Submitted:
01 March 2024
Posted:
01 March 2024
You are already at the latest version
Abstract
Verticillium wilt is a soil borne disease caused by distinct vegetative compatibility groups (VCG) of the fungus Verticillium dahliae. Two V. dahliae isolates were recovered from symptomatic cot-ton plants in Australian cotton fields and were assigned to two distinct vegetative compatibility groups (VCGs). One corresponding to the defoliating (VCG 1A) and the other non-defoliating (VCG 2A) pathotypes of Verticillium, have been transformed with Red and Green Fluorescent protein genes, respectively. The transformants maintained their ability to infect cotton and both strains were observed to colonise the xylem vessels of cotton plants. Moreover, we observed that the cotton V. dahliae strains could also infect some sampled non-Gossypium species found in the Australian landscape. The fluorescently labelled strains of V. dahliae will allow us to gain a thorough understanding of the infection processes of this important pathogen.
Keywords:
Reporter genes
; plant host and pathogen interactions
; Green Fluorescent Protein
; 'mCherry' Red Fluorescent Protein
; alternative weed host
; Nicotina benthamiana
; Vegetative Compatibility Groups 1A and 2A
; Defoliating and non-defoliating pathotypes
1. Introduction
Cotton (Gossypium L.) is a dicotyledonous, perennial shrub cultivated for its soft fibres that develop around the seeds of the cotton plant [1]. Grown in over 80 countries and in a range of ecological niches, cotton is the most economically important source of natural fibre worldwide [2]. The genus Gossypium contains over 50 species, of which four are cultivated globally as annual crops [3]. Gossypium hirsutum L., known as upland cotton, is the most widely grown of these species, representing 90% of cotton production globally [3,4,5]. In Australia, upland cotton production comprises an expanding multi-billion-dollar industry that employs over 12,000 people nationwide [6].
Verticillium dahliae Kleb. is an asexual, soil-borne member of the Ascomycota which acts as a monocyclic phytopathogen to over 400 plant species [7,8,9]. It is speculated that V. dahliae originated in Europe [7,10], and has since spread to major temperate cropping regions worldwide [11]. V. dahliae is the primary causal agent of Verticillium wilt in many economically important crop species, including upland cotton [12]. Verticillium wilt is a systemic disease that arises from the colonisation and subsequent occlusion of the host vasculature by the pathogen [13]. Resulting symptoms include wilting and stunting of the host, dropping of foliage, discolouration of vascular tissue, development of necrotic lesions, and plant death [14]. In Australia, Verticillium wilt is reported to cause reduction in cotton yields in the range of 10% to 62% under pathogen conducive conditions [15].
Eradication of V. dahliae is challenging once it becomes established in the field. The fungus can persist in soils for up to 14 years in the absence of a susceptible host as melanised resting structures called microsclerotia [16]. The longevity of these microsclerotia makes it difficult to eradicate the fungus through conventional disease management strategies, such as crop rotation or field fallowing [7]. Microsclerotia remain dormant in soils until they are triggered to germinate by the presence of plant root exudates [7,17]. Upon entering the host xylem tissues, hyaline, asexual conidiospores are produced, which disseminate systemically and colonise the above-ground host vasculature [13]. The accumulation of fungal material and the resulting host defence responses lead to occlusion of the xylem vessels, inducing the Verticillium wilt disease symptoms [13]. Upon senescence of the plant, the pathogen permeates the surface of the decaying tissues, producing microsclerotia, after which the fungus returns to dormancy in the soil [18].
Pathogenic strains of V. dahliae are generally categorised based on pathotype [19] or race [20]. In cotton, V. dahliae pathotypes are described as either defoliating or non-defoliating, depending on the symptoms induced in the host. The defoliating pathotype is generally considered more severe, whilst non-defoliating isolates may vary in disease aggression [21,22,23]. Populations within V. dahliae can be further classified on a sub-species level based on vegetative compatibility. This refers to the ability of isolates within a fungal species to exchange genetic information through the successful formation of vegetative heterokaryons [24]. Due to the clonal nature of the V. dahliae populations, hyphal anastomosis provides the only mechanism of genetic exchange between strains of V. dahliae [25]. Consequently, vegetative compatibility groups (VCGs) within V. dahliae have classically been considered as genetically isolated populations that may differ from each other in physiology, virulence, environmental response, and host range [25,26].
Prior to the 2013/14 cropping season, the only V. dahliae pathotype reported in Australian cotton fields was the mildly virulent VCG 4B, however, this changed following the confirmation of the non-defoliating VCG 2A pathotype in NSW from isolates collected in the 2009/10 season [27] and the 2011/2012 [28]. Chapman et al. (2016) used VCG classification with nit mutant testing and molecular assays performed on a set of eight historical isolates dating back as far as the 2009/2010 growing seasons in NSW Australia detected the presence of both VCG 4B and 2A as contributors of the non-defoliating pathotypes observed in Australian cotton fields [27]. In the following season, VCG 1A, a defoliating pathotype, was also detected for the first time. Internationally, VCG 1A has been shown to be a highly virulent pathotype [15,27] and has been associated with severe yield losses in cotton, as well as in olive production [21]. However, typical disease presentation and similar crop losses caused by VCG 1A internationally have not been widely observed in Australia [7]. On the other hand, VCG 2A has been associated with widespread disease and yield losses in Australian cotton fields, despite there being no reports of VCG 2A causing the same damage overseas [7,29]. The reasons behind the disparities occurring between these pathotypes are currently unknown.
It has been suggested that the dominating presence of V. dahliae VCG 2A observed in Australian cotton fields may be attributed to its ability to colonise and maintain inoculum capacity on weedy hosts common to these regions [7,30]. Alternate hosts have the potential to act as intermittent reservoirs for V. dahliae, increasing inoculum levels in the field [31]. Pathogenic V. dahliae VCG 2A strains have been reported infecting economically important weed species from cotton fields overseas [30]. In Australia, susceptibility to V. dahliae has been described previously in a range of native and non-native weedy hosts [32]; although these reports do not characterise the VCGs associated with these infections. The study by Evans in 1971 [32] also suggests the potential for V. dahliae to endophytically colonise weed species localised to Australian cotton-growing regions. Endophytic infection of hosts within the cotton field would pose an additional challenge to V. dahliae management by potentially enhancing carry over of inoculum. It is possible that V. dahliae VCG 2A has become the dominant strain in Australian cotton fields by endophytically adapting to a broad range of local hosts [7].
A better understanding of how V. dahliae moves between geographic areas will guide strategies to limit the spread of VCG 1A and 2A in Australian cotton-growing regions. Established modes of transmission for V. dahliae include movement of infected plant material or soil, irrigation, and use of contaminated equipment [8]. In cotton, there is evidence suggesting that seed-borne transmission of the pathogen is also possible. A study on commercially available seed lots reported widespread distribution of V. dahliae in cotton seed from Turkish fields [33]. The study identified V. dahliae isolates belonging to the VCGs 1A, 2A, 2B, and 4B from infected seeds. At present, only the defoliating pathotype of V. dahliae has been reported to infect seed from Australian cotton varieties at a very low incidence. The pathogen was recovered from 0.025% of seed collected from diseased plants [34]. The potential for the emerging Australian VCGs to be transmitted by Australian cotton seed therefore remains an ongoing concern for the industry.
The implementation of fluorescent reporter technologies can provide novel insights into the complex interactions between V. dahliae and its hosts. Integration of fluorescent proteins into plant pathogens has facilitated understanding of infection and colonisation patterns in studies on other filamentous fungi [35,36,37]. The Green Fluorescent Protein (GFP), isolated from Aequorea victoria [38], and the mCherry fluorescent protein, optimised from Dicosoma sp. [39], are two widely used fluorescent proteins suitable for expressing in V. dahliae [40,41]. V. dahliae isolates tagged with GFP or mCherry have been used to understand pathogen intercellular dissemination in cotton [40], antifungal activities of compounds [42], infection of seed in sunflower [43], infection responses in lettuce resistant or susceptible to Verticillium wilt [44], and pathogenic processes in the model species Nicotiana benthamiana [41].
The aim of this study was to use fluorescent proteins to observe infection by the emerging V. dahliae VCGs 1A and 2A in planta, and to investigate what has allowed these strains to persist in Australian cotton-growing regions. For this purpose, VCG 1A and VCG 2A isolates were stably transformed with mCherry or GFP, respectively. Furthermore, the potential for these strains to colonise weed species common to regions of cotton production in Australia were investigated. The results from this study contribute to the understanding of the epidemiology of Verticillium wilt in cotton and shed light on how this economically significant pathogen can survive on common weeds as alternative hosts, allowing disease control and management to be improved.
2. Materials and Methods
2.1. Fungal isolates
Two isolates, Verticillium dahliae strains Vd71171 (BRIP71171) and Vd71181 (BRIP71181) were used in this study (Table 1). They were obtained from the Queensland Department of Agriculture and Fisheries (Queensland Australia) phytopathology herbarium. Vd71171 was originally isolated from the Namoi Valley, NSW, Australia. This isolate was confirmed to group with the non-defoliating VCG 2A by nitrate-nonutilizing (nit) mutant complementation tests. Vd71181 originated from Gwydir Valley, NSW, Australia and was predicted to be V. dahliae based on the amplification of a specific ITS region sharing 100 % sequence identity to V. dahliae, as well as its defoliating pathotype typically associated with VCG 1A. Isolate Vd71172 and Vd71181 pathotypes were further confirmed by PCR using primers that can distinguish one pathotype from the other [45,46]. Whole genome SNP profile of Vd 71181 showed that it phylogenetically clusters with other strains in the VCG 1A subclade [47].
2.2. Plant materials used in this study
Untreated and de-linted cotton seeds from two Australian upland cotton cultivars, Siokra (full name: Siokra 1-4) and Sicot (full name: Sicot 714 B3F) were used in this study. Siokra has been shown to be susceptible to Fusarium oxysporum f. sp. vasinfectum, the Fusarium wilt pathogen of cotton, and V. dahliae (https://csd.net.au/variety-guide/; accessed on 26 Feb 2024), whilst Sicot is generally considered to be tolerant to both pathogens [48].
Seven weed species collected from cotton-growing region of Narrabri, NSW, Australia, were used in this study. They include flaxleaf fleabane (Conyza bonariensis), common sowthistle (Sonchus oleraceus), wild oats (Avena fatua), windmill grass (Chloris truncata), feathertop Rhodes grass (Chloris virgata), liverseed grass (Urochloa panicoides), and awnless barnyard grass (Echinochloa colona). Seeds from a native tobacco relative with known susceptibility to V. dahliae, Nicotiana benthamiana, were also used [41].
2.3. Vector construction and transformation
V. dahliae non-defoliating strain Vd71171 was previously transformed with a construct to express GFP driven by the strong constitutive translation elongation factor 1 alpha promoter from Aspergillus nidulans [42].
For transforming the defoliating VCG 1A strain (Vd71181), a vector designed to express the mCherry derivative of DsRed was built. Except for the mCherry coding sequence the final vector was the same as the GFP plasmid described previously [42]. Briefly, the cloning proceeded via isolation of the plasmid backbone from pPZPnat1-TEF-GFP-yeast using BamHI and use of PCR products encoding the TEF promoter, fungal codon-optimised mCherry coding sequence (synthesised by Integrated DNA Technologies, CA, USA) and the TEF terminator in yeast-mediated recombinatorial cloning. Primers are presented in Table S1. Agrobacterium tumefacien strains AGL1 and EHA105 were used to transfer the pPZPnat1-TEF-mCherry-yeast plasmid into V. dahliae using the method previously described [42,49,50].
2.4. Assessment of fungal transformants
Vd71171 and two GFP-expressing transformants (71T0003 and 71T0006) were inoculated on half-strength PDA plates (n = 5) and grown at 24°C under 12 h light / 12 h dark photoperiod before colony diameters were measured. At 7 dpi, plates were flooded with 10 - 15 mL of sterile water and spore concentration was determined using a hemocytometer. In R, a generalised linear model (with poisson regression) was used to model the spore count data and emmeans package used for pairwise comparison.
For the defoliating strain, after assessing the morphology of the transformants (81T0069, 81T0073, 81T0030, and 81T0028) on half-strength PDA plates, spore morphology, fluorescence, and growth rates were further assessed using the Cytation 1 Cell Imaging Multi-Mode Reader (BioTek, VT, USA) and Gen5 imaging software. Briefly, a spore suspension containing 2 x 105 spores/mL in PDB was aliquoted into a 96-well MicroWell™ flat plate (Thermo Fisher Scientific, MA, USA). Bright field and mCherry fluorescence images of spores were then taken over 30 hrs at 1-hr imaging intervals. The Object Sum Area (µm2), fluorescence total intensity (RFU), and cell count were calculated for each image. Maximal growth rate (µm2 / hr) was determined as the maximal slope along the Object Sum Area growth curve and was calculated using the BioTek Gen5 software. Fecundity of the transformants (n = 5) were determined as described in the previous paragraph.
2.6. Plant growth and pathogenicity assay
Siokra 1-4 and Sicot 714 B3F seeds without prior fungicide treatment were potted in seedling trays using steam-pasteurised UQ 23 mix (70% composted pine bark and 30% coco peat). Conditions were maintained at 28°C / 24°C (day/night, defoliating strains experiments) or 24°C / 20°C (non-defoliating strains experiment) with a 16 h photoperiod and 64% / 80% humidity until 8 days post germination. Upon emergence of true leaves, seedlings of similar size were uprooted, and the roots were washed with water and then dipped in an inoculum for 5 min at a spore concentration of 1 x 105 to 1 x 106 conidia/mL. A negative control was included where seedlings were root-dipped in SDW only. Plants were re-planted into 0.64 L pots using UQ 23 mix amended with NPK fertiliser (Osmocote®, Everris, The Netherlands) (4 g/L). Plants were watered every 2 - 4 days.
For non-defoliating Vd71171 or 71T0003 testing, inoculated Siokra and Sicot plants were moved to a temperature-controlled glasshouse set at 25°C. Pots were placed on saucers and contained within a bag to avoid cross-contamination. Pot positions were randomised on a single bench. Four plants per pot, 12 pots per treatment (n = 48) were included.
For the infection time course experiment using 71T0003, 2 - 4 plants were used for destructive examination under confocal microscope at 4 hpi, 1 dpi, 5 dpi, 7 dpi. Water-dipped plants were used as controls.
For the defoliating parent Vd71181 and the transformants, inoculated Sicot plants were moved back in the growth chamber and their positions were randomised. Growth conditions were adjusted to 24°C / 20°C (day / night) post inoculation.
Upon emergence of the second true leaf, weed species common to cotton growing regions in Australia were root-inoculated with transformant strains of V. dahliae VCGs 1A (81T0069) and 2A (71T0003). Plant stem tissue was surface sterilised and then plated onto half-strength PDA at 4 weeks post-inoculation.
2.7. Scoring
Disease severity was scored using a rating scale of 0 to 5 (Table 2) [51] for the non-defoliating strains or a Likert rating scale of 0 to 5 for the defoliating V. dahliae induced symptoms (Table 3) [52]. Plants were assessed for external symptoms visualised as chlorosis, necrosis, and wilting of leaves at 4 weeks post inoculation.
2.8. Re-isolation
At 4 weeks post inoculation, plants were destructively sampled for V. dahliae re-isolation using a previously described method [53]. A 10 - 15 cm segment from the base of the stem of each plant was sampled and then processed under sterile conditions. The tissues were surface sterilised in 70% ethanol for 5 seconds and were blotted dry. Approximately 5 mm x 2 mm pieces were excised from asymptomatic or symptomatic stems where discolouration within the vasculature was visible and were embedded into PDA containing 100 ppm streptomycin sulfate and nourseothricin 50 µg/mL (for transformants only). Plates were incubated in the dark at room temperature until colony growth was observed. Colonies emerging from transformant-inoculated plates were assessed for fluorescence under a Zeiss 700 Laser Scanning Microscope.
For plants inoculated with Vd71181 and the mCherry transformants, single spores were further obtained from the culture of stem sections on half-strength PDA. DNA was extracted from each isolate using a rapid extraction method [54] and PCR was performed using Verticillium specific ITS1 primers [55] to confirm their identity (Table S2).
2.9. Confocal microscopy
A Zeiss 700 laser scanning microscope was used to detect the transgenic fungi with excitation at 488 nm (GFP) and 555 nm (mCherry). GFP and mCherry were typically detected at 500 - 550nm, and 550 - 700 nm, respectively. Sections 0.5 - 1 mm in diameter were excised by using a sterile razor blade from the main root, root cap, lateral roots, lateral root junctions, basal stem, and petiole (Figure S1). GFP examination in plants inoculated with the non-defoliating V. dahliae strain were repeated across two experiments. Samples were examined within hours of sectioning to avoid autofluorescence from plant phenolic compounds, or the decline of fluorescence over time. Images were captured and processed in the software ZEN Blue v3.1 (Zeiss, Oberkochen, Germany).
2.10. Statistical analysis
For the non-defoliating strains associated analysis, results were conducted in R [56] using packages emmeans [57] and RVAideMemoire [58]. Two-way ordinal regression was performed in R and the ANOVA function from RVAideMemoire was used to analyse deviance between treatment groups.
For the defoliating strains, fluorescence data on each isolate (n=5) detected from Cytation 1 and the Biotek Gen5 software were assessed by analysis of variance (ANOVA). A negative binomial generalised linear model (GLM) and post hoc Tukey comparison were performed to compare transformant fecundity with the parent strain. Dispersion of fecundity data was assessed with the AER package in RStudio [59]. Disease scores from the pathogenicity assay and weed experiments were analysed across treatment groups using the non-parametric, rank based Kruskal Wallis H test. All datasets were tested for normality with the Shapiro-Wilk normality test, and homogeneity of variance using Levene’s test. The threshold for statistical significance was a p-value of p < 0.05.
Graphs were produced using the R package ggplot2 [60], or Microsoft Excel (2024).
3. Results
3.1. Transformation of V. dahliae strain Vd71171 does not alter agressiveness of the isolate
We previously described a transformant of V. dahliae that expressed strong constitutive GFP. Here we sought to establish that the transformation process did not affect the strains ability to cause disease for ultimate use in understanding the infection pathway of this pathogen. Of 21 stably transformed V. dahliae isolates, two, 71T0003 and 71T0006, were selected for futher charactersiation based on contrasting colony morphology (Figure 1A) and strength of GFP fluorescence (Figure 1B). Colonies of the Wildtype Vd71171 appeared white, with raised elevation from aerial hyphae, and circular. No microsclerotia were observed when colonies were examined under a microscope. The colony appearance of isolate 71T0003 was consistent with that of Vd71171 (Figure 1A). Isolate 71T0006 appeared dark and had an abundance of microsclerotia when examined under a microscope, flat in elevation, and with irregular colony margins. Colony diameter of all three isolates was measured over a period of 4 days (Figure 1C) and the differences between each pair were not significant (One-way ANOVA, p = 0.8779). In vitro fecundity, or the production of conidia in water, was reduced respectively by 27.3% and 54.5% in 71T0003 and 71T0006, when compared to the wildtype Vd71171 (p < 0.001) (Figure 1D).
Typical Verticillum wilt symptoms, including chlorosis, necrosis, leaf yellowing and wilting were observed on plants inoculated with Vd71171 or 71T0003 (Figure 1E-F). Symptomatic plants were often mixed with asymptomatic plants in the same pot (Figure 1E). No external symptoms were observed on uninoculated plants. There was no significant difference in disease severity between isolate Vd71171 and 71T0003 on either Sicot or Siokra at p = 0.05 (Figure 1G). This suggests that 71T0003 retained the virulence of the parental strain. Mean disease severity on Siokra inoculated with Vd71171 (3.3) or 71T0003 (2.9) was higher than Sicot inoculated with Vd71171 (2.5) or 71T0003 (1.9), respectively (Figure 1G), suggesting that Sicot is relatively more tolerant to these isolates than Siokra, consistent with Sicot 714B3F having a higher V-ranking than Siokra 1-4 which is considered susceptible [48]. This is further supported by the reisolation of these isolates at harvest (Table 4). Of 45 Siokra plants inoculated with Vd71171 or 71T0003, 33 (Vd71171) or 24 (71T0003) plants were symptomatic, whereas of 45 Sicot plants inoculated with Vd71171 or 71T0003, 24 (Vd71171) or 18 (71T0003) plants showed symptoms (Table 4). From these symptomatic plants, reisolation frequency of the wild type Vd71171 or transformant 71T0003 was higher in Siokra (94 - 100%) than in Sicot (39 - 83%) (Table 4). Vd71171 or 71T0003 were also occasionally reisolated from asymptomatic Siokra or Sicot plants inoculated with these strains. The pathology assays collectively suggest that the transformation process did not alter the ability of the fungus to cause disease on cotton.
3.2. Infection of cotton plants by V. dahliae progresses within hours of inoculation with vasculature colonisation and sporulation complete within one week
To assess the colonisation process of cotton by the non-defoliating strain of V. dahliae, a time course of infection was performed using 71T0003. As early as 4 hours post inoculation (hpi) spores expressing GFP were visualised attached to root tip epidermis of Sicot and Siokra. By 24 hpi, an abundant amount of GFP conidia were attached to the root tip epidermis in both Sicot and Siokra (Figure 2A-C), with germ tubes emerging from conidia (Figure 2A), presence of an infection peg (Figure 2B), and hyphal elongation (Figure 2C) clearly observed (Table S3). At 5 days post inoculation (dpi), mycelia were clearly observed on the root tips of both cultivars (Figure 2D-E). Mycelial networks were established along both the surface of and within the root epidermis (Figure 2E-F), appearing densely colonised at the base of the root (Figure 2E), and with intercellular movement of hyphae from the root surface towards the root cortex being evident (Figure 2F). At 7 dpi, advanced colonisation by the GFP-expressing fungus in the vasculature of lateral and main roots was observed (Figure 2G-J). Mycelia were observed in several vessels of the vasculature in the main root adjacent to root junction (Figure 2G). Conidia were also observed in these regions. Formation of dense mycelial networks was detected in the root vasculature (Figure 2F). In the same vessel, tightly packed fluorescent cells appeared to occlude a section of the xylem (Figure 2I). Cells observed using a single channel at an increased magnification suggest that they are ovoid, ranging from 2.2 to 3.8 um in length (Figure 2F). Similar observations were made on Siokra plants (Table S3). Furthermore, mycelia were detected in the vasculature and the adjacent vessels of basal stem sections of Siokra (Figure 2K). Hyphal tips were shown emerging through the vessel wall into an uninfected vessel. Consistent with this observation, hyphae were observed in the vessels of the petiole, and appeared to have penetrated the shared vessel wall (Figure 2L). Several germinating conidia were also observed within the colonised xylem vessel. The fungus was not observed in sections of the petiole from Sicot (Table S3).
3.3. Development of a defoliating V. dahliae strain expressing mCherry
Towards the aim of being able to compare and contrast the infection of cotton with different clonal lineages of V. dahliae we developed a strain of V. dahliae from VCG 1A that expressed mCherry fluorescent protein. Agrobacterium transformation yielded a total of 53 transformant colonies of V. dahliae strain Vd71181, and their red fluorescence was confirmed using confocal microscopy. Thirty-six of these were isolated from plates transformed with the AGL1 Agrobacterium strain while the rest were obtained on plates transformed with the EHA105 strain. Five isolates were selected for further analysis based on uniformity in colony morphology, spore size, and the level of fluorescence (Table S4, Figure 3A-C). Macroscopic morphology of the five transformants was consistent with the parent strain Vd71181 (Figure 3A). Both parental and transformant strains produced hyaline spores, cylindrical to ovate in shape, and approximately 5µm in length (Figure 3B). There was no significant differences (p = 0.101) in growth rates between Vd71181 and the transformants (Figure 3D). Quantification of total mCherry fluorescence in each transformant shows that 81T0069 had higher intensity (p < 0.001) in fluorescence than the rest (Figure 3E). There was no significant difference (p = 0.821) in fecundity between the parent Vd71181 and 81T00069 (Figure 3F). Another transformant, 81T0073 had a lower fecundity than both Vd71181 and 81T0069 (p < 0.001).
Disease severity in the form of leaf chlorosis, necrosis, and defoliation, and plant height of cotton seedlings were scored using a Likert rating scale of 0 to 5 (Table 3) for V. dahliae induced symptoms (Figure 4).
Sicot seedlings were root-inoculated with the parent Vd71181 isolate and transformants 81T0069 and 81T0073. Four weeks post inoculation, all V. dahliae-inoculated plants had a significantly higher mean disease rating (p < 0.001) compared to the water only inoculated negative control (Figure 5A). Internal red-brown discolouration was observed in the stem vasculature of plants inoculated with the transformants or the parent strain but was absent in the uninoculated plants (Figure 5B). Fungal colonies reisolated from plant tissues were white and resembled V. dahliae (Figure 5C). This was further confirmed by ITS PCR and sequencing on Vd71181 (Table S2) or detection of fluoresence in the transformed isolates. The parent Vd71181 strain was reisolated from 62.5% (n=24) of inoculated seedlings. The 81T0069 and 81T0073 transformants were reisolated from 45% (n=20) and 66.7% (n=21) of inoculated seedlings, respectively. V. dahliae was not recovered from any of the negative control plants inoculated with water only. All V. dahliae-inoculated plants had a significantly higher mean disease rating compared to the uninoculated plants (p < 0.001) (Figure 5D). There was no significant difference (p = 0.076) in the disease rating between seedlings inoculated with the Vd71181 parent isolate (mean score with standard error of the mean = 3.2 ± 0.12, n=24) and plants inoculated with either of the transformants 81T0069 (3.3 ± 0.23, n=20) (p = 0.887) or 81T0073 (3.9 ± 0.19, n=21) (Figure 5D). This suggests that the virulence of the transformants on cotton plants was not altered.
To determine if the defoliating strain of V. dahliae colonised cotton plants in a manner similar to the non-defoliating strain, longitudinal sections of the stem of Sicot inoculated with 81T0069 were dissected 28 days after inoculation and visualised under a confocal microscope. mCherry-tagged mycelia were clearly present in the xylem tissues of Sicot plants (Figure 6A-C) and colonising the root cortex (Figure 6A). A magnified view suggested the mycelia were within the parenchyma cells adjacent to the xylem in this region (Figure 6D-F). Although a side-by-side comparison has not been made, the colonisation process appeared similar for both the defoliating and non-defoliating strains.
3.4. Both defoliating and non-defoliating V. dahliae can colonise weed species and cause disease in Nicotiana benthamiana
V. dahliae transformant strains of 81T0069 (VCG 1A) and 71T0003 (VCG 2A) were used to inoculate seven common weed species found in Australian fields and the native tobacco N. benthamiana. At 4 weeks post inoculation, plants were visually examined (Figure 7). There were no noticeable differences in the size of the plants, the colour of the leaves, and stem height between 71T0003 or 81T0069 inoculated weeds and their respective uninoculated controls (Figure 7A-G). This was reflected in the disease severity ratings (Figure 8A). All weed species showed minimal symptoms (Figure 8A). C. bonariensis showed slightly elevated leaf yellowing on uninoculated plants and on plants inoculated with 81T0069 (Figure 8A). However, no V. dahliae was reisolated from the uninoculated controls and 81T0069 was detected in only one out of 26 C. bonariensis sampled, suggesting that the slight leaf yellowing of C. bonariensis was likely not due to the presence of V. dahliae (Figure 8A, Table 5). On the other hand, significantly elevated disease severity was detected (p < 0.05) in inoculated versus uninoculated N. benthamiana plants (Figure 8A). The impact of the inoculum on N. benthamiana was evident, with severe stunting observed on N. benthamiana plants inoculated with either 71T0003 or 81T0069 strain (Figure 7H).
Defoliating and non-defoliating V. dahliae transformants were reisolated from three and five weed species, respectively (Table 5). Where fungal colonies were recovered from C. bonariensis, S. oleraceus, C. virgata, E. colona, and U. panicoides, all were confirmed to be either 71T0003 or 81T0069 by fluorescence under confocal microscopy. V. dahliae was not reisolated from any of the uninoculated plants. The recovery rate of transformant 71T0003 and 81T0069 across all weeds (n = 190) were 17.4% and 8.9%, respectively (Table 5). The highest recovery percentage of the non-defoliating 71T0003 within weed species was 24% (n = 25) from U. panicoides belonging to the Poaceae family and 15.4% (n = 26) from Conyza bonariensis L. belonging to the Asteraceae family. The recovery rate of 81T0069 from the weeds was relatively low when compared to that of 71T0003, with a 7.7% recovery rate of the fungus from E. colona plants (Poaceae) being the highest (Table 5).
Recovery rates of 71T0003 (61.5%) and 81T0069 (50%) from stem sections of N. bethamiana was much higher than that for the weed species. Consistent with this observation was the detection of tagged isolates of V. dahliae in the stems or roots of N. benthamiana (Figure 8B-E). These observations suggest that N. benthamiana is a susceptible plant host for V. dahliae.
4. Discussion
Verticillium wilt is a major disease for the Australian cotton industry. In Australia, it is generally considered that the non-defoliating pathotypes of VCG 4B and the more recently detected VCG 2A are the prevalent disease-causing strains on cotton fields in Australia [7,27]. Recent field incidences of Verticillium wilt within the last decade have been low but were observed to be rising steadily in successive seasons [28]. This increased occurrence and the detection of VCG 1A pathotype from the NSW DPI culture collection [28] has raised some concerns about the cause of increased disease severity in the field. It is known that VCG 2A can infect weeds commonly found in cotton fields [30]. Whether its adaptation to survive on other plant species is what makes it the dominant pathotype in Australia is not clearly understood. However, the defoliating VCG 1A is not so widespread in Australia as it has been overseas in causing crop losses and complete defoliation of infected cotton plants [7]. This study addresses the pathogenicity of these two VCGs and paves the way for the evolution of these populations to be dissected and to aid the practical management of this disease in the Australian cotton industry.
Here we have used transformation of V. dahliae strains with gene cassettes to encode the expression of fluorescent proteins. This has allowed not only the pathogen to be tracked within the plant but also facilitated recovery of the pathogens from various hosts, including those that are non-symptomatic. The GFP-expressing VCG 2A and mCherry-expressing VCG 1A inoculated plants showed similar levels of disease severity when compared to their respective wildtype parents, indicating that the transformation did not alter the virulence of these strains on cotton.
The localisation of the GFP-expressing VCG 2A was visualised in Siokra and Sicot cotton plants during a period of 7 days post inoculation to study the early infection process. Conidia were observed on the root tips of both cultivars at 4 hpi. At 24 hpi, germ tubes were visible on approximately 50% of conidia observed on both cultivars. This is comparable to the germination timing of a GFP-expressing V. dahliae on lettuce, first observed at 12 to 48 h following inoculation [44]. Germination of conidia as early as 2 hpi on cotton has been reported [61]. A narrow infection peg was observed on the surface of the root tip at 24 dpi, with hyphal swelling appearing to narrow after penetration. While infection structures of V. dahliae in the form of appressoria were observed in penetrating the root surface of lettuce and fiber flax [44,62], it has not been observed in other plant species such as Nicotiana benthamiana [41]. V. dahliae only displayed slight hyphal swelling without a penetration peg observed before infection in oilseed rape and sunflower [43,63]. However, a cotton derived V. dahliae isolate showed slight hyphal swelling, followed by a narrow penetration peg on Arabidopsis roots [64]. This appears to be required for the isolate to breach the cell wall of cotton root epidermis during the initial colonisation [65]. This is consistent with our observations in this study.
The root tip was colonised by the fungus and its intercellular movement through the vascular tissues was evident at 5 dpi. This confirmed that the mechanism of Verticillium spp. infection is through establishing successful colonisation of the vascular tissues, particularly the xylem elements [61,66]. This also confirmed that root tips are sites of penetration for V. dahliae on cotton hosts [61,67].
At 7 dpi, advanced mycelia and mycelial networks became apparent. V. dahliae mycelia was mostly confined to the individual xylem vessels of the vasculature, with longitudinal movement in the xylem and the perforating tracheary elements. Colonisation of lateral root junctions was observed, as reported in a previous study [61]. Conidia and mycelia were detected in the stem and petiole of Siokra but not in Sicot. Above-ground colonisation by V. dahliae, specifically in the petiole base has been previously reported [61], although it was detected at 30 days post inoculation, using a virulent non-defoliating GFP-expressing isolate of V. dahliae. At 7 dpi, an intense fluorescence signal in the xylem of Sicot was identified as occlusion due to conidia within a xylem vessel. Vascular occlusions by fungal pathogens are often associated with the formation of plant structures such as tyloses to inhibit the movement of the fungus inside the host. Such occlusions were typically observed as densely clustered conidia in the tracheid of oilseed rape [63]. But sometimes occlusions can lead to the blockage of xylem vessels and instigate the classic wilt symptoms [67]. Restricting xylem vessels colonised by V. dahliae in the lateral roots was identified as an important response in wilt resistant lettuce cultivars [44]. Similarly, the cotton interaction observed here could point toward the restriction of the fungus at the border pit membranes of the xylem [8].
The typical defoliating symptoms were induced on cotton by the mCherry expressing VCG 1A isolate. The infection process through the vasculature appears similar to the non-defoliating strain. In another study, both defoliating and non-defoliating isolates recovered from the same stem showed comparable levels of virulence when cotton plants were inoculated with either or both isolates in a pot trial [53]. Although the detection of a defoliating pathotype even at a relatively low frequency compared to the nondefoliating pathotype complicates the landscape for disease management in the Australian cotton fields [53].
Mycelial networks with mCherry fluorescence were clearly visualised in the xylem vessels of plants inoculated with the defoliating strain. Interestingly, movement of mycelia were observed in the cortex region. However, the pattern of colonisation suggests that it was moving along the surface or in between cell layers of the cortex and endosperm. Unlike other hosts, intracellular colonisation was observed rarely in V. dahliae localisation studies on cotton and Arabidopsis [61,64]. Both intra- and intercellular movement of hyphae through the endoderm were proposed [68], however the movement from cortical cells into xylem vessels was observed in an intercellular manner [64].
The emergence and widespread prevalence of the Australian VCG 2A strain has prompted investigation into the capacity for Australian V. dahliae isolates to colonise common Australian hosts. Prior reports propose that VCG 2A may predominate in Australian cotton fields through its ability to infect and multiply in associated weed species [7]. The number of weeds colonised and their reisolation frequencies were higher when comparing VCG 2A and VCG 1A from the weeds selected here (Table 5), suggesting that VCG 2A clonal lineage are potentially better colonisers of weeds than the VCG 1A lineage. However, further analysis with more weed species and replications will need to be performed to confirm these observations. This highlights the need to investigate in much higher detail the local species, along with field cropping history, as potential reservoirs for V. dahliae VCG 2A in Australian environments.
V. dahliae was isolated from six of the eight species investigated, including the known host, Nicotiana benthamiana [41]. N. benthamiana, a native Australian relative of tobacco, has been used as a model species to understand V. dahliae infection in previous studies [41,69,70]. However, there is limited literature describing N. benthamiana susceptibilities to different V. dahliae VCGs. The study at hand re-isolated transformants 81T0069 and 71T0003 from N. benthamiana plants, suggesting that this species is a host for both V. dahliae VCGs 1A and 2A. It was also found that the VCG 1A transformant, 81T0069, induced higher severity of disease symptoms and frequency of plant death than VCG 2A transformant, 71T0003. These findings therefore provide a deeper insight into V. dahliae interactions with the model species N. benthamiana.
To our knowledge, this is the first report of Chloris virgata and Urochloa panicoides as hosts of V. dahliae. This is also the first description of V. dahliae infection of Conyza bariensis and Echinochloa colona, although closely related members within these genera have been reported as hosts of the pathogen elsewhere [32,71]. Reports on the status of Sonchus oleraceus as a host for V. dahliae are conflicting [30,72,73,74,75]. In the present study, S. oleraceus was found to be infected with V. dahliae 71T0003 at low frequencies (8%). This concurs with previous reports of VCG 2A infecting S. oleraceus internationally [71], as well as V. dahliae infecting this species in Australia [32].
V. dahliae could not be isolated from the native species Chloris truncata or from the globally distributed species Avena fatua. Conflictingly, A. fatua has previously been reported as a host of V. dahliae at low frequencies in olive orchards in Greece [73]. Furthermore, V. dahliae VCG 2A specifically has been associated with A. fatua in North American regions of potato production and has been observed to produce microsclerotia on host tissue [71]. However, given the low frequency of V. dahliae observed in these studies, it is unsurprising that V. dahliae was not isolated from A. fatua in the study at hand. Further investigation is needed to understand whether A. fatua is a susceptible host to V. dahliae in an Australian context. Indeed, in this study we have only used a single collection of all weed species so have not in any way assessed any standing host variation for interactions with V. dahliae. Monocotyledonous species are otherwise generally considered to be resistant to Verticillium wilt disease, however they may remain susceptible to endophytic colonisation by the fungus, which is symptomless [76]. Consistent with the literature, the study at hand found that infection by V. dahliae was asymptomatic for all weeds belonging to the Poaceae family. Similar results have been described previously in other grass species [77]. In S. oleraceus, a dicotyledonous weed, there was no difference in disease scores between inoculated and uninoculated groups, which also indicates asymptomatic colonisation of this species. This finding was consistent with other studies on V. dahliae infection of S. oleraceus in fields overseas [71].
V. dahliae 71T0003 and 81T0069 were both re-isolated from the dicotyledonous species C. bonariensis. Some of the C. bonariensis seedlings were exhibiting signs of stress throughout the duration of the experiment, which may explain the elevated disease scores of the uninoculated control plants and plants inoculated with 81T0069. However, tissue reisolations detected none or minimal presence of V. dahliae suggesting that V. dahliae was likely not the cause of leaf yellowing observed on these plants. Nevertheless, this warrants further investigation into whether C. bonariensis is susceptible to Verticillium wilt.
It is important to note that whilst this study identifies species that are susceptible to infection by V. dahliae VCGs 1A and 2A, it does not investigate the capacity of these hosts to increase pathogen inoculum levels in the field. V. dahliae microsclerotia are the primary fungal propagules that persist in soils and act as carry-over inoculum into subsequent cropping seasons [78]. Therefore, quantifying microsclerotia production that occurs in plant hosts is important for Verticillium wilt disease prediction and management [79]. Prior studies have demonstrated the potential for weed species to be infected by V. dahliae without increasing the microsclerotia inoculum load [32,78]. Evans (1971) isolated V. dahliae from the weed S. oleraceus in Australian cotton fields, however concluded the species as a non-host to V. dahliae because infection was confined to the plant roots and did not result in the production of microsclerotia. On the other hand, a study on the monocotyledonous weed A. fatua found that the species was host to a tomato-pathogenic V. dahliae strain from North America, and that production of microsclerotia was higher in this species compared to other local weed hosts [71]. Consequently, further investigations into the capacity of local species to increase inoculum load of V. dahliae will help to shape future Verticillium wilt management approaches.
5. Conclusions
Crop production regions around the globe are afflicted by different VCGs and pathotypes of V. dahliae owing to their genetic diversity and host range. Mounting evidence is suggesting that the pathogen is evolving and adapting to different geographical locations [14]. In Australia, the traditional landscape of Verticillium management needs to adapt to control the disease accelerated by the presence of both the non-defoliating and defoliating strains of V. dahliae. Fluorescently tagged V. dahliae not only provides a useful tool to gain deeper understanding of epidemiology of the infection process in cotton but also allows its survival on alternative plant hosts to be explored.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Figure S1: Anatomy of a cotton seedling approximately 10 -14 days after sowing; Table S1: Primers for yeast recombinantion-based cloning of to generate a plasmid for expression of mCherry in Verticillium dahliae; Table S2: Verticillium dahliae-specific primers [55] amplifying a 200 bp ITS product were used to confirm its identity; Table S3: Summary of rate of colonisation based on timing of initial observation at each infection stage throughout the confocal microscopy experiment; Table S4: mCherry transformant isolates selected for comparison against Verticillium dahliae VCG 1A parent, Vd71181, that originated from Gwydir Valley, NSW.
Author Contributions
Conceptualization, E.A.B.A. and D.M.G.; methodology, E.A.B.A. and D.M.G.; validation, S.M., A.G. and A.C.; formal analysis, S.M., A.G. and A.C.; investigation, S.M., A.G. and A.S.U.; resources, D.P.L., L.J.S. and A.S.U.; data curation, S.M. and A.G.; writing—original draft preparation, A.C.; writing—review and editing, S.M., A.G., D.P.L., A.S.U., L.J.S., E.A.B.A., D.M.G. and A.C.; supervision, D.M.G. and E.A.B.A.; project administration, D.M.G. and E.A.B.A.; funding acquisition, D.M.G. and E.A.B.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Australian Research Council Research Hub for Sustainable Crop Protection (project number IH190100022) and funded by the Australian Government. This research was also supported by the Cotton Research and Development Corporation through funding from the Australian Government Department of Agriculture as part of its Rural R&D for Profit program and the NSW Department of Primary Industries, grant number ‘DAN 1809’. Pathogen isolation and preliminary identification was supported by the Cotton Research and Development Corporation and the Australian Government Department of Agriculture as part of its Rural R&D for Profit program (project number RRDP1724).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Acknowledgments
We kindly thank Professor Rafael M. Jiménez-Díaz from the University of Córdoba for performing the complementation tests to confirm the VCG 2A and the non-defoliating pathotype of Vd71171. We also thank Dr Buddhi Dayananda from the University of Queensland for advice on the statistical analysis performed in this study.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Oosterhuis, D.M. Growth and Development of a Cotton Plant. In Nitrogen Nutrition of Cotton: Practical Issues; 1990; pp. 1-24.
- Rathore, K.S.; Sunilkumar, G.; Campbell, L.M. Cotton (Gossypium hirsutum L.). In Agrobacterium Protocols; Wang, K., Ed.; Humana Press: Totowa, NJ, 2006; pp. 267–280. [Google Scholar]
- Cope, R.B. Chapter 68 - Cottonseed Toxicity. In Veterinary Toxicology (Third Edition), Gupta, R.C., Ed.; Academic Press: 2018; pp. 967-980.
- Applequist, W.L.; Cronn, R.; Wendel, J.F. Comparative development of fiber in wild and cultivated cotton. Evolution & Development 2001, 3, 3–17. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, J.; Fang, L.; Zhang, Z.; Ma, W.; Niu, Y.; Ju, L.; Deng, J.; Zhao, T.; Lian, J.; et al. Gossypium barbadense and Gossypium hirsutum genomes provide insights into the origin and evolution of allotetraploid cotton. Nature Genetics 2019, 51, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Cotton Industry Australia Overview. Available online: https://cottonaustralia.com.au/industry-overview (accessed on 15 February 2024).
- Dadd-Daigle, P.; Kirkby, K.; Chowdhury, P.R.; Labbate, M.; Chapman, T.A. The Verticillium wilt problem in Australian cotton. Australasian Plant Pathology 2021, 50, 129–135. [Google Scholar] [CrossRef]
- Fradin, E.F.; Thomma, B.P.H.J. Physiology and molecular aspects of Verticillium wilt diseases caused by V. dahliae and V. albo-atrum. Molecular Plant Pathology 2006, 7, 71–86. [Google Scholar] [CrossRef]
- Wheeler, D.L.; Johnson, D.A. Verticillium dahliae infects, alters plant biomass, and produces inoculum on rotation crops. Phytopathology 2016, 106, 602–613. [Google Scholar] [CrossRef]
- Short, D.P.; Sandoya, G.; Vallad, G.E.; Koike, S.T.; Xiao, C.L.; Wu, B.M.; Gurung, S.; Hayes, R.J.; Subbarao, K.V. Dynamics of verticillium species microsclerotia in field soils in response to fumigation, cropping patterns, and flooding. Phytopathology 2015, 105, 638–645. [Google Scholar] [CrossRef]
- Requena-Mullor, J.M.; García-Garrido, J.M.; García, P.A.; Rodríguez, E. Climatic drivers of Verticillium dahliae occurrence in Mediterranean olive-growing areas of southern Spain. PLoS One 2020, 15, e0232648. [Google Scholar] [CrossRef]
- Martín-Sanz, A.; Rueda, S.; García-Carneros, A.B.; González-Fernández, S.; Miranda-Fuentes, P.; Castuera-Santacruz, S.; Molinero-Ruiz, L. Genetics, host range, and molecular and pathogenic characterization of Verticillium dahliae from sunflower reveal two differentiated groups in Europe. Frontiers in Plant Science 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Berlander, I.; Powelson, M.L. Verticillium wilt. The Plant Health Instructor. 2000. [Google Scholar] [CrossRef]
- Klimes, A.; Dobinson, K.F.; Thomma, B.P.H.J.; Klosterman, S.J. Genomics spurs rapid advances in our understanding of the biology of vascular wilt pathogens in the genus Verticillium. Annual Review of Phytopathology 2015, 53, 181–198. [Google Scholar] [CrossRef] [PubMed]
- Holman, S.; Kirkby, K.; Smith, L.; Kartnett, H. Vert update: The latest in vert research. CottonInfo. Available online: https://www.cottoninfo.com.au/publications/disease-vert-update-latest-verticillium-research (accessed on 15 February 2024).
- Klosterman, S.J.; Atallah, Z.K.; Vallad, G.E.; Subbarao, K.V. Diversity, pathogenicity, and management of Verticillium species. Annu Rev Phytopathol 2009, 47, 39–62. [Google Scholar] [CrossRef] [PubMed]
- Mol, L.; Termorshuizen, A.J. Life cycle and ecology of Verticillium dahliae in potato. In Potato Ecology And modelling of crops under conditions limiting growth: Proceedings of the Second International Potato Modeling Conference, Wageningen, 1994, Haverkort, A.J., MacKerron, D.K.L., Eds.; Springer Netherlands: Dordrecht. 1995; pp. 251–263. [Google Scholar]
- Toussoun, T.A. Root Diseases and Soil-Borne Pathogens; University of California Press: Berkeley, 1970. [Google Scholar]
- Bellahcene, M.; Assigbetsé, K.; Fortas, Z.; Geiger, J.-P.; Nicole, M.; Fernandez, D. Genetic diversity of Verticillium dahliae isolates from olive trees in Algeria. Phytopathologia Mediterranea 2005, 44, 266–274. [Google Scholar]
- Baroudy, F.; Putman, A.I.; Habib, W.; Puri, K.D.; Subbarao, K.V.; Nigro, F. Genetic diversity of Verticillium dahliae populations from olive and potato in Lebanon. Plant Disease 2019, 103, 656–667. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Díaz, R.M.; Mercado-Blanco, J.; Olivares-García, C.; Collado-Romero, M.; Bejarano-Alcázar, J.; Rodríguez-Jurado, D.; Giménez-Jaime, A.; García-Jiménez, J.; Armengol, J. Genetic and virulence diversity in Verticillium dahliae populations infecting artichoke in Eastern-Central Spain. Phytopathology 2006, 96, 288–298. [Google Scholar] [CrossRef]
- Korolev, N.; Katan, J.; Katan, T. Vegetative compatibility groups of Verticillium dahliae in Israel: their distribution and association with pathogenicity. Phytopathology 2000, 90, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Artés, E.; García-Pedrajas, M.D.; Bejarano-Alcázar, J.; Jiménez-Díaz, R.M. Differentiation of cotton-defoliating and nondefoliating pathotypes of Verticillium dahliae by RAPD and specific PCR analyses. European Journal of Plant Pathology 2000, 106, 507–517. [Google Scholar] [CrossRef]
- Leslie, J.F. Fungal vegetative compatibility. Annual Review of Phytopathology 1993, 31, 127–150. [Google Scholar] [CrossRef]
- Collado-Romero, M.; Mercado-Blanco, J.; Olivares-García, C.; Jiménez-Díaz, R.M. Phylogenetic analysis of Verticillium dahliae vegetative compatibility groups. Phytopathology 2008, 98, 1019–1028. [Google Scholar] [CrossRef]
- Milgroom, M.G.; Jiménez-Gasco, M.d.M.; Olivares García, C.; Drott, M.T.; Jiménez-Díaz, R.M. Recombination between clonal lineages of the asexual fungus Verticillium dahliae detected by genotyping by sequencing. PLOS ONE 2014, 9, e106740. [Google Scholar] [CrossRef]
- Chapman, T.A.; Chambers, G.A.; Kirkby, K.; Jiménez-Díaz, R.M. First report of the presence of Verticillium dahliae VCG1A in Australia. Australasian Plant Disease Notes 2016, 11, 13. [Google Scholar] [CrossRef]
- Smith, L.; Scheikowski, L.; Bauer, B.; Lehane, J.; Allen, S. Detection of new pathogens in Australian cotton. Available online: https://www.insidecotton.com/sites/default/files/article-files/poster_SMITH__Detection_of_new_pathogens_in_Australian_cotton.pdf (accessed on 15 February 2024).
- Dervis, S.; Kurt, S.; Soylu, S.; Erten, L.; Mine Soylu, E.; Yıldız, M.; Tok, F.M. Vegetative compatibility groups of Verticillium dahliae from cotton in the southeastern anatolia region of Turkey. Phytoparasitica 2008, 36, 74–83. [Google Scholar] [CrossRef]
- Yildiz, A.; Doğan, M.N.; Boz, Ö.; Benlioğlu, S. Weed hosts of Verticillium dahliae in cotton fields in Turkey and characterization of V. dahliae isolates from weeds. Phytoparasitica 2009, 37, 171–178. [Google Scholar] [CrossRef]
- Keykhasaber, M.; Thomma, B.P.H.J.; Hiemstra, J.A. Verticillium wilt caused by Verticillium dahliae in woody plants with emphasis on olive and shade trees. European Journal of Plant Pathology 2018, 150, 21–37. [Google Scholar] [CrossRef]
- Evans, G. Influence of weed hosts on the ecology of Verticillium dahliae in newly cultivated areas of the Namoi Valley, New South Wales. Annals of Applied Biology 1971, 67, 169–175. [Google Scholar] [CrossRef]
- Göre, M.E.; Erdoğan, O.; Caner, Ö.K.; Aydın, M.H.; Berk, S. VCG diversity and virulence of Verticillium dahliae from commercially available cotton seed lots in Turkey. European Journal of Plant Pathology 2014, 140, 689–699. [Google Scholar] [CrossRef]
- Smith, J.L. Improving the management of cotton diseases in Australian cotton farming systems. CRDC funded project RRDP1724. 2016. [Google Scholar]
- Eshel, D.; Beno-Moualem, D.; Lorang, J.M.; Dinoor, A.; Prusky, D. Induction of peroxidase during infection of unripe persimmon fruit by Alternaria alternata: a possible quiescence mechanism. Journal of Phytopathology 2002, 150, 357–362. [Google Scholar] [CrossRef]
- M., L.J.; P., T.R.; P., M.J.; L., S.T.; S., R.R.; A., R.J.; J., W.T.; B., J.K.; J., R.R.; B., D.M.; et al. Green fluorescent protein is lighting up fungal biology. Applied and Environmental Microbiology 2001, 67, 1987–1994. [CrossRef]
- Spellig, T.; Bottin, A.; Kahmann, R. Green fluorescent protein (GFP) as a new vital marker in the phytopathogenic fungus Ustilago maydis. Molecular and General Genetics 1996, 252, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, O.; Johnson, F.H.; Saiga, Y. Extraction, purification and properties of aequorin, a bioluminescent protein from the luminous hydromedusan, Aequorea. Journal of Cellular and Comparative Physiology 1962, 59, 223–239. [Google Scholar] [CrossRef] [PubMed]
- Shaner, N.C.; Campbell, R.E.; Steinbach, P.A.; Giepmans, B.N.; Palmer, A.E.; Tsien, R.Y. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat Biotechnol 2004, 22, 1567–1572. [Google Scholar] [CrossRef]
- Li, F.; Bibi, N.; Fan, K.; Ni, M.I.; Yuan, S.; Wang, M. Agrobacterium-mediated transformation of Verticillium dahliae with GFP gene to study cotton-pathogen interaction using a novel inoculation method. Pakistan Journal of Botany 2016, 48. [Google Scholar]
- Su, X.; Lu, G.; Rehman, L.; Li, X.; Sun, L.; Guo, H.; Cheng, H. mCherry-labeled Verticillium dahliae could be utilized to investigate its pathogenicity process in Nicotiana benthamiana. Genes 2018, 9, 508. [Google Scholar] [CrossRef] [PubMed]
- Sabburg, R.; Gregson, A.; Urquhart, A.S.; Aitken, E.A.B.; Smith, L.; Thatcher, L.F.; Gardiner, D.M. A method for high throughput image based antifungal screening. bioRxiv, 2008. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, J.; Gao, J.; Zhang, G.; Yu, Y.; Zhou, H.; Chen, W.; Zhao, J. The Colonization process of sunflower by a green fluorescent protein-tagged isolate of Verticillium dahliae and its seed transmission. Plant Disease 2018, 102, 1772–1778. [Google Scholar] [CrossRef] [PubMed]
- Vallad, G.E.; Subbarao, K.V. Colonization of resistant and susceptible lettuce cultivars by a green fluorescent protein-tagged isolate of Verticillium dahliae. Phytopathology 2008, 98, 871–885. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Díaz, R.M.; Olivares-García, C.; Landa, B.B.; del Mar Jiménez-Gasco, M.; Navas-Cortés, J.A. Region-wide analysis of genetic diversity in Verticillium dahliae populations infecting olive in southern Spain and agricultural factors influencing the distribution and prevalence of vegetative compatibility groups and pathotypes. Phytopathology 2011, 101, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Mercado-Blanco, J.; Rodríguez-Jurado, D.; Parrilla-Araujo, S.; Jiménez-Díaz, R. Simultaneous detection of the defoliating and nondefoliating Verticillium dahliae pathotypes in infected olive plants by duplex, nested polymerase chain reaction. Plant Disease 2003, 87, 1487–1494. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, D.M.; Smith, L.J.; Rusu, A.; Aitken, E.B.A. The genomes of two Australian isolates of Verticillium dahliae reccovered from cotton fields. Australas. Plant Pathol. 2024. submitted. [Google Scholar]
- Gaspar, Y.M.; McKenna, J.A.; McGinness, B.S.; Hinch, J.; Poon, S.; Connelly, A.A.; Anderson, M.A.; Heath, R.L. Field resistance to Fusarium oxysporum and Verticillium dahliae in transgenic cotton expressing the plant defensin NaD1. Journal of Experimental Botany 2014, 65, 1541–1550. [Google Scholar] [CrossRef]
- Gardiner, D.M.; Howlett, B.J. Negative selection using thymidine kinase increases the efficiency of recovery of transformants with targeted genes in the filamentous fungus Leptosphaeria maculans. Current Genetics 2004, 45, 249–255. [Google Scholar] [CrossRef]
- Urquhart, A.S.; Mondo, S.J.; Mäkelä, M.R.; Hane, J.K.; Wiebenga, A.; He, G.; Mihaltcheva, S.; Pangilinan, J.; Lipzen, A.; Barry, K.; et al. Genomic and genetic insights into a cosmopolitan fungus, Paecilomyces variotii (Eurotiales). Frontiers in Microbiology 2018, 9. [Google Scholar] [CrossRef]
- Zhang, J.; Fang, H.; Zhou, H.; Sanogo, S.; Ma, Z. Genetics, breeding, and marker-assisted selection for Verticillium wilt resistance in cotton. Crop Science 2014, 54, 1289–1303. [Google Scholar] [CrossRef]
- Cirulli, M.; Ciccarese, F.; Amenduni, M. Progress in the search for Verticillium wilt-resistant eggplant. Phytopathologia Mediterranea 1990, 29, 184–190. [Google Scholar]
- Le, D.P.; Gregson, A.; Tran, T.T.; Jackson, R. Co-occurrence of defoliating and non-defoliating pathotypes of Verticillium dahliae in field-grown cotton plants in New South Wales, Australia. Plants 2020, 9, 750. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.-H.; Yeh, Y.-L.; Shen, W.-C. Fast preparation of fungal DNA for PCR screening. Journal of Microbiological Methods 2011, 85, 170–172. [Google Scholar] [CrossRef] [PubMed]
- Lievens, B.; Brouwer, M.; Vanachter, A.C.R.C.; Cammue, B.P.A.; Thomma, B.P.H.J. Real-time PCR for detection and quantification of fungal and oomycete tomato pathogens in plant and soil samples. Plant Science 2006, 171, 155–165. [Google Scholar] [CrossRef]
- R Core Development Team 2019. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. (accessed on 15 February 2024).
- Singmann, R.L.H.; Love, J.; Buerkner, P.; Herve, M. Estimated Marginal Means, aka Least-Squares Means. CRAN. Available online: https://github.com/rvlenth/emmeans (accessed on 15 February 2024).
- Hervé, M. Testing and plotting procedures for biostatistics. CRAN. Available online: https://cran.r-project.org/web/packages/RVAideMemoire/index.html (accessed on 15 February 2024).
- Kleiber, C.; Zeileis, A. Applied econometrics with R. Springer-Verlag, New York. Available online: https://CRAN.R-project.org/package=AER (accessed on 15 February 2024).
- Wickham, H. ggplot2: Elegant graphics for data analysis. Springer-Verlag New York. Available online: https://ggplot2.tidyverse.org (accessed on 15 February 2024).
- Zhang, W.-W.; Jiang, T.-F.; Cui, X.; Qi, F.-J.; Jian, G.-L. Colonization in cotton plants by a green fluorescent protein labelled strain of Verticillium dahliae. European Journal of Plant Pathology 2013, 135, 867–876. [Google Scholar] [CrossRef]
- Blum, A.; Bressan, M.; Zahid, A.; Trinsoutrot-Gattin, I.; Driouich, A.; Laval, K. Verticillium wilt on fiber flax: symptoms and pathogen development in planta. Plant Disease 2018, 102, 2421–2429. [Google Scholar] [CrossRef] [PubMed]
- Eynck, C.; Koopmann, B.; Grunewaldt-Stoecker, G.; Karlovsky, P.; von Tiedemann, A. Differential interactions of Verticillium longisporum and V. dahliae with Brassica napus detected with molecular and histological techniques. European Journal of Plant Pathology 2007, 118, 259–274. [Google Scholar] [CrossRef]
- Zhao, P.; Zhao, Y.L.; Jin, Y.; Zhang, T.; Guo, H.S. Colonization process of Arabidopsis thaliana roots by a green fluorescent protein-tagged isolate of Verticillium dahliae. Protein Cell 2014, 5, 94–98. [Google Scholar] [CrossRef]
- Zhao, Y.L.; Zhou, T.T.; Guo, H.S. Hyphopodium-specific VdNoxB/VdPls1-dependent ROS-Ca2+ signaling is required for plant infection by Verticillium dahliae. PLoS Pathog 2016, 12, e1005793. [Google Scholar] [CrossRef]
- Bishop, C.D.; Cooper, R.M. An ultrastructural study of root invasion in three vascular wilt diseases. Physiological Plant Pathology 1983, 22, 15–IN13. [Google Scholar] [CrossRef]
- Pegg, G.F.; Brady, B.L. Verticillium Wilts; CABI Pub.: 2002.
- Garber, R.; Houston, B. Penetration and development of Verticillium alboatrum in the cotton plant. 1967.
- Faino, L.; de Jonge, R.; Thomma, B.P.H.J. The transcriptome of Verticillium dahliae-infected Nicotiana benthamiana determined by deep RNA sequencing. Plant Signaling & Behavior 2012, 7, 1065–1069. [Google Scholar] [CrossRef]
- Yin, Z.; Wang, N.; Pi, L.; Li, L.; Duan, W.; Wang, X.; Dou, D. Nicotiana benthamiana LRR-RLP NbEIX2 mediates the perception of an EIX-like protein from Verticillium dahliae. Journal of Integrative Plant Biology 2021, 63, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Frederick, Z.A.; Cummings, T.F.; Johnson, D.A. Susceptibility of weedy hosts from Pacific northwest potato production systems to crop-aggressive isolates of Verticillium dahliae. Plant Disease 2017, 101, 1500–1506. [Google Scholar] [CrossRef] [PubMed]
- Ligoxigakis, E.K.; Vakalounakis, D.J.; Thanassoulopoulos, C.C. Weed hosts ofVerticillium dahliae in crete: Susceptibility, symptomatology and significance. Phytoparasitica 2002, 30, 511–518. [Google Scholar] [CrossRef]
- Thanassoulopoulos, C.C.; Biris, D.A.; Tjamos, E.C. Weed hosts as inoculum source of Verticillium in olive orchards. Phytopathologia Mediterranea 1981, 20, 164–168. [Google Scholar]
- Vallad, G.E.; Bhat, R.G.; Koike, S.T.; Ryder, E.J.; Subbarao, K.V. Weedborne reservoirs and seed transmission of Verticillium dahliae in lettuce. Plant Disease 2005, 89, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Woolliams, G.E. Host range and symptomatology of Verticillium dahliae in economic, weed, and native plants in interrior British Columbia. Canadian Journal of Plant Science 1966, 46, 661–669. [Google Scholar] [CrossRef]
- Dung, J.K.S. Verticillium Wilt of Mint in the United States of America. Plants 2020, 9, 1602. [Google Scholar] [CrossRef]
- Krikun, J.; Bernier, C.C. Morphology of microsclerotia of Verticillium dahliae in roots of gramineous plants. Canadian Journal of Plant Pathology 1990, 12, 439–441. [Google Scholar] [CrossRef]
- Busch, L.; Smith, E.A.; Njoh-Elango, F. The effect of weeds on the value of rotation as a practical control for Verticillium wilt of potato. Canadian Plant Disease Survey 1978, 58, 61. [Google Scholar]
- Goud, J.C.; Termorshuizen, A.J. Quality of methods to quantify microsclerotia of Verticillium dahliae in soil. European Journal of Plant Pathology 2003, 109, 523–534. [Google Scholar] [CrossRef]
Figure 1.
The development and characterisation of a non-defoliating Verticillium dahliae strain of VCG 2A carrying GFP. (A) V. dahliae wild-type strain Vd71171 and the two GFP transformants derived from it, 71T0003 and 71T0006. (B) The green fluorescence of GFP is associated with the mycelia of 71T0003 imaged using BioTek Cytation 1 cell imaging reader. Scale bar = 200 µm. (C) Mean growth rate of the wildtype Vd71171 and the GFP transformants 71T0003 and 71T0006. Colony diameter (mm) is measured after 4 days of growth on potato dextrose plates (PDA). Median value (bar), lower and upper quartiles, minimum and maximum spore counts (whiskers) are shown. (D) Fecundity or spore concentration of Vd71171, 71T0003 and 71T0006 was determined using a hemocytomer in conidia per mL after spores were collected and then resuspended in 15mL of sterile water after 7 days of growth on PDA. (E) Siokra plants inoculated with wildtype Vd71171, 4 weeks post inoculation. Arrows indicate advanced necrosis spreading across the entire plant (left) and sometimes seen as severe wilting amongst healthy plants grown in the same pot (right). (F) Siokra plants inoculated with the GFP transformant 71T0003, 4 weeks post inoculation. Arrows indicate advanced necrosis on symptomatic plants (left) and a top-down view of a plant with necrosis spreading along the leaf margins (right). (G) Disease severity scored on Siokra and Sicot plants at 4 weeks post inoculation with either the wildtype Vd71171 or the GFP transformant 71T0003. Uninoculated plants served as negative controls. Least-squares means were estimated for each comparison using R package ‘emmeans’. Error bars indicate a 95% confidence interval. n = individual plants tested per isolate per cotton variety.
Figure 1.
The development and characterisation of a non-defoliating Verticillium dahliae strain of VCG 2A carrying GFP. (A) V. dahliae wild-type strain Vd71171 and the two GFP transformants derived from it, 71T0003 and 71T0006. (B) The green fluorescence of GFP is associated with the mycelia of 71T0003 imaged using BioTek Cytation 1 cell imaging reader. Scale bar = 200 µm. (C) Mean growth rate of the wildtype Vd71171 and the GFP transformants 71T0003 and 71T0006. Colony diameter (mm) is measured after 4 days of growth on potato dextrose plates (PDA). Median value (bar), lower and upper quartiles, minimum and maximum spore counts (whiskers) are shown. (D) Fecundity or spore concentration of Vd71171, 71T0003 and 71T0006 was determined using a hemocytomer in conidia per mL after spores were collected and then resuspended in 15mL of sterile water after 7 days of growth on PDA. (E) Siokra plants inoculated with wildtype Vd71171, 4 weeks post inoculation. Arrows indicate advanced necrosis spreading across the entire plant (left) and sometimes seen as severe wilting amongst healthy plants grown in the same pot (right). (F) Siokra plants inoculated with the GFP transformant 71T0003, 4 weeks post inoculation. Arrows indicate advanced necrosis on symptomatic plants (left) and a top-down view of a plant with necrosis spreading along the leaf margins (right). (G) Disease severity scored on Siokra and Sicot plants at 4 weeks post inoculation with either the wildtype Vd71171 or the GFP transformant 71T0003. Uninoculated plants served as negative controls. Least-squares means were estimated for each comparison using R package ‘emmeans’. Error bars indicate a 95% confidence interval. n = individual plants tested per isolate per cotton variety.
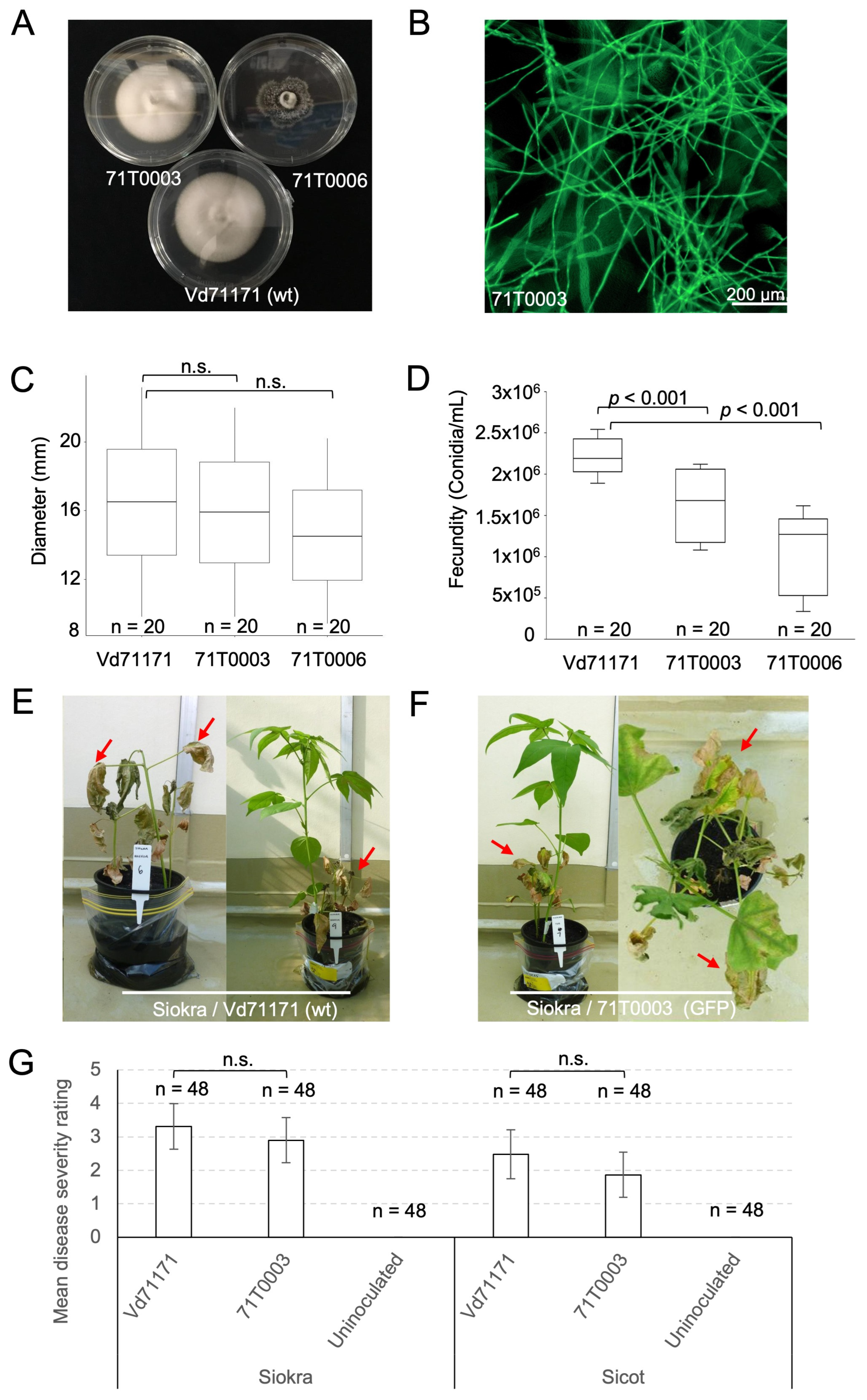
Figure 2.
Confocal laser scanning microscopy performed at 24 hpi, 1 dpi, 5 dpi and 7 dpi on Sicot and Siokra cotton cultivars inoculated with the GFP transformant 71T0003. (A) Conidia with germ tubes and hyphal elongation observed on the root tip of Sicot at 24 hpi. (B) Conidia and an infection peg observed on the root tip of Siokra at 24 hpi. Inset: magnified view of the infection peg under single-channel view. (C) Hyphal elongation and penetration into the root tip epidermis of Siokra at 24 hpi. (D) Mycelia growth on the root tip epidermis of Sicot at 5 dpi. (E) Mycelia visualised in the root cap (left) and at the base of the root tip (right) in Siokra at 5 dpi. (F) Intercellular movement of hyphae through lateral root epidermis on Siokra at 5 dpi. (G) Mycelia visualised in the xylem vessels of the main root in proximity to lateral root junctions in Sicot at 7 dpi. Circled area = free-moving spore observed in the xylem. (H) Mycelia visualised in an entire xylem vessel of the root vasculature in Sicot at 7 dpi. (I) Mycelia and conidia visualised in multiple xylem vessels of the root. Sites of vascular occlusion (o) were observed. (J) Single channel magnified view on the site of vascular occlusion in the xylem vessel densely packaged with conidia. (K) The movement of mycelia into the stem vasculature on Siokra at 7 dpi. (L) Presence of mycelia and free conidia was observed in the petiole of Siokra at 7 dpi. Inset = single channel view of the region containing mycelia and conidia (circled). m = mycelia, c = conidia. Scales are indicated by horizontal bars.
Figure 2.
Confocal laser scanning microscopy performed at 24 hpi, 1 dpi, 5 dpi and 7 dpi on Sicot and Siokra cotton cultivars inoculated with the GFP transformant 71T0003. (A) Conidia with germ tubes and hyphal elongation observed on the root tip of Sicot at 24 hpi. (B) Conidia and an infection peg observed on the root tip of Siokra at 24 hpi. Inset: magnified view of the infection peg under single-channel view. (C) Hyphal elongation and penetration into the root tip epidermis of Siokra at 24 hpi. (D) Mycelia growth on the root tip epidermis of Sicot at 5 dpi. (E) Mycelia visualised in the root cap (left) and at the base of the root tip (right) in Siokra at 5 dpi. (F) Intercellular movement of hyphae through lateral root epidermis on Siokra at 5 dpi. (G) Mycelia visualised in the xylem vessels of the main root in proximity to lateral root junctions in Sicot at 7 dpi. Circled area = free-moving spore observed in the xylem. (H) Mycelia visualised in an entire xylem vessel of the root vasculature in Sicot at 7 dpi. (I) Mycelia and conidia visualised in multiple xylem vessels of the root. Sites of vascular occlusion (o) were observed. (J) Single channel magnified view on the site of vascular occlusion in the xylem vessel densely packaged with conidia. (K) The movement of mycelia into the stem vasculature on Siokra at 7 dpi. (L) Presence of mycelia and free conidia was observed in the petiole of Siokra at 7 dpi. Inset = single channel view of the region containing mycelia and conidia (circled). m = mycelia, c = conidia. Scales are indicated by horizontal bars.
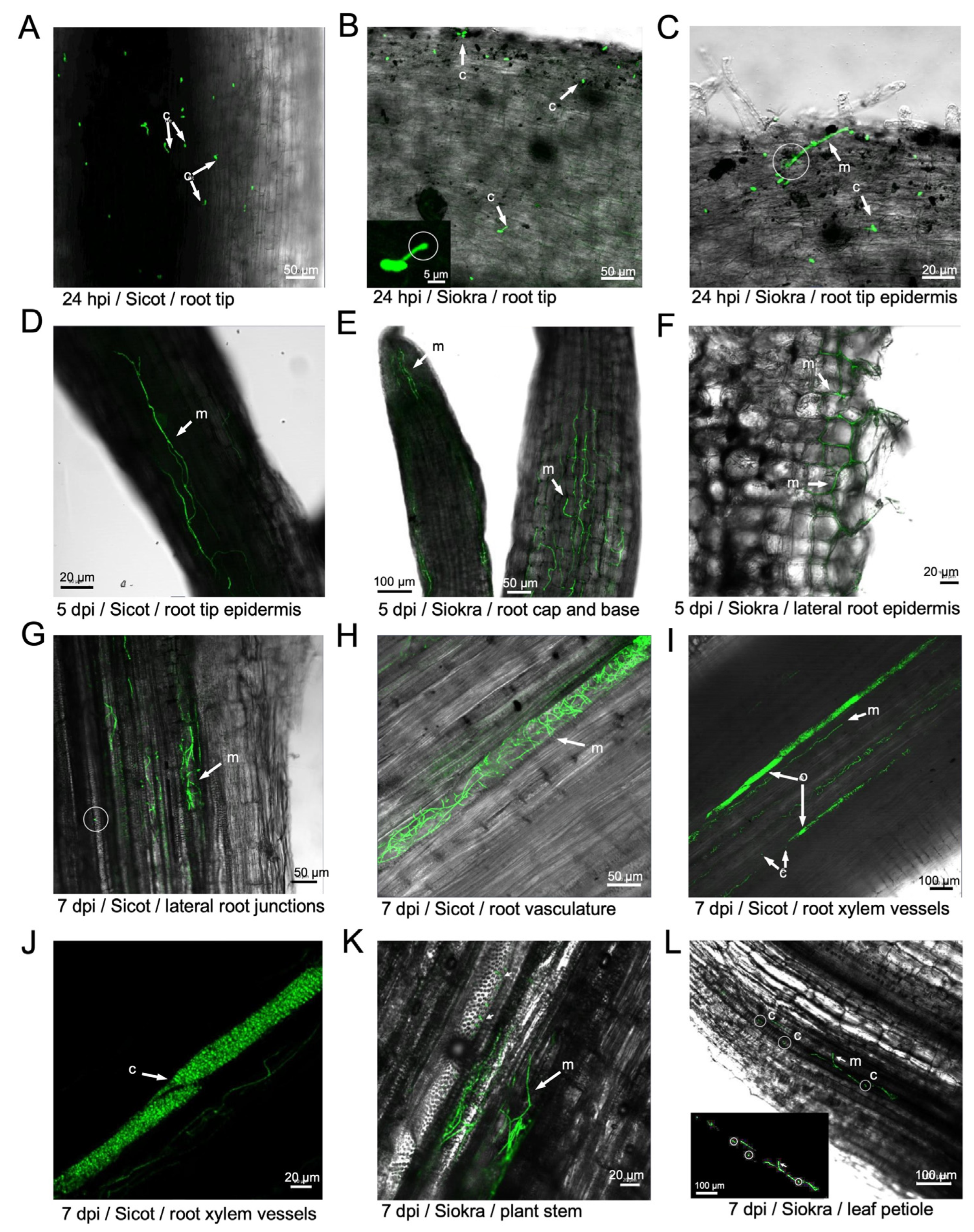
Figure 3.
The development and visualisation of defoliating Verticillium dahliae strains of VCG 1A carrying the mCherry fluorescent protein. (A) Colonies of mCherry transformants compared to the parent Vd71181 after 10 days of growth on half strength PDA. (B) Spores of the parent and the transformants visualised on a Biotek Cytation 1 imager. Scale bar = 200 µm. (C) Fluorescence of spores and hyphae of V. dahliae parent and transformant strains imaged using Biotek Cytation 1 Multi-Reader and Gen5 software. Scale bar = 200 µm. (D) Growth rates (µm2/hr) of the isolates in half-strength PDB media over a 30 h period. No significant differences in growth rates were detected (p = 0.101, one-way ANOVA). (E) mCherry total fluorescence intensity (RF) of the isolates were quantified using Biotek Cytation 1. (F) Fecundity of the isolates measured by spore concentration (spores / mL) after 10 days of growth on half-strength PDA media. Error bars show standard error of the mean (SEM). Statistics performed with a negative binomial GLM. D-E: Medians and IQRs of growth rates are represented by boxes with bars and taken from five replicates (n = 5). Maximum and minimum values are shown as whiskers. E-F: Letters indicate separation of means with significant differences (p < 0.001) detected between groups using One-way ANOVA followed by post-hoc Tukey test.
Figure 3.
The development and visualisation of defoliating Verticillium dahliae strains of VCG 1A carrying the mCherry fluorescent protein. (A) Colonies of mCherry transformants compared to the parent Vd71181 after 10 days of growth on half strength PDA. (B) Spores of the parent and the transformants visualised on a Biotek Cytation 1 imager. Scale bar = 200 µm. (C) Fluorescence of spores and hyphae of V. dahliae parent and transformant strains imaged using Biotek Cytation 1 Multi-Reader and Gen5 software. Scale bar = 200 µm. (D) Growth rates (µm2/hr) of the isolates in half-strength PDB media over a 30 h period. No significant differences in growth rates were detected (p = 0.101, one-way ANOVA). (E) mCherry total fluorescence intensity (RF) of the isolates were quantified using Biotek Cytation 1. (F) Fecundity of the isolates measured by spore concentration (spores / mL) after 10 days of growth on half-strength PDA media. Error bars show standard error of the mean (SEM). Statistics performed with a negative binomial GLM. D-E: Medians and IQRs of growth rates are represented by boxes with bars and taken from five replicates (n = 5). Maximum and minimum values are shown as whiskers. E-F: Letters indicate separation of means with significant differences (p < 0.001) detected between groups using One-way ANOVA followed by post-hoc Tukey test.
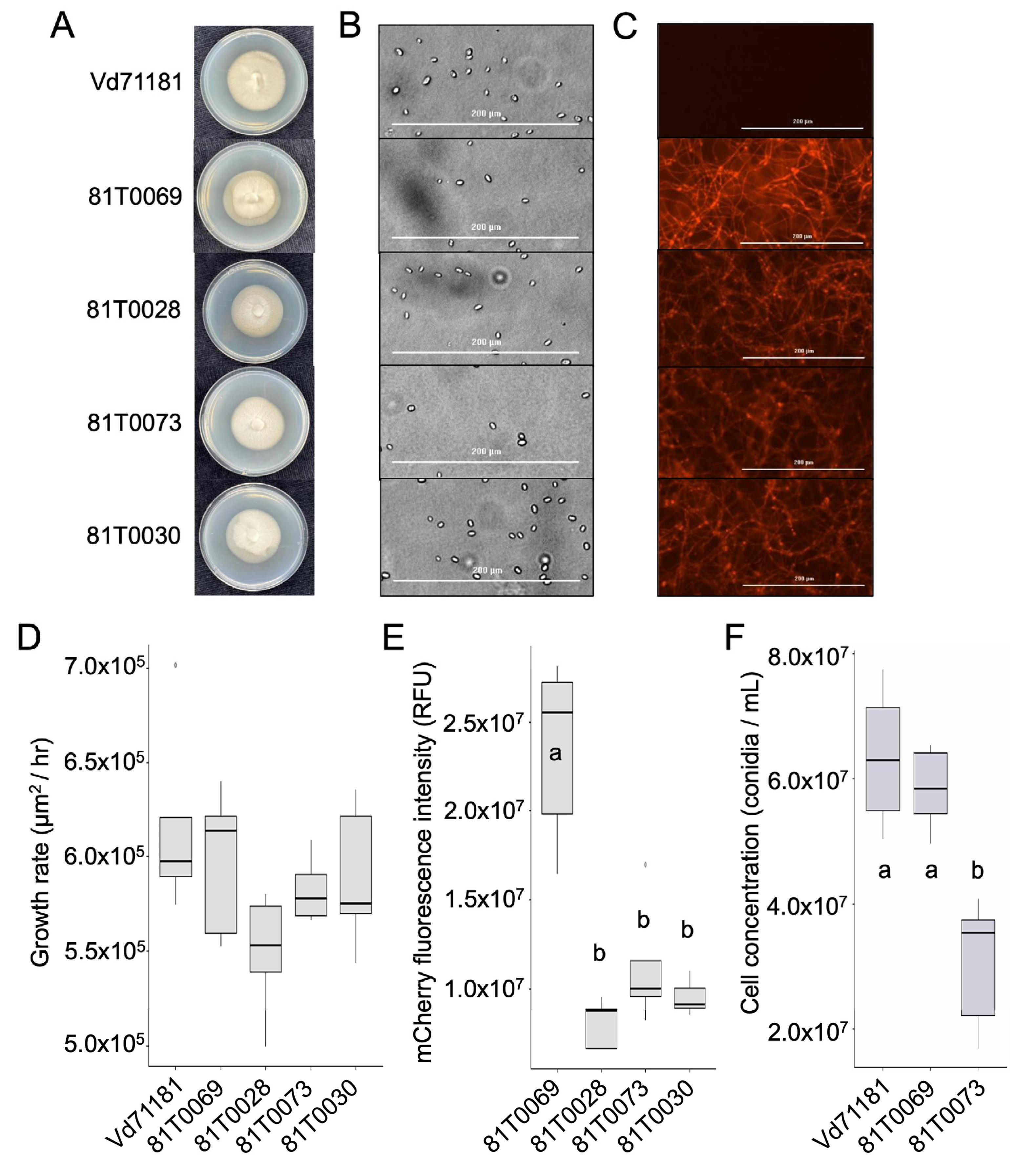
Figure 4.
Representative cotton plants (Sicot) challenged with a defoliating Verticillium dahliae strain Vd71181 from VCG 1A showing disease symptoms and progression. A scale of 0 to 4 depicts the severity of necrosis, chlorosis, and wilting of leaves, as well as stunting of plant stems when compared to the uninoculated plant. A disease score of 5 indicates a dead plant (not shown). Disease scores are based on a scoring guide adapted from Cirulli et al. (1990) (Table 3).
Figure 4.
Representative cotton plants (Sicot) challenged with a defoliating Verticillium dahliae strain Vd71181 from VCG 1A showing disease symptoms and progression. A scale of 0 to 4 depicts the severity of necrosis, chlorosis, and wilting of leaves, as well as stunting of plant stems when compared to the uninoculated plant. A disease score of 5 indicates a dead plant (not shown). Disease scores are based on a scoring guide adapted from Cirulli et al. (1990) (Table 3).

Figure 5.
Assessing the pathogenicity of mCherry-expressing transformants of Verticillium dahliae 81T0069 and 81T0073 on Sicot to compare with that of the parent strain Vd71181. (A) Assessment of symptoms in Sicot seedlings 4 weeks post inoculation. Plants were rated according to Cirulli et al. (1990) and were scored as the following. Uninoculated plant = 1, parental strain Vd71181 = 3, 81T0069 = 3 and 81T0073 = 4. (B) Transverse (t) and longitudinal (l) stem sections of symptomatic plants showing visible discolouration in the vasculature. Red discolouration is indicative of V. dahliae colonisation in the xylem vessels. (C) V. dahliae colonies recovered from cross-sections of the stem tissues of symptomatic Sicot seedlings after 10 days of incubation on half-strength PDA plates. Individual plates show four samples taken from one symptomatic seedling. (D) Mean disease scores in Sicot plants inoculated with Vd71181 (n = 24), 81T0069 (n = 20) and 81T0073 (n = 21). Sterile distilled water was used as the uninoculated control. Medians and IQRs of the growth rates are represented by bars and boxes. Statistical analysis was performed using the Kruskal-Wallis H test and multiple comparisons made using the post-hoc Dunn test adjusted with Benjamini-Hochberg method. Letters indicate the separation of means between the isolates at p < 0.001.
Figure 5.
Assessing the pathogenicity of mCherry-expressing transformants of Verticillium dahliae 81T0069 and 81T0073 on Sicot to compare with that of the parent strain Vd71181. (A) Assessment of symptoms in Sicot seedlings 4 weeks post inoculation. Plants were rated according to Cirulli et al. (1990) and were scored as the following. Uninoculated plant = 1, parental strain Vd71181 = 3, 81T0069 = 3 and 81T0073 = 4. (B) Transverse (t) and longitudinal (l) stem sections of symptomatic plants showing visible discolouration in the vasculature. Red discolouration is indicative of V. dahliae colonisation in the xylem vessels. (C) V. dahliae colonies recovered from cross-sections of the stem tissues of symptomatic Sicot seedlings after 10 days of incubation on half-strength PDA plates. Individual plates show four samples taken from one symptomatic seedling. (D) Mean disease scores in Sicot plants inoculated with Vd71181 (n = 24), 81T0069 (n = 20) and 81T0073 (n = 21). Sterile distilled water was used as the uninoculated control. Medians and IQRs of the growth rates are represented by bars and boxes. Statistical analysis was performed using the Kruskal-Wallis H test and multiple comparisons made using the post-hoc Dunn test adjusted with Benjamini-Hochberg method. Letters indicate the separation of means between the isolates at p < 0.001.
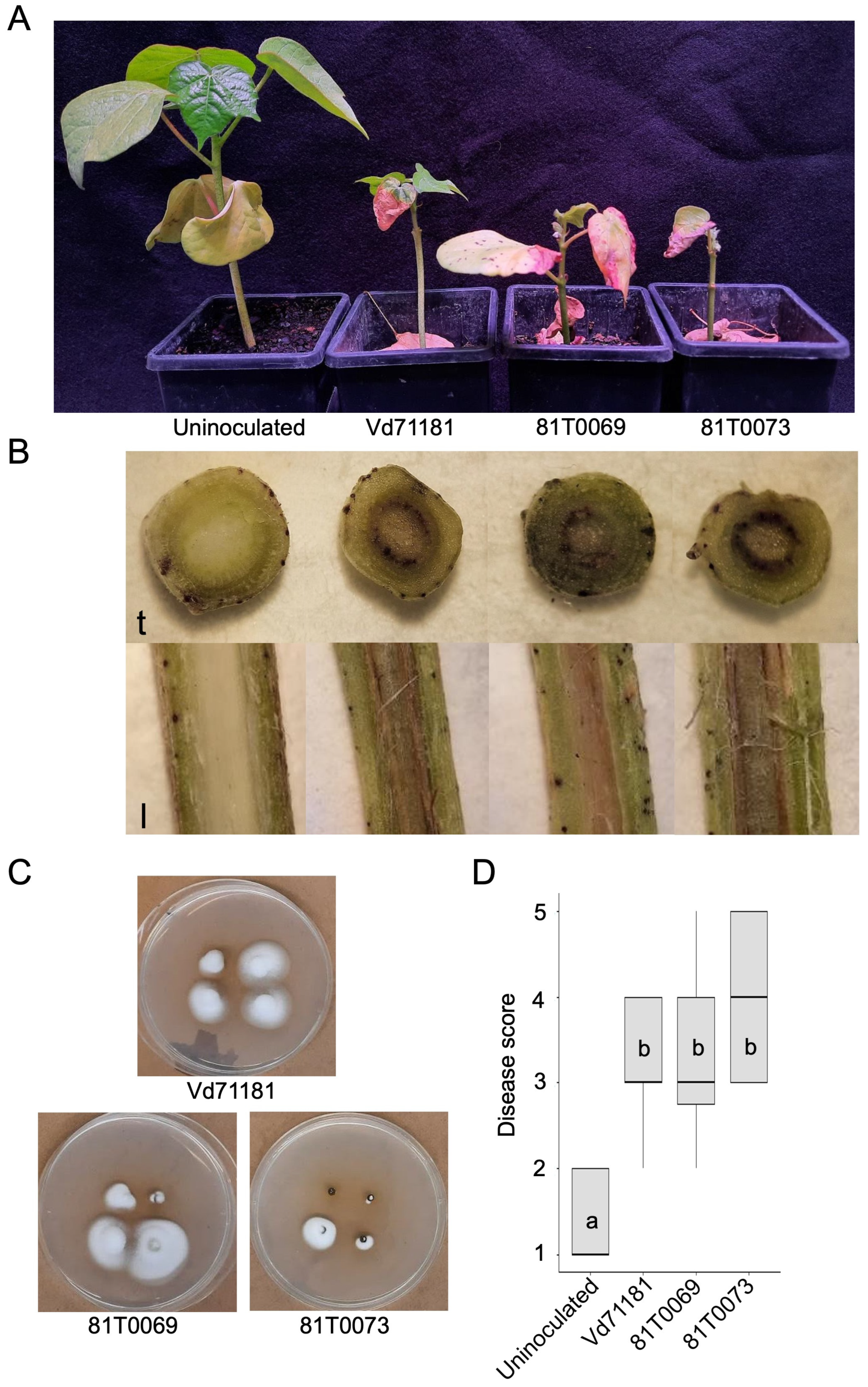
Figure 6.
Localisation of Verticillium dahliae in Sicot cotton seedlings inoculated with the mCherry expressing transformant, 81T0069 at 28 days post inoculation. (A) Confocal microscopy image of longitudinal stem section of Sicot showing the colonisation of host xylem tissues by mCherry expressing mycelia. Viewed at magnification 10x using EC Plan-Neofluar objective. Laser excitation = 555 nm, Master Gain = 809. m = mycelia. c = conidia. mCherry fluorescence is visualised with an overlay of plant tissue in T-PMT transmission illumination mode. (B) mCherry fluorescence visualised in single channel only. (C) T-PMT mode only showing the bright field of plant structure, without the laser scanning mode. (D) magnified view of the cortex region near the xylem and the proliferation of mCherry tagged mycelia in this region. (E) mCherry fluorescence in the cortex region visualised in single channel. (F) T-PMT mode only showing the bright field of cortex region in the stem. Bars indicate the scale used to capture each image.
Figure 6.
Localisation of Verticillium dahliae in Sicot cotton seedlings inoculated with the mCherry expressing transformant, 81T0069 at 28 days post inoculation. (A) Confocal microscopy image of longitudinal stem section of Sicot showing the colonisation of host xylem tissues by mCherry expressing mycelia. Viewed at magnification 10x using EC Plan-Neofluar objective. Laser excitation = 555 nm, Master Gain = 809. m = mycelia. c = conidia. mCherry fluorescence is visualised with an overlay of plant tissue in T-PMT transmission illumination mode. (B) mCherry fluorescence visualised in single channel only. (C) T-PMT mode only showing the bright field of plant structure, without the laser scanning mode. (D) magnified view of the cortex region near the xylem and the proliferation of mCherry tagged mycelia in this region. (E) mCherry fluorescence in the cortex region visualised in single channel. (F) T-PMT mode only showing the bright field of cortex region in the stem. Bars indicate the scale used to capture each image.
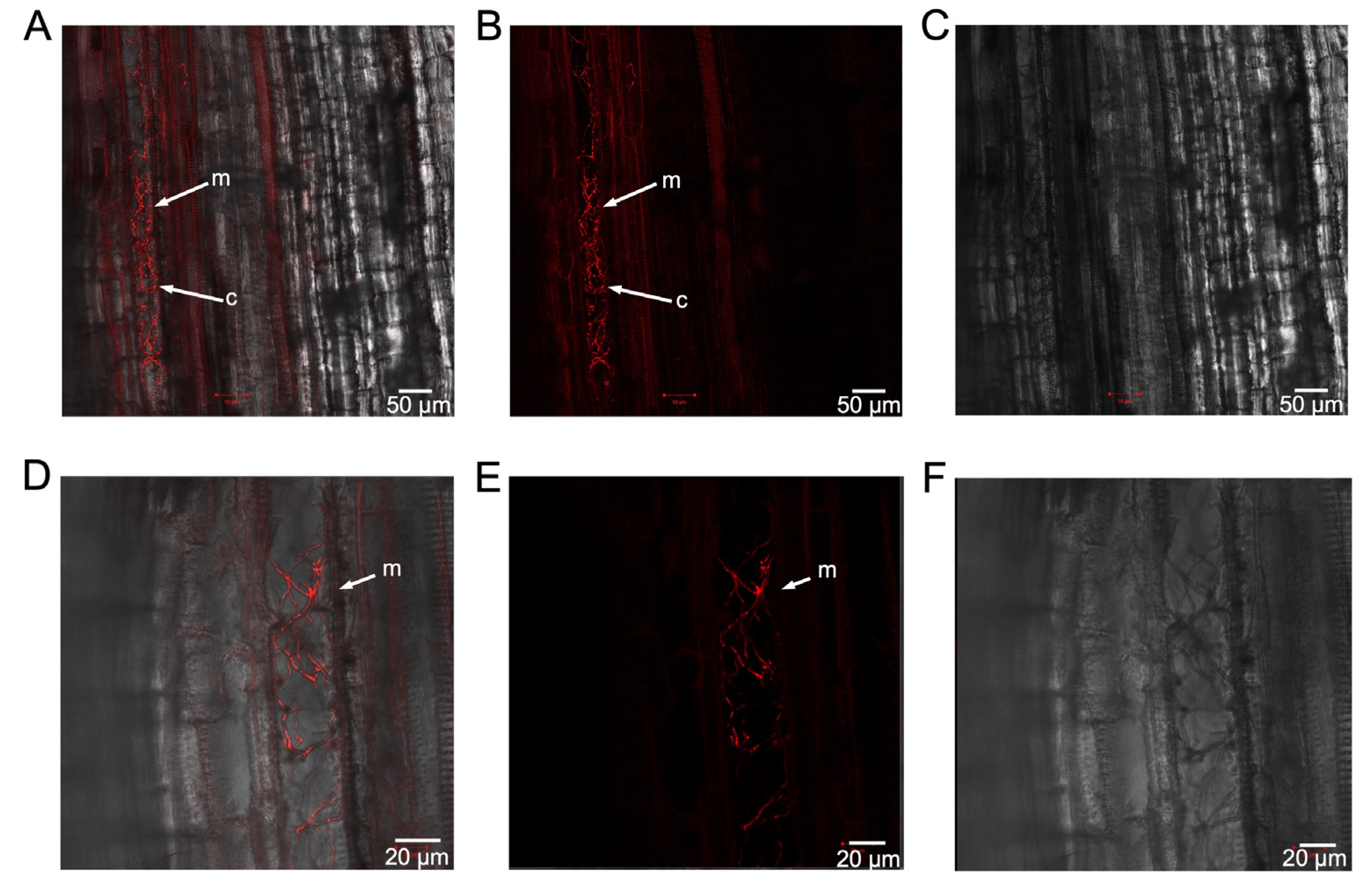
Figure 7.
Assessing different weed species and Nicotiana benthamiana for their potentials to house non-defoliating (VCG 1A) and defoliating (VCG 2A) transformant strains as alternative plant hosts. (A) Conyza bonariensis (flaxleaf fleabane) plants. (B) Sonchus oleraceus (common sowthistle) plants. (C) Avena fatua (wild oats) plants. (D) Chloris truncata (windmill grass) plants. (E) Echinochloa colona (awnless barnyard grass). (F) Chloris virgata (feathertop Rhodes grass) plants. (G) Urochloa panicoides (liverseed grass) plants. (H) Nicotiana benthamiana (native tobacco) plants. Plants were root-dipped in either water (uninoculated), GFP-fluorescing transformant 71T0003, or mCherry-expressing transformant 81T0069 and assessed for external symptoms 4 weeks post inoculation. N = 10 to 25 individual plants per treatment group.
Figure 7.
Assessing different weed species and Nicotiana benthamiana for their potentials to house non-defoliating (VCG 1A) and defoliating (VCG 2A) transformant strains as alternative plant hosts. (A) Conyza bonariensis (flaxleaf fleabane) plants. (B) Sonchus oleraceus (common sowthistle) plants. (C) Avena fatua (wild oats) plants. (D) Chloris truncata (windmill grass) plants. (E) Echinochloa colona (awnless barnyard grass). (F) Chloris virgata (feathertop Rhodes grass) plants. (G) Urochloa panicoides (liverseed grass) plants. (H) Nicotiana benthamiana (native tobacco) plants. Plants were root-dipped in either water (uninoculated), GFP-fluorescing transformant 71T0003, or mCherry-expressing transformant 81T0069 and assessed for external symptoms 4 weeks post inoculation. N = 10 to 25 individual plants per treatment group.
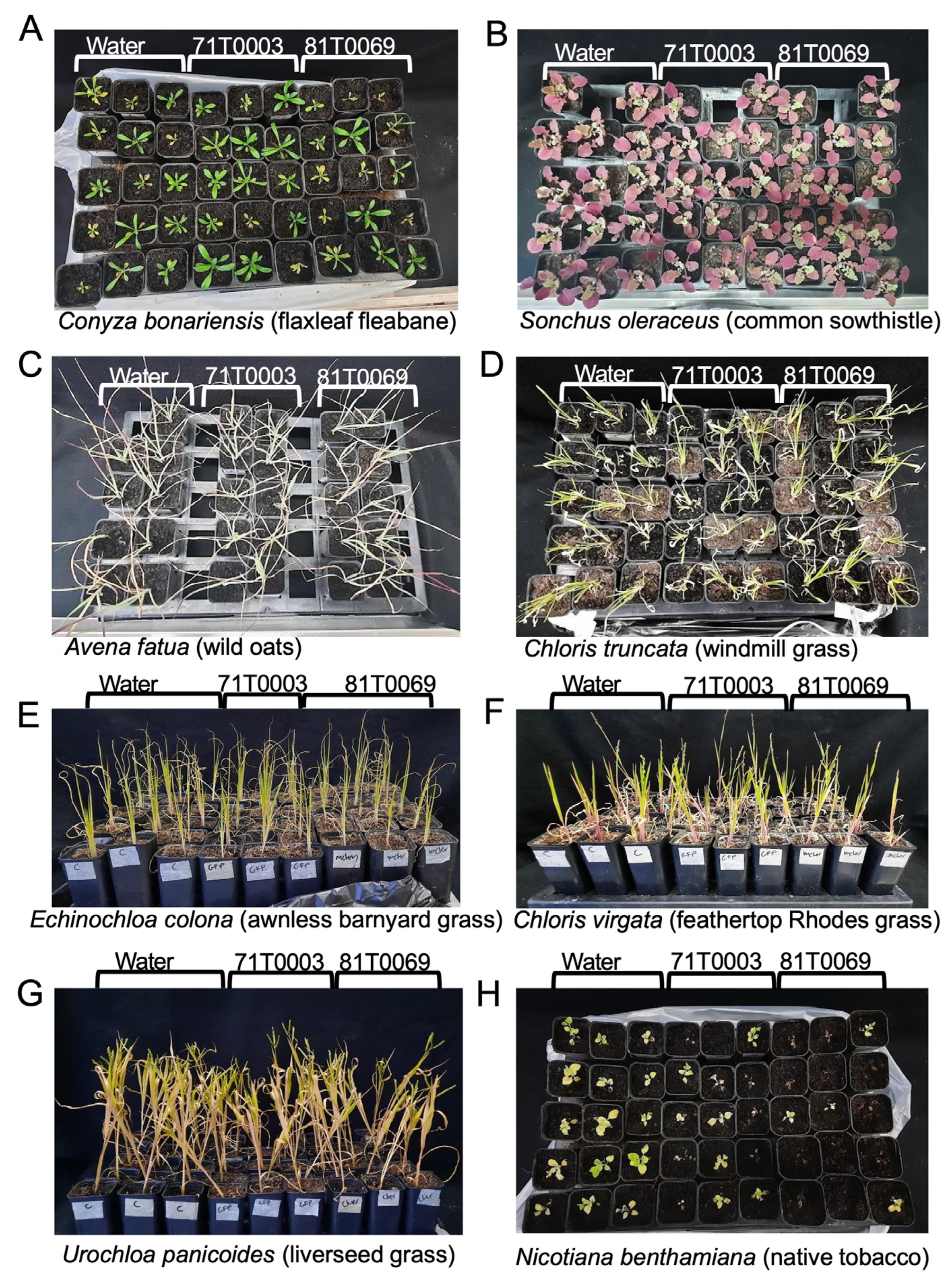
Figure 8.
Assessment of symptoms on weed species and Nicotiana benthamiana inoculated with 71T0003 (GFP, VCG 2A), 81T0069 (mCherry, VCG 1A). (A) Comparison of mean disease scores in weed species and N. benthamiana 4 weeks after inoculation. Scoring was performed according to the scale developed by Cirulli et al. (1990). Uninoculated plants were treated with sterile distilled water and served as a negative control. Statistical analyses were performed with a Kruskal-Wallis H test. Multiple comparisons were performed using a post-host Dunn test adjusted with Benjamini-Hochberg method. Letters indicate separation of means amongst treatment groups for Conyza bonariensis and N. benthamiana at p < 0.05. Error bars represent the standard errors of the mean (SEM). (B) Mycelia tagged with GFP fluorescence was observed in a longitudinal section of a leaf petiole from N. benthamiana plants inoculated with 71T0003. (C) Single channel view of GFP fluorescence. (D) Mycelia tagged with mCherry fluorescence was observed in a longitudinal section of roots of N. benthamiana inoculated with 81T0069. (E) Single channel view of mCherry fluorescence. Viewed at magnification 10x using EC Plan-Neofluar objective. Laser excitation = 555 nm (mCherry), 488 nm (GFP).
Figure 8.
Assessment of symptoms on weed species and Nicotiana benthamiana inoculated with 71T0003 (GFP, VCG 2A), 81T0069 (mCherry, VCG 1A). (A) Comparison of mean disease scores in weed species and N. benthamiana 4 weeks after inoculation. Scoring was performed according to the scale developed by Cirulli et al. (1990). Uninoculated plants were treated with sterile distilled water and served as a negative control. Statistical analyses were performed with a Kruskal-Wallis H test. Multiple comparisons were performed using a post-host Dunn test adjusted with Benjamini-Hochberg method. Letters indicate separation of means amongst treatment groups for Conyza bonariensis and N. benthamiana at p < 0.05. Error bars represent the standard errors of the mean (SEM). (B) Mycelia tagged with GFP fluorescence was observed in a longitudinal section of a leaf petiole from N. benthamiana plants inoculated with 71T0003. (C) Single channel view of GFP fluorescence. (D) Mycelia tagged with mCherry fluorescence was observed in a longitudinal section of roots of N. benthamiana inoculated with 81T0069. (E) Single channel view of mCherry fluorescence. Viewed at magnification 10x using EC Plan-Neofluar objective. Laser excitation = 555 nm (mCherry), 488 nm (GFP).
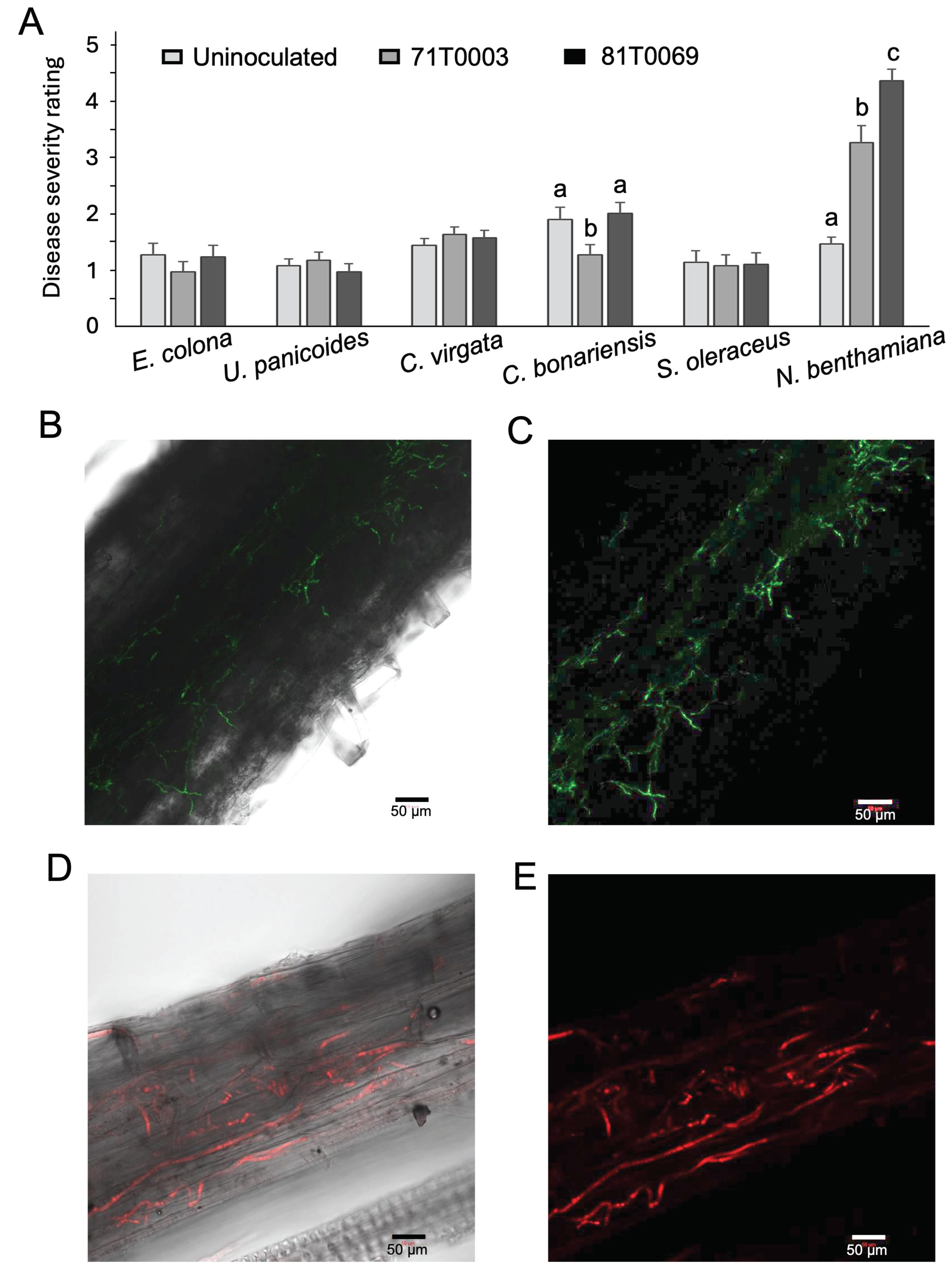

Table 1.
Australian Verticillium dahliae isolates obtained from Gossypium hirsutum (Upland cotton).
| Isolate | Accession | VCG | Pathotype | Locality |
|---|---|---|---|---|
| V. dahliae “Vd71171” | BRIP71171 | 2A1 | Non-defoliating | Namoi Valley, NSW, Australia |
| V. dahliae “Vd71181” | BRIP71181 | 1A2 | Defoliating | Gwydir Valley, NSW, Australia |
1 Confirmed to group with VCG 2A by nitrate-nonutilizing (nit) mutant complementation tests. 2 Predicted to be VCG 1A based on the defoliating pathotype. Its V. dahliae origin was further confirmed by the complete match of a specific ITS region to V. dahliae (Table S2).
Table 2.
Rating scale used to assess disease severity of cotton plants inoculated with a non-defoliating strain of Verticillium dahliae.
Table 2.
Rating scale used to assess disease severity of cotton plants inoculated with a non-defoliating strain of Verticillium dahliae.
| Score | Description of symptoms | Score |
|---|---|---|
| 0 | Healthy, no symptoms | 0 |
| 1 | 1-20% total leaf area affected | 1 |
| 2 | 21-40% total leaf area affected | 2 |
| 3 | 41-60% total leaf area affected | 3 |
| 4 | 61-80% total leaf area affected | 4 |
| 5 | 81-100% total leaf area affected, and or plant death | 5 |
Table 3.
Rating scale used to assess disease severity of cotton plants, weeds, and tobacco plants inoculated with a non-defoliating strain of Verticillium dahliae. This scoring system is adapted from a previous study [52].
Table 3.
Rating scale used to assess disease severity of cotton plants, weeds, and tobacco plants inoculated with a non-defoliating strain of Verticillium dahliae. This scoring system is adapted from a previous study [52].
| Symptoms | Affected leaves (%) | Degree of stunting compared to control1 | ||
|---|---|---|---|---|
| None or very slight (<10%) |
Moderate (11%-50%) |
Severe (>50%) |
||
| None. | 0 | 0 | - | - |
| Slight leaf chlorosis, flaccidity, necrosis. | 1-9 | 1 | 2 | 3 |
| Moderate leaf chlorosis, flaccidity, necrosis, slight defoliation. | 2 | 3 | 4 | |
| 10-24 | ||||
| Severe leaf chlorosis, flaccidity, necrosis, moderate defoliation. | 25-50 | 3 | 4 | 4 |
| Plants with severe or complete defoliation. | >50 | 4 | 4 | 4 |
| Dead plants. | – | 5 | 5 | 5 |
1 Scored based on the percentage in height reduced when compared to the uninoculated control plants.
Table 4.
The percentage of Verticillium dahliae isolates 71T0003and Vd71171 recovered upon termination (4 weeks post inoculation) of the pathogenicity assay.
Table 4.
The percentage of Verticillium dahliae isolates 71T0003and Vd71171 recovered upon termination (4 weeks post inoculation) of the pathogenicity assay.
| Siokra | Siokra | Sicot | Sicot | |
|---|---|---|---|---|
| Treatment | Recovery / Total Diseased1 | Recovery / Total Symptomless2 | Recovery / Total Diseased1 | Recovery / Total Symptomless2 |
| Uninoculated | 0 / 0 | 0 / 48 (0 %) | 0 / 0 | 0 / 48 (0 %) |
| Vd711713 | 31 / 33 (94 %) | 1 / 15 (7 %) | 20 / 24 (83 %) | 0 / 24 (0 %) |
| 71T00033 | 24 / 24 (100 %) | 1 / 24 (4 %) | 7 / 18 (39 %) | 1 / 30 (3 %) |
1 Diseased plants were determined based on V. dahliae symptoms such as wilting, leaf chlorosis and necrosis. 2Plants inoculated with V.dahliae with a disease severity score of 0 were considered symptomless. 3 Isolates recovered were confirmed to be positives by comparing their colony morphology to the parental strain (Vd71171) or by detecting GFP fluorescence (71T0003). Percentage (%) recovery is expressed as the number of plants from which the isolate was recovered from over the total number of diseased or symptomless plants.
Table 5.
Recovery into culture of Verticillium dahliae transformant isolates 71T0003 and 81T0069 from stem tissue of weed species 4 weeks post-inoculation. Colony identities confirmed on the basis of green or red fluorescence.
Table 5.
Recovery into culture of Verticillium dahliae transformant isolates 71T0003 and 81T0069 from stem tissue of weed species 4 weeks post-inoculation. Colony identities confirmed on the basis of green or red fluorescence.
| Family | Weed species | Isolate | VCG | Frequency1 |
|---|---|---|---|---|
| Asteraceae | Conyza bonariensis L. | 71T0003 | 2A | 4/26 (15.4 %) |
| 81T0069 | 1A | 1/26 (3.8 %) | ||
| Sonchus oleraceus L. | 71T0003 | 2A | 2/25 (8 %) | |
| 81T0069 | 1A | 0/25 (0 %) | ||
| Poaceae | Avena fatua L. | 71T0003 | 2A | 0/10 (0 %) |
| 81T0069 | 1A | 0/10 (0 %) | ||
| Chloris truncata R.Br. | 71T0003 | 2A | 0/26 (0 %) | |
| 81T0069 | 1A | 0/26 (0 %) | ||
| Chloris virgata Sw. | 71T0003 | 2A | 3/26 (11.5 %) | |
| 81T0069 | 1A | 0/26 (0 %) | ||
| Echinochloa colona L. | 71T0003 | 2A | 2/26 (7.7 %) | |
| 81T0069 | 1A | 2/26 (7.7%) | ||
| Urochloa panicoides P.Beauv. | 71T0003 81T0069 |
2A 1A |
6/25 (24 %) 1/25 (4 %) |
|
| Solanaceae | Nicotiana benthamiana Domin | 71T0003 81T0069 |
2A 1A |
16/26 (61.5 %) 13/26 (50 %) |
V. dahliae was not isolated from any of the water only control plants. 1 Number of plants from which V. dahliae was isolated / total plants inoculated.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



