
Submitted:
12 February 2024
Posted:
13 February 2024
You are already at the latest version
Abstract
Plant growth promoting rhizobacteria (PGPR) are a diverse group of microorganisms including genera such as Pseudomonas, Bacillus, Serratia, Enterobacter and Klebsiella. These bacteria have the ability to colonize roots externally and internally, and to directly enhance plant growth employing various mechanisms like phytohormone production and nutrient mineralization. These microbes can also promote plant growth indirectly by secreting antimicrobial compounds (e.g., siderophores, hydrogen cyanide) to antagonize root pathogens or by inducing defensive responses in the plant. Interestingly, there are also cases where production of antibiotic compounds such as 2,4-Diacetylphloroglucinol have the direct effect of enhancing plant growth and yield, irrespective of their biocontrol effects. The objective of this paper is to review publications showcasing PGPRs that have the capacity to both control root pathogens and stimulate plant growth, and to discuss what mechanisms might be involved.
Keywords:
biological control
; growth promotion
; antagonist
; antimicrobial compounds
; plant defense
; soilborne pathogens
1. Introduction
Many root pathogens can persist in and be dispersed by soil. Although root diseases are usually hard to diagnose and treat, they are economically important, estimated to cause 26% of the crop losses that are recorded around the world [1]. They are caused by a very broad group of microbes that include viruses, bacteria, fungi, oomycetes, and nematodes that invade and injure root tissues, often leading to a reduction in the plant’s ability to absorb water and nutrients [2]. Soil pathogens employ chemotaxis to sense and move towards roots which secrete exudates containing sugars and amino acids in order to attract and feed beneficial microbes [3,4]. Arriving at the plant/soil interface, pathogens attach, penetrate, and invade root tissues including epidermis and vasculature, causing root rotting, shoot wilting, or alterations in the germination and growth of the seedlings [2,5,6,7].
Fungi and oomycetes are the most common cause plant pathogens, responsible for some of the most severe plant diseases. One such soil transmitted fungus, Fusarium oxysporum enters the plant through the root and invades its vascular system, causing significant economic losses in a wide range of crops, including chickpeas, potatoes, strawberries, and tomatoes; for the latter, yield losses can be as high as 90% or 95% in Tunisian or Canadian farms, respectively [8,9,10,11,12]. Late blight can also be transmitted by soil and is caused by species of Phytophthora which can reduce potato plantation yields by up to 75% [13]. Between the years 1845-52, late blight of disease susceptible potatoes grown by Irish potato farmers led to the starvation of 1.5 million people [14]. Another soil transmitted pathogen, Pythium infects root tissues of both seedlings and adult plants, causing major economic losses exceeding 50% in soybean, maize and ginger crops in the United States and India [15,16]. Rhizoctonia solani has been reported to occur in 50-60% of rice plantations and can create infestations with losses approaching 20% [17,18], while Meloidogyne incognita also infects roots and can reduce yields up to 33% [19,20]. A few recent papers review root diseases and methods to manage them including chemical or biological control [21,22].
Given the severity of these diseases and the economic losses they cause, farmers have tried various techniques to control soilborne pathogens. Some techniques include organic amendments such as compost teas or liquid swine manure, while others attempt to treat the soil by withdrawing oxygen (anerobic soil disinfestation) or injecting hot steam to kill microbes [23,24]. More effective at killing soil microbes is the use of agrochemicals for disease control in crop fields [25,26]. These toxic chemicals are used to fumigate the soil before planting to reduce the inoculum of all bacteria, fungi and/or nematodes, and at the same time they can increase plant yields by improving traits such as root density and productivity [27]. Examples of these pesticides include pentachloronitrobenzene (PCNB), metalaxyl, triadimefon, etazol, propamocarb, furalaxyl, myclobutanil, nabam, chloropicrin and methyl bromide [6].
However effective at killing that these fumigants may be, excessive use of pesticides leads to problems such as economic waste, low nutrient use efficiency, reduction of soil quality (i.e. killing beneficial microbes, loss of organic matter), and environmental pollution [28,29,30]. It has been estimated that in some cases only 0.1% of the applied pesticide reaches the target pest, while the rest drifts into the surrounding environment where it can be incorporated into nutrient and biological cycles, thus persisting for long periods of time, and contaminating soil, water, and air where it continues to poison or affect non-target lifeforms [31,32,33]. Metalaxyl, triadimefon and chloropicrin have been observed to modify soil bacterial populations and fertility, reducing the function of biogeochemical cycles that are mediated by soil microorganisms [34,35,36,37,38]. Because methyl bromide has strongly negative effects on the ozone layer, its use has been banned in many countries in a campaign supported by the United Nations [39]. Chemicals like cypermethrin, chlorpyrifos, endosulfan, glyphosate, and 2,4-Dichlorophenoxyacetic acid, have been found to interfere with amphibian reproduction and survival [40]; flupyradifurone and neonicotinoids are pesticides that can also kill honey bees which are critical to the production of fruit crops [41,42,43]; pesticides like deltamethrin, hexachlorobenzene accumulate in animal tissues, leading to biomagnification throughout the food chain and early death in apex predators [44,45]. Chemical fumigants have also been reported to directly harm humans; according to the World Health Organization, more than 25 million farmers are poisoned each year by the use of agricultural pesticides [46]. Pentachloronitrobenzene (PCNB) is one example, considered by the EPA as a potential carcinogen [47], and propamocarb is another, which changes the intestinal microbiota and metabolism of mammals altering it normal function [48]. Toxic soil fumigants have many unintended negative effects, making it clearly necessary to look for alternatives that are more precise/targeted while retaining the efficiency at controlling soil pathogens.
A promising alternative to toxic fumigants is biological control, where plants are protected against root pathogens through soil application of antagonistic microorganisms, either fungal or bacterial, that kill or otherwise inhibit growth and activity of the infectious agent. Biocontrol of root pathogens may involve mechanisms such as the production of antibiotics, predation, parasitization, competitive exclusion of space or nutrients, production of enzymes that affect cellular components of the pathogen, or by the induction of defensive responses in the plant [49,50,51]. Many of the microorganisms that can be used for biological control in the soil are found in plant rhizospheres; a niche that is considered one of the most diverse ecosystems on Earth [52]. Many of these bacteria belong to genera such as Pseudomonas, Bacillus, Enterobacter, Klebsiella, Serratia, Azospirillum, Chrysobacterium, and Burkholderia, colonizing roots and stimulating plant growth through nitrogen fixation, phosphate solubilization, and production of growth-promoting hormones [53,54,55,56,57]. These microorganisms are known as plant growth-promoting rhizobacteria (PGPR), and within this group there are also some with biocontrol potential, as they can secrete siderophores, they can produce antimicrobial compounds and enzymes, or they can induce intrinsic plant defense responses such as systemic acquired resistance (SAR) [58,59,60]. Curiously, some antimicrobial compounds involved in pathogen biocontrol can also directly stimulate plant growth, thereby increasing plant productivity while aiding in plant defense [61,62,63]. PGPRs that offer the dual benefit of both controlling root pathogens and of increasing plant growth are likely to be ideal candidates for production of commercial inoculants, as big, healthy plants growing without pathogen pressure have higher yields than smaller plants under attack by soil pathogens [64,65]. The purpose of this review is to review the literature showing examples of PGPRs controlling root pathogens (especially soil-borne pathogens), while simultaneously stimulating plant growth.
2. Siderophore production
Iron is an essential mineral for the development of virtually all living organisms, acting as a cofactor approximately 140 different enzymes that catalyze biochemical reactions in primary metabolism, such as the reduction of ribonucleotides, the activation and decomposition of peroxides, and the transport of electrons through a variety of carriers [66,67]. In plants, iron is involved in the synthesis of chlorophyll, thylakoids, and chloroplasts [68] and is thus a critical mineral for growth. Under normal, aerobic conditions, soil iron is mostly present in an insoluble form (Fe3+) and is therefore not readily available for uptake by either roots or soil microbes [69,70]. To aid in uptake of soil iron, both microorganisms and plants secrete low molecular weight compounds called siderophores, which have a high affinity for this element and are easier to take up than the pure mineral [71]. One of the most common siderophores is pioverdine, a greenish-yellow, fluorescent pigment produced by bacteria of the genus Pseudomonas; cosmopolitan microorganisms that are frequently found in plant roots as growth promoters [72]. A Pseudomonas that has locked up a significant amount of soil iron using its pioverdine has increased the amount of iron available to itself, while blocking its access to other microbes which lack mechanisms to take up pioverdine. Next to Pseudomonas, Bacillus is the genus with the most examples of siderophore producing PGPR, where they secrete molecules like bacillibactin, hydroxamate, pyoverdine, pyochelin, 2,3- dihydroxybenzoic acid (DHBA), etc. to chelate soil iron [73,74,75,76]. Because of these properties, siderophores have become one of the best-known mechanisms of microbial biocontrol, depriving soil pathogens of this element, thus limiting their ability to proliferate and infect the root [77,78,79].
Siderophore based root pathogen biocontrol plus plant growth stimulation has been demonstrated by rhizobacteria such as Pseudomonas aeruginosa, Pseudomonas putida and Pseudomonas alcaligenes, which can inhibit up to 80% of wilt symptoms caused by F. oxysporum in tomato plants [80] or which significantly reduce nematode (Meloidogyne javanica) egg hatching, lentil root infection and disease gall formation when they are applied as a soil drench [81]. Bacteria of the genus Bacillus like B. amyloliquefaciens or B. subtilis also have the ability to produce siderophores such as bacillibactin, a molecule involved in the in vitro inhibition of pathogens like Rhizoctonia solani, F. oxysporum f. sp. radicis-lycopersici or Pseudomonas syringae pv. tomato [82]. Inoculation of siderophore producing strains of Bacillus onto roots infected with Pyricularia oryzae can reduce rice blast disease symptoms by up to 80% [83,84,85]; inoculation of biocontrol Bacillus into potato and banana roots inhibited disease caused by F. oxysporum and P. infestans by up to 88% and 65% respectively [86,87]. In addition to Pseudomonas and Bacillus, siderophores have also been documented in bacteria of the genera Serratia, Azospirillum, Enterobacter, Burkholderia and Klebsiella, where they have been implicated in the control of a wide range of plant pathogens such as Colletotrichum acutatum, F. oxysporum f. sp. radicis lycopersici (FORL), Botrytis cinerea, Cryphonectria parasitica, Rhizoctonia cerealis, Valsa sordida and R. solani, reducing plant disease symptoms by up to 48% in crops including rice, sunflower, strawberry and tomato [88,89,90,91,92,93,94,95,96,97,98,99]. Experiments with cell free, siderophore rich supernatants have various times been shown to have strong antimicrobial properties even compared with chemical fungicides like carbendazim [100,101] and have even found use as agricultural products in of themselves [72,102,103]. The use of siderophore producing strains of PGPR for biocontrol of root diseases have been reviewed especially for the application of Pseudomonas and Bacillus for controlling root pathogens [104,105,106] such as Cephalosporium maydis, R. solani and Colletotrichum gloeosporioides in maize and chilli [107,108]
Besides controlling disease, PGPR produced siderophores can also stimulate plant growth, most likely by alleviating disease pressure, reducing abiotic stress, improving plant mineral nutrition, and by influencing levels of phytohormones present in the rhizosphere. It has been shown experimentally that Streptomyces-derived, cell-free liquid supernatant containing siderophores and auxins, when added to heavy metal contaminated soil can increase the amount of iron absorbed by cowpeas, increase chlorophyll content, and reduce both lipid peroxidation and free radical formation, which in turn protected microbial auxins from degradation, allowing them to accumulate and enhance plant growth [109]. Inoculation of mung beans in both gnotobiotic and soil-based growth assays showed that the siderophore 17X overproducing mutant Pseudomonas fluorescens ATCC 13525 stimulated significantly more plant growth than inoculation with the wildtype strain did 61. Another study of a strongly wheat growth promoting and Pythium inhibiting strain of Pseudomonas showed that when siderophore production was eliminated by mutation, both biocontrol and plant growth promotion were also eliminated [110]. In several trials where Vigna radiata plants were inoculated with different strains of siderophore producing Pseudomonas, root and shoot growth stimulation correlated with an increase in chlorophyll synthesis and the iron available to the plant [61,111]. Inoculation of chickpea plants with P. fluorescens resulted in increases of dry root biomass of up to 70% [112]. Roots of potato plants colonized by strains of siderophore-producing P. fluorescens, enjoyed an enhanced tuber yield of up to 144% although the exact mechanism involved was unknown [113]. An endophytic, siderophore producing Streptomyces isolated from the roots of rice was able to significantly increase the length and biomass of pot grown rice and mungbean plants, in contrast to untreated controls and siderophore-deficient mutant treatments which didn’t – the authors speculate that this growth promotion was due to improved plant mineral nutrition [114]. Several reports of siderophore producing and plant growth promoting bacilli exist: a strain of B. subtilis in pepper plants where it enhanced plant development, resulting in an increase of up to 50% and 37% in size and fruit weight respectively [115]; Bacillus shackletonii in pot grown soybeans had up to 20% increases in germination rate, 20% increases in root length and 230% increases in shoot length [116]; bacillibactin-producing B. subtilis improved the growth of Sesamum indicum maintained in pots by up to 32% in leaf length, 39% in shoot length, 43% in root length and 27% in fresh weight [73]. The contribution of Bacillus to crop biocontrol and growth promotion has recently been reviewed [117]. Meta-analysis of over 300 publications on siderophore-producing bacteria found that they also consistently stimulate plant growth, particularly in the plant families Musaceae, Solanaceae, Cucurbitaceae, Cruciferae, and Poaceae, with an average observed increase of up to 30% in plant weight, height, and germination rate [118]. Another interesting review contrasts the biocontrol and plant growth potential of the two most important siderophore producing plant growth promoting rhizobacteria (PGPR); Pseudomonas and Bacillus [119].
3. Production of antimicrobial compounds
Many growth-promoting bacteria are known to also produce antimicrobial compounds [120]. The main way that rhizobacterial biocontrol could improve plant growth is the same way that soil fumigation with toxic compounds can benefit agricultural production: killing root pathogens liberates plants from wasting resources fighting disease, allowing root systems to function at 100% capacity, delivering the maximal amount of nutrients and water to the shoot [21,121]. The genus Pseudomonas has been most extensively found to display these two traits [122,123], promoting plant growth while also producing antimicrobial compounds such as hydrogen cyanide (HCN) [124,125], 2,4-diacetylphloroglucinol (DAPG) [126], phenazines [127,128], cyclic lipopeptides [129,130], pyrrolnitrin [131], and pyoluteorin [132]. There are also reports of Pseudomonas producing volatile organic compounds (VOCs) such as 2-nonanone, acetic acids, 2-undecanone, m-cymene, dl-limonene, dimethyl disulfide and dimethyl trisulfide, which inhibit plant pathogens such as Sclerotinia sclerotiorum [133]. Application of antimicrobial producing Pseudomonas to crop plants has increased the growth rate up to 33% for bean [134]. Several recent reviews also cover the application of antimicrobial producing pseudomonads to improve crop yields [135,136,137]
Besides Pseudomonas, there are also other genera of rhizobacteria producing antibiotics that control root pathogens. Biocontrol strains from the genus Bacillus possess genes encoding antimicrobial compounds such as surfactins, phengicins, kurstakins, bacilicin, bacillomycins or microcin that can limit the growth of plant pathogens such as R. solani, Fusarium solani, Schizophyllum commune, Macrophomina phaseolina and Fusarium graminearum [138,139,140,141]. Other substances that are toxic to the plant pathogenic nematode Meloidogyne incognita, have also been identified in Bacillus megaterium and Bacillus thuringiensis, including benzene acetaldehyde, 2-nonanone, decanal, 2-undecanone, dimethyl disulfide [142,143]. Other genera of PGPR such as Enterobacter, Chryseobacterium, Burkholderia, and Klebsiella contain strains having the ability to synthesize surfactins, kurstakins, phengicins, and bacillomycins, which have been shown to successfully inhibit pathogens such as Phytophthora capsici, R. solani, and others [144,145,146,147]. There are many good recent reviews about biocontrol courtesy of antibiotic producing PGPRs [104,148,149,150].
Amongst the antimicrobial compounds mentioned above, DAPG is of special interest as it has a dual effect on plant phenotype as shown in Figure 1. DAPG is thought to derive its antimicrobial properties by degrading barrier structures and permeabilizing membranes/walls [151,152]. This antibiotic is produced by many growth promoting pseudomonads such as P. protegens, P. chlororaphis and P. brassicacearum, which have strong inhibitory effects on root pathogens such as R. solani [123,153]. While serving to inhibit microbial growth, DAPG can also be perceived by the plant as a growth promoting hormone and can increase carbon availability in the rhizosphere by stimulating root exudation [154]. This compound has been shown to modify Arabidopsis thaliana and maize root architecture [155]. In tomato and pea plants, DAPG produced by P. fluorescens increased the biomass of plant roots by increasing their length and weight, and also stimulated lateral root development [62,156]. Similarly, in maize plants, this compound improved plant vigor while enhancing phosphorus and magnesium uptake [157].
There are reports that the use of DAPG-producing bacteria enhanced the formation of pea root nodules by Rhizobium leguminosarum; this plant phenotype which was lost when inoculating with mutant bacteria lacking DAPG production [158]. It was theorized by the authors that this effect occurred because DAPG increases the concentration of organic acids in the rhizosphere, providing chemotaxtic cues to help Rhizobium discover and migrate to the roots [159]. Yet another beneficial effect of DAPG is its ability to stimulate the growth and activity of other beneficial rhizobacteria. When DAPG producing Pseudomonas were co-inoculated with plant growth promoting Azospirillum into wheat rhizospheres, plants showed an increase in root growth relative to plants inoculated with any single strain or the DAPG- mutant Pseudomonas [160]. The authors observed that DAPG was stimulating Azospirillum brasilense Sp245-Rif by increasing its expression of phytostimulatory genes such as ppdC, flgE and nirK, improving the rhizobacteria’s establishment and colonization of the roots and the rhizoplane.
Like DAPG, HCN can have a dual effect on plants. This compound can be produced by bacteria of the genera Chryseobacterium and Pseudomonas, controlling tomato root infections (reducing symptoms up to 80%) caused by soilborne pathogens such as F. oxysporum [80,161,162]. As an antimicrobial, HCN is a strong inhibitor of metalloenzymes including cytochrome c oxidase and is therefore toxic for all forms of aerobic life [163]. In plants that have evolved mechanisms to avoid cyanide intoxication, HCN can also act as a plant growth promoter, as was found in tobacco plants where HCN produced by P. fluorescens stimulated lateral root growth [164]. Specifically, HCN can enhance rhizosphere microbiome production of indole acetic acid, antibiotics, fluorescent insecticidal toxins, and 1-aminocyclopropane-1-carboxylate deaminase, all of which can result in plant growth promotion [165]. HCN also appears to be able to increase the bioavailability of important plant nutrients, for example by forming complexes with soil phosphorus and increasing its solubility but the exact mechanism is unknown [166].
PGPR production of the antimicrobial VOCs undecanone and heptanol also has the dual effect of fungal biocontrol and plant growth promotion. B. subtilis GB03 and B. amyloliquefaciens producing 2-heptanol and 2-undecanone, promote the growth of Arabidopsis thaliana [167,168] and watermelon [169] while also being the biocontrol agents in Kodiak Flowable Fungicide and Kodiak Concentrate Fungicide, suppressing root diseases caused by Fusarium spp. and R. solani on canola, mustard, rapeseed, and soybeans [170] or reducing fusarium wilt symptoms in watermelon [169].
4. Induced Systemic Resistance (ISR) and Systemic Acquired Resistance (SAR)
PGPR can participate in the activation of intrinsic plant defense responses by releasing elicitor molecules (elicitins) or producing microbe-associated molecular patterns (MAMPs) [171,172] that modulate the plant’s physiology, triggering increased resistance to pathogen invasion; these phenomena are known as induced systemic resistance (ISR) or systemic acquired resistance (SAR). Once MAMPs and elicitins are perceived by the plant, ISR or SAR involves long (or short)-distance signaling within the plant which means it can manifest systematically throughout the plant [150,173]. Plant signaling cascades involve compounds including jasmonic acid (JA), ethylene (ET) for ISR, and for SAR the most common is salicylic acid (SA) [174,175]. Plants that have undergone ISR and/or SAR are primed to respond quickly and more vigorously to pathogen invasion, having accumulated large quantities of transcription factors and signaling molecules [176,177]. ISR primed plants are also better able to resist abiotic stress, which could explain how they could show growth promotion even without pathogen pressure [178]. It has been observed that the production of compounds such as reactive oxygen species (ROS) in response to ISR or SAR plays a pivotal role in enhancing the stress tolerance of plants. This phenomenon aids in bolstering their resistance to factors like heat or elevated salt concentrations [179]. This effect was notably evident in rice plants inoculated with the Pseudomonas strain BaC1-38. This particular strain demonstrated the ability to activate pathogenesis-related proteins (PRs) alongside ROS. This study revealed that, besides inhibiting the pathogenic bacteria Xanthomonas campestris, it also conferred increased tolerance to salt concentration in the plants [180]. Furthermore, experiments with mutant plants exhibiting alterations in ROS regulation showed heightened sensitivity to a diverse range of abiotic stresses [181].
Crop plants induced to exhibit Induced Systemic Resistance through inoculation with Plant Growth-Promoting Rhizobacteria like Serratia marcescens, Bacillus pumilus, Bacillus velezensis or Pseudomonas azotoformans can demonstrate reductions of up to 50% of disease symptoms caused by pathogens like P. capsici, F. oxysporum in tomato [182], Xanthomonas axonopodis pv. vesicatoria in sweet pepper [183] or F. graminearum and F. moniliforme in wheat [184]. In another example, the commercial strain B. subtilis MBI600 inoculated onto tomato plants triggered SAR and greatly enhancing resistance to the pathogens R. solani, Pythium ultimum and F. oxysporum f.sp. radicis-lycopersici-Forl; reducing tomato disease symptoms from 38 to 47% [185,186]. The strain B. amyloliquefaciens SQR9 inhibits B. cinerea on A. thaliana roots by activating signaling cascades related to the salicylic acid which is typical in SAR [187].
Many of the plant signaling compounds involved in ISR and SAR signaling can also stimulate plant growth. These molecules include salicylic acid which is an important intermediary in systemic acquired resistance which has been shown to improve vegetative growth, physiological\biochemical characteristics, and to increase the number of flowers, fruits, and seeds in plants such as Sinningia speciosa, Foeniculum vulgare and Artemisia annua [188,189,190]. Like salicylic acid, there is some evidence that jasmonic acid (important in activating induced systemic resistance) can also act as a growth regulator, stimulating the development of buds and adventitious roots in potato cuttings, while in in vitro cultures of garlic, jasmonic acid significantly enhances the development of shoots and bulbs [191,192,193].
Microbial induction of ISR has been reported for many groups of bacteria, although one of the first was P. fluorescens, which had the ability to induce defense responses in carnation (Dianthus caryophyllus), making them more resistant to wilting caused by Fusarium sp. [194]. Another strain of this genus, called EA105, triggered ISR to Magnaporthe oryzae involving a jasmonate- and ethylene-dependent pathway – resistant plants reduced the pathogen's formation of appressoria [195]. Inoculation of rice with two different strains of P. fluorescens led to induction of systemic resistance against sheath blight fungus R. solani, increased the seedling vigor, promoted plant growth, and increased yields under both glasshouse and field conditions [196]. Similarly, application of either P. putida or P. aeruginosa (isolated from roots of healthy tomatoes in Pakistan) to tomatoes triggering ISR, significantly reducing fusarium wilt (reducing symptoms by >60%) while significantly increasing vegetative biomass and fruit yield [197].
Bacillus also has the ability to induce such responses through jasmonate- and ethylene-dependent pathways that control pathogens such as Phytophthora infestans, fungi, and nematodes [198,199]. In sugarcane plants, Bacillus strains have been reported to increase the levels of ISR-related metabolites, particularly phenylalanine ammonia lyase (PAL), polyphenol oxidase (PPO), and peroxidase (PO), which could stimulate the synthesis of phytoalexins and thus control pathogens such as Colletotrichum falcatum [200]. Examples of a species of Bacillus both triggering ISR and promoting plant growth or yield includes a B. pumilus strain in pepper plants that stimulates induction of plant defense against Xanthomonas axonopodis pv. vesicatoria and at the same time the plants had a higher weight and chlorophyll content [183]. Similarly, soil application of the commercial strain B. subtilis MBI600 to tomatoes increased shoot and root lengths (perhaps by secreting auxin) while inhibiting the soilborne pathogens R. solani, P. ultimum, and F. oxysporum f.sp. radicis-lycopersici-Forl; mechanisms involved were ISR triggered by signaling cascades depending on jasmonic acid and ethylene [186].
There are some examples of other genera of bacteria besides Pseudomonas and Bacillus that have the ability to induce ISR in plants. These include Serratia, Azospirillum and Enterobacter, which can activate plant genes related to plant defense response (i.e. PR1a, PR1b, PR2, PR5, and PDF1. 2) which can result in the increased production of reactive oxygen species [201,202,203] and the successful inhibition of pathogens such as Heterodera glycines, F. oxysporum or R. solani [203,204,205]. Serratia marcescens 90-166 for example can both trigger ISR and increase A. thaliana growth and seed yield – these responses involved jasmonate and salicylic acid-dependent signaling [63,206].
5. Quorum Sensing System (QS)
A plant’s defensive responses can be triggered by the detection of molecules being released by nearby microbes. Many of these MAMPs are associated with microbial virulence, such as flagellin or the signals bacteria use to coordinate their invasion once enough cells are available; the so called “quorum sensing” (QS). QS or autoinduction, is a cellular communication mechanism mediated by the release of quorum sensing molecules, which in gram-negative bacteria are often N-acyl homoserine lactones (AHL) and in gram positive bacteria are often small peptides [207]. These systems have been reported in a wide variety of bacteria associated with plants, where they may participate in symbiotic or pathogenic relationships between microorganisms and their hosts [208,209]. Some PGPRs are known to depend on AHL for QS and to coordinate activities beneficial for plant growth. In one example, a QS- mutant Serratia stopped producing nucleases, lipases, chitinases, proteases, prodigiosin and beta-lactam antibiotics that are involved in their biocontrol activity [210]. Strains of endophytic Pseudomonas isolated from Salicornia europaea possess QS dependent bacterial phytopathogen biocontrol capacity when inoculated into tomato plants [211]. Similarly, when QS was compromised in mutant P. chlororaphis, its ability to control the pathogen S. sclerotiorum in canola was also diminished [212]. Many other reports exist of QS negative mutants such as P. putida, Serratia liquefaciens and Serratia plymuthica losing their capacity to produce antibiotics and inhibit pathogens such as Pythium aphanidermatum, Alternaria alternata and Verticillium sp. [213,214,215].
As many pathogens also employ QS to coordinate infections, it should come as no surprise that plants have evolved a sensitivity to the molecules involved. QS molecules such as AHLs can trigger ISR resulting in upregulation of PR1 and chitinases [203,213]. These molecules can induce plant membrane hyperpolarization, facilitating the entry of ions such as K+ into the root [216]. Additionally, QS molecules have been shown to influence the nitrogen cycle, enhance root development, and influence its architecture [207,217].
Plants so primed by exposure to QS molecules can much better survive biotic and abiotic stress [218,219]. In one example AHL-producing Burkholderia graminis can increase tomato salt stress tolerance [220]. The same phenomenon has been observed in wheat and A. thaliana, where the presence of AHL enhanced salt stress tolerance involving ABA-dependent and independent signaling pathways [221]. In another report, wheat plants inoculated with acyl homoserine lactone (AHL)-producing Aeromonas strains, enjoyed improved salt tolerance, and significant improvement in root growth [222].There are also examples where AHLs such as N-hexanoyl-L-homoserine lactone (C6-HL) or N-3-oxo-hexanoyl homoserine lactone (3-oxo-C6- HL) can promote root elongation by increasing expression of plant growth and auxin/cytokinin signaling genes [223,224,225,226]. Tomato and A. thaliana plants inoculated with Pseudomonas and Serratia strains that produce AHLs, experienced enhanced seedling vigor and also displayed modifications in their root architecture, stimulating the development of lateral roots [227,228]. In a notable study, seed application of C6-HSL (a specific type of AHL) to two Ukrainian wheat varieties followed by field growth over two years, significantly increased vegetative biomass, increased grain yield by up to 50% and improved grain quality by up to 30% [207].
6. Antibiotic enzyme Production
A mechanism employed by some growth-promoting bacteria to antagonize competing microbes is the secretion of antibiotic enzymes. This trait has been documented in a wide variety of bacterial groups such as Bacillus, Pseudomonas, Serratia, Burkholderia, Azospirillum and Enterobacter. Enzymes such as β-1,3-glucanases, proteases, chitinases, and cellulases can inactivate pathogens by degrading their structural components (i.e. cell wall) [229]. Extracellular enzymes might also interfere with pathogen quorum sensing [230] or with pathogen camouflage and chemotaxis [231]. Reports of rhizobacterially produced enzymes controlling phytopathogens include F. oxysporum, Pyricularia oryzae, R. solani, Aspergillus niger, F. solani, Schizophyllum commune, Macrophomina phaseolina and F. graminearum [80,83,99,141,201,232]. Several recent reviews also cover this subject well [233,234,235].
Glucanases are one powerful tool that biocontrol strains might use to control pathogens, since 1,3-glucan (commonly called laminarin) is a critical structural component of all fungal cell walls [236]. Fungi exposed to substantial quantities of microbe-derived glucanase undergo growth inhibition, as their cell walls weaken due to the degradation of polymers. This degradation ultimately results in cellular death, brought about by the loss of cellular integrity [237,238]. Examples where PGPR control fungal pathogens by secretion of glucanases include R. solani [239] and S. sclerotiorum [239,240,241]. There are also examples of sugarcane derived PGPR Bacillus secreting glucanases into the rhizosphere to control R. solani, F. oxysporum and Fusarium moniliforme [242,243]. The involvement of microbe derived hydrolytic enzymes in plant pathogen control has been reviewed recently [229]. Although there is not much published on the capacity of microbial glucanases to directly influence plant growth, there is emerging evidence that these enzymes are involved in plant-microbe interactions [244].
Chitinases are another set of enzymes that are widely reported to be produced by PGPR; among them strains of Klebsiella, Serratia, and Enterobacter [245,246]. Chitinases are especially able to antagonize fungi whose cell walls contain chitin, which is essential for structural stability providing anchor points for other components at the cell surface. Bacteria secreting chitinases have been shown control fungal pathogens including Botrytis cinerea and R. solani, reducing the incidence of these related crop diseases in field trials by 76% and 84% respectively [88,247]. In addition to their antagonistic activity, growth-promoting bacteria with chitinolytic activity have also been associated with plant growth; for example, bacteria of the genera Pseudomonas and Bacillus with high chitinolytic activity increased germination in pigeon pea (Cajanus cajan) and in another example, a root inoculation with a chitinolytic strain of Streptomyces was associated with increased biomass in chickpea [245,248]. Cell free extracts of Chromobacterium sp. C61 contain both extracellular chitinase and a cyclic lipopeptide which contribute to its biocontrol activity against a number of root pathogens (e.g. R. solani, F. oxysporum and Colletotrichum gloeosporioides) in tomato plants – chitinase negative mutants had significantly lower biocontrol activity [249]. Transgenic expression in tomato plants of the endochitinase and chitobiosidase genes from Streptomyces albidoflavus significantly increased flowers and fruit yield [250]. The mode of plant growth promotion employed by PGPR secreted chitinases likely involves the creation of elicitins like chitosan from the breakdown of fungal cell walls, which are in turn perceived by the plant, triggering ISR [251]. In an example of this principal, externally applied chitooligosaccharides enhance the growth of Brassica rapa by up to 27% under greenhouse conditions [252]. The influence of microbial chitinases on plant growth has recently been reviewed [253,254].
Many pathogens coordinate their attacks by molecular communication termed quorum sensing – bacteria that secrete enzymes to degrade these molecules can thus serve as biocontrol agents. The best-known molecule involved in quorum sensing is called N-acyl homoserine lactone (AHL) – it is used by bacterial phytopathogens such as Pectobacterium carotovorum. Enzymes involved in degradation of AHL (also known as quorum-quenching enzymes) include lactonases and acylases [255] and there are reports of bacteria with these enzymes both controlling phytopathogens and increasing crop yields or plant growth [256,257]. Interestingly, one study describes that inoculation of chickpea with growth promoting Bacillus triggered the plant to increase its own production of quorum quenching enzymes, although some of these may have been bacterially produced enzymes as well [258]. While the mechanism of biocontrol is quite strait forward (disrupting pathogen communication and coordination) it is harder to imagine how these enzymes could influence plant growth. One possibility implies that since the quorum quenching system may also be related to the communication among growth-promoting bacteria, it is possible that it could influence the formation and balance of these communities, thus impacting the growth and yield of plants [259,260,261].
The study of fungal quorum sensing is relatively new, but it has been shown to involve pheromones, lipids, lactones, volatile organic compounds, tyrosol and farnesol [262]. To our knowledge, no studies have yet been done on PGPR enzymes interfering with fungal quorum signaling.
7. Conclusions
Rhizosphere microbiology has for many years attracted much scientific and agronomic attention, promising to help protect roots from disease while stimulating plants to grow bigger and healthier, resulting in higher yields [56,263]. Bacteria in a plant’s rhizosphere that can both protect the roots against disease while stimulating plant growth and increasing yield, could be the “superheroes” that every farmer hopes will dominate in their soils. The mechanisms used by such PGPRs to both control pathogens and stimulate plant growth include production of siderophores, antipathogen enzymes, antimicrobial compounds, quorum sensing molecules (especially N-acyl homoserine lactones), and various elicitins, MAMPs and priming agents. The most common examples of these dual wielding microbes hail from the genera Pseudomonas and Bacillus, however there are some other examples including Serratia, Azospirillum and Enterobacter. Commercial products exist for some of these dual action PGPR, such as Bacillus strains MBI600 or GB03 in the products Integral® and Kodiak® respectively [264,265] and Pseudomonas strain MA342 in the product Cedomon® [266]. It is easy to understand that effective biocontrol can aid the plant by reducing pathogen pressure, but how these mechanisms of biocontrol are also able to stimulate plant growth is still an area of research that needs to be developed.
References
- Shafique, H. A.; Sultana, V.; Ehteshamul-Haque, S.; Athar, M. Management of Soil-Borne Diseases of Organic Vegetables. J. Plant Prot. Res. 2016, 56, 221–230. [Google Scholar] [CrossRef]
- Ownley, B. H.; Benson, D. M. Soilborne Plant Pathogens. In Plant Pathology Concepts and Laboratory Exercises; Ownley, B. H., Trigiano, R. N., Eds.; CRC Press Taylor & Francis: Boca Raton, 2017; p. 400. [Google Scholar]
- Haichar, F. el Z.; Santaella, C.; Heulin, T.; Achouak, W. Root Exudates Mediated Interactions Belowground. Soil Biol. Biochem. 2014, 77, 69–80. [Google Scholar] [CrossRef]
- Steinauer, K.; Chatzinotas, A.; Eisenhauer, N. Root Exudate Cocktails: The Link between Plant Diversity and Soil Microorganisms? Ecol. Evol. 2016, 6, 7387–7396. [Google Scholar] [CrossRef]
- Haas, D.; Défago, G. Biological Control of Soil-Borne Pathogens by Fluorescent Pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Agrios, G. N. Plant Pathology, 5th ed.; Elsevier academic press: San Diego, 2005; Volume 9780080473. [Google Scholar] [CrossRef]
- Vidhyasekaran, P. Concise Encyclopedia of Plant Pathology, 1st ed.; Food Products Press: Binghamton, 2004. [Google Scholar]
- Halila, M. H.; Strange, R. N. Identification of the Causal Agent of Wilt of Chickpea in Tunisia as Fusarium Oxysporum f. Sp. Ciceri Race 0. Phytopathol. Mediterr. 1996, 35, 67–74. [Google Scholar]
- Jones, J. P.; Woltz, S. S.; Scott, J. W. Fusarium Crown Rot of Tomato: Some Factors Affecting Disease Development. In Proceedings of the Florida tomato institute SS-VEG-01; Stall, W.., Ed.; Veg Crops Dept: Miami, 1991; pp. 74–79. [Google Scholar]
- Hibar, K.; Daami-Remadi, M.; El Mahjoub, M. Induction of Resistance in Tomato Plants against Fusarium Oxysporum f. Sp. Radicis-Lycopersici by Trichoderma Spp. Tunis. J. Plant Prot. 2007, 2, 47–58. [Google Scholar]
- Jarvis, W. R.; Thorpe, H. J.; Meloche, R. B. Survey of Greenhouse Management Practices in Essex County, Ontario, in Relation to Fusarium Foot and Root Rot of Tomato. Plant Dis. 1983, 67, 38–40. [Google Scholar] [CrossRef]
- Hassan, H. A. Biology and Integrated Control of Tomato Wilt Caused by Fusarium Oxysporum Lycopersici: A Comprehensive Review under the Light of Recent Advancements. J. Bot. Res. 2020, 3. [Google Scholar] [CrossRef]
- Oerke, E. C. Crop Losses to Pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Ristaino, J. B. Tracking Historic Migrations of the Irish Potato Famine Pathogen, Phytophthora Infestans. Microbes Infect. 2002, 4, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Dohroo, N. Diseases of Ginger. In Ginger, the genus Zingiber; Ravindran, P.., Nirmal Babu, K., Eds.; CRC Press Taylor & Francis: Boca Raton, 2005; pp. 305–340. [Google Scholar]
- Bickel, J. T.; Koehler, A. M. Review of Pythium Species Causing Damping-Off in Corn. Plant Heal. Prog. 2021, 22, 1–7. [Google Scholar] [CrossRef]
- Annou, M. M.; Wailes, E. J.; Thomsen, M. R. A Dynamic Decision Model of Technology Adoption under Uncertainty: Case of Herbicide-Resistant Rice. J. Agric. Appl. Econ. 2005, 37, 161–172. [Google Scholar] [CrossRef]
- Jayaweera, D. P.; Ray, R. V. Yield Loss and Integrated Disease Control of Rhizoctonia Solani AG2-1 Using Seed Treatment and Sowing Rate of Oilseed Rape. Plant Dis. 2023, 107, 1159–1165. [Google Scholar] [CrossRef]
- Mukhtar, T.; Kayani, M. Z. Comparison of the Damaging Effects of Meloidogyne Incognita on a Resistant and Susceptible Cultivar of Cucumber. Bragantia 2020, 79, 83–93. [Google Scholar] [CrossRef]
- Subedi, S.; Thapa, B.; Shrestha, J. Root-Knot Nematode (Meloidogyne Incognita) and Its Management: A Review. J. Agric. Nat. Resour. 2020, 3, 21–31. [Google Scholar] [CrossRef]
- Raymaekers, K.; Ponet, L.; Holtappels, D.; Berckmans, B.; Cammue, B. P. A. Screening for Novel Biocontrol Agents Applicable in Plant Disease Management – A Review. Biol. Control 2020, 144. [Google Scholar] [CrossRef]
- Enebe, M. C.; Babalola, O. O. The Impact of Microbes in the Orchestration of Plants’ Resistance to Biotic Stress: A Disease Management Approach. Appl. Microbiol. Biotechnol. 2019, 103, 9–25. [Google Scholar] [CrossRef]
- Panth, M.; Hassler, S. C.; Baysal-Gurel, F. Methods for Management of Soilborne Diseases in Crop Production. Agric. 2020, 10. [Google Scholar] [CrossRef]
- Ueki, A.; Kaku, N.; Ueki, K. Role of Anaerobic Bacteria in Biological Soil Disinfestation for Elimination of Soil-Borne Plant Pathogens in Agriculture. Appl. Microbiol. Biotechnol. 2018, 102, 6309–6318. [Google Scholar] [CrossRef] [PubMed]
- Rafiq, C. M.; Tariq Mahmood, M.; Ahmad, M.; Ali, I.; Kaukab, S.; Shafiq, M.; Saleem, M.; Iqbal, U. Fusarium Wilt’s Pathogenic Studies and Disease Management: A Review. Genet. Mol. Res. 2020, 19. [Google Scholar]
- Katan, J. Diseases Caused by Soilborne Pathogens: Biology, Management and Challenges. J. Plant Pathol. 2017, 99, 305–315. [Google Scholar]
- Yuen, G. Y.; Schroth, M. N.; Weinhold, A. R.; Hancock, J. G. Effects of Soil Fumigation with Methyl Bromide and Chloropicrin on Root Health and Yield of Strawberry. Plant Dis. 1991, 75, 416–420. [Google Scholar] [CrossRef]
- White, P. J.; Brown, P. H. Plant Nutrition for Sustainable Development and Global Health. Ann. Bot. 2010, 105, 1073–1080. [Google Scholar] [CrossRef]
- Sugiyama, A.; Vivanco, J. M.; Jayanty, S. S.; Manter, D. K. Pyrosequencing Assessment of Soil Microbial Communities in Organic and Conventional Potato Farms. Plant Dis. 2010, 94, 1329–1335. [Google Scholar] [CrossRef]
- Qing, W.; Daqi, Z.; Wensheng, F.; Yuan, L. I.; Qiuxia, W.; Dongdong, Y. A. N.; Aocheng, C. A. O. Research Progress on the Effect of Soil Fumigation on Soil Nitrogen Cycles and Functional Microorganisms. Chinese J. Pestic. Sci. 2021, 23, 1063–1072. [Google Scholar]
- Larson, S.; Capel, P.; Majewski, M. Pesticides in Surface Waters-Distribution, Trends, and Governing Factors. In Series of Pesticides in Hydrologic System; Gillion, R., Ed.; Ann Arbor Press: Chelsea, Michigan, 1997. [Google Scholar]
- Buccini, J. The Development of a Global Treaty on Persistent Organic Pollutants (POPs). In Tha handbook of environmental Chemistry; Barceló, D., Kostianoy, A.., Eds.; Springer Science & Business Media, 2003; pp 13–30.
- Sadana, D.; Didwania, N. Integrated Disease Management of Bull’s Eye Pathogen Infecting Lycopersicon Esculentum (Tomato). J. Microbiol. Biotechnol. Food Sci. 2019, 9, 58–62. [Google Scholar] [CrossRef]
- Sukul, P. Enzymatic Activities and Microbial Biomass in Soil as Influenced by Metalaxyl Residues. Soil Biol. Biochem. 2006, 38, 320–326. [Google Scholar] [CrossRef]
- Wang, F.; Zhu, L.; Wang, X.; Wang, J.; Wang, J. Impact of Repeated Applications of Metalaxyl on Its Dissipation and Microbial Community in Soil. Water. Air. Soil Pollut. 2015, 226, 1–14. [Google Scholar] [CrossRef]
- Yen, J. H.; Chang, J. S.; Huang, P. J.; Wang, Y. S. Effects of Fungicides Triadimefon and Propiconazole on Soil Bacterial Communities. J. Environ. Sci. Heal. - Part B Pestic. Food Contam. Agric. Wastes 2009, 44, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Tang, F. F.; Zhou, W.; Cao, Z. Y.; Wang, D. D.; Liu, K. L.; Wu, X. W.; Yu, Y. L. Persistence of Repeated Triadimefon Application and Its Impact on Soil Microbial Functional Diversity. J. Environ. Sci. Heal. - Part B Pestic. Food Contam. Agric. Wastes 2012, 47, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huang, B.; Wang, Q.; Li, Y.; Fang, W.; Yan, D.; Guo, M.; Cao, A. Effect of Fumigation with Chloropicrin on Soil Bacterial Communities and Genes Encoding Key Enzymes Involved in Nitrogen Cycling. Environ. Pollut. 2017, 227, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Engel, A.; Rigby, M.; Burkholder, J. .; Fernandez, R. P.; Froidevaux, L.; Hall, B.; Hossaini, R.; Saito, T.; Vollmer, M. K.; Yao, B. Update on Ozone-Depleting Substances (ODSs) and Other Gases of Interest to the Montreal Protocol. In Scientific Assessment of Ozone Depletion; Liang, Q., Reimann, S., Eds.; World Meteorological Organization, 2019; pp 1–91.
- Agostini, M. G.; Roesler, I.; Bonetto, C.; Ronco, A. E.; Bilenca, D. Pesticides in the Real World: The Consequences of GMO-Based Intensive Agriculture on Native Amphibians. Biol. Conserv. 2020, 241, 108355. [Google Scholar] [CrossRef]
- Tong, L.; Nieh, J. C.; Tosi, S. Combined Nutritional Stress and a New Systemic Pesticide (Flupyradifurone, Sivanto®) Reduce Bee Survival, Food Consumption, Flight Success, and Thermoregulation. Chemosphere 2019, 237, 124408. [Google Scholar] [CrossRef] [PubMed]
- Straub, L.; Villamar-Bouza, L.; Bruckner, S.; Chantawannakul, P.; Kolari, E.; Maitip, J.; Vidondo, B.; Neumann, P.; Williams, G. R. Negative Effects of Neonicotinoids on Male Honeybee Survival, Behaviour and Physiology in the Field. J. Appl. Ecol. 2021, 58, 2515–2528. [Google Scholar] [CrossRef]
- Toledo-Hernández, E.; Peña-Chora, G.; Hernández-Velázquez, V. M.; Lormendez, C. C.; Toribio-Jiménez, J.; Romero-Ramírez, Y.; León-Rodríguez, R. The Stingless Bees (Hymenoptera: Apidae: Meliponini): A Review of the Current Threats to Their Survival. Apidologie 2022, 53. [Google Scholar] [CrossRef]
- Hamadamin, A. Y.; Hassan, K. I. Gas Chromatography–Mass Spectrometry Based Sensitive Analytical Approach to Detect and Quantify Non-Polar Pesticides Accumulated in the Fat Tissues of Domestic Animals. Saudi J. Biol. Sci. 2020, 27, 887–893. [Google Scholar] [CrossRef] [PubMed]
- Boudh, S.; Singh, J. S. Pesticide Contamination: Environmental Problems and Remediation Strategies. In Emerging and Eco-friendly Approaches for Waste Management; Bharagava, R. N., Chowdhary, P., Eds.; Springer, 2018; pp 245–269. [CrossRef]
- Rajiv, K. S.; George, H.; Upendra, P.; Chandrajeet, K. Embarking on Second Green Revolution by Vermiculture for Production of Chemical Free Organic Foods, Protection of Crops and Farm Soils and Elimination of Deadly Agrochemicals from Earth: Meeting the Challenges of Food Security of 21st Century by Earthworm. In Agricultural Research Updates; Gorawala, P., Mandhatri, S., Eds.; Nova Science Publishers Inc: New York, 2016; pp. 1–49. [Google Scholar]
- U.S. Environmental Proteccion Agency. Waste minimization priority chemicals and chemical fact sheets website. Priority Chemicals. Available online: https://archive.epa.gov/epawaste/hazard/wastemin/web/html/priority.html (accessed on 18 June 2020).
- Wu, S.; Jin, C.; Wang, Y.; Fu, Z.; Jin, Y. Exposure to the Fungicide Propamocarb Causes Gut Microbiota Dysbiosis and Metabolic Disorder in Mice. Environ. Pollut. 2018, 237, 775–783. [Google Scholar] [CrossRef]
- Sturz, A. V; Christie, B. R.; Nowak, J. Bacterial Endophytes: Potential Role in Developing Sustainable Systems of Crop Production. CRC. Crit. Rev. Plant Sci. 2000, 19, 1–30. [Google Scholar] [CrossRef]
- Bouzoumita, A.; Metoui, M.; Jemni, M.; Kabaeir, N.; Belhouchette, K.; Ferchichi, A. The Efficacy of Various Bacterial Organisms for Biocontrol of Fusarium Root Rot of Olive in Tunisia. Polish J. Environ. Stud. 2020, 29, 11–16. [Google Scholar] [CrossRef]
- Sajeena, A.; Nair, D. S.; Sreepavan, K. Non-Pathogenic Fusarium Oxysporum as a Biocontrol Agent. Indian Phytopathol. 2020, 73, 177–183. [Google Scholar] [CrossRef]
- Barriuso, J.; Ramos Solano, B.; Lucas, J. A.; Lobo, A. P.; García-Villaraco, A.; Gutiérrez Mañero, F. J. Ecology, Genetic Diversity and Screening Strategies of Plant Growth Promoting Rhizobacteria (PGPR). In Plant-Bacteria Interactions: Strategies and Techniques to Promote Plant Growth; Ahmad, I., Pitchel, J., Hayat, S., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA, 2008; pp 1–17. [CrossRef]
- Bhattacharjee, R. B.; Singh, A.; Mukhopadhyay, S. N. Use of Nitrogen-Fixing Bacteria as Biofertiliser for Non-Legumes: Prospects and Challenges. Appl. Microbiol. Biotechnol. 2008, 80, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Yazdani, M.; Bahmanyar, M. A.; Pirdashti, H.; Esmaili, M. A. Effect of Phosphate Solubilization Microorganisms (PSM) and Plant Growth Promoting Rhizobacteria (PGPR) on Yield and Yield Components of Corn (Zea Mays L.). World Acad. Sci. Eng. Technol. 2009, 49, 90–92. [Google Scholar]
- Ngalimat, M. S.; Hata, E. M.; Zulperi, D.; Ismail, S. I.; Ismail, M. R.; Zainudin, N. A. I. M.; Saidi, N. B.; Yusof, M. T. Plant Growth-Promoting Bacteria as an Emerging Tool to Manage Bacterial Rice Pathogens. Microorganisms 2021, 9, 1–23. [Google Scholar] [CrossRef]
- Basu, A.; Prasad, P.; Das, S. N.; Kalam, S.; Sayyed, R. Z.; Reddy, M. S.; Enshasy, H. El. Plant Growth Promoting Rhizobacteria (Pgpr) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 13, 1–20. [Google Scholar] [CrossRef]
- ALKahtani, M. D. F.; Fouda, A.; Attia, K. A.; Al-Otaibi, F.; Eid, A. M.; El-Din Ewais, E.; Hijri, M.; St-Arnaud, M.; El-Din Hassan, S.; Khan, N.; Hafez, Y. M.; Abdelaal, K. A. A. Isolation and Characterization of Plant Growth Promoting Endophytic Bacteria from Desert Plants and Their Application as Bioinoculants for Sustainable Agriculture. Agronomy 2020, 10, 1–18. [Google Scholar] [CrossRef]
- Kloepper, J. W.; Lifshitz, R.; Zablotovicz, R. M. Free-Living Bacteria Inocula for Enhancing Crop Productivity. Trends Biotechnol. 1989, 7, 39–44. [Google Scholar] [CrossRef]
- Berg, G.; Smalla, K. Plant Species and Soil Type Cooperatively Shape the Structure and Function of Microbial Communities in the Rhizosphere. FEMS Microbiol. Ecol. 2009, 68, 1–13. [Google Scholar] [CrossRef]
- Jiao, X.; Takishita, Y.; Zhou, G.; Smith, D. L. Plant Associated Rhizobacteria for Biocontrol and Plant Growth Enhancement. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef]
- Katiyar, V.; Goel, R. Siderophore Mediated Plant Growth Promotion at Low Temperature by Mutant of Fluorescent Pseudomonad. Plant Growth Regul. 2004, 42, 239–244. [Google Scholar] [CrossRef]
- Brazelton, J. N.; Pfeufer, E. E.; Sweat, T. A.; McSpadden Gardener, B. B.; Coenen, C. 2,4-Diacetylphloroglucinol Alters Plant Root Development. Mol. Plant-Microbe Interact. 2008, 21, 1349–1358. [Google Scholar] [CrossRef] [PubMed]
- Shi, C. L.; Park, H. B.; Lee, J. S.; Ryu, S.; Ryu, C. M. Inhibition of Primary Roots and Stimulation of Lateral Root Development in Arabidopsis Thaliana by the Rhizobacterium Serratia Marcescens 90-166 Is through Both Auxin-Dependent and -Independent Signaling Pathways. Mol. Cells 2010, 29, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Liliane, T. N.; Charles, M. S. Factors Affecting Yield of Crops. In Agronomy climate change and food security; Amanullah, Ed.; InTechOpen: London, 2020; pp. 9–25. [Google Scholar]
- Ficke, A.; Cowger, C.; Bergstrom, G.; Brodal, G. Understanding Yield Loss and Pathogen Biology to Improve Disease Management: Septoria Nodorum Blotch - A Case Study in Wheat. Plant Dis. 2018, 102, 696–707. [Google Scholar] [CrossRef] [PubMed]
- Brittenham, G. . New Advances in Iron Metabolism, Iron Deficiency, and Iron Overload. Curr. Opin. Hematol. 1994, 1, 101–106. [Google Scholar] [PubMed]
- Pierre, J. L.; Fontecave, M.; Crichton, R. R. Chemistry for an Essential Biological Process: The Reduction of Ferric Iron. BioMetals 2002, 15, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Miller, G. W.; Huang, I. J.; Welkie, G. W.; Pushnik, J. C. Function of Iron in Plants with Special Emphasis on Chloroplasts and Photosynthetic Activity. In Iron Nutrition in Soils and Plants; Abadia, J., Ed.; Kluwer Academic Publishers: Dordrecht, 1995; pp. 19–28. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, D.; Sun, W.; Wang, T. The Adaptive Mechanism of Plants to Iron Deficiency via Iron Uptake, Transport, and Homeostasis. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef]
- Tsai, H. H.; Schmidt, W. One Way. Or Another? Iron Uptake in Plants. New Phytol. 2017, 214, 500–505. [Google Scholar] [CrossRef]
- Ansari, R.A.; Mahmood, I.; Rizvi, R.; Sumbul, A.; Safiuddin. Augmentation of Soil Health and Crop Productivity. In Probiotics in Agroecosystem; Kumar, V., Kumar, M., Sharma, S., Prasad, R., Eds.; Springer: Singapore, 2017; pp. 291–312. [Google Scholar] [CrossRef]
- Timofeeva, A. M.; Galyamova, M. R.; Sedykh, S. E. Bacterial Siderophores: Classification, Biosynthesis, Perspectives of Use in Agriculture. Plants 2022, 11. [Google Scholar] [CrossRef]
- Nithyapriya, S.; Lalitha, S.; Sayyed, R. Z.; Reddy, M. S.; Dailin, D. J.; El Enshasy, H. A.; Luh Suriani, N.; Herlambang, S. Production, Purification, and Characterization of Bacillibactin Siderophore of Bacillus Subtilis and Its Application for Improvement in Plant Growth and Oil Content in Sesame. Sustain. 2021, 13. [Google Scholar] [CrossRef]
- Yu, S.; Teng, C.; Bai, X.; Liang, J.; Song, T.; Dong, L.; Jin, Y.; Qu, J. Optimization of Siderophore Production by Bacillus Sp. PZ-1 and Its Potential Enhancement of Phytoextration of PB from Soil. J. Microbiol. Biotechnol. 2017, 27, 1500. [Google Scholar] [CrossRef]
- Rizzi, A.; Roy, S.; Bellenger, J. P.; Beauregard, P. B. Iron Homeostasis in Bacillus Subtilis Requires Siderophore Production and Biofilm Formation. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Doshi, H. .; Thakur, M.. Bacillus Spp.: A Prolific Siderophore Producer. In Bacilli and Agrobiotechnology; Rahman, M., Pandey, P., Jha, C., Aeron, A., Eds.; Springer, 2016; pp 309–323. [CrossRef]
- Aznar, A.; Dellagi, A. New Insights into the Role of Siderophores as Triggers of Plant Immunity: What Can We Learn from Animals? J. Exp. Bot. 2015, 66, 3001–3010. [Google Scholar] [CrossRef]
- Sadeghi, A.; Karimi, E.; Dahaji, P. A.; Javid, M. G.; Dalvand, Y.; Askari, H. Plant Growth Promoting Activity of an Auxin and Siderophore Producing Isolate of Streptomyces under Saline Soil Conditions. World J. Microbiol. Biotechnol. 2012, 28, 1503–1509. [Google Scholar] [CrossRef]
- Verma, V. C.; Singh, S. K.; Prakash, S. Bio-Control and Plant Growth Promotion Potential of Siderophore Producing Endophytic Streptomyces from Azadirachta Indica A. Juss. J. Basic Microbiol. 2011, 51, 550–556. [Google Scholar] [CrossRef]
- Paramanandham, P.; Rajkumari, J.; Pattnaik, S.; Busi, S. Biocontrol Potential against Fusarium Oxysporum f. Sp. Lycopersici and Alternaria Solani and Tomato Plant Growth Due to Plant Growth–Promoting Rhizobacteria. Int. J. Veg. Sci. 2017, 23, 294–303. [Google Scholar] [CrossRef]
- Siddiqui, Z. A.; Baghel, G.; Akhtar, M. S. Biocontrol of Meloidogyne Javanica by Rhizobium and Plant Growth-Promoting Rhizobacteria on Lentil. World J. Microbiol. Biotechnol. 2007, 23, 435–441. [Google Scholar] [CrossRef]
- Dimopoulou, A.; Theologidis, I.; Benaki, D.; Koukounia, M.; Zervakou, A.; Tzima, A.; Diallinas, G.; Hatzinikolaou, D. G.; Skandalis, N. Direct Antibiotic Activity of Bacillibactin Broadens the Biocontrol Range of Bacillus Amyloliquefaciens MBI600. mSphere 2021, 6, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Rais, A.; Shakeel, M.; Hafeez, F. Y.; Hassan, M. N. Plant Growth Promoting Rhizobacteria Suppress Blast Disease Caused by Pyricularia Oryzae and Increase Grain Yield of Rice. BioControl 2016, 61, 769–780. [Google Scholar] [CrossRef]
- Ait-Kaki, A.; Kacem-Chaouche, N.; Ongena, M.; Kara-Ali, M.; Dehimat, L.; Kahlat, K.; Thonart, P. In Vitro and in Vivo Characterization of Plant Growth Promoting Bacillus Strains Isolated from Extreme Environments of Eastern Algeria. Appl. Biochem. Biotechnol. 2014, 172, 1735–1746. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M. N.; Osborn, A. M.; Hafeez, F. Y. Molecular and Biochemical Characterization of Surfactin Producing Bacillus Species Antagonistic to Colletotrichum Falcatum Went Causing Sugarcane Red Rot. African J. Microbiol. Res. 2010, 4, 2137–2142. [Google Scholar]
- Caulier, S.; Gillis, A.; Colau, G.; Licciardi, F.; Liépin, M.; Desoignies, N.; Modrie, P.; Legrève, A.; Mahillon, J.; Bragard, C. Versatile Antagonistic Activities of Soil-Borne Bacillus Spp. and Pseudomonas Spp. against Phytophthora Infestans and Other Potato Pathogens. Front. Microbiol. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Li, S.; Li, S. Y.; Zhang, H.; Jiang, M. The Siderophore-Producing Bacterium, Bacillus Siamensis Gxun-6, Has an Antifungal Activity against Fusarium Oxysporum and Promotes the Growth of Banana. Egypt. J. Biol. Pest Control 2022, 32. [Google Scholar] [CrossRef]
- Kamensky, M.; Ovadis, M.; Chet, I.; Chernin, L. Soil-Borne Strain IC14 of Serratia Plymuthica with Multiple Mechanisms of Antifungal Activity Provides Biocontrol of Botrytis Cinerea and Sclerotinia Sclerotiorum Diseases. Soil Biol. Biochem. 2003, 35, 323–331. [Google Scholar] [CrossRef]
- Khoa, N. Đ.; Giàu, N. Đ. N.; Tuấn, T. Q. Effects of Serratia Nematodiphila CT-78 on Rice Bacterial Leaf Blight Caused by Xanthomonas Oryzae Pv. Oryzae. Biol. Control 2016, 103, 1–10. [Google Scholar] [CrossRef]
- Tortora, M. L.; Díaz-Ricci, J. C.; Pedraza, R. O. Azospirillum Brasilense Siderophores with Antifungal Activity against Colletotrichum Acutatum. Arch. Microbiol. 2011, 193, 275–286. [Google Scholar] [CrossRef]
- Bendaha, M. E. A.; Belaouni, H. A. Tomato Growth and Resistance Promotion by Enterobacter Hormaechei Subsp. Steigerwaltii EB8D. Arch. Phytopathol. Plant Prot. 2019, 52, 318–332. [Google Scholar] [CrossRef]
- Naureen, Z.; Hafeez, F. Y.; Hussain, J.; Harrasi, A. Al; Bouqellah, N.; Roberts, M. R. Suppression of Incidence of Rhizoctonia Solani in Rice By Siderophore Producing Rhizobacterial Strains Based on Competition for Iron. Eur. Sci. J. 2015, 11, 186–207. [Google Scholar]
- Gechemba, O. R.; Julius, M.; Makonde, H. M.; Bubambula, N. L. M.; Matiru, V. N. Potentially Beneficial Rhizobacteria Associated with Banana Plants in Juja Kenya. Res. Rev. J. Microbiol. Biotechnol. 2016, 5, 19–25. [Google Scholar]
- Ahemad, M.; Khan, M. S. Effects of Insecticides on Plant-Growth-Promoting Activities of Phosphate Solubilizing Rhizobacterium Klebsiella Sp. Strain PS19. Pestic. Biochem. Physiol. 2011, 100, 51–56. [Google Scholar] [CrossRef]
- El_Komy, M. H.; Hassouna, M. G.; Abou-Taleb, E. M.; Al-Sarar, A. S.; Abobakr, Y. A Mixture of Azotobacter, Azospirillum, and Klebsiella Strains Improves Root-Rot Disease Complex Management and Promotes Growth in Sunflowers in Calcareous Soil. Eur. J. Plant Pathol. 2020, 156, 713–726. [Google Scholar] [CrossRef]
- Liu, X.; Jia, J.; Atkinson, S.; Cámara, M.; Gao, K.; Li, H.; Cao, J. Biocontrol Potential of an Endophytic Serratia Sp. G3 and Its Mode of Action. World J. Microbiol. Biotechnol. 2010, 26, 1465–1471. [Google Scholar] [CrossRef]
- Thomas, M. S. Iron Acquisition Mechanisms of the Burkholderia Cepacia Complex. BioMetals 2007, 20, 431–452. [Google Scholar] [CrossRef] [PubMed]
- Araújo, W. L.; Creason, A. L.; Mano, E. T.; Camargo-Neves, A. A.; Minami, S. N.; Chang, J. H.; Loper, J. E. Genome Sequencing and Transposon Mutagenesis of Burkholderia Seminalis TC3.4.2R3 Identify Genes Contributing to Suppression of Orchid Necrosis Caused by B. Gladioli. Mol. Plant-Microbe Interact. 2016, 29, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, V. C.; Barbosa, L. de O.; Andrade, J. P.; Soares, A. C. F.; de Souza, J. T.; Marbach, P. A. S. Burkholderia Isolates from a Sand Dune Leaf Litter Display Biocontrol Activity against the Bole Rot Disease of Agave Sisalana. Biol. Control 2017, 112, 41–48. [Google Scholar] [CrossRef]
- Pellegrini, M.; Pagnani, G.; Bernardi, M.; Mattedi, A.; Spera, D. M.; Del Gallo, M. Cell-Free Supernatants of Plant Growth-Promoting Bacteria: A Review of Their Use as Biostimulant and Microbial Biocontrol Agents in Sustainable Agriculture. Sustainability 2020, 12, 1–22. [Google Scholar] [CrossRef]
- Sayyed, R. Z.; Chincholkar, S. B. Siderophore-Producing Alcaligenes Feacalis Exhibited More Biocontrol Potential Vis-à-Vis Chemical Fungicide. Curr. Microbiol. 2009, 58, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Dimkpa, C. Microbial Siderophores: Production, Detection and Application in Agriculture and Environment. Endocytobiosis Cell Res. 2016, 27, 7–16. [Google Scholar]
- Soares, E. V. Perspective on the Biotechnological Production of Bacterial Siderophores and Their Use. Appl. Microbiol. Biotechnol. 2022, 106, 3985–4004. [Google Scholar] [CrossRef]
- Tariq, M.; Noman, M.; Ahmed, T.; Hameed, A.; Manzoor, N.; Zafar, M. Antagonistic Features Displayed by Plant Growth Promoting Rhizobacteria (PGPR): A Review. J. Plant Sci. Phytopathol. 2017, 1, 038–043. [Google Scholar] [CrossRef]
- Prasad, K. Utilization of Siderophore Producing Plant Growth Promoting Rhizobacteria to Improve Crucial Nourishment and Management of Phytopathogen in Cash Crops for Sustainable Development. Am. J. Sci. Eng. Res. 2022, 5, 15–23. [Google Scholar]
- Shafi, J.; Tian, H.; Ji, M. Bacillus Species as Versatile Weapons for Plant Pathogens: A Review. Biotechnol. Biotechnol. Equip. 2017, 31, 446–459. [Google Scholar] [CrossRef]
- Ghazy, N.; El-Nahrawy, S. Siderophore Production by Bacillus Subtilis MF497446 and Pseudomonas Koreensis MG209738 and Their Efficacy in Controlling Cephalosporium Maydis in Maize Plant. Arch. Microbiol. 2021, 203, 1195–1209. [Google Scholar] [CrossRef]
- Sasirekha, B.; Srividya, S. Siderophore Production by Pseudomonas Aeruginosa FP6, a Biocontrol Strain for Rhizoctonia Solani and Colletotrichum Gloeosporioides Causing Diseases in Chilli. Agric. Nat. Resour. 2016, 50, 250–256. [Google Scholar] [CrossRef]
- Dimkpa, C. O.; Merten, D.; Svatoš, A.; Büchel, G.; Kothe, E. Metal-Induced Oxidative Stress Impacting Plant Growth in Contaminated Soil Is Alleviated by Microbial Siderophores. Soil Biol. Biochem. 2009, 41, 154–162. [Google Scholar] [CrossRef]
- Becker, J. O.; Cook, R. J. Role of Siderophores in Suppression of Pythium Species and Production of Increased-Growth Response of Wheat by Fluorescent Pseudomonads. Phytopathology, 1988, 78, 778–782. [Google Scholar] [CrossRef]
- Sharma, A.; Johri, B. N.; Sharma, A. K.; Glick, B. R. Plant Growth-Promoting Bacterium Pseudomonas Sp. Strain GRP3 Influences Iron Acquisition in Mung Bean (Vigna Radiata L. Wilzeck). Soil Biol. Biochem. 2003, 35, 887–894. [Google Scholar] [CrossRef]
- Syed, A.; Elgorban, A. M.; Bahkali, A. H.; Eswaramoorthy, R.; Iqbal, R. K.; Danish, S. Metal-Tolerant and Siderophore Producing Pseudomonas Fluorescence and Trichoderma Spp. Improved the Growth, Biochemical Features and Yield Attributes of Chickpea by Lowering Cd Uptake. Sci. Rep. 2023, 13, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Kloepper, J. W.; Leong, J.; Teintze, M.; Schroth, M. N. Enhanced Plant Growth by Siderophores Produced by Plant Growth-Promoting Rhizobacteria. Nature 1980, 286, 885–886. [Google Scholar] [CrossRef]
- Rungin, S.; Indananda, C.; Suttiviriya, P.; Kruasuwan, W.; Jaemsaeng, R.; Thamchaipenet, A. Plant Growth Enhancing Effects by a Siderophore-Producing Endophytic Streptomycete Isolated from a Thai Jasmine Rice Plant (Oryza Sativa L. Cv. KDML105). Antonie van Leeuwenhoek, Int. J. Gen. Mol. Microbiol. 2012, 102, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Ai, C.; Xin, L.; Zhou, G. The Siderophore-Producing Bacterium, Bacillus Subtilis CAS15, Has a Biocontrol Effect on Fusarium Wilt and Promotes the Growth of Pepper. Eur. J. Soil Biol. 2011, 47, 138–145. [Google Scholar] [CrossRef]
- Jikare, A. M.; Chavan, M. D. SiderophorepProduced by Bacillus Shackletonii. Gn-09 and Showed Its Plant Growth Promoting Activity. Int. J. Pharm. Biol. Sci. 2013, 3, 198–202. [Google Scholar]
- Miljaković, D.; Marinković, J.; Balešević-Tubić, S. The Significance of Bacillus Spp. In Disease Suppression and Growth Promotion of Field and Vegetable Crops. Microorganisms 2020, 8, 1–19. [Google Scholar] [CrossRef]
- Zhang, S.; Deng, Z.; Borham, A.; Ma, Y.; Wang, Y.; Hu, J.; Wang, J.; Bohu, T. Significance of Soil Siderophore-Producing Bacteria in Evaluation and Elevation of Crop Yield. Horticulturae 2023, 9, 1–10. [Google Scholar] [CrossRef]
- Santoyo, G.; del Orozco-Mosqueda, M. C.; Govindappa, M. Mechanisms of Biocontrol and Plant Growth-Promoting Activity in Soil Bacterial Species of Bacillus and Pseudomonas: A Review. Biocontrol Sci. Technol. 2012, 22, 855–872. [Google Scholar] [CrossRef]
- Ramakrishna, W.; Yadav, R.; Li, K. Plant Growth Promoting Bacteria in Agriculture: Two Sides of a Coin. Appl. Soil Ecol. 2019, 138, 10–18. [Google Scholar] [CrossRef]
- Niu, B.; Wang, W.; Yuan, Z.; Sederoff, R. R.; Sederoff, H.; Chiang, V. L.; Borriss, R. Microbial Interactions Within Multiple-Strain Biological Control Agents Impact Soil-Borne Plant Disease. Front. Microbiol. 2020, 11, 1–16. [Google Scholar] [CrossRef]
- Silby, M. W.; Winstanley, C.; Godfrey, S. A. C.; Levy, S. B.; Jackson, R. W. Pseudomonas Genomes: Diverse and Adaptable. FEMS Microbiol. Rev. 2011, 35, 652–680. [Google Scholar] [CrossRef]
- Wang, X.; Mavrodi, D. V.; Ke, L.; Mavrodi, O. V.; Yang, M.; Thomashow, L. S.; Zheng, N.; Weller, D. M.; Zhang, J. Biocontrol and Plant Growth-Promoting Activity of Rhizobacteria from Chinese Fields with Contaminated Soils. Microb. Biotechnol. 2015, 8, 404–418. [Google Scholar] [CrossRef] [PubMed]
- Nandi, M.; Selin, C.; Brawerman, G.; Fernando, W. G. D.; de Kievit, T. Hydrogen Cyanide, Which Contributes to Pseudomonas Chlororaphis Strain PA23 Biocontrol, Is Upregulated in the Presence of Glycine. Biol. Control 2017, 108, 47–54. [Google Scholar] [CrossRef]
- Strano, C. P.; Bella, P.; Licciardello, G.; Caruso, A.; Catara, V. Role of Secondary Metabolites in the Biocontrol Activity of Pseudomonas Corrugata and Pseudomonas Mediterranea. Eur. J. Plant Pathol. 2017, 149, 103–115. [Google Scholar] [CrossRef]
- Biessy, A.; Filion, M. Phloroglucinol Derivatives in Plant-Beneficial Pseudomonas Spp.: Biosynthesis, Regulation, and Functions. Metabolites 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Biessy, A.; Filion, M. Phenazines in Plant-Beneficial Pseudomonas Spp.: Biosynthesis, Regulation, Function and Genomics. Environ. Microbiol. 2018, 20, 3905–3917. [Google Scholar] [CrossRef]
- Serafim, B.; Bernardino, A. R.; Freitas, F.; Torres, C. A. V. Recent Developments in the Biological Activities, Bioproduction, and Applications of Pseudomonas Spp. Phenazines. Molecules 2023, 28. [Google Scholar] [CrossRef] [PubMed]
- Oni, F. E.; Geudens, N.; Onyeka, J. T.; Olorunleke, O. F.; Salami, A. E.; Omoboye, O. O.; Arias, A. A.; Adiobo, A.; De Neve, S.; Ongena, M.; Martins, J. C.; Höfte, M. Cyclic Lipopeptide-Producing Pseudomonas Koreensis Group Strains Dominate the Cocoyam Rhizosphere of a Pythium Root Rot Suppressive Soil Contrasting with P. Putida Prominence in Conducive Soils. Environ. Microbiol. 2020, 22, 5137–5155. [Google Scholar] [CrossRef] [PubMed]
- Geudens, N.; Martins, J. C. Cyclic Lipodepsipeptides from Pseudomonas Spp. - Biological Swiss-Army Knives. Front. Microbiol. 2018, 9, 1–18. [Google Scholar] [CrossRef]
- Bajpai, A.; Singh, B.; Joshi, S.; Johri, B. N. Production and Characterization of an Antifungal Compound from Pseudomonas Protegens Strain W45. Proc. Natl. Acad. Sci. India Sect. B - Biol. Sci. 2018, 88, 1081–1089. [Google Scholar] [CrossRef]
- Hansen, M. L.; Wibowo, M.; Jarmusch, S. A.; Larsen, T. O.; Jelsbak, L. Sequential Interspecies Interactions Affect Production of Antimicrobial Secondary Metabolites in Pseudomonas Protegens DTU9.1. ISME J. 2022, 16, 2680–2690. [Google Scholar] [CrossRef] [PubMed]
- Giorgio, A.; De Stradis, A.; Lo Cantore, P.; Iacobellis, N. S. Biocide Effects of Volatile Organic Compounds Produced by Potential Biocontrol Rhizobacteria on Sclerotinia Sclerotiorum. Front. Microbiol. 2015, 6, 1056. [Google Scholar] [CrossRef]
- Kalantari, S.; Marefat, A.; Naseri, B.; Hemmati, R. Improvement of Bean Yield and Fusarium Root Rot Biocontrol Using Mixtures of Bacillus, Pseudomonas and Rhizobium. Trop. Plant Pathol. 2018, 43, 499–505. [Google Scholar] [CrossRef]
- Mehmood, N.; Saeed, M.; Zafarullah, S.; Hyder, S.; Rizvi, Z. F.; Gondal, A. S.; Jamil, N.; Iqbal, R.; Ali, B.; Ercisli, S.; Kupe, M. Multifaceted Impacts of Plant-Beneficial Pseudomonas Spp. in Managing Various Plant Diseases and Crop Yield Improvement. ACS Omega 2023, 8, 22296–22315. [Google Scholar] [CrossRef]
- Guzmán-Guzmán, P.; Santoyo, G. Action Mechanisms, Biodiversity, and Omics Approaches in Biocontrol and Plant Growth-Promoting Pseudomonas: An Updated Review. Biocontrol Sci. Technol. 2022, 32, 527–550. [Google Scholar] [CrossRef]
- Raio, A.; Puopolo, G. Pseudomonas Chlororaphis Metabolites as Biocontrol Promoters of Plant Health and Improved Crop Yield. World J. Microbiol. Biotechnol. 2021, 37, 1–8. [Google Scholar] [CrossRef]
- Liu, K.; Newman, M.; McInroy, J. A.; Hu, C. H.; Kloepper, J. W. Selection and Assessment of Plant Growth-Promoting Rhizobacteria for Biological Control of Multiple Plant Diseases. Phytopathology 2017, 107, 928–936. [Google Scholar] [CrossRef]
- Vitullo, D.; Di Pietro, A.; Romano, A.; Lanzotti, V.; Lima, G. Role of New Bacterial Surfactins in the Antifungal Interaction between Bacillus Amyloliquefaciens and Fusarium Oxysporum. Plant Pathol. 2012, 61, 689–699. [Google Scholar] [CrossRef]
- Béchet, M.; Caradec, T.; Hussein, W.; Abderrahmani, A.; Chollet, M.; Leclére, V.; Dubois, T.; Lereclus, D.; Pupin, M.; Jacques, P. Structure, Biosynthesis, and Properties of Kurstakins, Nonribosomal Lipopeptides from Bacillus Spp. Appl. Microbiol. Biotechnol. 2012, 95, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Chenniappan, C.; Narayanasamy, M.; Daniel, G. M.; Ramaraj, G. B.; Ponnusamy, P.; Sekar, J.; Vaiyapuri Ramalingam, P. Biocontrol Efficiency of Native Plant Growth Promoting Rhizobacteria against Rhizome Rot Disease of Turmeric. Biol. Control 2019, 129, 55–64. [Google Scholar] [CrossRef]
- Huang, Y.; Xu, C. K.; Ma, L.; Zhang, K. Q.; Duan, C. Q.; Mo, M. H. Characterisation of Volatiles Produced from Bacillus Megaterium YFM3.25 and Their Nematicidal Activity against Meloidogyne Incognita. Eur. J. Plant Pathol. 2010, 126, 417–422. [Google Scholar] [CrossRef]
- Peng, D.; Chai, L.; Wang, F.; Zhang, F.; Ruan, L.; Sun, M. Synergistic Activity between Bacillus Thuringiensis Cry6Aa and Cry55Aa Toxins against Meloidogyne Incognita. Microb. Biotechnol. 2011, 4, 794–798. [Google Scholar] [CrossRef] [PubMed]
- Vial, L.; Groleau, M. C.; Dekimpe, V.; Deziel, E. Burkholderia Diversity and Versatility: An Inventory of the Extracellular Products. J. Microbiol. Biotechnol. 2007, 17, 1407–1428. [Google Scholar] [CrossRef]
- da Silva Araújo, D. F.; Araújo, W. L.; Eberlin, M. N. Potential of Burkholderia Seminalis TC3.4.2R3 as Biocontrol Agent Against Fusarium Oxysporum Evaluated by Mass Spectrometry Imaging. J. Am. Soc. Mass Spectrom. 2017, 28, 901–907. [Google Scholar] [CrossRef]
- Dey, S.; Dutta, P.; Majumdar, S. Biological Control of Macrophomina Phaseolina in Vigna Mungo L. by Endophytic Klebsiella Pneumoniae HR1. Jordan J. Biol. Sci. 2019, 12, 219–227. [Google Scholar] [CrossRef]
- Mandal, S. M.; Sharma, S.; Pinnaka, A. K.; Kumari, A.; Korpole, S. Isolation and Characterization of Diverse Antimicrobial Lipopeptides Produced by Citrobacter and Enterobacter. BMC Microbiol. 2013, 13, 152. [Google Scholar] [CrossRef] [PubMed]
- Oleńska, E.; Małek, W.; Wójcik, M.; Swiecicka, I.; Thijs, S.; Vangronsveld, J. Beneficial Features of Plant Growth-Promoting Rhizobacteria for Improving Plant Growth and Health in Challenging Conditions: A Methodical Review. Sci. Total Environ. 2020, 743. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, S.; Kabir, S.; Shabbir, U.; Batool, R. Plant Growth Promoting Rhizobacteria in Sustainable Agriculture: From Theoretical to Pragmatic Approach. Symbiosis 2019, 78, 115–123. [Google Scholar] [CrossRef]
- Aeron, A.; Khare, E.; Jha, C. K.; Meena, V. S.; Aziz, S. M. A.; Islam, M. T.; Kim, K.; Meena, S. K.; Pattanayak, A.; Rajashekara, H.; Dubey, R. C.; Maurya, B. R.; Maheshwari, D. K.; Saraf, M.; Choudhary, M.; Verma, R.; Meena, H. N.; Subbanna, A. R. N. S.; Parihar, M.; Shukla, S.; Muthusamy, G.; Bana, R. S.; Bajpai, V. K.; Han, Y. K.; Rahman, M.; Kumar, D.; Singh, N. P.; Meena, R. K. Revisiting the Plant Growth-Promoting Rhizobacteria: Lessons from the Past and Objectives for the Future. Arch. Microbiol. 2020, 202, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Julian, W. T.; Vasilchenko, A. V.; Shpindyuk, D. D.; Poshvina, D. V.; Vasilchenko, A. S. Bacterial-Derived Plant Protection Metabolite 2,4-Diacetylphloroglucinol: Effects on Bacterial Cells at Inhibitory and Subinhibitory Concentrations. Biomolecules 2021, 11, 1–16. [Google Scholar] [CrossRef]
- Stepanov, A. A.; Poshvina, D. V.; Vasilchenko, A. S. 2,4-Diacetylphloroglucinol Modulates Candida Albicans Virulence. J. Fungi 2022, 8. [Google Scholar] [CrossRef]
- Maketon, C.; Fortuna, A. M.; Okubara, P. A. Cultivar-Dependent Transcript Accumulation in Wheat Roots Colonized by Pseudomonas Fluorescens Q8r1-96 Wild Type and Mutant Strains. Biol. Control 2012, 60, 216–224. [Google Scholar] [CrossRef]
- Naseby, D. C.; Lynch, J. M. Impact of Wild-type and Genetically Modified Pseudomonas Fluorescens on Soil Enzyme Activities and Microbial Population Structure in the Rhizosphere of Pea. Mol. Ecol. 1998, 75, 617–625. [Google Scholar] [CrossRef]
- Vacheron, J.; Desbrosses, G.; Renoud, S.; Padilla, R.; Walker, V.; Muller, D.; PrigentCombaretlt, C. Differential Contribution of Plant-Beneficial Functions from Pseudomonas Kionensis Fl 13 to Root System Architecture Alterations in Arabidopsis Thaliana and Zea Mays. Mol. Plant-Microbe Interact. 2018, 31, 212–223. [Google Scholar] [CrossRef]
- De Leij, F. A. A. M.; Dixon-Hardy, J. E.; Lynch, J. M. Effect of 2,4-Diacetylphloroglucinol-Producing and Non-Producing Strains of Pseudomonas Fluorescens on Root Development of Pea Seedlings in Three Different Soil Types and Its Effect on Nodulation by Rhizobium. Biol. Fertil. Soils 2002, 35, 114–121. [Google Scholar] [CrossRef]
- Raudales, R. E.; Stone, E.; McSpadden Gardener, B. B. Seed Treatment with 2,4-Diacetylphloroglucinol-Producing Pseudomonads Improves Crop Health in Low-PH Soils by Altering Patterns of Nutrient Uptake. Phytopathology 2009, 99, 506–511. [Google Scholar] [CrossRef] [PubMed]
- Andrade, G.; De Leij, F. A. A. M.; Lynch, J. M. Plant Mediated Interactions between Pseudomonas Fluorescens, Rhizobium Leguminosarum and Arbuscular Mycorrhizae on Pea. Lett. Appl. Microbiol. 1998, 26, 311–316. [Google Scholar] [CrossRef]
- Lugtenberg, B. J. J.; Bloemberg, G. V.; Van Brussel, A. A. N.; Kijne, J. W.; Thomas-Oates, J. E.; Spaink, H. P. Signals Involved in Nodulation and Nitrogen Fixation. In Nitrogen Fixation: Fundamentals and Applicantions; Tikhonovich, I. A., Provorov, N. A., Romanov, V. I., Newton, W. E., Eds.; Springer: Dordrecht, 1995; pp. 37–48. [Google Scholar] [CrossRef]
- Combes-Meynet, E.; Pothier, J. F.; Moënne-Loccoz, Y.; Prigent-Combaret, C. The Pseudomonas Secondary Metabolite 2,4-Diacetylphloroglucinol Is a Signal Inducing Rhizoplane Expression of Azospirillum Genes Involved in Plant-Growth Promotion. Mol. Plant-Microbe Interact. 2011, 24, 271–284. [Google Scholar] [CrossRef]
- Kim, H. S.; Sang, M. K.; Jung, H. W.; Jeun, Y. C.; Myung, I. S.; Kim, K. D. Identification and Characterization of Chryseobacterium Wanjuense Strain KJ9C8 as a Biocontrol Agent of Phytophthora Blight of Pepper. Crop Prot. 2012, 32, 129–137. [Google Scholar] [CrossRef]
- Park, M. S.; Jung, S. R.; Lee, K. H.; Lee, M. S.; Do, J. O.; Kim, S. B.; Bae, K. S. Chryseobacterium Soldanellicola Sp. Nov. and Chryseobacterium Taeanense Sp. Nov., Isolated from Roots of Sand-Dune Plants. Int. J. Syst. Evol. Microbiol. 2006, 56, 433–438. [Google Scholar] [CrossRef]
- Anand, A.; Chinchilla, D.; Tan, C.; Mène-Saffrané, L.; L’haridon, F.; Weisskopf, L. Contribution of Hydrogen Cyanide to the Antagonistic Activity of Pseudomonas Strains against Phytophthora Infestans. Microorganisms 2020, 8, 1–10. [Google Scholar] [CrossRef]
- Voisard, C.; Keel, C.; Haas, D.; Dèfago, G. Cyanide Production by Pseudomonas Fluorescens Helps Suppress Black Root Rot of Tobacco under Gnotobiotic Conditions. EMBO J. 1989, 8, 351–358. [Google Scholar] [CrossRef]
- Sehrawat, A.; Sindhu, S. S.; Glick, B. R. Hydrogen Cyanide Production by Soil Bacteria: Biological Control of Pests and Promotion of Plant Growth in Sustainable Agriculture. Pedosphere 2022, 32, 15–38. [Google Scholar] [CrossRef]
- Rijavec, T.; Lapanje, A. Hydrogen Cyanide in the Rhizosphere: Not Suppressing Plant Pathogens, but Rather Regulating Availability of Phosphate. Front. Microbiol. 2016, 7, 1785. [Google Scholar] [CrossRef]
- Ryu, C. M.; Faragt, M. A.; Hu, C. H.; Reddy, M. S.; Wei, H. X.; Paré, P. W.; Kloepper, J. W. Bacterial Volatiles Promote Growth in Arabidopsis. Proc. Natl. Acad. Sci. U. S. A. 2003, 100, 4927–4932. [Google Scholar] [CrossRef]
- Kai, M.; Haustein, M.; Molina, F.; Petri, A.; Scholz, B.; Piechulla, B. Bacterial Volatiles and Their Action Potential. Appl. Microbiol. Biotechnol. 2009, 81, 1001–1012. [Google Scholar] [CrossRef]
- Wu, Y.; Zhou, J.; Li, C.; Ma, Y. Antifungal and Plant Growth Promotion Activity of Volatile Organic Compounds Produced by Bacillus Amyloliquefaciens. Microbiologyopen 2019, 8, 1–14. [Google Scholar] [CrossRef]
- Pest Management Regulatory Agency. Registration Decision RD2014-08, Bacillus subtilis strain GB03; Government of Canada. [CrossRef]
- Zhu, L.; Huang, J.; Lu, X.; Zhou, C. Development of Plant Systemic Resistance by Beneficial Rhizobacteria: Recognition, Initiation, Elicitation and Regulation. Front. Plant Sci. 2022, 13, 1–16. [Google Scholar] [CrossRef]
- Lü, P.; Liu, Y.; Yu, X.; Shi, C. L.; Liu, X. The Right Microbe-Associated Molecular Patterns for Effective Recognition by Plants. Front. Microbiol. 2022, 13, 1–10. [Google Scholar] [CrossRef]
- Tonelli, M. L.; Figueredo, M. S.; Rodríguez, J.; Fabra, A.; Ibañez, F. Induced Systemic Resistance -like Responses Elicited by Rhizobia. Plant Soil 2020, 448, 1–14. [Google Scholar] [CrossRef]
- Van der Ent, S.; Van Wees, S. C. M.; Pieterse, C. M. J. Jasmonate Signaling in Plant Interactions with Resistance-Inducing Beneficial Microbes. Phytochemistry 2009, 70, 1581–1588. [Google Scholar] [CrossRef]
- Altinok, H.; Yildiz, H. Induced Systemic Resistance By Plant Growth-Promoting Rhizobacteria in Control of Plant Diseases. Curr. Trends Nat. Sci. 2019, 8, 125–133. [Google Scholar]
- Pozo, M. J.; Van Der Ent, S.; Van Loon, L. C.; Pieterse, C. M. J. Transcription Factor MYC2 Is Involved in Priming for Enhanced Defense during Rhizobacteria-Induced Systemic Resistance in Arabidopsis Thaliana. New Phytol. 2008, 180, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Gui, Y.; Li, Z.; Jiang, C.; Guo, J.; Niu, D. Induced Systemic Resistance for Improving Plant Immunity by Beneficial Microbes. Plants 2022, 11, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Venegas-Molina, J.; Proietti, S.; Pollier, J.; Orozco-Freire, W.; Ramirez-Villacis, D.; Leon-Reyes, A. Induced Tolerance to Abiotic and Biotic Stresses of Broccoli and Arabidopsis after Treatment with Elicitor Molecules. Sci. Rep. 2020, 10, 1–17. [Google Scholar] [CrossRef]
- Maithani, D.; Singh, H.; Sharma, A. Stress Alleviation in Plants Using SAR and ISR: Current Views on Stress Signaling Network. In Microbes and Signaling Biomolecules Against Plant Stress. Rhizosphere Biology; Sharma, A., Ed.; Springer: Singapore, 2021; pp. 7–36. [Google Scholar] [CrossRef]
- Lucas, J. A.; García-Cristobal, J.; Bonilla, A.; Ramos, B.; Gutierrez-Mañero, J. Beneficial Rhizobacteria from Rice Rhizosphere Confers High Protection against Biotic and Abiotic Stress Inducing Systemic Resistance in Rice Seedlings. Plant Physiol. Biochem. 2014, 82, 44–53. [Google Scholar] [CrossRef]
- Davletova, S.; Rizhsky, L.; Liang, H.; Shengqiang, Z.; Oliver, D. J.; Coutu, J.; Shulaev, V.; Schlauch, K.; Mittler, R. Cytosolic Ascorbate Peroxidase 1 Is a Central Component of the Reactive Oxygen Gene Network of Arabidopsis. Plant Cell 2005, 17, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Arkhipov, A.; Carvalhais, L. C.; Schenk, P. M. PGPR Control Phytophthora Capsici in Tomato through Induced Systemic Resistance, Early Hypersensitive Response and Direct Antagonism in a Cultivar-Specific Manner. Eur. J. Plant Pathol. 2023, 167, 811–832. [Google Scholar] [CrossRef]
- Li, W.; Lee, S. Y.; Cho, Y. J.; Ghim, S. Y.; Jung, H. Y. Mediation of Induced Systemic Resistance by the Plant Growth-Promoting Rhizobacteria Bacillus Pumilus S2-3-2. Mol. Biol. Rep. 2020, 47, 8429–8438. [Google Scholar] [CrossRef] [PubMed]
- Singh, R. P.; Jha, P. N. The Multifarious PGPR Serratia Marcescens CDP-13 Augments Induced Systemic Resistance and Enhanced Salinity Tolerance of Wheat (Triticum Aestivum L.). PLoS One 2016, 11, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Samaras, A.; Kamou, N.; Tzelepis, G.; Karamanoli, K.; Menkissoglu-Spiroudi, U.; Karaoglanidis, G. S. Root Transcriptional and Metabolic Dynamics Induced by the Plant Growth Promoting Rhizobacterium (PGPR) Bacillus Subtilis Mbi600 on Cucumber Plants. Plants 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Samaras, A.; Roumeliotis, E.; Ntasiou, P.; Karaoglanidis, G. Bacillus Subtilis Mbi600 Promotes Growth of Tomato Plants and Induces Systemic Resistance Contributing to the Control of Soilborne Pathogens. Plants 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Liu, Y.; Xu, Y.; Zhang, G.; Shen, Q.; Zhang, R. Exploring Elicitors of the Beneficial Rhizobacterium Bacillus Amyloliquefaciens SQR9 to Induce Plant Systemic Resistance and Their Interactions with Plant Signaling Pathways. Mol. Plant-Microbe Interact. 2018, 31, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Hashmi, N.; Khan, M. M. A.; Moinuddin; Idrees, M.; Aftab, T. Exogenous Salicylic Acid Stimulates Physiological and Biochemical Changes to Improve Growth, Yield and Active Constituents of Fennel Essential Oil. Plant Growth Regul. 2012, 68, 281–291. [Google Scholar] [CrossRef]
- Aftab, T.; Masroor, M.; Khan, A.; Idrees, M.; Naeem, M.; Moinuddin. Salicylic Acid Acts as Potent Enhancer of Growth, Photosynthesis and Artemisinin Production in Artemisia Annua L. J. Crop Sci. Biotechnol. 2010, 13, 183–188. [Google Scholar] [CrossRef]
- Martín-Mex, R.; Nexticapan-Garcéz, Á.; Villanueva-Couoh, E.; Uicab-Quijano, V.; Vergara-Yoisura, S.; Larqué-Saavedra, A. Salicylic Acid Stimulates Flowering in Micropopagated Gloxinia Plants. Rev. Fitotec. Mex. 2015, 38, 115–118. [Google Scholar] [CrossRef]
- Ueda, J.; Miyamoto, K.; Sato, T.; Momotani, Y. Identification of Jasmonic Acid from Euglena Gracilis z as a Plant Growth Regulator. Agric. Biol. Chem. 1991, 55, 275–276. [Google Scholar] [CrossRef]
- Ravnikar, M.; Vilhar, B.; Gogala, N. Stimulatory Effects of Jasmonic Acid on Potato Stem Node and Protoplast Culture. J. Plant Growth Regul. 1992, 11, 29–33. [Google Scholar] [CrossRef]
- Ravnikar, M.; Žel, J.; Plaper, I.; Špacapan, A. Jasmonic Acid Stimulates Shoot and Bulb Formation of Garlic in Vitro. J. Plant Growth Regul. 1993, 12, 73–77. [Google Scholar] [CrossRef]
- Van Peer, R.; Niemann, G. J.; Schippers, B. Induced Resistance and Phytoalexin Accumulation in Biological Control of Fusarium Wilt of Carnation by Pseudomonas Sp. Strain WCS417r. Phytopathology 1991, 728–734. [Google Scholar] [CrossRef]
- Spence, C.; Alff, E.; Johnson, C.; Ramos, C.; Donofrio, N.; Sundaresan, V.; Bais, H. Natural Rice Rhizospheric Microbes Suppress Rice Blast Infections. BMC Plant Biol. 2014, 14, 1–17. [Google Scholar] [CrossRef]
- Nandakumar, R.; Babu, S.; Viswanathan, R.; Raguchander, T.; Samiyappan, R. Induction of Systemic Resistance in Rice against Sheath Blight Disease by Pseudomonas Fluorescens. Soil Biol. Biochem. 2001, 33, 603–612. [Google Scholar] [CrossRef]
- Fatima, S.; Anjum, T. Plant Pathology Potential of Rhizospheric Pseudomonas Strains To Manage Fusarium Wilt of Tomato. J. Agric. Res. 2017, 55, 525–536. [Google Scholar]
- Yan, Z.; Reddy, M. S.; Ryu, C. M.; McInroy, J. A.; Wilson, M.; Kloepper, J. W. Induced Systemic Protection against Tomato Late Blight Elicited by Plant Growth-Promoting Rhizobacteria. Phytopathology 2002, 92, 1329–1333. [Google Scholar] [CrossRef]
- Kloepper, J. W.; Ryu, C. M.; Zhang, S. Induced Systemic Resistance and Promotion of Plant Growth by Bacillus Spp. Phytopathology 2004, 94, 1259–1266. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M. N.; Namood-e-Sahar; Zia-Ul-Husnain Shah, S.; Afghan, S.; Hafeez, F.Y. Suppression of Red Rot Disease by Bacillus Sp. Based Biopesticide Formulated in Non-Sterilized Sugarcane Filter Cake. BioControl 2015, 60, 691–702. [Google Scholar] [CrossRef]
- Li, H.; Ding, X.; Wang, C.; Ke, H.; Wu, Z.; Wang, Y.; Liu, H.; Guo, J. Control of Tomato Yellow Leaf Curl Virus Disease by Enterobacter Asburiae BQ9 as a Result of Priming Plant Resistance in Tomatoes. Turkish J. Biol. 2016, 40, 150–159. [Google Scholar] [CrossRef]
- Fukami, J.; Ollero, F. J.; Megías, M.; Hungria, M. Phytohormones and Induction of Plant-Stress Tolerance and Defense Genes by Seed and Foliar Inoculation with Azospirillum Brasilense Cells and Metabolites Promote Maize Growth. AMB Express 2017, 7. [Google Scholar] [CrossRef]
- De Vleesschauwer, D.; Höfte, M. Using Serratia Plymuthica to Control Fungal Pathogens of Plants. CAB Rev. 2007, 2. [Google Scholar] [CrossRef]
- Liu, D.; Chen, L.; Zhu, X.; Wang, Y.; Xuan, Y.; Liu, X.; Chen, L.; Duan, Y. Klebsiella Pneumoniae SnebYK Mediates Resistance against Heterodera Glycines and Promotes Soybean Growth. Front. Microbiol. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Ji, S. H.; Gururani, M. A.; Chun, S. C. Isolation and Characterization of Plant Growth Promoting Endophytic Diazotrophic Bacteria from Korean Rice Cultivars. Microbiol. Res. 2014, 169, 83–98. [Google Scholar] [CrossRef]
- Wei, G.; Kloepper, J. W.; Tuzun, S. Induction of Systemic Resistance of Cucumber to Colletotrichum Orbiculare by Select Strains of Plant Growth-Promoting Rhizobacteria. Phytopathology 1991, 81, 1508–1512. [Google Scholar] [CrossRef]
- Majdura, J.; Jankiewicz, U.; Gałązka, A.; Orzechowski, S. The Role of Quorum Sensing Molecules in Bacterial–Plant Interactions. Metabolites 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Santoyo, G.; Rtis-Flores, C. A.; Loeza-Lara, P. D.; Orozco-Mosqueda, M. D. C.; Glick, B. R. Rhizosphere Colonization Determinants by Plant Growth-Promoting Rhizobacteria (PGPR). Biology (Basel). 2021, 10, 475. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Singh, P.; Sarmah, B. K.; Nandi, S. P. Quorum Sensing: Its Role in Microbial Social Networking. Res. Microbiol. 2020, 171, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Van Houdt, R.; Givskov, M.; Michiels, C. W. Quorum Sensing in Serratia. FEMS Microbiol. Rev. 2007, 31, 407–424. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, M.; Torres, M.; Blanco, L.; Béjar, V.; Sampedro, I.; Llamas, I. Plant Growth-Promoting Activity and Quorum Quenching-Mediated Biocontrol of Bacterial Phytopathogens by Pseudomonas Segetis Strain P6. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.; Gislason, A. S.; Becker, M.; Belmonte, M. F.; Fernando, W. G. D.; de Kievit, T. R. Investigation of the Quorum-Sensing Regulon of the Biocontrol Bacterium Pseudomonas Chlororaphis Strain PA23. PLoS One 2020, 15, e0226232. [Google Scholar] [CrossRef] [PubMed]
- Schuhegger, R.; Ihring, A.; Gantner, S.; Bahnweg, G.; Knappe, C.; Vogg, G.; Hutzler, P.; Schmid, M.; Van Breusegem, F.; Eberl, L.; Hartmann, A.; Langebartels, C. Induction of Systemic Resistance in Tomato by N-Acyl-L-Homoserine Lactone-Producing Rhizosphere Bacteria. Plant, Cell Environ. 2006, 29, 909–918. [Google Scholar] [CrossRef]
- Liu, X.; Bimerew, M.; Ma, Y.; Müller, H.; Ovadis, M.; Eberl, L.; Berg, G.; Chernin, L. Quorum-Sensing Signaling Is Required for Production of the Antibiotic Pyrrolnitrin in a Rhizospheric Biocontrol Strain of Serratia Plymuthica. FEMS Microbiol. Lett. 2007, 270, 299–305. [Google Scholar] [CrossRef]
- Pang, Y.; Liu, X.; Ma, Y.; Chernin, L.; Berg, G.; Gao, K. Induction of Systemic Resistance, Root Colonisation and Biocontrol Activities of the Rhizospheric Strain of Serratia Plymuthica Are Dependent on N-Acyl Homoserine Lactones. Eur. J. Plant Pathol. 2009, 124, 261–268. [Google Scholar] [CrossRef]
- Rankl, S.; Gunsé, B.; Sieper, T.; Schmid, C.; Poschenrieder, C.; Schröder, P. Microbial Homoserine Lactones (AHLs) Are Effectors of Root Morphological Changes in Barley. Plant Sci. 2016, 253, 130–140. [Google Scholar] [CrossRef]
- Moshynets, O. V.; Babenko, L. M.; Rogalsky, S. P.; Iungin, O. S.; Foster, J.; Kosakivska, I. V.; Potters, G.; Spiers, A. J. Priming Winter Wheat Seeds with the Bacterial Quorum Sensing Signal N-Hexanoyl-L-Homoserine Lactone (C6-HSL) Shows Potential to Improve Plant Growth and Seed Yield. PLoS One 2019, 14, 1–17. [Google Scholar] [CrossRef]
- Shrestha, A.; Grimm, M.; Ojiro, I.; Krumwiede, J.; Schikora, A. Impact of Quorum Sensing Molecules on Plant Growth and Immune System. Front. Microbiol. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Hartmann, A.; Klink, S.; Rothballer, M. Importance of N-Acyl-Homoserine Lactone-Based Quorum Sensing and Quorum Quenching in Pathogen Control and Plant Growth Promotion. Pathogens 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Barriuso, J.; Ramos Solano, B.; Fray, R. G.; Cámara, M.; Hartmann, A.; Gutiérrez Mañero, F. J. Transgenic Tomato Plants Alter Quorum Sensing in Plant Growth-Promoting Rhizobacteria. Plant Biotechnol. J. 2008, 6, 442–452. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Yang, X. Y.; Li, Y.; Liu, F.; Cao, X. Y.; Jia, Z. H.; Song, S. S. N-3-Oxo-Hexanoyl-Homoserine Lactone, a Bacterial Quorum Sensing Signal, Enhances Salt Tolerance in Arabidopsis and Wheat. Bot. Stud. 2020, 61. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M. S.; Arshad, A.; Rajput, L.; Fatima, K.; Ullah, S.; Ahmad, M.; Imran, A. Growth-Stimulatory Effect of Quorum Sensing Signal Molecule N-Acyl-Homoserine Lactone-Producing Multi-Trait Aeromonas Spp. on Wheat Genotypes Under Salt Stress. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Von Rad, U.; Klein, I.; Dobrev, P. I.; Kottova, J.; Zazimalova, E.; Fekete, A.; Hartmann, A.; Schmitt-Kopplin, P.; Durner, J. Response of Arabidopsis Thaliana to N-Hexanoyl-DL-Homoserine-Lactone, a Bacterial Quorum Sensing Molecule Produced in the Rhizosphere. Planta 2008, 229, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Li, M.; Jia, Z.; Liu, F.; Ma, H.; Huang, Y.; Song, S. AtMYB44 Positively Regulates the Enhanced Elongation of Primary Roots Induced by N-3-Oxo-Hexanoyl-Homoserine Lactone in Arabidopsis Thaliana. Mol. Plant-Microbe Interact. 2016, 29, 774–785. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Bian, Z.; Jia, Z.; Zhao, Q.; Song, S. The GCR1 and GPA1 Participate in Promotion of Arabidopsis Primary Root Elongation Induced by N-Acyl-Homoserine Lactones, the Bacterial Quorum-Sensing Signals. Mol. Plant-Microbe Interact. 2012, 25, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Castro, R.; Díaz-Pérez, C.; Martínez-Trujillo, M.; Del Río, R. E.; Campos-García, J.; López-Bucio, J. Transkingdom Signaling Based on Bacterial Cyclodipeptides with Auxin Activity in Plants. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 7253–7258. [Google Scholar] [CrossRef]
- Ortíz-Castro, R.; Martínez-Trujillo, M.; López-Bucio, J. N-Acyl-L-Homoserine Lactones: A Class of Bacterial Quorum-Sensing Signals Alter Post-Embryonic Root Development in Arabidopsis Thaliana. Plant, Cell Environ. 2008, 31, 1497–1509. [Google Scholar] [CrossRef]
- Hanif, M. K.; Malik, K. A.; Hameed, S.; Saddique, M. J.; Ayesha; Fatima, K. ; Naqqash, T.; Majeed, A.; Iqbal, M. J.; Imran, A. Growth Stimulatory Effect of AHL Producing Serratia Spp. from Potato on Homologous and Non-Homologous Host Plants. Microbiol. Res. 2020, 238, 126506. [Google Scholar] [CrossRef]
- Jadhav, H.; Shaikh, S.; Sayyed, R. Role of Hydrolytic Enzymes of Rhizoflora in Biocontrol of Fungal Phytopathogens: An Overview. In Rhizotrophs: Plant Growth Promotion to Bioremediation; Mehnaz, S., Ed.; Springer: Singapore, 2017; pp. 183–203. [Google Scholar] [CrossRef]
- Khalil, M. S. M.; Hassan, M. H. A. R.; Mahmoud, A. F.; Morsy, K. M. M. Involvement of Secondary Metabolites and Extracellular Lytic Enzymes Produced by Plant Growth Promoting Rhizobacteria in Inhibiting the Soilborne Pathogens in Faba Bean Plants. J. Trop. Plant Pests Dis. 2022, 22, 100–108. [Google Scholar] [CrossRef]
- Polonio, Á.; Vida, C.; de Vicente, A.; Cazorla, F. M. Impact of Motility and Chemotaxis Features of the Rhizobacterium Pseudomonas Chlororaphis PCL1606 on Its Biocontrol of Avocado White Root Rot. Int. Microbiol. 2017, 20, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Pan, R.; Shen, Y.; Yuan, J.; Wang, L.; Luo, X.; Raza, W.; Ling, N.; Huang, Q.; Shen, Q. Development of a Novel Bio-Organic Fertilizer for Plant Growth Promotion and Suppression of Rhizome Rot in Ginger. Biol. Control 2017, 114, 97–105. [Google Scholar] [CrossRef]
- Mishra, P.; Mishra, J.; Dwivedi, S. K.; Arora, N. K. Microbial Enzymes in Biocontrol of Phytopathogens. In Microbial Enzymes: Roles and Applications in Industries. Microorganisms for Sustainability; Arora, N., Mishra, J., Mishra, V., Eds.; Springer: Singapore, 2020; pp. 259–285. [Google Scholar] [CrossRef]
- Adhav, H.; Shaikh, S.; Sayyed, R. Role of Hydrolytic Enzymes of Rhizoflora in Biocontrol of Fungal Phytopathogens: An Overview. In Rhizotrophs: Plant Growth Promotion to Bioremediation. Microorganisms for Sustainability; Mehnaz, S., Ed.; Springer: Singapore, 2017; pp. 183–203. [Google Scholar] [CrossRef]
- Reddy, E. C.; Reddy, G. S.; Goudar, V.; Sriramula, A.; Swarnalatha, G. V; Al Tawaha, A. R. M.; Sayyed, R.Z. Hydrolytic Enzyme Producing Plant Growth-Promoting Rhizobacteria (PGPR) in Plant Growth Promotion and Biocontrol. In Secondary Metabolites and Volatiles of PGPR in Plant-Growth Promotion; Sayyed, R., Uarrota, V., Eds.; Springer, 2022; pp. 303–312. [Google Scholar] [CrossRef]
- Yoshimi, A.; Miyazawa, K.; Abe, K. Function and Biosynthesis of Cell Wall α-1,3-Glucan in Fungi. J. Fungi 2017, 3, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Ueki, A.; Takehara, T.; Ishioka, G.; Kaku, N.; Ueki, K. Production of β-1,3-Glucanase and Chitosanase from Clostridial Strains Isolated from the Soil Subjected to Biological Disinfestation. AMB Express 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Suyotha, W.; Yano, S.; Wakayama, M. α-1,3-Glucanase: Present Situation and Prospect of Research. World J. Microbiol. Biotechnol. 2016, 32, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gomma, M. A.; Radwan, F. I.; Kandil, E. E.; Gharib, A. F. Effect of Zinc Application on Growth and Yield of Rice ( Oryza Sativa L.) Effect of Zinc Application on Growth and Yield of Rice ( Oryza Sativa L.). Middle East J. Appl. Sci. 2015, 5, 913–919. [Google Scholar] [CrossRef]
- de Souza, R.; Ambrosini, A.; Passaglia, L. M. P. Plant Growth-Promoting Bacteria as Inoculants in Agricultural Soils. Genet. Mol. Biol. 2015, 38, 401–419. [Google Scholar] [CrossRef]
- Kalbe, C.; Marten, P.; Berg, G. Strains of the Genus Serratia as Beneficial Rhizobacteria of Oilseed Rape with Antifungal Properties. Microbiol. Res. 1996, 151, 433–439. [Google Scholar] [CrossRef]
- Zia, M. A.; Yasmin, H.; Shair, F.; Jabeen, Z.; Mumtaz, S.; Hayat, Z.; Shah, S. Z. ul H.; Afghan, S.; Hafeez, F. Y.; Hassan, M. N. Glucanolytic Rhizobacteria Produce Antifungal Metabolites and Elicit ROS Scavenging System in Sugarcane. Sugar Tech 2019, 21, 244–255. [Google Scholar] [CrossRef]
- Sayyed, R. Z.; Uarrota, V. G. Hydrolytic Enzyme Producing Plant Growth-Promoting Rhizobacteria (PGPR) in Plant Growth Promotion and Biocontrol. In Secondary Metabolites and Volatiles of PGPR in Plant-Growth Promotion; Sayyed, R., Uarrota, V., Eds.; Springer, 2022; pp. 303–312. [Google Scholar] [CrossRef]
- Perrot, T.; Pauly, M.; Ramírez, V. Emerging Roles of β-Glucanases in Plant Development and Adaptative Responses. Plants 2022, 11. [Google Scholar] [CrossRef]
- Dukare, A.; Paul, S.; Arambam, A. Isolation and Efficacy of Native Chitinolytic Rhizobacteria for Biocontrol Activities against Fusarium Wilt and Plant Growth Promotion in Pigeon Pea (Cajanus Cajan L.). Egypt. J. Biol. Pest Control 2020, 30, 1–12. [Google Scholar] [CrossRef]
- Mohammed, A. F. Optimization of Cellulase and Chitinase Enzymes Production by Plant Growth Promoting Rhizobacteria. Nov. Res. Microbiol. J. 2020, 4, 641–652. [Google Scholar] [CrossRef]
- Yuliar; Kartadi, S. F.; Salmah, A. Combined Use of Enterobacter Cloacae MB20 and the Microelements of Copper and Manganese to Control Damping-off of Tomato. Earth Environ. Sci. 2019, 308, 012025. [Google Scholar] [CrossRef]
- Singh, S. P.; Gaur, R. Evaluation of Antagonistic and Plant Growth Promoting Activities of Chitinolytic Endophytic Actinomycetes Associated with Medicinal Plants against Sclerotium Rolfsii in Chickpea. J. Appl. Microbiol. 2016, 121, 506–518. [Google Scholar] [CrossRef]
- Kim, H. J.; Choi, H. S.; Yang, S. Y.; Kim, I. S.; Yamaguchi, T.; Sohng, J. K.; Park, S. K.; Kim, J. C.; Lee, C. H.; Gardener, B. M.; Kim, Y. C. Both Extracellular Chitinase and a New Cyclic Lipopeptide, Chromobactomycin, Contribute to the Biocontrol Activity of Chromobacterium Sp. C61. Mol. Plant Pathol. 2014, 15, 122–132. [Google Scholar] [CrossRef]
- Gongora, C. E.; Broadway, R. M. Plant Growth and Development Influenced by Transgenic Insertion of Bacterial Chitinolytic Enzymes. Mol. Breed. 2002, 9, 123–135. [Google Scholar] [CrossRef]
- Fitza, K. N. E.; Payn, K. G.; Steenkamp, E. T.; Myburg, A. A.; Naidoo, S. Chitosan Application Improves Resistance to Fusarium Circinatum in Pinus Patula. South African J. Bot. 2013, 85, 70–78. [Google Scholar] [CrossRef]
- Zong, H.; Li, K.; Liu, S.; Song, L.; Xing, R.; Chen, X.; Li, P. Improvement in Cadmium Tolerance of Edible Rape (Brassica Rapa L.) with Exogenous Application of Chitooligosaccharide. Chemosphere 2017, 181, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, E. Z. Microbial Chitinases: Properties, Enhancement and Potential Applications. Protoplasma 2021, 258, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Brar, A.; Yadav, M.; Chawade, A.; Vivekanand, V.; Pareek, N. Chitinases—Potential Candidates for Enhanced Plant Resistance towards Fungal Pathogens. Agric. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Billot, R.; Plener, L.; Jacquet, P.; Elias, M.; Chabrière, E.; Daudé, D. Engineering Acyl-Homoserine Lactone-Interfering Enzymes toward Bacterial Control. J. Biol. Chem. 2020, 295, 12993–13007. [Google Scholar] [CrossRef]
- de Leon, V.; Orr, K.; Stelinski, L. L.; Mandadi, K.; Ibanez-Carrasco, F. Inoculation of Tomato With Plant Growth Promoting Rhizobacteria Affects the Tomato—Potato Psyllid—Candidatus Liberibacter Solanacearum Interactions. J. Econ. Entomol. 2023, 116, 379–388. [Google Scholar] [CrossRef]
- Mumtaz, M. Z.; Ahmad, M.; Zafar-Ul-Hye, M.; Saqib, M.; Akhtar, M. F. U. Z.; Zaheer, M. S. Seed-Applied Zinc-Solubilising Bacillus Biofertilisers Improve Antioxidant Enzyme Activities, Crop Productivity, and Biofortification of Maize. Crop Pasture Sci. 2022, 73, 503–514. [Google Scholar] [CrossRef]
- Saral, A.; Kanekar, S.; Koul, K. K.; Bhagyawant, S. S. Plant Growth Promoting Bacteria Induce Anti-Quorum-Sensing Substances in Chickpea Legume Seedling Bioassay. Physiol. Mol. Biol. Plants 2021, 27, 1577–1595. [Google Scholar] [CrossRef] [PubMed]
- Sikdar, R.; Elias, M. Quorum Quenching Enzymes and Their Effects on Virulence, Biofilm, and Microbiomes: A Review of Recent Advances. Expert Rev. Anti. Infect. Ther. 2020, 18, 1221–1233. [Google Scholar] [CrossRef] [PubMed]
- Bzdrenga, J.; Daudé, D.; Rémy, B.; Jacquet, P.; Plener, L.; Elias, M.; Chabrière, E. Biotechnological Applications of Quorum Quenching Enzymes. Chem. Biol. Interact. 2017, 267, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Achari, G. A.; Ramesh, R. Characterization of Quorum Quenching Enzymes from Endophytic and Rhizosphere Colonizing Bacteria. Biocatal. Agric. Biotechnol. 2018, 13, 20–24. [Google Scholar] [CrossRef]
- Mehmood, A.; Liu, G.; Wang, X.; Meng, G.; Wang, C.; Liu, Y. Fungal Quorum-Sensing Molecules and Inhibitors with Potential Antifungal Activity: A Review. Molecules 2019, 24, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Khatoon, Z.; Huang, S.; Rafique, M.; Fakhar, A.; Kamran, M. A.; Santoyo, G. Unlocking the Potential of Plant Growth-Promoting Rhizobacteria on Soil Health and the Sustainability of Agricultural Systems. J. Environ. Manage. 2020, 273. [Google Scholar] [CrossRef]
- Kumar, V. K. K.; Reddy, M. S.; Kloepper, J. W.; Yellareddygari, S. K. R.; Lawrence, K. S.; Zhou, X. G.; Sudini, H.; Miller, M. E.; Podile, A. .; Reddy, E..; Niranjana, S. R.; Nayaka, C. S. Plant Growth-Promoting Activities of Bacillus Subtilis Mbi 600 (Integral®) and Its Compatibility With Commonly Used Fungicides in Rice Sheath Blight Management. Int. J. Microbiol. Res. 2011, 3, 120–130. [Google Scholar] [CrossRef]
- Jang, S.; Choi, S. K.; Zhang, H.; Zhang, S.; Ryu, C. M.; Kloepper, J. W. History of a Model Plant Growth-Promoting Rhizobacterium, Bacillus Velezensis GB03: From Isolation to Commercialization. Front. Plant Sci. 2023, 14, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Mehnaz, S. An Overview of Globally Available Bioformulations. In Bioformulations: for Sustainable Agriculture; Arora, N., Mehnaz, S., Balestrini, R., Eds.; Springer: New Delhi, 2016; pp. 267–281. [Google Scholar] [CrossRef]
Figure 1.
Mechanisms of plant growth promotion and pathogen biocontrol mediated by DAPG produced by growth-promoting pseudomonads. DAPG can directly inhibit the growth of soilborne pathogens such as R. solani. At the same time, DAPG may also be directly associated with the enhancement of plant traits such as root biomass and lateral root development and may also contribute to nutrient availability. This compound is also beneficial in the recruitment of Rhizobium, which promotes the formation of root nodules and nitrogen fixation.
Figure 1.
Mechanisms of plant growth promotion and pathogen biocontrol mediated by DAPG produced by growth-promoting pseudomonads. DAPG can directly inhibit the growth of soilborne pathogens such as R. solani. At the same time, DAPG may also be directly associated with the enhancement of plant traits such as root biomass and lateral root development and may also contribute to nutrient availability. This compound is also beneficial in the recruitment of Rhizobium, which promotes the formation of root nodules and nitrogen fixation.
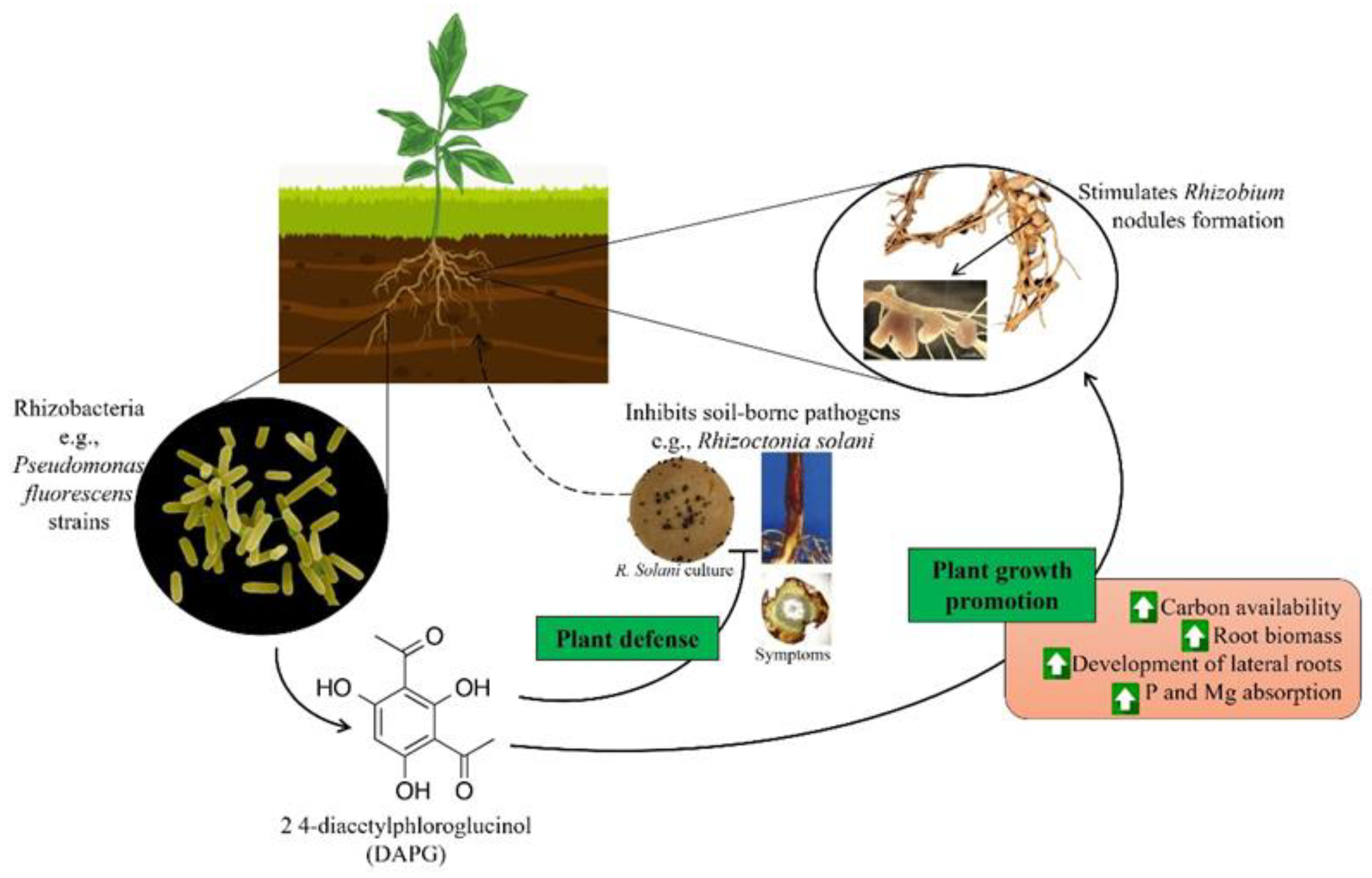
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



