
Submitted:
07 March 2024
Posted:
26 March 2024
You are already at the latest version
Abstract
In all mammals, the circulating pool of MLT is synthesized in the pineal gland, during the night darkness hours. Its main function is that of synchronizing the organism in the photoperiod. In contrast, extra-pineal MLT is synthesized in peripheral organs, does not follow any circadian rhythm nor circulates and plays a detoxifying and cytoprotective action. Circulating MLT may stimulate both innate and acquired immune responses by its circadian action and by activating high affinity receptors on immunocompetent cells. Extra-pineal MLT may have anti-oxidant and anti-inflammatory effects dampening the innate immune response. These two seemingly divergent roles may be considered as two faces of the same coin. In fact, the integration of both circulating and extra-pineal MLT functions might generate a balanced and effective immune response against microbial pathogens. The studies described in this review investigated the effect of exogenous MLT in various models of infectious diseases using extremely different doses and treatment schedules. None of them evaluated the possibility to integrate the non-circadian anti-inflammatory effect with the circadian immunoenhancing action of MLT. As a consequence, in spite that most studies agree that MLT has a beneficial effect against infections, it seems difficult to draw any definite conclusion about its possible therapeutic use.
Keywords:
melatonin
; circadian rhythm
; immunomodulation
; anti-inflammatory effect
; cytokine storm
; infection
1. Introduction
The current view about the origin of melatonin (MLT), chemically defined as N-acetyl-5-methoxytriptamine, suggests that MLT appeared on earth about 2.5 billions of years ago. Namely, it is proposed that at that time, anaerobic bacteria developed the ability to synthesize MLT as an adaptative response to increasing concentrations of oxygen in the atmosphere. Thus, the very first function of MLT is considered to be that of counteracting oxygen toxicity. These bacteria were then eventually phagocytized by eukaryotes where, according to the endosymbiotic theory, they established a symbiotic association and evolved in mitochondria or chloroplasts retaining the ability to synthesize MLT [1]. This could explain why MLT may be synthesized in many, if not in all, extra-pineal organs. In mammals, MLT has been identified in skin, gastrointestinal tract, liver, kidney, immune system, bone marrow, testis, skeletal muscles and all body fluids [2]. In general, the concentration of extra-pineal MLT is reported to be orders of magnitude higher than that of the circulating pool derived from the pineal gland. Nevertheless, it has been recently found that the gastrointestinal tract is not a major source of extra-pineal MLT as it was formerly believed [3]. It is therefore possible that some of the early studies in this field overestimated the MLT content because of methodological flaws.
Beyond of being able to act as an antioxidant [4], during evolution MLT acquired many other functions and became a pleiotropic molecule. In vertebrates MLT has got the basic function of synchronizing the organism physiology with the 24-h environmental cycle (circadian rhythm) caused by the daily rotation of our planet. This vital effect is carried out by the circadian oscillation of MLT synthesis in the pineal gland accompanied by its immediate release into the blood circulation [5]. In contrast, extra-pineal MLT does not show any rhythmicity nor is secreted into the blood in significant amounts [2]. The environmental cue driving the MLT circadian rhythm is the light/dark cycle of the day. In particular, light is sensed in retinal ganglion cells through a photopigment called melanopsin that is activated by photons of 460-480 nm wavelength (blue light). The ensuing nervous signal travels in the retino-hypothalamic tract and entrains the suprachiasmatic nucleus (SCN) of the hypothalamus, i.e. the central biological clock of the organism. In turn the SCN activates a nervous pathway involving the paraventricular nucleus (PVN) of the hypothalamus, the intermediolateral cell column and the superior cervical ganglia (SCG) regulating MLT synthesis in the pineal gland [5]. Remarkably, blue light inhibits MLT synthesis, hence MLT is synthesized and released during the darkness part of the photoperiod in all vertebrates [1,5] and this fact synchronizes the organism in the photoperiod by activating specific MLT receptors in both the SCN and peripheral biological clocks [1,5]. Basic biological rhythms such as the oscillations of the autonomic nervous system activity and of the hypothalamic pituitary adrenal (HPA) axis are also driven by the SCN upon the synchronizing action of MLT [6].
In vertebrates MLT synthesis is attained by two enzymatic steps: the first is accomplished by serotonin N-acetyl transferase (SNAT) that lead to the formation of N-acetyl-serotonin and the second transforms N-acetyl -serotonin in MLT by the enzyme hydroxyindole-O-methyl transferase (HIOMT) [1].
2. MLT as an Anti-Oxidant
Oxidative stress can be caused by excessive production of reactive oxygen species (ROS) or by a reduced activity of the antioxidant system. Oxidative stress is well known to increase inflammation and contribute to a variety of pathological conditions including cancer, cardiovascular diseases, neurodegenerative diseases, lung diseases, renal diseases and aging. MLT is considered to be a major player in the anti-oxidant machinery because of its direct scavenging of ROS and its stimulation of anti-oxidant enzymes and suppression of pro-oxidant enzymes [15]. The direct effect as scavenger of free radicals has been clearly demonstrated in cell cultures where MLT and its metabolites usually added at pharmacological concentrations may act by a variety of mechanisms including electron transfer, hydrogen transfer and metal chelation [16]. However, it has been recently pointed out that in living organisms the amount of substances that may react with MLT and its metabolites largely exceeds their concentration even considering that the concentration of extra-pineal MLT is usually orders of magnitude higher than that of plasma MLT [17]. This simple stoichiometric consideration casts some doubt on the view of MLT as an all-purpose in vivo scavenger of free oxygen or nitrogen radicals. However, a different interpretation suggests that MLT and its metabolites may act as stabilizers of the redox state of mitochondria when energy is produced by mitochondrial oxidative phosphorylation [16]. On the contrary, it is well recognized that activation of MT1 and MT2 receptors enhances the expression of anti-oxidant enzymes such as superoxide dismutase, catalase, glutathione peroxidase and glutathione reductase [15,18,19]. In addition, the MLT binding to QR2 inhibits its enzymatic activity reducing the generation of ROS [11].
3. Melatonin and the Immune System
Few reports associated the pineal gland to the immune system [20,21] before 1986 when it was shown for the first time that MLT could increase antibody production in mice and counteract the immunosuppressive effect of corticosterone and/or restrain stress via an opiatergic mechanism [22,23,24]. These findings were reproduced in different experimental models [25,26] and, in general, the immunoregulatory properties of MLT have been further extended in a variety of animal models and in humans [27,28,29,30,31,32,33,34,35]. Nevertheless, the overall picture about the immunological action of MLT is quite confused. Several reports indicate MLT as a powerful in vivo immunoenhancing factor suggesting its use as a therapeutic agent whenever it is needed to boost humoral and /or cellular immune responses, while others endow MLT with an anti-inflammatory effect [27,28,29,30,31,32,33,34,35]. This apparent contradiction might be due to the wide range of concentrations and dosages used possibly linked to the pleiotropic nature of the molecule or, more probably, to the fact that the immunological consequences of the circadian action exerted by MLT via its specific receptors were seldom discriminated from the other non-circadian effects [34].
It is well known that the immune system is under a circadian control exerted by the SCN which drives the activity of the sympathetic nervous system (SNS) and of the hypothalamo-pituitary adrenal (HPA) axis. Both the SNS and HPA axis conveys circadian information to peripheral organs by regulating clock genes expression [37,38] and basic immunological functions such as blood circulation of immunocompetent cells, their infiltration into peripheral organs and the circadian oscillation of their specific functions [39]. In general, circulating cells peak in the blood during the resting phase of the photoperiod while their migration into peripheral tissues occurs during the active phase. These phenomena are essential to ensure tissue homeostasis and activate the appropriate immune response in case of infection. For example, it has been reported that lymphocyte migration into lymph nodes peaked in phase with dendritic cells (DC) at the beginning of the active phase to optimize antigen presentation and the ensuing adaptive immune response [40]. In this context, MLT and its receptors play a fundamental role by their ability to synchronize the circadian output of the SCN and /or drive circadian rhythms directly in other brain structures [41]. Thus, the immunoenhancing action of MLT is linked to its circadian properties including receptor mediated modulation of cytokine production, cell migration and antigen presentation to immunocompetent cells [42,43]. Last but not least, MLT may suppress the nuclear translocation of glucocorticoid receptors [44] and hence modulate their effect on immunity [45].
In conclusion, the available evidence including exogenous administration and studies in pinealectomized animals [43], suggests that the immunoenhancing action of MLT is exerted at physiological or supraphysiological concentrations via activation of its specific receptors (Figure 1).
Blue light constitutes the major environmental cue regulating MLT synthesis in the pineal gland by inducing the nocturnal activation of preganglionic sympathetic neurons. MLT produced during the darkness hours is released into the blood and synchronizes the central biological clock of the organism, i.e. the SCN, with the photoperiod. In turn, the SCN drives the circadian oscillation of the sympathetic nervous system which may influence immunocompetent cells by adrenergic activation of a-and b-adrenoceptors. The SCN rhythm also induces the paraventricular nucleus to release corticotropin-releasing hormone which in turn stimulates adrenocorticotropic hormone production by the anterior hypophysis. This final stimulates the production of glucocorticoids(GCs ) by the adrenal cortex. All together these mechanisms influence circulation, migration and functions of immunocompetent cells. In addition, a robust circadian machinery is associated with regular sleep with positive effect on immunity. Last but not least, circulating MLT may act directly on MT1 and MT2 expressed on immunocompetent cells as well as on calmodulin (CAM) and modulate the expression of cells adhesion molecules , cytokine production and cell proliferation. Circadian administration of exogenous MLT using amounts that do not oversaturate its receptors may reinforce both the indirect and direct immunoenhancing effects of MLT.
On the contrary, non-circadian effects that comprise anti-inflammatory and mitochondria-related effects are exerted by MLT at concentrations of the same order of magnitude of extra-pineal MLT that physiologically does not contribute to the circulating pool. As such concentrations would oversaturate any receptor, the aforesaid effects are obviously receptor-independent [34].
Another important consideration concerning the immunological action of MLT relates to the widely used distinction between pro-inflammatory and anti-inflammatory effects with the former being used as synonymous of “immunostimulating”. This effect involves stimulation of cytokines such as IL-1β, IL-2, IL-6, IL-8, TNF-α , IFNγ, IL-17A and/or upregulation cyclooxygenase-2 (COX-2) and inducible NO synthase. Other effects are exerted on hematopoiesis by stimulating GM-CSF and differentiation of Th cells and NK cells [46]. Yet, an excessive inflammatory response may produce a paradoxical effect on immunity leading to activation of the coagulation system, organ failure and immunodeficiency by inducing T cell apoptosis [47]. Several studies bestow MLT with the capacity of exerting opposite actions according to the ongoing biological process [48]. In regard to the immune system, it has been proposed that the contrasting actions of MLT represents a system to guarantee the appropriate immune response according to the pathological situation [31,47]. The anti-inflammatory action of MLT includes several mechanisms in part related to its antioxidant properties. MLT may inhibit NF-kB activation , upregulates Nrf2 and inhibit TLR4 signaling Some of these MLT effects seem to be related to the activation of sirtuin 1(SIRT1) [46].
A bidirectional communication between the pineal gland and the immune system has been proposed as a mechanism integrating the immunological function of both pineal and extra-pineal MLT. This mechanism, termed immune-pineal axis, is based on the switch of MLT production from the pineal gland to local immunocompetent cells at the site of infection or tissue damage to control the inflammatory response and then, after resolution, back to the pineal gland again [49]. The transient inhibition of pineal MLT production and the induction of its synthesis in macrophage/microglia seems to depend on NFkB activation by circulating cytokines and/or pathogen associated molecular patter (PAMPs). Then, the inhibition of NFkB activity by extrapineal MLT and the activation of the HPA axis associated with the inflammatory response restore the circadian release of pineal MLT which is essential to optimize the acquired immune response and maintain the immune homeostasis [49]. Thus, one could infer that both the anti-inflammatory and pro-inflammatory effects of MLT are the sides of the same coin aimed at ensuring a successful immune response against the invading pathogen by balancing the innate with the acquired immune response.
Beside immunity, MLT can also influence hematopoiesis, i.e. the process responsible for the daily production of erythrocytes and immunocompetent cells. MLT was shown to rescue hematopoiesis in mice against the toxicity of anti-cancer drugs. This effect was apparently due to Th cell-derived opioid cytokines binding to k-opioid receptor on GM-CSF activated bone marrow (BM) stromal cells possibly resulting in IL-1 production [50]. The ability of MLT to counteract myelosuppression due to the toxic action of anti-cancer drugs was then amply confirmed in patients [51,52]. In the BM, hematopoietic stem and progenitors cells (HSPC) circadian mobilization and circulation is essential to replenish the blood with immunocompetent cells and ensure the immune homeostasis. In this process, BM-derived MLT play a major role by inducing HSPC quiescence and retention [53].
Finally, MLT may influence the immune system by its well known effect on sleep. Sleep and immunity are tightly linked. Regular sleep is crucial for the immune system and immune derived factors are needed for regular sleep. Thus, the effect of MLT on inflammatory cytokines might be linked to its sleep-facilitating action which in turn contributes to maintain a healthy immune system [54].
4. MLT and Viral Infections
The firs evidence about an antiviral activity of MLT was shown against encephalomyocarditis virus (ECMV), a highly pathogenic virus that produces encephalitis and myocarditis in rodents. Exogenous MLT could prevent paralysis and dead of mice infected with EMCV [55]. Other encephalitogenic viruses proved to be affected by MLT. Normal mice were infected with the Semliki Forest virus (SFV) and stressed mice were injected with the attenuated non-invasive West Nile virus (WNV). SFV can produce viral encephalitis in normal mice while the attenuated form of WNV can do it only in immunosuppressed mice. In both models, administration of MLT significantly postponed onset of the disease and reduced mortality [56]. A similar effect was then reported in mice infected with Venezuelan equine encephalomyelitis virus (VEEV) [57]. The protective effect of MLT in this model was shown to depend on increased IL-1β production as it was abolished by IL-1β neutralization [58]. An inverse correlation between MLT and IL-12 plasma levels and disease progression has been described in HIV-1 infected individuals suggesting a direct relationship between MLT and Th1 cell function [59]. MLT has proven to be effective also against respiratory syncytial virus (RSV). In vitro infection of macrophages with RSV activated TLR3 and NFkB and the subsequent inflammatory response. In this model MLT was able to inhibit the response by suppressing NFkB activation [60]. This effect was reproduced in mice infected with RSV where MLT could inhibit lung oxidative stress [61]. The anti-inflammatory and regenerative effect of MLT was evident also in rabbit with fulminant hepatitis of viral origin [62]. In the same model, another study showed that MLT could inhibit mitophagy and the innate immune response while restoring the circadian dysregulation induced by the virus, recommending the use of MLT as a therapeutic option in human fulminant hepatic failure [63]. In an in vitro model of Hemorrhagic Shock Syndrome by the EBOLA virus , MLT was suggested to be a promising therapeutic agent because of its ability to neutralize endothelial cell disruption [64].
With the advent of the Covid-19 pandemic, an impressive number of studies tested MLT for possible therapeutic and prophylactic effects. A PubMed search with the terms MLT and COVID-19 retrieved 138 publications including many clinical randomized studies. However, in spite of this outsized number of publications it is difficult to draw any definite conclusion about the therapeutic efficacy of MLT in Covid-19 patients. In fact, there are reports showing a positive therapeutic effect of exogenous MLT [65,66,67,68,69,70], while others deny any effect [71,72,73]. Amid the beneficial effects exerted by MLT against Covid-19, we can find prevention of complications and reduction of mortality in severely ill patients [67,70], improvement of respiratory symptoms by reduction of lungs involvement [69] and reduced requirement for invasive mechanical ventilation as well as overall improvement of clinical status [70]. On the other hand, a randomized retrospective study negates any effect of MLT on survival of Covid-19 patients [71] and another contemporaneous randomized clinical trial reached the same conclusion [72]. Perhaps this drastic discrepancy is due to the wide array of doses and treatment schedules used in these studies which continue to perpetuate the misperception about the real therapeutic properties of MLT. For example, MLT has been administered once per day in the evening at a 10 mg dose for 14 days [67] or twice a day without mentioning the timetable at a dose-pro-dose of 3 mg [69] or 5 mg [70]. In the rationale of the studies none considered a distinction between the circadian and non-circadian effects of MLT that could be related to its conceivable therapeutic effect against Sars-Cov-2. Most studies just mentioned in a general fashion the immunomodulatory and anti-inflammatory effects of MLT. Moreover, some new and peculiar mechanism of action have been highlighted to explain the observed effect of MLT. Thus it has been reported an influence of MLT on the pathogenic enzyme p21-activated kinase 1 whose activation is involved in a variety of pathological conditions including viral infections [65] , on cluster differentiation 147 [66], on viral phase separation and epitranscriptomics [68] and on the coagulation system [67].
The emergency linked to the Covid-19 pandemic has somewhat boosted the interest for the putative anti-viral potential of MLT generating studies about its effect on influenza infection. Even in this case, MLT has been administered at very high doses and in some cases with treatment schedules ignoring completely its circadian nature. A report claims that MLT ameliorates influenza A H1N1 infection in mice by virtue of its ability to inhibit pro-inflammatory cytokines while enhancing the anti-inflammatory cytokine IL-10. MLT was administered subcutaneously either 6 hours before infection and/or 2,4 and 6 days post infection at a concentration of 200 mg/kg b.w. without specifying any timetable [74]. Another study in mice infected with influenza A H3N2, MLT was administered intraperitoneally at 30 mg/kg b.w. for 7 days in the evening. In this case MLT proved to diminish pulmonary damage, leukocyte infiltration, edema and to switch the polarization of alveolar macrophages from M1 to M2 phenotype [75]. A third study provided the interesting observation that MLT-deficient mice show a significantly higher mortality rate in comparison to their wild type counterpart after infection with influenza A H1N1 virus. In other experiments BALB/c mice were pretreated for 3 days with intranasal administration of MLT (3,10 and 30 mg/kg b.w.) before virus inoculation. The MLT treated animals were significantly protected from the virus apparently by suppression of mast cell activation and inhibition of cytokine storm [76].
Again, we are faced with results that are difficult to integrate in a clear understanding of the MLT action. In particular, it seems rather problematic to gather together the interesting observation of an augmented vulnerability to influenza infection of MLT-deficient mice with the effects of exogenous MLT administered at very high doses and by extremely different treatments. Table 1. shows the accessible preclinical studies investigating the possible therapeutic effect of MLT against viral diseases.
It seems noteworthy the observation that early studies used supraphysiological doses of MLT administered according to a circadian schedule while the recent ones employed high pharmacological doses and typically did not follow any circadian administration. Probably this divergence reflects a different conceptual approach connected to the MLT property to be exploited in fighting the infection. In the initial studies the authors investigated whether the immunoenhancing action could be used to fight the disease while in the latest ones, the authors mostly focused on the anti-oxidant and anti-inflammatory effect.
5. MLT and Bacterial Infections
The first evidence suggesting that MLT could influence the outcome of bacterial infection was its protective effect in an animal model of septic shock. A single injection of MLT, few hours after intraperitoneal inoculation of a lethal dose of LPS in mice, was able to protect the animals. The doses used were 1,2,3,4,5 and 10 mg/kg b.w, and the protective effect which involved a reduction of NO synthesis, was significant in the 2-5 mg range but was lost at 10 mg [77]. This finding was then confirmed and extended in a variety of animal models and in humans with sepsis [78]. In particular, MLT could ameliorate the clinical status and increase survival of human newborns with sepsis [79]. Doses and treatment schedule ranged from oral administration of 2 single doses of 10 mg within 12 hours from the diagnosis of sepsis to one injection of 20 mg/kg in septic newborns treated with antibiotics. In general, the effect of MLT is suggested to depend on suppression of prooxidant and proinflammatory pathways [78,79]. However, a recent study shows that polymicrobial sepsis in mice enhanced the expression of MT2 receptors in neutrophils and that MLT administration protected the mice by enhancing the bactericidal effects of neutrophils [80]. In this study MLT was used at the massive doses of 50 mg/ kg in vivo and 100 ug/ml in vitro [80]. Another study reports that in mice exposed to short photoperiod and infected with Staphylococcus aureus or Escherichia coli, MLT administration at 10 mg/kg resulted in an improved clearance of bacteria from blood [81]. A further in vitro study using porcine macrophages claims that the impracticable concentration of 1 mM MLT may improve bacterial clearance of enterotoxigenic Escherichia coli and suggests the MLT is important to control this type of infection [82]. Similarly, in a model of Escherichia coli meningitis, mice were treated for 7 days with MLT at 10, 30 and 60 mg/kg and the claim was that MLT may prevent meningitis by acting on the intestinal microbiota [83]. Also bacterial mastitis and infection by Klebsiella pneumoniae are among the bacterial diseases in which MLT is suggested to exert a therapeutic action by virtue of its anti-inflammatory and anti-oxidant effects [84,85]. The doses and concentrations of MLT used in these studies are in line with the above reported citations.
Antimicrobial resistance is a growing emergency in public health. In particular, the transferable resistance-nodulation-division efflux pump TMexCD1-TOprJ1, conferring resistance to tigecycline is becoming a serious health problem. A potentially very interesting and novel approach to combat resistance to tigecycline used MLT either in vitro or in vivo in an infection model using tmexCD1-toprJ1-positive Klepsiella pneumoniae with encouraging results [86]. However, even in this study MLT was used at extremely high concentrations (2-8 mg/ml) and doses (50 mg/kg) [86]. Table 2. lists the preclinical in vivo studies about the effect of MLT against bacterial infections.
As for viral diseases, also in these studies the rationale for using gigantic amounts of MLT is not mentioned. It is somewhat surprising that in most studies the well known concept that circadian rhythms influence the outcome of and the susceptibility to infections [87], was completely ignored. In addition, both viral and bacterial infections may disrupt the circadian machinery [87] but whether such effect involves the immune-pineal axis or it is exerted only on peripheral circadian clocks is still obscure. In my opinion, this is probably the crucial point that has to be carefully pondered in future studies aimed at improving the therapeutic approach by MLT in infectious diseases.
6. MLT and Parasitic Infections
The available evidence suggests that MLT may affect parasitic infections by acting directly on the biology of protozoan parasites and/ or the host’s immune response.
6.1. Malaria
Malaria is caused by parasites of the genus Plasmodium. The infection is transmitted by female Anophele mosquitos which inject the parasite sporozoites into the host during blood feeding. The sporozoites establish the primary infection in the liver where they proliferate and become merozoites. These are then released from the hepatocytes and infect erythrocytes where they develop through a series of different stages and finally are released to infect more erythrocytes. The periodic rupture of erythrocytes releases cytosolic substances and parasite metabolites that elicit the host response causing the malaria symptoms which follow a circadian cycle. In this rhythm, MLT plays a major role synchronizing the Plasmodium cell cycle by acting on the cAMP-PKA and IP3-Ca2+ pathways to favor the synchronous egress of the merozoites which enhances their capacity to invade circulating erythrocytes. The development of MLT-related analogues capable of disrupting this cycle shows a promising therapeutic potential against malaria which affect several millions of people worldwide [88]. Remarkably, a recent report claims that MLT administered at doses of 5 and 10 mg/kg may prevent brain damage and cognitive impairment in an animal model of cerebral malaria [89]. In this model, mice were infected with Plasmodium berghei which develops an asynchronous pattern of infection because its life cycle is not influenced by MLT [90].
6.2. Trypanosomiasis
T.cruzi is transmitted by insect vector or blood transfusion and organ transplantation. In humans causes Chagas’ disease, an extremely debilitating illness that has spread thru migration from Latin America to the rest of the world, especially to United States and Europe [91].
The possible therapeutic effect of MLT against T. cruzi have been studied in rodents.
In a series of studies by Santello and coworkers, administration of MLT at 5 mg/kg either before infection as a pretreatment or during T.cruzi infection in rats proved to protect the animals against the disease by increasing the Th1 response while suppressing the Th2 response as evidenced by enhanced production of TNF-α, IFN-γ, IL-12, IL-2 and increased leukocytes counts [92,93,94].
In the same model, another study showed MLT and zinc treatment increased the plasma level of IL-2, IL-10 as well as thymocyte proliferation counteracting the parasite induced immune alterations [95]. A more recent study using mice reports that MLT decreased the circulating load of parasitemia without affecting the parasite replication. At the cellular level MLT rather enhanced the parasite release, a potentially dangerous effect [96].
Human African trypanosomiasis is caused by the protozoan parasites Trypanosoma brucei and it is transmitted by the bite of the tsetse fly. The disease is also called sleeping sickness because it is associated with a disruption of the circadian rhythms possibly because it elicits a Th1 skewed immune response [97]. However, apart from a publication showing that MLT administration restored a normal sleep pattern in rats infected with T. brucei [98], no study addressed a possible therapeutic action of MLT against the infection.
6.2. Leishmaniases
Leishmaniases are caused by protozoan parasites of the Leishmania genus transmitted by the bite of a sand-fly and are characterized by cutaneous, mucocutaneous or visceral lesions. Leishmania amastigotes are obligatory intracellular parasites which can live and reproduce within macrophages. By doing so the parasite is able to modulate the host immunity reducing inflammation and the adaptive immune response [99].
The first evidence of a possible MLT action in Leishmaniases was provided in 2014 by in vitro study showing that pharmacological concentration of MLT could reduce the number of viable Leishmania infantum promastigotes by altering Ca++ distribution in the parasite [100]. An interesting study in rodents shows that the Leishmania/host interaction varies following the circadian rhythm of MLT production and that MLT treatment during day time reduced the macrophage uptake of arginine by 40% inhibiting parasites replication [101]. More recently, a report shows that pharmacological concentrations of MLT reduced Leishmania amazonensis infection in murine macrophages and modulated host microRNA expression as well as production of cytokines such as IL-6.MCP-1/CCL2 and RANTES/CCL9 [102]. A remarkable study combined MLT with amphotericin B in solid lipid nanoparticles and treated Leishmania donovani infected BALB/c mice inducing a 98.89% decrease of the intracellular parasite load in liver tissue. It was therefore emphasized that MLT would be effective in combating visceral Leishmaniases [103].
6.2. Toxoplasmosis
Toxoplasmosis is caused by the obligate intracellular parasite Toxoplasma gondii and is one of the most common parasitic infection in humans. Toxoplasmosis is very frequent and usually asymptomatic but it may cause a fatal disease in presence of immunodeficiency. In spite of this risk, the therapeutic options against severe Toxoplasmosis are still inadequate [104].
As far as it concerns MLT, few studies addressed its possible role against Toxoplasmosis. In a model of retinochoroiditis, rats were infected with Toxoplasma gondii and treated with MLT (3mg/kg) for one month. The experimental groups included also pinealectomized mice and zinc supplementation. The highest cellular infiltration by CD3+, CD4+ and CD8+ cells was observed in the choroid and retina of rats treated with MLT in combination with zinc supplementation. On the contrary, the minor infiltration was found in pinealectomized animals not supplemented with zinc [105]. Similar findings were obtained in other animal studies by the same group [106,107].
Although at concentrations in the mM range , MLT has been described to inhibit Toxoplasma gondii growth in a monkey kidney cells line culture without affecting host cells viability. The authors suggested that MLT reduced parasites growth inducing both apoptosis and necrosis by modifying energy metabolism [108]. An opposite finding has been reported in a human colon adenocarcinoma cell line, where 0.2 nM MLT proved to boost parasite replication. It was also suggested that Toxoplasma parasites may degrade IDO1 and convert tryptophan to MLT which in turn suppress ROS generation and favors their growth. The presence of IFNγ prevents IDO1 degradation so that tryptophan is catabolized into kynurenine, inducing cell death [109].
The impractical high concentrations of MLT used in these in vitro studies and the conflicting results obtained cast doubts on the biological meaning of these results.
Curiously, in most in vivo studies related to parasitic infections MLT was administered at pharmacological doses ( few mg/kg) without, however, reaching the exaggerated doses used in the models of viral and bacterial infection. Perhaps, this peculiarity reflects a more specific and higher sensitivity of parasites to MLT in comparison to pathogenic viruses and bacteria.
6. Conclusion
There is no doubt that being a multitasking molecule, MLT may influence infectious diseases. In this sense, the more persuasive studies are those showing that MLT-deficient animals show higher susceptibility and/or higher mortality when infected with influenza virus H1N1 [76], Leishmania amazonensis [101] and Toxoplasma gondii [105]. Other outstanding studies are those showing that MLT may overcome tigecycline resistance in a model of Klebsiella pneumoniae infection [86] and its synergy with amphotericin B in combating Visceral Leishmaniases [103]. These observations suggest that combining MLT with antiviral drugs or with antibiotics might be a very promising approach to fight infections. In fact, due to its equilibrating action on the immune response and its anti-inflammatory effect, MLT might improve the therapeutic index of the drugs and decrease their toxicity. On the other hand, in oncology MLT has been already shown to improve the effectiveness of anti-cancer treatments while counteracting their negative side effects [110]. The majority of the experimental studies agree that MLT exerts a positive therapeutic effect in viral, bacterial and parasitic infections. This evidence encouraged clinical studies in septic patients and in Covid-19 patients. The results obtained confirmed the therapeutic potential of MLT against sepsis [78,79] but were controversial in case of Covid-19[65,66,67,68,69,70,71,72,73]. In general, the beneficial action of MLT was ascribed to its immunomodulating and/or anti-inflammatory properties. Nevertheless, no investigation challenged the hypothesis that these properties might be two faces of the same coin, i.e. of the immune-pineal axis whose physiological role is that of balancing innate and adaptive immune responses to the sake of optimizing the defense against the invading pathogen. In fact, in all relevant in vivo studies, MLT has been administered by a variety of treatment schedules and using very different doses mostly in the high pharmacological range. Similarly, MLT was often tested in vitro for possible direct effects on infectious agents at very high, almost unreasonable concentrations. Moreover, no study asked the question whether the effects observed were due, at least in part, to the ability of MLT to synchronize the circadian machinery and hence all neuroendocrine, immune and hematopoietic cycles influencing either the susceptibility or the resistance to infectious agents. Thus, the state of the art in this potentially important topic is far from being clear and does not provide a sound basis for clinical applications. It is in fact very possible that to be most effective,, MLT have to be administered at different doses and different schedules according to the infection stage. It might be that in the early stages when the host reaction is mostly of the innate type, MLT should be administered at pharmacological doses without taking in consideration any circadian rhythmicity, while later, when adaptive immunity takes place, the MLT doses should be lower and administered in the evening in accordance with its endogenous circadian rhythm. This schedule is schematized in Figure 2.
On the basis of the available evidence about the possible therapeutic actions of MLT in combating infections, various treatments schedules are proposed. The scheme represents different experimental possibilities to investigate the therapeutic effect of MLT. The effect of pre-treatment and treatment might be evaluated either separately or in combination and, within the treatment protocol, one could evaluate separately the effect of the non-circadian high doses vs that of the circadian low doses and so on. Hopefully, various doses of MLT should be investigated with low doses possibly ranging from 10 ug to 1000 ug / kg administered in a circadian fashion in the evening and high doses from 3 mg to 50 mg /kg administered in the morning. Of course, all relevant control groups should be incluided.
In this way, the treatment schedule would mimic the physiological response of the immune-pineal axis, reinforcing bot the anti-inflammatory action early during the infection and, later, the ensuing adaptive immune response. To verify this hypothesis further studies investigating doses and treatment schedules in the various types of infection are clearly needed.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Zhao, D.; Yu, Y.; Shen, Y.; Liu, Q.; Zhao, Z.; Sharma, R.; Reiter, R.J. Melatonin Synthesis and Function: Evolutionary History in Animals and Plants. Front Endocrinoly 2019, 10, 249. [Google Scholar] [CrossRef] [PubMed]
- Acuña-Castroviejo, D.; Escames, G.; Venegas, C.; Diaz-Casado, M.; Lima-Cabello, E.; Lopez, L.; Rosales-Corral, S.; Tan, D.; Reiter, R. Extra-pineal melatonin: Sources, regulation, and potential functions. Cell. Mol. Life Sci. 2014, 71, 2997–3025. [Google Scholar] [CrossRef]
- Kennaway, D. J. The mammalian gastro-intestinal tract is a NOT a major extra-pineal source of melatonin. J Pineal Res 2023, 75, e12906. [Google Scholar] [CrossRef] [PubMed]
- Carretero, V.J.; Ramos, E.; Segura-Chama, P.; Hernández, A.; Baraibar, A.M.; Álvarez-Merz, I.; Muñoz, F.L.; Egea, J.; Solís, J.M.; Romero, A.; Hernández-Guijo, J.M. Non-Excitatory Amino Acids, Melatonin, and Free Radicals: Examining the Role in Stroke and Aging. Antioxidants (Basel, Switzerland) 2023, 12, 1844. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Galano, A. Melatonin: exceeding expectations. Physiology (Bethesda, Md.), 2014, 29, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Kalsbeek, A.; Yi, C.X.; Cailotto, C.; la Fleur, S.E.; Fliers, E.; Buijs, R.M. (2011). Mammalian clock output mechanisms. Essays in biochemistry 2014, 49, 137–151. [Google Scholar] [CrossRef]
- Reppert, S.M.; Weaver, D.R.; Ebisawa, T. Cloning and characterization of a mammalian melatonin receptor that mediates reproductive and circadian responses. Neuron 1994, 13, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Reppert, S.M.; Weaver, D.R.; Ebisawa, T.; Mahle, C.D.; Kolakowski, L.F.; Jr. Cloning of a melatonin-related receptor from human pituitary. FEBS letters 1996, 386, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Jockers, R.; Maurice, P.; Boutin, J.A.; Delagrange, P. Melatonin receptors, heterodimerization, signal transduction and binding sites: what's new? British journal of pharmacology, 2008, 154, 1182–1195. [Google Scholar] [CrossRef] [PubMed]
- Levoye, A.; Dam, J.; Ayoub, M.A.; Guillaume, J.L.; Couturier, C.; Delagrange, P.; Jockers, R. The orphan GPR50 receptor specifically inhibits MT1 melatonin receptor function through heterodimerization. The EMBO J 2006, 25, 3012–3023. [Google Scholar] [CrossRef] [PubMed]
- Boutin, J. A. Quinone reductase 2 as a promising target of melatonin therapeutic actions. Expert opinion on therapeutic targets 2016, 20, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Argueta, J.; Solís-Chagoyán, H.; Estrada-Reyes, R.; Constantino-Jonapa, L.A.; Oikawa-Sala, J.; Velázquez-Moctezuma, J.; Benítez-King, G. Further Evidence of the Melatonin Calmodulin Interaction: Effect on CaMKII Activity. Int J Mol Sci 2022, 23, 2479. [Google Scholar] [CrossRef] [PubMed]
- Targhazeh, N.; Reiter, R.J.; Rahimi, M.; Qujeq, D.; Yousefi, T.; Shahavi, M.H.; Mir, S.M. Oncostatic activities of melatonin: Roles in cell cycle, apoptosis, and autophagy. Biochimie 2022, 200, 44–59. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Kang, J.; Fan, W.; He, H.; Huang, F. ROR: Nuclear Receptor for Melatonin or Not? Molecules (Basel, Switzerland) 2021, 26, 2693. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Mayo, J.C.; Tan, D.X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: under promises but over delivers. J Pin Res 2016, 61, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Reiter, R.J. Melatonin and its metabolites vs oxidative stress: From individual actions to collective protection. J Pineal Res. 2018, 65, e12514. [Google Scholar] [CrossRef] [PubMed]
- Boutin, J.A.; Kennaway, D.J.; Jockers, R. Melatonin: Facts, Extrapolations and Clinical Trials. Biomolecules 2023, 13, 943. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R.; Pandi-Perumal, S.R. Melatonin, a potent agent in antioxidative defense: actions as a natural food constituent, gastrointestinal factor, drug and prodrug. Nutrition & metabolism 2005, 2, 22. [Google Scholar] [CrossRef]
- Ferlazzo, N.; Andolina, G.; Cannata, A.; Costanzo, M.G.; Rizzo, V.; Currò, M.; Ientile, R.; Caccamo, D. Is Melatonin the Cornucopia of the 21st Century? Antioxidants 2020, 9, 1088. [Google Scholar] [CrossRef] [PubMed]
- Csaba, G.; Bodoky, M.; Fischer, J.; Acs, T. The effect of pinealectomy and thymectomy on the immune capacity of the rat. Experientia 1966, 22, 168–169. [Google Scholar] [CrossRef] [PubMed]
- Janković, B.D.; Isaković, K.; Petrović, S. Effect of pinealectomy on immune reactions in the rat. Immunology 1970, 1970 18, 1–6. [Google Scholar]
- Maestroni, G.J.; Conti, A.; Pierpaoli, W. Role of the pineal gland in immunity. Circadian synthesis and release of melatonin modulates the antibody response and antagonizes the immunosuppressive effect of corticosterone. J. Neuroimmunol. 1986, 13, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Maestroni, G.J.; Conti, A.; Pierpaoli, W. Role of the pineal gland in immunity: II. Melatonin enhances the antibody response via an opiatergic mechanism. Clinical Exp Immunol 1987, 68, 384–391. [Google Scholar]
- Maestroni, G.J.; Conti, A.; Pierpaoli, W. Role of the pineal gland in immunity. III. Melatonin antagonizes the immunosuppressive effect of acute stress via an opiatergic mechanism. Immunology 1988, 63, 465–469. [Google Scholar] [PubMed]
- Moore, C.B.; Siopes, T.D. Melatonin enhances cellular and humoral immune responses in the Japanese quail (Coturnix coturnix japonica) via an opiatergic mechanism. Gen Comp Endocrinol 2003, 131, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Majewski, P.; Dziwinski, T.; Pawlak, J.; Waloch, M.; Skwarlo-Sonta, K. (2005). Anti-inflammatory and opioid-mediated effects of melatonin on experimental peritonitis in chickens. Life Sci 2005, 76, 1907–1920. [Google Scholar] [CrossRef] [PubMed]
- Liebmann, P.M.; Wölfler, A.; Felsner, P.; Hofer, D.; Schauenstein, K. Melatonin and the immune system. Intern arch aller Immunol 1997, 112, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Calvo, J.R.; Karbownik, M.; Qi, W.; Tan, D.X. Melatonin and its relation to the immune system and inflammation. Annals NY Acad Sci 2000, 917, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.B.; Siopes, T.D. Melatonin enhances cellular and humoral immune responses in the Japanese quail (Coturnix coturnix japonica) via an opiatergic mechanism. Gen Comp Endocrinol 2003, 131, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, D.P.; Esquifino, A.I.; Srinivasan, V.; Pandi-Perumal, S.R. Melatonin and the immune system in aging. Neuroimmunomodulation 2008, 15, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Vico, A.; Lardone, P.J.; Alvarez-Sánchez, N.; Rodríguez-Rodríguez, A.; Guerrero, J.M. Melatonin: buffering the immune system. Int J Mol Sci 2013, 14, 8638–8683. [Google Scholar] [CrossRef] [PubMed]
- Claustrat, B.; Leston, J. Melatonin: Physiological effects in humans. Neuro-Chirurgie 2015, 61, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Aging, Melatonin, and the Pro- and Anti-Inflammatory Networks. Int J Mol Sci 2019, 20, 1223. [Google Scholar] [CrossRef] [PubMed]
- Fiore, A.; Murray, P.J. Tryptophan and indole metabolism in immune regulation. Curr Opin Immunol 2021, 70, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Moslehi, M.; Moazamiyanfar, R.; Dakkali, M.S.; Rezaei, S.; Rastegar-Pouyani, N.; Jafarzadeh, E.; Mouludi, K.; Khodamoradi, E.; Taeb, S.; Najafi, M. Modulation of the immune system by melatonin; implications for cancer therapy. Int Immunopharmacol 2022, 108, 108890. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Redox Biology of Melatonin: Discriminating Between Circadian and Noncircadian Functions. Antioxid Redox Signal 2022, 37, 704–725. [Google Scholar] [CrossRef] [PubMed]
- García-García, A.; Méndez-Ferrer, S. The Autonomic Nervous System Pulls the Strings to Coordinate Circadian HSC Functions. Front Immunol. 20 May. [CrossRef]
- Shimba, A.; Ikuta, K. Glucocorticoids Regulate Circadian Rhythm of Innate and Adaptive Immunity. Front Immunol 2020, 11, 2143. [Google Scholar] [CrossRef] [PubMed]
- Scheiermann, C.; Kunisaki, Y.; Frenette, P.S. Circadian control of the immune system. Nat Rev Immunol 2013, 13, 190–198. [Google Scholar] [CrossRef]
- Holtkamp, S.J.; Ince, L.M.; Barnoud, C.; Schmitt, M.T.; Sinturel, F.; Pilorz, V.; Pick, R.; Jemelin, S.; Mühlstädt, M.; Boehncke, W.H.; Weber, J.; Laubender, D.; Philippou-Massier, J.; Chen, C.S.; Holtermann, L.; Vestweber, D.; Sperandio, M.; Schraml, B.U.; Halin, C.; Dibner, C.; … Scheiermann, C. Circadian clocks guide dendritic cells into skin lymphatics. Nat Immunol 2021, 22, 1375–1381. [Google Scholar] [CrossRef] [PubMed]
- Pevet, P.; Challet, E. Melatonin: both master clock output and internal time-giver in the circadian clocks network. J Physiol Paris 2011, 105, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.C.; Pandi-Perumal, S.R.; Esquifino, A.I.; Cardinali, D.P.; Maestroni, G.J. The role of melatonin in immuno-enhancement: potential application in cancer. Intern J Exp Pathol 2006, 87, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Csaba, G. The pineal regulation of the immune system: 40 years since the discovery. Acta Microbiol Immunol Hung 2013, 60, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G. Melatonin, BAG-1 and cortisol circadian interactions in tumor pathogenesis and patterned immune responses. Expl Target Antitumor Ther 2023, 4, 962–993. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Haldar, C. Physiological crosstalk between melatonin and glucocorticoid receptor modulates T-cell mediated immune responses in a wild tropical rodent, Funambulus pennanti. J Steroid Biochem Mol Bio 2013, 2013 134, 23–36. [Google Scholar] [CrossRef]
- Hardeland, R. (2019). Aging, Melatonin, and the Pro- and Anti-Inflammatory Networks. Int J Mol Sci 2019, 20, 1223. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Huang, S.Y.; Sun, J.H.; Zhang, H.C.; Cai, Q.L.; Gao, C.; Li, L.; Cao, J.; Xu, F.; Zhou, Y.; Guan, C.X.; Jin, S.W.; Deng, J.; Fang, X.M.; Jiang, J.X.; Zeng, L. Sepsis-induced immunosuppression: mechanisms, diagnosis and current treatment options. Mil Med Res 2022, 9, 56. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Opposite effects of melatonin in different systems and under different conditions. Curr. Top. Biochem. Res 2016, 17, 57–69. [Google Scholar]
- Markus, R.P.; Fernandes, P.A.; Kinker, G.S.; da Silveira Cruz-Machado, S.; Marçola, M. Immune-pineal axis - acute inflammatory responses coordinate melatonin synthesis by pinealocytes and phagocytes. Br J Pharm 2018, 175, 3239–3250. [Google Scholar] [CrossRef] [PubMed]
- Maestroni, G.J. Neurohormones and catecholamines as functional components of the bone marrow microenvironment. Ann NY Acad Sci 2000, 917, 29–37. [Google Scholar] [CrossRef]
- Lissoni, P.; Paolorossi, F.; Ardizzoia, A. : Barni, S., Chilelli, M.; Mancuso, M., Tancini, G.,; Conti, A.; Maestroni, G.J. A randomized study of chemotherapy with cisplatin plus etoposide versus chemoendocrine therapy with cisplatin, etoposide and the pineal hormone melatonin as a first-line treatment of advanced non-small cell lung cancer patients in a poor clinical state. J Pineal Res 1997, 23(1), 15–19. [CrossRef]
- Golan, K.; Kumari, A.; Kollet, O.; Khatib-Massalha, E.; Subramaniam, M.D.; Ferreira, Z.S.; Avemaria, F.; Rzeszotek, S.; García-García, A.; Xie, S.; Flores-Figueroa, E.; Gur-Cohen, S.; Itkin, T.; Ludin-Tal, A.; Massalha, H.; Bernshtein, B.; Ciechanowicz, A.K.; Brandis, A.; Mehlman, T.; Bhattacharya, S.; … Lapidot, T. Daily Onset of Light and Darkness Differentially Controls Hematopoietic Stem Cell Differentiation and Maintenance. Cell stem cell. [CrossRef]
- Mafi, A.; Rezaee, M.; Hedayati, N.; Hogan, S.D.; Reiter, R.J.; Aarabi, M.H.; Asemi, Z. Melatonin and 5-fluorouracil combination chemotherapy: opportunities and efficacy in cancer therapy. Cell Comm Signal : CCS. [CrossRef]
- Maestroni, G. Circadian regulation of the immune-hematopoietic system. Expl Neurosci 2023, 2, 123–139. [Google Scholar] [CrossRef]
- Maestroni, G.J.; Conti, A.; Pierpaoli, W. (1988). Pineal melatonin, its fundamental immunoregulatory role in aging and cancer. Ann NY Acad Sci 1988, 521, 140–148. [CrossRef]
- Ben-Nathan, D.; Maestroni, G.J.; Lustig, S.; Conti, A. Protective effects of melatonin in mice infected with encephalitis viruses. Archi Virol 1995, 140, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, E.; Valero-Fuenmayor, N.; Pons, H.; Chacín-Bonilla, L. Melatonin protects mice infected with Venezuelan equine encephalomyelitis virus. Cell Mol Life Sci 1997, 53, 430–434. [Google Scholar] [CrossRef]
- Valero, N.; Bonilla, E.; Pons, H.; Chacin-Bonilla, L.; Añez, F.; Espina, L.M.; Medina-Leendertz, S.; García Tamayo, J. Melatonin induces changes to serum cytokines in mice infected with the Venezuelan equine encephalomyelitis virus. Trans R Soc Trop Med Hyg 2002, 96, 348–351. [Google Scholar] [CrossRef] [PubMed]
- Nunnari, G.; Nigro, L.; Palermo, F.; Leto, D.; Pomerantz, R.J.; Cacopardo, B. Reduction of serum melatonin levels in HIV-1-infected individuals' parallel disease progression: correlation with serum interleukin-12 levels. Infection 2003, 31, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.H.; Cao, X.J.; Wei, W. Melatonin decreases TLR3-mediated inflammatory factor expression via inhibition of NF-kappa B activation in respiratory syncytial virus-infected RAW264.7 macrophages. J Pineal Res 2008, 45, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.H.; Cao, X.J.; Liu, W.; Shi, X.Y.; Wei, W. Inhibitory effect of melatonin on lung oxidative stress induced by respiratory syncytial virus infection in mice. J Pineal Res 2010, 48, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Laliena, A.; San Miguel, B.; Crespo, I.; Alvarez, M.; González-Gallego, J.; Tuñón, M.J. Melatonin attenuates inflammation and promotes regeneration in rabbits with fulminant hepatitis of viral origin. J Pineal Res 2012, 53, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Crespo, I.; Fernández-Palanca, P.; San-Miguel, B.; Álvarez, M.; González-Gallego, J.; Tuñón, M.J. Melatonin modulates mitophagy, innate immunity and circadian clocks in a model of viral-induced fulminant hepatic failure. J Cell Mol Med 2020, 24, 7625–7636. [Google Scholar] [CrossRef] [PubMed]
- Junaid, A.; Tang, H.; van Reeuwijk, A.; Abouleila, Y.; Wuelfroth, P.; van Duinen, V.; Stam, W.; van Zonneveld, A.J.; Hankemeier, T.; Mashaghi, A. Ebola Hemorrhagic Shock Syndrome-on-a-Chip. iScience 2020, 23, 100765. [Google Scholar] [CrossRef] [PubMed]
- Bahrampour Juybari, K.; Pourhanifeh, M.H.; Hosseinzadeh, A.; Hemati, K.; Mehrzadi, S. (2020). Melatonin potentials against viral infections including COVID-19: Current evidence and new findings. Virus Res 2020, 287, 198108. [Google Scholar] [CrossRef] [PubMed]
- Sehirli, A.O.; Sayiner, S.; Serakinci, N. Role of melatonin in the treatment of COVID-19; as an adjuvant through cluster differentiation 147 (CD147). Mol Biol Rep 2020, 47, 8229–8233. [Google Scholar] [CrossRef] [PubMed]
- Hasan, Z.T.; Atrakji, D.M.Q.Y.M.A.A.; Mehuaiden, D.A.K. The Effect of Melatonin on Thrombosis, Sepsis and Mortality Rate in COVID-19 Patients. Inter J Inf Dis , 2022, 114, 79–84. [CrossRef]
- Loh, D.; Reiter, R.J. Melatonin: Regulation of Viral Phase Separation and Epitranscriptomics in Post-Acute Sequelae of COVID-19. Int J Mol Sci 2022, 23, 8122. [Google Scholar] [CrossRef] [PubMed]
- Farnoosh, G.; Akbariqomi, M.; Badri, T.; Bagheri, M.; Izadi, M.; Saeedi-Boroujeni, A.; Rezaie, E.; Ghaleh, H.E.G.; Aghamollaei, H.; Fasihi-Ramandi, M.; Hassanpour, K.; Alishiri, G. Efficacy of a Low Dose of Melatonin as an Adjunctive Therapy in Hospitalized Patients with COVID-19: A Randomized, Double-blind Clinical Trial. Arch Med Res 2022, 53, 79–85. [Google Scholar] [CrossRef]
- Ameri, A.; Frouz Asadi, M.; Ziaei, A.; Vatankhah, M.; Safa, O.; Kamali, M.; Fathalipour, M.; Mahmoodi, M.; Hassanipour, S. (2023). Efficacy and safety of oral melatonin in patients with severe COVID-19: a randomized controlled trial. Inflammopharmacology 2023, 31(1), 265–274. [CrossRef]
- Sánchez-Rico, M.; de la Muela, P.; Herrera-Morueco, J.J.; Geoffroy, P.A.; Limosin, F.; Hoertel, N. ; AP-HP/Université de Paris/INSERM COVID-19 Research Collaboration/AP-HP COVID CDR Initiative/Entrepôt de Données de Santé AP-HP Consortium Melatonin does not reduce mortality in adult hospitalized patients with COVID-19: a multicenter retrospective observational study. J Travel Med, 2022, 29(3), taab195.
- https://doi.org/10.1093/jtAlizadeh, N.; Dianatkhah, M.; Alimohamadi, Y.; Moradi, H.; Akbarpour, S.; Akrami, M.; Mansouri, F.; Faraji, N.; Rezaie, Z.; Alizadeh, M.; Hosamirudsari, H. High dose melatonin as an adjuvant therapy in intubated patients with COVID-19: A randomized clinical trial. J Taibah Uni Med Sci 2022,17(3), 454–460. https://doi.org/10.1016/j.jtumed.2022.04.012.
- Darban, M.; Malek, F.; Memarian, M.; Gohari, A.; Kiani, A.; Emadi, A.; Bagheri, B. Efficacy of high dose vitamin C, melatonin and zinc in Iranian patients with acute respiratory syndrome due to coronavirus infection: a pilot randomized trial. J. Cell. Mol. Anesth 2021, 6, 164–167. [Google Scholar]
- Hing-Hwa,H.;, Ching-Len,L.; Shyi-Jou,C.; Li-Ge,S.; Li,L.;, Yuan-Wu,C.;, Chia-Pi,C.; Huey-Kang,S.; Shih-Ta,S.;, Gu-Jiun, L. Melatonin possesses an anti-influenza potential through its immune modulatory effect,J Fun Foods 2019, 58, 189-198. [CrossRef]
- Xu, M.M.; Kang, J.Y.; Ji, S.; Wei, Y.Y.; Wei, S.L.; Ye, J.J.; Wang, Y.G.; Shen, J.L.; Wu, H.M.; Fei, G.H. Melatonin Suppresses Macrophage M1 Polarization and ROS-Mediated Pyroptosis via Activating ApoE/LDLR Pathway in Influenza A-Induced Acute Lung Injury. Ox Med Cell Longev 2022, 2022, 2520348. [Google Scholar] [CrossRef] [PubMed]
- Huo, C.; Tang, Y.; Li, X.; Han, D.; Gu, Q.; Su, R.; Liu, Y.; Reiter, R.J.; Liu, G.; Hu, Y.; Yang, H. Melatonin alleviates lung injury in H1N1-infected mice by mast cell inactivation and cytokine storm suppression. PLoS pathogens 2023, 19, e1011406. [Google Scholar] [CrossRef] [PubMed]
- Maestroni, G. J. Melatonin as a therapeutic agent in experimental endotoxic shock. J Pineal Res. [CrossRef]
- Colunga Biancatelli, R.M.L.; Berrill, M.; Mohammed, Y.H.; Marik, P.E. Melatonin for the treatment of sepsis: the scientific rationale. J Thoracic Dis 2020, 12 (Suppl 1), S54–S65. [Google Scholar] [CrossRef] [PubMed]
- Poggi, C.; Dani, C. Sepsis and Oxidative Stress in the Newborn: From Pathogenesis to Novel Therapeutic Targets. Oxid Med Cell Long 2018, 9390140. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhang, W.; Kwak, M.; Zhang, L.; Lee, P.C.W.; Jin, J.O. Protective Effect of Melatonin Against Polymicrobial Sepsis Is Mediated by the Anti-bacterial Effect of Neutrophils. Front Immunol 2019, 10, 1371. [Google Scholar] [CrossRef] [PubMed]
- Bishayi, B.; Adhikary, R.; Nandi, A.; Sultana, S. Beneficial Effects of Exogenous Melatonin in Acute Staphylococcus aureus and Escherichia coli Infection-Induced Inflammation and Associated Behavioral Response in Mice After Exposure to Short Photoperiod. Inflammation 2016, 39, 2072–2093. [Google Scholar] [CrossRef]
- Du, L.; Liu, B.; Han, Z.; Xia, Y.; Wu, M.; Liu, S. Melatonin shapes bacterial clearance function of porcine macrophages during enterotoxigenic Escherichia coli infection. Anim Nut 2022, 11, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Xu, S.; Wu, H.; Liu, J.; Wang, Y.; Zhu, G. (2022). Melatonin Is Neuroprotective in Escherichia coli Meningitis Depending on Intestinal Microbiota. Int J Mol Sci 2023, 24, 298. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Sun, P. Insight of Melatonin: The Potential of Melatonin to Treat Bacteria-Induced Mastitis. Antioxidants (Basel, Switzerland), 2022, 11, 1107. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Liu, J.; Zhao, X.; Yang, W. Melatonin ameliorates lung cell inflammation and apoptosis caused by Klebsiella pneumoniae via AMP-activated protein kinase. Inflammopharmacol 2022, 30, 2345–2357. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, T.; Lv, X.; Sun, N. , Li, F.; Luo, L.; Zhuge, X.; Huang, J.; Wang, L. (A Potential Nontraditional Approach To Combat tmexCD1-toprJ1-Mediated Tigecycline Resistance: Melatonin as a Synergistic Adjuvant of Tigecycline. Antimicr Agent Chemother 2023, 67, e0004723. [Google Scholar] [CrossRef]
- Huang, H.; Mehta, A.; Kalmanovich, J.; Anand, A.; Bejarano, M.C.; Garg, T.; Khan, N.; Tonpouwo, G.K.; Shkodina, A.D.; Bardhan, M. Immunological and inflammatory effects of infectious diseases in circadian rhythm disruption and future therapeutic directions. Mol Biol Rep 2023, 50, 3739–3753. [Google Scholar] [CrossRef]
- Singh, M.K.; Dias, B.K.M.; Garcia, C.R. S Role of Melatonin in the Synchronization of Asexual Forms in the Parasite Plasmodium falciparum. Biomolecules 2020, 10, 1243. [Google Scholar] [CrossRef] [PubMed]
- Ataide, B.J.A.; Kauffmann, N.; Mendes, N.S.F.; Torres, M.L.M.; Dos Anjos, L.M.; Passos, A.D.C.F.; de Moraes, S.A.S.; Batista, E.J.O.; Herculano, A.M.; Oliveira, K.R.H.M. (Melatonin Prevents Brain Damage and Neurocognitive Impairment Induced by Plasmodium Berghei ANKA Infection in Murine Model of Cerebral Malaria. Front Cell Infect Microbiol 2020, 10, 541624. [Google Scholar] [CrossRef] [PubMed]
- Bagnaresi, P.; Alves, E.; Borges da Silva, H.; Epiphanio, S.; Mota, M.M.; Garcia, C.R. (Unlike the synchronous Plasmodium falciparum and P. chabaudi infection, the P. berghei and P. yoelii asynchronous infections are not affected by melatonin. Int J Gen Med. [CrossRef]
- Bocchi, E.A.; Bestetti, R.B.; Scanavacca, M.I.; Cunha Neto, E.; Issa, V.S. Chronic Chagas Heart Disease Management: From Etiology to Cardiomyopathy Treatment J Am Coll Cardiol 2017, 70, 1510–1524. [CrossRef]
- Santello, F.H.; Frare, E.O.; dos Santos, C.D.; Toldo, M.P.; Kawasse, L.M.; Zucoloto, S.; do Prado, J.C. Jr. Melatonin treatment reduces the severity of experimental Trypanosoma cruzi infection. J Pineal Res 2007, 42, 359–363. [Google Scholar] [CrossRef]
- Santello, F.H.; Frare, E.O.; dos Santos, C.D.; Caetano, L.C.; Alonso Toldo, M.P.; do Prado, J.C. , Jr. Suppressive action of melatonin on the TH-2 immune response in rats infected with Trypanosoma cruzi. J Pineal Res 2008, 45(3), 291–296. [CrossRef]
- Santello, F.H.; Frare, E.O.; Caetano, L.C.; AlonsoToldo, M.P.; do Prado, J.C. Jr Melatonin enhances pro-inflammatory cytokine levels and protects against Chagas disease. J Pineal Res 2008, 45, 79–85. [Google Scholar] [CrossRef]
- Brazão, V.; Del Vecchio Filipin, M.; Santello, F.H.; Caetano, L.C.; Abrahão, A.A.; Toldo, M.P.; do Prado, J.C. Jr.Melatonin and zinc treatment: distinctive modulation of cytokine production in chronic experimental Trypanosoma cruzi infection. Cytokine 2011, 56(3), 627–632. [CrossRef]
- Providello, M.V. , Portapilla, G.B., Oliveira, P.A.S., da Silva, C.B.P., Anchieta, N.F., Tirapelli, C.R., de Albuquerque, S. Melatonin decreases circulating Trypanosoma cruzi load with no effect on tissue parasite replication. Can J Physiol Pharmacol 2021, 99, 795–802. [Google Scholar] [CrossRef]
- Hunter, F.K.; Butler, T.D.; Gibbs, J.E. Circadian rhythms in immunity and host-parasite interactions. Parasite Immunol 2022, 44, e12904. [Google Scholar] [CrossRef]
- Grassi-Zucconi, G.; Semprevivo, M.; Mocaer, E.; Kristensson, K.; Bentivoglio, M. Melatonin and its new agonist S-20098 restore synchronized sleep fragmented by experimental trypanosome infection in the rat. Brain Res Bull 1996, 39, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat Rev Immunol 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Elmahallawy, E.K.; Jiménez-Aranda, A.; Martínez, A.S.; Rodriguez-Granger, J.; Navarro-Alarcón, M.; Gutiérrez-Fernández, J.; Agil, A. Activity of melatonin against Leishmania infantum promastigotes by mitochondrial dependent pathway. Chemico-biol Interac 2014, 220, 84–93. [CrossRef]
- Laranjeira-Silva, M.F.; Zampieri, R.A.; Muxel, S.M.; Floeter-Winter, L.M.; Markus, R.P. Melatonin attenuates Leishmania (L.) amazonensis infection by modulating arginine metabolism. J Pineal Res 2015, 59(4), 478–487. [CrossRef]
- Fernandes, J.C.R.; Aoki, J.I.; Maia Acuña, S.; Zampieri, R.A.; Markus, R.P.; Floeter-Winter, L.M.; Muxel, S.M. Melatonin and Leishmania amazonensis Infection Altered miR-294, miR-30e, and miR-302d Impacting on Tnf, Mcp-1, and Nos2 Expression. Front Cell Inf Microbiol 2019, 9, 60. [Google Scholar] [CrossRef]
- Parvez, S.; Yadagiri, G.; Arora, K.; Javaid, A.; Kushwaha, A.K.; Singh, O.P.; Sundar, S.; Mudavath, S.L. (Coalition of Biological Agent (Melatonin) With Chemotherapeutic Agent (Amphotericin B) for Combating Visceral Leishmaniasis via Oral Administration of Modified Solid Lipid Nanoparticles. ACS Bio,mater Sci Eng 2023, 9(6), 2902–2910. [CrossRef]
- Szewczyk-Golec, K.; Pawłowska, M.; Wesołowski, R.; Wróblewski, M.; Mila-Kierzenkowska, C. Oxidative Stress as a Possible Target in the Treatment of Toxoplasmosis: Perspectives and Ambiguities. Int J Mol Sci 2021, 22, 5705. [Google Scholar] [CrossRef] [PubMed]
- Avunduk, A.M.; Avunduk, M.C.; Baltaci, A.K.; Moğulkoç, R. Effect of melatonin and zinc on the immune response in experimental Toxoplasma retinochoroiditis. Ophthalmologica. Int J Ophtal. 2007, 221, 421–425. [Google Scholar] [CrossRef]
- Baltaci, A.K.; Bediz, C.S.; Mogulkoc, R.; Kurtoglu, E.; Pekel, A. Effect of zinc and melatonin supplementation on cellular immunity in rats with toxoplasmosis. Biol Trace Elem Res 2003, 96(1-3), 237–245. [CrossRef]
- Baltaci, A.K.; Mogulkoc, R.; Bediz, C.S. , Pekel, A.Effects of zinc deficiency and pinealectomy on cellular immunity in rats infected with Toxoplasma gondii. Biol Trace Elem Res 2005, 104, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Machado, N.I.; Dos Santos, T.A.T.; de Souza, W.; DaMatta, R.A.; Seabra, S.H. Treatment with melatonin induces a reduction of Toxoplasma gondii development in LLC-MK2 cells. Parasitol Res 2020, 119, 2703–2711. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, T.; Sharma, S.; Kumar, M.; Hussain, M.A.; Chauhan, N.; Kalia, I.; Sahu, A.K.; Rana, V.S.; Bharti, R.; Haldar, A.K.; Singh, A.P.; Mazumder, S. Tryptophan-kynurenine pathway attenuates β-catenin-dependent pro-parasitic role of STING-TICAM2-IRF3-IDO1 signalosome in Toxoplasma gondii infection. Cell Death Dis 2019, 10, 161. [Google Scholar] [CrossRef] [PubMed]
- Talib, W.H.; Alsayed, A. R.; Abuawad, A.; Daoud, S.; Mahmod, A.I. Melatonin in Cancer Treatment: Current Knowledge and Future Opportunities. Molecules 2021, 26, 2506. [CrossRef]
Figure 1.
The indirect and direct immunoenhancing action of MLT.
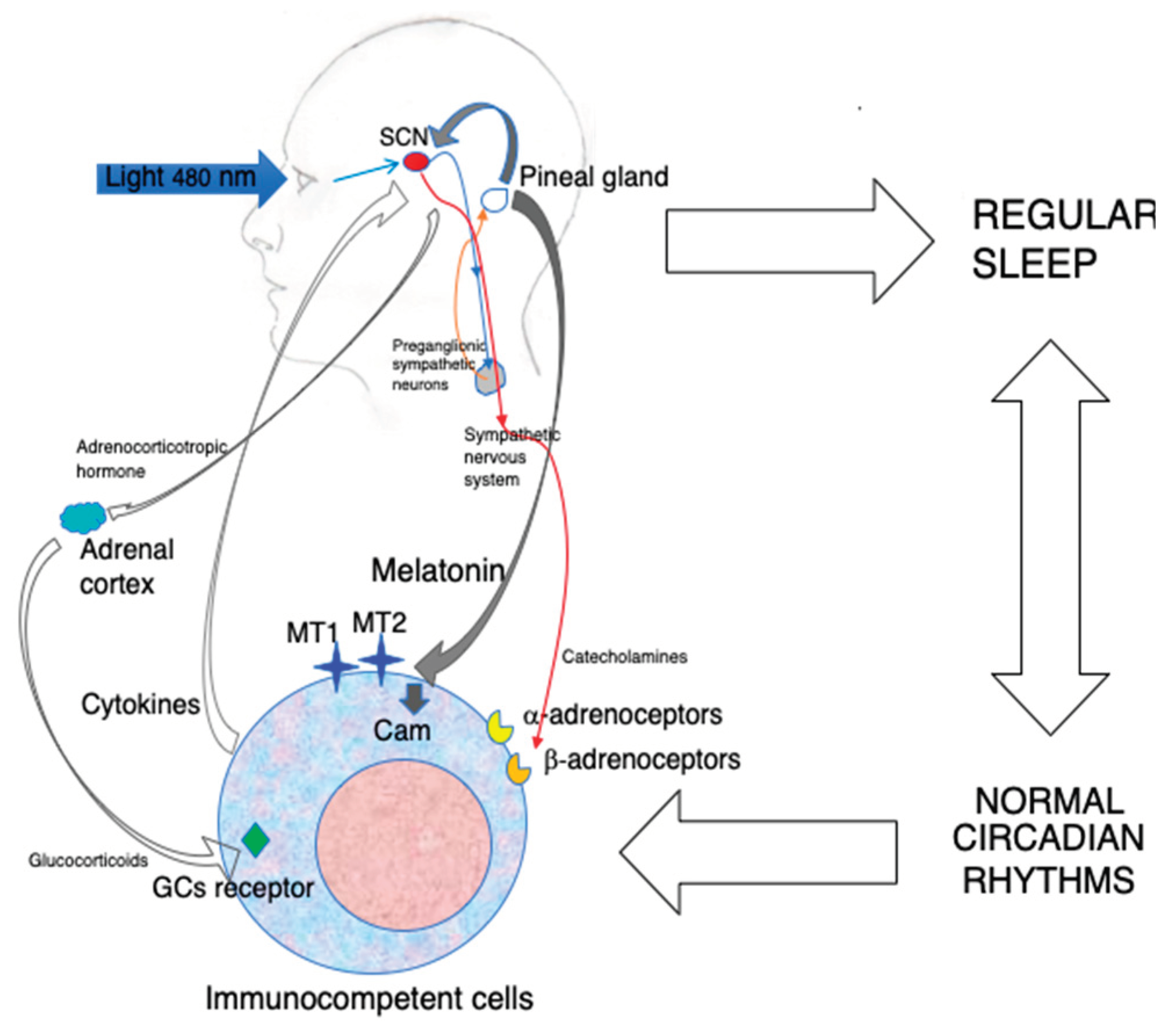
Figure 2.
A possible experimental approach integrating the anti-inflammatory and immunoenhancing actions of MLT against infections.
Figure 2.
A possible experimental approach integrating the anti-inflammatory and immunoenhancing actions of MLT against infections.
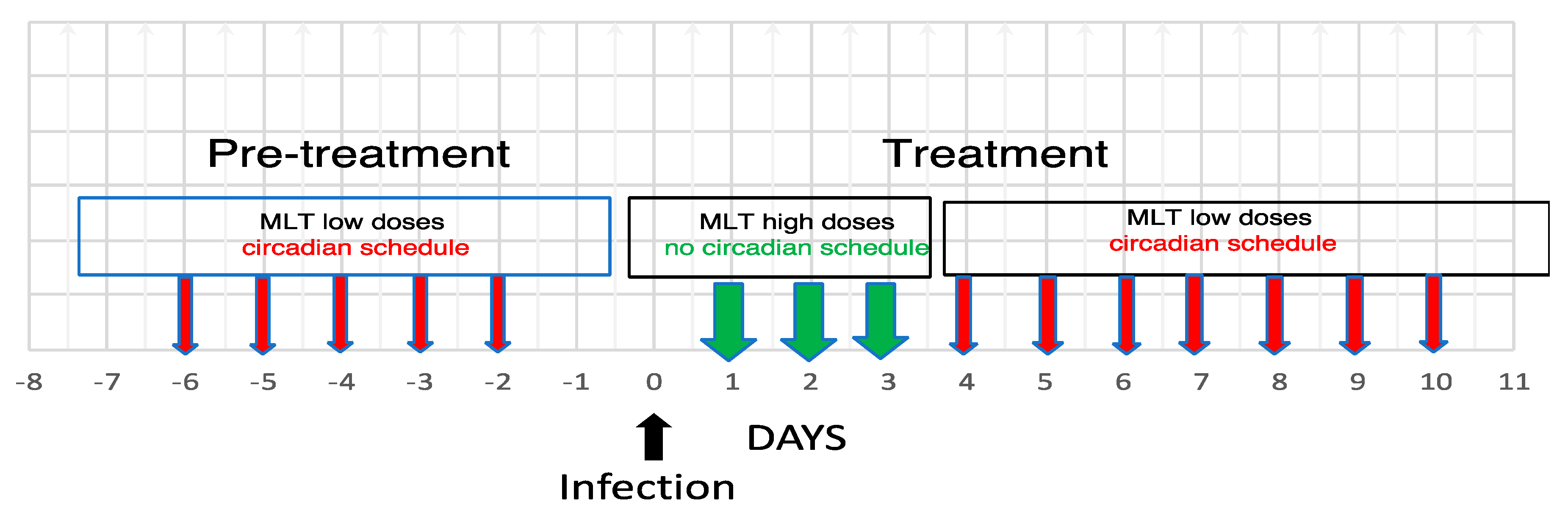

Table 1.
In vivo preclinical studies on the anti-viral activity of MLT.
| Pathogen | Species | MLT dose | Treatment | Outcome | Ref. |
|---|---|---|---|---|---|
| EMCV | Mice | 1ug /mouse | 10 days at 4 pm | Reversal of stress -induced death | [55] |
| SFV | Mice | 500 ug /kg | From 3 days before until 10 days after infection at 4 pm | Increased survival and decreased viremia | [56] |
| aWNV | Mice | 5ug /mouse | From 2 days before until 8 days after infection at 4 pm | Reduced mortality | [56] |
| VEEV | MIce | 1 mg/kg | From 3 days before until 10 days after infection at 6 pm | Increased survival, decreased viremia, increased antibody response | [57] |
| RSV | Mice | 5mg/kg | Twice daily for 3 days | Reduced oxidative damage of the lung | [61] |
| RHDV | Rabbits | 20 mg/kg | 0, 12, 24 h after infection | Decreased mitophagy, inflammation and innate immunity | [62,63] |
| H1N1 | Mice | 3, 10, 30 mg/kg | Pretreatment for 3 days before infection | Decreased lung injury by inhibition of mast cells and cytokine storm | [76] |
| H1N1 | Mice | 200 mg/kg | 6 h before and 2,4 and 6 days post infection | Inhibition of pro-inflammatory cytokines and stimulation of IL-10; Synergy with an antiviral drug | [74] |
| H3N2 | Mice | 30 mg/kg | 7 days at 6 pm | Attenuated pulmonary damage, leukocyte infiltration and edema | [75] |
The features of the existing preclinical studies on the possible therapeutic effect of MLT in viral infections are reported.EMCV: Encephalomyocarditis virus; SFV: Semliki forest virus; aWNV: attenuated West Nile Virus; VEEV: Venezuelan Equine Encephalitis Virus; RSV: Respiratory Syncitial Virus; RHDV: rabbit hemorrhagic disease virus; H1N1, Influenza A H1N1; H3N2: influenza A H3N2.
Table 2.
Preclinical and clinical studies on the anti-bacterial effect of MLT.
| Pathogen | Species | MLT dose | Treatment | Outcome | Ref. |
|---|---|---|---|---|---|
| Lethal dose of LPS | Mice | 1,2,3,4,5,10 mg/kg | 3 or 6 h after LPS injection | 2,3,4, 5 mg/kg reduced mortality and NO synthesis | [77] |
| Sepsis | HumanNewborns | 2 x 10 mg | Oral administration within 12 hours after diagnosis | Increased survival and improved clinical status | [78] |
| Sepsis | Human newborns | 20 mg/kg | One injection plus antibiotics | Increased survival and improved clinical status | [79] |
| Polymicrobial sepsis | Mice | 50 mg/kg | Two doses , 30 min before and 30 min after cecal ligation puncture | Protection of mice by induction of neutrophil extracellular trap | [80] |
| Staphylococcus aureus, Escherichia coli | Mice | 10mg/kg | Once daily for 7 days | Improved clearance of bacteria from blood, reduced iNOS, plasma C-reactive protein, COX2 expression in the hypothalamus . | [81] |
| Escherichia coli | Mice | 30mg/kg | Pretreatment for 7 consecutive days before infection | Prevention of and protection from bacterial meningitis by modulating the intestinal microbiota | [83] |
| Tigecyclin resistant Klebsiella pneumoniae | Mice | 50 mg/kg | One dose after infection | Restoring tigecycline activity | [86] |
The available preclinical and clinical studies on the therapeutic effect of MLT against bacterial infections are reported. NO. nitric oxide; iNOS: inducible nitric oxide synthetase; COX2: cyclooxygenase.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



