Submitted:
22 February 2024
Posted:
23 February 2024
You are already at the latest version
Abstract
Pseudomonas aeruginosa is an opportunistic pathogen that poses a significant threat to individuals with chronic respiratory ailments, such as cystic fibrosis (CF). The pathogen is highly prevalent in CF individuals and is responsible for chronic infection, resulting in severe tissue damage and poor patient outcome. However, early detection and immediate treatment are critical to prevent bacterial persistence and establishment of chronic infection. Presently, antibiotics including aminoglycosides, beta-lactams, polymyxins, and fluoroquinolones along with anti-inflammatory medications are being used to treat P. aeruginosa infections in CF patients. However, prolonged antibiotic administration has led to the emergence of widespread multidrug resistance in P. aeruginosa (CF isolates), contributing to antimicrobial resistance (AMR) and failure of antimicrobial therapies. Thus, exploring viable alternatives such as antivirulence therapeutics can be vital towards managing pseudomonal infections in CF patients and mitigating the risk of AMR. In this direction, antivirulence strategies achieving targeted inhibition of bacterial virulence pathways/factors, including quorum sensing, efflux pumps, lectins, and iron chelators, have been widely explored against CF isolates of P. aeruginosa. Simultaneously, there have been discussions about the sufficiency of in vitro findings and their successful translation in pulmonary infection models (in vivo) for the clinical implementation of antivirulence therapies hence, this review article presents a bird’s eye view on the pulmonary infections involving P. aeruginosa in CF patients by laying emphasis on factors contributing to bacterial colonization, persistence, and disease progression along with the current line of therapeutics against P. aeruginosa in CF while underscoring the limitation of antimicrobial approaches. We further collate scientific literature and comprehensively discuss the various antivirulence strategies that have been successfully tested against P. aeruginosa isolates from CF patients. This could pave the way towards a paradigm shift towards clinical therapy of pseudomonal infections in CF and can potentially override conventional antimicrobial treatments.
Keywords:
Cystic fibrosis
; Pseudomonas aeruginosa
; Antivirulence therapy
; Quorum quenching and biofilm inhibition
; Lectin and efflux pump inhibitors
; Iron chelation
1. Introduction: a brief overview on cystic fibrosis
Cystic fibrosis (CF) is an autosomal recessive disorder induced due to mutations in CF transmembrane regulator gene (CFTR), which is located on chromosome 7 [1]. The disease is a growing concern in today’s time as it affects 1 out of 3500 children in the Caucasian population [2]. In the past, this disease was primarily limited to childhood fatalities. However, with recent advancements in diagnostics and aggressive therapeutic approaches, disease prognosis has improved significantly, thereby increasing the survival rate in CF patients (10.1016/j.jcf.2013.12.002). The CFTR gene, first identified in 1989, is associated with the functioning of anion channels, which conducts chloride and bicarbonate at the apical membrane of differential epithelia [3]. Furthermore, it maintains ion and water channels for regulating epithelial surface hydration. The bicarbonate anion plays a crucial role in unfolding of mucins (airway), neutralizing gastric acidity, and activating pancreatic enzymes [4]. CFTR is also important for many physiological mechanisms related to natural immunity, including internalization of adhered bacterial pathogens and mitigating local inflammatory responses in bronchial epithelial cells [5]. The CFTR gene is expressed by a multitude of organs such as lungs, liver, pancreas, bones, intestine, etc., and hence its dysregulation can functionally affect their pivotal activities. Several complications including malignancies, CF-related diabetes, CF-related bone disorders, male infertility, bowel cancer and leukemia can result from mutations in the CFTR gene [6,7]. Based on the mechanism of dysfunction and repercussions on the protein function, CFTR mutations have been classified into six groups [8]: (i) Class I (protein production mutations), arising from premature termination codon due to frameshift mutations, thereby resulting in the complete loss of CFTR gene function; (ii) Class II (protein processing mutations), which are responsible for ineffective processing and transportation of CFTR protein. The most common mutant allele identified is the F508del (∆F508); (iii) Class III (gating mutations) that result in irregular opening of ion channels; (iv) Class IV (conduction mutations) that prevent the translocation of ions through the transport channel; (v) Class V (insufficient protein mutations), wherein the production of CFTR protein is abnormally reduced beyond regular levels; and (vi) Class VI (protein destabilizing mutations) that lead to reduced conformational stability and premature recycling of the unstable CFTR protein [9]. While genetic modulation of the CFTR gene is a primary factor responsible for CF, there are other predisposing factors such as polycystic kidney disease and secretory diarrhea [10]. In such cases, the activity of CFTR protein is observed to be higher than normal levels, which may ultimately lead to development of tumors [11].
Additionally, malfunctioning of the CFTR gene in bronchial epithelia has been shown to promote host natural defenses, exacerbating inflammatory responses which result in progressive lung damage [12]. Unregulated sodium absorption via gated channels also stimulates lowered airway-surface liquid volume and dysfunction of mucociliary clearance, thereby inhibiting the mechanisms for bacterial clearance in the lungs and upper airways [13]. Subsequently, colonization of the respiratory tract by mixed bacterial and fungal communities is promoted, which in-turn stimulates the decline of pulmonary function in CF patients [14,15]. Bacterial colonizers include Pseudomonas aeruginosa, Staphylococcus aureus, Haemophilus influenzae, Burkhloderia cepacia, and non-tuberculosis Mycobacterium sp.; while fungal invaders primarily consist of Aspergillus sp., Candida sp., Exophiala sp., and Scedosporium sp. [16]. Moreover, the prevalence of viral pathogens like adenoviruses, human rhinovirus, influenza A and B, parainfluenza and respiratory syncytial virus in CF patients is also well documented [17,18]. However, P. aeruginosa is the most prevalent microorganism associated with accelerated loss of lung function in CF patients [19]. The pathogen is acquired at an early stage from environmental sources, primarily exhibiting non-mucoid phenotype with considerable antibiotic sensitivity [20]. Nevertheless, due to extensive multidrug resistance (intrinsic and extrinsic), strong biofilm-forming abilities, and the presence of a sophisticated quorum sensing (QS)-regulated virulence artillery, the management of P. aeruginosa infection in CF patients becomes even more challenging [21]. Although there are antimicrobial therapies in place for preventing acquisition of bacterial infection, early eradication, timely control, and lowering pulmonary aggravation in CF patients, their long-term effectiveness and potency against P. aeruginosa has been a matter of serious concern among clinicians [22]. With the spurring incidence of antimicrobial resistance (AMR) among clinical pseudomonal strains [23], there is a pressing need to explore alternate strategies and therapeutics to combat the menace of P. aeruginosa infections in CF individuals. Hence, this review recapitulates the role of pseudomonal infections in aggravating CF, briefly discussing the mechanisms/factors for bacterial persistence and virulence. It also focuses on the current line of treatment (antimicrobial therapies) and collates scientific literature regarding the clinical limitations of existing strategies against P. aeruginosa. Furthermore, it also highlights the imperative need for devising next-generation approaches such as antivirulence therapy for the management of pseudomonal infections in CF patients.
2. Pseudomonas aeruginosa: the grim instigator in CF patients
2.1. Reservoir of infection and disease transmission
P. aeruginosa is a Gram-negative opportunistic microbe, frequently found in damp settings like drainage systems, vegetative fields, tap water, and surface soils [24]. Its prevalence is not only limited to general environmental settings, but also it extends to hospitals, home environment, cross infection between patients and family members, and nebulizers [24,25]. Hence, the ubiquitous existence of P. aeruginosa makes it hard to recognize the possible source of pseudomonal infection in CF patients.
Majority of P. aeruginosa infections are either community- or environmentally-acquired infections. In this regard, a study was conducted on children found in rural and metropolitan areas of Australia [26]. It revealed that children living in countryside had higher chances of acquiring P. aeruginosa infections. About 63% of the isolated strains obtained, originated likely from environment rather than being transmitted via nosocomial settings [26]. Cases of cross infection among CF patients living in close contact such as siblings, or those admitted in rehabilitation centres for longer periods of time have been reported [27,28]. Further, there are a handful of studies which have demonstrated that effective infection control methods reduce P. aeruginosa transmission amongst patients [29,30]. Despite advancements in preventing and controlling infections, two recent investigations have reported possible patient-to-patient transmission among 14-15% of patients at CF centers stipulated by the epidemiological ties between the pseudomonal strains isolated in each case. [31].
2.2. Colonization of P. aeruginosa in CF patients
The onset of pseudomonal infection in CF patients occurs in early childhood. Among the newly born CF individuals screened, approximately 53% exhibit positive P. aeruginosa cultures by the time they reach the age of 5 [20]. The establishment of such a notorious pathogen in the lungs is an outcome of multiple factors. In the respiratory tract of CF patients, compromised mucociliary clearance leads to diminished functionality of innate and non-cellular defense mechanisms. This significantly contributes as being one of the cardinal factors facilitating the colonization of P. aeruginosa in lungs. Additionally, defective CFTR gene is associated with reduced clearance of the pathogen from the respiratory tract [32]. CFTR impairment influences both the production as well as the quality of airway surface liquid (ASL). Due to reduced hydration of ASL, there is alteration in its composition resulting in thicker mucus which becomes less elastic and more challenging to clear, thereby creating a favorable environment for the colonization of pathogens in the lungs [33]. Furthermore, the impaired antimicrobial properties of ASL compromise its ability to detect pseudomonal flagella, facilitating colonization. Additionally, the flagella exhibit a pronounced affinity to asialoGM1, a surface molecule on airway cells with CFTR deficiency, further promoting infection in these cells [34].
The initial colonization of P. aeruginosa is marked by elevated inflammation resulting in tissue damage, thereby increasing the vulnerability of CF patients to pseudomonal infections [28]. Moreover, P. aeruginosa exacerbates airway inflammation and tissue damage via release of exoproducts, such as proteases, mucoid exopolysaccharides and exotoxin and cellular components like DNA, flagella, and lipopolysaccharide (LPS). Shedding of heparan sulfate Syndecan1 from epithelial cells induced by LasA further compounds this process [28,35]. Subsequent to tissue injury, P. aeruginosa utilizes type IV pili and flagella, for attachment to epithelial cells (Table 1). Conversely, chronic colonization by P. aeruginosa in CF is characterized by continuous persistance of the microbe for over six months and this results in provocation of a heightened antibody response [36]. It is also identified by the reduction of virulence factors and a transition to a biofilm growth. Further, a distinctive feature of persistent P. aeruginosa infection is production of mucoid alginate layer and formation of microcolonies in the lungs of patients [37]. Biofilm formation is an adaptive survival strategy employed by environmental bacteria, with alginate being a major component of the P. aeruginosa biofilm matrix. Mucoid layers may occur in patients with long term colonization of the pathogen, these are notably distinctive in CF patients.
2.3. Persistence of infection and adverse effects
Host factors such as socio-economic status, gender, impaired CFTR gene, and compromised immune defenses are mainly attributed to failure in eradicating P. aeruginosa. A funtional CFTR gene is a pre-requisite for monocytes and alveolar macrophages to acheive proper complement-mediated opsonization and lysosomal and phagosomal acidification [38]. Additionally, defective intraphagolysosomal HOCl production [39] and dysregulated Toll-like receptors (TLR) signaling are some of the ascertained clinical manifestations in CF pateints. TLR2, TLR4, and TLR5 are the primary TLR’s responsible for initiating an immune response to P. aeruginosa which identify bacterial cell wall components [40].
P. aeruginosa harbours a considerably larger genome comprising a higher proportion of genes that facilitate the bacteria to adapt and thrive in the lung environment [41]. It produces an array of bacterial components like toxins and virulence factors during the course of its infection which lead to its persistence in lungs [42]. The initiation of chronic respiratory tract infections can be influenced by some toxins. Particularly, virulence hallmarks like elastase and alkaline protease, which not only interfere with non-specific defense mechanisms (phagocytes) rather also affect specific immunologic defenses [43]. Certain bacterial pigments like pyocyanin can cause neutrophil apoptosis, making it harder for the body to eliminate P. aeruginosa (Table 1). The early colonization in CF patients is associated with lower levels of mannose-binding lectin 2 (MBL2), as the MBL2 gene transcribes a protein that aids bacterial opsonization and phagocytosis.
A unique defense mechanism employed by P. aeruginosa is quorum sensing (QS) which includes the Las, Rhl, and Pqs pathways [21]. They regulate the synthesis of virulence factors and the formation of biofilms in conjunction with each other. For instance, the LasIR quorum sensing system regulates the synthesis of the secondary metabolite pyocyanin [44]. This compound interferes with the macrophage’s ability to efficiently engulf apoptotic neutrophils, thereby intensifying the inflammatory process. The exopolysaccharides, including alginate, Psl, and Pel, help in formation of biofilm structure and safeguard the pathogen from the action of antibiotics and innate defense system. [45]. The role of alginate is to scavenge the hypochlorite and inhibit complement activation, neutrophil migration, chemotaxis, as well as macrophage phagocytosis and killing (Table 1). Psl functions by inhibiting complement-mediated opsonization of P. aeruginosa [46]. Also, it is instrumental in pseudomonal adherence to airway cells, thereby promoting enhanced cell cytotoxicity and cell death. Pel is crucial for pellicle formation at the air-liquid interface [47] and is associated with the wrinkled colony phenotype upon its elevated production (Table 1). Recent findings also highlight the role of Pel as a structural scaffold during the preliminary stages of P. aeruginosa PA14 biofilm formation [48]. The diverse virulence factors of P. aeruginosa that are primarily responsible for instigating chronic infection in CF patients have also been depicted in Figure 1.
It is evident that P. aeruginosa stands as the primary contributor to increased morbidity and fatality rates in CF patients. Its presence is linked to heightened respiratory symptoms [28,49] an increased frequency of exacerbations, elevated inflammation characterized by higher concentrations of neutrophil elastase in the sputum, elevated serum C-reactive protein (CRP) levels, and diminished monocyte responsiveness to various stimuli [50].

Table 1.
The role of QS-regulated pseudomonal virulence factors in chronic infection and their clinical implications in CF.
Table 1.
The role of QS-regulated pseudomonal virulence factors in chronic infection and their clinical implications in CF.
| S. No. | Virulence Factors | Clinical implications | References |
|---|---|---|---|
| 1 | Flagella/Pili | Binding of pili to asialoGM1 glycolipid residue, present in the respiratory epithelial cells of CF patients facilitates the attachment of P. aeruginosa, thereby leading to its colonization in the lungs | [34] |
| 2 | Exotoxin A (ETA) | ETA harbors ADP-ribosyltransferase (ADPRT) activity which catalyzes the transfer of ADP moiety to elongation factor 2, thereby stalling protein elongation and synthesis, resulting in cell intoxication | [51] |
| 3 | Alginate | P. aeruginosa CF isolates produces mucoid alginate and forms microcolonies in lungs (biofilms). Such alginate biofilms are resistant to antibiotics. Thus, it plays an important role in persistence of P. aeruginosa in the lungs of CF patients | [37] |
| 4 | Pyocyanin | Prevents clearance of P. aeruginosa infection in CF by mitigating ciliary beat frequency in airway epithelium. It inhibits catalase activity inducing oxidative stress on the respiratory epithelium further impairing CFTR function | [52] |
| 5 | LasB elastase (protease) | Important virulence factor for successful establishment of chronic infection. It provokes lung tissue damage and dysregulates host immune responses (down-regulation of CXCR1 on human neutrophils surface), and prevents the mucocilliary clearance of pathogen from the lung airway | [53,54] |
| 6 | Exopolysaccharides (Alginate, Psl, Pel) | Alginate serves as a chelators for hypochlorite and inhibits complement activation, neutrophil migration, chemotaxis, as well as macrophage phagocytosis and killing Psl enhances the attachment of P. aeruginosa to airway cells. Pel acts as a structural anchor in initial stages of biofilm formation and it enhances the resistance to aminoglycoside antibiotics |
[28,48] |
| 7 | Alkaline protease | It is responsible for the persistence of pathogen in the lungs by interfering with non-specific defense mechanisms like phagocytosis of the pathogen and specific immune responses such as T cells, natural killer cells and immunoglobulins. It inhibits neutrophil and monocyte function especially chemotaxis, which helps P. aeruginosa to escape the front line defenses of the host | [43] |
| 8 | Lectins | LecA and LecB, extracellular proteins primarily mediate adherence of P. aeruginosa to host cell surface during infection. LecA is involved in host cell invasion and cytotoxicity, while LecB reduces ciliary beating of airway epithelium. The presence of LecB leads to increased retention of cells and EPS in the biofilm. Along with Psl, LecB helps in establishment and stabilization of biofilm | [55] |
| 9 | Rhamnolipids | They play a major role in suppression of polymorphonuclear neutrophilic leucocytes (PMN’s) predominantly present during immune response against CF. Therefore, it eliminates the cellular host immune responses | [56] |
| 10 | Hydrogen cyanide | Acts as a repressor for cytochrome c oxidase in human cell mitochondria and inhibits several metalloenzymes. It is also used as a virulence biomarker (detected in exhaled breath) for patients who have acquired P. aeruginosa in cystic fibrosis infection as it is the only organism frequently found in the CF lung that produces hydrogen cyanide | [57,58] |
| 11 | Siderophores (Pyoverdine and Pyochelin) | Pyoverdine is responsible for chronic P. aeruginosa infection in CF patients. It not only act as an iron chelator but also stimulates septicemia, surface motility and biofilm maturation Pyochelin interacts with pyocyanin forming hydroxyl free radicals. These radicals damage the pulmonary artery endothelial and airway epithelial cells in humans, thereby contributing to pathogenicity in CF lungs |
[28,59,60] |
3. Current line of antimicrobial and alternative therapies: where we stand today
The primary objective of treating CF is to decrease mortality and reduce adverse effects of the disease, consequently preventing disease advancement. Presently, antibiotics including beta-lactams, aminoglycosides and polymyxins and fluoroquinolones, along with anti-inflammatory medications, are employed to manage pseudomonal infection and inflammatory responses in the respiratory tract [61]. In CF, antibiotics serve four main purposes: (i) preventing infection acquisition, (ii) eradicating early infections, (iii) controlling chronic infections, and (iv) treating pulmonary exacerbations. These antibiotics are administered through three primary routes: oral, intravenous, and via inhalation [62]. In managing pseudomonal infection among CF patients, a common strategy involves employing a combination of at least two antibiotics. This typically includes administering intravenous beta-lactams, intravenous or oral fluoroquinolones, and intravenous aminoglycosides (along with nebulized tobramycin) [61]. Tobramycin belongs to aminoglycoside class of antibiotics which is used as an accepted standard for CF patients. The aerosol nebulized formulation of tobramycin, approved for use in 1998, is widely accessible [63]. It is administered as 300 mg, twice daily in alternate 28 days on/off cycles to improvise pulmonary health. Tobramycin when employed in combination with fosfomycin (anti-pseudomonal antibiotic), synergistic effects are ascertained. This combination utilizes a lesser amount of tobramycin, hence reducing its long term toxic effects on the patients [64]. Ciprofloxacin, a second generation fluoroquinolone can be used as a successful line of treatment during the off cycle of tobramycin. Introduced in 1985, it is administered as oral and intravenous preparation. Despite having adverse drug reactions like sunlight sensitivity rash and cytotoxicity, ciprofloxacin is regarded as a safe and an effective alternative to already existing traditional approaches that are used to treat CF [65]. Among cephalosporins, intravenous administration of cefepime and ceftazidime has also demonstrated anti-infective properties against P. aeruginosa [66]. Further, beta-lactams utilized to combat pseudomonal infections in CF patients consist of penicillins (ticarcillin-clavulanate and piperacillin-tazobactam), monobactums (aztreonam), cephalosporins (cefepime and ceftazidime), and carbapenems (meropenem, imipenem-cilastatin, doripenem) [66]. The above-mentioned penicillins have broad-range antibacterial activity against diverse bacteria, including P. aeruginosa, and are given intravenously during acute pulmonary exacerbations [67]. Aztreonam available as both aerosolized and intravenous forms, has numerous advantages including effective sputum penetration, a prolonged time to the next exacerbation, and enhanced lung function [68]. Also, intravenous administration of carbapenems has been extensively deployed against a plethora of acute pulmonary pathogens.
Besides conventional antibiotic pipeline, alternative therapeutics include vaccination and CFTR modulators. Nowadays, anti-pseudomonal vaccines (bivalent P. aeruginosa flagella vaccine) delivered via nasal or oral route are being considered as viable options for eliciting a humoral response in the pulmonary tissues of CF patients [69]. Interestingly, effector molecules (CFTR modulators) that enhance the channel activity of CFTR proteins (potentiators) and increase the availability of functional CFTR protein at the apical membrane (correctors), have also been documented in the literature. In this regard, modulators are known to function specifically against a particular class of CFTR mutation [70]. The initial success was achieved with ivacaftor, a CFTR potentiator medication, which proved to be effective for individuals harboring class 3 or ’gating’ mutations. Ivacaftor has recently been approved for use in children upto 2 years old and is further undergoing trials in even neonates with CF. A combination of ivacaftor and lumacaftor (corrector + potentiator) showcased slight improvement in lung function as compared to administration of ivacaftor alone in patients harbouring class 3 mutations [71].
4. Limitations of antimicrobial approach: the fault in our stars
The intrinsic drug resistance in P. aeruginosa has widely been correlated with the expression of different efflux pumps and porins in the bacterial cell membrane. These efflux pumps can expel a multitude of antibiotics such as fluoroquinolones, aminoglycosides, and β-lactams, but not polymyxins [72]. Further, conjugation across bacterial types is a potent mechanism that contributes to acquired drug resistance through passage of transferable enzymes that modify or hydrolyze antibiotics. Contrastingly, de novo mutational events seldom occur as a consequence of β-lactamase and efflux pump overexpression, ultimately leading to acquired resistance [73].
In the realm of CF strains, there is a notable prevalence of acquired resistance via adaptive mutations. This occurs because of the prolonged selection pressure arising from prolonged antibiotic therapies against persistent infections, as well as interactions with the host immune system [74]. Adaptive resistance, induced by external stimuli (stress factors and specific antibiotics), is distinct from acquired resistance mechanisms. Unlike mutations, it is unstable and transient, as it gets inactivated upon eliminating the stress factor [75]. Adaptive resistance mechanisms often involve regulatory or metabolic pathways, resulting in differential expression of bacterial genes, altered protein synthesis, or changes in the target entity. Biofilm formation, MexXY induction, and the two-component signaling systems (TCSS) are a few illustrative examples of adaptive resistance in P. aeruginosa [76,77]
While the prolonged use of antibiotics has been crucial in improving the survival rates of CF patients, it introduces the complication of patients grappling with the negative consequences of antibiotic toxicity. Specifically, aminoglycosides are commonly linked to nephrotoxicity and ototoxicity, posing additional challenges for patients [78]. Additionally, antibiotic treatment can impact not only the lung microbiome but also microbial communities in other parts of the body [79]. Allergic reactions to antibiotics are also a significant concern, affecting up to 30% of CF patients [80].
Another challenge in dealing with antimicrobials include targeted drug delivery systems. Owing to low potency of current antimicrobials, there is need for the inhaled drugs to be highly soluble in water accompanied with administration of high concentration of doses up to 100 mg. Moreover, the extracellular matrix within the pseudomonal biofilm, primarily consisting of extrapolymeric substances (rhamnolipids, alginate), complex polysaccharides (Pel and Psl), and DNA, acts as a substantial obstacle and significantly limit the entry of antimicrobial drugs [81]. Specifically, the efficacy of aminoglycosides, notably tobramycin, has been observed to diminish due to robust interactions with the biofilm components. These interactions result in slower penetration through the biofilm matrix and is usually incomplete or partial [82].
5. Antivirulence strategies to combat P. aeruginosa in CF: the next-generation therapies?
Besides antimicrobial approaches, novel therapeutics which aim to disrupt or inhibit the production of virulence factors is emerging as an attractive alternative in treating Pseudomonas infection. These antivirulence therapeutics interrupt vital pathways for pathogenesis without impacting bacterial growth [83]. The forthcoming sections provide an insight into the prospective antivirulence strategies such as inhibition of quorum sensing mechanism and biofilm formation, iron uptake, lectin, and efflux pump. The different antivirulence strategies employed against P. aeruginosa have been enlisted in Table 2. Based on the existing literature on antivirulence approaches targeting CF isolates of P. aeruginosa, a schematic overview has also been illustrated in Figure 2.
5.1. Targeting QS mechanisms and biofilm inhibition: the communal approach
Quorum sensing (QS) is an intra-cellular signaling mechanism which enables microorganims to detect variability in cell density and accordingly adjust their gene expression patterns, as well as coordinate species behaviour within a microbial population. It regulates various processes, such as the activation of pathogen virulence factors (immune-evasion factors, proteases, toxins) and production of biofilm. The concentration of autoinducers (QS signal molecules) released by bacteria are a deciding factor for initiation of such responses [84].
P. aeruginosa harbors three quorum sensing (QS) pathways, namely Las, Rhl and Pqs [83]. These pathways rely on distinct autoinducers: N-(3-oxododecanoyl)-l-HSL (OdDHL) produced in case of Las system, it is synthesized by LasI and recognized by LasR receptor. N-butanoyl-L-homoserine lactone (C4-HSL) produced by RhlI synthase, and detected by RhlR [85]. 2-heptyl-3-hydroxy-4(1H)-quinolone (PQS) is formed in case of the pqs system and identified by PqsR receptor [86]. A cascade of cellular communication events is triggered above the limiting concentration of the signal, which in turn modulates expression of numerous genes governing variety of physiological processes related to bacterial pathogenicity [84].
Biofilm formation refers to specific clustered configurations of bacteria enclosed in extracellular polymeric substances (EPS). This arrangement serves as a protective mechanism, enabling pathogenic bacteria to escape adverse conditions like temperature variations, nutrient scarcity, and antibiotic exposure. The formation of biofilm emerges as the key feature of P. aeruginosa, with highly organized biofilms frequently identified in individuals suffering from persistent infections [87]. QS along with virulence factors serves as an important factor in strengthening and increasing the thickness of biofilms. Nevertheless, both QS and biofilm present themselves as potential targets for antivirulence therapies as their inhibition can substantially reduce pathogen virulence, thus impeding its pathogenecity. Numerous plant extracts have been found to curb QS signals and impede the formation of biofilm such as coumarin (a phenolic plant compound). It is effective in suppressing protease and pyocyanin production as well as subduing biofilm formation. Additionally, a trancriptomic study based on coumarin treated P. aeruginosa PAO1 biofilm, showed a possible downregulation in QS genes involved in persistence of the organism in the lungs [88]. Another organic compound baicalin, derived from Scutellaria baicalensis, hampers the formation of P. aeruginosa biofilm below the minimum inhibitory concentrations (sub-MICs) and promises enhanced in vitro efficacy of bactericidal compounds. Furthermore, it diminishes the expression levels of quorum-sensing regulatory genes (lasI, lasR, rhlI, rhlR, pqsR, and pqsA). Baicalin produces an enhanced Th1-induced immune response, facilitating the clearance of pathogen in C. elegans, thereby reducing the disease causing potential of P. aeruginosa [89]. A recent study on the essential oil extracted from Corydothymus capitatus, highlighted its potential as an innovative agent targeting virulence pathways exhibited by P. aeruginosa infection in clinical isolates from individuals with CF [90]. The Corydothymus capitatus essential oil (CCEO) significantly proved to reduce pyocyanin production (84% to 100%) in all tested P. aeruginosa strains. Further, it showed biofilm inhibition as well as restricted swimming and swarming motility of all the tested strains. Hence CCEO stands out as a prospective candidate to mitigate P. aeruginosa virulence [28]. In a similar direction, diminished expression of four QS genes (lasI, lasR, rhlI, rhlR) and ndvB gene (responsible for biofilm formation) in P. aeruginosa isolates was observed on treatment with a plant extract obtained from Dioon spinulosum. It also helped in effectively reducing extracellular polymeric substance (EPS), altered cell surface hydrophobicity, and biofilm formation [91]. Besides natural products, many synthetic compounds have proved to be inhibitors of QS system and biofilm formation. A compound with glycine ester branch tested in P. aeruginosa strain MH 602, blocked the activity of LasR. Also, it consequently lowered pyocyanin production [92]. LasI represents an intriguing drug target, and various studies have pinpointed molecules capable of inhibiting its activity. Trans-cinnamaldehyde has been known as a potent inhibitor of AHL synthases, leading to reduction in pyocyanin production in P. aeruginosa PAO1. The binding of trans-cinnamaldehyde to the substrate binding pocket of LasI has been confirmed by molecular docking analysis [93]. A research study analyzed a range of synthetic molecules and revealed their ability to inhibit the QS receptor LasR and RhlR. The most potent molecule was identified to be an analog of a natural autoinducer known as meta-bromothiolactone (mBTL). This analog was highly effective in impeding the production of biofilm and virulence factors (mainly pyocyanin). Furthermore, it demonstrated protective effects against P. aeruginosa infections in both human respiratory epithelial cells and C. elegans [94].
PqsR is commonly targeted by QS Inhibitors (QSIs). Among the inhibitors targeting Pqs, one notable example is a derivative of HHQ. Once inside the cell, synthase PqsH is responsible for the conversion of the derivative into a powerful PqsR agonist. Further optimization was achieved by introducing a CONH2 group that helped in increasing the chances of survival in P. aeruginosa infections [95]. The suppression of Pqs- related virulence traits in P. aeruginosa was attained using inhibitors such as M34 and Clofactol M34, identified through high-throughput screening, protected mice from pseudomonal infection by preventing the binding of PQS-PqsR [96]. Clofoctol, similar in action to M34, is also an inhibitor of the Pqs system and increases the life span of G. mellonella larvae infected by P. aeruginosa [97]. The synthetic derivative, Furanone C-30 is an inhibitor of LasR which disrupts the Las pathway. This interference not only leads to increment in biofilm susceptibility to tobramycin, but also causes a repression in levels of virulence factors of P. aeruginosa [98]. Additionally, two synthetic compounds, MHY1383 and MHY1387, harbour the ability to impede QS-regulated virulence in P. aeruginosa. These compounds bring about a substantial reduction in biofilm production by declining the intracellular c-diGMP levels and act as competitive inhibitors, preventing the binding of OdHSL to LasR [99]. Recently, some newly devised molecules such as furothiazole derivative 5, derivative 4, amino benzene sulfonamide, pyridine derivative 3 and derivative 2 have shown anti-fouling property against P. aeruginosa. Notably, derivative 3 demonstrates comparable anti-fouling efficacy against the P. aeruginosa ATCC 27853 to drugs like ciprofloxacin and ampicillin [100]. Moreover, nitric oxide (NO) serves as a crucial regulator in P. aeruginosa bacterial dispersal and biofilm production owing to its potential in reducing the expression levels of c-di-GMP [101]. Recently, it has been discovered that benzoxazolone derivatives functioning as QS inhibitors, structurally mimic the AHL autoinducers. Specifically, a benzoxazolone derivative (B20) [102] illustrated pyocyanin inhibition in PAO1 without impacting bacterial growth. Further, exposure to B20 led to a dose-dependent decrease in QS genes levels (pqsA, rhlA, and lasB), suggesting its role in reduced pyocyanin levels [102]. Furthermore, Blanco-Cabra et al. carried out investigations and proposed that P. aeruginosa possess the ability to neutralize antimicrobial threats by sequestering antibiotics within the biofilm. In this regard, DNaseI-linked dextran-based single-chain polymer nanoparticles (SCPNs) (coupled with tobramycin) counterbalanced the ionic interactions consequently dispersing the biofilm matrix [103].
5.2. Targeting iron acquisition pathways: provoking nutrient deficiency
Bacterial persistence, survival, and biofilm formation in P. aeruginosa are quintessentially dependent on iron, which may be acquired from haem or non-haem iron sources. Two iron uptake systems are present in P. aeruginosa namely Has and Phu. The Has system releases a hemoglobin-binding protein (HasAp) that is taken back up through the Has receptor (HasR) once it binds to hemoglobin [104]. Contrastingly, Phu system functions by directly attaching hemoglobin or hemoglobin-containing proteins to a receptor embedded in the membrane. Although the genome of P. aeruginosa includes a third gene (hxuC) encoding a hemoglobin receptor, its regulatory mechanisms have not been characterized to date [105].
P. aeruginosa encounters limited heme as an iron source in its natural environment and, under aerobic conditions. Consequently, it must efficiently scavenge non-haem iron, predominantly in the less soluble ferric (Fe3+) state. To address this need, Pseudomonas synthesizes and releases high-affinity iron-chelating siderophores, specifically pyoverdine and pyochelin, to capture iron from host iron-binding proteins [106]. Pyoverdine, the predominant siderophore in P. aeruginosa, exhibits unique greenish-yellow fluorescence, with over 50 characterized subtypes. Each strain typically produces one of three sub-classes of pyoverdine [107]. In comparison to pyoverdine, pyochelin, has a lower iron sequestering ability [105,106].
The iron acquisition mechanisms of P. aeruginosa are strictly regulated by the ferric uptake regulator (Fur). Fur exercise control both directly and indirectly, involving extracytoplasmic sigma factors such as PvdS, to restrict iron absorption [108]. In iron rich environments, Fur protein binds to ferrous iron and adheres to a consensus sequence (Fur-box) located in the promoter region of genes essential for iron acquisition. As a consequence, transcription of these genes is suppressed [109]. Conversely, in environments with scarce iron, there is an upregulation of siderophore synthesis, and non-essential processes that consume iron are downregulated.
Currently, there is an increasing emphasis on development of newer therapeutic strategies to target the iron acquisition systems vital for P. aeruginosa to establish successful infections. One such approach is iron mimetics. Gallium nitrate [Ga (NO3)3] has received FDA approval for various medical applications such as intravenous administration for cancer-associated hypercalcemia.Ga3+ is emerging as an anti-pseudomonal agent because it shares a nearly identical ionic radius with Fe3+, making it practically indistinguishable for P. aeruginosa [110] This similarity allows Ga3+ to act as a "Trojan horse," disrupting iron metabolism. Unlike Fe3+, P. aeruginosa cannot undergo the necessary redox reactions with Ga3+. A research led by Kaneko et al. (Kaneko et al., 2007) revealed suppression of PvdS (transcriptional regulator of pyoverdine synthesis) by Ga(NO3)3 lead to a reduction in iron uptake in P. aeruginosa PAO1 [110]. Nevertheless, biological iron chelators can also be used to inhibit P. aeruginosa infections. For instance, Lactoferrin, an antimicrobial glycoprotein chelates iron in turn, increasing bacterial motility and diminishing biofilm production at sub-MIC [111].
Further, biofilm-dwelling bacteria, like Pseudomonas depends on iron, and scientists are exploring ways to "hijack" this iron transport system. They have developed siderophore–antibiotic conjugates (SACs), which could act like "Trojan horses" to get past the biofilm’s defenses and deliver antibiotics effectively [112,113]. In vitro investigations have elucidated that the cooperative administration of desferrioxamine (iron chelator) and tobramycin (antibiotic) significantly hinders the formation of biofilms and also decreases the biomass of preformed biofilms on CF epithelial cells [114].
5.3. Lectin inhibitors: outmaneuvering the battle for cell adhesion
Lectins are external membrane proteins required by certain microorgansms to establish biofilms [115]. They also form a barrier against host mediated responses by inhibiting the action of mucociliary clearance of the pathogen in the airway [116] Significance of these extracellular proteins lies in initiating interactions by attaching to carbohydrate epitopes present on bacterial exopolysaccharides, along with binding to both bacterial and host glycocalyces. LecA and LecB refer to two extracellular lectins expressed by P. aeruginosa. These lectins play pivotal roles in the biofilm formation process. These have specific binding sites for fucose and galactose respectively [55]. Consequently, these binding sites make them susceptible to inhibition by competitive agents. In vitro studies have shown that these inhibitors, either independently or in conjunction with antibiotics, effectively facilitate the disintegration and disruption of biofilms [117,118].
A small randomized trial involving patients with CF who underwent galactose/fucose inhalation treatment showcased a positive trend towards improvement in disease [119]. This trial specifically recruited subjects with persistent pseudomonal infection amidst infection flare-up. The participants randomly received aerosol sugar inhalation treatment or in combination with antibiotics, administered intravenously. Both groups exhibited a considerable reduction in levels of tumour necrosis factor alpha and colony-forming units of P. aeruginosa in sputum. More recently, higher affinities for multivalent dendrimers carrying these sugars as compared to monovalent fucose was observed, highlighting their use as substantial therapeutic agents [120]. In a separate investigation, glycocluster comprising tetravalent thio- and selenogalactopyranosides were synthesized using the azide-alkyne click strategy. The resulting compounds had the potential to act as ligands for the P. aeruginosa lectin LecA [121]. The hemagglutination inhibition assay was employed to explore the interactions between LecA and tetravalent glycoconjugates and comparison was made with digalactosyl diselenide, and digalactosyl disulfide (mono- and divalent galactosides) and propargyl 1-thio- and 1-seleno-β-d-galactopyranoside. Additionally, carbohydrate-lectin interactions were examined using the NMR technique. Both thio- and seleno-tetravalent glycoconjugates demonstrated a significantly higher inhibition (approximately 64 times higher) of LecA compared to simple D-galactose. Given the utilization of L-fucose and D-galactose in managing CF during a clinical trial [119], the newly developed S- and Se-galactoconjugates could emerge as potential candidates for anti-adhesion therapy in CF. These compounds could potentially be perceived as inhalational drugs for the management of CF. Hence, inhibiting the activities of both LecA and LecB is an emerging technique to mitigate the pathogenesis of P. aeruginosa and disrupt its antimicrobial resistance in CF patients.
5.4. Efflux pump inhibitors: eroding the pathways for drug extrusion
Efflux pumps are transmembrane proteins, actively involved in transporting a variety of substances such as toxins and antimicrobials between bacterial cells and the extracellular environment. Notable, resistance-nodulation-cell-division (RND) family of efflux pump systems in P. aeruginosa include MexXY-OprM, MexEF-OprN MexCD-OprJ and MexAB-OprM [122]. These pumps confer intrinsic antibiotic resistance in P. aeruginosa. These systems consist of various proteins that form complexes facilitating the recognition and extrusion of different molecules [122]. In addition, QS also relies on efflux mechanisms, as the OdDHL requires efflux pumps for its active transportation as it is not capable of independent diffusion through cell wall, thereby affecting pathogen virulence [28]. The effects can lead to both positive and negative outcomes. While heightened activity of the efflux pumps might lead to enhanced transport of QS molecules, it could also result in a substantial amount of molecules exiting the cell, thereby reducing the quorum signal. Consequently, this reduction in quorum signal may further contribute to decreased virulence [123]. Furthermore, efflux pumps contribute either directly or indirectly in imparting resistance against antimicrobial agents, therefore, they become a compelling and promising target for antivirulence therapy.
Efflux pump inhibitors (EPIs) are compounds having the capacity to interfere with the activity of efflux pumps. Researchers are investigating these compounds as promising candidates for restoring the effectiveness of antibiotics that have encountered resistance in bacterial strains. A plethora of studies have provided evidence that the concurrent use of EPIs can successfully address inherent antibiotic resistance, reverse acquired antimicrobial resistance, and impede the development of MDR strains when given in combination with existing antibiotics [28]. For instance, a study demonstrated that RND efflux pumps of P. aeruginosa PAO1 in an insect model of infection are responsible for conferring resistance to already existing antimicrobials. Phe-Arg-β-naphthylamide (PAβN), showed inhibitory activity against RND efflux pumps such as MexEF-OprN and MexCD-OprJ. Additionally, it reduced the virulence of both P. aeruginosa PAO1 and clinical isolates in vivo [124]. After comprehensive phenotypic and transcriptomic analyses, it was revealed that by suppression of crucial virulence genes (regulating iron and phosphate starvation), PAβN successfully defended against P. aeruginosa PAO1 infection in animal models. PAβN when used above 50-200 µM concentrations can drastically lead to a decrement in bacterial growth [125] In another study, the impact of 27µM PAβN on clinical strains of P. aeruginosa strains (eleven) was investigated on the production of virulence-related phenotypes such as swarming motility, pyocyanin and OdDHL. Intriguingly, PAβN showed varying effects amidst the seven strains that produced OdDHL. In one of the strains the trait was increased, while in the other two no effects were observed and further the production of this signal molecule was inhibited to varying degree in the remaining strains. Then, out of four CF isolates producing pyocyanin, two showed reduced production in response to PAβN, while the other two exhibited increased production. In the absence of PAβN, only five CF isolates showcased swarming motility, whereas this attribute was eliminated in all of them upon PAβN treatment [124]. Similarly, a research study evaluated the effectiveness of free as well as liposomal forms of erythromycin and gentamicin coupled with PAβN against various pseudomonal strains [126]. Liposomes showed an average diameter of 3086.35 nm for erythromycin and 562.67 nm for gentamicin with encapsulation efficiencies of 51.58% and 13.89% respectively. Furthermore, MIC and MBC were reduced by 4–32 fold overall upon combining antibiotics with PAβN, thereby, enhancing their potency. This amalgamation markedly suppressed biofilm formation and variedly reduced the production of virulence factors. Surprisingly, drug treatments did not impact the production of QS signals. Overall, the findings suggest that PAβN acts synergistically along with liposomal aminoglycosides and macrolides in tackling pulmonary infections in CF patients [126]
To conquer efflux-mediated multidrug resistance, one approach involves the development of indirect antimicrobial agents. These agents restore the activity of antimicrobials that had been previously neutralized by efflux mechanisms. In this regard, TXA09155 (conformationally constrained indole carboxamide) is known to be a potent EPI that diminishes intrinsic resistance levels and markedly reverses acquired resistance besides reducing the frequency of emerging anthropogenic resistance. The efficacy of TXA09155 in conjunction with levofloxacin was examined to assess its impact on the frequency of resistance (FoR) of P. aeruginosa ATCC 27853. The results showed that TXA09155 significantly decreased the FoR to levofloxacin by over 149-fold [127]. Negligible resistance levels observed in this combination could be immensely beneficial in a clinical context, especially for CF patients harboring P. aeruginosa. Other EPI’s including, EA-371δ and EA-371α were screened from microbial fermentation extracts archives comprising 78,000 samples [128]. These are natural products that exhibit inhibitory effects on MDR efflux pumps, particularly the MexAB-OprM system. These compounds, derived from a Streptomyces strain, demonstrate significant potency as MexAB-OprM inhibitors, with 2.15 μM (EA-371δ) and MPC8 values of 4.29 μM (EA-371α) for levofloxacin against strain PAM 103.However, potential of EA-371α’s as a lead compound is hindered by its moderate cytotoxicity, highlighting the need for further exploration in the search for effective and safe MDR efflux pump inhibitors [128]. Another distinct efflux pump inhibitor, D13-9001(pyridopyrimidine derivative) has been discovered. This compound effectively hinders MexAB-OprM both in vivo and in vitro, showcasing promising antivirulence properties. Notably, D13-9001 demonstrates a favorable low toxicity profile [129]. It acts by interacting with the hydrophobic trap of the pump and interfering with structural changes important for the pump to function. Simultaneously, D13-9001 acts to obstruct substrate binding to MexB, contributing to its inhibitory efficacy. This compound thus represents a valuable addition to the repertoire of efflux pump inhibitors, offering a dual mechanism for potential therapeutic applications [130].
Table 2.
Presently-reported anti-virulence strategies targeting CF isolates of P. aeruginosa.
| Anti-virulence approaches | Therapeutic used | Effect/outcome on disease progression | References |
|---|---|---|---|
| Quorum sensing and biofilm inhibition | Coumarin | It is described as a potent QSI. Significant reduction in pyocyanin and protease levels as well as biofilm formation were observed in vitro. Decline in intracellular C-di-GMP levels suggested inhibition of biofilm. However, its biofilm-inhibitory effects in wound healing model and Lucilia sericata maggots were reduced. Hence, the therapeutic potential of coumarin in treating wound infections and helping with maggot-based debridement therapy may be limited | [88] |
| Corydothymus capitatus essential oil (CCEO) | CCEO demonstrated a significant decrease in pyocyanin production, ranging from 84% to 100%, across all tested strains. In nearly half of the strains, inhibition and reduction of mature biofilm was reported. It also displayed a substantial impact on the swarming and swimming motility of P. aeruginosa for nearly all strains examined | [28] |
|
| Baicalin | Baicalin exhibited potent anti-QS and anti-biofilm properties. The study showcased its dose-dependent inhibitory effect on virulence phenotypes (LasA protease, LasB elastase, pyocyanin, rhamnolipid, motilities and exotoxin A) which are regulated by QS. Decrease in signaling molecule 3-oxo-C12-HSL and C4-HSL was also observed due to repressed QS regulatory gene expression level. In vivo treatment with baicalin in C. elegans and mouse peritoneal implant infection model resulted in lowered P. aeruginosa pathogenicity | [89] | |
| Dyer Ex Eichler extract (DSE) | Remarkable reduction in P. aeruginosa biofilm formation was observed both in vitro as well as in vivo rat model. Further it subdued surface hydrophobicity and extracellular polymeric substance (EPS) accumulation. qRT-PCR results revealed reduction in four QS genes (lasI, lasR, rhlI, rhlR) and biofilm related gene ndvB | [91] | |
| M34 and Clofoctol | M34 and its derivatives potentially affects the transcription factor PqsR, impeding the binding between PQS and pqsR, disrupting the production of pqs-dependent signaling molecules, and offering protection against P. aeruginosa infection in mice Clofoctol, FDA-approved compound; acts by inhibiting the pqs system and decreasing the mortality rate of Galleria mellonella larvae infected with P. aeruginosa by targeting PqsR. PqsR inhibition leads to decreased production of 2-alkyl-4-quinolones (AQs) and hence of AQs-dependent virulence factors, with consequent attenuation of P. aeruginosa infectivity |
[28,97] | |
| Furanone C-30 | When used in combination with tobramycin, Furanone C-30 effectively reduced protease production. It was also used in combination with gallium (a siderophore) which resulted in reduction of virulence factor and QS genes. Additionally it disrupted Las pathway (especially LasR) and increased the susceptibility of P.aeruginosa biofilms towards tobramycin | [98] | |
| Niclosamide | Niclosamide acts on the 3OC12-HSL signaling pathway through a mechanism that remains to be fully characterized. It reduces the synthesis of 3OC12-HSL as well as QS-dependent virulence factors (pyocyanin and elastase), resulting in decreased virulence in the Galleria mellonella infection model. The efficacy of niclosamide against CF isolates in inhibiting QS and virulence was highly variable and strain-dependent. It demonstrated low range of effectiveness and its inhibition of las signal production did not correspond to a decrease in the production of virulence factors | [131] |
|
| Luteolin | Biofilm formation, production of virulence factors, and motility (swimming, swarming, twitching) of P. aeruginosa was effectively inhibited by luteolin. It attenuated the accumulation of the QS-signaling molecules N-(3-oxododecanoyl)-L-homoserine lactone (OdDHL) and N-butanoyl-L-homoserine lactone (BHL). In addition, it downregulated the QS genes (lasR, lasI, rhlR, and rhlI) | [132] | |
| Inhibiting iron uptake |
Gallium nitrate (Ga(NO3)3) |
Gallium suppressed the growth and formation of biofilms in P. aeruginosa and eliminated both planktonic and biofilm bacteria in vitro. Its mechanism involves reducing bacterial iron uptake and disrupting iron signaling mediated by the transcriptional regulator pvdS required for pyoverdine synthesis. Furthermore, the study demonstrated the efficacy of Gallium in murine lung infection models | [110] |
| Koelreuteria paniculata leaf extract silver nanoparticles | The research demonstrated that the Ka-AgNPs significantly mitigates QS-regulated virulence factors in PAO1 and effectively suppress the formation of biofilm of PAO1. Further, the expression of QS-regulated virulence genes was reduced as well. These results suggest that the phyto-synthesized AgNPs could be used as promising anti-infective agents for treating drug-resistant P. aeruginosa | [133] |
|
| Lactoferrin | Inhibitory/destructive effects of lactoferrin (2 mg/ml) on biofilm formation as well as pre-formed biofilm were observed. Further, pre-treatment with FeCl2 partially restored biofilm formation, suggesting its role as an iron-chelator that may be implicated to the inhibitory mechanism of lactoferrin | [111] |
|
| N,N’-bis (2-hydroxybenzyl) ethylenediamine-N,N’-diacetic acid (HBED) | HBED showed inhibitory effects on growth and biofilm formation in all clinical strains of P. aeruginosa isolated from CF patients, under aerobic or anaerobic conditions. The addition of HBED significantly decreased the biomass and when combined with colistin, HBED notably augmented the microcolony-killing effects of colistin, leading to nearly complete eradication of the biofilm. Thus, the combination of HBED and colistin demonstrates high efficacy in vitro against biofilms formed by clinical strains of P. aeruginosa | [134] | |
| Lectin inhibitors |
Fucose/galactose inhalation | Lectin specific sugars, fucose and Galactose prevented binding of P. aeruginosa lectins I and II responsible for adherence of bacteria in the airway cells. The competitive inhibition of P. aeruginosa lectins by the sugars may overcome particular mechanisms of bacterial resistance in patients with airway infection. Reductions in tumor necrosis factor alpha and colony growth of P. aeruginosa were also reported | [119] |
| Thio- and Seleno-tetravalent glycoconjugates | Both thio- and seleno-tetravalent glycoconjugates act as ligands for lectin PA-IL (one of the major virulence factor in P. aeruginosa associated with CF) consequently inhibiting binding of LecA. It also demonstrated approximately 64 times higher supression of LecA compared to d-galactose | [121] | |
| Efflux pump inhibitors (EPI) | Phe-Arg-β-naphthylamide (PAβN) | PAβN functions as an anti-QS and anti-biofilm agent against P. aeruginosa CF isolates. It has been shown to permeabilize the bacterial cell membrane in a dose-dependent manner, thereby increasing the efficacy of bulky β-lactam antibiotics (incapable of crossing the membranes). It also repressed the activity of efflux pumps like MexCD-OprJ and MexEF-OprN and negatively impacted bacterial growth and virulence factors in insect model of infection | [124,125] |
| TXA09155 (Conformationally Constrained Indole carboxamide + levofloxacin | TXA09155 served as a potential EPI in P. aeruginosa. When used at concentration of 6.25 µg/mL it increased potency of antibiotics by 8 times. TXA09155 is known to improvise killing dynamics of moxifloxacin and decrease frequency of resistance (FoR) to levofloxacin. Combination of TXA09155 and levofloxacin was used effectively in treatment of burn wound victims, as it significantly reduced emergence of MDR strains of P. aeruginosa against carbapenams | [127] |
6. Conclusions
To sum up, P. aeruginosa airway infection presents considerable challenges in CF, with current diagnostic and treatment approaches inadequately preventing the progression to chronic infection in the majority of patients by adulthood. The relentless nature of these infections, coupled with the rise of antimicrobial resistance, underscores the pressing need for innovative strategies. In this comprehensive review, emphasis has been made on the critical significance of P. aeruginosa infection in patients diagnosed with CF. Various mechanisms facilitating the pathogen’s persistence within the host are thoroughly examined. The review looks into existing techniques and antimicrobial strategies aimed at addressing P. aeruginosa infections. Additionally, novel antivirulence approaches, including the inhibition of quorum sensing (QS), biofilm formation, iron uptake, LecA and LecB, and efflux pumps, are discussed in detail. Ongoing researchers aim to enhance early detection methods, potentially enabling real-time identification of pseudomonal infections in the future. Conventional antibiotic treatments are only partially effective and contribute to resistance issues. Hence, innovative antibiotic formulations and combinations are currently undergoing clinical trials. This review places particular emphasis on diverse antivirulence therapeutics, offering a fresh perspective to confront the prevailing resistance crisis and improve the overall well-being of individuals diagnosed with CF.
Author Contributions
Umang Mudgil: Investigation, Literature survey, Writing-original draft. Lavanya Khullar: Investigation, Validation, Data curation, Writing- review, and editing. Jatin Chadha: Idea conceptualization, Investigation, Data curation, Supervision, Writing- original draft, review, and editing. Prerna: Literature survey, Formal analysis. Kusum Harjai: Idea conceptualization, Supervision, Validation, Continuous motivation, Writing-review and editing.
Data Availability Statement
All the datasets generated and analyzed during the current study have all been cited in this manuscript.
Acknowledgements
Lavanya Khullar and Jatin Chadha would like to thank the Indian Council of Medical Research (ICMR), New Delhi, for providing Senior Research Fellowship (SRF).
Conflict of Interest
The authors declare that there is no conflict of interests.
Abbreviations
AHL: Acyl homoserine lactone, AMR: antimicrobial resistance, CF: cystic fibrosis, CFTR: cystic fibrosis transmembrane regulator, LPS: Lipopolysaccharide, OdDHL: N-(3-oxododecanoyl)-l-HSL, PAβN: Phe-Arg-β-naphthylamide, QS: Quorum sensing, QSI: Quorum sensing inhibitor.
References
- Davis, P.B. Cystic Fibrosis Since 1938. American Journal of Respiratory and Critical Care Medicine 2006, 173, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Mehta, G.; Macek, M.; Mehta, A. Cystic fibrosis across Europe: EuroCareCF analysis of demographic data from 35 countries. Journal of Cystic Fibrosis 2010, 9, S5–S21. [Google Scholar] [CrossRef]
- Boucher, R.C. An overview of the pathogenesis of cystic fibrosis lung disease. Advanced Drug Delivery Reviews 2002, 54, 1359–1371. [Google Scholar] [CrossRef] [PubMed]
- Shah, V.S.; Meyerholz, D.K.; Tang, X.X.; Reznikov, L.; Abou Alaiwa, M.; Ernst, S.E.; Karp, P.H.; Wohlford-Lenane, C.L.; Heilmann, K.P.; Leidinger, M.R.; et al. Airway acidification initiates host defense abnormalities in cystic fibrosis mice. Science 2016, 351, 503–507. [Google Scholar] [CrossRef]
- Döring, G.; Gulbins, E. Cystic fibrosis and innate immunity: how chloride channel mutations provoke lung disease. Cellular Microbiology 2009, 11, 208–216. [Google Scholar] [CrossRef]
- Ciro, D.O.; Padoan, R.; Blau, H.; Marostica, A.; Fuoti, M.; Volpi, S.; Pilotta, A.; Meyerovitch, J.; Sher, D.; Assael, B.M. Growth retardation and reduced growth hormone secretion in cystic fibrosis. Clinical observations from three CF centers. Journal of Cystic Fibrosis 2013, 12, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Maisonneuve, P.; Marshall, B.C.; Knapp, E.A.; Lowenfels, A.B. Cancer Risk in Cystic Fibrosis: A 20-Year Nationwide Study From the United States. JNCI: Journal of the National Cancer Institute 2013, 105, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Derichs, N. Targeting a genetic defect: cystic fibrosis transmembrane conductance regulator modulators in cystic fibrosis. European Respiratory Review 2013, 22, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Veit, G.; Avramescu, R.G.; Chiang, A.N.; Houck, S.A.; Cai, Z.; Peters, K.W.; Hong, J.S.; Pollard, H.B.; Guggino, W.B.; Balch, W.E.; et al. From CFTR biology toward combinatorial pharmacotherapy: expanded classification of cystic fibrosis mutations. Molecular Biology of the Cell 2016, 27, 424–433. [Google Scholar] [CrossRef]
- Azimi, A. “Cystic fibrotics could survive cholera, choleraics could survive cystic fibrosis”; hypothesis that explores new horizons in treatment of cystic fibrosis. Medical Hypotheses 2015, 85, 715–717. [Google Scholar] [CrossRef]
- Xu, J.; Yong, M.I.N.; Li, J.I.A.; Dong, X.; Yu, T.; Fu, X.; Hu, L. High level of CFTR expression is associated with tumor aggression and knockdown of CFTR suppresses proliferation of ovarian cancer in vitro and in vivo. Oncology Reports 2015, 33, 2227–2234. [Google Scholar] [CrossRef]
- Khan, T.Z.; Wagener, J.S.; Bost, T.; Martinez, J.; Accurso, F.J.; Riches, D.W.H. Early Pulmonary Inflammation in Infants with Cystic Fibrosis. American Journal of Respiratory and Critical Care Medicine 1995, 151, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Tang, A.C.; Turvey, S.E.; Alves, M.P.; Regamey, N.; Tummler, B.; Hartl, D. Current concepts: host-pathogen interactions in cystic fibrosis airways disease. European Respiratory Review 2014, 23, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, S.; Hayes, D.; Wozniak, D.J. Cystic Fibrosis and Pseudomonas aeruginosa: the Host-Microbe Interface. Clinical Microbiology Reviews 2019, 32. [Google Scholar] [CrossRef] [PubMed]
- Hector, A.; Kirn, T.; Ralhan, A.; Graepler-Mainka, U.; Berenbrinker, S.; Riethmueller, J.; Hogardt, M.; Wagner, M.; Pfleger, A.; Autenrieth, I.; et al. Microbial colonization and lung function in adolescents with cystic fibrosis. Journal of Cystic Fibrosis 2016, 15, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.J.; LiPuma, J.J. The Microbiome in Cystic Fibrosis. Clinics in Chest Medicine 2016, 37, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Smyth, A.R.; Smyth, R.L.; Tong, C.Y.; Hart, C.A.; Heaf, D.P. Effect of respiratory virus infections including rhinovirus on clinical status in cystic fibrosis. Archives of Disease in Childhood 1995, 73, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Collinson, J.; Nicholson, K.G.; Cancio, E.; Ashman, J.; Ireland, D.C.; Hammersley, V.; Kent, J.; O’Callaghan, C. Effects of upper respiratory tract infections in patients with cystic fibrosis. Thorax 1996, 51, 1115–1122. [Google Scholar] [CrossRef]
- Blanchard, A.C.; Waters, V.J. Opportunistic Pathogens in Cystic Fibrosis: Epidemiology and Pathogenesis of Lung Infection. Journal of the Pediatric Infectious Diseases Society 2022, 11, S3–S12. [Google Scholar] [CrossRef]
- Kidd, T.J.; Ramsay, K.A.; Vidmar, S.; Carlin, J.B.; Bell, S.C.; Wainwright, C.E.; Grimwood, K.; Francis, P.W.; Dakin, C.; Cheney, J.; et al. Pseudomonas aeruginosa genotypes acquired by children with cystic fibrosis by age 5-years. Journal of Cystic Fibrosis 2015, 14, 361–369. [Google Scholar] [CrossRef]
- Chadha, J.; Harjai, K.; Chhibber, S. Revisiting the virulence hallmarks of Pseudomonas aeruginosa: a chronicle through the perspective of quorum sensing. Environmental Microbiology 2021, 24, 2630–2656. [Google Scholar] [CrossRef] [PubMed]
- Losito, A.R.; Raffaelli, F.; Del Giacomo, P.; Tumbarello, M. New Drugs for the Treatment of Pseudomonas aeruginosa Infections with Limited Treatment Options: A Narrative Review. Antibiotics 2022, 11, 579. [Google Scholar] [CrossRef] [PubMed]
- AbdulWahab, A.; Zahraldin, K.; Sid Ahmed, M.; Jarir, S.; Muneer, M.; Mohamed, S.; Hamid, J.; Hassan, A.I.; Ibrahim, E. The emergence of multidrug-resistant Pseudomonas aeruginosa in cystic fibrosis patients on inhaled antibiotics. Lung India 2017, 34, 527. [Google Scholar] [CrossRef] [PubMed]
- Farrell, P.M.; Shen, G.; Splaingard, M.; Colby, C.E.; Laxova, A.; Kosorok, M.R.; Rock, M.J.; Mischler, E.H. Acquisition ofPseudomonas aeruginosain Children With Cystic Fibrosis. Pediatrics 1997, 100, e2. [Google Scholar] [CrossRef]
- Frederiksen, B.; Koch, C.; Hiby, N. Changing Epidemiology ofPseudomonas aeruginosa Infection in Danish Cystic Fibrosis Patients (1974-1995). Pediatric Pulmonology 1999, 28, 159–166. [Google Scholar] [CrossRef]
- Ranganathan, S.C.; Skoric, B.; Ramsay, K.A.; Carzino, R.; Gibson, A.-M.; Hart, E.; Harrison, J.; Bell, S.C.; Kidd, T.J. Geographical Differences in First Acquisition of Pseudomonas aeruginosa in Cystic Fibrosis. Annals of the American Thoracic Society 2013, 10, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Denton, M.; Kerr, K.; Mooney, L.; Keer, V.; Rajgopal, A.; Brownlee, K.; Arundel, P.; Conway, S. Transmission of colistin-resistant Pseudomonas aeruginosa between patients attending a pediatric cystic fibrosis center. Pediatric Pulmonology 2002, 34, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Starkey, M.; Lepine, F.; Maura, D.; Bandyopadhaya, A.; Lesic, B.; He, J.; Kitao, T.; Righi, V.; Milot, S.; Tzika, A.; Rahme, L. Identification of anti-virulence compounds that disrupt quorum-sensing regulated acute and persistent pathogenicity. PLoS Pathogens, 1004. [Google Scholar] [CrossRef]
- Wiehlmann, L.; Cramer, N.; Ulrich, J.; Hedtfeld, S.; Weißbrodt, H.; Tümmler, B. Effective prevention of Pseudomonas aeruginosa cross-infection at a cystic fibrosis centre – Results of a 10-year prospective study. International Journal of Medical Microbiology 2012, 302, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Conway, S. Segregation is good for patients with cystic fibrosis. Journal of the Royal Society of Medicine 2008, 101, 31–35. [Google Scholar] [CrossRef]
- Stapleton, P.J.; Izydorcyzk, C.; Clark, S.; Blanchard, A.; Wang, P.W.; Yau, Y.; Waters, V.; Guttman, D.S. Pseudomonas aeruginosa Strain-sharing in Early Infection Among Children With Cystic Fibrosis. Clinical Infectious Diseases 2021, 73, e2521–e2528. [Google Scholar] [CrossRef]
- Pier, G.B.; Grout, M.; Zaidi, T.S.; Olsen, J.C.; Johnson, L.G.; Yankaskas, J.R.; Goldberg, J.B. Role of Mutant CFTR in Hypersusceptibility of Cystic Fibrosis Patients to Lung Infections. Science 1996, 271, 64–67. [Google Scholar] [CrossRef] [PubMed]
- Smith, A. Pathogenesis of bacterial bronchitis in cystic fibrosis. The Pediatric Infectious Disease Journal 1997, 16, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Saiman, L.; Prince, A. Pseudomonas aeruginosa pili bind to asialoGM1 which is increased on the surface of cystic fibrosis epithelial cells. Journal of Clinical Investigation 1993, 92, 1875–1880. [Google Scholar] [CrossRef] [PubMed]
- Cohen, T.S.; Prince, A. Cystic fibrosis: a mucosal immunodeficiency syndrome. Nature Medicine 2012, 18, 509–519. [Google Scholar] [CrossRef]
- Sainz-Mejías, M.; Jurado-Martín, I.; McClean, S. Understanding Pseudomonas aeruginosa–Host Interactions: The Ongoing Quest for an Efficacious Vaccine. Cells 2020, 9, 2617. [Google Scholar] [CrossRef] [PubMed]
- Hiby, N. Prospects for the Prevention and Control of Pseudomonal Infection in Children with Cystic Fibrosis. Paediatric Drugs 2000, 2, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Van de Weert–van Leeuwen, P.B.; Van Meegen, M.A.; Speirs, J.J.; Pals, D.J.; Rooijakkers, S.H.M.; Van der Ent, C.K.; Terheggen-Lagro, S.W.J.; Arets, H.G.M.; Beekman, J.M. Optimal Complement-Mediated Phagocytosis of Pseudomonas aeruginosa by Monocytes Is Cystic Fibrosis Transmembrane Conductance Regulator–Dependent. American Journal of Respiratory Cell and Molecular Biology 2013, 49, 463–470. [Google Scholar] [CrossRef]
- Hill, P.J.; Scordo, J.M.; Arcos, J.; Kirkby, S.E.; Wewers, M.D.; Wozniak, D.J.; Torrelles, J.B. Modifications of Pseudomonas aeruginosa cell envelope in the cystic fibrosis airway alters interactions with immune cells. Scientific Reports 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Vencken, S.F.; Greene, C.M. Toll-Like Receptors in Cystic Fibrosis: Impact of Dysfunctional microRNA on Innate Immune Responses in the Cystic Fibrosis Lung. Journal of Innate Immunity 2016, 8, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.L.; Hufnagle, W.O.; Kowalik, D.J.; Lagrou, M.; et al. Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 2000, 406, 959–964. [Google Scholar] [CrossRef]
- Pedersen, S.S. Lung infection with alginate-producing, mucoid Pseudomonas aeruginosa in cystic fibrosis. APMIS Suppl 1992, 28, 1–79. [Google Scholar] [PubMed]
- Kharazmi, A. Interactions of Pseudomonas aeruginosa Proteases with the Cells of the Immune System. Antibiot. Chemother. 1989; 42, 42–49. [Google Scholar] [CrossRef]
- Lu, H.D.; Pearson, E.; Ristroph, K.D.; Duncan, G.A.; Ensign, L.M.; Suk, J.S.; Hanes, J.; Prud’homme, R.K. Pseudomonas aeruginosa pyocyanin production reduced by quorum-sensing inhibiting nanocarriers. International Journal of Pharmaceutics 2018, 544, 75–82. [Google Scholar] [CrossRef]
- Howell, P.L.; Weadge, J.T.; Nivens, D.E.; Franklin, M.J. Biosynthesis of the Pseudomonas aeruginosa Extracellular Polysaccharides, Alginate, Pel, and Psl. Frontiers in Microbiology 2011, 2. [Google Scholar] [CrossRef]
- Byrd, M.S.; Pang, B.; Mishra, M.; Swords, W.E.; Wozniak, D.J.; Pier, G. The Pseudomonas aeruginosa Exopolysaccharide Psl Facilitates Surface Adherence and NF-κB Activation in A549 Cells. mBio 2010, 1. [Google Scholar] [CrossRef] [PubMed]
- Friedman, L.; Kolter, R. Genes involved in matrix formation in Pseudomonas aeruginosa PA14 biofilms. Molecular Microbiology 2003, 51, 675–690. [Google Scholar] [CrossRef] [PubMed]
- Ausubel, F.M.; Colvin, K.M.; Gordon, V.D.; Murakami, K.; Borlee, B.R.; Wozniak, D.J.; Wong, G.C.L.; Parsek, M.R. The Pel Polysaccharide Can Serve a Structural and Protective Role in the Biofilm Matrix of Pseudomonas aeruginosa. PLoS Pathogens 2011, 7, e1001264. [Google Scholar] [CrossRef]
- Sawicki, G.S.; Rasouliyan, L.; McMullen, A.H.; Wagener, J.S.; McColley, S.A.; Pasta, D.J.; Quittner, A.L. Longitudinal assessment of health-related quality of life in an observational cohort of patients with cystic fibrosis. Pediatric Pulmonology 2010, 46, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Clemente, M.; de la Rosa, D.; Máiz, L.; Girón, R.; Blanco, M.; Olveira, C.; Canton, R.; Martinez-García, M.A. Impact of Pseudomonas aeruginosa Infection on Patients with Chronic Inflammatory Airway Diseases. Journal of Clinical Medicine 2020, 9, 3800. [Google Scholar] [CrossRef] [PubMed]
- Gallant, C.V.; Raivio, T.L.; Olson, J.C.; Woods, D.E.; Storey, D.G. Pseudomonas aeruginosa cystic fibrosis clinical isolates produce exotoxin A with altered ADP-ribosyltransferase activity and cytotoxicity The GenBank accession numbers for the toxA sequences are: strain 4384, AF227419; strain 5154, AF227420; strain 5166, AF227421; strain 5552, AF227422; strain 5585, AF227423; strain 5588, AF227424. Microbiology 2000, 146, 1891–1899. [Google Scholar] [CrossRef]
- Moser, C.; Jensen, P.Ø.; Thomsen, K.; Kolpen, M.; Rybtke, M.; Lauland, A.S.; Trøstrup, H.; Tolker-Nielsen, T. Immune Responses to Pseudomonas aeruginosa Biofilm Infections. Frontiers in Immunology 2021, 12. [Google Scholar] [CrossRef]
- Sandri, A.; Ortombina, A.; Boschi, F.; Cremonini, E.; Boaretti, M.; Sorio, C.; Melotti, P.; Bergamini, G.; Lleo, M. Inhibition of Pseudomonas aeruginosa secreted virulence factors reduces lung inflammation in CF mice. Virulence 2018, 9, 1008–1018. [Google Scholar] [CrossRef] [PubMed]
- Llanos, A.; Achard, P.; Bousquet, J.; Lozano, C.; Zalacain, M.; Sable, C.; Revillet, H.; Murris, M.; Mittaine, M.; Lemonnier, M.; et al. Higher levels of Pseudomonas aeruginosa LasB elastase expression are associated with early-stage infection in cystic fibrosis patients. Scientific Reports 2023, 13. [Google Scholar] [CrossRef]
- Passos da Silva, D.; Matwichuk, M.L.; Townsend, D.O.; Reichhardt, C.; Lamba, D.; Wozniak, D.J.; Parsek, M.R. The Pseudomonas aeruginosa lectin LecB binds to the exopolysaccharide Psl and stabilizes the biofilm matrix. Nature Communications 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Kreindler, J.L.; Bjarnsholt, T.; Jensen, P.Ø.; Jakobsen, T.H.; Phipps, R.; Nielsen, A.K.; Rybtke, M.T.; Tolker-Nielsen, T.; Givskov, M.; Høiby, N.; et al. Quorum Sensing and Virulence of Pseudomonas aeruginosa during Lung Infection of Cystic Fibrosis Patients. PLoS ONE 2010, 5, e10115. [Google Scholar] [CrossRef]
- Cooper, C.E.; Brown, G.C. The inhibition of mitochondrial cytochrome oxidase by the gases carbon monoxide, nitric oxide, hydrogen cyanide and hydrogen sulfide: chemical mechanism and physiological significance. Journal of Bioenergetics and Biomembranes 2008, 40. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, F.J.; Belcher, J.; Jones, A.M.; Smith, D.; Smyth, A.R.; Southern, K.W.; Španěl, P.; Webb, A.K.; Lenney, W. Exhaled breath hydrogen cyanide as a marker of earlyPseudomonas aeruginosainfection in children with cystic fibrosis. ERJ Open Research 2015, 1, 00044–02015. [Google Scholar] [CrossRef] [PubMed]
- Britigan, B.E.; Rasmussen, G.T.; Cox, C.D. Augmentation of oxidant injury to human pulmonary epithelial cells by the Pseudomonas aeruginosa siderophore pyochelin. Infection and Immunity 1997, 65, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Banin, E.; Vasil, M.L.; Greenberg, E.P. Iron and Pseudomonas aeruginosa biofilm formation. Proceedings of the National Academy of Sciences 2005, 102, 11076–11081. [Google Scholar] [CrossRef]
- Ibrahim, B.M.; Tsifansky, M.D.; Yang, Y.; Yeo, Y. Challenges and advances in the development of inhalable drug formulations for cystic fibrosis lung disease. Expert Opinion on Drug Delivery 2011, 8, 451–466. [Google Scholar] [CrossRef]
- Rosenfeld, M.; Rayner, O.; Smyth, A.R. Prophylactic anti-staphylococcal antibiotics for cystic fibrosis. Cochrane Database of Systematic Reviews 2020, 2020. [Google Scholar] [CrossRef]
- Dudley, M.N.; Loutit, J.; Griffith, D.C. Aerosol antibiotics: considerations in pharmacological and clinical evaluation. Current Opinion in Biotechnology 2008, 19, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, B.C.; McColley, S.A.; Kissner, D.G.; Rolfe, M.W.; Rosen, J.M.; McKevitt, M.; Moorehead, L.; Montgomery, A.B.; Geller, D.E. Fosfomycin/Tobramycin for Inhalation in Patients with Cystic Fibrosis with Pseudomonas Airway Infection. American Journal of Respiratory and Critical Care Medicine 2012, 185, 171–178. [Google Scholar] [CrossRef]
- Banerjee, D.; Stableforth, D. The Treatment of Respiratory Pseudomonas Infection in Cystic Fibrosis. Drugs 2000, 60, 1053–1064. [Google Scholar] [CrossRef] [PubMed]
- Zobell, J.T.; Young, D.C.; Waters, C.D.; Stockmann, C.; Ampofo, K.; Sherwin, C.M.T.; Spigarelli, M.G. Optimization of anti-pseudomonal antibiotics for cystic fibrosis pulmonary exacerbations: I. aztreonam and carbapenems. Pediatric Pulmonology 2012, 47, 1147–1158. [Google Scholar] [CrossRef] [PubMed]
- Zobell, J.T.; Waters, C.D.; Young, D.C.; Stockmann, C.; Ampofo, K.; Sherwin, C.M.T.; Spigarelli, M.G. Optimization of anti-pseudomonal antibiotics for cystic fibrosis pulmonary exacerbations: II. cephalosporins and penicillins. Pediatric Pulmonology 2012, 48, 107–122. [Google Scholar] [CrossRef] [PubMed]
- Oermann, C.M.; Retsch-Bogart, G.Z.; Quittner, A.L.; Gibson, R.L.; McCoy, K.S.; Montgomery, A.B.; Cooper, P.J. An 18-month study of the safety and efficacy of repeated courses of inhaled aztreonam lysine in cystic fibrosis. Pediatric Pulmonology 2010, 45, 1121–1134. [Google Scholar] [CrossRef] [PubMed]
- Döring, G.; Meisner, C.; Stern, M. A double-blind randomized placebo-controlled phase III study of a Pseudomonas aeruginosa flagella vaccine in cystic fibrosis patients. Proceedings of the National Academy of Sciences 2007, 104, 11020–11025. [Google Scholar] [CrossRef]
- Pettit, R.S.; Fellner, C. CFTR Modulators for the Treatment of Cystic Fibrosis. P T 2014, 39, 500–511. [Google Scholar] [PubMed]
- Wainwright, C.E.; Elborn, J.S.; Ramsey, B.W.; Marigowda, G.; Huang, X.; Cipolli, M.; Colombo, C.; Davies, J.C.; De Boeck, K.; Flume, P.A.; et al. Lumacaftor–Ivacaftor in Patients with Cystic Fibrosis Homozygous for Phe508delCFTR. New England Journal of Medicine 2015, 373, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Lister, P.D.; Wolter, D.J.; Hanson, N.D. Antibacterial-ResistantPseudomonas aeruginosa: Clinical Impact and Complex Regulation of Chromosomally Encoded Resistance Mechanisms. Clinical Microbiology Reviews 2009, 22, 582–610. [Google Scholar] [CrossRef]
- Meletis, G.; Bagkeri, M. Pseudomonas aeruginosa: Multi-Drug-Resistance Development and Treatment Options. 2013. [Google Scholar] [CrossRef]
- Dettman, J.R.; Sztepanacz, J.L.; Kassen, R. The properties of spontaneous mutations in the opportunistic pathogen Pseudomonas aeruginosa. BMC Genomics 2016, 17. [Google Scholar] [CrossRef]
- Moradali, M.F.; Ghods, S.; Rehm, B.H.A. Pseudomonas aeruginosa Lifestyle: A Paradigm for Adaptation, Survival, and Persistence. Frontiers in Cellular and Infection Microbiology 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Sun, E.; Gill, E.E.; Falsafi, R.; Yeung, A.; Liu, S.; Hancock, R.E.W. Broad-Spectrum Adaptive Antibiotic Resistance Associated with Pseudomonas aeruginosa Mucin-Dependent Surfing Motility. Antimicrobial Agents and Chemotherapy 2018, 62. [Google Scholar] [CrossRef]
- Coleman, S.R.; Blimkie, T.; Falsafi, R.; Hancock, R.E.W. Multidrug Adaptive Resistance of Pseudomonas aeruginosa Swarming Cells. Antimicrobial Agents and Chemotherapy 2020, 64. [Google Scholar] [CrossRef]
- Prayle, A.; Watson, A.; Fortnum, H.; Smyth, A. Side effects of aminoglycosides on the kidney, ear and balance in cystic fibrosis. Thorax 2010, 65, 654–658. [Google Scholar] [CrossRef] [PubMed]
- Patangia, D.V.; Anthony Ryan, C.; Dempsey, E.; Paul Ross, R.; Stanton, C. Impact of antibiotics on the human microbiome and consequences for host health. MicrobiologyOpen 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Burrows, J.A.; Toon, M.; Bell, S.C. Antibiotic desensitization in adults with cystic fibrosis. Respirology 2003, 8, 359–364. [Google Scholar] [CrossRef]
- Tseng, B.S.; Zhang, W.; Harrison, J.J.; Quach, T.P.; Song, J.L.; Penterman, J.; Singh, P.K.; Chopp, D.L.; Packman, A.I.; Parsek, M.R. The extracellular matrix protects Pseudomonas aeruginosa biofilms by limiting the penetration of tobramycin. Environmental Microbiology 2013, 15, 2865–2878. [Google Scholar] [CrossRef]
- Ho, D.-K.; Nichols, B.L.B.; Edgar, K.J.; Murgia, X.; Loretz, B.; Lehr, C.-M. Challenges and strategies in drug delivery systems for treatment of pulmonary infections. European Journal of Pharmaceutics and Biopharmaceutics 2019, 144, 110–124. [Google Scholar] [CrossRef]
- Dickey, S.W.; Cheung, G.Y.C.; Otto, M. Different drugs for bad bugs: antivirulence strategies in the age of antibiotic resistance. Nature Reviews Drug Discovery 2017, 16, 457–471. [Google Scholar] [CrossRef]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal–response systems in Gram-negative bacteria. Nature Reviews Microbiology 2016, 14, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Schuster, M.; Peter Greenberg, E. A network of networks: Quorum-sensing gene regulation in Pseudomonas aeruginosa. International Journal of Medical Microbiology 2006, 296, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Heeb, S.; Fletcher, M.P.; Chhabra, S.R.; Diggle, S.P.; Williams, P.; Cámara, M. Quinolones: from antibiotics to autoinducers. FEMS Microbiology Reviews 2011, 35, 247–274. [Google Scholar] [CrossRef]
- Thi, M.T.T.; Wibowo, D.; Rehm, B.H.A. Pseudomonas aeruginosa Biofilms. International Journal of Molecular Sciences 2020, 21, 8671. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sass, A.; Van Acker, H.; Wille, J.; Verhasselt, B.; Van Nieuwerburgh, F.; Kaever, V.; Crabbé, A.; Coenye, T. Coumarin Reduces Virulence and Biofilm Formation in Pseudomonas aeruginosa by Affecting Quorum Sensing, Type III Secretion and C-di-GMP Levels. Frontiers in Microbiology 2018, 9. [Google Scholar] [CrossRef]
- Seleem, M.N.; Luo, J.; Dong, B.; Wang, K.; Cai, S.; Liu, T.; Cheng, X.; Lei, D.; Chen, Y.; Li, Y.; et al. Baicalin inhibits biofilm formation, attenuates the quorum sensing-controlled virulence and enhances Pseudomonas aeruginosa clearance in a mouse peritoneal implant infection model. PLoS ONE 2017, 12, e0176883. [Google Scholar] [CrossRef]
- Vrenna, G.; Artini, M.; Ragno, R.; Relucenti, M.; Fiscarelli, E.V.; Tuccio Guarna Assanti, V.; Papa, R.; Selan, L. Anti-Virulence Properties of Coridothymus capitatus Essential Oil against Pseudomonas aeruginosa Clinical Isolates from Cystic Fibrosis Patients. Microorganisms 2021, 9, 2257. [Google Scholar] [CrossRef] [PubMed]
- Elekhnawy, E.; Negm, W.A.; El-Aasr, M.; Kamer, A.A.; Alqarni, M.; Batiha, G.E.-S.; Obaidullah, A.J.; Fawzy, H.M. Histological assessment, anti-quorum sensing, and anti-biofilm activities of Dioon spinulosum extract: in vitro and in vivo approach. Scientific Reports 2022, 12. [Google Scholar] [CrossRef]
- Nizalapur, S.; Kimyon, Ö.; Biswas, N.N.; Gardner, C.R.; Griffith, R.; Rice, S.A.; Manefield, M.; Willcox, M.; Black, D.S.; Kumar, N. Design, synthesis and evaluation of N-aryl-glyoxamide derivatives as structurally novel bacterial quorum sensing inhibitors. Organic & Biomolecular Chemistry 2016, 14, 680–693. [Google Scholar] [CrossRef]
- Chang, C.Y.; Krishnan, T.; Wang, H.; Chen, Y.; Yin, W.F.; Chong, Y.M.; Tan, L.Y.; Chong, T.M.; Chan, K.-G. Non-antibiotic quorum sensing inhibitors acting against N-acyl homoserine lactone synthase as druggable target. Scientific Reports 2014, 4. [Google Scholar] [CrossRef]
- O’Loughlin, C.T.; Miller, L.C.; Siryaporn, A.; Drescher, K.; Semmelhack, M.F.; Bassler, B.L. A quorum-sensing inhibitor blocks Pseudomonas aeruginosa virulence and biofilm formation. Proceedings of the National Academy of Sciences 2013, 110, 17981–17986. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Maurer, C.K.; Kirsch, B.; Steinbach, A.; Hartmann, R.W. Overcoming the Unexpected Functional Inversion of a PqsR Antagonist in Pseudomonas aeruginosa: An In Vivo Potent Antivirulence Agent Targeting pqs Quorum Sensing. Angewandte Chemie International Edition 2013, 53, 1109–1112. [Google Scholar] [CrossRef] [PubMed]
- Whiteley, M.; Starkey, M.; Lepine, F.; Maura, D.; Bandyopadhaya, A.; Lesic, B.; He, J.; Kitao, T.; Righi, V.; Milot, S.; et al. Identification of Anti-virulence Compounds That Disrupt Quorum-Sensing Regulated Acute and Persistent Pathogenicity. PLoS Pathogens 2014, 10, e1004321. [Google Scholar] [CrossRef]
- D’Angelo, F.; Baldelli, V.; Halliday, N.; Pantalone, P.; Polticelli, F.; Fiscarelli, E.; Williams, P.; Visca, P.; Leoni, L.; Rampioni, G. Identification of FDA-Approved Drugs as Antivirulence Agents Targeting the pqs Quorum-Sensing System of Pseudomonas aeruginosa. Antimicrobial Agents and Chemotherapy 2018, 62. [Google Scholar] [CrossRef]
- Schulenburg, H.; Rezzoagli, C.; Archetti, M.; Mignot, I.; Baumgartner, M.; Kümmerli, R. Combining antibiotics with antivirulence compounds can have synergistic effects and reverse selection for antibiotic resistance in Pseudomonas aeruginosa. PLOS Biology 2020, 18, e3000805. [Google Scholar] [CrossRef]
- Hwang, H.-J.; Choi, H.; Hong, S.; Moon, H.R.; Lee, J.-H.; Oglesby, A.G. Antipathogenic Compounds That Are Effective at Very Low Concentrations and Have Both Antibiofilm and Antivirulence Effects against Pseudomonas aeruginosa. Microbiology Spectrum 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Dawoud, N.T.A.; El-Fakharany, E.M.; Abdallah, A.E.; El-Gendi, H.; Lotfy, D.R. Synthesis, and docking studies of novel heterocycles incorporating the indazolylthiazole moiety as antimicrobial and anticancer agents. Scientific Reports 2022, 12. [Google Scholar] [CrossRef]
- Cutruzzolà, F.; Frankenberg-Dinkel, N.; O’Toole, G.A. Origin and Impact of Nitric Oxide in Pseudomonas aeruginosa Biofilms. Journal of Bacteriology 2016, 198, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhang, P.; Guo, T.; Gu, X.; Bai, B.; Zhang, S.; Chang, X.; Wang, Y.; Ma, S. Design, synthesis and evaluation of oxazolopyridinone derivatives as quorum sensing inhibitors. Bioorganic Chemistry 2023, 130, 106266. [Google Scholar] [CrossRef]
- Blanco-Cabra, N.; Movellan, J.; Marradi, M.; Gracia, R.; Salvador, C.; Dupin, D.; Loinaz, I.; Torrents, E. Neutralization of ionic interactions by dextran-based single-chain nanoparticles improves tobramycin diffusion into a mature biofilm. npj Biofilms and Microbiomes 2022, 8. [Google Scholar] [CrossRef]
- Alontaga, A.Y.; Rodriguez, J.C.; Schönbrunn, E.; Becker, A.; Funke, T.; Yukl, E.T.; Hayashi, T.; Stobaugh, J.; Moënne-Loccoz, P.; Rivera, M. Structural Characterization of the Hemophore HasAp from Pseudomonas aeruginosa: NMR Spectroscopy Reveals Protein−Protein Interactions between Holo-HasAp and Hemoglobin. Biochemistry 2008, 48, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, P. Iron uptake and metabolism in pseudomonads. Applied Microbiology and Biotechnology 2010, 86, 1637–1645. [Google Scholar] [CrossRef] [PubMed]
- Gifford, A.H.; Moulton, L.A.; Dorman, D.B.; Olbina, G.; Westerman, M.; Parker, H.W.; Stanton, B.A.; O’Toole, G.A. Iron Homeostasis during Cystic Fibrosis Pulmonary Exacerbation. Clinical and Translational Science 2012, 5, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Visca, P.; Imperi, F.; Lamont, I.L. Pyoverdine siderophores: from biogenesis to biosignificance. Trends in Microbiology 2007, 15, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, P.; Matthijs, S.; Van Oeffelen, L. Iron uptake regulation in Pseudomonas aeruginosa. BioMetals 2009, 22, 15–22. [Google Scholar] [CrossRef]
- Prince, R.W.; Cox, C.D.; Vasil, M.L. Coordinate regulation of siderophore and exotoxin A production: molecular cloning and sequencing of the Pseudomonas aeruginosa fur gene. Journal of Bacteriology 1993, 175, 2589–2598. [Google Scholar] [CrossRef]
- Kaneko, Y.; Thoendel, M.; Olakanmi, O.; Britigan, B.E.; Singh, P.K. The transition metal gallium disrupts Pseudomonas aeruginosa iron metabolism and has antimicrobial and antibiofilm activity. Journal of Clinical Investigation 2007, 117, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K. Iron sequestration by human lactoferrin stimulates P. aeruginosa surface motility and blocks biofilm formation. BioMetals 2004, 17, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Braun, V. Active transport of iron and siderophore antibiotics. Current Opinion in Microbiology 2002, 5, 194–201. [Google Scholar] [CrossRef]
- Negash; Norris; Hodgkinson. Siderophore–Antibiotic Conjugate Design: New Drugs for Bad Bugs? Molecules 2019, 24, 3314. [Google Scholar] [CrossRef]
- Moreau-Marquis, S.; O’Toole, G.A.; Stanton, B.A. Tobramycin and FDA-Approved Iron Chelators Eliminate Pseudomonas aeruginosa Biofilms on Cystic Fibrosis Cells. American Journal of Respiratory Cell and Molecular Biology 2009, 41, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Tielker, D.; Hacker, S.; Loris, R.; Strathmann, M.; Wingender, J.; Wilhelm, S.; Rosenau, F.; Jaeger, K.-E. Pseudomonas aeruginosa lectin LecB is located in the outer membrane and is involved in biofilm formation. Microbiology 2005, 151, 1313–1323. [Google Scholar] [CrossRef] [PubMed]
- Adam, E.C.; Mitchell, B.S.; Schumacher, D.U.; Grant, G.; Schumacher, U. Pseudomonas aeruginosa II lectin stops human ciliary beating: therapeutic implications of fucose. American Journal of Respiratory and Critical Care Medicine 1997, 155, 2102–2104. [Google Scholar] [CrossRef] [PubMed]
- Kadam, R.U.; Bergmann, M.; Hurley, M.; Garg, D.; Cacciarini, M.; Swiderska, M.A.; Nativi, C.; Sattler, M.; Smyth, A.R.; Williams, P.; et al. A Glycopeptide Dendrimer Inhibitor of the Galactose-Specific Lectin LecA and of Pseudomonas aeruginosa Biofilms. Angewandte Chemie International Edition 2011, 50, 10631–10635. [Google Scholar] [CrossRef] [PubMed]
- Johansson, E.M.V.; Crusz, S.A.; Kolomiets, E.; Buts, L.; Kadam, R.U.; Cacciarini, M.; Bartels, K.-M.; Diggle, S.P.; Cámara, M.; Williams, P.; et al. Inhibition and Dispersion of Pseudomonas aeruginosa Biofilms by Glycopeptide Dendrimers Targeting the Fucose-Specific Lectin LecB. Chemistry & Biology 2008, 15, 1249–1257. [Google Scholar] [CrossRef]
- Hauber, H.P.; Schulz, M.; Pforte, A.; Mack, D.; Zabel, P.; Schumacher, U. Inhalation with Fucose and Galactose for Treatment of Pseudomonas Aeruginosa in Cystic Fibrosis Patients. International Journal of Medical Sciences 2008, 371–376. [Google Scholar] [CrossRef]
- Kolomiets, E.; Swiderska, M.A.; Kadam, R.U.; Johansson, E.M.V.; Jaeger, K.E.; Darbre, T.; Reymond, J.L. Glycopeptide Dendrimers with High Affinity for the Fucose-Binding Lectin LecB from Pseudomonas aeruginosa. ChemMedChem 2009, 4, 562–569. [Google Scholar] [CrossRef]
- Illyés, T.Z.; Malinovská, L.; Rőth, E.; Tóth, B.; Farkas, B.; Korsák, M.; Wimmerová, M.; Kövér, K.E.; Csávás, M. Synthesis of Tetravalent Thio- and Selenogalactoside-Presenting Galactoclusters and Their Interactions with Bacterial Lectin PA-IL from Pseudomonas aeruginosa. Molecules 2021, 26, 542. [Google Scholar] [CrossRef] [PubMed]
- Lorusso, A.B.; Carrara, J.A.; Barroso, C.D.N.; Tuon, F.F.; Faoro, H. Role of Efflux Pumps on Antimicrobial Resistance in Pseudomonas aeruginosa. International Journal of Molecular Sciences 2022, 23, 15779. [Google Scholar] [CrossRef]
- Sanchez, P. Fitness of in vitro selected Pseudomonas aeruginosanalB and nfxB multidrug resistant mutants. Journal of Antimicrobial Chemotherapy 2002, 50, 657–664. [Google Scholar] [CrossRef]
- Rampioni, G.; Pillai, C.R.; Longo, F.; Bondì, R.; Baldelli, V.; Messina, M.; Imperi, F.; Visca, P.; Leoni, L. Effect of efflux pump inhibition on Pseudomonas aeruginosa transcriptome and virulence. Scientific Reports 2017, 7. [Google Scholar] [CrossRef]
- Webber, M.A.; Lamers, R.P.; Cavallari, J.F.; Burrows, L.L. The Efflux Inhibitor Phenylalanine-Arginine Beta-Naphthylamide (PAβN) Permeabilizes the Outer Membrane of Gram-Negative Bacteria. PLoS ONE 2013, 8, e60666. [Google Scholar] [CrossRef]
- Gbian, D.L.; Omri, A. The Impact of an Efflux Pump Inhibitor on the Activity of Free and Liposomal Antibiotics against Pseudomonas aeruginosa. Pharmaceutics 2021, 13, 577. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Rosado-Lugo, J.D.; Datta, P.; Sun, Y.; Cao, Y.; Banerjee, A.; Yuan, Y.; Parhi, A.K. Evaluation of a Conformationally Constrained Indole Carboxamide as a Potential Efflux Pump Inhibitor in Pseudomonas aeruginosa. Antibiotics 2022, 11, 716. [Google Scholar] [CrossRef]
- Lee, M.D.; Galazzo, J.L.; Staley, A.L.; Lee, J.C.; Warren, M.S.; Fuernkranz, H.; Chamberland, S.; Lomovskaya, O.; Miller, G.H. Microbial fermentation-derived inhibitors of efflux-pump-mediated drug resistance. Il Farmaco 2001, 56, 81–85. [Google Scholar] [CrossRef]
- Yoshida, K.-i.; Nakayama, K.; Ohtsuka, M.; Kuru, N.; Yokomizo, Y.; Sakamoto, A.; Takemura, M.; Hoshino, K.; Kanda, H.; Nitanai, H.; et al. MexAB-OprM specific efflux pump inhibitors in Pseudomonas aeruginosa. Part 7: Highly soluble and in vivo active quaternary ammonium analogue D13-9001, a potential preclinical candidate. Bioorganic & Medicinal Chemistry 2007, 15, 7087–7097. [Google Scholar] [CrossRef]
- Nakashima, R.; Sakurai, K.; Yamasaki, S.; Hayashi, K.; Nagata, C.; Hoshino, K.; Onodera, Y.; Nishino, K.; Yamaguchi, A. Structural basis for the inhibition of bacterial multidrug exporters. Nature 2013, 500, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Collalto, D.; Giallonardi, G.; Fortuna, A.; Meneghini, C.; Fiscarelli, E.; Visca, P.; Imperi, F.; Rampioni, G.; Leoni, L. In vitro Activity of Antivirulence Drugs Targeting the las or pqs Quorum Sensing Against Cystic Fibrosis Pseudomonas aeruginosa Isolates. Frontiers in Microbiology 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.F.; Yang, C.; Zhang, Y.; Tao, S.N.; Mei, J.; Zhang, X.C.; Sun, Y.J.; Zhao, B.T. An innovative role for luteolin as a natural quorum sensing inhibitor in Pseudomonas aeruginosa. Life Sciences 2021, 274, 119325. [Google Scholar] [CrossRef]
- Kumar, S.; Paliya, B.S.; Singh, B.N. Superior inhibition of virulence and biofilm formation of Pseudomonas aeruginosa PAO1 by phyto-synthesized silver nanoparticles through anti-quorum sensing activity. Microbial Pathogenesis 2022, 170, 105678. [Google Scholar] [CrossRef]
- Mettrick, K.; Hassan, K.; Lamont, I.; Reid, D. The Iron-chelator, N,N’-bis (2-hydroxybenzyl) Ethylenediamine-N,N’-diacetic acid is an Effective Colistin Adjunct against Clinical Strains of Biofilm-Dwelling Pseudomonas aeruginosa. Antibiotics 2020, 9, 144. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of virulence factors produced by P. aeruginosa that play a critical role in aggravating CF.
Figure 1.
Schematic representation of virulence factors produced by P. aeruginosa that play a critical role in aggravating CF.
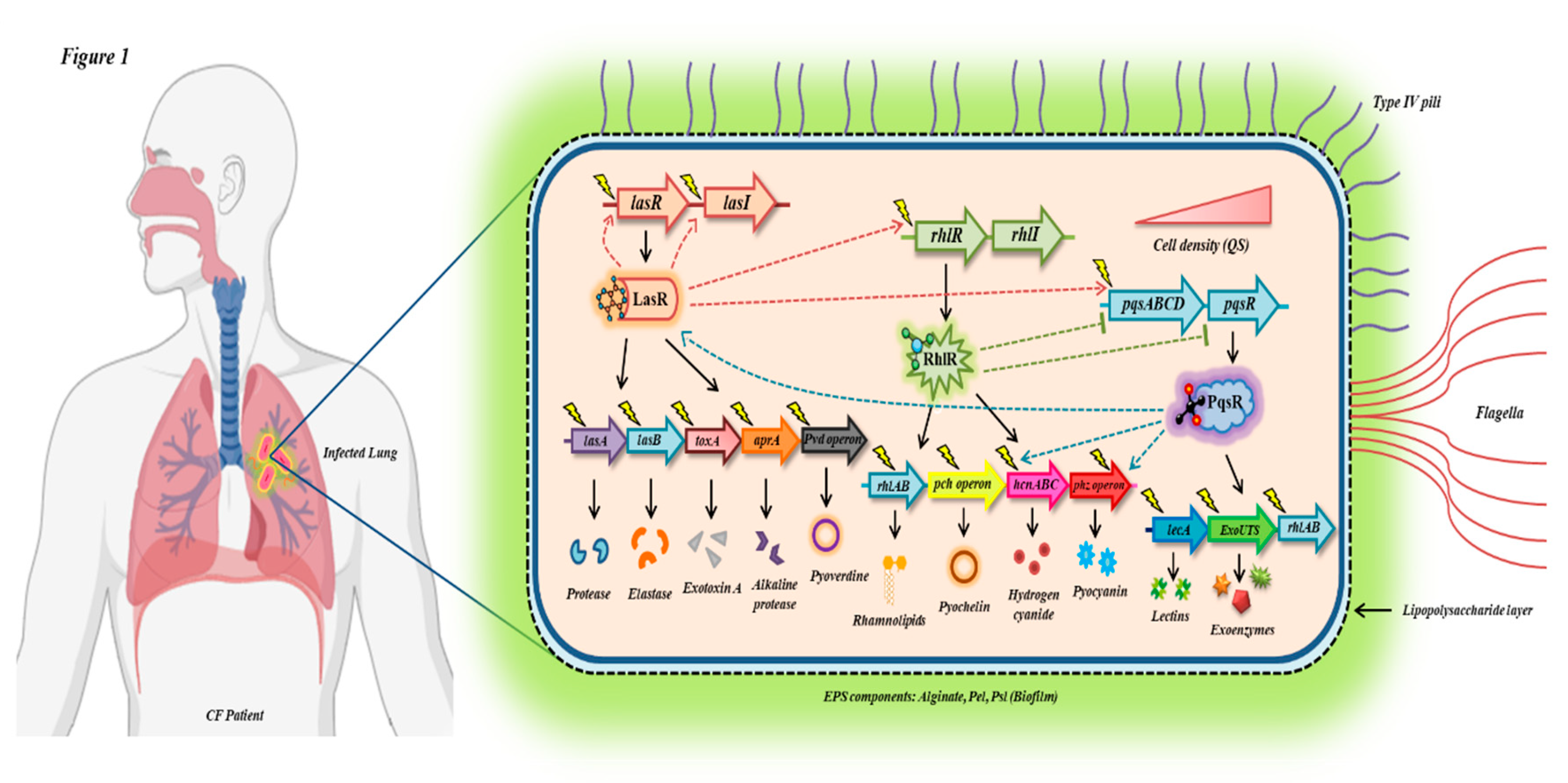
Figure 2.
Depiction of various antivirulence strategies being employed against CF isolates of P. aeruginosa for disarming bacterial virulence and lowering disease severity.
Figure 2.
Depiction of various antivirulence strategies being employed against CF isolates of P. aeruginosa for disarming bacterial virulence and lowering disease severity.
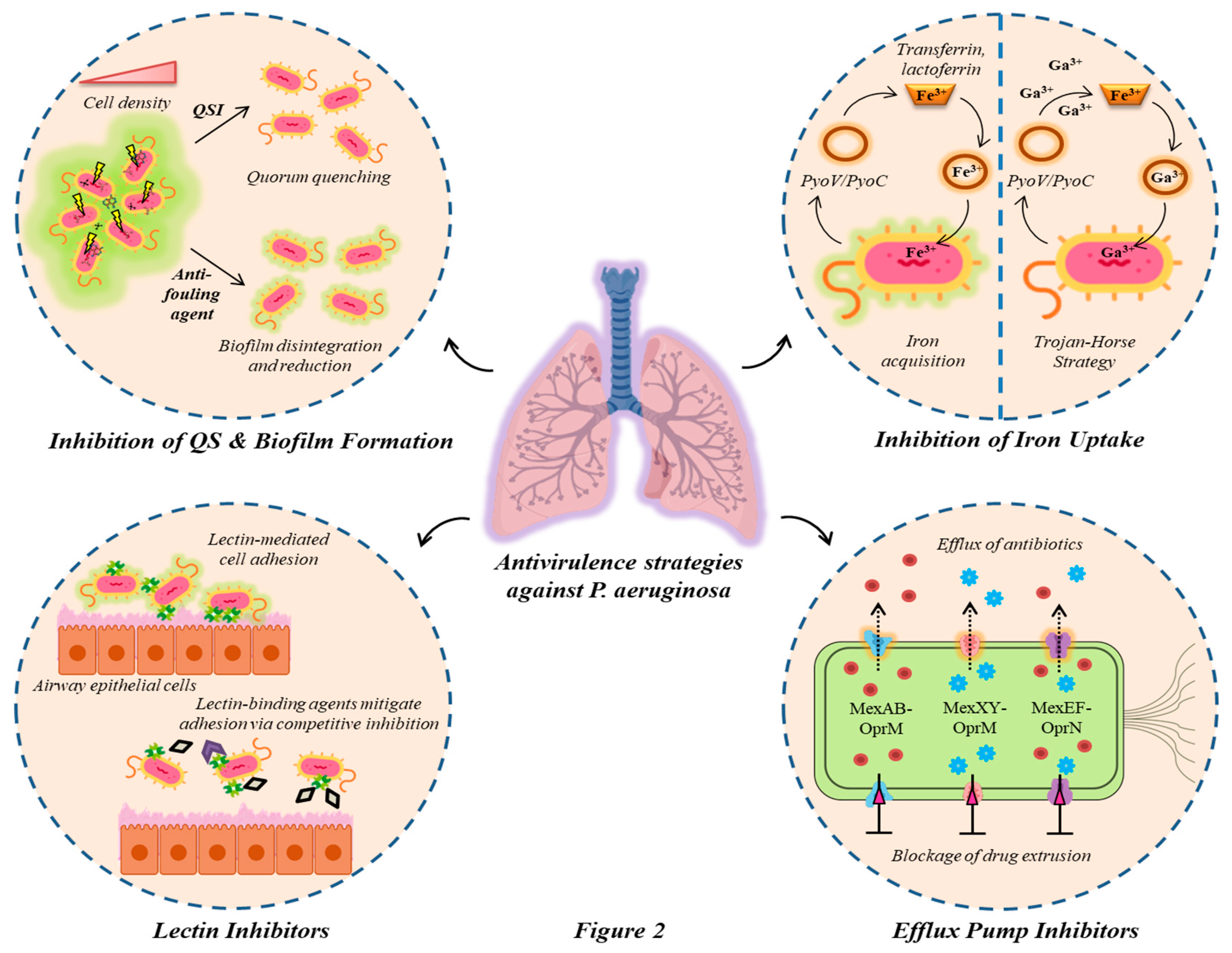
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



