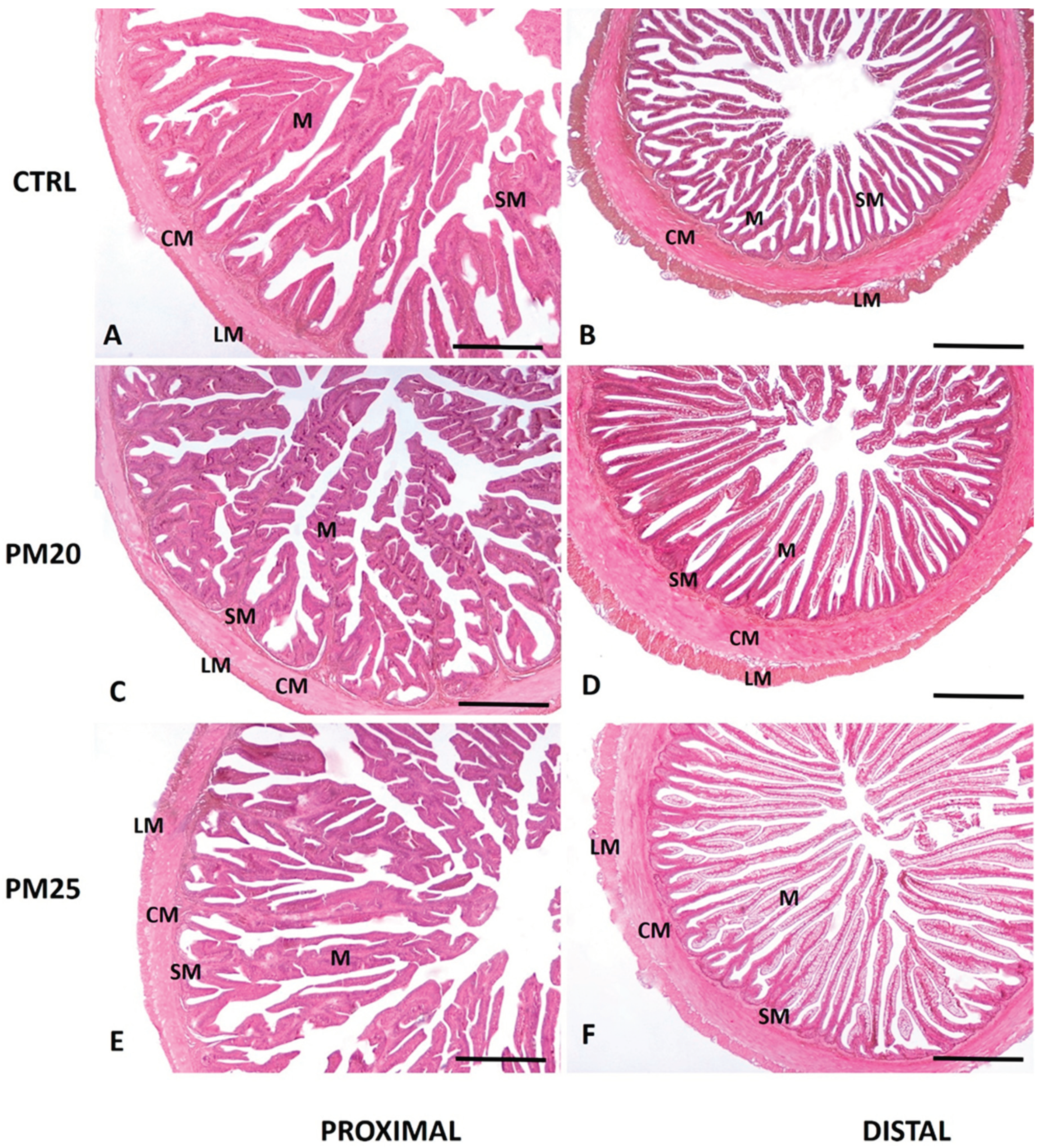
Figure 2.
Standard hematoxylin-eosin (H&E) histochemical analysis of liver of representative images of fish fed Control, PM20, and PM25 diets. 5 µm cross sections were obtained from the liver paraffin-embedded intestine of CTRL (panels A and B), PM20 (panels C and D), and PM25 (panels E and F) fish. Scale bar = 100 µm.
Figure 2.
Standard hematoxylin-eosin (H&E) histochemical analysis of liver of representative images of fish fed Control, PM20, and PM25 diets. 5 µm cross sections were obtained from the liver paraffin-embedded intestine of CTRL (panels A and B), PM20 (panels C and D), and PM25 (panels E and F) fish. Scale bar = 100 µm.
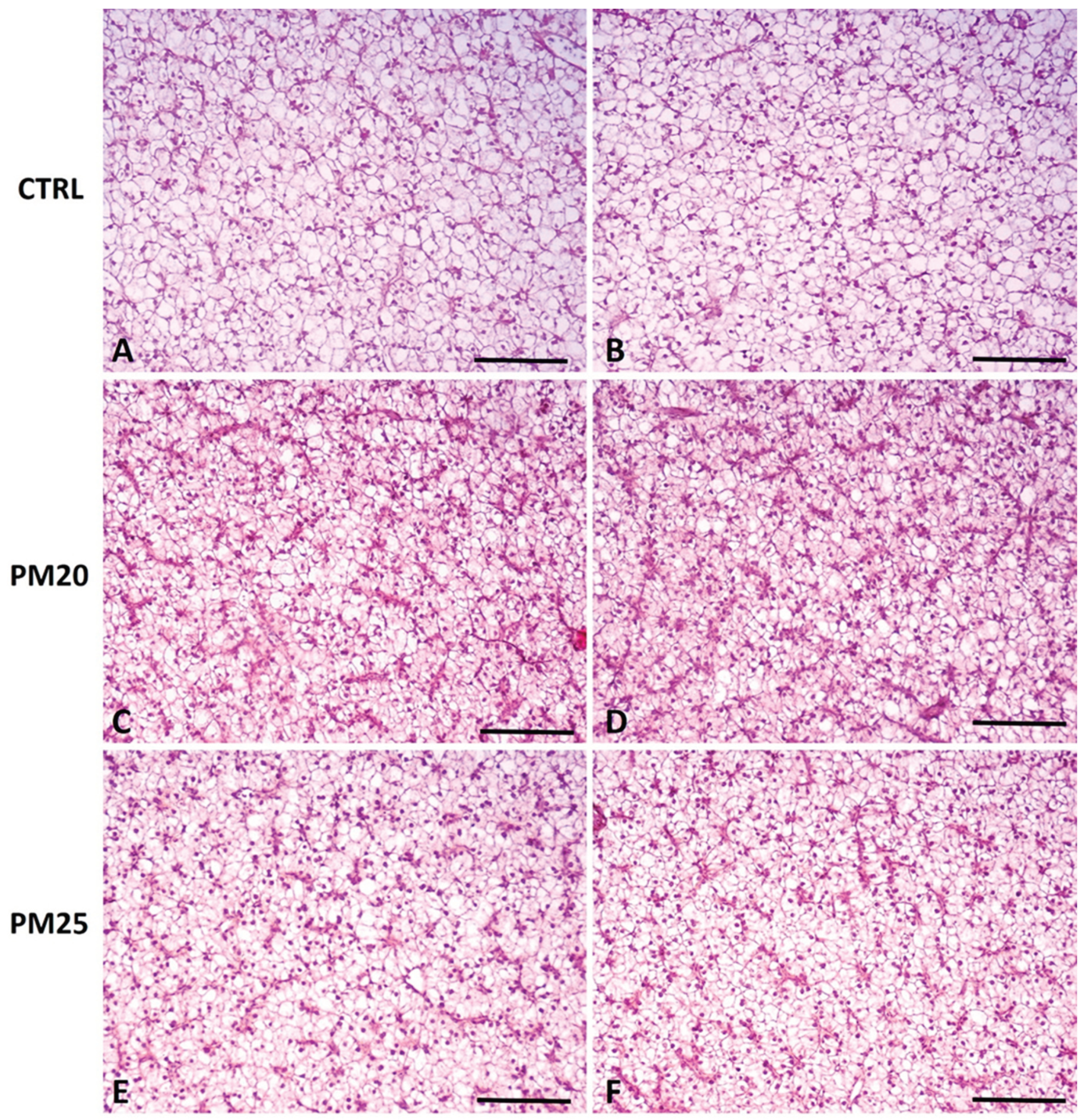
Figure 3.
Stacked bar chart of the mean relative abundances (%) of the most abundant classified bacterial phyla (A) and families (B) most frequently found in feed samples.
Figure 3.
Stacked bar chart of the mean relative abundances (%) of the most abundant classified bacterial phyla (A) and families (B) most frequently found in feed samples.
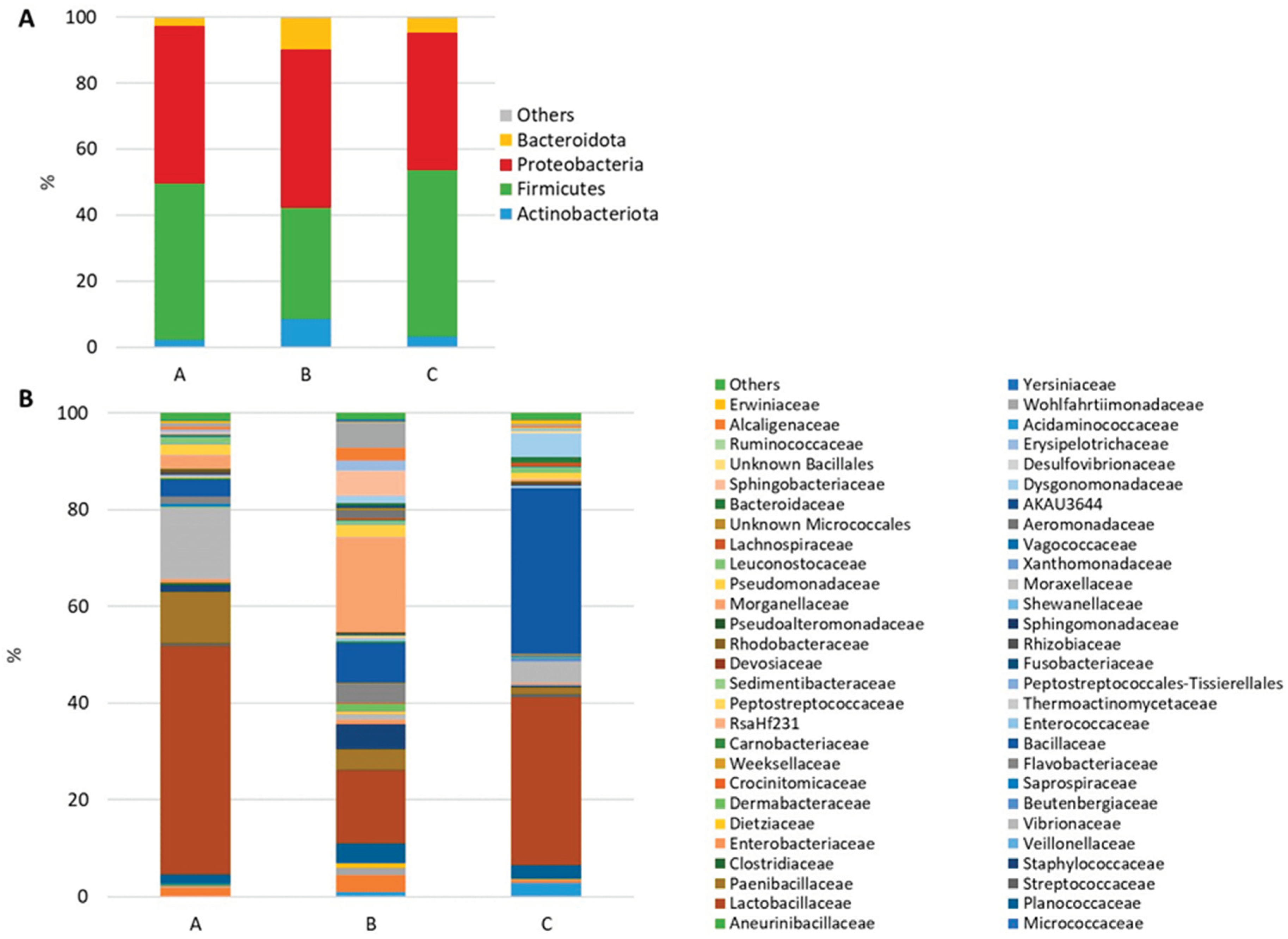
Figure 4.
Stacked bar chart of the mean relative abundances (%) of the most abundant classified bacterial phyla (A) and families (B) found in gut samples.
Figure 4.
Stacked bar chart of the mean relative abundances (%) of the most abundant classified bacterial phyla (A) and families (B) found in gut samples.
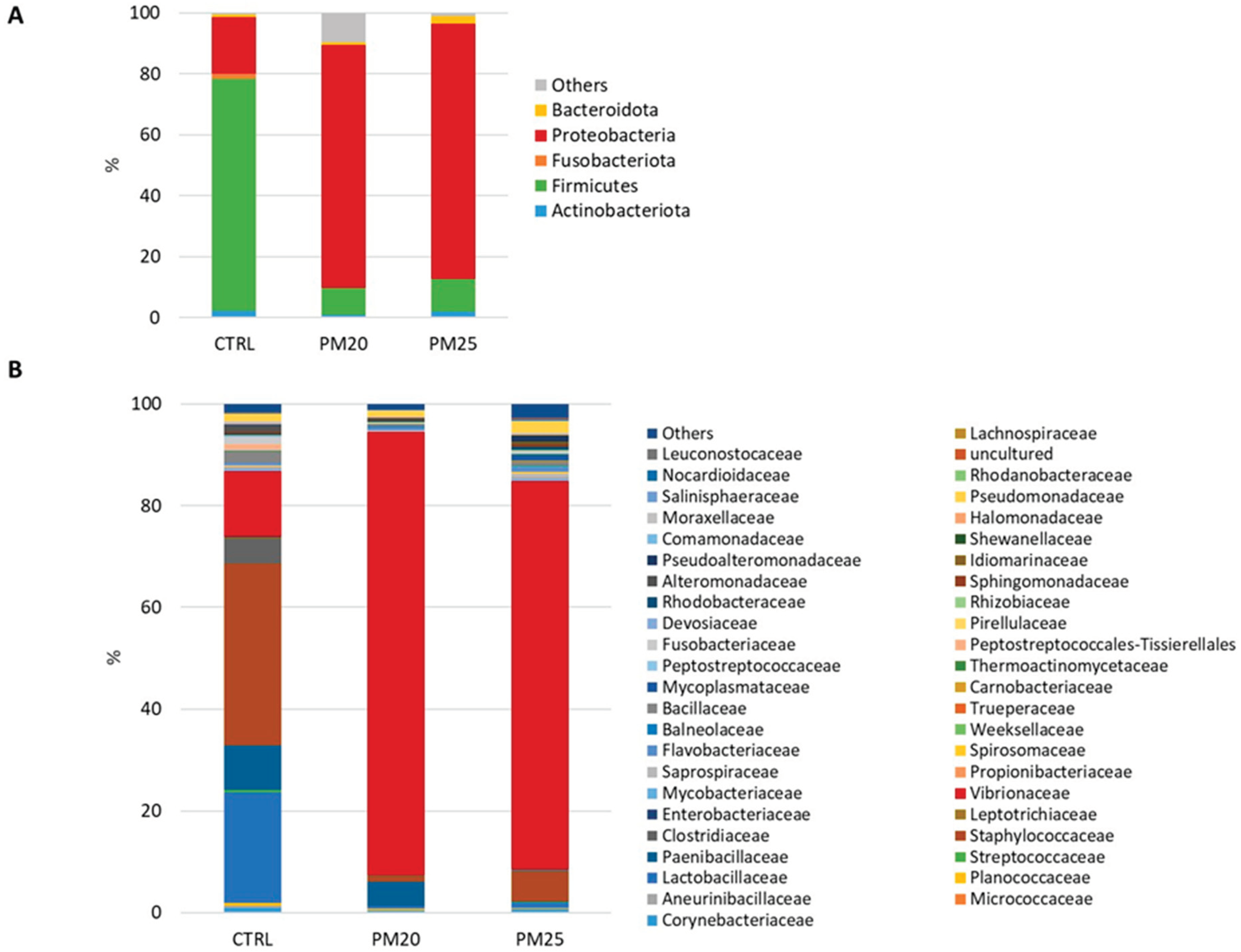
Figure 5.
Plot of principal coordinate analysis (PCoA) using weighted (A) and unweigheted (B) UniFrac distance matrices of gut microbial communities at the genus level.
Figure 5.
Plot of principal coordinate analysis (PCoA) using weighted (A) and unweigheted (B) UniFrac distance matrices of gut microbial communities at the genus level.
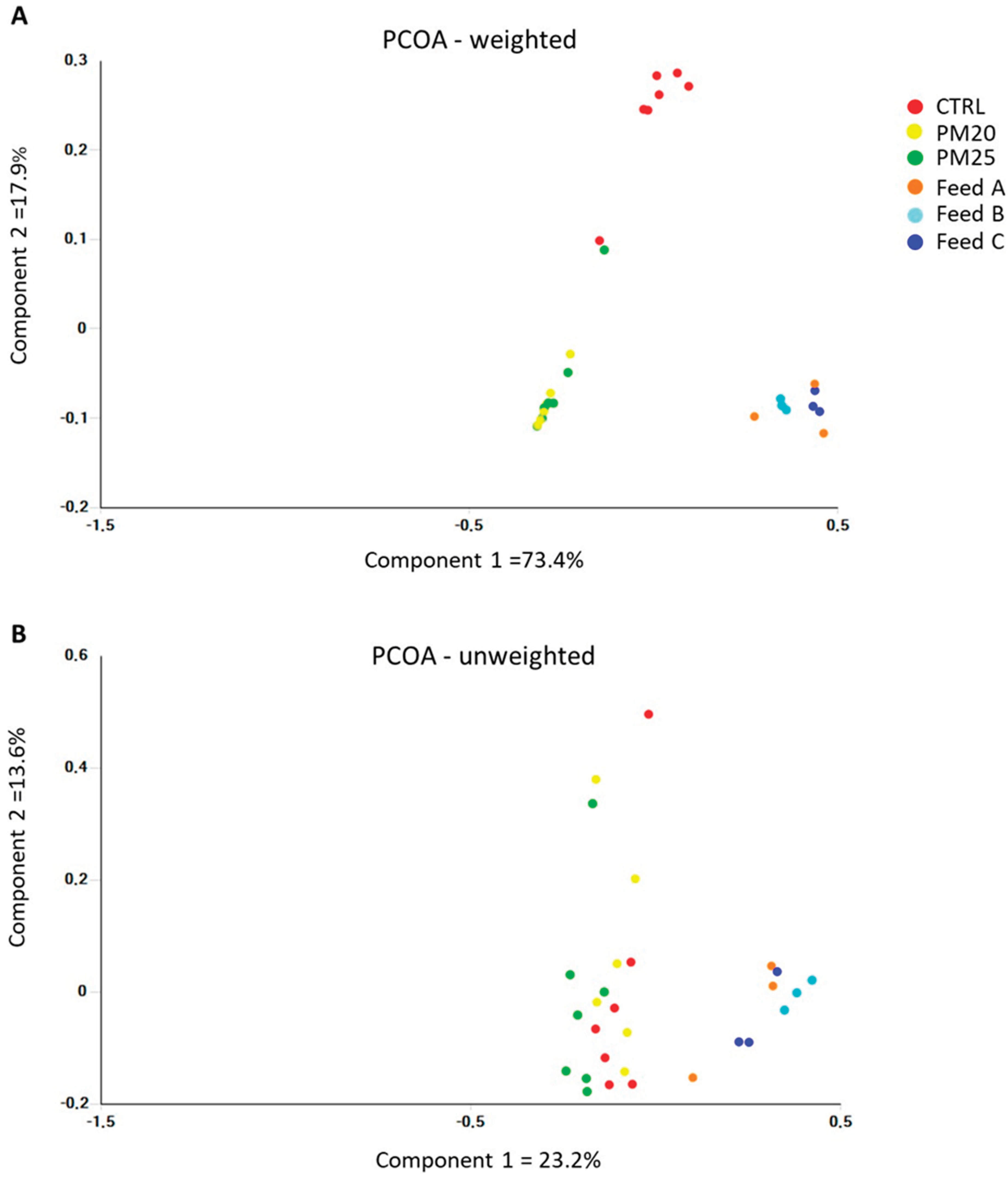
Figure 6.
Comparison of the relative abundance of the PICRUSt functional profile of the gut microbiota between the control and PM20 experimental groups. Only the predicted functional pathways that differ significantly (p < 0.05) are shown.
Figure 6.
Comparison of the relative abundance of the PICRUSt functional profile of the gut microbiota between the control and PM20 experimental groups. Only the predicted functional pathways that differ significantly (p < 0.05) are shown.
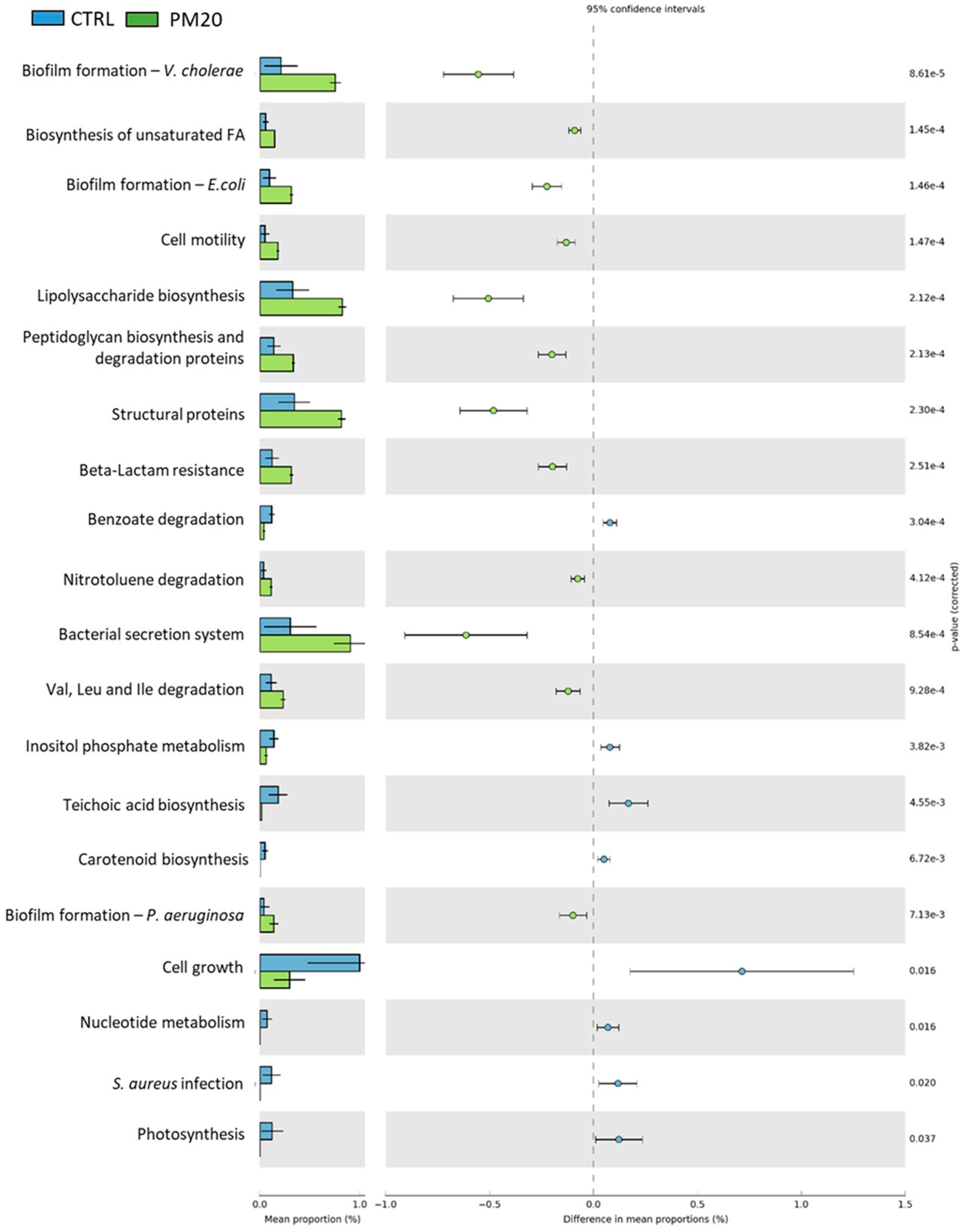
Figure 7.
Comparison of the relative abundance of PICRUSt functional profile of the gut microbiota between the control and PM25 experimental groups. Only the predicted functional pathways that differ significantly (p < 0.05) are shown.
Figure 7.
Comparison of the relative abundance of PICRUSt functional profile of the gut microbiota between the control and PM25 experimental groups. Only the predicted functional pathways that differ significantly (p < 0.05) are shown.
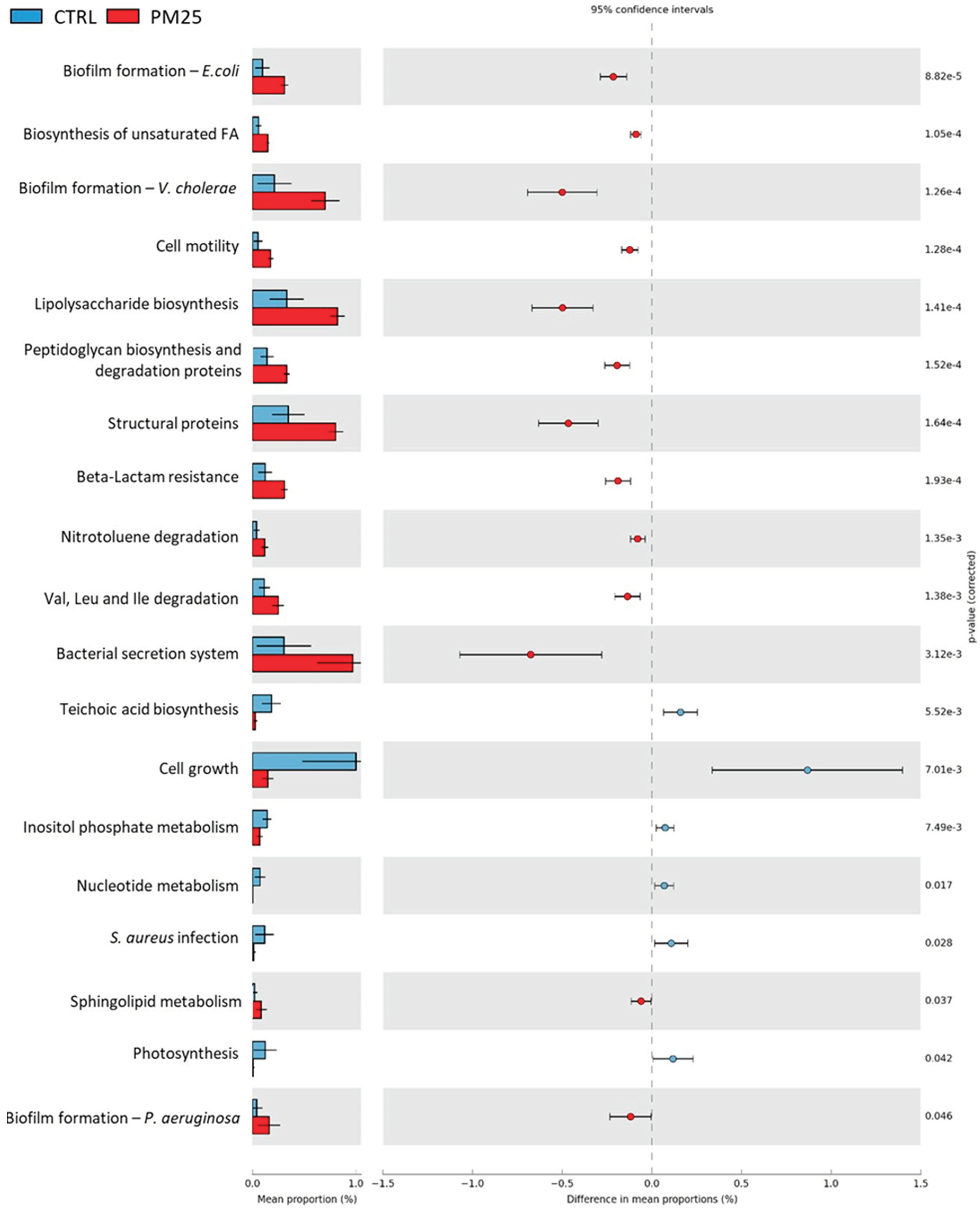

Table 1.
Diet formulations.
| INGREDIENTS (%) | A | B | C |
|---|---|---|---|
| Fishmeal | 20 | 10 | 10 |
| Poultry by-product | 0 | 20 | 25 |
| Fish oil | 3 | 3 | 3 |
| Soybean meal | 11 | 11 | 11 |
| Exuviae meal | 0 | 0 | 0.5 |
| Insect meal | 0 | 5.0 | 0 |
| Corn gluten meal | 21.9 | 7.9 | 7.9 |
| Vital wheat gluten | 6 | 6 | 6 |
| Soy Protein Concentrate | 6 | 4 | 2 |
| Rapeseed meal | 11 | 11 | 12.5 |
| Soybean oil | 2 | 2 | 2 |
| Minerals and Vitamins | 2 | 2 | 2 |
| Premix antioxidants | 0.1 | 0.1 | 0.1 |
Table 2.
Proximate composition of diets.
| A | B | C | |
|---|---|---|---|
| Gross Energy (MJ/kg) | 17.34 | 17.55 | 17.58 |
| DE (MJ/kg) | 15.60 | 15.66 | 15.66 |
| DE (%) | 89.98 | 89.23 | 89.09 |
| Crude Fat (g/100 g) | 10.63 | 10.77 | 10.82 |
| Crude Protein (g/100 g) | 45.08 | 45.00 | 45.04 |
| DP (%) | 40.17 | 39.35 | 39.28 |
| Digestible Protein (%) | 89.12 | 87.44 | 87.22 |
| Fish Protein (%) | 13.20 | 6.60 | 6.60 |
| Animal Protein (%) | 13.20 | 22.98 | 23.85 |
| FP/TP (%) | 29.28 | 14.67 | 14.65 |
| DP/DE (mg/Kj or g/MJ) | 25.75 | 25.14 | 25.09 |
| AP/TP (%) | 29.28 | 51.06 | 52.96 |
| Fiber (g/100 g) | 3.03 | 3.41 | 3.04 |
| EI (g/100 g) | 35.48 | 32.29 | 33.01 |
| Amido (g/100 g) | 15.95 | 15.55 | 15.55 |
| NSP (g/100 g) | 22.57 | 21.15 | 20.51 |
| Protein-to-lipid ratio | 4.50 | 4.18 | 4.16 |
| Ash (g/100 g) | 6.38 | 7.53 | 8.09 |
| DE (kcal/kg) | 3729.34 | 3741.83 | 3742.61 |
| Crude En (kcal/kg) | 4304.66 | 4243.40 | 4233.13 |
Notes: FP/TP: Fish Protein/Total Protein; DP/DE: Digestible Protein/Digestible Energy, AP/TP: Animal Protein/Total Protein, NFE: Nitrogen-Free Extracts; NSP: Non-Starch Polysaccharides.
Table 3.
Growth performance of seabass fed with three experimental diets. Different letters in the same row indicate a significant difference between the mean values (p<0.05; N(A) = 70; N(B) = 67; N(C) = 69).
Table 3.
Growth performance of seabass fed with three experimental diets. Different letters in the same row indicate a significant difference between the mean values (p<0.05; N(A) = 70; N(B) = 67; N(C) = 69).
| CTRL | PM20 | PM25 | |
|---|---|---|---|
| IBW (g) | 52.88 ± 7.24 | 52.16 ± 6.14 | 52.04 ± 5.13 |
| FBW (g) | 135.88 ± 20.59a | 120.26 ± 19.76b | 128.37 ± 23.01ab |
| SGR (% day-1) | 1.1 | 0.96 | 1.03 |
| FCR | 1.76 | 2.17 | 1.94 |
Notes: initial body weight (IBW); final body weight (FBW); standard growth rate (SGR); feed conversion ratio (FCR). Values are given as mean ± standard deviation.
Table 4.
Intestinal morphometric parameters of seabass fed experimental diets.
| CTRL | PM20 | PM25 | p-value | |
|---|---|---|---|---|
| Proximal intestine | ||||
| ViH (μm) | 1008.78 ± 21.50 | 1064.90 ± 37.14 | 1035.87 ± 31.11 | 0.435 |
| ViW (μm) | 78.35 ± 2.57 | 83.18 ± 3.05 | 84.79 ± 3.42 | 0.300 |
| LPW (μm) | 28.44 ± 1.00 | 29.64 ± 1.42 | 27.3 ± 1.78 | 0.585 |
| SMT (μm) | 352.65 ± 14.93 | 365.94 ± 18.56 | 332.02 ± 9.22 | 0.266 |
| Distal intestine | ||||
| ViH (μm) | 1008.78 ± 21.50 | 1064.90 ± 3 7.14 | 1035.87 ± 31.11 | 0.435 |
| ViW (μm) | 78.35 ± 2.57 | 83.18 ± 3.05 | 84.79 ± 3.42 | 0.300 |
| LPW (μm) | 28.44 ± 1.00 | 29.64 ± 1.42 | 27.3 ± 1.78 | 0.585 |
| SMT (μm) | 352.65 ± 14.93 | 365.94 ± 18.56 | 332.02 ± 9.22 | 0.266 |
Notes: villus height (ViH), villus width (ViW), lamina propria width (LPW), and submucos layer thickness (SMT) Values are given as mean ± standard error.
Table 5.
Results of histological scoring of the liver.
| CTRL | PM 20% | PM 25% | |
|---|---|---|---|
| HV | 2.00 | 1.67 | 2.00 |
| ND | 2.00 | 1.33 | 1.67 |
| NS | 1.00 | 1.00 | 1.00 |
| CH | 1.00 | 1.00 | 1.00 |
| Total | 6.00 | 5.00 | 5.67 |
Notes: HV, hepatocytes vacuolization; ND, nuclear displacement; NS, irregular nuclei shapes; CH, cellular hypertrophy.
Table 6.
Volatile SCFA content in fecal samples from three test groups. The results are given in mmol/L, mean ± SD (N = 6).
Table 6.
Volatile SCFA content in fecal samples from three test groups. The results are given in mmol/L, mean ± SD (N = 6).
| Acetate (C 2:0) | Propionate (C 3:0) | Butyrate (C 4:0) | |
|---|---|---|---|
| CTRL | 14.67 ± 0.66b | 5.93 ± 0.62c | 2.13 ± 0.27b |
| PM20 | 17.05 ± 0.94a | 6.43 ± 0.46b | 2.00 ± 0.13b |
| PM25 | 15.19 ± 0.42b | 7.41 ± 0.18a | 3.03 ± 0.13a |
Table 7.
Alpha diversity analysis of bacterial communities associated with feeds.
| A | B | C | p-value | |
|---|---|---|---|---|
| Observed OTUs | 1325 ± 216b | 1721 ± 33a | 1224 ± 168b | 0.019 |
| Chao1 | 1541 ± 250ab | 1893 ± 47a | 1382 ± 226b | 0.047 |
| Faith-PD | 21.7 ± 4.9 | 23.9 ± 1.3 | 21.8 ± 3.3 | > 0.05 |
| Shannon | 5.8 ± 0.4b | 7.1 ± 0.1a | 5.9 ± 0.2b | 0.004 |
| Simpson | 0.92 ± 0.02 | 0.95 ± 0.00 | 0.92 ± 0.00 | > 0.05 |
Table 8.
The list of bacterial genera that differ between experimental feeds A (control) and B.
| PHYLUM | CLASS | ORDER | FAMILY | GENUS | A (%) | SD (%) | B (%) | SD (%) | p-value |
|---|---|---|---|---|---|---|---|---|---|
| Actinobacteriota | Actinobacteria | Micrococcales | AKAU3644 | AKAU3644 | 0.01 | 0.01 | 0.64 | 0.02 | 0.0000 |
| Actinobacteriota | Actinobacteria | Micrococcales | Dermabacteraceae | Brachybacterium | 0.16 | 0.06 | 1.44 | 0.07 | 0.0001 |
| Actinobacteriota | Actinobacteria | Actinomycetales | Actinomycetaceae | Actinomyces | 0.19 | 0.11 | 1.20 | 0.10 | 0.0006 |
| Actinobacteriota | Actinobacteria | Micrococcales | Brevibacteriaceae | Brevibacterium | 0.28 | 0.13 | 1.70 | 0.06 | 0.0012 |
| Actinobacteriota | Actinobacteria | Micrococcales | Microbacteriaceae | Leucobacter | 0.06 | 0.02 | 0.43 | 0.04 | 0.0019 |
| Actinobacteriota | Actinobacteria | Micrococcales | Micrococcaceae | Enteractinococcus | 0.05 | 0.02 | 0.19 | 0.01 | 0.0023 |
| Actinobacteriota | Actinobacteria | Corynebacteriales | Dietziaceae | Dietzia | 0.06 | 0.01 | 0.66 | 0.07 | 0.0070 |
| Actinobacteriota | Actinobacteria | Corynebacteriales | Corynebacteriaceae | Corynebacterium | 1.62 | 0.74 | 3.72 | 0.19 | 0.0497 |
| Bacteroidota | Bacteroidia | Bacteroidales | Dysgonomonadaceae | Dysgonomonadaceae | 0.03 | 0.01 | 0.36 | 0.02 | 0.0000 |
| Bacteroidota | Bacteroidia | Sphingobacteriales | Sphingobacteriaceae | Sphingobacterium | 0.30 | 0.13 | 2.62 | 0.10 | 0.0001 |
| Bacteroidota | Bacteroidia | Flavobacteriales | Flavobacteriaceae | Myroides | 1.06 | 0.23 | 3.89 | 0.13 | 0.0004 |
| Bacteroidota | Bacteroidia | Bacteroidales | Dysgonomonadaceae | Dysgonomonas | 0.17 | 0.12 | 1.33 | 0.07 | 0.0009 |
| Bacteroidota | Bacteroidia | Sphingobacteriales | Sphingobacteriaceae | Pedobacter | 0.02 | 0.02 | 0.13 | 0.02 | 0.0034 |
| Bacteroidota | Bacteroidia | Flavobacteriales | Flavobacteriaceae | Ulvibacter | 0.02 | 0.01 | 0.38 | 0.05 | 0.0059 |
| Bacteroidota | Bacteroidia | Bacteroidales | Bacteroidaceae | Bacteroides | 0.04 | 0.02 | 0.51 | 0.07 | 0.0066 |
| Desulfobacterota | Desulfovibrionia | Desulfovibrionales | Desulfovibrionaceae | Desulfovibrio | 0.00 | 0.01 | 0.14 | 0.01 | 0.0002 |
| Firmicutes | Clostridia | Peptostreptococcales-Tissierellales | Sedimentibacteraceae | Sedimentibacter | 0.01 | 0.01 | 0.11 | 0.00 | 0.0007 |
| Firmicutes | Bacilli | Bacillales | Bacillaceae | Pseudogracilibacillus | 0.70 | 0.23 | 2.72 | 0.18 | 0.0008 |
| Firmicutes | Bacilli | Lactobacillales | Enterococcaceae | Enterococcus | 0.10 | 0.05 | 0.63 | 0.06 | 0.0009 |
| Firmicutes | Bacilli | Staphylococcales | Staphylococcaceae | Staphylococcus | 1.43 | 0.44 | 5.38 | 0.23 | 0.0014 |
| Firmicutes | Bacilli | RsaHf231 | RsaHf231 | RsaHf231 | 0.01 | 0.01 | 0.16 | 0.02 | 0.0020 |
| Firmicutes | Bacilli | Bacillales | Bacillaceae | Bacillus | 1.15 | 0.12 | 2.09 | 0.16 | 0.0032 |
| Firmicutes | Bacilli | Erysipelotrichales | Erysipelotrichaceae | Erysipelothrix | 0.41 | 0.32 | 2.20 | 0.21 | 0.0043 |
| Firmicutes | Bacilli | Bacillales | Bacillaceae | Amphibacillus | 0.08 | 0.04 | 0.31 | 0.02 | 0.0062 |
| Firmicutes | Clostridia | Peptostreptococcales-Tissierellales | Peptostreptococcaceae | Clostridioides | 0.07 | 0.04 | 0.25 | 0.01 | 0.0153 |
| Firmicutes | Clostridia | Lachnospirales | Lachnospiraceae | Lachnoclostridium | 0.02 | 0.02 | 0.11 | 0.03 | 0.0273 |
| Firmicutes | Clostridia | Oscillospirales | Ruminococcaceae | Incertae_Sedis | 0.01 | 0.01 | 0.08 | 0.02 | 0.0300 |
| Proteobacteria | Gammaproteobacteria | Enterobacterales | Morganellaceae | Providencia | 1.14 | 0.62 | 12.29 | 0.56 | 0.0001 |
| Proteobacteria | Gammaproteobacteria | Pseudomonadales | Pseudomonadaceae | Oblitimonas | 0.19 | 0.12 | 1.32 | 0.14 | 0.0010 |
| Proteobacteria | Gammaproteobacteria | Cardiobacteriales | Wohlfahrtiimonadaceae | Ignatzschineria | 0.83 | 0.38 | 5.12 | 0.54 | 0.0013 |
| Proteobacteria | Gammaproteobacteria | Enterobacterales | Yersiniaceae | Serratia | 0.09 | 0.06 | 0.64 | 0.03 | 0.0017 |
| Proteobacteria | Gammaproteobacteria | Burkholderiales | Alcaligenaceae | Paenalcaligenes | 0.24 | 0.11 | 1.28 | 0.15 | 0.0019 |
| Proteobacteria | Gammaproteobacteria | Aeromonadales | Aeromonadaceae | Aeromonas | 0.15 | 0.09 | 1.97 | 0.20 | 0.0022 |
| Proteobacteria | Gammaproteobacteria | Burkholderiales | Alcaligenaceae | Alcaligenes | 0.26 | 0.02 | 1.47 | 0.12 | 0.0035 |
| Proteobacteria | Gammaproteobacteria | Enterobacterales | Morganellaceae | Proteus | 1.49 | 0.83 | 7.20 | 0.24 | 0.0068 |
| Proteobacteria | Gammaproteobacteria | Enterobacterales | Morganellaceae | Morganella | 0.12 | 0.07 | 1.99 | 0.33 | 0.0122 |
| Proteobacteria | Alphaproteobacteria | Rhizobiales | Rhizobiaceae | Paenochrobactrum | 0.08 | 0.05 | 0.41 | 0.06 | 0.0044 |
Table 9.
The list of bacterial genera that differ between experimental feeds A (control) and C.
| PHYLUM | CLASS | ORDER | FAMILY | GENUS | A (%) | SD (%) | C (%) | SD (%) | p-value |
|---|---|---|---|---|---|---|---|---|---|
| Actinobacteriota | Actinobacteria | Actinomycetales | Actinomycetaceae | Actinomyces | 0.19 | 0.11 | 3.01 | 0.22 | 0.0006 |
| Bacteroidota | Bacteroidia | Bacteroidales | Bacteroidaceae | Bacteroides | 0.04 | 0.02 | 1.27 | 0.05 | 0.0001 |
| Bacteroidota | Bacteroidia | Bacteroidales | Dysgonomonadaceae | Dysgonomonadaceae | 0.03 | 0.01 | 1.93 | 0.06 | 0.0002 |
| Bacteroidota | Bacteroidia | Bacteroidales | Dysgonomonadaceae | Dysgonomonas | 0.17 | 0.12 | 2.90 | 0.44 | 0.0090 |
| Bacteroidota | Bacteroidia | Flavobacteriales | Flavobacteriaceae | Myroides | 1.06 | 0.23 | 0.10 | 0.04 | 0.0244 |
| Firmicutes | Clostridia | Oscillospirales | Ruminococcaceae | Incertae_Sedis | 0.01 | 0.01 | 0.23 | 0.01 | 0.0000 |
| Firmicutes | Bacilli | Bacillales | Planococcaceae | Sporosarcina | 0.41 | 0.17 | 2.54 | 0.21 | 0.0004 |
| Firmicutes | Clostridia | Lachnospirales | Lachnospiraceae | Lachnoclostridium | 0.02 | 0.02 | 0.17 | 0.02 | 0.0019 |
| Firmicutes | Clostridia | Peptostreptococcales-Tissierellales | Sedimentibacteraceae | Sedimentibacter | 0.01 | 0.01 | 0.11 | 0.01 | 0.0040 |
| Firmicutes | Bacilli | Bacillales | Bacillaceae | Bacillus | 1.15 | 0.12 | 21.97 | 1.98 | 0.0044 |
| Firmicutes | Bacilli | Bacillales | Bacillaceae | Ureibacillus | 0.23 | 0.10 | 12.00 | 1.37 | 0.0065 |
| Firmicutes | Negativicutes | Acidaminococcales | Acidaminococcaceae | Phascolarctobacterium | 0.01 | 0.01 | 0.14 | 0.04 | 0.0303 |
| Firmicutes | Clostridia | Clostridiales | Clostridiaceae | Clostridium_sensu_stricto_18 | 0.10 | 0.03 | 0.02 | 0.02 | 0.0416 |
| Firmicutes | Bacilli | RsaHf231 | RsaHf231 | RsaHf231 | 0.01 | 0.01 | 0.06 | 0.02 | 0.0443 |
| Firmicutes | Bacilli | Staphylococcales | Staphylococcaceae | Staphylococcus | 1.43 | 0.44 | 0.16 | 0.12 | 0.0460 |
| Proteobacteria | Gammaproteobacteria | Burkholderiales | Alcaligenaceae | Alcaligenes | 0.26 | 0.02 | 0.05 | 0.01 | 0.0027 |
Table 10.
The list of bacterial genera that differ between experimental feeds B and C.
| PHYLUM | CLASS | ORDER | FAMILY | GENUS | B (%) | SD (%) | C (%) | SD (%) | p-value |
|---|---|---|---|---|---|---|---|---|---|
| Actinobacteriota | Actinobacteria | Micrococcales | Brevibacteriaceae | Brevibacterium | 1.70 | 0.06 | 0.07 | 0.03 | 4.41E-05 |
| Actinobacteriota | Actinobacteria | Corynebacteriales | Corynebacteriaceae | Corynebacterium | 3.72 | 0.19 | 0.46 | 0.17 | 5.54E-05 |
| Actinobacteriota | Actinobacteria | Micrococcales | Dermabacteraceae | Brachybacterium | 1.44 | 0.07 | 0.08 | 0.03 | 0.0003 |
| Actinobacteriota | Actinobacteria | Micrococcales | Micrococcaceae | Enteractinococcus | 0.19 | 0.01 | 0.01 | 0.01 | 0.0004 |
| Actinobacteriota | Actinobacteria | Micrococcales | AKAU3644 | AKAU3644 | 0.64 | 0.02 | 0.00 | 0.00 | 0.0004 |
| Actinobacteriota | Actinobacteria | Micrococcales | Microbacteriaceae | Leucobacter | 0.43 | 0.04 | 0.10 | 0.03 | 0.0015 |
| Actinobacteriota | Actinobacteria | Actinomycetales | Actinomycetaceae | Actinomyces | 1.20 | 0.10 | 3.01 | 0.22 | 0.0025 |
| Actinobacteriota | Actinobacteria | Corynebacteriales | Dietziaceae | Dietzia | 0.66 | 0.07 | 0.02 | 0.03 | 0.0032 |
| Bacteroidota | Bacteroidia | Flavobacteriales | Crocinitomicaceae | Fluviicola | 0.16 | 0.01 | 0.00 | 0.00 | 2.09E-05 |
| Bacteroidota | Bacteroidia | Sphingobacteriales | Sphingobacteriaceae | Sphingobacterium | 2.62 | 0.10 | 0.09 | 0.04 | 0.0001 |
| Bacteroidota | Bacteroidia | Bacteroidales | Dysgonomonadaceae | Dysgonomonadaceae | 0.36 | 0.02 | 1.93 | 0.06 | 0.0002 |
| Bacteroidota | Bacteroidia | Flavobacteriales | Flavobacteriaceae | Myroides | 3.89 | 0.13 | 0.10 | 0.04 | 0.0003 |
| Bacteroidota | Bacteroidia | Bacteroidales | Bacteroidaceae | Bacteroides | 0.51 | 0.07 | 1.27 | 0.05 | 0.0005 |
| Bacteroidota | Bacteroidia | Flavobacteriales | Flavobacteriaceae | Flavobacterium | 0.17 | 0.02 | 0.03 | 0.02 | 0.0021 |
| Bacteroidota | Bacteroidia | Sphingobacteriales | Sphingobacteriaceae | Pedobacter | 0.13 | 0.02 | 0.01 | 0.01 | 0.0041 |
| Bacteroidota | Bacteroidia | Flavobacteriales | Flavobacteriaceae | Ulvibacter | 0.38 | 0.05 | 0.09 | 0.06 | 0.0055 |
| Bacteroidota | Bacteroidia | Bacteroidales | Dysgonomonadaceae | Dysgonomonas | 1.33 | 0.07 | 2.90 | 0.44 | 0.0343 |
| Firmicutes | Clostridia | Peptostreptococcales-Tissierellales | Peptostreptococcaceae | Clostridioides | 0.25 | 0.01 | 0.01 | 0.01 | 0.0001 |
| Firmicutes | Bacilli | Staphylococcales | Staphylococcaceae | Staphylococcus | 5.38 | 0.23 | 0.16 | 0.12 | 0.0001 |
| Firmicutes | Bacilli | Bacillales | Bacillaceae | Pseudogracilibacillus | 2.72 | 0.18 | 0.85 | 0.12 | 0.0005 |
| Firmicutes | Bacilli | Bacillales | Bacillaceae | Amphibacillus | 0.31 | 0.02 | 0.05 | 0.01 | 0.0005 |
| Firmicutes | Bacilli | Bacillales | Bacillaceae | Virgibacillus | 0.23 | 0.01 | 0.14 | 0.01 | 0.0008 |
| Firmicutes | Bacilli | Bacillales | Planococcaceae | Sporosarcina | 0.74 | 0.11 | 2.54 | 0.21 | 0.0015 |
| Firmicutes | Clostridia | Oscillospirales | Ruminococcaceae | Incertae_Sedis | 0.08 | 0.02 | 0.23 | 0.01 | 0.0031 |
| Firmicutes | Clostridia | Clostridiales | Clostridiaceae | Clostridium_sensu_stricto_7 | 0.00 | 0.00 | 0.04 | 0.00 | 0.0033 |
| Firmicutes | Bacilli | Bacillales | Bacillaceae | Geobacillus | 0.04 | 0.01 | 0.10 | 0.01 | 0.0038 |
| Firmicutes | Bacilli | Lactobacillales | Enterococcaceae | Enterococcus | 0.63 | 0.06 | 0.14 | 0.02 | 0.0038 |
| Firmicutes | Bacilli | Bacillales | Bacillaceae | Bacillus | 2.09 | 0.16 | 21.97 | 1.98 | 0.0047 |
| Firmicutes | Bacilli | Erysipelotrichales | Erysipelotrichaceae | Erysipelothrix | 2.20 | 0.21 | 0.40 | 0.05 | 0.0047 |
| Firmicutes | Bacilli | Bacillales | Planococcaceae | Savagea | 2.62 | 0.27 | 0.16 | 0.04 | 0.0052 |
| Firmicutes | Bacilli | Lactobacillales | Streptococcaceae | Streptococcus | 0.30 | 0.03 | 0.72 | 0.07 | 0.0057 |
| Firmicutes | Bacilli | RsaHf231 | RsaHf231 | RsaHf231 | 0.16 | 0.02 | 0.06 | 0.02 | 0.0060 |
| Firmicutes | Bacilli | Bacillales | Bacillaceae | Ureibacillus | 0.19 | 0.05 | 12.00 | 1.37 | 0.0066 |
| Firmicutes | Bacilli | Lactobacillales | Leuconostocaceae | Weissella | 0.69 | 0.02 | 1.17 | 0.07 | 0.0070 |
| Firmicutes | Bacilli | Lactobacillales | Carnobacteriaceae | Atopostipes | 0.36 | 0.05 | 0.07 | 0.02 | 0.0077 |
| Firmicutes | Bacilli | Bacillales | Bacillaceae | Cerasibacillus | 0.14 | 0.01 | 0.07 | 0.01 | 0.0094 |
| Firmicutes | Bacilli | Lactobacillales | Lactobacillaceae | Lactobacillus | 16.56 | 0.76 | 35.81 | 3.29 | 0.0111 |
| Firmicutes | Bacilli | Staphylococcales | Staphylococcaceae | Nosocomiicoccus | 0.13 | 0.02 | 0.00 | 0.00 | 0.0159 |
| Firmicutes | Bacilli | Bacillales | Bacillaceae | Oceanobacillus | 0.24 | 0.04 | 0.09 | 0.01 | 0.0251 |
| Fusobacteriota | Fusobacteriia | Fusobacteriales | Fusobacteriaceae | Cetobacterium | 0.06 | 0.01 | 0.13 | 0.01 | 0.0042 |
| Proteobacteria | Gammaproteobacteria | Enterobacterales | Yersiniaceae | Serratia | 0.64 | 0.03 | 0.03 | 0.02 | 6.01E-05 |
| Proteobacteria | Gammaproteobacteria | Enterobacterales | Morganellaceae | Proteus | 7.20 | 0.24 | 0.07 | 0.02 | 0.0005 |
| Proteobacteria | Gammaproteobacteria | Enterobacterales | Morganellaceae | Providencia | 12.29 | 0.56 | 0.17 | 0.06 | 0.0009 |
| Proteobacteria | Gammaproteobacteria | Pseudomonadales | Moraxellaceae | Acinetobacter | 0.23 | 0.02 | 0.05 | 0.02 | 0.0010 |
| Proteobacteria | Gammaproteobacteria | Burkholderiales | Alcaligenaceae | Alcaligenes | 1.47 | 0.12 | 0.05 | 0.01 | 0.0032 |
| Proteobacteria | Gammaproteobacteria | Burkholderiales | Alcaligenaceae | Paenalcaligenes | 1.28 | 0.15 | 0.31 | 0.08 | 0.0038 |
| Proteobacteria | Gammaproteobacteria | Aeromonadales | Aeromonadaceae | Aeromonas | 1.97 | 0.20 | 0.06 | 0.02 | 0.0050 |
| Proteobacteria | Gammaproteobacteria | Pseudomonadales | Pseudomonadaceae | Oblitimonas | 1.32 | 0.14 | 0.04 | 0.01 | 0.0052 |
| Proteobacteria | Gammaproteobacteria | Cardiobacteriales | Wohlfahrtiimonadaceae | Ignatzschineria | 5.12 | 0.54 | 0.46 | 0.02 | 0.0066 |
| Proteobacteria | Gammaproteobacteria | Enterobacterales | Morganellaceae | Morganella | 1.99 | 0.33 | 0.08 | 0.04 | 0.0134 |
| Proteobacteria | Alphaproteobacteria | Rhizobiales | Rhizobiaceae | Paenochrobactrum | 0.41 | 0.06 | 0.07 | 0.01 | 0.0139 |
| Proteobacteria | Gammaproteobacteria | Xanthomonadales | Xanthomonadaceae | Stenotrophomonas | 0.18 | 0.04 | 0.05 | 0.02 | 0.0169 |
Table 11.
Result of the alpha diversity analysis of the microbial communities in the gut.
| CTRL | PM20 | PM25 | p-value | |
|---|---|---|---|---|
| Observed OTUs | 1144 ± 380 | 756 ± 329 | 945 ± 445 | > 0.05 |
| Chao1 | 1276 ± 440 | 914 ± 386 | 1086 ± 471 | > 0.05 |
| Faith-PD | 18.5 ± 6.7 | 15.8 ± 5.6 | 18.5 ± 5.9 | > 0.05 |
| Shannon | 6.20 ± 1.06 | 4.69 ± 0.65 | 5.18 ± 1.23 | 0.048 |
| Simpson | 0.93 ± 0.04 | 0.89 ± 0.02 | 0.89 ± 0.05 | > 0.05 |
Table 12.
Validation of the PCoA analysis.
| Unweighted UniFrac PCoA | |||||
|---|---|---|---|---|---|
| Anosim | |||||
| Sample size | Permutations | R | p-value | ||
| CTRL | PM20 | 13 | 999 | 0.316 | 0.005 |
| CTRL | PM25 | 14 | 999 | 0.312 | 0.005 |
| PM20 | PM25 | 13 | 999 | 0.184 | 0.037 |
| CTRL | A | 10 | 999 | 0.496 | 0.030 |
| PM20 | B | 9 | 999 | 0.728 | 0.014 |
| PM25 | C | 10 | 999 | 0.639 | 0.007 |
| Permanova | |||||
| Sample size | Permutations | pseudo-F | p-value | ||
| CTRL | PM20 | 13 | 999 | 1.922 | 0.010 |
| CTRL | PM25 | 14 | 999 | 1.979 | 0.003 |
| PM20 | PM25 | 13 | 999 | 1.567 | 0.024 |
| CTRL | A | 10 | 999 | 2.486 | 0.013 |
| PM20 | B | 9 | 999 | 4.473 | 0.012 |
| PM25 | C | 10 | 999 | 3.749 | 0.007 |
| Weighted UniFrac PCoA | |||||
| Anosim | |||||
| Sample size | Permutations | R | p-value | ||
| CTRL | PM20 | 13 | 999 | 0.947 | 0.003 |
| CTRL | PM25 | 14 | 999 | 0.892 | 0.002 |
| PM20 | PM25 | 13 | 999 | -0.009 | 0.437 |
| CTRL | A | 10 | 999 | 1 | 0.009 |
| PM20 | B | 9 | 999 | 1 | 0.013 |
| PM25 | C | 10 | 999 | 1 | 0.009 |
| Permanova | |||||
| Sample size | Permutations | pseudo-F | p-value | ||
| CTRL | PM20 | 13 | 999 | 38.427 | 0.002 |
| CTRL | PM25 | 14 | 999 | 26.332 | 0.001 |
| PM20 | PM25 | 13 | 999 | 1.441 | 0.233 |
| CTRL | A | 10 | 999 | 23.772 | 0.012 |
| PM20 | B | 9 | 999 | 268.127 | 0.012 |
| PM25 | C | 10 | 999 | 75.875 | 0.011 |
Table 13.
The list of genera of intestinal bacteria that differ between CTRL and PM20 fish.
| PHYLUM | CLASS | ORDER | FAMILY | GENUS | CTRL (%) | SD (%) | PM20 (%) | SD (%) | p-values |
|---|---|---|---|---|---|---|---|---|---|
| Actinobacteriota | Actinobacteria | Corynebacteriales | Mycobacteriaceae | Mycobacterium | 0.69 | 0.51 | 0.04 | 0.05 | 0.0210 |
| Bacteroidota | Bacteroidia | Flavobacteriales | Flavobacteriaceae | Ulvibacter | 0.11 | 0.08 | 0.01 | 0.02 | 0.0196 |
| Firmicutes | Bacilli | Staphylococcales | Staphylococcaceae | Staphylococcus | 36.87 | 24.78 | 1.14 | 1.04 | 0.0123 |
| Firmicutes | Bacilli | Bacillales | Bacillaceae | Geobacillus | 0.84 | 0.62 | 0.00 | 0.00 | 0.0168 |
| Firmicutes | Clostridia | Clostridiales | Clostridiaceae | Clostridium_sensu_stricto_7 | 1.23 | 0.96 | 0.01 | 0.01 | 0.0208 |
| Firmicutes | Clostridia | Clostridiales | Clostridiaceae | Clostridium_sensu_stricto_1 | 0.53 | 0.36 | 0.07 | 0.14 | 0.0228 |
| Firmicutes | Clostridia | Peptostreptococcales-Tissierellales | Peptostreptococcales-Tissierellales | Anaerosalibacter | 1.08 | 0.89 | 0.02 | 0.03 | 0.0259 |
| Firmicutes | Clostridia | Clostridiales | Clostridiaceae | Clostridium_sensu_stricto_18 | 0.28 | 0.24 | 0.01 | 0.01 | 0.0364 |
| Firmicutes | Bacilli | Bacillales | Bacillaceae | Oceanobacillus | 0.25 | 0.21 | 0.04 | 0.04 | 0.0494 |
| Firmicutes | Bacilli | Lactobacillales | Lactobacillaceae | Lactobacillus | 22.47 | 22.09 | 0.35 | 0.36 | 0.0496 |
| Fusobacteriota | Fusobacteriia | Fusobacteriales | Fusobacteriaceae | Cetobacterium | 1.47 | 0.81 | 0.00 | 0.01 | 0.0044 |
| Proteobacteria | Gammaproteobacteria | Vibrionales | Vibrionaceae | Photobacterium | 6.66 | 5.30 | 43.79 | 10.80 | 0.0002 |
| Proteobacteria | Gammaproteobacteria | Vibrionales | Vibrionaceae | Vibrio | 6.70 | 11.52 | 44.16 | 13.29 | 0.0006 |
Table 14.
The list of genera of intestinal bacteria that differ between CTRL and PM25 fish.
| PHYLUM | CLASS | ORDER | FAMILY | GENUS | CTRL (%) | SD (%) | PM25 (%) | SD (%) | p-value |
|---|---|---|---|---|---|---|---|---|---|
| Firmicutes | Clostridia | Clostridiales | Clostridiaceae | Clostridium sensu stricto 1 | 0.53 | 0.36 | 0.06 | 0.08 | 0.0188 |
| Firmicutes | Bacilli | Paenibacillales | Paenibacillaceae | Paenibacillus | 9.33 | 7.21 | 0.06 | 0.07 | 0.0197 |
| Firmicutes | Bacilli | Bacillales | Bacillaceae | Geobacillus | 0.84 | 0.62 | 0.04 | 0.05 | 0.0209 |
| Firmicutes | Bacilli | Staphylococcales | Staphylococcaceae | Staphylococcus | 36.87 | 24.78 | 6.14 | 8.80 | 0.0226 |
| Firmicutes | Bacilli | Bacillales | Bacillaceae | Oceanobacillus | 0.25 | 0.21 | 0.00 | 0.01 | 0.0288 |
| Firmicutes | Clostridia | Peptostreptococcales-Tissierellales | Peptostreptococcales-Tissierellales | Anaerosalibacter | 1.08 | 0.89 | 0.06 | 0.09 | 0.0300 |
| Firmicutes | Bacilli | Mycoplasmatales | Mycoplasmataceae | Mycoplasma | 0.02 | 0.05 | 1.23 | 1.14 | 0.0413 |
| Firmicutes | Clostridia | Clostridiales | Clostridiaceae | Clostridium sensu stricto 18 | 0.28 | 0.24 | 0.03 | 0.05 | 0.0452 |
| Fusobacteriota | Fusobacteriia | Fusobacteriales | Fusobacteriaceae | Cetobacterium | 1.47 | 0.81 | 0.08 | 0.16 | 0.0052 |
| Proteobacteria | Gammaproteobacteria | Vibrionales | Vibrionaceae | Vibrio | 6.70 | 11.52 | 43.21 | 30.77 | 0.0273 |
| Proteobacteria | Gammaproteobacteria | Vibrionales | Vibrionaceae | Photobacterium | 6.66 | 5.30 | 34.77 | 26.22 | 0.0393 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



