
Submitted:
01 February 2024
Posted:
02 February 2024
You are already at the latest version
Abstract
The normal ageing process significantly affects resistance arteries, leading to various biological and physiological consequences. Systolic hypertension is a common occurrence in ageing individuals, accompanied by increased large artery stiffening, heightened pulsatility, small artery remodeling, and damage to critical microvascular structures. Starting from young adulthood, there is a progressive elevation in mean arterial pressure, supported by clinical and epidemiological evidence as well as findings from animal models. The myogenic response, a protective mechanism for the microcirculation, may face disruptions during ageing. Dysregulation of calcium entry channels (T-type, L-type, TRP channels), dysfunction in intracellular calcium storage and extrusion mechanisms, altered expression of potassium channels, and a change in smooth muscle calcium sensitization may contribute to age-related dysregulation of myogenic tone. Flow-mediated vasodilatation, a hallmark of endothelial function, is compromised in ageing. This result of endothelial dysfunction is related to oxidative stress, lower nitric oxide bioavailability and a low-grade inflammatory response, further exacerbating vascular dysfunction. Resistance artery remodeling in ageing emerges as a hypertrophic response of the vessel wall, often seen in conjunction with outward remodeling (in normotension), but can also present as inward hypertrophic remodeling (in hypertension). The remodeling process involves oxidative stress, inflammation, reorganization of extracellular matrix fiber proteins and actin cytoskeletal components. Reactive oxygen species (ROS) signaling and chronic low-grade inflammation, often referred to as "inflammaging”, play substantial roles in age-related vascular dysfunction. Due to its role in regulation of vascular tone as well as structural proteins, the RhoA/Rho-kinase pathway becomes a new target in age-related vascular dysfunction and associated diseases. Understanding the intricate interplay of these factors is crucial for developing targeted interventions to mitigate the consequences of ageing on resistance arteries and enhance overall vascular health.
Keywords:
aging
; resistance artery
; myogenic tone
; flow-mediated vasodilation
; structural remodeling
; hypertension
; cardiovascular disease
; neurodegenerative disease
; proteomics
Introduction
Aging is associated with an increase in the occurrence of arterial hypertension and organ blood flow dysregulation, which are major risk factors for cardiovascular diseases, renal failure, retinopathy, stroke, and cognitive impairment in both sexes. Active diameter regulation (vasoconstriction or vasodilatation, remodeling) of resistance arteries (small arteries and arterioles) plays a paramount role in the control of mean arterial blood pressure (MAP) and organ blood flow. Thus, organ blood flow regulation is achieved as an interplay between neurohormonal regulation, metabolic regulation, myogenic tone/myogenic response (basal tone vs. autoregulation), flow-mediated vasodilation (+ other endothelium-dependent dilating mechanisms), structural remodeling, and control of capillarity (angiogenesis and rarefaction). In the present paper, the discussion of resistance artery ageing will be limited to the roles of myogenic tone, flow-mediated vasodilation, and structural remodeling. Proteomics and bioinformatics will also be discussed since it provides a promising approach to obtain new insight into vascular ageing. The emphasis is on normal “healthy aging” in resistance arteries, but it is inevitable to also provide examples of hypertension and other diseases arising during ageing. The cited papers were selected based on consistent use of a simple search string on PubMed containing the relevant terms incl. ageing, vascular, myogenic tone OR flow-mediated vasodilation OR structural remodeling. In some cases the search was narrowed down by including the term resistance artery and/or the organ of interest (e.g., brain or cerebral). The cited information from relevant papers was selected based on clear presentation of data concerning a direct comparison of relevant age groups in terms of the different vascular functions being discussed. Thus, papers were omitted if they merely presented their data as “age-related” functions or diseases without a direct comparison between age groups. A thorough understanding of the molecular mechanisms involved in dysregulation of vascular tone and remodeling in ageing will provide a better basis for developing new targets and treatments for vascular complications involving dysregulation of organ blood flow in the elderly population. Moreover, as sexual dimorphism in these responses may arise during the course of ageing, future studies will need to take this into careful consideration.
Systolic hypertension in ageing
Arterial stiffening evolving in most humans during the ageing process causes isolated systolic hypertension, which may progress to systolic-diastolic hypertension through endothelial dysfunction, enhanced contractility, and structural remodeling in resistance vessels [1]. The isolated systolic hypertension seen with advanced age is distinct from the midlife hypertension with elevated diastolic and systolic blood pressures, which are due to increased total peripheral resistance [2]. The increased pulsatility in large stiff arteries is augmented by an early systolic arrival of the reflected pressure wave due to increased impedance in the microcirculation [1,3]. As the pressure wave is transmitted longer distally in the aging systemic circulation, the microcirculation may be compromised. This is observed especially in renal glomeruli, blood-brain barrier, coronary microcirculation and in retinal capillaries, which are the site of common complications of age-related increases in large artery pulsatility and systolic hypertension [4-6]. Arterial stiffness in ageing is related to changes in extracellular matrix (ECM) properties with loss of elasticity and remodeling of the elastin, collagen, and fibronectin content of the arterial wall [4,7]. Molecular determinants of large artery stiffness in ageing may thus be ECM proteins governing collagen breakdown and production, such as matrix metalloproteinases (MMPs), tissue inhibitors of MMPs (TIMPs), transforming growth factor-β1 (TGF-β1), and transglutaminases (TGMs). In addition, proteins responsible for elastin or elastic fiber expression, coherence, and functional characteristics, such as fibrillins and fibulins [8,9], may be involved in age-related arterial stiffness. However, increasing attention is given to smooth muscle phenotypic changes related to various stressors, such as mechano-transduction, oxidative stress, genetic and epigenetic factors [10-12]. It is beyond the scope of this review to provide a complete list of the possible molecular determinants of large artery stiffness in ageing.
Increased Mean Arterial Pressure in ageing
The Framingham large population-based clinical study tracking hemodynamic variables from young adulthood to old age showed that mean arterial pressure (MAP) in both men and women increases over the lifetime from early adulthood to ~60 years of age, and stays high albeit with a tendency to a decline towards old age [13,14]. When exploring sex-dependent differences at the population level in 4 community-based US cohort studies, it became evident that MAP (unadjusted or adjusted for multi-variables) is higher in men than in women throughout their lifetime, but the elevation in MAP from baseline at young adulthood is significantly higher in females than in males [15]. An advantage of using mice for vascular ageing studies is that mice age rather fast, i.e., reaching middle age at 10-14 months and old age at 18-24 months of age [16,17]. Rats were defined as middle-aged at 12-18 months, and old age at 22-30 months [16,18]. Another important point is that normal laboratory C57BL6 wildtype mice at 12 months of age do present with age-dependent hypertension [19]. However, it should be noted that the tendency to develop hypertension with ageing must be associated with a high degree of inter-individual variability in mice and rats. In another study, male wildtype laboratory mice displayed increased MAP at 14 and 18 months of age compared to 3 and 5 months, whereas female mice only were hypertensive at 18 months of age [20]. A later study from the same authors showed that estrogen-replacement therapy in aged female mice lowered MAP significantly to pre-hypertensive levels, but this was dependent on simultaneous increase in renal Angiotensin AT2-Receptor expression [21]. The molecular determinants of age-dependent increases in MAP in humans have not been clarified, but are a subject of intense investigation due to the large clinical advantage of targeting human hypertension specifically in the elderly, whilst taking into account the differences between males vs. females.
Myogenic response protects the microcirculation, but may be disrupted in ageing
The myogenic response is a smooth muscle-derived, mechano-activated vasoconstriction or vasodilatation to an increase or decrease of the intravascular pressure, respectively. This response is important for establishing basal (myogenic) tone (MT) in resistance vessels, and for blood flow autoregulation in the brain, kidney and most other organs [22,23]. In human pial arteries of different calipers, the level of myogenic tone was not dependent on the age of the donor [24]. In contrast, the level of myogenic tone in human posterior ciliary arteries of the eye was inversely proportional to the age of the subject [25]. As the information about age-dependent changes in myogenic tone in human resistance vessels is scarce, we have to turn to data from rodent models. In mouse middle cerebral arteries subjected to a pulsatile pressure ex vivo, myogenic tone is significantly reduced at old age [26], and non-significantly reduced under static pressure at old age [26-28]. During Ang-II induced hypertension, myogenic tone was dramatically reduced in old mice compared to young hypertensive mice [27,28]. In rat middle cerebral arteries, myogenic tone was increased at old age in endothelium-intact preparations, but was significantly decreased in endothelium-denuded preparations from old rats [29]. In endothelium-intact mouse parenchymal arterioles, myogenic tone was increased at old age, whereas no age-dependent effect was observed using endothelium-denuded arterioles [30]. In a study elucidating sex-dependent effects in rat middle cerebral arteries, myogenic tone was increased in old females compared to young females but reduced in old males compared to young males [31]. However, these divergent age-dependent effects were only significant at low pressures (40-60 mm Hg), which is at the lower end of or below the autoregulatory range of blood pressure in the brain. These data from the cerebral vascular bed suggest that the endothelium releases a contractile factor (e.g., Endothelin-1, Thromboxane A2 or Superoxide) that enhances spontaneous vascular tone at old age, whereas “classical” endothelium-independent myogenic tone derived by mechanical activation of smooth muscle constriction is in general reduced at old age, possibly in a sex-dependent manner. In mice, renal autoregulation was impaired in the kidney of old mice, due to impairments in myogenic response and pressure-induced calcium increase in afferent arterioles [32]. In addition, the impaired autoregulation could be explained by a loss of the tubuloglomerular feedback response and down-regulation of A1-type adenosine receptors in afferent arterioles [32]. In skeletal muscle arterioles of rat soleus and gastrocnemius muscles, myogenic tone was reduced at old age [33-35]. In mouse superior epigastric arteries, however, myogenic tone was not different between young and old mice [36]. In coronary arterioles from Fischer 344 rats, myogenic constriction was reduced at old age in endothelium-intact preparations, but not in endothelium-denuded vessels [37]. Myogenic tone in 3rd order mesenteric arteries from control rats was decreased at middle age (18 months) vs. at young age (3 months) [38]. In mouse 2nd- 3rd order mesenteric arteries, we previously found that myogenic tone was increased in mature adult mice (6-7 months) vs. young mice (2-3 months), but in contrast myogenic tone was markedly decreased in middle aged mice (13-14 months) compared to mature adult [39]. The disruption of myogenic tone in small mesenteric arteries from middle-aged mice is supported by our own unpublished data as well as a previous study showing a gradual age-dependent loss of myogenic tone in five age groups from 2 to 12 months old mice [40]. In the latter study, no sex-dependent differences in MT were observed. Thus, in 13 out of 15 studies, myogenic tone was decreased in middle aged or old mice and rats compared to young. In cerebral and coronary vessels, the endothelium seems to play a modulating role to alter myogenic tone in aging, and in cerebral resistance arteries, there may be a sex-dependent effect of aging. To summarize, most if not all vascular beds display a progressive loss or deficiency in myogenic tone at middle age and old age. The disruption of myogenic reactivity may have deleterious effects on the integrity of the microcirculation and the organ function at middle age and old age. When the basal tone is lost or weak, the vessels can no longer dilate and increase the perfusion to vasodilatory stimuli (i.e., reduced flow reserve), causing a mismatch in demand and supply. Probably even more important, the autoregulation of blood flow is disrupted in critical organs, such that they will face too little supply of oxygen and nutrients when the arterial pressure is low, whereas the capillaries will face mechanical disruption under high arterial pressures, due to excessive microcirculatory flow. This might trigger serious diseases such as stroke, neurodegenerative diseases, cardiac ischemia, renal failure, and retinopathy just to mention a few.
Molecular determinants of reduced myogenic tone in aging?
What are then the molecular determinants of age-dependent alterations in myogenic tone in resistance vessels? In fact only limited and dispersed information is available, making this subject an important area for future investigations. It is well known that Ca2+ entry channels (L-type, T-type and TRP channels) as well as intracellular Ca2+ handling mechanisms (Ca2+ store release and extrusion) in vascular smooth muscle are dysregulated during aging. [16,18]
Voltage-gated calcium channels
In small mesenteric arteries from middle-aged mice, two successive studies from the same authors showed that smooth muscle mineralocorticoid receptor (MR) is the driver of expression of a single micro-RNA, miR-155, which negatively regulates expression of L-type channel (CaV1.2) and AT1-receptor expression [41,42]. In middle-aged mice a decreased expression of miR-155 was responsible for an increase in L-type channel expression and activity as well as increased AT1-R expression and vasoconstriction [42]. This model serves to explain the enhanced vasoconstrictor responses to Ang-II and hypertension in middle-aged mice, but cannot account for the general reduction in myogenic tone with aging. Other studies have shown a decrease in L-type channel expression and function in cerebral arteries from old mice [43] and in mesenteric arteries from middle-aged rats [44]. Interestingly the reduced L-type channel expression in rat mesenteric arteries was mirrored by increased expression of miR-328, which negatively regulates L-type channel expression [44]. Yet other studies failed to find an age-dependent difference in L-type channel expression and function in small coronary arteries from old rats [45], or in small mesenteric arteries from mature adult [46] and old [47] mice. Due to the lack of consistency between the studies, changes in L-type channel expression and activity cannot be the primary molecular determinant for decreased myogenic tone with aging. CaV3.1 T-type channels, which are responsible for myogenic tone in the low pressure range from 40-80 mm Hg [48], are dramatically downregulated by aging [46]. However, the precise role of CaV3.1 channels in the age-dependent reduction of myogenic tone, if any, remains to be determined. The CaV3.2 T-type channel localized in smooth muscle plasma membrane caveolae suppresses myogenic tone in small mesenteric arteries from young mice by activating the SR Ca2+ release channel RYR2 and nearby BKCa K+ channels [18,46,49]. Although the CaV3.2 expression was unaffected by aging, we found that this protective role of the CaV3.2 channels against excessive myogenic tone was absent in mature adult and middle-aged mice, suggesting a pathophysiological role in age-related vascular dysfunction [46]. Subsequently, it was found that smooth muscle plasma membrane caveoale are also disrupted by aging, which could explain the loss of the CaV3.2 channel-mediated activation of the RYR2/BKCa axis and suppression of myogenic tone in middle-aged mice [50]. A loss of BKCa-mediated negative feedback on myogenic tone in aging might also be due to a weaker physical coupling between SR/RYR2 complex and BKCa channels, for example via reduced junctophilin/microtubular coupling between SR and the plasma membrane with less peripheral location of SR in aged arteries [18]. Finally, administration for 4 months of the senolytic cocktail Dasatinib + Quercetin was able to rescue the CaV3.2/RYR2/BKCa-mediated suppression of myogenic tone in middle-aged mice [51]. Nevertheless, the loss of the CaV3.2/SR/RYR2/BKCa axis by aging does not serve to explain the general age-dependent decrease in myogenic tone seen across most studies.
TRP channels
The transient receptor potential cation channel subfamily C member 6 (TRPC6) is involved in myogenic tone development in rat cerebellar and posterior cerebral arteries [52]. We showed that pressure-dependent activation of Phospholipase A2 activity and 20-HETE production causes an increase in agonist-activated TRPC6 currents, and this was critically involved in the myogenic response in rat small mesenteric arteries preconstricted with low concentration of α1-agonist and neuropeptide Y [53]. Comparing Ang-II induced hypertension in young vs. old mice, Toth and colleagues observed an increased TRPC6 expression in middle cerebral arteries of young hypertensive vs. normotensive mice, however, this compensatory upregulation of TRPC6 channel expression was not seen in aged hypertensive mice [28]. Myogenic tone and pressure-induced increase of smooth muscle intracellular [Ca2+] in cerebral arteries were increased by Ang-II hypertension in young mice, but were decreased by more than 50% in old Ang-II hypertensive mice [27,28]. These studies suggest that the loss of a compensatory TRPC6 channel upregulation in RAS-induced hypertension during ageing could be involved in the mechanism for the disruption of myogenic tone.
Intracellular calcium handling proteins
Due to a preserved peak magnitude in caffeine/phenylephrine-induced SR Ca2+ store release but a reduced plateau phase Ca2+ signal in old vs. mature adult murine mesenteric artery myocytes, it was suggested that “storage-operated” non-selective cation channels (TRPC; Orai channels) might be involved in age-related disruption of myogenic tone [47]. In mesenteric artery myocytes from old mice, a reduction in expression of several intracellular Ca2+ handling proteins (CaV1.2, RYR2, SERCA2, PLB, STIM1) caused a global decrease in the amplitude of Ca2+ signals, whereas the duration was increased due to a reduction in the rate of Ca2+ store refilling [43]. This might also be a contributing factor to the reduced myogenic tone in aged arteries. Mice with a missense mutation in Collagen IV alpha 1 (Col4a1), associated with small vessel disease-like brain pathology, had reduced myogenic tone and pressure-induced depolarization at middle age. In contrast, young mice with the mutation, and middle-aged control mice without the mutation, had normal myogenic tone [54]. Furthermore, the middle-aged Col4a1 mutant mice had disrupted SR Ca2+ release and reduced IP3-receptor mediated, Ca2+-dependent TRPM4 channel activation, which can explain the disruption of pressure-induced membrane depolarization and myogenic tone development in the middle-aged mutant mice [54]. Thus, disruption of SR Ca2+ store release may be involved in the reduced myogenic tone development in patients with cerebral small vessels disease and/or vascular dementia.
Potassium channels
BKCa channel expression and function were decreased in coronary arteries from old rats and from old human subjects [55]. Likewise, BKCa channel expression in soleus muscle arterioles was decreased in old rats [34]. However, the BKCa channel blocker iberiotoxin did not eliminate the age-dependent reduction in myogenic tone in cerebral arteries from aged hypertensive mice [27]. The coupling between CaV3.2 T-type channels and BKCa mediated negative feedback on myogenic tone was lost in small mesenteric arteries from middle-aged mice [46]. The KV channel blocker 4-aminopyridine increased the myogenic tone from a reduced level in aged rat soleus and gastrocnemius muscle arterioles to the same level as in young arterioles [34]. A recent study suggested that Kir2.x channels are upregulated in smooth muscle cells of cerebral parenchymal arterioles in old mice [30]. Whereas the above effects involving decreased BKCa channel expression and function cannot explain the disruption of myogenic tone in ageing, an increased activity or expression of smooth muscle KV and Kir2.x channels can. However, more data on the function and expression of KV and Kir2.x channels in resistance vessels with myogenic tone will be necessary to elucidate their role in aging.
RhoA/Rho-kinase pathway
In addition to the Ca2+ signaling pathways, we aimed to investigate the possible role of altered Ca2+ sensitivity of smooth muscle contraction in aging. Phosphorylation of the regulatory subunit of Myosin Light Chain is a balance between the activities of the Ca2+/Calmodulin-activated Myosin Light Chain Kinase and the Myosin Light Chain Phosphatase, the latter being negatively regulated by phosphorylation via Rho-kinase. Initially, we found that the expression of Rho-kinase 2 (ROCK2) was increased in mature adult mice compared to young mice, and that the ROCK2 inhibitor KD025 elicited stronger inhibition of myogenic tone in the mature adult mice [39]. The increased myogenic tone in mature adult mice was paralleled by our observation of increased noradrenaline-induced tone in this age group [46]. However, these data could only serve to explain the modest increase in myogenic tone in mature adult mice, whereas we needed more data to explain the decline in myogenic tone at middle age and old age. This prompted us to investigate the proteomic effects of aging by comparing middle cerebral arteries (MCA) and 2nd-3rd order mesenteric arteries (MRA) from 8 young (3 months old) vs. 7 middle aged (14 months old) mice using mass spectrometry-based proteomics [56]. In total 207 proteins were significantly differentially expressed and/or hierarchically clustered in an age-dependent manner. Out of 1833 proteins that were uniquely identified and quantified in the mass spec analyses of all four groups of vessels (young MCA, middle-aged MCA, young MRA, middle-aged MRA), proteins that were affected by aging thus corresponds to ~11% of the identified and quantified proteome [56]. In our KEGG analysis of biological pathway representations, we discovered that the Regulation of actin cytoskeleton pathway was one of the top significantly enriched pathways. Central in this pathway is the RhoA/Rho-kinase pathway, confirming our initial hypothesis that this pathway was important for age-dependent changes in vascular function. Several proteins connected with this pathway was on our list of age-dependent proteome changes: ARHGEF7, PAK2, ROCK1, M-RIP, ML12B, MYO5B, ACTB [56]. Work is underway to try to understand if any of these changes are underlying the changes in myogenic tone with aging, but in fact, down-regulation of ROCK1 and ML12B could explain this alone.
Wnt3a of the canonical Wnt pathway is activated during advanced arterial aging and is associated with vascular injury [57,58]. Furthermore, the non-canonical Wnt pathway member Wnt5a, capable of activating RhoA, is involved in excess vasoconstriction and salt-sensitive hypertension in middle-aged mice, a phenotype that was rescued by supplementation with the anti-aging factor Klotho [59]. Interestingly, Wnt10a of the canonical Wnt/β-catenin pathway was significantly upregulated in the proteome of middle cerebral and small mesenteric arteries from middle-aged mice [56]. There is a paucity of data concerning the role of Wnt signaling in myogenic tone, but mechanical stretch in human bronchi caused an increase in expression of a large number of genes in the Wnt-mediated signaling pathway [60]. It is tempting to speculate that some components or mediators of the canonical or non-canonical Wnt pathways might be involved in the disruption of myogenic tone in aging.
Flow-mediated vasodilatation—indicator of endothelial (dys-)function in ageing
Flow-mediated vasodilation (FMVD) is an endothelium-dependent, shear stress-activated vasodilatation found in both large and small arteries [46,61,62]. The FMVD responses are regarded as the hallmark of endothelial function, which is directly correlated with cardiovascular health [63]. Non-invasive FMVD measurements in humans are routinely measured on brachial arteries using the post-occlusion increase in blood flow to calculate the percent increase in FMVD [61,64]. In children and adolescents, there was a trend to an age-dependent decline in FMVD, but there were no significant sex-differences [65]. Conversely, FMVD in large conduit arteries is markedly reduced in old subjects compared to young adults [66-71]. In a large, community-based study, FMVD in both sexes was negatively correlated with age with men having lower FMVD than women, but women having a steeper age-dependent decline in FMVD with age [72]. In brachial arteries from young and aged otherwise healthy subjects, FMVD correlated negatively with the arterial endothelial expression of ROS-derived nitrotyrosine [69]. In the aged subjects the expression of mitochondrial ROS-inducing NAD(P)H oxidase and the inflammation-induced transcription factor NFκB was increased in the arteries, pointing to a role of oxidative stress and vascular low-grade inflammation in the reduced FMVD in aging [69]. Supplementation with MitoQ to quench mitochondrial Superoxide production improved the FMVD in brachial artery in old (60-79 years) otherwise healthy subjects [73]. In human coronary arterioles, there was an age-dependent transition in the mediator of FMVD from prostacyclin (PGI2) in children to NO in young (18-55 years) and older subjects (>55 years) without coronary artery disease, whereas in older patients with coronary artery disease the remaining FMVD was mediated via mitochondrial H2O2 production [74].
Whereas in humans FMVD is primarily a measure of large (brachial) artery endothelial function, in rodent models FMVD can be measured in resistance arteries and arterioles using ex vivo pressure myography. FMVD measured using ultrasonography in femoral arteries in mice was significantly reduced in old compared to young mice, showing that it is possible to assess FMVD in vivo in murine conduit arteries [75]. In old male rats, FMVD measured ex vivo in soleus muscle arterioles was reduced by ~50% in old male compared to young rats, and this difference was eliminated after partial blockade of NO production using the nitric oxide synthase (NOS) inhibitor L-NAME [76]. FMVD in mouse femoral arteries was reduced significantly in old mice, but this decline was rescued by activating endothelial P2Y1-receptors using 2-Me-ATP [77]. In rat coronary arterioles, aging reduced FMVD and flow-induced NO and H2O2 production, which were linked with an increased Superoxide production in coronary arterioles from old rats [78]. In old female rats, estrogen replacement therapy could improve age-dependent loss of NO bioavailability and FMVD in coronary arterioles [79]. Overall, there was a gradual shift from FMVD being dependent on NO production at young age towards being dependent on the vasodilator H2O2 at old age in rat coronary arterioles [78,79]. Treatment with stromal vascular fraction cells or adipose-derived stem cells was able to reverse oxidative stress and rescue the age-dependent decline in FMVD in coronary arterioles from aged female rats [80].
In small mesenteric arteries from middle-aged rats, FMVD was reduced compared to young rats, and this age-dependent decline was improved by TNFα blockade [81]. There was a shift from a flow-induced rise in NO production in young rats to a decreased NO production with an increased NAD(P)H oxidase-derived H2O2 production in middle-aged rats. [82] In small mesenteric arteries from middle-aged rats, there were no sex-dependent differences in maximal total FMVD, but there was a shift from NO-mediated to EDHF-mediated vasodilation in middle-aged female rats [83]. In small mesenteric arteries from old rats, FMVD was reduced compared to young rats [84]. In old rats, the decrease in shear stress-induced release of NO in mesenteric resistance vessels was caused by an increased production of ROS, a reduced activity of SuperOxide Dismutase (SOD), and an impaired shear stress-induced activation of eNOS [85]. In mouse small mesenteric arteries, FMVD was reduced by ~50% in young mice deficient in the T-type Ca2+ channel CaV3.2, whereas no difference was observed in mature adult and middle-aged CaV3.2KO vs. wildtype mice, suggesting the loss of a protective role of endothelial CaV3.2 channels in aging [46].
To summarize the data on FMVD and ageing, all studies from human and rodents and from all vascular beds so far examined (comprising large conduit arteries as well as resistance vessels) point to a gradual impairment of FMVD from middle age to old age. An abundance of the studies point to a role of oxidative stress and low-grade inflammatory response in the age-dependent decline in FMVD, involving a reduction in NO bioavailability and a partial compensation by increased role of EDHF, such as H2O2. The molecular determinants are thus likely to include changes in expression and function of NO, H2O2, Superoxide, eNOS, NAD(P)H oxidase, SOD, catalase, TNFα, and/or NFκB. Deficiency of protective mechanisms such as estrogen in females, and endothelial CaV3.2 T-type channels or P2Y1-receptors may also play a role.
Resistance artery remodeling—a hypertrophic response to ageing
Structural remodeling of resistance arteries, causing either an inward or outward change of the maximal lumen diameter and a hyper- or hypotrophic change in the vascular wall material, is a chronic adaptation to long-term changes in pressure and/or flow as well as disease states [86,87]. The relevant structural/dimensional parameters to consider in vascular remodeling in fully relaxed (passive) vessels are: Lumen diameter; media:lumen-ratio (alternatively wall/lumen-ratio); cross-sectional area of vessel wall, distensibility and/or incremental elastic modulus (slope or β-value of stress/strain curve). In terms of clinical relevance, an increased wall/lumen-ratio has been shown to be an independent risk factor for serious cardiovascular events in humans [88,89]. To provide a short overview, the typical remodeling patterns are as follows: 1) a persistent increase in intraluminal pressure (i.e., hypertension) will cause an inward remodeling without hypertrophia (inward eutrophic remodeling with increased media:lumen-ratio) to normalize wall stress. 2) Persistent increase in intraluminal flow (i.e., normal growth, pregnancy, physical activity) will cause an outward hypertrophic remodeling, to normalize shear stress. With no change in media:lumen-ratio, this remodeling response is healthy, but if the media:lumen-ratio is increased, it is associated with increased risk of cardiovascular disease. 3) A persistent stimulation in vascular growth factors will cause a hypertrophic growth in wall material (e.g., under obesity, diabetes). These vessels will tend to remodel outward to normalize flow and shear stress (hence outward hypertrophic remodeling with no change in media:lumen-ratio), but in case of coexisting hypertension or increased stimulation of local vasoconstriction, an inward hypertrophic remodeling is seen (with a large increase in media:lumen-ratio). 4) Persistent anti-hypertensive treatment with a vasodilator will cause an outward remodeling. Since ageing causes long-term changes in blood pressure, myogenic tone, flow-mediated vasodilation, and possibly in other vasomotor responses, it can be expected that ageing will also elicit structural remodeling of resistance vessels.
In aging a consistent enlargement of the aortic media:lumen-ratio (M/L-ratio) is noted in humans and rodent models [90-92]. In human renal arteries, the media layer showed outward hypertrophic remodeling, whereas the intima layer showed inward hypertrophic remodeling with aging [93]. Expression of Matrix Metallo-Proteinase 2 (MMP2) and MMP9 are increased in an age-dependent manner in aortae of rats, non-human primates, and humans [94-96]. MMP2 activation leads to proteolysis of extracellular matrix proteins, such as collagen and elastin, thus enabling the remodeling process. However, in old rat aortae MMP2 also increases expression of the growth factor TGFβ1 and activates TGFBR2-receptor signaling, which leads to an increase in fibronectin and collagen expression, and promotes fibrosis [97]. Plasma levels of MMP9 were increased in old male and female mice [98]. Vascular stiffness and pulse wave velocity correlated significantly with plasma MMP9 levels in patients with isolated systolic hypertension, suggesting MMP9 as an age-related plasma biomarker [99,100]. In a rat model of isolated systolic hypertension, arterial stiffening was associated with the aortic activity of transglutaminase 2 (TGM2) [101]. TGM2 is secreted into the interstitial space and functions to crosslink extracellular matrix proteins, such as collagen, to complete the remodeling process [87].
In human subcutaneous resistance arteries, there is a positive correlation between M/L-ratio and age in both normotensive and hypertensive subjects irrespective of sex, but with a steeper age-dependent increase in the hypertensives [4,102]. In coronary arterioles from old rats, the wall:lumen-ratio (W/L-ratio) was increased while the elastic modulus and vessel stiffness were decreased [103]. In old rats, feed arteries and arterioles from soleus and gastrocnemius muscles displayed outward hypertrophic remodeling with no change in W/L-ratio [104]. In basilar arteries from old rats, an increased W/L-ratio but no significant change of lumen diameter was observed, showing age-dependent hypertrophic remodeling in basilar arteries [105,106]. In mature (11 month) rats, posterior cerebral arteries displayed an inward hypertrophic remodeling with no change in distensibility compared to young (3 month) rats [107]. In 5 different intracranial arteries (BA, ACA, MCA, ICA, PCA), outward remodeling was observed in old mice [108]. Likewise in old mice, posterior cerebral arteries and parenchymal arterioles showed outward remodeling but no change in wall cross-sectional area (CSA), and consequently a decrease of the W/L-ratio [109]. In pudential arteries from old Spontaneously Hypertensive Rats (SHR) with erectile dysfunction, luminal diameter was decreased while cross-sectional area and W/L-ratio were increased compared to young SHR [110]. The latter study indicated inward hypertrophic remodeling instead of outward, likely due to the existence of hypertension in the SHR model. In uterine arteries from old mice, outward hypertrophic remodeling and increased vessel stiffness were observed [111]. Mesenteric small arteries from old (>8 years) vs. young (1.5–2 years) sheep showed age-dependent outward hypertrophic remodeling with no change in M/L-ratio or blood pressure [112]. In middle-aged rats, 1st-order mesenteric arteries showed an increased CSA indicating hypertrophic remodeling [113]. In old rats, mesenteric arteries displayed outward hypertrophic remodeling with an increased thickness of the media layer and unchanged adventitial layer. Moreover, the number of smooth muscle cells in the media layer was unchanged [114]. The hypothesis that the vascular smooth muscle cell size is increased in ageing is supported by another study, in which the cell capacitance and cell circumference of mesenteric artery myocytes were increased in old mice [47]. The flow-induced outward remodeling in small mesenteric arteries was absent in old rats compared to young, but the hypertrophic remodeling in old rats was similar under normal flow and high-flow conditions [115]. These data indicate an increase in W/L-ratio under high-flow conditions in old rats. Moreover, mesenteric arteries from old rats released more Superoxide than those from young rats, while the old arteries also showed a higher expression of Superoxide Dismutase [115]. With no change of eNOS expression in old rats, these data suggest ROS-induced decrease in NO bioavailability as the mechanism for the lack of flow-induced outward remodeling in old rats [115]. In mature adult and middle-aged wildtype and CaV3.2-KO mice, small mesenteric arteries had increased CSA, indicating that the hypertrophic remodeling was not dependent on CaV3.2 T-type Ca2+ channels [46]. In middle-aged mice, mesenteric small arteries displayed outward hypertrophic remodeling, lowered distensibility and increased elastic modulus (β-value) vs. young mice, indicating age-dependent structural remodeling and increased vessel stiffness [116]. In the same study, middle-aged mice deficient in Caveolin-1 showed outward hypotrophic remodeling and lowered distensibility, suggesting that caveolae and Cav-1 are required for the hypertrophic remodeling response but not the increased stiffness [116]. The lack of flow-induced outward remodeling in small mesenteric arteries from old mice was restored by selective agonism of the Angiotensin AT2-receptor [117]. In the mouse model of transgenic familial Alzheimer’s Disease (3xFAD), middle-aged female mice, but not male, showed outward hypertrophic remodeling with increased M/L-ratio mice in small mesenteric arteries compared to non-transgenic control mice [118]. This could be related to the fact that females are usually more susceptible to Alzheimer’s Disease than males. In old female rats, but not in male rats, the flow-induced outward remodeling response in mesenteric small arteries was preserved [119]. The latter two studies also point to a role of female sex hormones in structural remodeling, but more data is needed to unravel the sex-dependency of the remodeling response in aging.
To summarize the reported diversity of remodeling responses in aging, nearly all studies show a hypertrophic remodeling response to aging. Some studies did not measure passive lumen diameter, and in such case it is impossible to evaluate the inward/outward direction of remodeling. However, most of the remaining studies suggest that outward hypertrophic remodeling is the most common occurrence in aging, with the alternative possibility that high blood pressure or other comorbidities may cause an inward hypertrophic remodeling response. A majority of studies reporting the distensibility or elastic modulus point to an increased stiffness of resistance arteries with aging, however, there are also studies reporting the opposite. Sex-dependent changes are also observed in aged arteries, and it will be important to unravel the exact occurrence and mechanisms of these changes in future studies.
Potential molecular mechanisms in age-dependent structural remodeling
From the studies above, it is clear that reduced NO signaling/bioavailability and increased oxidative stress is a key component in age-dependent remodeling responses. The oxidative stress activates gelatinases (MMP2; MMP9) that digest ECM fibers and activate TGFβ1 signaling, which via TGFBR2 receptors promotes growth and/or fibrosis in aging. Transglutaminases (TGM2) may be involved in crosslinking of the degraded protein fibers. In addition, caveolae containing Cav-1 seem to play a role in age-dependent remodeling, which is not surprising given the many receptors and signaling molecules that are concentrated in caveolae. Estrogen receptors and a reduced estrogen production associated with menopause likely plays a role in the sex-dependency of remodeling seen with aging. The involvement of receptors for angiotensin II, aldosterone, prostaglandins, thromboxane, estrogen and endothelin-1 in age-dependent structural remodeling and arterial stiffening has been extensively reviewed elsewhere [1,120-123]. It is also clear that pro-inflammatory signaling and so-called “sterile” inflammation plays an important role in the age-associated vascular remodeling process [1].
Thrombospondin-1 (TSP-1) is involved in structural remodeling in large and small arteries [124,125]. Physiological levels of TSP-1 increased Superoxide production and decreased NO-dependent vasodilation more in coronary arterioles from old female rats compared to young females [126]. The vascular remodeling index after hindlimb ischemia was significantly lower in old compared to young wildtype mice. However, in TSP-1 deficient mice, the remodeling index during ischemia was increased in both young and old mice, and the age-dependent difference was eliminated [127]. These data confirm a role for TSP-1 to suppress physiologically important remodeling responses in aging. In an investigation of age-dependent changes in the proteome of murine and human left ventricular cardiomyocytes, lactadherin (MFGE8) protein was significantly increased by aging [128]. In coronary arteries, lactadherin accumulation was observed in the interstitial space between endothelial and smooth muscle cells [128]. Coronary artery endothelial cells cultured on immobilized lactadherin showed increased expression of the pro-inflammatory cytokine IL6, and activation of age-related signaling cascades by stimulating the activities of GSK3B, MAPK, and MTOR and decreasing the activity of CDK [128].
Our proteomic investigation of age-dependent changes of the proteome in middle cerebral arteries and small mesenteric arteries in middle-aged mice [56] showed significant up- or down-regulation of the following ECM or cytoskeletal structural components in aged arteries: MFAP2, LAMA3, FBLN4, and TBB1. Furthermore, the proteins ZCHC3 and KV6AB involved in immune system activation or signaling were significantly upregulated, and the oxidases AOXA and COX5B were downregulated in aged arteries [56]. KEGG biological pathways with potential relevance to vascular structure and remodeling, and showing significant enrichment in our proteomics study, were: Vitamin B6 metabolism, Biosynthesis of antibiotics, Regulation of actin cytoskeleton, Endocytosis, Focal adhesion, and ECM-receptor interaction [56]. These data highlight the potential usefulness of mass spec-based proteomics in elucidating known as well as hitherto unknown mechanisms and pathways involved in age-dependent changes in resistance arterial structure and function.
Protein-Protein interactions and pathway analysis of age-dependent mechanisms
All proteins mentioned in this review with a potential age-dependent role are listed in Table 1. Based on this list, pathway enrichment analyses are shown in Table 2 (KEGG biological pathways) and Table 3 (REACTOME pathways). A protein-protein interaction network (STRING database) is shown in Figure 1. Furthermore, the STRING analysis yielded a list of predicted functional partners of the proteins in the network, as shown in Table 4.
The list of age-dependent proteins in Table 1 should serve as inspiration for future work regarding the role of specific proteins in the age-dependent deteriorations in resistance artery function, specifically in myogenic tone, flow-mediated vasodilation, and structural remodeling. Most frequent among the top 15 enriched KEGG pathways are unsurprisingly cellular signaling pathways, such as calcium signaling, cGMP-PKG signaling, Relaxin signaling, Advanced Glycation Endproducts (AGE) and receptor for AGE (RAGE) signaling, Focal adhesion pathway, and Oxytocin signaling. Second most enriched KEGG pathways are brain disease pathways, where Neurodegenerative diseases in general and Alzheimer’s Disease pathways appear, stressing the fact that aging is the biggest risk factor for neurodegenerative disorders. Other disease pathways are cancer-related, diabetic complications, dilated cardiomyopathy, and a hormonal disorder (Cushing’s Syndrome). Among the top 15 REACTOME pathways enriched, 7 of them are cellular signaling pathways, and 6 denote physiological functions (contraction, homeostasis, conduction, hemostasis). Overall, cellular signaling pathways are affected by aging in resistance arteries, and disease pathways are well represented indicating that aging confers a high risk of developing these diseases. However, processes involved in intercellular communication, ECM-cell interaction and cell-cell adhesion processes, such as focal adhesion, EPH-Ephrin signaling, and electrical conduction also seem to play a role in the aging vasculature. In the STRING interaction network (Figure 1), the most busy interaction cluster consist of regulators of structural proteins, showing close interactions between MMP9, MMP2, TGFβ1, TNFα, and Decorin. Decorin, which is a regulator of fibril formation and interacts with several other ECM proteins, was predicted by the STRING database as a functional interaction partner with proteins in Table 1. Another small interaction cluster is centered on the L-type voltage-gated Ca2+ channels (CaV1.2), with intracellular Ca2+ handling proteins, T-type channels, TRP channels, and K+ channels also being prominent in this cluster. The G-protein coupled receptors (AGTR1, AGTR2, EDNRA, EDNRB, TBXA2R, PTGFR, P2YR1 and ADORA1) form a similar cluster aside from the main network, with several connections to the other clusters and the main network. The actin cytoskeleton regulatory proteins ROCK1, ROCK2, MYL12B, PAK2, and ARHGEF7, form an additional cluster at the side of the largest cluster with the structural matrix regulating proteins. These pathways and interactions point to central roles of intracellular calcium, extracellular matrix structural proteins, actin-cytoskeleton regulation, and G protein-coupled receptors as being major targets for understanding and targeting age-dependent changes in resistance artery structure and function. By aiming to solve the complex role of aging in resistance arteries, it would be instrumental to obtain more mass-spec based proteomics studies, including special applications for posttranslational modifications and membrane proteins.
Discussion and concluding remarks
Figure 2 presents an overview of the changes found regarding myogenic tone, flow-mediated vasodilation, and structural remodeling in aging resistance arteries. The decline in myogenic tone would tend to cause vasodilation, in the absence of endothelial dysfunction and a loss of FMVD. Myogenic tone per se is not a direct cause of changes in blood pressure, but rather myogenic tone is adjusting the basal tone and autoregulation of blood flow at the prevailing blood pressure. Therefore, with normal blood pressure (normotension), a loss of myogenic tone would tend to cause outward remodeling [129,130]. By simultaneously stimulating vessel wall growth, i.e., outward hypertrophic remodeling, the aged artery under normotension will maintain the M/L-ratio unchanged. This remodeling response is reminiscent of the remodeling occurring with increased blood flow and a general increase in cardiac output, such as during pregnancy, normal growth, physical activity, obesity, etc. Conversely, if the predominant change in aging is a disruption of endothelial function and FMVD with little change in myogenic tone, the vessel will undergo acute vasoconstriction as shown in the lower part of Figure 2. Endothelial dysfunction often leads to hypertension, and with prolonged vasoconstriction due to disruption of FMVD and an increased wall stress due to hypertension, the vessel will respond by inward hypertrophic remodeling in aging (see lower part of Figure 2). In addition, Figure 2 summarizes the major mechanisms that might be involved in disruption of myogenic tone, FMVD and the changes in arterial structure. The reader is referred to Table 1, Table 2, Table 3 and Table 4 for a deeper insight into the concrete molecular mechanisms and pathways that become activated or deficient by aging. However, it is worthwhile considering the proteomic and bioinformatics strategy to advance our understanding of the age-dependent changes in resistance artery structure and function. For example, the STRING analysis shown in Figure 1 enables an overview of the physical and functional interactions between the proteins with a suggested age-dependent role. The same analysis also led to a suggestion of additional interacting protein partners (see Table 4) to the existing protein-protein interaction network, and therefore offers new insight not previously discussed in the review. For example, a literature search indicated that the proteins Decorin, NFKFBIA, RelB, TGFB3, TIMP1, TNFRSF1A and UQCRC1 might be involved in vascular ageing.
This review focused on age-dependent changes in three types basal vascular responses causing local changes in vasomotor tone and with consequences for an individual’s risk of developing serious age-related diseases. Other mechanisms, such as local metabolic regulation, neuro-hormonal regulation, capillary rarefaction, or angiogenesis, might also be crucial for the age-dependent changes in resistance artery structure and function. It is my hope that this review will stimulate more work and new approaches in the study of the effects and possible therapeutic management options of an age-dependent decline in resistance artery function.
Author Contributions
Only the author has contributed to the manuscript.
Acknowledgments
The title and content of this review reflects the subject of an invited lecture that I gave at Department of Pharmacology and Pharmacy, The University of Hong Kong in August 2023. I therefore thank the guest editor, Professor Yu Wang, Department of Pharmacology and Pharmacy, The University of Hong Kong, for the invitation to submit this review based on my lecture.
Conflicts of Interest
None.
Abbreviations
AGE advanced glycation end-products; CSA cross-sectional area; DL lumen diameter; ECM extracellular matrix; FMVD flow-mediated vasodilation; KEGG Kyoto encyclopedia of genes and genomes; MAP mean arterial pressure; MCA middle cerebral artery; M/L media:lumen-ratio; MRA mesenteric resistance artery; MT myogenic tone; NAD(P)H nicotinamide adenine dinucleotide phosphate (NAD(P)H oxidase); NO nitric oxide; RAGE receptor for advanced glycation end-products; ROS reactive oxygen species; SHR spontaneously hypertensive rat; SR sarcoplasmic reticulum; TRP transient receptor potential (cation channel); VGCC voltage-gated calcium channel; W/L wall:lumen-ratio; Wnt wingless-related integration site (Wnt-signaling); 3xFAD triple-transgenic familial Alzheimer’s Disease mouse model. The reader is referred to Table 1 for a definition of individual proteins.
References
- Schiffrin, E.L. How Structure, Mechanics, and Function of the Vasculature Contribute to Blood Pressure Elevation in Hypertension. Can. J. Cardiol. 2020, 36, 648–658. [Google Scholar] [CrossRef] [PubMed]
- Smulyan, H.; Mookherjee, S.; Safar, M.E. The two faces of hypertension: role of aortic stiffness. J. Am. Soc. Hypertens. 2016, 10, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Laurent, S.; Boutouyrie, P. Arterial Stiffness and Hypertension in the Elderly. Front Cardiovasc Med 2020, 7, 544302. [Google Scholar] [CrossRef] [PubMed]
- Rizzoni, D.; Rizzoni, M.; Nardin, M.; Chiarini, G.; Agabiti-Rosei, C.; Aggiusti, C.; Paini, A.; Salvetti, M.; Muiesan, M.L. Vascular Aging and Disease of the Small Vessels. High Blood Press Cardiovasc. Prev 2019, 26, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Thorin-Trescases, N.; de Montgolfier, O.; Pincon, A.; Raignault, A.; Caland, L.; Labbe, P.; Thorin, E. Impact of pulse pressure on cerebrovascular events leading to age-related cognitive decline. Am. J. Physiol. Heart Circ. Physiol. 2018, 314, H1214–H1224. [Google Scholar] [CrossRef] [PubMed]
- Laurent, S.; Agabiti-Rosei, C.; Bruno, R.M.; Rizzoni, D. Microcirculation and Macrocirculation in Hypertension: A Dangerous Cross-Link? Hypertension 2022, 79, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Gong, Z.; Li, Z.; Li, L.; Kong, W. Vascular Extracellular Matrix Remodeling and Hypertension. Antioxid Redox Signal 2021, 34, 765–783. [Google Scholar] [CrossRef] [PubMed]
- Fonck, E.; Feigl, G.G.; Fasel, J.; Sage, D.; Unser, M.; Rufenacht, D.A.; Stergiopulos, N. Effect of aging on elastin functionality in human cerebral arteries. Stroke 2009, 40, 2552–2556. [Google Scholar] [CrossRef] [PubMed]
- Wagenseil, J.E.; Mecham, R.P. Vascular extracellular matrix and arterial mechanics. Physiol. Rev. 2009, 89, 957–989. [Google Scholar] [CrossRef]
- Huveneers, S.; Daemen, M.J.; Hordijk, P.L. Between Rho(k) and a hard place: the relation between vessel wall stiffness, endothelial contractility, and cardiovascular disease. Circ. Res. 2015, 116, 895–908. [Google Scholar] [CrossRef]
- Lacolley, P.; Regnault, V.; Segers, P.; Laurent, S. Vascular Smooth Muscle Cells and Arterial Stiffening: Relevance in Development, Aging, and Disease. Physiol. Rev. 2017, 97, 1555–1617. [Google Scholar] [CrossRef]
- Lacolley, P.; Regnault, V.; Laurent, S. Mechanisms of Arterial Stiffening: From Mechanotransduction to Epigenetics. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 1055–1062. [Google Scholar] [CrossRef]
- Franklin, S.S.; Gustin, W.t.; Wong, N.D.; Larson, M.G.; Weber, M.A.; Kannel, W.B.; Levy, D. Hemodynamic patterns of age-related changes in blood pressure. The Framingham Heart Study. Circulation 1997, 96, 308–315. [Google Scholar] [CrossRef]
- Cheng, S.; Xanthakis, V.; Sullivan, L.M.; Vasan, R.S. Blood pressure tracking over the adult life course: patterns and correlates in the Framingham heart study. Hypertension 2012, 60, 1393–1399. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Kim, A.; Ebinger, J.E.; Niiranen, T.J.; Claggett, B.L.; Bairey Merz, C.N.; Cheng, S. Sex Differences in Blood Pressure Trajectories Over the Life Course. JAMA Cardiol 2020, 5, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Harraz, O.F.; Jensen, L.J. Vascular calcium signalling and ageing. J. Physiol. 2021, 599, 5361–5377. [Google Scholar] [CrossRef] [PubMed]
- Flurkey, K.; Currer, J.M.; Harteneck, C. Mouse Models in Aging Research. In The Mouse in Biomedical Research: Normative Biology, Husbandry, and Models, 2nd ed.; Fox, J.G., Barthold, S.W., Davisson, M.T., Newcomer, C.E., Quimby, F.W., Smith, A.L., Eds.; Academic Press: Burlington, MA, 2007; pp. 637–672. [Google Scholar]
- Harraz, O.F.; Jensen, L.J. Aging, calcium channel signaling and vascular tone. Mech. Ageing Dev 2020, 191, 111336. [Google Scholar] [CrossRef]
- Wirth, A.; Wang, S.; Takefuji, M.; Tang, C.; Althoff, T.F.; Schweda, F.; Wettschureck, N.; Offermanns, S. Age-dependent blood pressure elevation is due to increased vascular smooth muscle tone mediated by G-protein signalling. Cardiovasc. Res 2016, 109, 131–140. [Google Scholar] [CrossRef]
- Barsha, G.; Denton, K.M.; Mirabito Colafella, K.M. Sex- and age-related differences in arterial pressure and albuminuria in mice. Biol. Sex Differ. 2016, 7, 57. [Google Scholar] [CrossRef]
- Barsha, G.; Mirabito Colafella, K.M.; Walton, S.L.; Gaspari, T.A.; Spizzo, I.; Pinar, A.A.; Hilliard Krause, L.M.; Widdop, R.E.; Samuel, C.S.; Denton, K.M. In Aged Females, the Enhanced Pressor Response to Angiotensin II Is Attenuated By Estrogen Replacement via an Angiotensin Type 2 Receptor-Mediated Mechanism. Hypertension 2021, 78, 128–137. [Google Scholar] [CrossRef]
- Jensen, L.J.; Nielsen, M.S.; Salomonsson, M.; Sorensen, C.M. T-type Ca(2+) channels and autoregulation of local blood flow. Channels (Austin) 2017, 11, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Holmberg, J.; Bhattachariya, A.; Alajbegovic, A.; Rippe, C.; Ekman, M.; Dahan, D.; Hien, T.T.; Boettger, T.; Braun, T.; Sward, K.; et al. Loss of Vascular Myogenic Tone in miR-143/145 Knockout Mice Is Associated With Hypertension-Induced Vascular Lesions in Small Mesenteric Arteries. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Thorin-Trescases, N.; Bartolotta, T.; Hyman, N.; Penar, P.L.; Walters, C.L.; Bevan, R.D.; Bevan, J.A. Diameter dependence of myogenic tone of human pial arteries. Possible relation to distensibility. Stroke 1997, 28, 2486–2492. [Google Scholar] [CrossRef]
- Nyborg, N.C.; Nielsen, P.J. The level of spontaneous myogenic tone in isolated human posterior ciliary arteries decreases with age. Exp. Eye Res. 1990, 51, 711–715. [Google Scholar] [CrossRef] [PubMed]
- Springo, Z.; Toth, P.; Tarantini, S.; Ashpole, N.M.; Tucsek, Z.; Sonntag, W.E.; Csiszar, A.; Koller, A.; Ungvari, Z.I. Aging impairs myogenic adaptation to pulsatile pressure in mouse cerebral arteries. J. Cereb. Blood Flow Metab 2015, 35, 527–530. [Google Scholar] [CrossRef] [PubMed]
- Toth, P.; Csiszar, A.; Tucsek, Z.; Sosnowska, D.; Gautam, T.; Koller, A.; Schwartzman, M.L.; Sonntag, W.E.; Ungvari, Z. Role of 20-HETE, TRPC channels, and BKCa in dysregulation of pressure-induced Ca2+ signaling and myogenic constriction of cerebral arteries in aged hypertensive mice. Am. J. Physiol Heart Circ. Physiol 2013, 305, H1698–H1708. [Google Scholar] [CrossRef] [PubMed]
- Toth, P.; Tucsek, Z.; Sosnowska, D.; Gautam, T.; Mitschelen, M.; Tarantini, S.; Deak, F.; Koller, A.; Sonntag, W.E.; Csiszar, A.; Ungvari, Z. Age-related autoregulatory dysfunction and cerebromicrovascular injury in mice with angiotensin II-induced hypertension. J. Cereb. Blood Flow Metab. 2013, 33, 1732–1742. [Google Scholar] [CrossRef] [PubMed]
- Geary, G.G.; Buchholz, J.N. Selected contribution: Effects of aging on cerebrovascular tone and [Ca2+]i. J Appl Physiol (1985) 2003, 95, 1746–1754. [Google Scholar] [CrossRef]
- Polk, F.D.; Hakim, M.A.; Silva, J.F.; Behringer, E.J.; Pires, P.W. Endothelial K(IR)2 channel dysfunction in aged cerebral parenchymal arterioles. Am. J. Physiol. Heart Circ. Physiol. 2023. [CrossRef]
- Reed, J.T.; Pareek, T.; Sriramula, S.; Pabbidi, M.R. Aging influences cerebrovascular myogenic reactivity and BK channel function in a sex-specific manner. Cardiovasc. Res. 2020, 116, 1372–1385. [Google Scholar] [CrossRef]
- Wei, J.; Zhu, J.; Zhang, J.; Jiang, S.; Qu, L.; Wang, L.; Buggs, J.; Tan, X.; Cheng, F.; Liu, R. Aging Impairs Renal Autoregulation in Mice. Hypertension 2020, 75, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Muller-Delp, J.; Spier, S.A.; Ramsey, M.W.; Lesniewski, L.A.; Papadopoulos, A.; Humphrey, J.D.; Delp, M.D. Effects of aging on vasoconstrictor and mechanical properties of rat skeletal muscle arterioles. Am. J. Physiol. Heart Circ. Physiol. 2002, 282, H1843–1854. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.S.; Kim, S.; Dominguez, J.M.; Sindler, A.L.; Dick, G.M.; Muller-Delp, J.M. Aging and muscle fiber type alter K+ channel contributions to the myogenic response in skeletal muscle arterioles. J. Appl. Physiol 2009, 107, 389–398. [Google Scholar] [CrossRef]
- Ghosh, P.; Mora Solis, F.R.; Dominguez, J.M., 2nd; Spier, S.A.; Donato, A.J.; Delp, M.D.; Muller-Delp, J.M. Exercise training reverses aging-induced impairment of myogenic constriction in skeletal muscle arterioles. J Appl Physiol (1985) 2015, 118, 904–911. [Google Scholar] [CrossRef] [PubMed]
- Hayoz, S.; Pettis, J.; Bradley, V.; Segal, S.S.; Jackson, W.F. Increased amplitude of inward rectifier K(+) currents with advanced age in smooth muscle cells of murine superior epigastric arteries. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H1203–H1214. [Google Scholar] [CrossRef]
- Shipley, R.D.; Muller-Delp, J.M. Aging decreases vasoconstrictor responses of coronary resistance arterioles through endothelium-dependent mechanisms. Cardiovasc. Res. 2005, 66, 374–383. [Google Scholar] [CrossRef]
- Vessieres, E.; Dib, A.; Bourreau, J.; Lelievre, E.; Custaud, M.A.; Lelievre-Pegorier, M.; Loufrani, L.; Henrion, D.; Fassot, C. Long Lasting Microvascular Tone Alteration in Rat Offspring Exposed In Utero to Maternal Hyperglycaemia. PLoS One 2016, 11, e0146830. [Google Scholar] [CrossRef]
- Björling, K.; Joseph, P.D.; Egebjerg, K.; Salomonsson, M.; Hansen, J.L.; Ludvigsen, T.P.; Jensen, L.J. Role of age, Rho-kinase 2 expression, and G protein-mediated signaling in the myogenic response in mouse small mesenteric arteries. Physiol Rep 2018, 6, e13863. [Google Scholar] [CrossRef] [PubMed]
- Gros, R.; Van, W.R.; You, X.; Thorin, E.; Husain, M. Effects of age, gender, and blood pressure on myogenic responses of mesenteric arteries from C57BL/6 mice. Am. J. Physiol Heart Circ. Physiol 2002, 282, H380–H388. [Google Scholar] [CrossRef]
- McCurley, A.; McGraw, A.; Pruthi, D.; Jaffe, I.Z. Smooth muscle cell mineralocorticoid receptors: role in vascular function and contribution to cardiovascular disease. Pflugers Arch. 2013, 465, 1661–1670. [Google Scholar] [CrossRef]
- DuPont, J.J.; McCurley, A.; Davel, A.P.; McCarthy, J.; Bender, S.B.; Hong, K.; Yang, Y.; Yoo, J.K.; Aronovitz, M.; Baur, W.E.; et al. Vascular mineralocorticoid receptor regulates microRNA-155 to promote vasoconstriction and rising blood pressure with aging. JCI. Insight 2016, 1, e88942. [Google Scholar] [CrossRef] [PubMed]
- Georgeon-Chartier, C.; Menguy, C.; Prevot, A.; Morel, J.L. Effect of aging on calcium signaling in C57Bl6J mouse cerebral arteries. Pflugers Arch. 2013, 465, 829–838. [Google Scholar] [CrossRef]
- Liao, J.; Zhang, Y.; Ye, F.; Zhang, L.; Chen, Y.; Zeng, F.; Shi, L. Epigenetic regulation of L-type voltage-gated Ca(2+) channels in mesenteric arteries of aging hypertensive rats. Hypertens. Res. 2017, 40, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Korzick, D.H.; Muller-Delp, J.M.; Dougherty, P.; Heaps, C.L.; Bowles, D.K.; Krick, K.K. Exaggerated coronary vasoreactivity to endothelin-1 in aged rats: role of protein kinase C. Cardiovasc. Res. 2005, 66, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, M.F.; Bjorling, K.; Jensen, L.J. Age-dependent impact of CaV 3.2 T-type calcium channel deletion on myogenic tone and flow-mediated vasodilatation in small arteries. J. Physiol 2016, 594, 5881–5898. [Google Scholar] [CrossRef] [PubMed]
- del Corsso, C.; Ostrovskaya, O.; McAllister, C.E.; Murray, K.; Hatton, W.J.; Gurney, A.M.; Spencer, N.J.; Wilson, S.M. Effects of aging on Ca2+ signaling in murine mesenteric arterial smooth muscle cells. Mech. Ageing Dev. 2006, 127, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Björling, K.; Morita, H.; Olsen, M.F.; Prodan, A.; Hansen, P.B.; Lory, P.; Holstein-Rathlou, N.H.; Jensen, L.J. Myogenic tone is impaired at low arterial pressure in mice deficient in the low-voltage-activated CaV 3.1 T-type Ca(2+) channel. Acta Physiol. (Oxf.) 2013, 207, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Harraz, O.F.; Abd El-Rahman, R.R.; Bigdely-Shamloo, K.; Wilson, S.M.; Brett, S.E.; Romero, M.; Gonzales, A.L.; Earley, S.; Vigmond, E.J.; Nygren, A.; et al. Ca(V)3.2 channels and the induction of negative feedback in cerebral arteries. Circ. Res 2014, 115, 650–661. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.; Kassmann, M.; Cui, Y.; Matthaeus, C.; Kunz, S.; Zhong, C.; Zhu, S.; Xie, Y.; Tsvetkov, D.; Daumke, O.; et al. Age attenuates the T-type CaV 3.2-RyR axis in vascular smooth muscle. Aging Cell 2020, 19, e13134. [Google Scholar] [CrossRef]
- Lin, J.; Guo, W.; Luo, Q.; Zhang, Q.; Wan, T.; Jiang, C.; Ye, Y.; Lin, H.; Fan, G. Senolytics prevent caveolar Ca(V) 3.2-RyR axis malfunction in old vascular smooth muscle. Aging Cell 2023, 22, e14002. [Google Scholar] [CrossRef]
- Welsh, D.G.; Morielli, A.D.; Nelson, M.T.; Brayden, J.E. Transient receptor potential channels regulate myogenic tone of resistance arteries. Circ. Res 2002, 90, 248–250. [Google Scholar] [CrossRef]
- Inoue, R.; Jensen, L.J.; Jian, Z.; Shi, J.; Hai, L.; Lurie, A.I.; Henriksen, F.H.; Salomonsson, M.; Morita, H.; Kawarabayashi, Y.; et al. Synergistic activation of vascular TRPC6 channel by receptor and mechanical stimulation via phospholipase C/diacylglycerol and phospholipase A2/omega-hydroxylase/20-HETE pathways. Circ. Res 2009, 104, 1399–1409. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, E.; Thakore, P.; Ali, S.; Sanchez Solano, A.; Wang, X.; Gao, X.; Labelle-Dumais, C.; Chaumeil, M.M.; Gould, D.B.; Earley, S. Impaired intracellular Ca(2+) signaling contributes to age-related cerebral small vessel disease in Col4a1 mutant mice. Sci Signal 2023, 16, eadi3966. [Google Scholar] [CrossRef] [PubMed]
- Marijic, J.; Li, Q.; Song, M.; Nishimaru, K.; Stefani, E.; Toro, L. Decreased expression of voltage- and Ca(2+)-activated K(+) channels in coronary smooth muscle during aging. Circ. Res 2001, 88, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Rabaglino, M.B.; Wakabayashi, M.; Pearson, J.T.; Jensen, L.J. Effect of age on the vascular proteome in middle cerebral arteries and mesenteric resistance arteries in mice. Mech. Ageing Dev. 2021, 200, 111594. [Google Scholar] [CrossRef]
- Kawarazaki, W.; Fujita, T. Role of Rho in Salt-Sensitive Hypertension. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Marchand, A.; Atassi, F.; Gaaya, A.; Leprince, P.; Le, F.C.; Soubrier, F.; Lompre, A.M.; Nadaud, S. The Wnt/beta-catenin pathway is activated during advanced arterial aging in humans. Aging Cell 2011, 10, 220–232. [Google Scholar] [CrossRef]
- Kawarazaki, W.; Mizuno, R.; Nishimoto, M.; Ayuzawa, N.; Hirohama, D.; Ueda, K.; Kawakami-Mori, F.; Oba, S.; Marumo, T.; Fujita, T. Salt causes aging-associated hypertension via vascular Wnt5a under Klotho deficiency. J. Clin. Invest. 2020, 130, 4152–4166. [Google Scholar] [CrossRef]
- Faisy, C.; Pinto, F.M.; Le Guen, M.; Naline, E.; Grassin Delyle, S.; Sage, E.; Candenas, M.L.; Devillier, P. Airway response to acute mechanical stress in a human bronchial model of stretch. Crit. Care 2011, 15, R208. [Google Scholar] [CrossRef]
- Thijssen, D.H.; Black, M.A.; Pyke, K.E.; Padilla, J.; Atkinson, G.; Harris, R.A.; Parker, B.; Widlansky, M.E.; Tschakovsky, M.E.; Green, D.J. Assessment of flow-mediated dilation in humans: a methodological and physiological guideline. Am. J. Physiol. Heart Circ. Physiol. 2011, 300, H2–12. [Google Scholar] [CrossRef]
- Gradel, A.K.J.; Salomonsson, M.; Sorensen, C.M.; Holstein-Rathlou, N.H.; Jensen, L.J. Long-term diet-induced hypertension in rats is associated with reduced expression and function of small artery SK(Ca), IK(Ca), and Kir2. 1 channels. Clin. Sci. (Lond.) 2018, 132, 461–474. [Google Scholar] [CrossRef] [PubMed]
- Thijssen, D.H.J.; Bruno, R.M.; van Mil, A.; Holder, S.M.; Faita, F.; Greyling, A.; Zock, P.L.; Taddei, S.; Deanfield, J.E.; Luscher, T.; et al. Expert consensus and evidence-based recommendations for the assessment of flow-mediated dilation in humans. Eur. Heart J. 2019, 40, 2534–2547. [Google Scholar] [CrossRef] [PubMed]
- Jordan, J.; Nilsson, P.M.; Kotsis, V.; Olsen, M.H.; Grassi, G.; Yumuk, V.; Hauner, H.; Zahorska-Markiewicz, B.; Toplak, H.; Engeli, S.; Finer, N. Joint scientific statement of the European Association for the Study of Obesity and the European Society of Hypertension: Obesity and early vascular ageing. J. Hypertens. 2015, 33, 425–434. [Google Scholar] [CrossRef]
- Hopkins, N.D.; Dengel, D.R.; Stratton, G.; Kelly, A.S.; Steinberger, J.; Zavala, H.; Marlatt, K.; Perry, D.; Naylor, L.H.; Green, D.J. Age and sex relationship with flow-mediated dilation in healthy children and adolescents. J Appl Physiol (1985) 2015, 119, 926–933. [Google Scholar] [CrossRef]
- Eskurza, I.; Monahan, K.D.; Robinson, J.A.; Seals, D.R. Effect of acute and chronic ascorbic acid on flow-mediated dilatation with sedentary and physically active human ageing. J. Physiol. 2004, 556, 315–324. [Google Scholar] [CrossRef]
- Franzoni, F.; Ghiadoni, L.; Galetta, F.; Plantinga, Y.; Lubrano, V.; Huang, Y.; Salvetti, G.; Regoli, F.; Taddei, S.; Santoro, G.; Salvetti, A. Physical activity, plasma antioxidant capacity, and endothelium-dependent vasodilation in young and older men. Am. J. Hypertens. 2005, 18, 510–516. [Google Scholar] [CrossRef]
- Parker, B.A.; Ridout, S.J.; Proctor, D.N. Age and flow-mediated dilation: a comparison of dilatory responsiveness in the brachial and popliteal arteries. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H3043–3049. [Google Scholar] [CrossRef]
- Donato, A.J.; Eskurza, I.; Silver, A.E.; Levy, A.S.; Pierce, G.L.; Gates, P.E.; Seals, D.R. Direct evidence of endothelial oxidative stress with aging in humans: relation to impaired endothelium-dependent dilation and upregulation of nuclear factor-kappaB. Circ. Res. 2007, 100, 1659–1666. [Google Scholar] [CrossRef] [PubMed]
- Lane-Cordova, A.D.; Ranadive, S.M.; Kappus, R.M.; Cook, M.D.; Phillips, S.A.; Woods, J.A.; Wilund, K.R.; Baynard, T.; Fernhall, B. Aging, not age-associated inflammation, determines blood pressure and endothelial responses to acute inflammation. J. Hypertens. 2016, 34, 2402–2409. [Google Scholar] [CrossRef]
- Rossman, M.J.; Groot, H.J.; Garten, R.S.; Witman, M.A.; Richardson, R.S. Vascular function assessed by passive leg movement and flow-mediated dilation: initial evidence of construct validity. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H1277–H1286. [Google Scholar] [CrossRef]
- Benjamin, E.J.; Larson, M.G.; Keyes, M.J.; Mitchell, G.F.; Vasan, R.S.; Keaney, J.F., Jr.; Lehman, B.T.; Fan, S.; Osypiuk, E.; Vita, J.A. Clinical correlates and heritability of flow-mediated dilation in the community: the Framingham Heart Study. Circulation 2004, 109, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Rossman, M.J.; Santos-Parker, J.R.; Steward, C.A.C.; Bispham, N.Z.; Cuevas, L.M.; Rosenberg, H.L.; Woodward, K.A.; Chonchol, M.; Gioscia-Ryan, R.A.; Murphy, M.P.; Seals, D.R. Chronic Supplementation With a Mitochondrial Antioxidant (MitoQ) Improves Vascular Function in Healthy Older Adults. Hypertension 2018, 71, 1056–1063. [Google Scholar] [CrossRef] [PubMed]
- Beyer, A.M.; Zinkevich, N.; Miller, B.; Liu, Y.; Wittenburg, A.L.; Mitchell, M.; Galdieri, R.; Sorokin, A.; Gutterman, D.D. Transition in the mechanism of flow-mediated dilation with aging and development of coronary artery disease. Basic Res. Cardiol. 2017, 112, 5. [Google Scholar] [CrossRef] [PubMed]
- Schuler, D.; Sansone, R.; Freudenberger, T.; Rodriguez-Mateos, A.; Weber, G.; Momma, T.Y.; Goy, C.; Altschmied, J.; Haendeler, J.; Fischer, J.W.; et al. Measurement of endothelium-dependent vasodilation in mice--brief report. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2651–2657. [Google Scholar] [CrossRef] [PubMed]
- Delp, M.D.; Behnke, B.J.; Spier, S.A.; Wu, G.; Muller-Delp, J.M. Ageing diminishes endothelium-dependent vasodilatation and tetrahydrobiopterin content in rat skeletal muscle arterioles. J. Physiol. 2008, 586, 1161–1168. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.M.; Park, S.K.; Kwon, O.S.; Taylor La Salle, D.; Cerbie, J.; Fermoyle, C.C.; Morgan, D.; Nelson, A.; Bledsoe, A.; Bharath, L.P.; et al. Activating P2Y1 receptors improves function in arteries with repressed autophagy. Cardiovasc. Res. 2023, 119, 252–267. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.S.; Reyes, R.A.; Muller-Delp, J.M. Aging impairs flow-induced dilation in coronary arterioles: role of NO and H(2)O(2). Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H1087–1095. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.S.; Chen, B.; Reyes, R.A.; Leblanc, A.J.; Teng, B.; Mustafa, S.J.; Muller-Delp, J.M. Aging and estrogen alter endothelial reactivity to reactive oxygen species in coronary arterioles. Am. J. Physiol. Heart Circ. Physiol. 2011, 300, H2105–2115. [Google Scholar] [CrossRef] [PubMed]
- Tracy, E.P.; Dukes, M.; Rowe, G.; Beare, J.E.; Nair, R.; LeBlanc, A.J. Stromal Vascular Fraction Restores Vasodilatory Function by Reducing Oxidative Stress in Aging-Induced Coronary Microvascular Disease. Antioxid Redox Signal 2023, 38, 261–281. [Google Scholar] [CrossRef] [PubMed]
- Arenas, I.A.; Xu, Y.; Davidge, S.T. Age-associated impairment in vasorelaxation to fluid shear stress in the female vasculature is improved by TNF-alpha antagonism. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H1259–1263. [Google Scholar] [CrossRef]
- Zhou, X.; Bohlen, H.G.; Unthank, J.L.; Miller, S.J. Abnormal nitric oxide production in aged rat mesenteric arteries is mediated by NAD(P)H oxidase-derived peroxide. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H2227–2233. [Google Scholar] [CrossRef] [PubMed]
- Morton, J.S.; Rueda-Clausen, C.F.; Davidge, S.T. Flow-mediated vasodilation is impaired in adult rat offspring exposed to prenatal hypoxia. J Appl Physiol (1985) 2011, 110, 1073–1082. [Google Scholar] [CrossRef] [PubMed]
- Pinaud, F.; Bocquet, A.; Dumont, O.; Retailleau, K.; Baufreton, C.; Andriantsitohaina, R.; Loufrani, L.; Henrion, D. Paradoxical role of angiotensin II type 2 receptors in resistance arteries of old rats. Hypertension 2007, 50, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Huang, A.; Yan, E.H.; Wu, Z.; Yan, C.; Kaminski, P.M.; Oury, T.D.; Wolin, M.S.; Kaley, G. Reduced release of nitric oxide to shear stress in mesenteric arteries of aged rats. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, H2249–2256. [Google Scholar] [CrossRef]
- Mulvany, M.J. Vascular remodelling of resistance vessels: can we define this? Cardiovasc. Res 1999, 41, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Ludvigsen, T.P.; Olsen, L.H.; Pedersen, H.D.; Christoffersen, B.O.; Jensen, L.J. Hyperglycemia-induced transcriptional regulation of ROCK1 and TGM2 expression is involved in small artery remodeling in obese diabetic Gottingen Minipigs. Clin. Sci. (Lond) 2019, 133, 2499–2516. [Google Scholar] [CrossRef]
- Buus, N.H.; Mathiassen, O.N.; Fenger-Gron, M.; Praestholm, M.N.; Sihm, I.; Thybo, N.K.; Schroeder, A.P.; Thygesen, K.; Aalkjaer, C.; Pedersen, O.L.; et al. Small artery structure during antihypertensive therapy is an independent predictor of cardiovascular events in essential hypertension. J. Hypertens 2013, 31, 791–797. [Google Scholar] [CrossRef]
- Agabiti-Rosei, E.; Rizzoni, D. Microvascular structure as a prognostically relevant endpoint. J. Hypertens 2017, 35, 914–921. [Google Scholar] [CrossRef]
- Lakatta, E.G. The reality of aging viewed from the arterial wall. Artery Res 2013, 7, 73–80. [Google Scholar] [CrossRef]
- Nilsson, P.M. Hemodynamic Aging as the Consequence of Structural Changes Associated with Early Vascular Aging (EVA). Aging Dis. 2014, 5, 109–113. [Google Scholar] [CrossRef]
- Harvey, A.; Montezano, A.C.; Touyz, R.M. Vascular biology of ageing-Implications in hypertension. J. Mol. Cell. Cardiol. 2015, 83, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Miura, K. Tunica intima compensation for reduced stiffness of the tunica media in aging renal arteries as measured with scanning acoustic microscopy. PLoS One 2020, 15, e0234759. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Froehlich, J.; Galis, Z.S.; Lakatta, E.G. Increased expression of matrix metalloproteinase-2 in the thickened intima of aged rats. Hypertension 1999, 33, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Takagi, G.; Asai, K.; Resuello, R.G.; Natividad, F.F.; Vatner, D.E.; Vatner, S.F.; Lakatta, E.G. Aging increases aortic MMP-2 activity and angiotensin II in nonhuman primates. Hypertension 2003, 41, 1308–1316. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, J.; Jiang, L.Q.; Spinetti, G.; Pintus, G.; Monticone, R.; Kolodgie, F.D.; Virmani, R.; Lakatta, E.G. Proinflammatory profile within the grossly normal aged human aortic wall. Hypertension 2007, 50, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhao, D.; Spinetti, G.; Zhang, J.; Jiang, L.Q.; Pintus, G.; Monticone, R.; Lakatta, E.G. Matrix metalloproteinase 2 activation of transforming growth factor-beta1 (TGF-beta1) and TGF-beta1-type II receptor signaling within the aged arterial wall. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1503–1509. [Google Scholar] [CrossRef] [PubMed]
- Longtine, A.G.; Venkatasubramanian, R.; Zigler, M.C.; Lindquist, A.J.; Mahoney, S.A.; Greenberg, N.T.; VanDongen, N.S.; Ludwig, K.R.; Moreau, K.L.; Seals, D.R.; Clayton, Z.S. Female C57BL/6N mice are a viable model of aortic aging in women. Am. J. Physiol. Heart Circ. Physiol. 2023, 324, H893–H904. [Google Scholar] [CrossRef] [PubMed]
- Yasmin; McEniery, C.M.; Wallace, S.; Dakham, Z.; Pulsalkar, P.; Maki-Petaja, K.; Ashby, M.J.; Cockcroft, J.R.; Wilkinson, I.B. Matrix metalloproteinase-9 (MMP-9), MMP-2, and serum elastase activity are associated with systolic hypertension and arterial stiffness. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 372. [Google Scholar] [CrossRef] [PubMed]
- Zieman, S.J.; Melenovsky, V.; Clattenburg, L.; Corretti, M.C.; Capriotti, A.; Gerstenblith, G.; Kass, D.A. Advanced glycation endproduct crosslink breaker (alagebrium) improves endothelial function in patients with isolated systolic hypertension. J. Hypertens. 2007, 25, 577–583. [Google Scholar] [CrossRef]
- Chabot, N.; Moreau, S.; Mulani, A.; Moreau, P.; Keillor, J.W. Fluorescent probes of tissue transglutaminase reveal its association with arterial stiffening. Chem. Biol. 2010, 17, 1143–1150. [Google Scholar] [CrossRef]
- Bruno, R.M.; Duranti, E.; Ippolito, C.; Segnani, C.; Bernardini, N.; Di, C.G.; Chiarugi, M.; Taddei, S.; Virdis, A. Different Impact of Essential Hypertension on Structural and Functional Age-Related Vascular Changes. Hypertension 2017, 69, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Hanna, M.A.; Taylor, C.R.; Chen, B.; La, H.S.; Maraj, J.J.; Kilar, C.R.; Behnke, B.J.; Delp, M.D.; Muller-Delp, J.M. Structural remodeling of coronary resistance arteries: effects of age and exercise training. J Appl Physiol (1985) 2014, 117, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Behnke, B.J.; Prisby, R.D.; Lesniewski, L.A.; Donato, A.J.; Olin, H.M.; Delp, M.D. Influence of ageing and physical activity on vascular morphology in rat skeletal muscle. J. Physiol. 2006, 575, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Moreau, P.; d’Uscio, L.V.; Luscher, T.F. Structure and reactivity of small arteries in aging. Cardiovasc. Res. 1998, 37, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Tumer, N.; Toklu, H.Z.; Muller-Delp, J.M.; Oktay, S.; Ghosh, P.; Strang, K.; Delp, M.D.; Scarpace, P.J. The effects of aging on the functional and structural properties of the rat basilar artery. Physiol Rep 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Mandala, M.; Pedatella, A.L.; Morales Palomares, S.; Cipolla, M.J.; Osol, G. Maturation is associated with changes in rat cerebral artery structure, biomechanical properties and tone. Acta Physiol. (Oxf.) 2012, 205, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Rzechorzek, W.; Zhang, H.; Buckley, B.K.; Hua, K.; Pomp, D.; Faber, J.E. Aerobic exercise prevents rarefaction of pial collaterals and increased stroke severity that occur with aging. J. Cereb. Blood Flow Metab. 2017, 37, 3544–3555. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Otero, J.M.; Garver, H.; Fink, G.D.; Jackson, W.F.; Dorrance, A.M. Aging is associated with changes to the biomechanical properties of the posterior cerebral artery and parenchymal arterioles. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H365–375. [Google Scholar] [CrossRef] [PubMed]
- Hannan, J.L.; Blaser, M.C.; Pang, J.J.; Adams, S.M.; Pang, S.C.; Adams, M.A. Impact of hypertension, aging, and antihypertensive treatment on the morphology of the pudendal artery. J. Sex. Med. 2011, 8, 1027–1038. [Google Scholar] [CrossRef]
- Nicholson, C.J.; Sweeney, M.; Robson, S.C.; Taggart, M.J. Estrogenic vascular effects are diminished by chronological aging. Sci. Rep. 2017, 7, 12153. [Google Scholar] [CrossRef]
- Ohanian, J.; Liao, A.; Forman, S.P.; Ohanian, V. Age-related remodeling of small arteries is accompanied by increased sphingomyelinase activity and accumulation of long-chain ceramides. Physiol Rep 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Freidja, M.L.; Vessieres, E.; Clere, N.; Desquiret, V.; Guihot, A.L.; Toutain, B.; Loufrani, L.; Jardel, A.; Procaccio, V.; Faure, S.; Henrion, D. Heme oxygenase-1 induction restores high-blood-flow-dependent remodeling and endothelial function in mesenteric arteries of old rats. J. Hypertens. 2011, 29, 102–112. [Google Scholar] [CrossRef]
- Briones, A.M.; Salaices, M.; Vila, E. Mechanisms underlying hypertrophic remodeling and increased stiffness of mesenteric resistance arteries from aged rats. J. Gerontol. A Biol. Sci. Med. Sci. 2007, 62, 696–706. [Google Scholar] [CrossRef]
- Dumont, O.; Pinaud, F.; Guihot, A.L.; Baufreton, C.; Loufrani, L.; Henrion, D. Alteration in flow (shear stress)-induced remodelling in rat resistance arteries with aging: improvement by a treatment with hydralazine. Cardiovasc. Res. 2008, 77, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Hausman, N.; Martin, J.; Taggart, M.J.; Austin, C. Age-related changes in the contractile and passive arterial properties of murine mesenteric small arteries are altered by caveolin-1 knockout. J. Cell. Mol. Med. 2012, 16, 1720–1730. [Google Scholar] [CrossRef]
- Caillon, A.; Grenier, C.; Grimaud, L.; Vessieres, E.; Guihot, A.L.; Blanchard, S.; Lelievre, E.; Chabbert, M.; Foucher, E.D.; Jeannin, P.; et al. The angiotensin II type 2 receptor activates flow-mediated outward remodelling through T cells-dependent interleukin-17 production. Cardiovasc. Res. 2016, 112, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Altayo, F.; Sanchez-Ventura, J.; Vila, E.; Gimenez-Llort, L. Crosstalk between Peripheral Small Vessel Properties and Anxious-like Profiles: Sex, Genotype, and Interaction Effects in Mice with Normal Aging and 3xTg-AD mice at Advanced Stages of Disease. J. Alzheimers Dis. 2018, 62, 1531–1538. [Google Scholar] [CrossRef]
- Tarhouni, K.; Freidja, M.L.; Guihot, A.L.; Vessieres, E.; Grimaud, L.; Toutain, B.; Lenfant, F.; Arnal, J.F.; Loufrani, L.; Henrion, D. Role of estrogens and age in flow-mediated outward remodeling of rat mesenteric resistance arteries. Am. J. Physiol. Heart Circ. Physiol. 2014, 307, H504–514. [Google Scholar] [CrossRef]
- Wang, M.; Jiang, L.; Monticone, R.E.; Lakatta, E.G. Proinflammation: the key to arterial aging. Trends Endocrinol. Metab. 2014, 25, 72–79. [Google Scholar] [CrossRef]
- Harvey, A.; Montezano, A.C.; Lopes, R.A.; Rios, F.; Touyz, R.M. Vascular Fibrosis in Aging and Hypertension: Molecular Mechanisms and Clinical Implications. Can. J. Cardiol. 2016, 32, 659–668. [Google Scholar] [CrossRef]
- Chehaitly, A.; Vessieres, E.; Guihot, A.L.; Henrion, D. Flow-mediated outward arterial remodeling in aging. Mech. Ageing Dev. 2021, 194, 111416. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cheng, C.K.; Yi, M.; Lui, K.O.; Huang, Y. Targeting endothelial dysfunction and inflammation. J. Mol. Cell. Cardiol. 2022, 168, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Lemkens, P.; Boari, G.; Fazzi, G.; Janssen, G.; Murphy-Ullrich, J.; Schiffers, P.; De Mey, J. Thrombospondin-1 in early flow-related remodeling of mesenteric arteries from young normotensive and spontaneously hypertensive rats. Open Cardiovasc. Med. J. 2012, 6, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Yamashiro, Y.; Thang, B.Q.; Ramirez, K.; Shin, S.J.; Kohata, T.; Ohata, S.; Nguyen, T.A.V.; Ohtsuki, S.; Nagayama, K.; Yanagisawa, H. Matrix mechanotransduction mediated by thrombospondin-1/integrin/YAP in the vascular remodeling. Proc. Natl. Acad. Sci. U. S. A. 2020, 117, 9896–9905. [Google Scholar] [CrossRef]
- Nevitt, C.; McKenzie, G.; Christian, K.; Austin, J.; Hencke, S.; Hoying, J.; LeBlanc, A. Physiological levels of thrombospondin-1 decrease NO-dependent vasodilation in coronary microvessels from aged rats. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H1842–1850. [Google Scholar] [CrossRef] [PubMed]
- Isenberg, J.S.; Hyodo, F.; Pappan, L.K.; Abu-Asab, M.; Tsokos, M.; Krishna, M.C.; Frazier, W.A.; Roberts, D.D. Blocking thrombospondin-1/CD47 signaling alleviates deleterious effects of aging on tissue responses to ischemia. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2582–2588. [Google Scholar] [CrossRef] [PubMed]
- Santinha, D.; Vilaca, A.; Estronca, L.; Schuler, S.C.; Bartoli, C.; De Sandre-Giovannoli, A.; Figueiredo, A.; Quaas, M.; Pompe, T.; Ori, A.; Ferreira, L. Remodeling of the Cardiac Extracellular Matrix Proteome During Chronological and Pathological Aging. Mol. Cell. Proteomics 2024, 23, 100706. [Google Scholar] [CrossRef]
- Jacobsen, J.C.; Mulvany, M.J.; Holstein-Rathlou, N.H. A mechanism for arteriolar remodeling based on maintenance of smooth muscle cell activation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R1379–1389. [Google Scholar] [CrossRef]
- Klein, A.; Joseph, P.D.; Christensen, V.G.; Jensen, L.J.; Jacobsen, J.C.B. Lack of tone in mouse small mesenteric arteries leads to outward remodeling, which can be prevented by prolonged agonist-induced vasoconstriction. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H644–H657. [Google Scholar] [CrossRef]
Figure 1.
Protein-protein interaction network. The proteins in Table 1 were analyzed using STRING database to show their respective physical and functional interactions with medium confidence level (Score ≥ 0.400). See Table for definition of symbols.
Figure 1.
Protein-protein interaction network. The proteins in Table 1 were analyzed using STRING database to show their respective physical and functional interactions with medium confidence level (Score ≥ 0.400). See Table for definition of symbols.
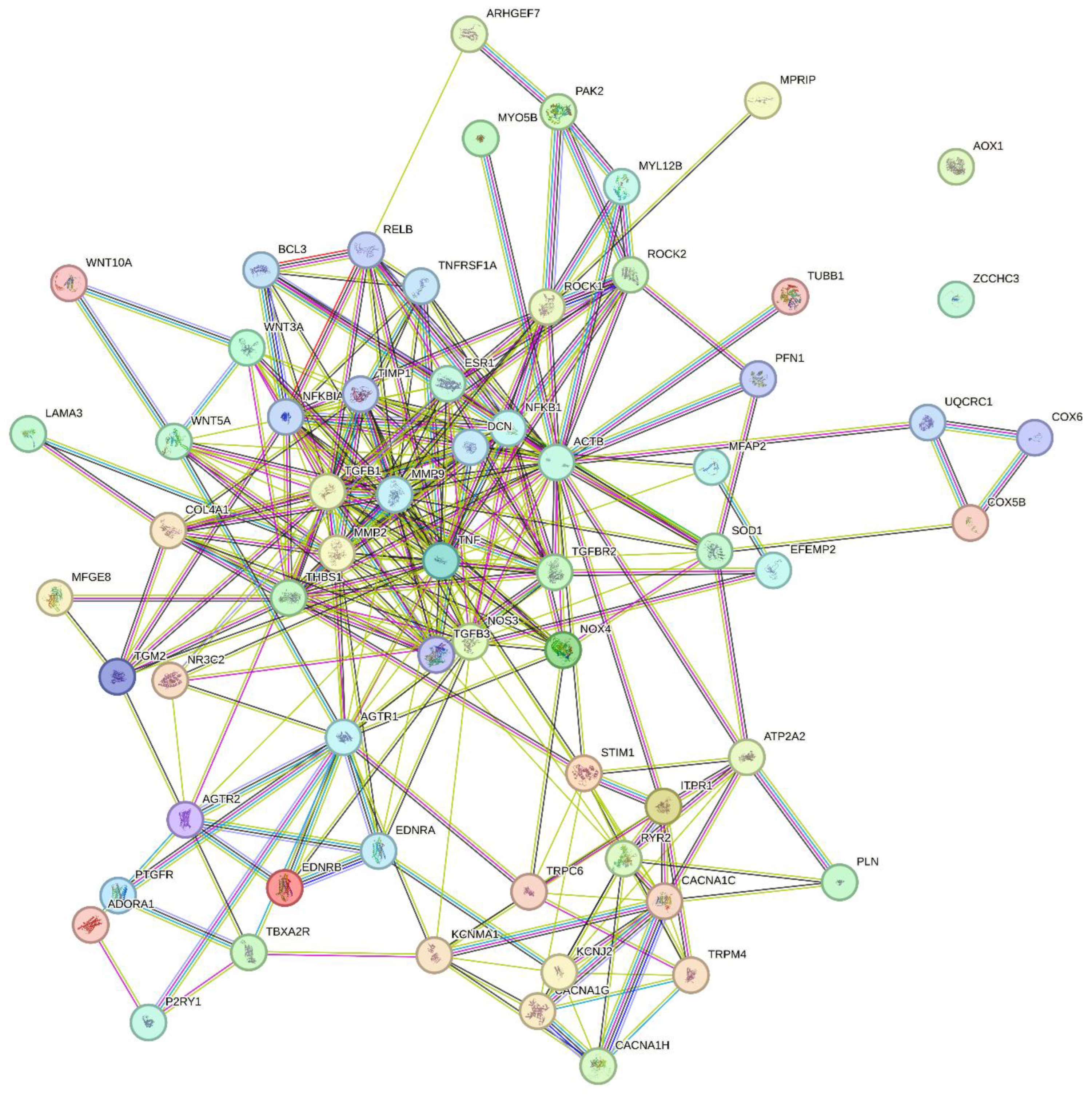
Figure 2.
Summary of age-dependent changes in myogenic tone, flow-mediated vasodilation and structural remodeling in resistance arteries. See text for explanations. The inner endothelial (intima) layer is shown as gray-blue, the smooth muscle (media) layer is shown as pink, and the connective tissue (adventitia) layer is shown in yellow color. Arrows denote the direction of change in a function, factor or parameter. MT (myogenic tone); FMVD (flow-mediated vasodilation); DL (passive lumen diameter); CSA (cross-sectional area); M/L-ratio (media:lumen-ratio); ROS (reactive oxygen species); NO (nitric oxide); VGCC (voltage-gated calcium channel); TRP (transient receptor potential channel).
Figure 2.
Summary of age-dependent changes in myogenic tone, flow-mediated vasodilation and structural remodeling in resistance arteries. See text for explanations. The inner endothelial (intima) layer is shown as gray-blue, the smooth muscle (media) layer is shown as pink, and the connective tissue (adventitia) layer is shown in yellow color. Arrows denote the direction of change in a function, factor or parameter. MT (myogenic tone); FMVD (flow-mediated vasodilation); DL (passive lumen diameter); CSA (cross-sectional area); M/L-ratio (media:lumen-ratio); ROS (reactive oxygen species); NO (nitric oxide); VGCC (voltage-gated calcium channel); TRP (transient receptor potential channel).
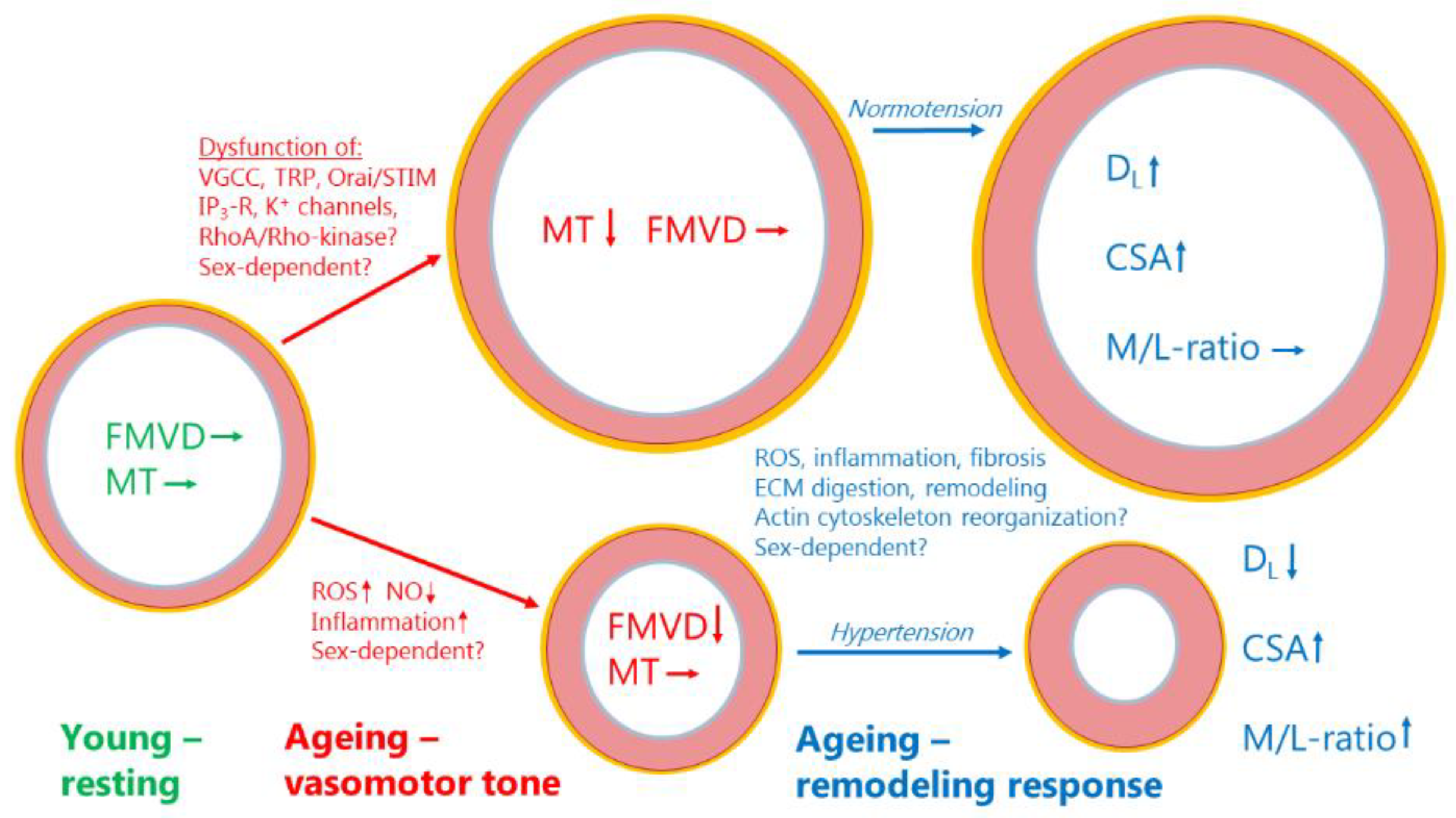

Table 1.
List of proteins with possible age-dependent role in resistance arteries (see text).
| Uniprot ID (Homo sapiens) | Gene name | Protein (short name) | Protein name |
|---|---|---|---|
| P60709 | ACTB | Beta-actin | Actin, cytoplasmic 1 |
| P30542 | ADORA1 | A1-R | Adenosine receptor A1 |
| P30556 | AGTR1 | AT1-R | Type-1 angiotensin II receptor |
| P50052 | AGTR2 | AT2-R | Type-2 angiotensin II receptor |
| Q06278 | AOX1 | AOXA | Aldehyde oxidase |
| Q14155 | ARHGEF7 | RhoGEF7/β-Pix | Rho guanine nucleotide exchange factor 7 |
| P16615 | ATP2A2 | SERCA2 | Sarcoplasmic/endoplasmic reticulum calcium ATPase 2 |
| O73707 | Cacna1C | CaV1.2 | Voltage-dependent L-type calcium channel subunit alpha-1C |
| O43497 | Cacna1g | CaV3.1 | Voltage-dependent T-type calcium channel subunit alpha-3G |
| O95180 | Cacna1h | CaV3.2 | Voltage-dependent T-type calcium channel subunit alpha-3H |
| P02462 | COL4A1 | Col4a1 | Collagen alpha-1(IV) chain |
| P10606 | COX5B | COX5B | Cytochrome c oxidase subunit 5B, mitochondrial |
| P25101 | EDNRA | ETA-R | Endothelin receptor type A |
| P24530 | EDNRB | ETB-R | Endothelin receptor type B |
| P03372 | ESR1 | ER | Estrogen receptor |
| O95967 | FBLN4 | Fibulin-4 | EGF-containing fibulin-like extracellular matrix protein 2 |
| Q14643 | ITPR1 | InsP3R1 | Inositol 1,4,5-trisphosphate receptor type 1 |
| P63252 | KCNJ2 | Kir2.1 | Inward rectifier potassium channel 2 |
| Q12791 | KCNMA1 | BKCa α-subunit | Calcium-activated potassium channel subunit alpha-1 |
| Q16787 | LAMA3 | Laminin α3 | Laminin subunit alpha-3 |
| P55001 | MFAP2 | MFAP-2 | Microfibrillar-associated protein 2 |
| Q08431 | MFGE8 | MFG-E8 | Lactadherin |
| P08253 | MMP2 | MMP-2 | Matrix metalloproteinase-2 |
| P14780 | MMP9 | MMP-9 | Matrix metalloproteinase-9 |
| Q6WCQ1 | MPRIP | M-RIP/p116RIP | Myosin phosphatase Rho-interacting protein |
| O14950 | MYL12B | MLC20 | Myosin regulatory light chain 12B |
| Q9ULV0 | MYO5B | MYO5B | Unconventional myosin-Vb |
| P19838 | NFKB1 | NF-κB | Nuclear factor NF-kappa-B p105 subunit |
| P29474 | NOS3 | NOS3/eNOS | Nitric oxide synthase 3 |
| Q9NPH5 | NOX4 | NOX4 | NADPH oxidase 4 |
| P08235 | NR3C2 | MR | Mineralocorticoid receptor |
| P47900 | P2RY1 | P2Y1-R | P2Y purinoceptor 1 |
| Q13177 | PAK2 | PAK-2/p58 | Serine/threonine-protein kinase PAK 2 |
| P26678 | PLN | PLB | Cardiac phospholamban |
| P43088 | PTGFR | PGF2α-R | Prostaglandin F2-alpha receptor |
| Q13464 | ROCK1 | ROCK1 | Rho-associated protein kinase 1 |
| O75116 | ROCK2 | ROCK2 | Rho-associated protein kinase 2 |
| Q92736 | RYR2 | RYR2 | Ryanodine receptor 2 |
| P00441 | SOD1 | Sod1 | Superoxide dismutase [Cu-Zn] |
| Q13586 | STIM1 | STIM1 | Stromal interaction molecule 1 |
| P21731 | TBXA2R | TXA2-R | Thromboxane A2 receptor |
| A0A499FJK2 | TGFB1 | TGFβ1 | Transforming growth factor beta |
| P37173 | TGFBR2 | TGFR-2 | TGF-beta receptor type-2 |
| P21980 | TGM2 | TGM2 | Protein-glutamine gamma-glutamyltransferase 2 (Transglutaminase 2) |
| P07996 | THBS1 | TSP-1 | Thrombospondin-1 |
| P01375 | TNF | TNFα | Tumor necrosis factor |
| Q9Y210 | TRPC6 | TrpC6 | Short transient receptor potential channel 6 |
| Q8TD43 | TRPM4 | TrpM4 | Transient receptor potential cation channel subfamily M member 4 |
| Q9H4B7 | TUBB1 | Tubulin β1 | Tubulin beta-1 chain |
| Q9GZT5 | WNT10A | Wnt-10a | Protein Wnt-10a |
| P56704 | WNT3A | Wnt-3a | Protein Wnt-3a |
| P41221 | WNT5A | Wnt-5a | Protein Wnt-5a |
| Q9NUD5 | ZCCHC3 | ZCHC3 | Zinc finger CCHC domain-containing protein 3 |
Table 2.
List of significantly most enriched KEGG pathways (ranked according to P-value).
| Pathway term | % occurrence* | P-value | FDR** |
|---|---|---|---|
| Calcium signaling pathway | 26 | 2,80E-09 | 2,80E-07 |
| Proteoglycans in cancer | 24 | 3,90E-09 | 2,80E-07 |
| cGMP-PKG signaling pathway | 20 | 1,20E-07 | 5,60E-06 |
| AGE-RAGE signaling pathway in diabetic complications | 16 | 6,10E-07 | 1,80E-05 |
| Diabetic cardiomyopathy | 20 | 6,30E-07 | 1,80E-05 |
| Pathways in cancer | 28 | 1,40E-06 | 3,20E-05 |
| Platelet activation | 16 | 2,60E-06 | 5,20E-05 |
| Renin secretion | 12 | 2,40E-05 | 4,20E-04 |
| Relaxin signaling pathway | 14 | 4,40E-05 | 6,80E-04 |
| Pathways of neurodegeneration—multiple diseases | 22 | 1,10E-04 | 1,50E-03 |
| Cushing syndrome | 14 | 1,20E-04 | 1,50E-03 |
| Dilated cardiomyopathy | 12 | 1,60E-04 | 1,90E-03 |
| Focal adhesion | 14 | 5,20E-04 | 5,60E-03 |
| Alzheimer disease | 18 | 6,10E-04 | 6,10E-03 |
| Oxytocin signaling pathway | 12 | 1,00E-03 | 9,70E-03 |
*: % of age-dependent proteins occurring in the respective pathways. **: False Discovery Rate.
Table 3.
List of significantly most enriched REACTOME pathways (ranked according to P-value).
| Pathway term | % occurrence* | P-value | FDR** |
|---|---|---|---|
| Muscle contraction | 5 | 1.40e-07 | 5.81e-05 |
| EPH-Ephrin signaling | 8 | 6.89e-07 | 1.43e-04 |
| Extra-nuclear estrogen signaling | 8 | 4.91e-06 | 6.77e-04 |
| Signal Transduction | 1 | 7.76e-06 | 8.07e-04 |
| Ion homeostasis | 9 | 1.17e-05 | 9.67e-04 |
| Signaling by GPCR | 2 | 2.06e-05 | 0.001 |
| GPCR ligand binding | 2 | 3.82e-05 | 0.002 |
| Signal amplification | 11 | 4.11e-05 | 0.002 |
| Cardiac conduction | 5 | 7.34e-05 | 0.003 |
| Smooth Muscle Contraction | 9 | 9.89e-05 | 0.004 |
| Nuclear Receptor transcription pathway | 6 | 1.05e-04 | 0.004 |
| Hemostasis | 2 | 1.11e-04 | 0.004 |
| Platelet homeostasis | 6 | 1.16e-04 | 0.004 |
| Signaling by Receptor Tyrosine Kinases | 2 | 1.57e-04 | 0.005 |
| Axon guidance | 2 | 1.75e-04 | 0.005 |
*: % of age-dependent proteins occurring in the respective pathways. **: False Discovery Rate.
Table 4.
Predicted Functional Partners with proteins in STRING network (very high confidence):.
| Gene name (Homo sapiens) | Protein name | Score |
|---|---|---|
| BCL3 | B-cell lymphoma 3 protein | 0.999 |
| COX6A1 | Cytochrome c oxidase subunit 6A1, mitochondrial | 0.999 |
| DCN | Decorin | 0.999 |
| NFKBIA | NF-kappa-B inhibitor alpha | 0.999 |
| PFN1 | Profilin-1 | 0.999 |
| RELB | Transcription factor RelB | 0.999 |
| TGFB3 | Transforming growth factor beta-3 proprotein | 0.999 |
| TIMP1 | Metalloproteinase inhibitor 1 | 0.999 |
| TNFRSF1A | Tumor necrosis factor receptor superfamily member 1A, membrane form | 0.999 |
| UQCRC1 | Cytochrome b-c1 complex subunit 1, mitochondrial | 0.999 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



