
Submitted:
05 February 2024
Posted:
06 February 2024
You are already at the latest version
Abstract
Salinity is one of the major factors responsible for the deterioration of soils, making them unsuitable for agriculture. The application of halotolerant and halophilic plant growth promoting bacteria (PGPR: Plant Growth Promoting Rhizobacteria) with biological control activities as an inoculant of cultivated plants offers a biological alternative to the use of agrochemicals, particularly when subjected to salt stress. In this perspective, 70 bacterial strains were isolated from saline soils (sebkha) in arid and semi-arid areas of eastern Algeria. Three isolates were selected based on their ability to produce bioactive molecules allowing them to promote plant growth, such as hydrolytic enzymes, indole acetic acid, HCN, NH3, etc. Two of these isolates belong to the genus Serratia and the third is a halophilic Halomonas bacteria. These bacteria were identified based on the 16S rDNA sequence. Antagonism tests against phytopathogenic fungi were carried out. The identification of the antifungal molecules produced by these bacteria was done by high performance liquid chromatography. These bacteria are capable of inhibiting mycelial development against phytopathogenic fungi with rates reaching 80.67% against Botrytis cinerea, 76.22% against Aspergillus niger and 66.67% against Fusarium culmorum for Serratia sp. The strain Halomonas sp. inhibited mycelial growth by production of volatile substances of Aspergillus niger at 71.29%, Aspergillus flavus at 75.49% and Penicillium glabrum at a rate of 72.22%. The identification by HPLC of the antifungal molecules produced by these three bacteria reveals that they are polyphenols, which makes these strains the first rhizobacteria capable of producing phenolic compounds. Finally, pot tests to determine the effectiveness of these strains in promoting wheat growth under salinity stress (125 mM, 150 mM and 200 mM) were carried out. The results revealed that a consortium of two isolates (Serratia sp. and Halomonas sp.) performed best at 125 mM. But, at higher concentrations it was the halophilic bacteria Halomonas sp which gave the best result. In all cases, there was a significant improvement in the growth of wheat seedlings inoculated with the bacteria, compared to non-inoculated controls.
Keywords:
halophilic bacteria
; salt stress
; biological control
; polyphenols
; Plant Growth Promoting Bacteria (PGPB)
1. Introduction
Soil salinization is a major challenge in arid and semi-arid regions, which seriously affects agricultural production [1]. 20% of total cultivated land and 33% of irrigated agricultural land worldwide are affected [2]. According to the Food and Agriculture Organization (FAO), around 1 billion hectares of the world's land surface are affected by salinization, which represents around 8.7% of the planet's land surface, this causes an estimated 27,3 billion dollars in economic losses [3]. Each year, approximately 10 million hectares of the world's agricultural land are destroyed by salt accumulation [4].
The increase in salinity level has negative repercussions on soil properties and plant physiology leading to deterioration in plant development [5]. Saline soils have high electrical conductivity (EC) and impose ionic toxicity, osmotic and oxidative stress, nutrient deficiency on plants and thus limit water absorption from the soil [2].
Faced with this saline stress, numerous physical, chemical and biological strategies such as plant breeding, genetic engineering of plants and a whole series of agricultural techniques. Recently, the application of rhizobacteria has been developed to improve the plant defense system [6]. The application of PGPR is one of the most potential biological measures for improving agricultural yield. [7,8]. Many studies have shown that PGPR directly or indirectly promotes plant growth. Direct mechanisms refer to the ability of PGPR to fix nitrogen, dissolve organic or inorganic phosphorus, produce phytohormones, such as auxin or indole-3-acetic acid (IAA), and other bioactive molecules that stimulate the growth and development of plants [9,10,11,12,13]. For indirect mechanisms, PGPR participates in preventing the development of phytopathogens, either through competition for space and nutrient occupation, or by producing metabolites such as antibiotics, inducing plant resistance [14,15,16].
To deal with salt stress, in recent decades there has been interest in the application of halotolerant and halophilic bacteria. These PGPRs promote plant growth under salt stress through complex mechanisms, such as enhancing antioxidant enzymes to regulate reactive oxygen (ROS), enhancing the accumulation of intracellular osmolytes to ensure ionic balance and through the accumulation of secondary metabolites [17,18,19].
On the other hand, plant pathogens have a devastating effect on agricultural crops. The use of chemical substances has a negative effect on health and the environment. Therefore, the use of plant growth-promoting rhizobacteria (PGPR) as growth-promoting and plant disease-fighting agents is a better alternative [20].
In this study, we isolated three bacteria from the rhizosphere of halophytic plants from two arid and semi-arid regions of eastern Algeria. These bacteria have been selected for their ability to produce hydrolytic enzymes, hydrogen cyanide, ammonia, indole acetic acid (IAA), etc. After identification, the PGP isolates were tested for their in vitro ability to inhibit fungal growth and their ability to enhance the growth of wheat plants under salt stress.
2. Materials and methods
2.1. Soil Sampling
Sampling was carried out at two sites; Sebkhet Bazer (36°03′16′′N 5°40′35′′E) and Chott El Hodna (35°25’34’’N 4°44’39’’E), in Eastern Algeria. 10 soil samples were collected from each site in the rhizosphere of wild halophilic plants.
The samples were collected in sterile plastic bags and then transported to the laboratory at 4°C. 10g of soil from each sample was homogenized in sterile physiological water. 100µl of each suspension were inoculated into culture media for halophiles with a NaCl concentration ranging from 0.5M to 3M in triplicate. After 72 hours of incubation at 28°C, the bacteria were isolated on the same medium supplemented with agar. 70 bacterial isolates were purified then preserved for subsequent studies.
2.2. Production of extracellular lytic enzymes:
2.2.1. Cellulase:
The search for extracellular cellulase enzymes was carried out using agar disks 6 mm in diameter containing young bacterial cultures and placed inside Petri dishes containing Carder's medium (1986); composition in g/L: Na2HPO4 (6); KH2PO4 (3); NaCl (0.5); NH4Cl (1); Yeast extract (3); CMC (carboxymethylcellulose) (7); Agar (15); at a pH of 7.2 ± 0.02. The boxes are subsequently incubated at 28°C for 8 days [21]. After incubation, a solution of Congo red (0.1%) is added to the surface of the media. After 20 minutes, the surface of the box is flooded with a 1M NaCl solution, then left to stand overnight. The appearance of a clear halo around the colonies indicates the degradation of the CMC and reflects the presence of an extracellular cellulase.
2.2.2. Amylase:
Discs of young bacterial cultures are placed on starch-based culture medium in order to demonstrate their ability to degrade starch. The medium was prepared according to the method described by Vinoth et al. [22] and whose composition in g/L is: KNO3 (0.5); K2HPO4 (1.0); MgSO4 (0.2); CaCl2 (0.1); FeCl3 (0.001); soluble starch (10.0); agar (15.0); 7.2 ± 0.02 pH. The boxes are incubated for a period ranging from 48-72 h at 28°C. After incubation, a solution of (lugol) is used to flood the culture medium inside the boxes. After a few minutes of contact, the plates are washed with distilled water, and the appearance of clear zones around the agar discs reflects the presence of extracellular amylase in the medium. Areas where the starch is not degraded appear blackish blue.
2.2.3. Lipase:
Discs of young bacterial cultures are placed on culture medium containing g/l: peptone (10); NaCl (5.0); CaCl2 2H2O (0.1); Tween 20 (1%, v/v) and Agar (18). The pH is adjusted to 7.4. After 48 hours of incubation at 28°C, a positive result results in the appearance of a clear halo around the colonies [21, 23]
2.2.4. Protease:
2.2.5. Urease:
According to Christensen [25], the revelation of bacterial urease is carried out by the agar disk method on a medium with a pH of 6.8 prepared as follows: Add (in g) to 950 ml of distilled water: Peptone (1);Glucose (1); NaCl (5); Na2HPO4 (1.2); KH2PO4 (0.8); Phenol red (0.012); Agar (15).After autoclaving, add 50 ml of a sterile urea solution (40%) and pour the medium into Petri dishes. Isolates having produced urease show a pink halo around their colonies.
2.2.6. Chitinase:
The following culture medium is used, it is composed as follows in g/l:
Colloidal chitin (0.8 to 0.6); K2HPO4 (2.7); KH2PO4(0.3); MgSO47H2O (0.7); NaCl (0.5); KCl (0.5); Yeast extract (0.13); Agar (15). The medium is inoculated by the disk method and incubated at 28°C for a minimum of 7 days. Chitinasic activity is manifested by the appearance of a transparent halo around these discs [26,27].
2.3. PGPR features:
The isolates with the best scores were tested to select the best performing strains.
2.3.1. Nitrogen fixation:
The bacterial isolates were streaked on nitrogen-free “Burk’s N-free” culture medium containing in g/l: glucose (10); NaCl (30); KH2PO4 (0.4); K2HPO4 (0.5); Na2SO4 (0.05); CaCl2 (0.2); MgSO4.7H2O (0.005); Na2MoO4 (0.003); agar (15). The development of bacterial colonies demonstrates the capacity of the strain to fix atmospheric nitrogen.
2.3.2. Solubilization of phosphates:
The bacterial isolates were incubated at 28°C for 7 days on modified Pikovskaya medium containing in g/l: glucose (10); Ca3(PO4)2 (5); (NH4)2SO4, (0.5); MgSO4-7H2O (0.1); KCl (0.2); yeast extract (0.5); MnSO4-H2O (0.002); FeSO4-7H2O (0.002); NaCl (50) and agar (15).
2.3.3. Production of ammonia (NH3):
The bacterial isolates were inoculated in tubes containing 5ml of peptone water (peptone 20g/l and NaCl 30g/l) then incubated at 28°C for 48h. After incubation, 0.25ml of Nessler reagent is added to each tube and the appearance of a brown to yellow color indicates the production of ammonia.
2.3.4. Production of hydrogen cyanide (HCN):
The production of HCN is determined following the method of Lorck [28]. The bacteria are streaked on nutrient agar supplemented with glycine at a rate of 4.4 g/l. A sterile disk of whatman paper is placed on the lid of the Petri dish and impregnated with a solution of sodium picrate (2% anhydrous sodium carbonate in 5% picric acid). The petri dish is sealed with Parafilm and incubated at 28°C for 96 hours. The color change to orange or brown indicates the production of HCN.
2.3.5. Production of indole 3-acetic acid (IAA)
Demonstration of the capacity of each isolate to produce IAA was determined by the Salkowski colorimetric method [29]. Each isolate is inoculated in modified HM culture broth containing 1 mg.ml-1 and 5% glucose, then incubated with shaking (100 rpm) at 28°C for 3 days. After incubation, 5ml of each culture is centrifuged. 2ml of the supernatant is added with 2ml of Salkowski's reagent (2% FeCl3 0.5M in 35% HClO4) and incubated in the dark at room temperature for 25 min. optical density is measured at 530 nm.
2.4. Molecular identification:
The selected strains were identified based on the 16S rDNA sequence. DNA transmission was followed by amplification of this gene using the forward primer (356F) 5'ACWCCTACGGGWGGCWGC and the reverse primer (1064R) 5'AYCTCACGRCACGAGCTGAC. Polymerase chain reactions (PCR) were carried out in a final volume of 20 μl. The reaction mixture contains: 10µl of Xpert Fast Hotstar Mastermix 2x (Grisp), 1µl of DNA sample and 7µl of ultra pure water. The mixture was placed in the T100 PCR thermocycler (BIO-RAD), programmed as follows: initial denaturation at 95°C for 3 min, then 35 cycles each composed of three steps: denaturation at 95°C for 15 s, annealing at 55°C for 15 s, elongation at 72°C for 15 s, then a final elongation at 72°C for 3 min. Aliquots of PCR reactions were resolved on 1% agarose gels stained with ethidium bromide. The purified PCR products were sequenced in one direction on StabVida (Caprica, Portugal) using the previously mentioned primers.
2.5. Antifungal activity
The antifungal activity of the selected bacterial strains was tested against four fungi: Aspergillus niger, Aspergillus flavus, Penicillium glabrum Fusarium culmurum and Botrytis cinerea.
2.5.1. In-vitro tests:
Directed inhibition of mycelial growth (direct confrontation test):
The direct confrontation method on agar [30,31] was used. A disc of young mycelium (5 days old) from each mushroom is placed in the center of a Petri dish containing PDA (Potato-Dextrose-Agar) medium. 3 disks of a young culture of each bacterial strain are placed 2.5 cm from the fungus. Plates with sterile agar disks served as controls. After 7 days of incubation at 25°C, the percentage of inhibition is calculated according to the following formula: PGI% = (KR-R1) /KR x 100 where KR is the average diameter between the point of deposition of the fungus and the margin of the colony contained in the control (mm), R1 is the average diameter between the point of deposition of the fungus and the margin of the colony contained in the treated Petri dish (mm) [32,33].
Indirect inhibition of mycelial growth (indirect confrontation test):
This test was carried out against A. niger. Bacterial filtrates were prepared by the modified method of Li et al. [34]. The bacterial strains were inoculated on HM agar at 28°C for 24 hours then subcultured in the same liquid medium. Three categories of cultures were prepared. The first pure, containing the bacterial strains alone. The second mixed, where disks of the pathogen (A. niger) were added. In the third, the bacterial strains were cultivated in HM culture medium free of glucose. After incubation with shaking (100 rpm) at 28°C for 72 h, these media were centrifuged at 6000 rpm for 30 min, and the supernatant was filtered through a sterile 0.22 µm multi-pore membrane.
Petri dishes containing the PDA medium are inoculated in tight streaks with the sporal suspension of the phytopathogen, then sterile paper disks soaked in the filtrate are placed aseptically to test the antifungal activity. Sterile discs soaked in sterile HM medium will serve as controls. The boxes are incubated at 25°C for 2 days. This test is carried out following the modified method of Aghighi et al. [35] and Mahdieh et al. [36].
Indirect inhibition of mycelial growth by production of volatile substances:
The test for inhibition of mycelial growth by production of volatile substances was carried out following the protocol described by Dennis and Webster [37]. A Petri dish containing the HM medium is inoculated in tight streaks with the bacterial antagonist. Another Petri dish containing the PDA medium is inoculated with a 5mm diameter disk of the pathogenic fungus (A. niger, A. flavus, P. glabrum or F. culmorum). The bottom of the two Petri dishes containing the seeded culture media are placed face to face and sealed with parafilm. Petri dishes without bacteria are used as a control. The radial growth of the mushrooms is measured after 7 days of incubation and is compared to the control.
2.5.2. Characterization of bacterial antifungal metabolites:
The filtrates obtained were freeze-dried at 40°C and 200 KPa. [38]. The residue obtained is dissolved in 50 ml of methanol then adjusted to 100 ml with distilled water. The mixture obtained is centrifuged (1535 x g) for 20 min then made up to 100 ml with a methanol-water solution (50:50 v/v) following the modified method of Kim et al. [39] and N’Guessan et al. [40].
A Hypersil BDS-C18 reversed-phase column (250 × 4.6 mm, 5 μm) was used for chromatographic separations. The mobile phase is composed of 0.2% acetic acid/acetonitrile (ACN) in a gradient, starting with 95% 0.2% acetic acid and ending with 100% ACN. The flow rate was 1.5 ml min-1 for 30 min.
The injection volume of the SPCs (standard polyphenol compounds) was 20 μl, while that of the samples and its standard solutions was 10 μl. The column temperature was maintained at 30 °C. The UV detection wavelengths were 230-255-280-300 and 355nm, consistent with the UV scanning results [41]
The standards used in this study were: Gallic acid, Hydroxy quinone, Resorcinol, Catechin, 1-2di hydroxy benzene, Syringic acid, Naringenin 7 glucoside, 3 4 5 Trimethoxy benzoic acid, M anisic acid, Hesperidine.
2.5.3. In-vivo test:
Biological test on fruits (apples):
This test is carried out according to the modified protocol of Xiao and Kim [42]. Fresh apples of the “Golden Delicious” variety of approximately the same size and at the same level of maturity were sterilized with bleach (2%) for 2 min, then rinsed three times with sterile distilled water. Four wounds (3 mm wide x 3 mm deep) were made in the equatorial zone of each fruit. Each well is inoculated with 30 µl of the bacterial antagonist (108 CFU/ml for each strain tested) and 30 µl of sterile distilled water for the control [43]. After 2 hours at room temperature, 15 µl of the spore suspension of the pathogen (2x104 spores / ml for Botrytis cinerea, and 105 spores / ml for Aspergillus niger) were added. The fruits were incubated at 25°C at 90 to 95% relative humidity for 6 days [44]. The experiment was carried out in triplicate with 6 fruits in each repetition. Injuries are assessed daily. Disease severity was calculated by the incidence and size of lesions around the wounds. Wound diameters were measured 3 and 6 days after inoculation [45].
2.6. Plant Growth Stimulation Tests
2.6.1. Potting
Sterilization of the surface of durum wheat seeds (Triticum durum, Simeto variety) with 70% ethanol and 3% sodium hypochlorite then rinsed 5 times with sterile distilled water according to the Orhan method [46].
Seed inoculation: CH11, SB6 and SB39 bacteria were cultured in HM medium at 28°C for 18 hours. The bacterial suspensions were centrifuged (6000 rpm/10 min) and the supernatant was discarded. The cells were washed 3 times with 20ml of sterile phosphate-buffered saline (PBS, pH 7.2) then re-suspended in sterile PBS (108 CFU/ml). The sterile seeds are soaked in the bacterial suspensions of the three isolates selected either alone or in combination two by two or all three together for 1 hour. Sterile seeds soaked in sterile PBS served as controls [47].
Growing wheat: wheat seeds are sown in plastic pots filled with sterilized or unsterilized soil, at a rate of three seeds per pot. The pots were divided into batches (9 pots per batch) and each batch was irrigated with the same volume of saline solutions of different concentrations (125, 150 and 200 Mm NaCl) on the 1st day and the 8th day. The other waterings were done with the same volume of sterile tap water (10 ml/pot) to avoid dehydration. The experiment was carried out under semi-controlled conditions with temperatures of 20/25 °C night/day and a photoperiod of 16/8 h light/dark and relative humidity of 60% for two weeks. A block randomization plan was carried out.
2.6.2. Evaluation of plant growth factors under salt stress
After 14 days of growth, the seedlings were harvested and the growth of the plants was observed. Thus, the morphological properties of the seedlings were measured under the different experimental growth conditions: the height of the aerial part, the length of the roots, the fresh weight. and dry the aerial part and the roots of the seedlings. The fresh weight was obtained just after harvest. On the other hand, the dry weight is obtained after drying in a hot oven at 45°C until the weight stabilizes.
2.7. Statistical analysis
Variance analysis one way (ANOVA) was carried out using the Statistical Package for the Social Sciences (IBM SPSS V.26, USA). The Tukey test is used to compare mean values between treatments at the 5% significance level (p = 0.05).
3. Results
3.1. Researching traits that promote plant growth
In this study, 70 halotolerant bacteria were isolated from saline soils. These bacteria have been studied for their ability to produce enzymes of agricultural interest and promoting plant growth. The preliminary test of enzymatic activities allowed the selection of six named bacterial strains (SB6, CH11, CH12, SB29, SB39, CH42) which produce cellulases, amylases, lipases, proteases, ureases and chitinases. All of these bacteria have the ability to fix atmospheric nitrogen and produce ammonia (NH3). 53.33% have the power to solubilize inorganic phosphate, and 33.33% produce hydrogen cyanide (HCN). On the other hand, only SB39 and CH42 bacteria have the power to produce small quantities of IAA at a rate of 2.031 μg/ml ± 0.01 and 1.027 μg/ml ± 0.1 respectively. (Table 1)
3.2. Molecular identification
Following the study of the characteristics promoting plant growth, three bacteria (SB6, CH11 and SB39) were selected for molecular identification and further studies.
An analysis of 16S rDNA was carried out. After PCR amplification and gene sequencing, the results showed that the SB6 and CH11 strains are closely related to Serratia sp with a similarity rate of 98.84% for SB6, and 98.98% for CH11. On the other hand, strain SB39 gave 98.98% similarity with the halophilic bacterium Halomonas sp. Still, these results only allowed an unambiguous attribution of gender.
3.3. Antifungal activity
3.3.1. Direct inhibition of mycelial growth
Figure 1.a shows the results of direct comparison on agar of the selected bacterial strains against the four phytopathogenic fungi, namely Aspergillus niger, Aspergillus flavus, Penicillium glabrum, Fusarium culmurum and Botrytis cinerea. The results show that the CH11 strain obtained the best activity of inhibition rates which vary from 51.04% to 80.67%. The SB6 strain gave an inhibition rate of 71.28% against A. niger. On the other hand, isolate SB39 recorded a maximum rate of 57.44% against A. flavus. All results show that the fungi were inhibited by isolates SB6, CH11 and SB39 but to different degrees (Figure 1b).
3.3.2. Indirect inhibition of mycelial growth
The results of the effect of the filtrates of the bacterial cultures showed the presence of an antifungal activity against A. niger for the CH11 isolate in the three environments (with and without biotic and abiotic stress) which indicates that this isolate produces antifungal molecules in a constitutive manner. As for isolate SB39, the results show that there is an inducible production of antifungal molecules, that is to say only when this bacteria is subjected to biotic stress (presence of the fungus) or abiotic (environment free of glucose). On the other hand, for isolate SB6, the zone of inhibition is very negligible (Figure 2).
3.3.3. Inhibition of mycelial growth by volatile substances
The effect of volatile substances produced by bacteria on mycelial growth was determined by calculating the percentage inhibition compared to the negative control. Volatiles reduced the mycelial growth of phytopathogenic fungi. For A. niger and P. glabrum the reduction was 71.29% and 72.22% respectively for SB39, 56.93% and 63.33% for SB6 and only 3.96% and 27.78% for isolate CH11. Concerning F. culmorum the reduction varied from 12.5% to 26.67% for the three isolates (Fig. 1c).
3.3.4. Bioassay on apples
In this experiment, apple fruits that were inoculated with the pathogens (A. niger or B. cinerea) plus the antagonist developed much smaller areas of infection compared to fruits that were inoculated with the pathogens alone (Figure 3). Thus, the severity of the disease was reduced by more than 50% in the case of A. niger and by more than 70% for B. cinerea (Figure 4).
3.3.5. Characterization of bacterial antifungal metabolites
The antifungal molecules produced by the bacteria SB6, CH11 and SB39 were identified by RP-HPLC by matching their retention times to that of the corresponding standard. The quantification of the compounds was determined by comparing their surface area to that of the corresponding standard of known concentration.
The analysis of the filtrates of the culture broths which showed positive antifungal activity allowed the identification of polyphenols produced by the bacterial isolates. The results obtained are represented in Table 2. Thus, the three bacterial isolates produce gallic acid, catechins and syringic acid. In the presence of the pathogen, the SB39 strain is capable of producing gallic acid and syringic acid with concentrations that exceed 20 mg/ml. And it is the only one that produces hydroxyquinone.
3.4. Tests to stimulate the growth of wheat plants subjected to salt stress
The results of the treatment of wheat seeds with the three bacterial isolates and their different combinations show a significant improvement in the growth parameters considered compared to those obtained with the control under salt stress (Figure 5). To evaluate the harmful effects of salt stress, different concentrations of NaCl were used (125, 150 and 200 mM). The results obtained indicate that the increase in the NaCl concentration causes a reduction in the length of stems and roots and consequently in their fresh and dry weight in control plants. However, the inoculation of the three bacterial isolates showed a significant attenuation of the harmful effects of salinity (Figure 6). Thus, the inoculation of the bacterial strains caused an increase of 30% for the length of the stems to 125 Mm with l The association of the two strains SB6 and SB39 (Serratia sp. and Halomonas sp.) and increases of 34% and 24% at 150 and 200 Mm respectively with the strain Halomonas sp. Concerning the length of the stems, the best result was obtained with the Halomonas sp. strain with an increase of 39%, 45% and 53% for the concentrations of 125 mM, 150 and 200 mM.
4. Discussion
The salinization of agricultural soils constitutes one of the major problems in agriculture which limits plant growth and agricultural productivity by up to 50% [48,49,50]. Around 1 billion hectares of the world's land surface are affected by salinization, which represents around 7% of the planet's land surface [31]. On the other hand, the use of fungicidal chemicals has a negative effect on health and the environment. Therefore, the application of plant growth-promoting rhizobacteria (PGPR) as growth-promoting and plant disease-fighting agents is a better alternative [20,51,52].
In this study, halotolerant rhizobacteria capable of promoting plant growth were isolated. Three bacterial strains were selected and then identified based on the similarity of their 16S rDNA genes. Two of them belong to the genus Serratia (SB6 and CH11) while isolate SB39 belongs to the genus Halomonas. Bacteria of the genus Serratia and Halomonas have previously been reported for their plant growth promoting and antagonistic effects against phytopathogens [53,54,55,56].
Antifungal activity was observed in all three bacterial strains. Noting that the Serratia sp strain showed better antagonistic potential by direct confrontation on agar. On the other hand, the strain Halomons sp. gave the best results for the inhibition test by production of volatile substances. Similar results were obtained by [57,58,59,60]. According to several studies, one of the mechanisms of action of soil bacteria against phytopathogens is linked to the production of biomolecules (HCN, NH3) and lytic enzymes such as cellulase, proteases, chitinases. These enzymes play an important role in degrading the cell walls of fungi [48,49,50]. The production of HCN by PGPRs played an important role in the biological control of several soil pathogenic fungi, as reported by Azeem al. [64]. And as our results show (Table 1), these bacteria produce, to different degrees, these bioactive molecules which inhibit the growth of fungi. These results are similar to the work reported by [56,60].
One of the problems of harvests is the contamination of products during storage, which leads to significant losses [42]. In our study, the application of the three bacterial strains to apples previously contaminated by phytopathogenic fungi made it possible to limit the expansion of the rot zone induced by these fungi. The results obtained show that there is a significant difference between the tests and the negative control. Furthermore, disease severity after application of the three bacterial antagonists reduced rot in apples contaminated with A. niger and B. cinerea. Similar results were reported by Sun et al. [59]and Jimtha et al. [65]with bacteria of the genus Serratia.
Mixed cultures in liquid HM medium containing the bacterial antagonist and the phytopathogen A. niger (biotic stress), or the bacteria alone in a depleted medium (abiotic stress) revealed that the production of antifungal molecules only took place in case of stress in the case of isolate SB39 (Halomonas sp.). This means that these molecules are inducible by environmental stress of the bacteria which corresponds to the results obtained by Tabli et al. (2017), who reported that the antifungal activity was only demonstrated when the Psudomonas and Serratia strains were grown in the presence of A. niger. On the other hand, for the CH11 strain, antifungal molecules are synthesized in a constitutive manner (in the presence and absence of stress).
In order to identify these antifungal molecules, HPLC was carried out. The results reveal that our three bacterial strains have the capacity to produce polyphenols. According to Skorokhod et al. [66], only plants and certain microorganisms are capable of synthesizing the precursors of phenolic compounds. Thus, Livinska et al. [67] studied the production of polyphenols by some lactic acid bacteria in different environments. This present study is the first to demonstrate the capacity of bacteria of the genus Serratia and Halomonas to produce polyphenols. Endophytes can produce secondary metabolites identical or similar to those of their host [68]. Knowing that Serratia and Halomonas can be endophytic bacteria, this could explain our results.
Enzymes of microbial origin are considered biofertilizers because they degrade organic polymers, and make essential elements such as nitrogen and phosphorus accessible to the plant [69]. In this study, we selected halotolerant and halophilic bacteria in order to test their beneficial effects on the growth of durum wheat under saline stress. Several studies have shown the beneficial effect of halotolerant and halophilic bacteria on plant growth [46,70,71]. Our studied strains demonstrated the ability to produce hydrolytic enzymes (cellulase, amylase, lipase, proteases, urease and chitinase), to solubilize phosphate, to fix nitrogen and to moderately produce the phytohormone AIA (table 1). The action of microbial enzymes intervenes in the physicochemical of the soil by improving the availability of elements essential for plant growth [72]. Thus, the simultaneous expression of different PGP traits, acting synergistically, promotes plant growth [73]. IAA is a phytohormone involved in root initiation, cell division and enlargement. Therefore, IAA-producing microorganisms increase plant growth and root length, which allows the plant to obtain more nutrients from the soil [74]. A study by Dasele et al. [75] revealed that Halomonas sp. had AIA production potential. This corresponds to the results we found where SB39 was the only isolate selected to produce IAA, despite the fact that work reports the ability of bacteria of the genus Serratia to synthesize this phytohormone [76,77]. IAA-producing rhizobacteria allow the plant to increase water permeability causing better water absorption in the leaves, thus inducing an improvement in cellular metabolism linked to environmental stress [78,79]. In addition, PGPR can fix atmospheric nitrogen essential to the metabolism of various molecules such as amino acids, enzymes, NADH, chlorophyll, etc. The production of ammonia is also important because it represents another source of nitrogen. Soluble phosphates directly support plant growth because they act as a macronutrient. It notably promotes root development and intervenes in other processes such as the ripening of flowers or fruits, the mobilization of nutrient reserves... Recently, most research has focused on the isolation and study of microorganisms. halotolerant and halophilic for their PGP potential [55,60,80]
Inoculation of halophilic/halotolerant bacteria improves plant growth under salt stress. Thus, the strains selected in this study are capable of synthesizing a large number of these metabolites, which could explain their effectiveness against target pathogenic fungi and the stimulation of wheat growth.
In the results we obtained, we note a significant improvement in the growth stimulated by bacteria under saline stress compared to the control. It is interesting to point out that the consortia gave good results especially at 200 mM NaCl. These results are perhaps due to the synthesis of phytohormones by these bacteria as well as to the improvement in the availability of mineral elements in the soil [81].
These results showed us that NaCl reduces the weight of plant shoots and roots, but that the presence of PGP reduces the harmful effects of stress caused by NaCl. The bacterial strains studied promoted plant growth under salt stress from 120 mM to 200 mM and the results are in agreement with previous studies [46,47,54,82]
5. Conclusions
Salinity is a big challenge for agricultural production. The three halotolerant and halophilic strains isolated from Serratia sp. (SB6 and CH11) and Halomonas sp. (SB39) gave interesting results on the growth of wheat under salinity stress. They have also shown their effectiveness in inhibiting the development of phytopathogenic fungi. These bacteria have the capacity to produce biofertilizers and secondary metabolites such as polyphenols. Based on these results, we can consider these bacteria as effective agents for stimulating plant growth and in biocontrol. It would be interesting to verify these potentialities in vivo and in the field, and to understand the mechanisms used by these bacteria as PGP.
Author Contributions
Conceptualization, Karim Houali and El-hafid Nabti; Methodology, Karima Ould Ouali, Karim Houali and Zahia Madani; Validation, El-hafid Nabti; Formal analysis, Juliana Melo; Investigation, Cristina Cruz; Data curation, Yasmina Benakli; Writing – original draft, Lila Ousmer; Writing – review & editing, Karima Ould Ouali; Supervision, Cristina Cruz and El-hafid Nabti
Data Availability Statement
The data is contained within the article.
Conflict of Interest
The authors declare no conflict of interest.
References
- El Hasini, S.; Iben. Halima, O.; El. Azzouzi, M.; Douaik, A.; Azim, K.; Zouahri, A. Organic and inorganic remediation of soils affected by salinity in the Sebkha of Sed El Mesjoune – Marrakech (Morocco), Soil and Tillage Research. 2019, 193, 153-160. 2019, 193, 153–160. [CrossRef]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation, Saudi Journal of Biological Sciences. 2015, 22 (2), 123-131. [CrossRef]
- Cheik, S. La salinisation des sols, un défi majeur pour la sécurité alimentaire mondiale. 2021. Project No. IC 18CT 96-0055, pp: 152-226.
- Pimentel, D.; Berger, B.; Filiberto, D.; Newton, M.; Wolfe, B.; Karabinakis, E.; Clark, S.; Poon, E.; Abbett, E.; Nandagopal, S. Water Resources: Agricultural and Environmental Issues, BioScience. 2004, 54 (10), 909–918. [CrossRef]
- Machado, R.M.A.; Serralheiro, R.P. Soil Salinity: Effect on Vegetable Crop Growth. Management Practices to Prevent and Mitigate Soil Salinization. Horticulturae. 2017, 3, 30. [CrossRef]
- Roy, S.; Chowdhury, N. Salt Stress in Plants and Amelioration Strategies: A Critical Review. Abiotic Stress in Plants. 2020, Chapter 19. [CrossRef]
- Egamberdieva, D.; Wirth, S.; Bellingrath-Kimura, S.D.; Mishra, J.; Arora, N.K. Salttolerant plant growth promoting rhizobacteria for enhancing crop productivity of saline soils. Front. Microbiol. 2019, 10, 2791. [CrossRef]
- Bhat, M.A.; Kumar, V.; Bhat, M.A.; Wani, I.A.; Dar, F.L.; Farooq, I.; Bhatti, F.; Koser, R.; Rahman, S.; Jan, A.T. Mechanistic insights of the interaction of plant growthpromoting rhizobacteria (PGPR) with plant roots toward enhancing plant productivity by alleviating salinity stress. Front. Microbiol. 2020, 11, 1952. [Google Scholar] [CrossRef]
- Qin, S.; Miao, Q.; Feng, W.W.; Wang, Y.; Zhu, X.; Xing, K.; Jiang, J.H. Biodiversity and plant growth promoting traits of culturable endophytic actinobacteria associated with Jatropha curcas L. growing in Panxi dry-hot valley soil. Appl. Soil Ecol. 2015, 93, 47–55. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; et al. Plant growth-promoting rhizobacteria: context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef]
- Wang, W.; Wu, Z.; He, Y.; Huang, Y.; Li, X.; Ye, B.C. Plant growth promotion and alleviation of salinity stress in Capsicum annuum L. by Bacillus isolated from saline soil in Xinjiang. Ecotoxicol. Environ. Saf. 2018, 164, 520–529. [Google Scholar] [CrossRef]
- Lobo, C.B.; Tomás, M.S.J.; Viruel, E.; Ferrero, M.A.; Lucca, M.E. Development of low-cost formulations of plant growth-promoting bacteria to be used as inoculants in beneficial agricultural technologies. Microbiol. Res. 2019, 219, 12–25. [Google Scholar] [CrossRef]
- Vaishnav, A.; Singh, J.; Singh, P.; Rajput, R.S.; Singh, H.B.; Sarma, B.K. Sphingobacterium sp. BHU-AV3 induces salt tolerance in tomato by enhancing antioxidant activities and energy metabolism. Front. Microbiol. 2020, 11, 443. [Google Scholar] [CrossRef]
- Li, X.; Li, B.; Cai, S.; and al. Identification of rhizospheric actinomycete Streptomyces lavendulae SPS-33 and the inhibitory effect of its volatile organic compounds against Ceratocystis Fimbriata in postharvest sweet potato (Ipomoea Batatas (L.) Lam.). Microorganisms. 2020, 8 (3), 319. [CrossRef]
- Zainab, N.; Amna, Din, B.U.; et al. Deciphering metal toxicity responses of flax (Linum usitatissimum L.) with exopolysaccharide and ACC-deaminase producing bacteria in industrially contaminated soils. Plant Physiol. Biochem. 2020, 152, 90–99. [CrossRef]
- Orozco-Mosqueda, M.D.C.; Glick, B.R.; Santoyo, G. ACC deaminase in plant growth-promoting bacteria (PGPB): an efficient mechanism to counter salt stress in crops. Microbiol. Res. 2020, 235, 126439. [Google Scholar] [CrossRef]
- El-Esawi, M.; Glascoe, A.; Engle, D.; Ritz, T.; Link, J.; Ahmad, M. Cellular metabolites modulate in vivo signaling of Arabidopsis cryptochrome-1. Plant Signal. Behav. 2015, 10(9), e1063758. [Google Scholar] [CrossRef]
- Qin, S.; Feng, W.W.; Zhang, Y.J.; Wang, T.T.; Xiong, Y.W.; Xing, K. Diversity of bacterial microbiota of coastal halophyte Limonium sinense and amelioration of salinity stress damage by symbiotic plant growth-promoting actinobacterium Glutamicibacter halophytocola KLBMP 5180. Appl. Environ. Microbiol. 2018, 84 (19) e01533-18. [CrossRef]
- Xiong, Y.W.; Gong, Y.; Li, X.W.; Chen, P.; Ju, X.Y.; Zhang, C.M.; Yuan, B.; Lv, Z.P.; Xing, K.; Qin, S. Enhancement of growth and salt tolerance of tomato seedlings by a natural halotolerant actinobacterium Glutamicibacter halophytocola KLBMP 5180 isolated from a coastal halophyte. Plant Soil. 2019, 445, 307–322. [Google Scholar] [CrossRef]
- Tabli, N.; Nabti, E.H.; Dahel, D.; Mokrane, N.; Manyani, H.; Dary, M.; Megias, M. G. Impact of Diazotrophic Bacteria on Germination and Growth of Tomato, with Bio-control Effect, Isolated from Algerian Soil.J. Eco. Heal. Env. 2014, 2: 1, 1-7. [CrossRef]
- Carrim, A. J. I.; Barbosa, E. C.; Vieira, J. D. G. Enzymatic activity of endophytic bacterial isolates of Jacaranda decurrens Cham.(Carobinha-do-campo). Brazilian Archives of Biology and Technology. 2006,49(3), 353-359. [CrossRef]
- Vinoth, R. S.; Kanikkai, R. A.; Babu, V. A.; Manoj, G. T.; Naman, H. S.; Johnson, A. J.; Infant, S. B.; Sathiyaseelan, K. Study of starch degrading bacteria from kitchen waste soil in the production of amylase by using paddy straw. Recent. res. sci. Technol. 2009, 1(1): 008–013.
- Sierra, G.A. A simple method for the detection of lypolytic activity of microorganisms and some observations on the influence of the contact between cells and fatty substrates. A. Van Leeuw. 1957, 28: 15-22.
- Kumar, S.; Karan, R.; Kapoor, S.; Singh, S. P.; & Khare, S. K. Screening and isolation of halophilic bacteria producing industrially important enzymes. Brazilian Journal of Microbiology. 2012, 43, 1595-1603.
- Christensen, W.B. Urea decomposition as a means of differentiating proteus and paracolon cultures from each other and from Salmonella and Shigella types. J. Bacteriol. 1946, 52, 461–466. [Google Scholar] [CrossRef]
- Hsu, S. C., & Lockwood, J. Powdered chitin agar as a selective medium for enumeration of actinomycetes in water and soil. Applied microbiology. 1975, 29(3), 422-426. 29.
- Kopečný, J.; Hodrová, B.; Stewart, C.S. The isolation and characterization of a rumen chitinolytic bacterium. Lett. Appl. Microbiol. 1996, 23, 195–198. [Google Scholar] [CrossRef]
- Lorck, H. Production of hydrocyanic acid by bacteria. Physiol. Plant. 1948, 1, 142–146. [Google Scholar] [CrossRef]
- Bric, J.M., Bostock, R.M.; Silverston, S.E. Rapid in situ assay for indole acetic acid production by bacteria immobilization on a nitrocellulose membrane. Appl. Environ. Microbiol. 1991, 57, 535–538. 1991; 57, 535–538.
- Fokkema, N.J. Fungal antagonism in the phylosphere. Ann. Appl. Biol. 1978, 89, 115–117. [Google Scholar] [CrossRef]
- Sagahón, I.P.; Reyes, M.A.A.; Rojas, H.V.S.; Cuenca, A.A.; Jurado, A.T.; Alvarez, I.O.C.; Flores, Y.M. Isolation of bacteria with antifungal activity against the phytopathogenic fungi Stenocarpella maydis and Stenocarpella macrospora. Int. J. Mol. Sci. 2011, 12, 5522–5537. [Google Scholar] [CrossRef] [PubMed]
- Korsten, L.; De Jager, E.S. Mode of action of Bacillus subtilis for control of avocado postharvest pathogens. S. Afr. Avocado Growers Assoc. Yearb. 1995, 18, 124–130. [Google Scholar] [CrossRef]
- Lee, J.Y.; Hwang, B.K. Diversity of antifungal Actinomycetes in various vegetative soils of Korea. Can. J. Microbiol. 2002, 48, 407–417. [Google Scholar] [CrossRef]
- Li, Q. ; Ning, P.; Zheng, L.; Huang, J.; Li, G.; Hsiang, T. Fumigant activity of volatiles of Streptomyces globisporus JK-1 against Penicillium italicum on Citrus microcarpa. Postharvest Biol. Technol. 2010,58:157–165. [CrossRef]
- Aghighi, S.; Shahidi Bonjar, G.H.; Saadoun, I.; Rawashdeh, R.; Batayneh, S. First report of antifungal spectra of activity of Iranian actinomycetes strains against Alternaria solani, Alternaria alternata, Fusarium solani, Phytophthora megasperma, Verticillium dahliae and Saccharomyces cerevisiae. Asian J. Plant Sci. 2004, 3, 463–471. [Google Scholar] [CrossRef]
- Mahdieh, S.; Gholam, H.S.B.; Gholam, R.S.S. Post-harvest biological control of apple bitter rot by soil-borne Actinomycetes and molecular identification of the active antagonist. Postharvest Biol. Technol. 2016, 112, 46–54. [Google Scholar]
- Dennis, C.; Webster, J. Antagonistic properties of species groups of Trichoderma, II. Production of volatile antibiotics. Trans. Br. Mycol. Soc. 1971, 57, 363–369. [Google Scholar] [CrossRef]
- Jorjandi, M.; Shahidi Bonjar, G.H.; Baghizadeh, A.; Sharifi Sirchi, G.R.; Massumi, H.; Baniasadi, F.; Aghighi, S.; Rashid Farokhi, P. Biocontrol of Botrytis allii Munn the causal agent of neck rot, the post-harvest disease in onion, by use of a new Iranian Iisolate of Streptomyces. Am. J. Agric. Biol. Sci. 2009, 4(1), 72–78. [Google Scholar] [CrossRef]
- Kim, D.O.; Jeong, S.W.; Lee, C.Y. Antioxidant capacity of phenolic phytochem-icals from various cultivars from pulms. Food Chem. 2002, 81, 321–326. [Google Scholar] [CrossRef]
- N’Guessan, A.H.O.; Dago Déliko, C.E.; Akhanovna, J.; Békro, M.; Békro, Y.A. Teneurs en composés phénoliques de 10 plantes médicinales employées dansla tradithérapie de l’hypertension artérielle, une pathologie émergente en Côted’Ivoire. Génie industriel. 2011, 6, 55–61. [Google Scholar]
- Zhao, Y.; Wang, Y.; Jiang, Z-T. ; Li, R. Screening and evaluation of active compounds in polyphenol mixtures by HPLC coupled with chemical methodology and its application, Food Chemistry. 2017, 227, 187-193. [CrossRef]
- Xiao, C.L.; and Kim, Y.K. Postharvest fruit rots in apples caused by Botrytis cinerea, Phacidiopycnis washingtonensis, and Sphaeropsis pyriputrescens. Online. Plant Health Progress. 2008, 9(1), 34. [Google Scholar] [CrossRef]
- Lu, L.; Lu, H.; Wu, C.; Fang, W.; Yu, C.; Ye, C.; & al. Rhodosporidium paludigenum induces resistance and defense-related responses against Penicillium digitatum in citrus fruit. Postharvest Biol. Technol. 2013, 85, 196–202. [CrossRef]
- Huang, Y.; Sun, C.C.; Guan, X.N.; Lian, S.; Li, B.; Wang, C. Butylated hydroxytoluene induced resistance against Botryosphaeria dothidea in apple fruit. Front. Microbiol. 2021, 11, 599062. [Google Scholar] [CrossRef]
- Zihao, S.; Baihui H.; Cuicui, W.; Shiyu, L.; Yuxin, X.; Baohua, L.; and Caixia, W. Biocontrol features of Pseudomonas syringae B-1 against Botryosphaeria dothidea in apple fruit. Front. Microbiol. 2023, 11 : 599062. [CrossRef]
- Orhan, F. Alleviation of salt stress by halotolerant and halophilic plant growth-promoting bacteria in wheat (Triticum aestivum), Braz. J. Microbiol. 2016, 47, 621–627. [Google Scholar] [CrossRef]
- Bensidhoum, L.; Nabti, E.H.; Tabli, N.; Kupferschmied, P.; Weiss, A.; Rothballer, M.; Schmid, M.; Keel, C.; Hartmann, A. Heavy metal tolerant Pseudomonas protegens isolates from agricultural well water in northeastern Algeria with plant growth promoting, insecticidal and antifungal activities, European Journal of Soil Biology. 2016, 75, 38-46. [CrossRef]
- Vimal, S.R.; Singh, J.S.; Arora, N.K.; Singh, D.P. PGPR-an effective bioagent in stress agriculture management. In: Sarma, B.K., Jain, A. (Eds.), Microbial Empowerment in Agriculture: A Key to Sustainability and Crop Productivity. Biotech Books, New Delhi), 2016, 1–108.
- Farahmand, N.; Sadeghi, V. Estimating soil salinity in the dried lake bed of urmia lake using optical sentinel-2 images and nonlinear regression models. J. Indian Soc. Rem. Sens. 2020, 48 (4), 675–687. April 2020. [CrossRef]
- Li, H.; La, S.; Zhang, X.; Gao, L.; Tian, Y. Salt-induced recruitment of specific root-associated bacterial consortium capable of enhancing plant adaptability to salt stress. ISME J. 2021, 15, 2865–2882. [Google Scholar] [CrossRef]
- Djebaili, R.; Marika, P.; Claudia, E.; Beatrice, F.; Mahmoud, K.; and Maddalena, D.G. "Biocontrol of Soil-Borne Pathogens of Solanum lycopersicum L. and Daucus carota L. by Plant Growth-Promoting Actinomycetes: In Vitro and In Planta Antagonistic Activity" Pathogens. 2021, 10, 1305. [Google Scholar] [PubMed]
- Noumavo, P.A.; Agbodjato, N.A.; Baba-Moussa, F.; Adjanohoun, A.; Baba-Moussa, L. Plant growth promoting rhizobacteria: Beneficial effects for healthy and sustainable agriculture. Afr. J. Biotechnol. 2016, 15(27):1452-1463.
- Tabli, N.; Rai, A.; Bensidhouma, L.; Palmierib, G.; Gogliettinob, M.; Coccab, E.; Consigliob, C.; Cilloc, F.; Bubicic, G. ; Nabti, El-H. Plant growth promoting and inducible antifungal activities of irrigation well water-bacteria. J. Biocontrol. 2017, 10.010.
- Desoky, E.S.M.; Saad, A M.; El-Saadony, M.T.; Merwad, A.R.M.; Rady, M.M. Plant growth-promoting rhizobacteria: Potential improvement in antioxidant defense system and suppression of oxidative stress for alleviating salinity stress in Triticum aestivum (L.) plants. j.bcab. 2020, 30, 1878-8181. [CrossRef]
- Oliva, G.; Di Stasio, L.; Vigliotta, G.; Guarino, F.; Cicatelli, A.; Castiglione, S. Exploring the potential of four novel halotolerant bacterial strains as plant-growth-promoting rhizobacteria (PGPR) under saline conditions. Appl. Sci. 2023, 13, 4320. [Google Scholar] [CrossRef]
- Mukherjee, P.; Mitra, A.; Roy, M. Halomonas rhizobacteria of avicennia marina of indian sundarbans promote rice growth under saline and heavy metal stresses through exopolysaccharide production. Front. Microbiol. 2019, 10:1207. [CrossRef]
- Alijani, Z.; Amini, J.; Karimi, K.; Pertot, I. Characterization of the Mechanism of action of Serratia rubidaea Mar61-01 against Botrytis cinerea in Strawberries. Plants. 2023, 12, 154. [Google Scholar] [CrossRef]
- Kamou, N.N.; Dubey, M.; Tzelepis, G.; Menexes, G.; Papadakis, E.N.; Karlsson, M.; agopodi, A.L.; Jensen, D.F. Investigating the compatibility of the biocontrol agent Clonostachys rosea IK726 with prodigiosin-producing Serratia rubidaea S55 and phenazineproducing Pseudomonas chlororaphis ToZa7. Arch. Microbiol. 2016, 198, 369–377. [Google Scholar] [CrossRef]
- Sun, M.; Liu, J.; Li, J.; Huang, Y. Endophytic Bacterium Serratia plymuthica From Chinese Leek Suppressed Apple Ring Rot on Postharvest Apple Fruit. Front. Microbiol. 2022, 12:802887. [CrossRef]
- Cherif, H.; Sghaier, I.; Hassen, W.H.; Amara, C.; Masmoudi, A.S.; Cherif, A.; Neifar, M. Halomonas desertis G11, Pseudomonas rhizophila S211 and Oceanobacillus iheyensis E9 as biological control agents against wheat fungal pathogens: PGPB consorcia optimization through mixture design and response surface analysis. Int Clin Pathol J. 2022, 9(1):20‒28.
- Fillinger, S.; Elad, Y. Botrytis - The Fungus, the Pathogen and Its Management in Agricultural Systems; Springer International Publishing: Cham, Switzerland. 2016, 488.
- Xun, F.; Xie, B.; Liu, S.; Guo, C. Effect of plant growth-promoting bacteria (PGPR) and arbuscular mycorrhizal fungi (AMF) inoculation on oats in saline-alkali soil contaminated by petroleum to enhance phytoremediation. Environ. Sci. Pollut. Res. 2015, 22, 598–608. [Google Scholar] [CrossRef]
- Trejo-López, J.A.; Rangel-Vargas, E.; Gómez-Aldapa, C.A.; Villagómez-Ibarra, J.R.; Falfán-Cortes, R.N.; Acevedo-Sandoval, O.A.; Castro-Rosas, J. Isolation and Molecular Identification of Serratia Strains Producing Chitinases, Glucanases, Cellulases, and Prodigiosin and Determination of Their Antifungal Effect against Colletotrichum siamense and Alternaria alternata In Vitro and on Mango Fruit. Int. J. Plant Biol. 2022, 13, 281–297. [Google Scholar] [CrossRef]
- Azeem, S.; Agha, S.I.; Jamil, N.; Tabassum, B.; Ahmed, S.; Raheem, A.; Jahan, N.; Ali, N.; Khan, A. Characterization and survival of broad-spectrum biocontrol agents against phytopathogenic fungi. Rev Argent Microbiol. 2022, 54(3), 233–242. [Google Scholar] [CrossRef]
- Jimtha, J.C.; Jishma, P.; Sreelekha, S.; Chithra, S.; Radhakrishnan, E. K. Antifungal properties of prodigiosin producing rhizospheric Serratia sp. Rhizosphere. 2017, 3, 105–108. [Google Scholar] [CrossRef]
- Skorokhod, I.; &Kurdysh, I. The low-molecular weight antioxidants of microorganisms.Mikrobiology, 2014, 76: 48–59.
- Livinska, O.; Ivaschenko, O.; Garmasheva, I.; Kovalenko, N. The screening of lactic acid bacteria with antioxidant properties. Microbiology. 2016, 2(4), 447–459. [Google Scholar] [CrossRef]
- Puri, S.C.; Verma, V.; Amna, T.; Qazi, G.N.; Spiteller, M. An endophytic fungus fromNothapodytes foetida that produces camptothecin. Journal of Natural Products. 2005, 68:1717–9.
- Garcia-Fraile, P.; Menéndez, E.; Rivas, R. Role of bacterial biofertilizers in agriculture and forestery. AIMS Bioengeneering. 2015, 2, 183–205. [Google Scholar] [CrossRef]
- Qurashi, A.W.; Sabri, A.N. Osmolyte accumulation inmoderately halophilic bacteria improves salt tolerance ofchickpea. Pak J Bot. 2013, 45:1011–1016.30.
- Essghaier, B.; Dhieb, C.; Rebib, H.; Ayari, S.; Boudabous, A.R.A.; Sadfi-Zouaoui, N. Antimicrobial behavior of intracellularproteins from two moderately halophilic bacteria: strain J31 of Terribacillus halophilus and strain M3-23 of Virgibacillusmarismortui. J Plant Pathol Microb. 2014, 5, 214. [Google Scholar] [CrossRef]
- Rai, A.; Bensidhoum, L.; Tabli, N.; Bouaoud, Y.; Naili, F.; Cruz, C.; Nabti, E. A Pseudomonas Protegens with high antifungal activity protects apple fruits against Botrytis Cinerea Gray Mold. Int. J. Sci. Res. Sci. Tech. 2016, 2, 227–237.
- Masmoudi, F.; Abdelmaleka, N.; Tounsia, S.; Dunlapb, C.A.; Triguia, M. Abiotic stress resistance, plant growth promotion and antifungal potential of halotolerant bacteria from a Tunisian solar saltern. Microbiological Research. 2019, 229, 126331. [Google Scholar] [CrossRef]
- Boiero, L.; Perrig, D.; Masciarelli, O.; Penna, C.; Cassan, F.; Luna, V. Phytohormone production by three strains of Bradyrhizobiumjaponicum and possible physiological and technologicalimplications. Appl Microbiol Biotechnol. 2007,74:874–880.
- Desale, P.; Patel, B.; Singh, S.; Malhotra, A.; Nawani, N. Plantgrowth promoting properties of Halobacillus sp. and Halomonas sp. in presence of salinity and heavy metals. JBasic Microbiol. 2014, 54: 781–791.
- Duca, D.R. & Glick, B.R. Indole-3-acetic acid biosynthesis and its regulation in plant-associated bacteria. Applied microbiology and biotechnology. 2020,104, 8607-8619. [CrossRef]
- Singh, R. P.; & Jha, P. N. The multifarious PGPR Serratia marcescens CDP-13 augments induced systemic resistance and enhanced salinity tolerance of wheat (Triticum aestivum L.). PLos one. 2016, 11(6), e0155026. [CrossRef]
- Etesami, H.; Maheshwari, D.K. Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicol. Environ. Saf. 2018, 156, 225–246. [Google Scholar] [CrossRef]
- Ha-Tran, D.M.; Nguyen, T.T.M.; Hung, S.-H.; Huang, E.; Huang, C.-C. Roles of Plant Growth-Promoting Rhizobacteria (PGPR) in stimulating salinity stress defense in plants: A review. Int. J. Mol. Sci. 2021, 22, 3154. [Google Scholar] [CrossRef]
- Alijani, Z.; Amini, J.; Ashengroph, M. et al. Antifungal Activity of Serratia rubidaea Mar61-01 Purified Prodigiosin Against Colletotrichum nymphaeae, the Causal Agent of Strawberry Anthracnose. J Plant Growth Regul. 2022, 41, 585–595. [CrossRef]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.L.; Touraine, B.; Moënne-Loccoz, Y.; Muller, D.; Prigent-Combaret, C. Plant growth-promoting rhizobacteria and root system functioning. Frontiers in plant science. 2013, 4, 356. [Google Scholar] [CrossRef]
- Nawaz, A.; Shahbaz, M.; Asadullah, Imran, A.; Marghoob, M.U.; Imtiaz, M.; Mubeen, F. Potential of Salt Tolerant PGPR in Growth and Yield Augmentation of Wheat (Triticum aestivum L.) Under Saline Conditions. Front. Microbiol. 2020, 11:2019.
Figure 1.
(a) Effect of isolates SB6, CH11 and SB39 on mycelial growth ; (b) Percentage growth inhibition (PGI %) of phyopathogenic fungi by isolates (plate confrontation tests) ; (c) Effect of volatiles substances produced by the three strains on growth inhibition (PGI%).
Figure 1.
(a) Effect of isolates SB6, CH11 and SB39 on mycelial growth ; (b) Percentage growth inhibition (PGI %) of phyopathogenic fungi by isolates (plate confrontation tests) ; (c) Effect of volatiles substances produced by the three strains on growth inhibition (PGI%).

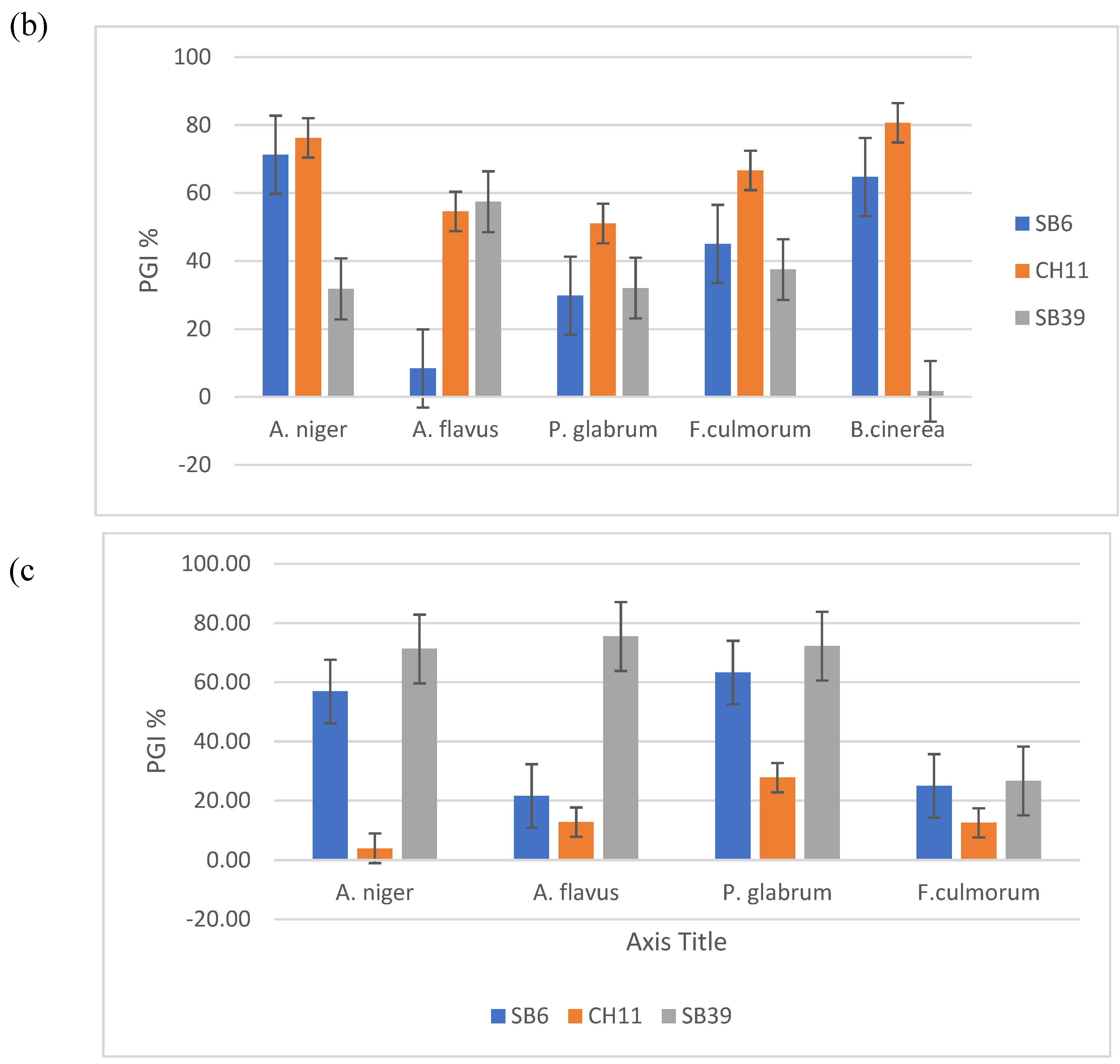
Figure 2.
Effect of culture broth filtrate on mycelial growth (a. SB6 ; b. CH11 ; c. SB39).
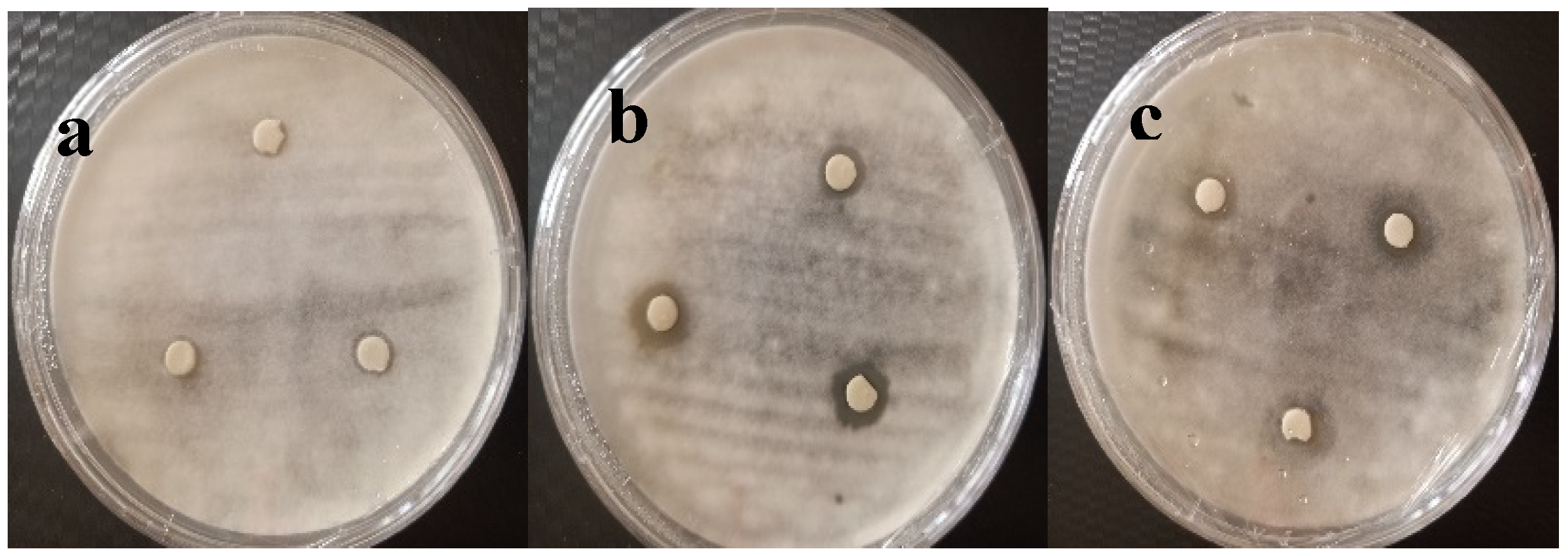
Figure 3.
Effect of fungal mold on apple fruit treated (b) or not (a) with the bacterial antagonist after 6 days of incubation.
Figure 3.
Effect of fungal mold on apple fruit treated (b) or not (a) with the bacterial antagonist after 6 days of incubation.
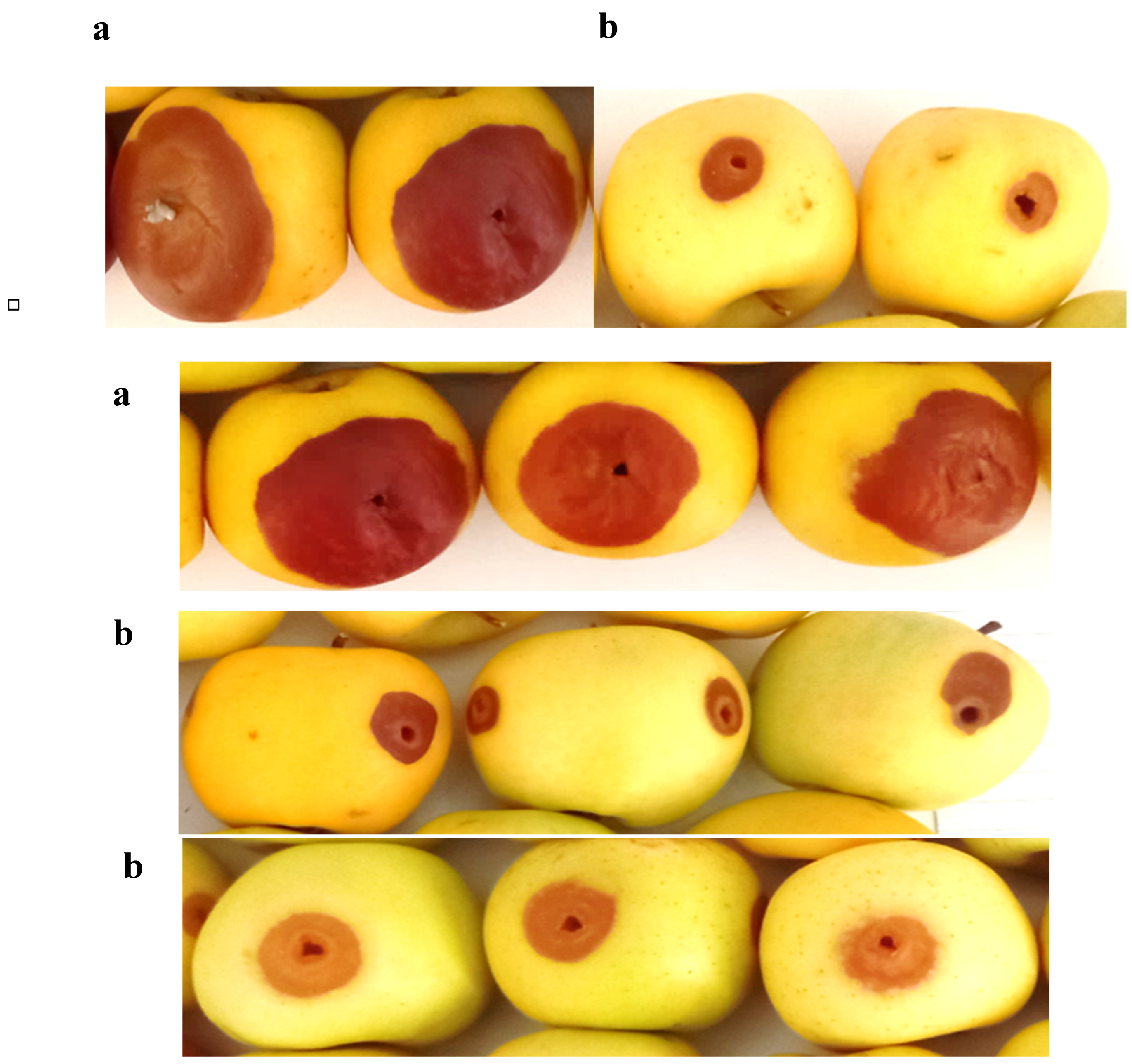
Figure 4.
Efficiency of bacterial isolates (SB6, CH11 and SB39) against A. niger and B. cinerea in apple fruit.
Figure 4.
Efficiency of bacterial isolates (SB6, CH11 and SB39) against A. niger and B. cinerea in apple fruit.
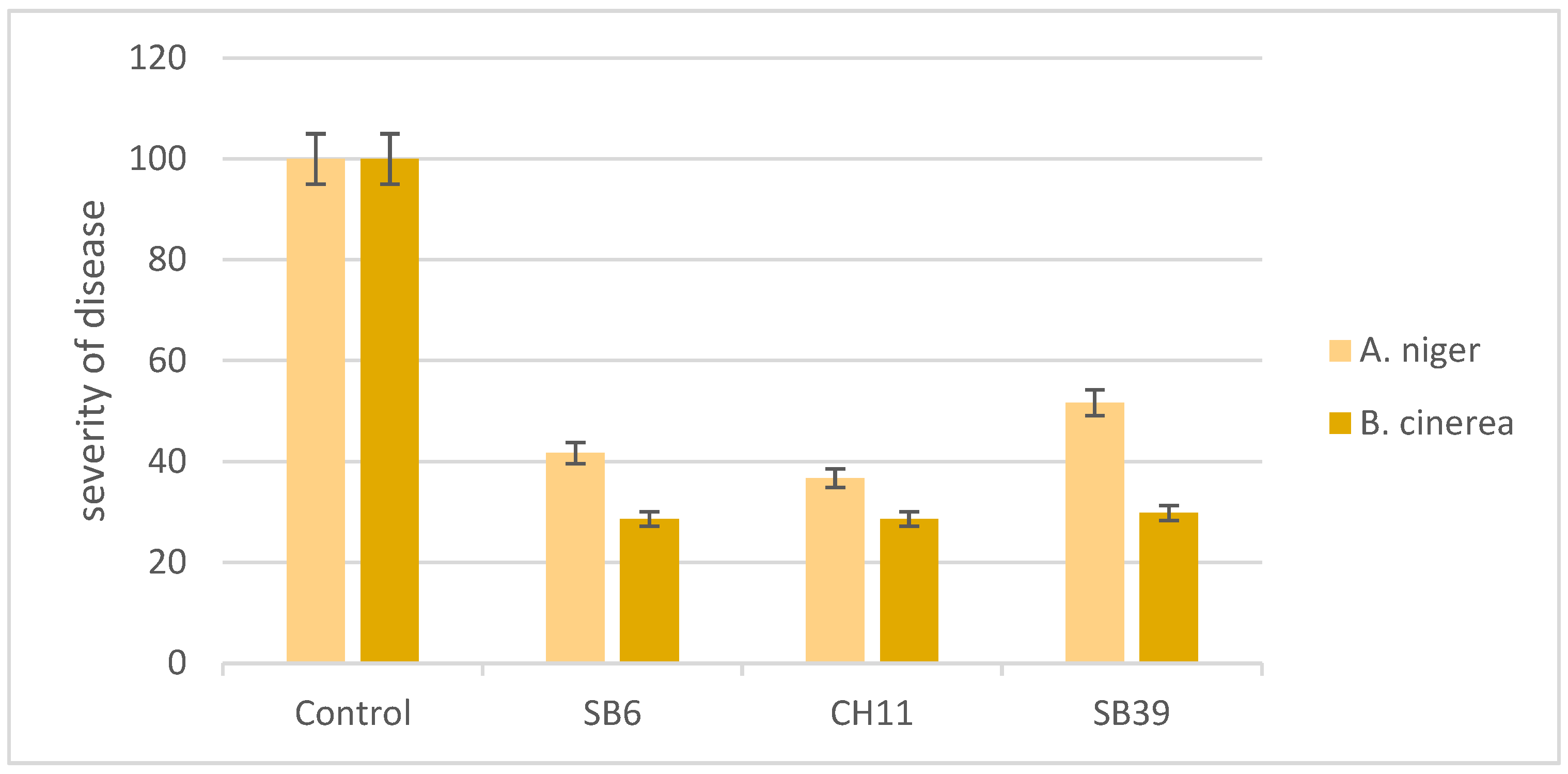
Figure 5.
Effects of bacterial strains treatment on the growth of wheat seedlings under salt stress conditions (125 mM) 1) Control ; 2) SB6 ; 3) CH11 ; 4) SB39 ; 5) SB6-SB39 ; 6) SB6-CH11 ; 7) CH11-SB39 ; 8) SB6-CH11-SB39.
Figure 5.
Effects of bacterial strains treatment on the growth of wheat seedlings under salt stress conditions (125 mM) 1) Control ; 2) SB6 ; 3) CH11 ; 4) SB39 ; 5) SB6-SB39 ; 6) SB6-CH11 ; 7) CH11-SB39 ; 8) SB6-CH11-SB39.

Figure 6.
The effect of inoculation with halotolerant rhizobacteria (SB6 : Serratia sp; CH11 : Serratia sp; SB39 : Halomonas sp) ongrowth (a : shoot lenght; b : root lenght ; c : shoot fresh weight; d :root fresh weight; e :shoot dry weight; f :root dry weight) of wheatplants grown under salt stress of 125mM, 150 or 200 mM of NaCl. The bars on the top of the columns indicate standard error of the mean.
Figure 6.
The effect of inoculation with halotolerant rhizobacteria (SB6 : Serratia sp; CH11 : Serratia sp; SB39 : Halomonas sp) ongrowth (a : shoot lenght; b : root lenght ; c : shoot fresh weight; d :root fresh weight; e :shoot dry weight; f :root dry weight) of wheatplants grown under salt stress of 125mM, 150 or 200 mM of NaCl. The bars on the top of the columns indicate standard error of the mean.
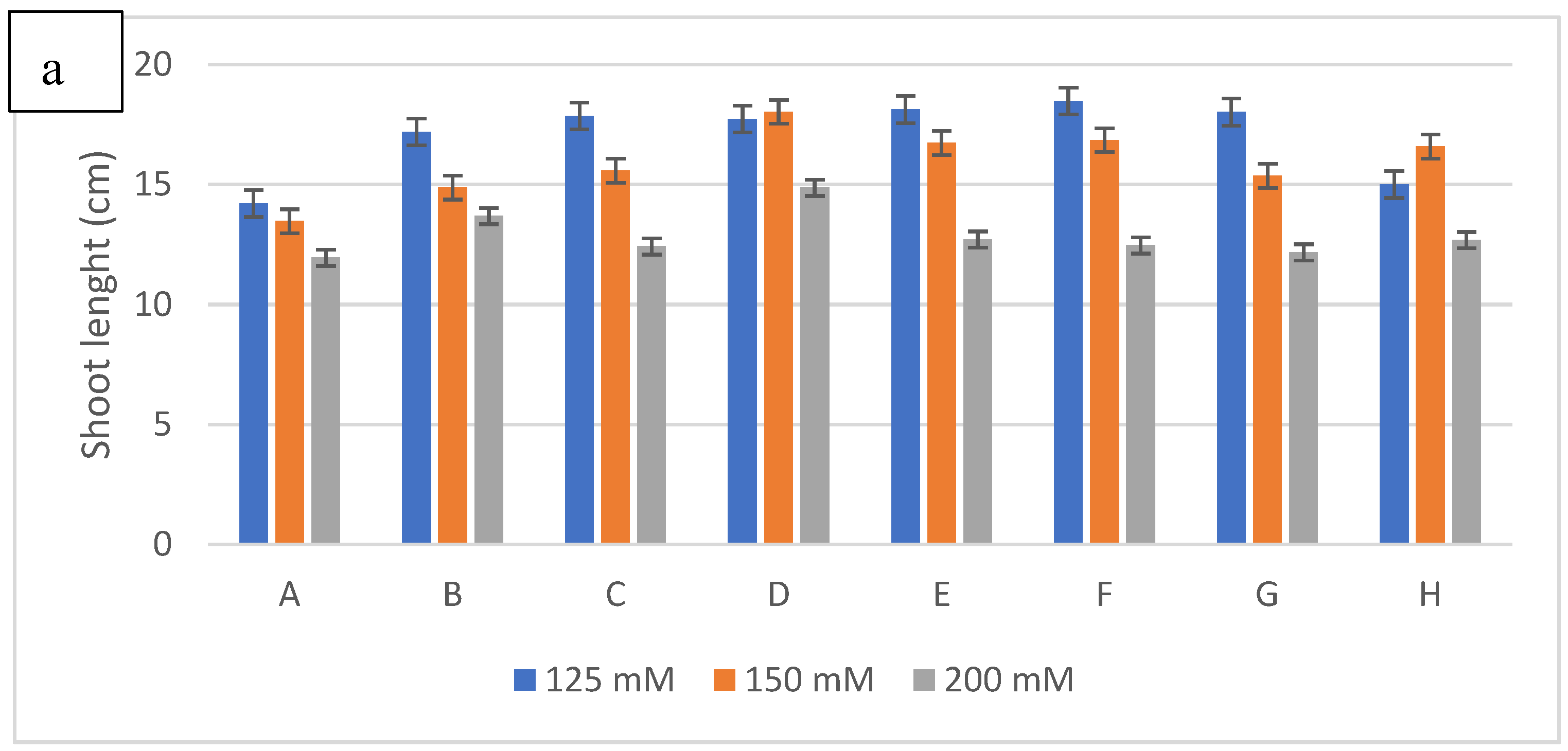
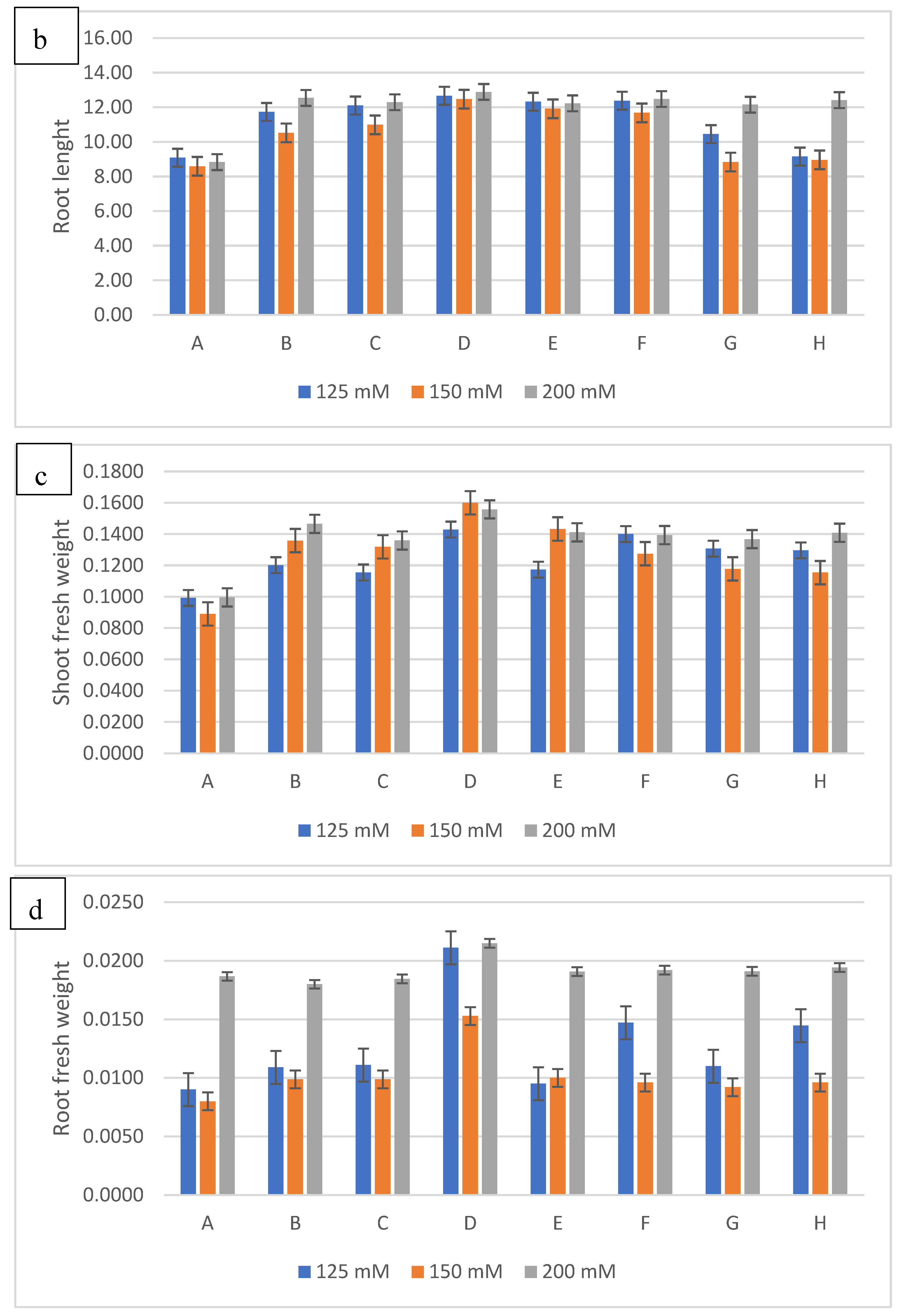
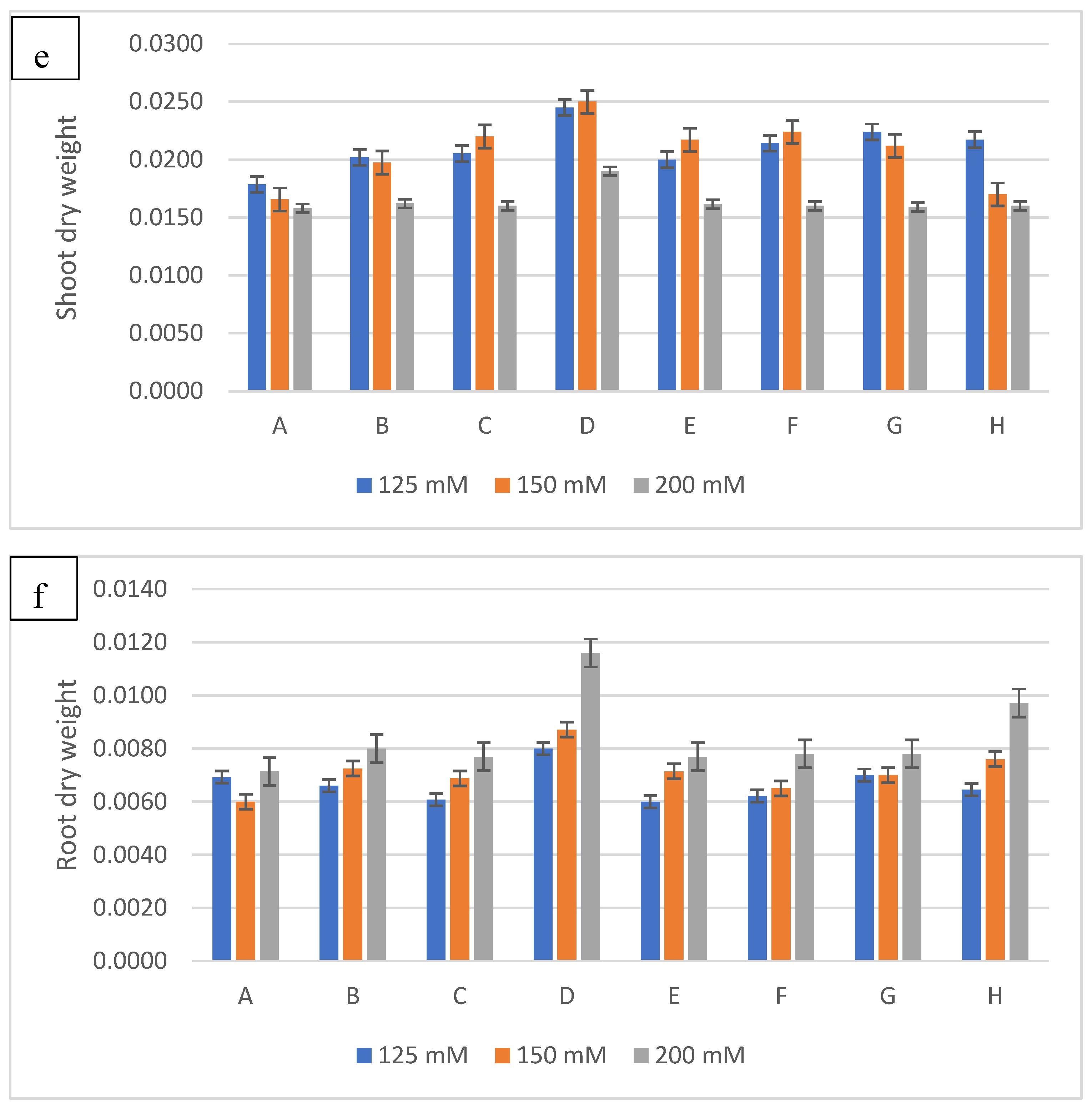

Table 1.
Plant growth promoting traits of selected isolates.
| SB6 | CH11 | CH12 | SB29 | SB39 | CH42 | |
| Cellulase Amylase Lipase Protease Urease Chitinase Nitrogen fixation Phosphate solubilization IAA (µg/mL) NH3 HCN |
+ +++ - +++ - +++ + ++ - ++ + |
+ - ++ +++ - +++ + ++ - + - |
++ - - +++ + +++ + - - + - |
- - ++ +++ - +++ ++ + - ++ - |
+ +++ - +++ ++ ++ ++ - +++ +++ |
+ - + - ++ ++ ++ - ++ - |
+++: high activity ++ : intermediate activity + : low activity - : no activity.
Table 2.
Phenolic compounds identified in the filtrate culture broth and their concentrations.
| RRT(min) | Coumpound name | Broth filtrate (mg/ml) | |||||
| SB6 - | CH11 - | CH11 + | CH11 dep | SB39 + | SB39 dep | ||
| 3,15 ± 0,05 | Gallic acid | 13,77 | 20,79 | 12,76 | 13,43 | 21,56 | 9,53 |
| 3,61 ± 0,00 | Hydroxy quinone | - | - | - | - | 6,31 | - |
| 5,47 ± 0,23 | Resorcinol | 2,24 | 2,82 | 2,29 | 2,88 | 5,38 | - |
| 6,10 ± 0,17 | Catechin | 3,81 | 4,84 | 4,27 | 1,29 | 5,17 | 3,50 |
| 6,23 ± 0,41 | 1,2-hydroxy benzen | - | 8,16 | 3,13 | 3,55 | - | - |
| 7,21 ± 0,05 | Syringic acid | 16,65 | 13,24 | 18,54 | 11,72 | 22,88 | 12,23 |
| 10,63 ± 0,00 | Naringenin-7-glucoside | - | - | 4,56 | - | - | - |
| 11,08 ± 0,00 | 3,4,5-Trimethoxy benzoic acid | 2,51 | - | - | - | - | - |
| 11,36 ± 0,01 | m-anisic acid | - | 1,82 | 1,01 | 2,39 | - | - |
| 15,07 ±0,01 | Hesperidine | 9,69 | 1,91 | - | - | - | - |
(-) : in the absence of the phytopathogen ; (+) :in the presence of the phytopathogen ; (dep) :in a depleted culture medium.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



