Submitted:
30 January 2024
Posted:
31 January 2024
You are already at the latest version
Abstract
This overview initially describes insect immune reactions and then brings together present knowledge of the interactions of vector insects with their invading parasites and pathogens. It is a way of introducing this special issue with subsequent papers presenting the latest details of these interactions in each particular group of vectors. Hopefully, this paper will fill a void in the literature since brief descriptions of vector immunity have now been brought together in one publication and could form a starting point for those interested and new to this important area. Descriptions are given on the immune reactions of mosquitoes, blackflies, sandflies, tsetse flies, lice, fleas and triatomine bugs. Cellular and humoral defences are described separately but emphasis is made on the co-operation of these processes in the completed immune response. The paper also emphasises the need of great care in extracting haemocytes for subsequent study as appreciation of their fragile nature is sometimes overlooked with non-sterile media, smearing techniques and excessive centrifugation sometimes used. The potential vital role of eicosanoids in the instigation of many of the immune reactions described is also discussed. Finally, the priming of the immune system, mainly in mosquitoes, is considered and one possible mechanism is presented
Keywords:
insect vector immunity
; Anopheles
; Aedes
; Culex
; blackflies
; sandflies
; tsetse flies
; lice
; fleas
; Rhodnius
; mosquitoes
; malaria
; sleeping sickness
; leishmaniasis
; Chagas disease
; filariasis
; onchocer
1. Introduction*- The Good, the Bad and the Incredible about the Immune Reactions of Vector Insects against Parasites and Pathogens
The “good” thing about insect immunity is that rapid progress has been made in understanding its functional roles at the molecular level. This increased knowledge has also been applied to understanding the interaction of vector species with invading pathogens and parasites.
There are a large number of reviews in the last 10 years related to insect vector immunity including e.g. [1,2,3,4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31]. Most of these deal with a single group, genus or species of vector insects or even just with one key aspect of immunity such as the antimicrobial peptides (AMP), signal molecules or infections caused by one particular parasite or pathogen. Herein, by way of introduction, we briefly describe the immune reactions of a range of vector insects to invading parasites and pathogens in order to gain some idea about possible common reactions and responses as a result of infection. Subsequently, many of these interactions are described with greater details in other papers in this special issue, and should help identify where additional research is required.
The general view of vector insects is that they are all “bad” due to the diseases they transmit. There are, however, some “good” aspects of their existence. For example, although there are about 3,700 species of mosquitoes [32], the majority do not bite or transmit disease but, especially in wetlands, form key elements in the food chain of aquatic and terrestrial predators including fish, newts, bats and birds [33]. In addition, during their quest for nectar as food-sources, mosquitoes are effective plant pollinators [34]. Furthermore, the saliva of blood-feeding vector insects, such as anopheline mosquitoes, tsetse flies and triatomines produce peptidic anticoagulant molecules with unique trivalent properties to inhibit thrombin and potentially act as sources of unique drugs for treating clotting diseases in humans [35].
Despite these positive aspects of insect vectors, and as their name indicates, they are regarded as bad due to disease transmission to humans and animals resulting from adaptation of invading organisms to manipulate and overcome the vector immune defences. Actual numbers of insect vectors species are probably confined to hundreds [36], in contrast to the recent estimated 5.5 million insect species on Earth [37].
2. Diseases Vectored by Insects
The orders Diptera and Hemiptera contain many species transmitting parasites and pathogens causing diseases such as malaria, leishmaniasis, Chagas disease, sleeping sickness, filariasis, onchocerciasis and arboviruses (Table 1). The dipterans include the mosquitoes (Culicidae) as well as blackflies (Simuliidae), sandflies (Phlebotominae), tsetse flies (Glossinidae) and gnats (eg. Sciaridae). Mosquitoes belonging to the genera Anopheles, Aedes and Culex, vector many etiologic agents of diseases, including malaria, and the arboviruses, namely, yellow fever, dengue, zika, chikungunya, West Nile fever, Japanese encephalitis, and filarial nematodes (Table 1).
Mosquitoes alone kill ca. 1 million each year, with those due to malaria having occurred for many decades, with deprived children < 5 years particularly vulnerable [38,39]. Mosquitoes also vector dengue fever which is the most common viral disease, transmitted by Aedes spp., and infecting 3.9 billion people each year in 129 countries, resulting in about 40,000 deaths [38,40]. Sandflies are vectors of Leishmania transmitted in Africa, Americas (Brazil), Middle East, South Asia, and the Mediterranean. Leishmaniasis, together with onchocerciasis and filariasis transmitted by blackflies and mosquitoes, respectively, result in permanent disfigurement in infected people. In Africa, the tsetse flies, Trypanosoma brucei rhodesiense and T. b. gambiense, cause sleeping sickness in those infected (Table 1). The economic burden and human suffering caused by these diseases are enormous [41].
Other important diseases vectored by dipterans include lymphatic filariasis and river blindness (onchocerciasis) vectored by mosquitoes and blackflies, respectively (Table 1). Filarial worms are nematodes transmitted by mosquitoes and include 3 species, Wuchereria bancrofti, Brugia malayi and Brugia timori. Infection occurs when parasite larvae are deposited in the skin following biting by mosquitoes vectoring the parasites. The parasites penetrate into the lymphatic vessels to become adults and result in inflammation and tissue damage leading to elephantiasis and other symptoms [42]. In 2000, WHO began a ”Global Programme to Eliminate Lymphatic Filariasis” which has reduced the need to use preventative chemotherapy for 740 million people [42]. Blackflies, Simulium spp., transmit onchcerciasis caused by the filarial nematode Onchocerca volvulus. When infected blackflies bite humans, they transmit parasite microfilariae and the resulting adults reside under the skin in nodules. The adults produce masses of microfilaiae which migrate through the body and can infect the eyes and optic nerves leading to blindness [43]. Great progress is being made in controlling and eliminating this dreadful disease in many countries such as Columbia, Ecuador, Mexico, Uganda and Sudan [44].
Hemipterans infect fewer people than dipterans and result in a reduced disease impact with the parasites vectored. Triatominae, including Rhodnius prolixus and Triatoma infestans, transmit the flagellate protozoan Trypanosoma cruzi, resulting in Chagas disease throughout Latin America and more recently the USA (Table 1). The disease pathology includes chronic inflammation of the heart, colon and nervous system. Of the ca. 6 million people infected with T. cruzi, about one third will die [45].
Fleas and lice (Table 1) have been particularly devastating insect vectors in the past. Plague, vectored by rat fleas, has resulted in the most calamitous pandemics of mankind. “The Black Death” occurred over centuries in China and then swept through Europe leading to the Great Plague of London in 1665-66 [46]. The bacteria immobilise human innate immunity leading to multi-organ failure, lung infections, and death. Foci of plague still remain throughout the World with recent outbreaks recorded [47]. Lice have also been associated with humans for millions of years and epidemic typhus caused by the body louse, Pediculus humanus, resulted in some of the worst pandemics recorded. Nowadays, body lice are mainly associated with allergic reactions in deprived populations although modern wars still pose a risk for the re-emergence of epidemic typhus [48].
Unfortunately, global warming has increased the emergence and/or spread of insect vector-borne parasitic/pathogenic diseases. In 1990, for example, the highly invasive Asian tiger mosquito, Aedes albopictus, was probably introduced into Europe via Italy with imported vehicle tyres. The climate was suitable and the mosquitoes spread and vectored outbreaks of chickungunya and dengue imported into Europe by international travellers [49]. The rapid spread of emerging pathogens can also be explosive, as with the Zika virus pandemic in Brazil in which the virus first appeared in 2015 but by 2016 had infected ca. 211,700 people [50].

Table 1.
The main insect vectors, their distribution and the diseases transmitted .
| Vectors | Diseases | Pathogens | Distribution | At Risk |
|---|---|---|---|---|
| Aedes, Anopheles , Culex, Mansonia | Lymphatic filariasis | Nematode worms Brugia spp., and Wucheria bancrofti | Tropical and subtropical regions of SE Asia, Central and South America, Africa, West Pacific | 882 million |
| Aedes | Dengue | Flavivirus | Tropical, subtropical and spreading to Europe | 3.9 billion in 129 countries |
| Aedes | Yellow fever | Flavivirus | Endemic in tropical regions of Africa and Latin America | 900 million |
| Aedes | Chikungunya | Alphavirus | Tropical, subtropical and temperate regions. | ¾ of the World population at risk. |
| Aedes | Zika | Flavivirus | The Americas, Europe, India and 89 countries | Over 2 billion at risk |
| Anopheles complex with 484 recognised species but An.gambiae carries the deadliest form. | Malaria | Protozoan parasite with 5 Plasmodium species. | In 2021, the African region carried 95% of cases | Nearly half the World was at risk of malaria in 2021 |
| Culex spp. | Arboviruses | West Nile Virus, (Both Flaviviruses) Japanese encephalitis |
USA, Canada, Caribbean, Central and South America. South East Asia and West Pacific |
These and other arboviruses (Zika etc) risk emerging pandemics |
| Blackflies Simulium spp |
Onchocerciasis River blindness | Vector-borne nematode worms eg. Onchocercas volvulus | 99% in Africa but also foci in Brazil, Venezuela and Yemen | 123 million |
| Sandflies Phlebotomus spp and Lutzomyia spp. | Leishmaniasis | Protozoan parasite more than 20 species | Africa, Americas (Brazil), Middle East, South Asia, Mediterranean | 99 countries |
| Tsetse flies Glossina spp. |
African trypanosomiasis | Protozoan parasites, Trypanosoma brucei | Sub-Saharan Africa | 55 million people but control now and less than 1000 cases in 2022 |
| Lice Pediculus humanus and Pthirus pubis |
Louse-borne typhus and severe allergic reactions | Rickettsiae Rickettsia prowazekii |
Epidemics in wars, prisons and refugee camps in colder regions | Global diseases of poverty |
| Fleas Xenopsylla cheopis |
Plague | Gram-negative bacterium Yersinia pestis | Associated with close living in deprived areas with rat infestations | Global distribution |
| Triatomine bugs Rhodnius and Triatoma spp. | Chagas disease | Protozoan Trypanosoma cruzi | Mainly South, Central America but also North America now | 70 million in the Americas |
The “incredible” aspect of insect immunity, which embraces vector species too, is derived from studies of non-self recognition processes involved in insect interactions with parasites and pathogens. In particular, work with Drosophila showed that the insect Toll transmembrane receptor was involved in antifungal defence [51], and this led to the discovery of mammalian Toll homologues [52]. Thus, immune recognition and signalling processes in insects have close parallels with innate immune reactions in humans, and this is leading to innovative human disease therapies. For example, the field of human adjuvants for the production of vaccines is being revolutionised by the discovery of Toll-like receptors in humans with implications not only in vaccine production but also in immuno-therapies for many diseases including Alzheimer´s, allergies, cancer, and drug addiction [53]. Who would have imagined that diminutive Drosophila fruit flies could be the source of major new information on immune functioning and innovative therapies in human diseases including COVID-19 [54]? This also emphasises the wisdom of using Drosophila and developing new insect-based models, such as Galleria, Sarcophaga, Manduca and Rhodnius, as powerful tools for dissecting out many aspects of the human immune responses eg. [55].
Finally, the incredible complexity of vector – parasite/pathogen interactions must be emphasized and “The Biology of Blood-Sucking in Insects” [56] is still highly recommended for more background details of this. For example, variations in immune reactivity can occur with the same parasite species in different members of the same host population [57]. There are also great variations in immunity in the same host species exposed to different pathogens [58], probably related, in part, to co-infections.
There are numerous determinants of successful infection or parasitisation (Figure 1). These include resistance to environmental stressors such as changes in the ambient temperature and exposure to xenobiotics, as well as previous invasions by parasites/pathogens [59]. The extent of networking occurring with other species in complex ecological communities will also affect host finding, biting and disease transmission [36]. Other determinants include the sex and developmental stage of the vector [60,61], as well as physiological factors such as the host nutrition [62] and the fitness costs of immunity [63]. In addition, the route of entry and survival in the host will also dictate the nature of the immune response [64]. The immune process itself against the invader is also not simple but, as shown in mosquitoes, composed of a tripartite response involving interplay between the gut microbiome, immune defence reactions and invading parasites [18,29]. Recently too, evidence is accumulating in insects for the presence of immune priming whereby an initial low dose infection provides protection against a subsequent potentially lethal dose [65]. This latter could partially explain variations in immune capability within a population composed of insects of different ages.
3. Typical Insect Immune Scenario
To understand the dynamics of vector immunity to invading parasites and pathogens it is essential to understand the initial processes involved in gaining access to particular insect host tissues. Different invaders adopt different strategies for entry although routes are mainly limited to the outer cuticle and various epithelia lining the cuticle, gut, reproductive system and tracheae. Most human parasites/pathogens are ingested during blood feeding by vectors with biting mouthparts, but the subsequent events in the host vary significantly. Thus, African trypanosomes, T. cruzi, Leishmania spp. and Yersinia pestis remain and develop within the confines of the midgut, while Plasmodium spp., Trypanosoma rangeli, arboviruses and human parasitic nematodes, migrate out of the gut and colonise and develop in other vector organs [66]. Problems of parasite/ pathogen survival due to physiological incompatibilities and robust host innate immune responses have to be overcome and this topic is considered in more detail in and in other papers in this special issue. One example of the effect of these problems is illustrated by African trypanosomes in which, in some host/parasite combinations, less than 1% of parasites survive in the tsetse fly vector by 3 days post-infection [67].
The basic insect immune response can be elicited in response to foreign invasion by wounding or in the gut, the hemocoel, the haemocytes, the fat body, the salivary glands and other tissues following ingestion. Recognition is mediated by means of Pattern Recognition Receptors (PRRs) located on/in immune tissues which bind to Pathogen Associated Molecular Patterns (PAMPs) of invading parasites and microbes. This recognition and binding can directly activate innate immune responses like phagocytosis and also initiate various signalling pathways leading to transcription of immune effector genes and the secretion of immune proteins/cytokines for eliminating invaders [68]. The insect effector immune factors include antibacterial peptides (AMPs), lectins, prophenoloxidases (PPO, for melanisation), reactive oxygen species (ROS), nitric oxide (NO), antiviral factors, cytokines and many more [17,68,69]. Much of the knowledge of these processes was gleaned by work on Drosophila eg. [70,71,72] while valuable contributions have also been made with other species including Aedes/Anopheles mosquitoes eg. [18,29,73,74,75], Hyalophia cecropia eg. [76], Manduca eg. [77,78], Galleria [79,80,81] and Bombyx [82,83] moths, Sarcophaga fleshflies [84,85] and Rhodnius assassin bugs [86,87].
Insect immunity is usually described as consisting of interacting humoral and cellular elements. The humoral components, often derived from the immune tissues, involve immune proteins and pathways, mentioned above, such as the antimicrobial peptides and melanisation reaction, while cellular elements include coagulation, phagocytosis, nodule formation and encapsulation-type responses [17,68,69].
4. Vector Cellular Immunity
Central to many of the immune responses are the haemocytes which can be free in the circulation or sessile and associated with tissues such as the heart ostia of mosquitoes [68]. One recurring problem for many workers has been the extraction and identification of the types of haemocytes present. Previous work on haemocyte classification [88,89,90] is still helpful in overcoming these problems. Some haemocytes types are extremely fragile, including those from vector species, usually degranulate and breakdown rapidly on exposure to air. The use of an anticoagulant solution stabilises the cells and allows examination under phase optics to observe both the structure and some functions of these cells. In addition, with an anticoagulant it has been possible to separate Galleria mellonella cell types [91], study their interactions in vitro [92] and stain specific cell types with monoclonal antibodies [93]. Smearing the insect blood and staining with Giemsa, as reported constantly, is not recommended.
4.1. Haemocyte Types
The huge diversity of insects is also reflected in variations in the form of the different types of haemocytes present, although the use of phase observations of stabilised cells can distinguish which cells are phagocytic or involved in coagulation reactions due to granule discharge. Studies on 15 insect orders identified the main haemocyte types as prohemocytes, plasmatocytes, granular cells, oenocytoids, spherule cells and coagulocytes, with the number of these classes present varying with different insect species and during various stages of development [89]. The recent ability to study the function of single haemocytes [16,94,95,96,97], using single cell RNA sequencing (scRNA-seq) combined with gene silencing and functional studies, is a breakthrough in typing these cells. In Drosophila, scRNA-seq has identified many subpopulations of plasmatocytes at different stages of development [94,95], while in mosquitoes immune cell subsets, their differentiation and lineages have also been defined [16,96,97].
Haemocytes in vectors have mainly been studied in mosquitoes and triatomines. In mosquitoes, 3 main cell types, ie. prohaemocytes, granulocytes and oenocytoids, have previously been described (Table 2), but the study of haemocyte classification, as mentioned above, has been revolutionized by RNA-seq and other modern molecular techniques. Thus, subsequently, in mosquitoes, the haemocytes identified are in subtypes of these 3 haemocyte classes. Thus, Kwon et al. [97] identified 7 haemocyte subtypes in Anopheles gambiae with 4 different populations of granulocytes, 2 populations of oenocytoids, and the prohaemocytes. These populations were identified using scRNA-seq analysis combined with homology studies, phagocytosis and phagocyte-depletion assays. Severo et al. [96] and Raddi et al. [16] used similar methods in classifying mosquito haemocyte subtypes. Severo et al. [96] only detected 2 subtypes, granulocytes and oenocytoids, while the results of Raddi et al. [16] complemented those of Kwon et al. [97], with the addition of another cell type designated as megacytes.
We have now entered a new era with the use of advanced molecular, labelling and functional techniques. In addition, detailed analysis of haemocyte ontogeny is available showing the functional plasticity of subpopulations of previously statically classified haemocyte types. With these new techniques great care must be taken to ensure that the perfused haemocytes remain stable, especially in manipulations involving labelling and cell counting techniques. In addition, checks should be made to confirm that perfusion removes any large populations of sessile haemocytes attached to various organs in the body. Examination of the methods of some recent papers clearly indicates that the haemocytes are often perfused with non-steriile solutions, magnetic beads etc and placed for many minutes to attach on slides, and then treated with various antibodies or non-sterile probes. Even if perfusion is made with anticoagulant solutions, some haemocyte types will still react with any PAMPS in solutions and the environment to discharge their contents and elicit coagulation-like responses. The cells also often breakdown after degranulation with attachment to or ingestion by other cells. This will likely result in errors in identification of cell types and shows a basic lack of understanding of the fragility of some insect immune cells. The literature is full of accounts of the rapid discharge and breakdown of insect cell types on exposure to environmental factors like the PAMPS. One source of PAMPS such as endotoxin could be the water from stills and deionizing columns which may not fully remove endotoxin [98]. Endotoxin from Gram-negative bacteria can activate mammalian leukocytes at concentrations as low as 0.01 ng/ml. Anticoagulant solutions also reduce the viability and attachment of haemocytes in other arthropod immune systems [99].
Table 2.
Haemocyte types* in vector insects mainly based on morphological and functional studies.
 |
* The papers cited are mainly those with some identification of the types of haemocytes studied. 1. Mosquitoes, Hall [100]; Castillo et al. [101]; Hyllier and Strand, [102]. Kown et al. [97] used RNA-seq analysis and showed 7 subpopulations of hemocytes in An. gambiae. 2. Blackflies, Silva et al. [103]; Luckhart et al. [104] also described spherulocytes; Cupp et al. [105] reported small and large granule cells. 3. Sandflies, De Albuquerque et al. [106]. 4. Tsetse flies, Matetovici et al. [12] record melanin-producing crystal cells; East et al. [107] and Kaaya et al. [108] described spindle cells and thrombocytoids too. 5. Lice, Coulaud et al. [109] described only phagocytes which are likely to be plasmatocytes or granulocytes. 6. Fleas, Kozlov et al. [110]; Munoz et al. [111] described only phagocytes which are likely to be plasmatocytes or granulocytes. 7. Triatomines, Azambuja et al. [112]; Moyetta et al. [113]. The granular cells were often giant cells and maybe ingested other cells.
The haemocytes of triatomines have been described in detail by some authors although there can be some variations from species to species and according to the preparation technique used [112]. In most studies, prohaemocytes, plasmatocytes, granular cells, oenocytoids and adipohaemocytes are recognised eg. [112,113] with giant cells and cystocytes also identified in Rhodnius prolixus and Dipetalogaster maxima [112,113] (Table 2). The more recent study of the haemocytes of D. maxima also used immunofluorescence and flow cytometry analysis and seemed to confirm the classification of cell types described above. Results of research on the haemocytes of triatomines using using scRNA-seq analysis combined with homology studies, phagocytosis and phagocyte-depletion assays are awaited, as utilised for Drosophila and mosquitoes [16,94,95,96,97].
Regarding blackflies, different species of blackflies are reported to have four types of haemocytes although these varied with the species studied (Table 2). Thus, Luckhart et al. [104] described prohaemocytes, plasmatocytes, granulocytes, and spherulocytes in Simulium, vittatum, while Silva et al. [103] described a similar profile except oenocytoids occurred instead of spherulocytes in Ectemnaspis rorotaense and Ectemnaspis trombetense. For these studies, an anticoagulant solution was perfused before harvesting the haemolymph and placing on slides for air drying before staining. A process very likely inducing haemocyte degranulation and lysis, as described above.
Studies on sandfly haemocytes are wanting except for that of Albuquerque [106] on Lutzomyia migonei for which five haemocyte types were identified, namely, prohaemocytes, plasmatocytes, granulocytes, adipohaemocytes, and “eonocites” (oenocytoids?) (Table 2). The insects were perfused with anticoagulant and slides air dried and stained with Giemsa, so this is yet another example of possible loss of cell types. Anticoagulant is not a fixative and haemocytes can attach to slides in such solutions and combined with air drying change dramatically.
Tsetse fly haemocytes have been described by East et al. [107] and Kaaya et al. [108]. These are prohaemocytes, plasmatocytes, granulocytes, and adipohaemocytes in all stages, as well as thrombocytoids and spindle cells in adults (Table 2). These papers were published before the introduction of anticoagulants. Despite the apparent lack of oenocytoids, the haemolymph of tsetse flies does melanise (see section 4. 2. Cellular Defence Mechanisms, below), and subsequently melanin-producing crystal cells have been recorded (equivalent to oenocytoids of other insects) by Matetovici et al. [12].
Regarding lice, little is known about haemocyte types, all of which are described as phagocytes by Coulaud et al. [109] who collected the haemolymph using an insulin syringe and then added this to various culture media obviously under sterile conditions. Cells survived well and phagocytosed microbes but probably did not include all haemocyte types present in vivo. In fleas, Kozlov et al. [110] identified proleukocytes, leukocytes, trophic cells and oenocytoids while Munoz et al. [111] termed all flea haemocytes as phagocytes and did not distinguish cell types.
4.2. Cellular Defence Mechanisms
These include coagulation, phagocytosis, nodule formation/encapsulation, apoptosis, autophagy and extracellular traps. Dividing immunity into cellular and humoral reactions is often arbitrary as these components frequently interact in most immune responses.
4.2.1. Coagulation
Coagulation of insect haemolymph occurs in most studies of different species and is an essential process limiting the excess loss of haemolymph and preventing the entry of parasites and pathogens. Haemolymph coagulation has been reported in mosquitoes [114], triatomines [112,113], blackflies [104], tsetse flies [12], and lice [115]. It basically occurs in two phases, initially involving the cross-linking of clot components from the plasma and haemocytes and depending on transaminase activity followed by phenoloxidase hardening and melanising the soft clot material [116]. In mosquitoes, the process involves aplolipophorin-1 and phenoloxidase with the latter more important than in Drosophila [114]. Recent studies on haemolymph coagulation in vector insects are very limited and in blackflies not mentioned directly but can be interpreted in the figures published [104]. In addition, in lice, the possibility of haemolymph coagulation is inferred by transcription studies detecting the humoral immune-related genes hemocytin and noduler [115], since these are associated with sticky fibrous structures exocytosed from granulocytes and nodule formation in other insects [117]. The presence of haemolymph clotting in fleas has also been implied [111]
4.2.2. Phagocytosis
Phagocytosis by insect haemocytes has been widely reported as an important defence process in vector insects. Detailed studies of the interaction of the haemocyte cell surface and free pattern recognition receptors (PRRs) with the pathogen-associated molecular patterns (PAMPs), as well as the role of plasma opsonins, have mainly been confined to mosquitoes. The comprehensive studies published on the immune system of the mutants of Drosophila melanogaster provide useful guides for comparison with mosquitoes as does RNA interference (RNAi) utilising the multiple mosquito genome sequences available [118]. Some modest progress, however, has recently been made in understanding haemocyte-mediated phagocytosis in other vector insects [104,105,110,111,115,119].
Details of mosquito phagocytic interactions with parasites and pathogens are given in other papers in this special issue and have been published previously [15,102,118,120,121,122,123,124,125]. Granulocytes are the main phagocytes in mosquitoes and comprise about 90% of the haemogram of which there are ca.2-5,000 cells [126]. These haemocytes are either free in circulation or form sessile concentrations, the periosteal cells, around the heart valves (ostia) in adults, and in tracheal tuffs in larvae [121]. Both free and sessile haemocytes rapidly phagocytose small particulates including viruses, bacteria, fungal elements and protozoan parasites.
The recognition and phagocytosis of invading pathogens and parasites depends upon a variety of putative PRRs (more details of recognition are given in section 6 on Recognition, below). These include the thioester-containing proteins (TEPs) identified in D. melanogaster, An. gambiae, and Aedes aegypti and which are mostly free in the haemolymph. One of these, TEP1, is secreted into the haemolymph and enhances phagocytosis following its activation by LRIM1 and APL1C to form a complex binding to and destroying bacteria and Plasmodium ookinetes [122]. Numerous other components of the mosquito immune system, including families of proteins, such as fibrinogen-related proteins (FREPs), C-type lectins and Gram-negative binding proteins (GNBPs), as well as the signalling pathways, have been characterised in mosquitoes and influence the activity of the immune response to parasite/pathogen invasion [122] (see section 5. Vector Humoral Immunity, below).
A fascinating study on the mosquito responses to Plasmodium has shown that infection results in recruitment of additional periosteal haemocytes to amplify the heart-related immune response. This results from the upregulation of the IMD and JNK signalling pathways (see section 6 on Recognition, below) which enhances this periosteal haemocyte aggregation, activating phagocytosis and melanisation on the heart, thereby demonstrating the integration of the response between the immune and circulatory systems [125]. The freely circulating haemocytes were not affected in a similar way. Finally, recent work with dengue and Zika infected Ae. aegypti mosquitoes showed that phagocytosis by haemocytes is not needed for controlling viral infection in the midgut but is vital for restricting systemic viral dissemination [124].
Information on phagocytosis in other vector insects has been published, although the details available are often limited, and may be related to whether the invading parasites/pathogens are confined to the gut or invade other tissues in the body.
- In blackflies, phagocytosis of erythrocytes and bacteria is mediated by plasmatocytes and granulocytes with evidence of these cells also being involved in remodelling tissues [104].
- With sandflies, Leishmania development occurs exclusively in the gut with research mainly confined to this organ and interaction with the microbiota [8], so that consideration of the possible role of the haemocytes has been neglected. Lutzomyia longipalpis embryonic cell lines used for studying innate immunity in sandflies, however, have active Toll and Imd pathways and internalisation of Leishmania parasites was reported [127], so that the haemocytes may well be activated following infection. In lice and fleas more information on phagocytosis has been published recently.
- In head and body lice, the relative phagocytic activities of the haemocytes have been compared following injections of Escherichia coli or Staphylococcus aureus and showed that the body lice had a reduced immune response compared to the head lice [115]. In addition, haemocytes have been identified engulfing endosymbionts during their migrations around the body of the lice [128]. The reduced phagocytic competence of the body lice may be related to the increased pathogen vectoring capacity of these insects [128]. The presence in lice of the genes for the main signalling pathways, except Imd, may indicate the potential for activation of the haemocytes [129].
- The phagocytic activity of fleas has been the subject to similar research to that in lice [111], so that following inoculations of E. coli the phagocytic activity of the haemocytes increased significantly. This was accompanied by a general enhancement of antimicrobial resistance of the haemolymph, probably also involving humoral immune factors induced via signalling pathways [130] (see section 5. Vector Humoral Immunity, below).
- ∙ Two other major vectors in which phagocytosis has been recorded are the tsetse flies, Glossina spp. and the triatomines, Rhodnius and Triatoma. The main parasites involved in these insects are the African trypanosomes and T. cruzi, respectively. These parasites are mainly confined to the vector gut, although African trypanosomes do migrate in the vector during maturation [12], and Rhodnius also hosts T. rangeli which invades the haemocoel [131].
The tsetse fly association with the invading African trypanosome is extremely complex since the phagocytic ability of the haemocytes against bacteria has been recorded [132], as has a specific genomic expansion within the thioester-containing protein family (TEPs) [133] which are usually regarded as opsonic for phagocytosis.
In a series of fascinating experiments, comparing tsetse fly survival following E. coli challenge of antibiotic-treated, aposymbiotic tsetse flies with wild-type flies transfused with either wild-type fly haemolymph separated into its cellular or soluble fractions, showed that all those flies receiving the soluble fraction died by 12 days while 62% of those with the cellular fraction survived. In this innovative study, much additional evidence confirmed the importance of cellular immunity in tsetse defences against foreign invaders [132]. The importance of this study with tsetse flies (6-16 mm length) should not be overlooked as the ability to repeat such experiments with smaller vector insects such as sandflies (ca. 3 mm), fleas (1–3.2 mm), lice (0.5–5 mm), blackflies (2-6 mm) or mosquitoes (3-6 mm) would be challenging.
The immune reaction of the tsetse flies, however, to trypanosome infection also includes other factors such as peptidoglycan recognition proteins (PGRPs), antimicrobial peptides (AMPs), a tsetse EP-protein, interactions with the Imd pathway, and the role of the symbiont, Wigglesworthia, on immune efficacy eg. [12,134] (see section 5. Vector Humoral Immunity, below, and in other papers in this special issue).
Most research on triatomines has been carried out on Rhodnius prolixus and considerable progress in understanding the complex interaction of T. cruzi with the insect vector is being made eg. [4,24,135,136,137,138,139]. Phagocytosis by the haemocytes in Rhodnius has been described following T. rangeli invasion from the gut into the haemolymph and following injection of bacteria into the haemolymph [140]. However, what is particularly interesting are experiments showing the induction in the haemolymph of an early systemic immune response following the colonization by T. cruzi of the vector gut. Therefore, communication of parasite invasion is transmitted rapidly to the innate immune system following oral infection [24]. This, of course, has implications for many other vector-parasite associations in which the parasites remain confined in the gut (see section 5. 7. Triatomines, below).
4.2.3. Nodules and Capsules
Similar to phagocytosis, nodule formation and encapsulation (often referred together as capsules) occur rapidly following invasion of a range of insects by parasites and pathogens. The encapsulation response may be humoral and melanotic, as in some dipterans with limited numbers of haemocytes, solely cellular as in lepidopterans like Galleria, or even formed by a combination of humoral and cellular responses [141].
The formation of these structures is controlled by interacting cytokines and effector molecules, details of which have been identified mainly in Drosophila, lepidopterans and non-vector insects eg. [17,72,142,143]. Nodules/capsules have been described as forming as a result of insect invasion by large numbers of microbes or by parasites too large to be ingested by a single haemocyte [141]. Recently, however, it has been shown that the initial haemocyte aggregation process, typical of capsules, can occur following contact with one or a small numbers of labelled yeast cells [142]. During this process, granule-containing cells stick together due to their release of a viscous material identified as hemocytin which is a homolog of the mammalian von Willebrand factor [142,143]. The degranulation of the hemocytes occurs after recognition of PAMPs by PRRs, activation of a haemolymph serine proteinase and Spätzle1, and with Toll signalling releasing 5 -HT and eicosanoids to aggregate the cells [142]. In addition, although good progress has been made in understanding aspects of phagocytosis and capsule formation in model insects, like Drosophila, Galleria and Bombyx, how exactly these early events trigger subsequent stages in capsule formation and humoral responses from the fat body and other tissue needs clarification. The work of Sato [142] discusses some of these issues in detail (see section 5. 7. Triatomines, below).
Regarding capsule formation in vector insects, details of this process are mainly confined to mosquitoes and triatomines. In mosquitoes, the invasion by Plasmodium and nematodes results in encapsulation-type responses surrounding the parasites. In refractory mosquitoes, the ookinetes become surrounded by a capsule which melanises and the parasites are lysed by a complement-related thioester-containing protein (TEP1) [144]. The capsule is composed of eumelanin and pheomelanin with 14 host proteins embedded including AgMesh with domains indicating a recognition function [145]. This is interesting as perhaps the capsule is not just an inert structure walling off lysed ookinetes but during formation may communicate with the immune system and activate a host response.
There have also been numerous studies on the encapsulation of filarial nematodes in mosquitoes, since these parasites include medically important species resulting in lymphatic filariasis, such as Wuchereria bancrofti, causing 90% of human cases, and Brugia malayi, and Brugia timori. These are transmitted mainly by Culex in urban regions, Anopheles in rural areas, and by Aedes in islands of the Pacific [42]. Another filarial nematode, Onchocerca volvulus, is transmitted by Simulium spp. blackflies and causes river blindness.
The nematode microfilariae are ingested by the Insect vectors during feeding on the blood of mammalian hosts and then penetrate the insect midgut epithelium to enter the haemolymph. They subsequently migrate to the thoracic musculature, invading the flight muscles before moulting twice into third-stage infective larvae. These migrate to the head to reach the proboscis where they can infect another human host during the wounding process of a blood meal [146]. It is during the migration of the microfilariae around the body in the haemolymph that the parasites encounter the cellular and humoral encapsulation reactions of the vector. These reactions include melanotic encapsulation of the microfilariae, often involving an initial deposition of melanin granules and cellular debris on the parasite surface, followed by plasmatocyte attachment in later stages [147].
The importance of melanin in mosquito defence against invading microfilariae may be indicated by the RNAi knockdown of phenylalanine hydroxylase (a melanin precursor) in Ae. aegypti and Armigeres subalbatus resulting in the reduced melanisation of invading Dirofilaria immitis microfilariae [148]. There are also many other studies confirming the important role of melanin and its precursor phenoloxidase (PO) enzymes in insect immunity. For example, in Drosophila, the genome encodes 3 precursor POs, designated as PPO1, PPO2 and PPO3, and flies were generated with deletions for PPO1 and/or PPO2. Analysis of these mutants alone and in combination identified the functions of both PPO1 and PPO2, but not PPO3, in melanisation of the haemolymph and in encapsulation of parasitoid eggs. This study also showed an essential role of melanisation in defence against some Gram-positive bacteria and fungi [149]. In addition, Ae.aegypti has 10 PPO genes, 4 of which are transcriptionally activated by Cactus silencing which also leads to the arrested development and death of Plasmodium gallinaceum. This research indicates that the PPO gene expression and its RUNT-related transcription factor 4 (RUNX4), are controlled by the Toll pathway (see section 6 on Recognition, below) and are important in restricting parasite development [150].
Here, we have included the encapsulation of nematodes in mosquitoes within section 4. 2. Cellular Defence Mechanisms. In fact, these capsules probably are generated by interactions between the cellular and humoral immune defences of the mosquito.
Recent work has revealed the true complexity of the mosquito immune response to nematode invasion. For example, very useful studies on mosquito vectors during the first 12 h of infection have compared the transcriptomic profiles of resistant and susceptible Ae. aegypti to B. malayi filarial nematodes. Differentially regulated genes were found including those involved in antimicrobial peptides, recognition proteins including lectins, signalling components of the Imd, Toll and JAK/STAT pathways, and serine proteases [151]. Other immune-related factors such as the generation of reactive oxygen and nitrogen radicals, the influence of symbiotic bacteria like Wolbachia on immunity, the role of parasite extracellular vesicles and avoidance mechanisms of the host response also need consideration [17,152,153,154]. Some of these are mentioned in section 5. Vector Humoral Immunity, below.
In triatomines, T. cruzi and T. rangeli form aggregates in the anterior midgut and in the haemolymph, respectively, of R. prolixus. The T. cruzi aggregates are formed entirely of parasites which develop confined to the gut and result from lectin present in the gut, and may serve to protect the parasites from the lytic activity present in this organ [155]. True melanising nodules consisting of parasites and haemocytes are only formed after invasion of T. rangeli from the gut into the haemocoel or by injection of bacteria [135]. In R. prolixus haemolymph, a galactose-binding lectin enhances nodule formation in vitro by T. rangeli [156]. Subsequently, this molecule was identified as a rhamnose-binding lectin with potential importance in the host defence of Rhodnius against T. cruzi and T. rangeli infections [157]. Although certain strains of T. rangeli appear to be broken down within nodules others survive and multiply modulating the host response [135]. This process may involve the effects of eicosanoids and platelet activating factor (PAF) on the PPO system and haemocyte aggregation [135]. More recently, a quantitative proteomics study of T. cruzi analysing the expression profiles of Rhodnius haemolymph proteins from 6 h to 24 h post- infection identified 12 novel immune proteins of unknown functions [24]. A similar approach following early T. rangeli infection would probably be just as rewarding and identify the differential expression of haemolymph proteins in response to T. rangeli compared with T. cruzi (see the detailed paper by Schaub et al. on triatomines in this special issue).
In Glossina, there are reports of encapsulation-type reactions towards injected bacteria or implants [12,132,158] with few details given except that the reaction appears to just involve a few haemocytes and with melanin deposition occurring.
Apart from the classical haemocyte defence reactions against invading parasites, described above for insect vectors, more recently, additional immune responses have been reported. These responses include RNA interference (RNAi), apoptosis, autophagy and extracellular traps [159,160]. RNAi, apoptosis and autophagy occur in response to viral infections whilst extracellular traps in Rhodnius deal with a range of microbes [159,160]. These reactions are not confined to haemocytes but more generally involve other tissue in the body such as the intestine, Malpighian tubules, fat body and salivary glands. More information on some these is available in the above references as well as in section 6 on Recognition.
5. Vector Humoral Immunity
Table 3 shows a summary of some of the more important aspects of the humoral immune factors reported in vector insects. These factors are conservative throughout the insects but are expressed and utilised to a greater or lesser extent from species to species and even within related species. The majority of the papers published on vector immune responses to parasites and pathogens are those dealing with humoral responses in mosquitoes, due to their transmission of malaria, with triatomines in second place a long way behind. The majority of the remaining vector insects have received scant attention and those published are mainly confined to humoral immunity since the vectors are either very small and contain few haemocytes (eg. sandflies) or else the parasites vectored do not invade the haemocoel (eg, sandflies, tsetse flies). One important aspect of the vector insect-parasite/pathogen interactions, rapidly becoming more evident, is the complexity of these associations so that only basic information can be summarised here with the reader referred to the additional papers in this special issue for details.
Table 3.
Basic summary of some important aspects of the humoral immune factors identified in vector insects*.
Table 3.
Basic summary of some important aspects of the humoral immune factors identified in vector insects*.
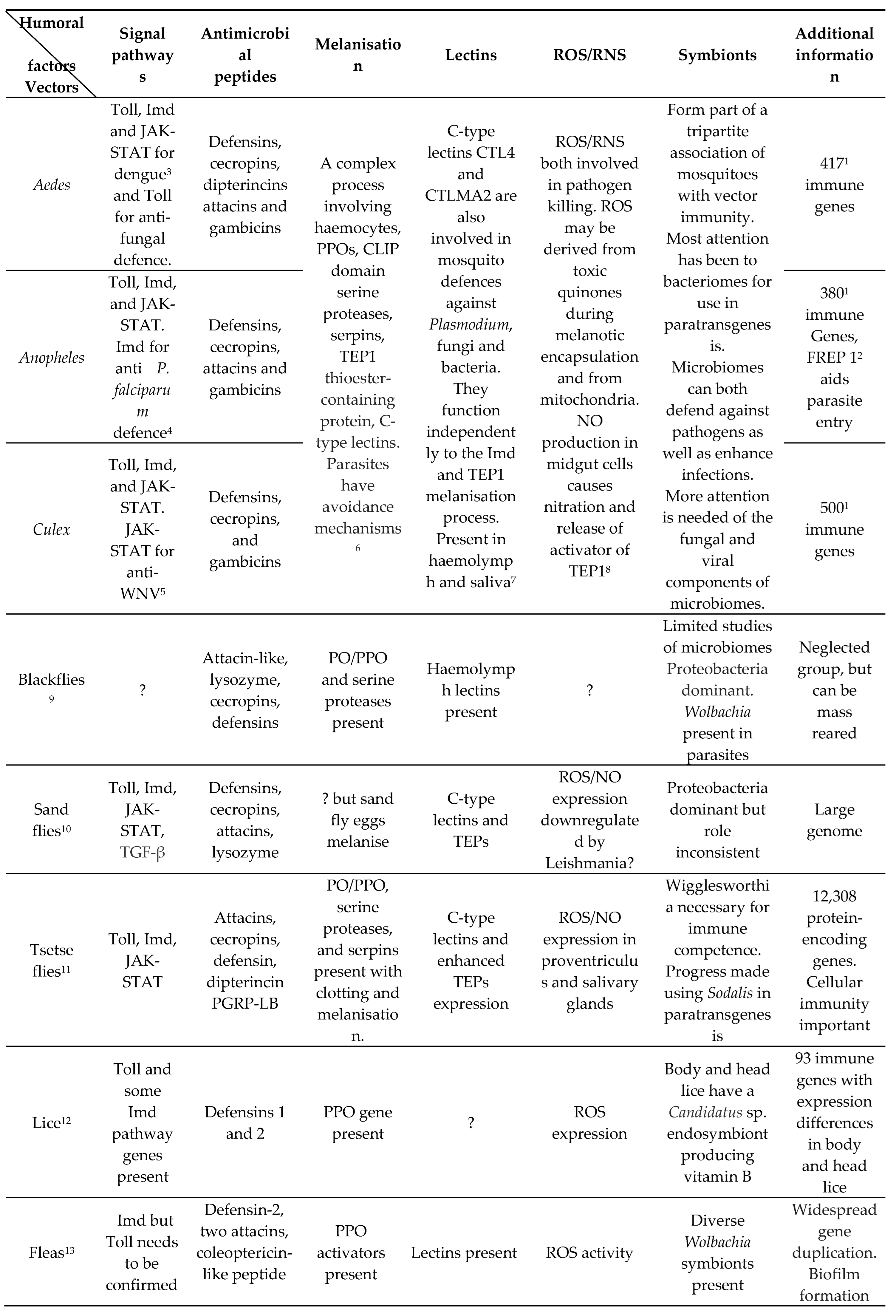 
|
* See text for details.
In contrast to Vector Cellular Immunity, in which the main immune processes were described separately, here the humoral immunity of each vector group is briefly described. This allows a more dynamic and interactive summary of this complex process.
5.1. Mosquitoes (Table 1 and Table 3)
The sequencing of the genomes of An. gambiae, Ae. aegypti and Culex quinquefasciatus [161,162,163] provided great opportunities for new approaches for studying the biology of these important vectors and has led to rapid progress in understanding the vector-parasite/pathogen associations. Ae. aegypti are responsible for vectoring yellow fever, dengue, Zika and Chikungunya, and filarial nematodes, An. gambiae for malaria and filarial worms, and C. quinquefasciatus for West Nile and Saint Louis encephalitis viruses, and filarial worms (Table 1).
The mosquito responses to these diverse pathogens may well vary with new PAMPS to be recognised on/in these, as well as with the different vector species whose immune competence is affected by the composition of the microbiota [18,164]. For example, in Aedes, Anopheles and Culex, 417, 380 and 500 immunity genes have been recorded, respectively, with expansions in Culex of C-type lectins, fibrinogen-related proteins (FREPs), and serine protease inhibitors (SRPNs) accounting for some of the increases in immunity gene numbers [165].
Following ingestion with the blood meal, the pathogens enter the midgut where they face numerous host-derived factors which may inhibit/kill the invaders or else assist in their development. The epithelial cells of the midgut, however, are protected by a chitinous/glycoprotein peritrophic membrane, although in An. gambiae, FREP1 anchors Plasmodium to the peritrophic matrix and assists parasite penetration of this structure [166]. Regarding arboviruses, commensal bacteria may assist the infection process into the mosquito midgut [167], while disrupting the formation of the peritrophic membrane by RNAi of chitin synthase expression has no effect on B. pahangi development or on the spread of dengue virus [168].
Once in the midgut, many factors are involved in the survival or elimination of the parasite including:-
- i.
- upregulation of immune effector genes to antimicrobial peptides (AMPs) via signalling pathways
- ii.
- the vector PpO system, melanisation and serine proteases
- iii.
- cytotoxic/stimulatory lectin molecules
- iv.
- nitric oxide and ROS killing of parasites
- v.
- specific peptides stimulating parasite differentiation
- vi.
- glycoprotein receptors on the surface of the midgut for parasite attachment
- vii.
- role of bacterial symbionts
Recognition by the PRRs of the PAMPs of invading parasites and pathogens occurs shortly after entry of the blood meal into the gut and other tissues of vectors, and results in the systemic upregulation of genes and immune effector molecules [24]. These are produced by triggering of the three signalling pathways, Toll, Imd and JAK-STAT and details of these are given in section 6 on Recognition, below, and in other papers in this special issue.
i. Important groups of effector molecules are the antimicrobial peptides against bacteria, fungi, Plasmodium, viruses and nematodes [24,126,151]. Four important AMPs in mosquitoes are defensins, cecropins, attacins and gambicins (Table 3). An example of the AMPs role against invaders is provided by RNAi silencing in Ae. aegypti of the Toll factor, Cactus, resulting in enhanced expression of the defensin gene and helping to control/neutralize dengue virus. Some bacterial species in the gut can also promote the expression of AMPs as with Serratia. marcescens in An. stephensi against Plasmodium berghei [169]. In mosquitoes, it is not clear which pathway induces which AMP but some are modulated by both Toll and Imd and this may indicate that these two signalling pathways partially converge downstream in response to infection with different pathogens [19]. The role mosquito AMPs play against bacteria, fungi, Plasmodium and viruses is recorded but interaction with nematodes is not fully understood, although previously primed Ae. aegypti exposed to B. malayi, significantly reduces infection intensity [172].
Ii/iii. The vector PpO, melanisation, serine proteases and C-type lectins (CTLs), are all involved in the mosquito immune defences to pathogens (Table 3). Melanisation of nematode microfilariae and Plasmodium ookinetes are described above in section 4. 2. 3. Nodules and Capsules and the mosquito haemocytes have been shown to be significantly involved eg. [102]. There are, however, reports of melanisation of nematode larvae in the midgut before entry into the haemolymph [173], and other confirmations that the interaction of this key defence process with pathogens is not fully understood in mosquitoes [174]. For example, in Ae. aegypti, the filarial nematode, B. malayi, has recently been shown to produce extracellular vesicles that downregulate the AAEL002590 gene encoding a serine protease involved in mosquito PO activity [152]. In addition, activation of the An. gambiae melanisation response is mediated by complex extracellular hierarchical cascades involving CLIP-domain serine proteases (CLIP-SPs), namely, SPCLIP1, CLIPA8, and CLIPA28 upstream of CLIPC9 [174]. To further emphasise the complexity of melanisation, C-type lectins (CTLs) in the genomes of Ae. aegypti, An. gambiae and C. quinquefasdatus have 52, 55 and 25 CTLs predicted, respectively [175], and have also previously been shown to be involved in PPO activation in other Insects [176,177]. In An. gambiae too, CTLs play an important role in interactions with Plasmodium falciparum parasites with CTL4 protecting the human parasite from a killing mechanism that is independent of the normal TEP1 mediated Imd pathway melanisation process [177]. In contrast, defence against the rodent parasite, P. berghei, involves TEP1 and the Imd pathway so that the mosquito immune system has different roles depending upon the pathogen [177]. The literature, however, also shows that in Anopheles coluzzii, the immune genes APL1C, LRIM1, TEP1, and TEP3 and the signalling pathways influence susceptibility to both fungal and P. falciparum infections [178].
Iv. In addition, reactive oxygen (ROS) and nitric oxide (RNS) species are also involved in mosquito killing of pathogens [122,173]. The blood meal alone results in ROS/RNS production but when Plasmodium passes through the midgut epithelium this is enhanced further [6]. Strains of An. gambiae resistant to bacteria also have higher levels of ROS and the application of antioxidants decreases mosquito survival [179]. Furthermore, P. berghei infection often produces high levels of mosquito killing which can be reduced by the oral application of the antioxidant uric acid [179]. It has been proposed that toxic quinones generated during melanotic encapsulation also generate high levels of ROS to kill parasites [180] and that in the mosquito midgut epithelium responses to Plasmodium are modulated by ROS from mitochondria [181]. Regarding RNS activity, a nitric oxide synthase (NOS) gene is transcriptionally activated in An. gambiae by both bacteria and Plasmodium parasites and is particularly active in the midgut wall where the synthesis of nitric oxide (NO) occurs by inducible NOS (AsNOS) [182,183]. Furthermore, mosquitoes fed with the NOS substrate, L-arginine, have reduced infection rates of 28 %, while the NOS inhibitor, L-NAME, increases significantly oocyst numbers in the midgut wall [183]. More recent research has revealed the role of RNS in the mosquito defences against invading Plasmodium parasites. The invasion of An. gambiae (G3) midgut epithelial cells by ookinetes causes a wounding response resulting in apoptosis and also induction of NOS, heme peroxidase (HPX2) and NADPH oxidase 5 (NOX5) enzymes in these cells. The NO produced is toxic and results in protein nitration of the midgut basal lamina which attracts haemocytes. Upon contact with the midgut nitrated surface these release haemocyte-derived microvesicles (HdMv) which activate the mosquito (TEP1) complement system to lyse the parasites [184,185].
v/vi. There have been many reports of parasite proteins required for Plasmodium development and invasion of mosquitoes but fewer on the role of mosquito midgut proteins assisting parasite invasion [186]. Such proteins include Anopheles alanyl aminopeptidase N (AnAPN1), fibrinogen-related protein1(FREP1), and AgPfs47Rec (An. gambiae Plasmodium falciparum P47 receptor). AnAPN1 is a midgut lumen surface glycoprotein functioning to digest the blood meal and binding to ookinetes to facilitate their entry into midgut cells for continuation of sporogenesis [186,187]. FREP1, as mentioned previously, anchors Plasmodium to the peritrophic matrix and assists parasite penetration of this structure [166]. AgPfs47Rec is an Anopheles, midgut receptor for P. falciparum protein Pfs47. This interaction mediates the parasite avoidance of mosquito immunity by disruption of the JNK (c-Jun-N-terminal kinase) signalling pathway leading to inhibition of the midgut nitration process and the TEP1 complement system [186,188].
vii. Research on the mosquito microbiome is most important since it has been shown that the component microbes have roles in the physiology, nutrition, metabolism, immunity, reproduction, longevity and behaviour of these vector insects eg. [26,189]. The microbiome also modulates the interaction of the vector with invading parasites and pathogens [18,26,189]. The reviews by Gabrieli et al. and Vinayagam et al. [18,29] describe the trilogy of interactions between the mosquito gut microbiome, the vector immune system and invading pathogens. The blood meal in female mosquito triggers the proliferation of the midgut microbiota whose cell wall peptidoglycan is recognised by the peptidoglycan recognition protein LC (PGRP-LC) in the anterior midgut, resulting in activation of the Imd pathway [18,29]. The activated Imd pathway eventually induces the expression of antimicrobial peptides (AMPs) via the NF-κB transcription factor Relish. This priming of mosquito immunity by the gut bacteria to express AMPs explains one way by which the microbiome defends against invading parasites and pathogens. For example, S. marcescens in An. stephensi defends against P. berghei [169] and Proteus sp. in Ae. aegypti protects against dengue [190].
Much work has concentrated on the bacteriomes of Aedes spp. and Anopheles although the mosquito microbiomes also contain fungi, viruses, archaea and protozoans [26,191,192]. The bacteriomes of these mosquitoes are composed mainly of Gram-negative species. In anophelines as many as 98 genera have been recorded [193], although few taxa, the core microbiota usually dominate [26,194]. The origin of the microbiome seems partially to depend on the ecology as different mosquito species from comparable environments have similar core bacteria [195]. The fact that these bacteria can reduce Plasmodium infections has stimulated interest in developing paratransgenesis (genetic manipulation of the insect vector´s native microbiome to inhibit or kill invading disease pathogens) to control mosquito-transmitted diseases, such as malaria, and maybe mosquitoes too. The main bacteria of interest include Asaia, Pantoea, Escherichia, Serratia, Enterobacter, Chromobacterium and Pseudomonas [26]. For example, Asaia strains inhibit Plasmodium development by producing toxic proteins [196], reduce parasite numbers by activating Anopheles immunity after infective feeding [197], and also inhibit competing Wolbachia infections [198]. Wang and Jacobs-Lorena [199] recognised four classes of anti-Plasmodium effector molecules: (i) parasite killers; (ii) those engaging with parasites; (iii) those engaging with epithelia of the mosquito midgut or salivary glands; and (iv) modulators of the mosquito immune system. These effector molecules with different modes of action, can potentially be engineered in combination into symbiotic bacteria, such as Pantoea agglomerans, to kill parasites and prevent the development of resistance [199].
5.2. Blackflies (Table 1 and Table 3).
There are more than 2,200 species of blackflies of which the largest genus, Simulium, has at least 26 species that are vectors of Onchocerca volvulus [200]. Many recent papers on this important group are concerned with “Prevention, Control and Elimination” [43,44] together with taxonomy, infection rates and composition of the saliva eg. [201,202]. Unfortuneately, since the pioneering research of Ham, Hagen et al. in the 1990s and early 2000s [203,204], there have been few papers on the details of the interaction of the blackfly immune system in the midgut and haemolymph with the microfilariae following parasitisation by O. volvulus. Therefore, vaccine development against O. volvulus has concentrated on antigens associated with the parasites rather than on elements of the vector immune response [43].
The usual comment about the life cycle of the microfilariae in the blackfly vector is that following an infected blood meal the parasites “develop further in the black fly and are then transmitted to the next human host” eg. [44]. In fact, Ham et al. [203] described 4 types of molecules modulated by ingested microfilariae in black flies, namely, antimicrobial peptides (AMPs), proteases, phenoloxidases (PO) and haemolymph lectins. The AMPs included attacin-like molecules (23 kDa), lysozyme (14 kDa), cecropins and defensins (4-8 kDa). Antibodies raised against some of these peptides inhibited immune killing of Onchocerca sp. microfilariae in Simulium haemolymph [205]. The proteases were both serine and cysteine proteases, and it was postulated that some of these are infection-specific in response to both bacteria and microfilariae, and may have been involved in PPO activation [203]. Levels of PO in microfilariae-infected black flies were reduced compared with controls possibly due to binding to the parasites and internal vector tissues. The haemolymph lectins recorded by Ham et al. [203] were hypothesised to be involved with PO/PPO in the recognition of non-self resulting in signal transduction to the nucleus and the induction of genes [203]. More recently, a defensin and a cecropin were identified and characterised from the salivary glands of Simulium bannaense [202]. The defensin, SibaDef, had high antimicrobial properties against Gram-positive bacteria while the cecropin, SibaCec, possessed potent activity against Gram-negative bacteria. SibaCec also had low cytotoxicity towards mammalian cells, neutralised LPS and exhibited strong anti-inflammatory activity [202]. Finally, progress has been made in identifying the composition of the bacteriome of black flies. The phylum Proteobacteria predominates in the black fly core bacteriome with Wolbachia the most dominant genus [206]. Importantly, the infection status of the blackflies had a significant association with the abundance of certain bacterial genera such as Serratia [206]. With further research, these bacteria have potential for the development of innovative control techniques including paratransgenesis.
5.3. Sandflies (Table 1 and Table 3).
Approximately, 500 species of sandflies (phlebotomine) have been described with more than 90 transmitting leishmaniasis. Species and subspecies of Phlebotomus in the Old World and Lutzomyia in the New World are the main vectors of human leishmaniasis [207]. There are numerous papers on various aspects of sandfly biology with some consideration of the role of the vector immune response as a determinant of infection by Leishmania parasites. Examples of relevant publications on sandfly innate immunity include Dillon et al. [208], Boulanger et al. [209], Telleria et al. [8], Coutinho-Abreu et al. [210], Kykalová et al. [211], Omondi et al. [212] and Campolina et al. [213]. Dillon et al. [208] undertook an analysis of expressed sequence tags (ESTs) derived from a whole-body cDNA library from Lutzomyia longipalpis sandflies, some of which were infected with Leishmania infantum, and revealed putative proteins involved in the barrier function of the vector gut, digestive physiology, and the immune response. The immune factors identified included Gram-negative binding proteins, galectins, thioester proteins, scavenger receptors, signalling pathway factors, serpins, caspases, and peroxidases. The detection of these proteins indicated the presence of an active innate immune system in sandflies capable of interacting with invading parasites/pathogens like Leishmania. Evidence for this has also been provided by transcriptomic studies of changes in immune gene expression following infection with Leishmania. The genes affected included members of the Toll, Imdand JNK pathways and the antioxidants, catalase, glutathione s-transferase, superoxide dismutase and peroxiredoxin, controlling ROS levels [reviewed in 8]. The upregulation of the Dorsal and Relish genes, that are positive modulators of the Toll and Imd pathways, respectively, also occurred following Leishmania challenge and led to the expression of the AMPs attacin, cecropin and defensin 2 increases at different time points in the sand fly LL5 cell line [127]. In contrast, Leishmania infected L. longipalpis show no significant changes in ROS gut levels compared with controls. Since reductions in numbers of Leishmania in the sand fly gut occur following silencing of the sand fly antioxidant, catalase, this may indicate manipulation of vector antioxidative elements by the parasite [127].
The above implies the involvement of components of the sandfly innate immune response following Leishmania infections. Consideration, however, of the published work leads to some contradictions in observations recorded since minimal responses to the presence of Leishmania in the sandfly gut have also been published [210]. Similar inconsistences have also been noted in research on mosquito immunity in which the vector insects or parasites used were derived from different vector or parasite populations or used in alternative combinations. In addition, variations in insect physiological states or parasite developmental stages and the use of alternative sampling and analysis techniques can all affect the results [26]. Leishmania, in contrast to Plasmodium in mosquitoes, is confined to the gut of sandflies where it undergoes several developmental changes. The amastigotes in the blood meal develop into to weakly motile procyclic promastigotes in the peritrophic membrane, then to strongly motile long nectomonad promastigotes in the midgut lumen that transform into short nectomonad promastigotes, also called leptomonads, which eventually form the infective metacyclic stage which are regurgitated and transmitted during blood feeding [214]. All these various forms probably present unique molecular surface challenges to the sandfly immune system and result in variations in the vector response [8]. This emphasises the complexity of the sandfly-parasite association which is made even more complex by differences in the microbiomes of the vectors.
Studies of sandfly microbiomes have identified Gram-negative members belonging to the phylum Proteobacteria as dominant with the Gram-positive phyla Firmicutes and Actinobacteria also present. The Proteobacteria include Serratia and Enterobacter in the family Enterobacteriales and Pseudomonas as core taxa eg. [8,26,215]. The role of the bacteriome in priming mosquito immunity and reducing Plasmodium infections has been described above (see section 5. 1. Mosquitoes), and similar reports exist for sandflies. For example, the effects of in vitro and in vivo co-cultivation for 24 h of each of 13 native bacteria isolated from L. longipalpis, at different developmental stages and physiological conditions, with promastigotes of Leishmania infantum chagasi, Leishmania major, Leishmania amazonensis, and Leishmania braziliensis were analysed. After co-cultivation, a reduction in growth was recorded with all parasite species [213]. In contrast, again with L. longipalpis, but infected with L. Infantum, antibiotic-mediated perturbation of the midgut microbiome rendered sandflies unable to support parasite growth and metacyclogenesis. This suggests that an intact sand fly midgut microbiome is necessary for Leishmania development to its infective stage [216].
The reasons for such variations in experimental results has been discussed in detail in other papers eg. [8,26,213], and confirms the complexity of the sand fly-Leishmania relationship. This complexity emphasises the need for further studies, for example, of the sandfly haemocytes (see section 4.1. Hemocyte Types), of the parasite evasion processes eg. [8,210,213,218], of the potential for development of paratransgenesis [26] and vaccines [23,219], and of the role of the vector microbiome co-egested with the parasites during sandfly feeding on the establishment of Leishmania in the mammalian host [23,218].
The reasons for such variations in experimental results has been discussed in detail in other papers eg. [8,26,213], and confirms the complexity of the sand fly-Leishmania relationship. This complexity emphasises the need for further studies, for example, of the sandfly haemocytes (see section 4.1. Hemocyte Types), of the parasite evasion processes eg. [8,210,213,218], of the potential for development of paratransgenesis [26] and vaccines [23,219], and of the role of the vector microbiome co-egested with the parasites during sandfly feeding on the establishment of Leishmania in the mammalian host [23,218].
5.4. Tsetse flies (Table 1 and Table 3)
Tsetse flies (Glossina spp) are viviparous and include 30–33 species and subspecies and usually divided into the Morsitans, Palpalis and Fusca groups which are particularly important medically and economically due to transmission of African trypanosomes in humans and animals [220]. An excellent review and original papers on tsetse fly innate immunity are provided by Matetovici et al. [12], while Weiss et al. [132] emphasize the importance of tsetse cellular immunity, as described above (see section 4. 2. 2. Phagocytosis).
The Glossina genome was published in 2014 and is almost twice the size of the Drosophila genome containing ca. 12,308 protein-encoding genes [221]. The competence of the tsetse flies as vectors is determined by many factors such as nutrition, age, sex and symbionts [220,221]. Glossina has a reduced component of some humoral immunity related genes, for example, some AMPs, lysozyme, C-type lectins, peptidoglycan recognition proteins (PGRPs), glucan binding proteins, serine proteases and serpins, while other immune genes are expanded such as those encoding for the AMPs, attacin A and attacin B, and for the thioester-containing protein family (TEPs) [12,221,222]. In addition, tsetse EP protein, reactive intermediates of oxygen and nitrogen species (ROS, RNS), coagulation, melanisation, phagocytosis and the peritrophic membrane all contribute to the tsetse defence response [12,132]. The AMPs include the attacins A and B as well as cecropins A1, A2, B and C that are induced through the Toll and Imd pathways [12,222].
Particularly significant is the role of the symbiont, Wigglesworthia, in the development of the immune system in the tsetse larvae in order for the immune system to function normally in adult flies. In a basic simplification of the research by Weiss et al. [132,223], pregnant female tsetse were fed a diet containing tetracycline. The antibiotic removes all symbionts from the flies, resulting iñ aposymbiotic adult Glossina morsitans morsitans [GmmApo]), with a severely compromised immune system without phagocytic haemocytes and with abnormal expression of immunity-related genes. Subsequently, these flies rapidly succumbed to infection with normally non-pathogenic Escherichia coli. Furthermore, the process of immune system development can be restored in intrauterine GmmApo larvae when their mothers receive a diet supplemented with Wigglesworthia cell extracts. Therefore, molecular components of Wigglesworthia have immunostimulatory activity within tsetse, and this represents a novel evolutionary adaptation that links an obligate symbiont with its host [132,223]. Wigglesworthia also enhances, in the gut of intrauterine tsetse larvae, an odorant binding protein 6 that stimulates the hematopoietic RUNX transcription factor, lozenge, causing larval haemocyte precursors to develop into functional crystal cells and initiate the melanisation cascade, via prophenoloxidase release [224], and haemolymph clotting. In Wigglesworthia-free tsetse flies cuticular wounds also fail to clot [223].
The above is a basic description of tsetse fly innate immunity since many other aspects of this process have been hardly mentioned. For example, the role of the PGRPs in tsetse immunity is important with the Glossina genome containing six PGRPs genes, four in the long (pgrp-la, -lb, -lc, -ld) and two in the short (pgrp-sa, -sb) subfamilies [221]. Of these, RNAi silencing of PGRP-LC in G. morsitans suppresses the Imd pathway resulting in a strong inhibition of attacin expression and an enhancement of midgut trypanosome infections [12,225]. Tsetse PGRP-LB, however, functions to degrade peptidoglycan from microbes, preventing an overactive immune response and avoids damage to the essential Wigglesworthia symbionts in the tsetse bacteriome [12,225]. PGRP-LB, together with the other AMPs, has strong trypanocidal activity against procyclic and bloodstream trypanosomes [226]. In addition, the peritrophic matrix forms the first barrier to the parasites and determines the immunologically detection of the invader. In summary, this immune detection together with the Imd-pathway associated PGRP-LB, the AMP attacin, and the cellular immune system are key components of the tsetse fly / trypanosome interaction that leads either to parasite establishment or elimination from the midgut [12,132].
5.5. Lice (Table 1 and Table 3)
There are about 4,000 species of lice but only 3 species infest humans, namely, the body louse, Pediculus humanus (= P. humanus humanus), the head louse, Pediculus capitis, and the pubic or crab louse, Pthirus pubis [56]. The body lice and head lice are closely related and may belong to the same species. The human body louse, P. humanus, vectors Rickettsia prowazekii, causing epidemic typhus, as well as Borrelia recurrentis, causing epidemic relapsing fever, and Bartonella quintana, the causative agent of trench fever [56]. Body lice may also be the vectors of plague caused by Yersinia pestis [227]. It is generally regarded that body lice act as vectors of these diseases and not the head lice although there is increasing evidence that head lice may be vectors too [227].
Lice are ectoparasites with infections often spread from the faeces or from crushed insect bodies, with blood meals taken several times per day by members of the suborder Anoplura [56]. The genome sequence of the body louse and a symbiont were published in 2010 [228] and showed that the body louse has the smallest genome of any hemimetabolous insect reported up to 2019 [229]. Comparisons have been made between the immune systems of body and head lice to identify any differences resulting in the primary vector role of the former and not of the latter [115,129]. For example, following bacterial challenge with Bartonella quintana, several genes in the body lice are downregulated compared with head lice and the bacteria multiply at a higher rate in the body lice too [230]. A transcriptional analysis of the basal immune response of the guts of body lice and head lice, showed that the transcript levels of important immune genes, such as the peptidoglycan recognition protein and defensins, were reduced in body lice while the defensin 1 transcription following B. quintana oral infection was only up-regulated in head lice [230]. Furthermore, the level of ROS produced by epithelial cells was significantly lower in body lice [230]. These results seem to indicate that the higher vector capacity of the body lice may be related to the reduced expression of certain key immune genes [129,229,230]. Immune genes associated with the Toll pathway have been identified in lice including those for the AMPs, Scavenger receptor A, Fibrinogen-like Protein, and Spaetzle. Lice also lack an Imd gene but retain some other Imd pathway genes [115] and can still respond to Gram-negative bacteria like E. coli [129}. Regarding symbiotic bacteria, both body and head lice have an endosymbiont, Candidatus Riesia pediculicola, in special enlarged midgut cells called mycetocytes forming the mycetoma and producing certain B vitamins not present in the blood meal [231].
5.6. Fleas (Table 1 and Table 3)
There are ca. 2574 species of fleas with 16 families and 238 genera, but only a minority is synanthropic ie. live intimately with humans [232]. Common synanthropic species are Pulex irritans (human flea), Ctenocephalides felis (cat flea), and Xenopsylla cheopis (rat flea). Diseases vectored by fleas include plague, caused by Yersinia pestis, murine typhus (endemic typhus, Rickettsia typhi), rural epidemic typhus (Rickettsia prowazekii) in the USA, spotted fever agent Rickettsia felis, and Bartonella spp, including Bartonella henselae, the agent of cat-scratch disease. P. irritans is an inefficient vector of plague bacteria, Y. pestis, compared with the rodent flea, X. cheopis. Furthermore, fleas also carry the helminths, Dipylidium caninum and Hymenolepis diminuta that, respectively, are parasites of carnivores and rats. Finally, tungiasis in the tropics is a human disease linked to the parasitism of humans by fleas [233].
Often, pathogen transmission by fleas occurs orally through regurgitation of blood meals or from contaminated faeces [233]. The outcome of infection of fleas by bacterial pathogens also varies according to both the species of the flea and of the infecting bacteria [234]. Thus, Y. pestis is usually confined to the gut of different flea species while rickettsial pathogens, such as R. felis, may penetrate the midgut epithelium to migrate rapidly through the haemocoel to the salivary glands [235]. Therefore, any description of the immune response of fleas to bacterial invasion needs to consider events occurring in the midgut, haemolymph and salivary glands. This process may be complicated in C. felis, and probably X. cheopis, by the widespread gene duplication in the genome with genome sizes ranging from 433-551 Mb for individual fleas in different populations [236]. Therefore, every cat flea has a unique genome sequence with gene duplication a source of genetic innovation creating problems in gene-targeting pest control measures and complicating comparative transcriptomics analysis [236].
In the flea midgut following an infected blood meal, the bacteria need to avoid being excreted and so they form biofilm aggregates or bind to the gut by receptor/ligand interaction [9]. For example, in X. cheopis, Y. pestis forms a biofilm which blocks the proventricular valve causing regurgitation of the flea gut contents and pathogen transmission into the wound [237, 238}. Studies on the gut transcriptome of X. cheopis showed that the initial immune response to Y. pestis infection was the upregulation of AMPs. Coincidentally, Relish, an NF-κB-like transcription factor controlling gene expression in the Imd pathway, was also upregulated in infected fleas. In addition. the peptidoglycan recognition proteins (PGRP-LC and PGRP-LB), which are also activators and regulators of the Imd pathway, were upregulated too. The AMPs detected included two attacins and a coleoptericin-like peptide, [237]. Gene expression of antibacterial ROS, however, was limited, so that that Y. pestis fails induction of a strong initial ROS response in X. cheopis [237]. However, previously, ROS have been described in response to Y. pestis infection in the gut of X. cheopis [130,237] and this illustrates either the variable nature of flea immunity and/or differences in the experimental protocols used. Cactus, the negative regulator of Toll, was upregulated by infected and sterile blood meals, so the Toll pathway probably does not participate in the flea immune response to Y. pestis [237]. Other transcripts modulated in the midgut by infection and /or a blood meal include serine proteases in C. felis midguts infected by R. typhi [239], while serpins and PPO activators have also been detected [9].
Regarding the invasion of the flea haemocoel, salivary glands and other tissues by rickettsial pathogens, such as R. felis and R. typhi, these organs have also been shown to have active immune responses to bacteria [111,235,240]. For example, utilising model bacteria injected into the haemolymph of the cat flea. C. felis, Muñoz et al. [111] found that the haemolymph had enhanced antibacterial activity and increased numbers of haemocytes so that additional studies with pathogens were justified. Likewise with studies on C. felis injected with R. felis and using PCR showed that defensin-2, glycine-rich AMPs and several flea antigens were modulated by rickettsial infection [235]. Finally, apart from the bacterial pathogens vectored, fleas are also known to have other bacteria in their microbiomes, but information is limited except that a diverse range of Wolbachia strains have been reported of unknown interactions with flea immunity and vector competence [241].
5.7. Triatomines (Table 1 and Table 3).
There are ca. 150 species of triatomine bugs [242,243] distributed mainly in South and Central America and southern USA. Most triatomine species are capable of vectoring the haemoflagellate protozoan parasite, Trypanosoma cruzi. In contrast, the metacyclic form of the closely related species, Trypanosoma rangeli, is reported to be mainly confined to Rhodnius species [244]. T. cruzi infection occurs after a blood meal from the human host by the triatomine vector results in transmission of Chagas disease, an often fatal condition of the heart and gut [242]. The T. cruzi development is confined within the gut, from stomach to rectum, while T. rangeli invades the haemocoel and haemocytes before colonising the vector salivary glands [245]. For T. cruzi, Rhodnius, Triatoma, and Panstrongylus species are epidemiologically most important in Latin America, as they colonize peridomestic and domestic areas close to humans and their animals [243,246].
There are considerable advantages of using triatomines for research into the immune reactions of insect vectors to their parasites and pathogens. First, the large size of many triatomines means that tissue samples, such as haemolymph, are easily obtained. For example, the most commonly researched triatomine, Rhodnius prolixus, grows up to 34 mm in length which is ca. 10 times the size of sandflies and fleas, and at least 5-7 times that of lice, blackflies and mosquitoes (personal observations). Secondly, R. prolixus is relatively easily cultured, and there is already much information on its physiology gleaned from decades of research by entomologists of this vector as a model insect [247].
In this Special Issue, Schaub et al. present a detailed overview of “Triatomine innate immunity against parasites” so that only a basic description of triatomine humoral immunity is given here. Emphasis will therefore be made of some of the more recent research since the publication of the R. prolixus genome in 2015 [248]. In addition, many details of research progress in understanding triatome immune responses following infection are also given in a number of the most up to date reviews [21,24,27,135,137,138,243,249,250,251,252,253].
The humoral immune defences of triatomines have a number of factors/processes produced by the haemocytes, fat body, gut and other tissues, including antimicrobial peptides (AMPs), reactive oxygen species (ROS) and nitric oxide (NO), prophenoloxidase (PPO), melanisation, lysins, lectins and enzymes. All of these come together to combat would-be invaders, denying infection so that only a minority of vectors usually succumb to transmit disease.
Regarding the AMPs, an arsenal of these has been detected in triatomines including defensins A, B and C, lysozymes A and B, prolixicin, attacin, diptericin and trialysin [137]. The PRGPs recognise the PAMPs on the surface of the invaders and the Toll, Imd and JAK/STAT pathways are activated to induce the expression of the AMPs. Previously, there was some doubt about whether the Imd pathway in Rhodnius was fully functional, but using knockdown experiments of one of the PGRP genes reduced AMP expression induced by Gram- bacteria and confirmed the role of the Imd pathway in AMP expression [119]. There has also been interesting research on Triatoma pallidipennis in which the Toll and Imd pathways were silenced separately or together and then insects challenged with bacteria. This resulted in insect survival rates of ca. 62 – 73 % when silencing occurred separately, but this fell to ca. 36% for Escherichia coli and 41% for Micrococcus luteus) after silencing both pathways This indicated that the Imd and Toll pathways participate jointly to eliminate Gram+ and Gram- bacteria [139]. Challenging with T. cruzi or T. rangeli and monitoring parasite numbers might have yielded interesting results regarding the participation of the Imd and Toll pathways in immune reactions towards parasites.
Using quantitative proteomics for the analysis of the R. prolixus haemolymph from 6 to 24 hr following T. cruzi infection also identified novel proteins with possible roles in immune reactivity to parasites [24]. These included an immunoglobulin I-set domain containing protein (T1HCN4) for the first time in triatomines and of unknown function but which like hemolin might be involved in inhibition of haemocyte aggregation [24]. Another protein identified was a C1q-like protein which was strongly induced by T. cruzi infection compared with uninfected blood alone and may also be involved in the vector immune response to parasitisation [24].
A complex of interacting factors from the R. prolixus vector as well as from the invading trypanosomes determine the outcome of infection and the subsequent survival or killing of the parasites [135,136,137,138]. One key factor that seems to be involved in the invasion and survival of T. cruzi in R. prolixus, and other vector insects, is nitric oxide (NO), with reactive nitrogen species (RNS) resulting from activation of nitric oxide synthase (NOS) to yield NO. It has been shown that NOS gene activity and NO production are specifically modulated in R. prolixus according to the nature of the invading organism so that differential responses occur following infection with T. cruzi and T. rangeli [254,255,256]. Results of Whitten et al. [255] together those of Baptista et al. [257] showed an augmentation of NOS gene expression by treatment with L-arginine, an activator of NO, and a reduction of NOS expression with L-NAME, an inhibitor of NOS. These modulations coincided not only with different levels of parasites in the gut but also with changes in levels of phenoloxidase, superoxide anion and antimicrobial activity but also with reductions in intestinal microbes. Such reductions in numbers of bacteria following infection with T. cruzi and T. rangeli were confirmed in a recent metagenomic shotgun sequencing study of the gut microbiome of R. prolixus [252]. These studies confirm several aspects of the complexity of the triatomine immune response with no real explanation of the factors governing these changes. It is essential that the responses occur early on in the invasion of the vector insect by the parasites since the anterior gut is a hostile environment with the majority of parasites often failing to infect the host insect. It has previously been postulated that surface components of T. rangeli directly or indirectly cause rapid modulation of the insect vector immune system after parasite invasion into the haemocoel [258]. In addition, T. cruzi has been reported to spontaneously secrete extracellular vesicles [259]. Such vesicles have been shown in humans to play key roles in pathological and physiological functions [260]. In protozoan parasites causing malaria and Chagas disease, there is also evidence that extracellular vesicles enhance growth and promote transmission, helping to avoid host immunity and to modulate the microenvironment [260].
Potential factors modulating the complex immune interactions between the vector insect and trypanosome parasites are the prostaglandins (PGs) and other eicosanoids. These molecules have been shown to play important roles in insect innate immunity eg. [142,250,261,262,263] but, with notable exceptions, seem to be have had less attention recently. This may be due to the fact that cyclooxygenase (COX) genes responsible for converting C20 PUFAs (polyunsaturated fatty acids) into PGs seem to be absent from published insect genomes [262]. There is now confirmed evidence that eicosanoids mediate both cellular and humoral immunity in insects including much research in R. prolixus and some in mosquitoes [263,264]. Significantly, eicosanoids have their effects early in the immune response and are involved in phagocytosis, nodule formation, clotting, haemocyte chemotaxis and aggregation, ROS formation, melanisation by PPO, mediation of AMP gene expression in both Toll and Imd pathways, interactions with the fat body, and Toll/Imd signalling to trigger NO to activate PLA2 (phospholipase A2) and synthesise eicosanoids [142,264]. The research by Barletta et al. [263] with An. gambiae mosquitoes invaded by Plasmodium parasites is classic and beautifully designed and executed and confirms the role of the midgut production of prostaglandin E2 to attract haemocytes and induce long-term systemic cellular immune responses to infection by Plasmodium parasites (see also section 6.3.6 Immune Priming for more details). Finally, it is well known in vertebrates that eicosanoids effect the expression of iNOS and NO production [265] so much can also be gained by reading this literature.
6. Recognition, Signalling, and Priming in Vector Innate Immunity
Previously, mention was made of recognition, signalling, RNA inhibition and priming of vector immune responses to invading pathogens/parasites but little detail was provided of these processes. In this section, additional consideration is given to these responses with reference to Drosophila which provides a comparative model and guide for investigations into these processes in vector insects by assumptions that the innate immune responses of Drosophila are well conserved. These approaches include identification of orthologous genes, quantifying transcript abundance of these genes after pathogen challenge, and RNA interference of key pathway regulators combined with quantifying the effect of this gene silencing on pathogen abundance in the manipulated vector. While for some vector insects the data underlying these assumptions are consistent with the canonical pathways in Drosophila, for others, such as triatomines, the available evidence points to the existence of non-canonical signalling pathways, as discussed below.
6.1. Pathogen Recognition
As in other animals and plants, insects solve the conundrum of recognition of self versus non-self with the help of proteins termed pattern recognition receptors (PRRs). These recognise pathogen-associated molecular patterns (PAMPs) that are not features of insect cells, thus enabling discrimination between self versus non-self. These PAMPs are typically components of the cell surface of parasites/pathogens and can be categorised as polysaccharide PAMPs such as peptidoglycan (PGN), lipopolysaccharide (LPS), capsular polysaccharide, β-1,3-glucan and zymosan. Apart from LPS, other lipid-containing PAMPs include lipoteichoic acid and lipoarabinomannan, while proteinaceous PAMPs include capsid proteins and flagellins. Finally, pathogen-released nucleic acids are additional PAMPs, composed of viral RNA, both double- and single-stranded, and DNA [266].
PAMPs alert the insect to a possible infection, triggering the innate immune response. PRRs are the sensing proteins that initiate this response with some PRRs functioning to directly elicit responses such as phagocytosis and the PPO cascade, whereas others trigger signal transduction pathways that regulate immunity. We describe the principal PRR protein families involved in vector insect immunity; other candidate PRRs are reviewed elsewhere [267,268].
6.2. Pattern Recognition Receptors (PPRs).
6.2.1. Peptidoglycan Recognition Proteins (PGRPs)
PGRP is a fundamental PRR conserved from insects to mammals, regulating the immune response to bacteria. Insects can express multiple PGRPs with each regulating specific immune responses. For example, in D. melanogaster, two extracellular PGRPs, PGRP-SA and PGRP-SD, recognise bacterial Lys-type PGN and stimulate the Toll pathway after infection by Gram-positive bacteria, with the latter also upregulating the Imd pathway [269,270,271]. The Imd pathway is also stimulated by extracellular PGRP-LE and a membrane-bound PGRP-LC [273,274]. Aside from eliciting these responses, PGRP-LE also participates in the PPO cascade [275,276] converting prophenoloxidase (PPO) to active phenoloxidase (PO) that leads to localised wound healing and melanisation. To counter cell-invasive pathogens such as Listeria monocytogenes, intracellular PGRP-LE also stimulates autophagy [277].
In an in-silico study conducted using the genome of An. gambiae, approximately 150 putative PRRs were identified, the majority of which were found to be secreted proteins having adhesive domains to interact with PAMPs. These PRRs were found to cluster as members of large families of genes [278].
6.2.2. Immunolectins
Typical insect immunolectins (IMLs) are calcium-dependent C-type lectins that contain either single or dual carbohydrate receptor domains. They can bind to LPS, lipoteichoic acid and β-1,3-glucan and participate in a variety of immune responses, including encapsulation, phagocytosis, nodulation and activation of the PPO cascade [279] Another important family of carbohydrate binding proteins in the mosquito’s innate immune response are the fibrinogen-related proteins (FREPs = FBNs). In An. gambiae, they are the largest pattern-recognition protein family consisting of 59 putative members, most of which showed immune responsive transcription when challenged by infection with bacteria, fungi, or Plasmodium [280,281]. Using RNAi-mediated gene-silencing assays, showed that members of the FREP family play a central role in the innate immune response and maintenance of immune homeostasis in mosquitoes [282]. Thus, FBN9, FBN 30 and 39 inhibit Plasmodium infection in Anopheles midgut epithelial cells [186]. In contrast, however, FREP1 functions as an anchor in the peritrophic membrane helping invasion of the mosquito midgut by P. falciparum ookinetes, with CRISPR-Cas9 knockout of FREP1 mosquitoes resulting in suppression of infection by both oocysts and sporozoites [186].
6.2.3. Thioester-Containing Proteins (TEPs)
TEPs are secreted glycoproteins containing a highly reactive thioester motif that covalently binds to microbial surfaces [283] and function as opsonins, promoting phagocytosis, lysis and the PPO cascade [284]. Studies in Drosophila indicate that several different TEPs acting as PRRs can activate the immune Toll, Imd, and JAK/STAT signalling pathways [285,286]. The mosquito Anopheles gambiae encodes up to 15 TEPs, and of these, TEP1 plays important roles in (i) the phagocytosis of bacteria [287], (ii) the lysis of Plasmodium ookinetes [288], and (iii) the lysis and melanisation of entomopathogenic fungi [289]. TEP1 of Aedes aegypti limits infection by Dengue virus and is required for activation of the Imd signalling pathway and AMP synthesis [290].
6.3. Signal Transduction Pathways
Pathogen recognition activates signal transduction pathways to amplify and coordinate the immune responses including synthesis of antimicrobial factors. PAMP recognition by PRRs leads to activation of protein kinases, proteases and other enzymes in pathways that ultimately act on core transcription factors that, once translocated into the nucleus, activate expression of immune-related genes. Different immune signalling pathways have been described in insects, including Toll, Imd, JAK/STAT, JNK and RNAi [267,291]. The JAK/STAT pathway regulates immune response genes related to viral and bacterial infections and the RNAi pathway mainly controls virus replication. The Toll and Imd pathways are inflammatory responses that activate expression of a wide spectrum of antimicrobial peptides (AMPs) through the activation of the NF-kB-like (Nuclear Factor-kappa B-like) transcription factors. The activated NF-κB is translocated from the cytosol into the nucleus of the cell, binding to specific sequences of DNA and inducing the transcription of AMP genes [292].
6.3.1. The Toll Signalling Pathway
Insect Toll receptors perform critical functions in both embryogenesis and innate immunity. Toll receptors have an extracellular leucine-rich repeat region, a transmembrane domain and an intracellular Toll/interleukin-1 receptor (TIR) domain and are activated by a complementary family of six cytokine-like molecules called Spaetzle (Spz) [293]. The terminal proteinase, Spaetzle processing enzyme (SPE), has trypsin-like specificity and cleaves the Spz precursor The released C-terminal polypeptide can dimerize and bind to Toll receptors in the membrane [294], triggering a conformational change to the cytoplasmic TIR domain so that it can recruit a heterotrimeric complex of three proteins: MyD88, Tube and the kinase Pelle [295]. The protein kinase Pelle promotes the phosphorylation and degradation of Cactus, a cytoplasmic ankyrin-repeat-containing protein that retains NF-κB family transcription factor Dorsal and Dorsal-related immune factor (Dif) in the cytoplasm [295,296]. After degradation of Cactus and translocation to the nucleus, Dorsal and/or Dif can then activate expression of antimicrobial factors such as antimicrobial peptides (AMPs). In Drosophila, the Toll pathway regulates both cellular and humoral immune responses and these may respond to an interplay between the Toll pathway and eicosanoid signalling (see below).
In An. gambiae, the Toll pathway controls expression of the anti-parasitic proteins TEP1, a thioester-containing protein, and LRIM1 (leucine-rich repeat immune protein 1). Basal level expression of these factors and their secretion from haemocytes, resulting from an immune priming mechanism, reduces initial parasite infection [297]. Moreover, post-infection, the expression of these factors increases, presumably to modulate parasite numbers. Experimental RNAi of Cactus, the inhibitor of Relish1 (the NF-κB gene controlling the Toll response in A. gambiae), results in overexpression of these factors, completely blocking parasite development in the mosquito. Commensal Asaia bacteria can prime Anopheles immunity to limit subsequent Plasmodium infection by upregulating expression of TEP1 [197].
6.3.2. The Imd Signalling Pathway
In insects such as flies, mosquitoes and beetles, PGRPs including PGRP-LC and PGRP-LE form complexes with DAP-type PGN of Gram-negative bacteria. Following recognition of the PGRPs results in the recruitment of the Imd protein [298] and then an intracellular signalling cascade is activated by the interaction of Imd with other intracellular signalling proteins [299]. This leads to the translocation of the transcription factor Relish into the nucleus where it binds to the transcription response elements of AMP genes and promotes their expression [300].
In Aedes mosquitoes, the Imd pathway is activated by bacteria and Plasmodium, and an indirect effect of the Imd pathway has been shown on viral load [301]. Although the Toll and JAK-STAT pathways contribute to the immune response against Plasmodium, the Imd pathway is considered the primary response [302]. The NF-kB transcription factor in the Imd pathway is represented by the protein Rel-2. It plays the central role in Imd signalling and regulates a major AMP, cecropin1. Multiple studies have shown that Caspar functions as a negative regulator of the Imd pathway and that the immune response against Plasmodium could be exaggerated by the silencing of the Caspar gene [302].
In sandflies, gene silencing via RNAi of Caspar [303] led to a reduction in Leishmania population in the gut of L. longipalpis, while the knockdown of Relish (the transcription factor gene of the Imd pathway) resulted in the increase in Leishmania and bacteria in P. papatasi [304].
The cat flea Ctenocephalides felis contains homologs to Drosophila Imd pathway genes and inactivation of relish or Imd genes increases Rickettsia typhi burden in the gut of infected fleas [305]. However, mutant strains of Yesinia pestis that are highly sensitive to AMPs in vitro exhibited normal growth in the digestive tract of fleas, which suggests that the presence of this pathogen in vivo does not automatically induce AMP gene expression [306], indicating that fleas are permissive hosts to this bacterium.
Tsetse flies have a full complement of Imd signalling pathway genes and this pathway responds to trypanosome infection. RNAi silencing of the Imd pathway transcriptional factor Relish (GmmRel) prevents the activation of the pathway, blocking the expression of both attacin and cecropin AMPs and increasing the establishment of trypanosome infection in the tsetse midgut [225,307].
Some insect species appear to lack key Imd signalling components. For example, the triatomine, Rhodnius prolixus, lacks both Imd and Kenny genes [136], and the function of other identified candidate Imd pathway genes has yet to be determined. RNA interference, inhibition studies and qRT-PCR experiments implicate remaining Imd pathway genes, such as Relish, being involved in a possible non-canonical Imd-Toll pathway regulating expression of AMPs as a result of challenges by either Gram + or Gram – bacteria [308,309,310]. Infection with the parasite T. cruzi can result in increased AMP expression that, in turn, impacts competing bacterial numbers in the midgut, enabling the parasite to establish an infection.
Lice also lack an Imd gene but retain some other Imd pathway genes [115]. They also have a remarkably small complement of effector AMP genes, namely 2 defensin genes. Challenge with Gram-positive Staphylococcus aureus resulted in increased expression of these two AMP genes in the absence of upregulation of Toll or Imd pathway genes [311]. However, no responses were detected after challenges with Gram-negative E. coli. These results suggest that lice are somewhat permissive to bacterial infection, underlining their role as important vectors of bacterial pathogens. This functional crosstalk across Imd and Toll pathways may be more common than generally recognized among different insect species and could be an evolutionarily intermediate step towards the loss of Imd-related genes [312]. This hypothesis suggests repeated and independent losses of Imd pathway components occurring during arthropod evolution. Alternatively, hemimetabolous insects and other arthropods that characteristically do not liberate their gut bacteria during metamorphosis, may possess an ancestral incomplete (relative to holometabolous vectors and Drosophila) Imd pathway. For example, the tick Ixodes scapularis lacks genes encoding upstream regulators of the Imd pathway, including Imd, and FADD [313]. Despite the absence of these regulators, the remaining Imd pathway is active against infection by tick-borne pathogenic bacteria such as Borrelia burgdorferi and is triggered by cellular stress responses centring on the evolutionary conserved Unfolded Protein Response (UPR) [314]. We speculate that this could also be true for hemimetabolous vector insects.
6.3.3. The JAK/STAT Pathway
The JAK/STAT signalling cascade takes place in the fat body and gut cells and is elicited upon injury and microbial infections. It is activated in response to Upd (unpaired) cytokines released by haemocytes [315,316]. In Drosophila, Upd3 binds to the membrane Domeless receptor [316] which dimerises, causing phosphorylation of the Janus kinase specific to Drosophila called Hopscotch [316,317,318]. In turn, this allows phosphorylation at conserved tyrosine residues of the STAT (signal transducer and activator of transcription) transcription factor. Phosphorylated STAT dimerizes and translocates to the nucleus where it activates expression of target genes [316,318]. These genes include TotA, Tep1 and Tep2 [318] whose products are released into the haemolymph and promote wound healing, resistance to stress, and phagocytosis [287,299]. The JAK/STAT pathway contributes to cellular immune responses, such as haemocyte differentiation, and renewal of intestinal epithelial cells. Aberrant activation of the JAK/STAT pathway can result in nodulation in which the PPO cascade is activated [319]. STAT knockdown by RNAi increased the number of Plasmodium vivax in the midgut of Anopheles aquasalis, and enhanced P. berghei and P. falciparum infection in the digestive tract of An. gambiae [320,321].
The JAK–STAT pathway in mosquitoes also responds to viral infections. Knockdown of the Domeless and Hop genes in An. aegypti enhances the burden of Dengue virus (DENV), while silencing PIAS (a negative regulator of the JAK–STAT pathway) results in enhanced resistance to DENV infection [322]. STAT gene silencing mediated by RNAi reduced the expression of inducible nitric oxide synthase (iNOS) and favoured Leishmania growth in sandflies [323]. Based on gene conservation, other vectors have JAK/STAT pathways but their function in relation to pathogens has yet to be determined in detail.
6.3.4. JNK Signalling
Jun N-terminal kinase (JNK) represents a subgroup of mitogen-activated protein kinases which are evolutionarily conserved in eukaryotic cells and activated by environmental stresses and inflammatory cytokines [324]. Upon activation, JNK phosphorylates the transcription factors Jun and Fos, leading to formation of the Jun/Fos dimer, which activates transcription of target genes. The JNK pathway is required for AMP gene expression in Drosophila [325]. JNK signalling is a key regulator in the mosquito An. gambiae limiting Plasmodium infection [326]. DENV2, ZIKV or CHIKV activates the JNK pathway which reduces viral infection by inducing complement and apoptosis in the salivary glands of Ae. aegypti mosquitoes [327].
6.3.5. Eicosanoid Signalling
Eicosanoids are oxygenated metabolites of polyunsaturated fatty acids known to act as signalling molecules in animals. They are derived from arachidonic acid (AA) released from phospholipids by hydrolytic cleavage mediated by phospholipase A2 (PLA2). In Drosophila, the Imd signalling pathway can be elicited by exposure to bacterial LPS, activating expression of AMPs, and this response can be suppressed by exposure to specific PLA2 inhibitors [328]. As this suppression can be reversed by supplementation with AA, this suggests that eicosanoids are important mediators for relaying the Imd response. Eicosanoids are implicated in both humoral and cellular immune responses, including nodulation, the PPO cascade and haemocyte spreading [250].
In An. gambiae a receptor for the eicosanoid prostoglandin E2 (PGE2) is expressed in oenocytes, differentiated haemocytes implicated in the production of phenoloxidases (POs) leading to melanisation responses. PGE2 regulates both the PPO cascade and also AMP expression in these cells to limit both bacterial infection and Plasmodium oocyte survival [329]. In An. aegypti, experimentally reduced prostaglandin synthesis is associated with decreased expression of components of the Toll and Imd immune pathways, thereby rendering mosquitoes more susceptible to both bacterial and viral (Dengue) infections [330]. It is suggested that prostaglandins control the amplitude of the immune response to guarantee an efficient pathogen clearance. In Rhodnius prolixus, the survival of the parasite T. rangeli in the haemolymph involves the modulation of eicosanoid production (the hormone ecdysone also plays a crucial role in parasite survival in the haemolymph). The parasite inhibits phagocytosis by modulation of the phospholipase A2 and PAF (platelet activating factor)-acetylhydrolase activities [331]. Both enzymes have key roles in eicosanoids and PAF pathways, respectively. Exogenous supplementation of AA prevented parasite infection.
6.3.6. Antiviral Immunity
Viral infections can induce Toll, Imd and JAK/STAT pathways in different insect species, although details on how specific viruses are sensed in this context is unclear even for Drosophila. Infection with Drosophila C virus (DCV) leads to induction of 130 genes, with many of these dependent on the hop gene encoding the JAK kinase [332]. Consequently, hop mutant flies are very susceptible to DCV infection. In addition, DCV can induce the Pelle-dependent nuclear translocation of the Toll-pathway specific NF-kB transcription factor, Dorsal [332]. A functioning Toll pathway is required for resistance to Drosophila X virus [332], and the Imd pathway is implicated in resistance to infections by Sindbis virus [333]. In Ae. aegypti, both Toll and Imd pathways are stimulated by the endosymbiont Wolbachia, and this immune priming is crucial in the subsequent inhibition of dengue infection (DENV) [334]. In addition, the JAK/STAT pathway regulates innate immunity during (DENV) infection of An. aegypti by upregulating the expression of JAK, STAT, and DENV response factors 1 (DENV-1) and 2 (DENV-2) [335]. Suppression of Zika virus (ZIKV) infection in An. aegypti requires the activation of both the Toll and JAK/STAT pathways [336]. RNA interference is also a major immune pathway for limiting arbovirus infection in mosquitoes. Disruption of this defence by CRISPR/Cas9-based knockout of the Argonaute (Ago) gene, encoding the major component of the RNA-induced silencing complex (RISC), impaired the mosquitoes' ability to degrade arbovirus RNA, leading to hyper-infection accompanied by cell lysis and tissue damage [337]. The recently characterized cGAS-STING pathway in Drosophila is required to control viral infections, for example by DCV and Cricket paralysis virus (CrPV) (91), but we are not aware of studies on this pathway in vector insects.
6.3.7. Immune Priming.
As described herein and by numerous previous papers, exposure of insects for the first time to invading organisms, like parasites or pathogens, activates an effective immune response, but now evidence indicates that upon second exposure to the same organism, a more rapid and elevated immune response occurs. This process is called immune priming and has the hallmarks of acquired immunity in vertebrates but without the presence of specific memory cells and lymphoid tissue eg. [339,340,341,342,343,344,345]. The possibility of immune priming in insects is not new and was shown by Karp and his co-workers, many years ago, using grafting and bacterial immunisation experiments [339,340]. The priming effect can be short- or long-term and can be pathogen specific or non-specific [345]. Priming can also be prolonged throughout different developmental stages so that primed larvae can enhance adult immunity [346]. In addition, transgenerational immune priming also occurs by which the adults transfer immune effectors like AMPs or microbial components to the eggs and produces immune priming in the next generation This subject is dealt with in detail by an excellent article by Andreas Vilcinskas [343].
Except for mosquitoes, however, much of the work on insect immune priming has been carried out with non-vector insects. In Drosophila, for example, a recent study with Enterococcus faecalis for immune priming showed that a low dose primes flies to survive longer with a subsequent high dose infection for at least 7 days. This increased survival did not result from a more efficient bacterial clearance [347]. In addition, phagocytosis was required for full priming, since decreased priming occurred after bead-blocking experiments and in flies deficient in Eater. Since primed flies, however, failed to clear bacteria more effectively than non-primed controls then the haemocytes must be playing a none canonical role in the priming process. A clue to this was indicated by phagocytic reprogramming and transcriptional changes in the metabolic pathways of the phagocytes from primed flies [347]. Fortunately, the overview of Gomes et al. [345] on immune priming in Anopheles by Plasmodium provides more information about the role of phagocytes in this process. Plasmodium infection produces primed immunity to subsequent infections and is dependent on increases in circulating granulocytes that mediate immune memory [345]. This enhancement is permanent, with the gut microbiome needed for establishment and the memory of the priming process, and not resulting from the continuation of the initial response to infection This priming process involves different tissues, and eicosanoids are shown to be involved as systemic signaling molecules (see section 5. 7. Triatomines, above). More details are provided in Gomes et al. [345]. Briefly Plasmodium infection results in production of two heme-peroxidases which mediate the midgut synthesis of prostaglandin E2 (PGE2). The systemic PGE2 release results in the production of a haemocyte differentiation factor (HDF), enhancing haemocyte development into granulocytes. Subsequently, ookinete invasion and damage of midgut epithelial cells induces a nitration process and the cells undergo apoptosis. The systemic release of the PGE2 attracts granulocytes to the basal midgut and when the haemocytes contact the nitrated surfaces, they produce microvesicles that result in mosquito complement to destroy the ookinetes [345].
7. Final Comments
There has been an enormous explosion of the literature on insect immunity although, apart from mosquitoes and triatomines, the majority of vector insect species have been neglected. Herein, we give examples of key and also the most relevant recent references and reviews on vector immunity. We have also cited papers on Drosophila for comparative purposes and some of the more original work on insect immunity to acknowledge the contributions of pioneers in this important field without whom insect immunity would still be regarded an irrelevant branch of immunity. We apologise sincerely to authors whose papers have not been included. There is much work still required on vector insect interactions with parasites and pathogens, and areas needing particular attention are indicated. Recently developed innovative molecular techniques should facilitate progress in the study of neglected vector systems such as those of fleas, lice, sandflies and blackflies. After all, if scientists can now analyse the molecular biology and functioning of single haemocytes then insect vectors with limited numbers of blood cells will be open to investigation too. The results of such research should identify important molecules and processes which potentially could be the subject of new control measures.
Author Contributions
N.A.R conceived the project with Professor Gunter Schaub (co-editor). N.A.R, P.D, and M.F. wrote the first draft and C.B.M. and H.C.C. provided critical reviews of the text and additional papers for inclusion. All authors have read and agreed to the publication of the final version of the submitted manuscript.
Funding
The work of P.D. and M.F. was supported by a BBSRC grant: Grant Ref: BB/Y001125/1. Fellowships were provided by Universidade Federal Fluminense (UFF), the Carlos Chagas Filho Foundation for Research Support in the State of Rio de Janeiro (FAPERJ), and National Council for Scientific and Technological Development (CNPq).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The authors confirm that the data supporting the conclusions of this study are cited within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Patramool, S.; Choumet, V.; Surasombatpattana, P.; Sabatier, L.; Thomas, F.; Thongrungkiat, S.; Rabilloud, T.; Boulanger, N.; Biron, D.G.; Missé, D. Update on the proteomics of major arthropod vectors of human and animal pathogens. Proteomics 2012, 12, 3510–3523. [Google Scholar] [CrossRef]
- Soumana, I.H.; Simo, G.; Njiokou, F.; B. ; Abd-Alla, A.M.; Cuny, G.; Geiger, A. The bacterial flora of tsetse fly midgut and its effect on trypanosome transmission. J. Invertebr. Pathol. 2013, 112. Suppl, S89-93. [Google Scholar]
- Gulley, M.M.; Zhang, X.; Michel. K. The roles of serpins in mosquito immunology and physiology. J. Insect Physiol. 2013, 59, 138–147. [Google Scholar] [CrossRef]
- Flores-Villegas, A.L.; Salazar-Schettino, P.M.; Córdoba-Aguilar, A.; Gutiérrez-Cabrera, A.E.; Rojas-Wastavino, G.E.; Bucio-Torres, M.I. , Cabrera-Bravo, M. Immune defence mechanisms of triatomines against bacteria, viruses, fungi and parasites. Bull. Entomol. Res. 2015, 105, 523-532. 105.
- Garcia, G.R.; Maruyama, S.R.; Malardo, T.; Zangirolamo, A.F.; Gardinassi, L.G. The biology of hematophagous arthropods addressed by molecular high-throughput approaches. Austin J. Trop. Med. & Hyg. 2015; 1, 1004. [Google Scholar]
- Saraiva, R.G.; Kang, S.; Simões, M.L.; Angleró-Rodríguez, Y.I.; Dimopoulos, G. Mosquito gut antiparasitic and antiviral immunity. Dev. Comp. Immunol. 2016, 64, 53–64. [Google Scholar] [CrossRef]
- Wang, J.; Song, X.; Wang, M. Peptidoglycan recognition proteins in hematophagous arthropods. Dev. Comp. Immunol. 2018, 83, 89–95. [Google Scholar] [CrossRef]
- Telleria, E.L.; Martins-da-Silva, A.; Tempone, A.J.; Traub-Csekö, Y.M.M. Leishmania, microbiota and sand fly immunity. Parasitology 2018, 145, 1336–1353. [Google Scholar] [CrossRef]
- Brown, L.D. Immunity of fleas (Order Siphonaptera). Dev. Comp. Immunol. 2019, 98, 76–79. [Google Scholar] [CrossRef]
- Koenraadt, C.J.M. Infectious Diseases and Arthropods. The Lancet, Infectious Diseases 2019, 19, 1298. [Google Scholar] [CrossRef]
- Guarneri, A.A. , Schaub, G.A. Interaction of Triatomines with Their Bacterial Microbiota and Trypanosomes. In: Guarneri, A., Lorenzo, M. eds; Triatominae - The Biology of Chagas Disease Vectors. Entomology in Focus, vol 5. Springer, Cham. 2019, pp. 345-386.
- Matetovici, I.; De Vooght, L.; Van Den Abbeele, J. Innate immunity in the tsetse fly (Glossina), vector of African trypanosomes. Dev. Comp. Immunol. 2019, 98, 181–188. [Google Scholar] [CrossRef]
- Salcedo-Porras, N.; Lowenberger, C. The Immune System of Triatomines. In: Guarneri, A., Lorenzo, M. eds; Triatominae - The Biology of Chagas Disease Vectors. Entomology in Focus, vol 5. Springer, Cham, 2019; pp.307-344.
- Tawidian, P.; Rhodes, V.L.; Michel, K. Mosquito-fungus interactions and antifungal immunity. Insect Biochem. Mol. Biol. 2019, 111, 103182. [Google Scholar] [CrossRef]
- King, J.G. Developmental and comparative perspectives on mosquito immunity. Dev. Comp. Immunol. 2020, 103, 103458. [Google Scholar] [CrossRef]
- Raddi, G. , Barletta, A.B.F., Efremova, M., Ramirez, J.L., Cantera, R., Teichmann, S.A., et al.; Mosquito cellular immunity at single-cell resolution. Science 2020, 369, 1128–1132. [Google Scholar] [CrossRef]
- Eleftherianos, I.; Heryanto, C.; Bassal, T.; Zhang, W.; Tettamanti, G.; Mohamed, A. Haemocyte-mediated immunity in insects: Cells, processes and associated components in the fight against pathogens and parasites. Immunology 2021, 164, 401–432. [Google Scholar] [CrossRef]
- Gabrieli, P.; Caccia, S.; Varotto-Boccazzi, I.; Arnoldi, I.; Barbieri, G.; Comandatore, F.; Epis, S. Mosquito trilogy: microbiota, immunity and pathogens, and their implications for the control of disease transmission. Front. Microbiol. 2021, 12, 630438. [Google Scholar] [CrossRef]
- Rosendo Machado, S.; van der Most, T.; Miesen, P. Genetic determinants of antiviral immunity in dipteran insects - compiling the experimental evidence. Dev. Comp. Immunol. 2021, 119, 104010. [Google Scholar] [CrossRef]
- Bahia, A.C.; Barletta, A.B.F.; Lopes, A.H.; De Niz, M. Editorial: Parasite interactions with insect hosts in tropical diseases. Front. Trop. Dis. 2022, 3, 992277. [Google Scholar] [CrossRef]
- Cardoso, M.A.; Brito, T.F.; Brito, I.A.dA.; Berni, M.A.; Coelho, V.L.; Pane, A. The neglected virome of triatomine Insects. Front. Trop. Dis. 2022, 3, 828712. [Google Scholar] [CrossRef]
- Cardoso-Jaime, V.; Tikhe, C.V.; Dong, S.; Dimopoulos, G. The role of mosquito hemocytes in viral infections. Viruses 2022, 14, 2088. [Google Scholar] [CrossRef]
- Cecílio, P. , Cordeiro-da-Silva, A.; Oliveira, F. Sand flies: basic information on the vectors of leishmaniasis and their interactions with Leishmania parasites. Commun. Biol. 2022, 5, 305. [Google Scholar] [CrossRef]
- Ouali, R.; Vieira, L.R.; Salmon, D.; Bousbata, S. Rhodnius prolixus hemolymph immuno-physiology: deciphering the systemic immune response triggered by Trypanosoma cruzi establishment in the vector using quantitative proteomics. Cells, 2022; 11, 1449. [Google Scholar]
- Ramirez, J.L.; Hampton, K.J.; Rosales, A.M.; Muturi, E.J. Multiple mosquito AMPs are needed to potentiate their antifungal effect against entomopathogenic fungi. Front. Microbiol. 2022, 13, 1062383. [Google Scholar] [CrossRef]
- Ratcliffe, N.A.; Furtado Pacheco, J.P. ; Dyson, P,; Castro, H.C.; Gonzalez, M.S.; Azambuja, P.; Mello.; C.B. Overview of paratransgenesis as a strategy to control pathogen transmission by insect vectors. Parasit. Vectors, 2022; 15, 112. [Google Scholar]
- Mwangi, V.I.; Martinez, E.G.; Leda, R.L.; Catunda, M.E.S.L.A.; Dias, A.S.; Padron Antonio, Y.; Guerra, M.D.G.V.B. Resisting an invasion: A review of the triatomine vector (Kissing bug) defense strategies against a Trypanosoma sp infection. Acta Trop. 2023, 238, 106745. [Google Scholar] [CrossRef]
- Parres-Mercader, M.; Pance, A.; Gomez-Dıaz, E. Novel systems to study vector-pathogen interactions in malaria. Front. Cell. Infect. Microbiol. 2023, 13, 1146030. [Google Scholar] [CrossRef]
- Vinayagam, S.; Rajendran, D.; Sekar, K.; Renu, K.; Sattu, K. The microbiota, the malarial parasite, and the mosquito [MMM] - A three-sided relationship. Mol. Biochem. Parasitol. 2023, 253, 111543. [Google Scholar] [CrossRef]
- Wang, J.; Gao, L.; Aksoy, S. Microbiota in disease-transmitting vectors. Nat. Rev. Microbiol. 2023, 21, 604–618. [Google Scholar] [CrossRef]
- Zheng, R. , Wang, Q., Wu, R.; Paradkar, P.N.; Hoffmann, A.A.; Wang G-H. Holobiont perspectives on tripartite interactions among microbiota, mosquitoes, and pathogens. ISME J. 2023; 17, 1143–1152. [Google Scholar]
- Wilkerson, R.C.; Linton, Y.-M.; Strickman, D. Mosquitoes of the World; Johns Hopkins University Press, Baltimore; 2021, 1-2, 1332 pages; ISBN 978-1-421438-14-6.
- Brühl, C.A.; Després, L.; Frör, O. ; Patil. C.D.; Poulin, B.; Tetreau, G.; Allgeier S. Environmental and socioeconomic effects of mosquito control in Europe using the biocide Bacillus thuringiensis subsp. israelensis (Bti). Science of the Total Environment, 2020; 724, 137800. [Google Scholar]
- Lahondère, C.; Vinauger, C.; Okubo, R.P.; Wolff, G.H.; Chan, J.K.; Akbari, O.S.; Riffell, J.A. The olfactory basis of orchid pollination by mosquitoes. Proc. Natl Acad. Sci. USA. 2020, 117, 708–716. [Google Scholar] [CrossRef]
- Agten, S. M, Watson, E.E.; Ripoll-Rozada, J.; Dowman, L.J.; Wu, M.C.L.; Alwis, I.; Jackson, S.P.; Pereira, P.J.B.; Payne, R.J. Potent trivalent inhibitors of thrombin through hybridization of salivary sulfopeptides from hematophagous arthropods. Angew Chem. Int. Ed. Engl. 2021, 60, 5348–5356. [Google Scholar]
- Bellekom, B.; Hackett, T.D.; Lewis, O.T. A network perspective on the vectoring of human disease. Trends Parasitol. 2021, 37, 391–400. [Google Scholar] [CrossRef]
- Stork, N.E. How many species of insects and other terrestrial arthropods are there on Earth? Annu. Rev. Entomol. 2018, 63, 31–45. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Vector Borne Diseases. https://www.who.int/news-room/fact-sheets/detail/vector-borne-diseases. Accessed 1st June, 2023.
- Shaw, W.R.; Catteruccia, F. Vector biology meets disease control: using basic research to fight vector-borne diseases. Nat. Microbiol. 2019, 4, 20–34. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Ethics and vector-borne diseases: WHO guidance. 2020. https://apps.who.int/iris/handle/10665/336075. Accessed 1st June 2023.
- U.S. Center for Disease Control and Prevention (CDC). Malaria‘s impact worldwide. 25 Feb 2020. https://www.cdc.gov›malaria›impact. Accessed 1st June 2023.
- World Health Organization (WHO). Lymphatic filariasis. 2023. https://www.who.int/news-room/fact-sheets/detail/lymphatic-filariasis. Accessed 2nd June 2023.
- Frallonardo, L.; Di Gennaro, F.; Panico, G.G.; Novara, R.; Pellara, E.; Cotugno, S. , Guido, G.; De Vita, E.; Ricciardi A.; Totaro, V.; et al. Onchocerciasis: current knowledge and future goals. Front. Trop. Dis, 2022, 3, 986884. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Onchocerciasis. 2022. https://www.who.int/news-room/fact-sheets/detail/onchocerciasis. Accessed 4th June 2023.
- Bern, C. Chagas’ disease. N. Engl. J. Med. 2015, 373, 456–466. [Google Scholar] [CrossRef]
- Howard, J. Plague (Black Death) bacterial infection information and facts. https://www.nationalgeographic.com/science/article/the-plague. 2020.
- Demeure, C.E.; Dussurget, O.; Mas Fiol, G.; Le Guern, A.S.; Savin, C.; Pizarro-Cerda, J. Yersinia pestis and plague: an updated view on evolution, virulence determinants, immune subversion, vaccination and diagnostics . Genes Immun. 2019, 20, 357–370. [Google Scholar]
- Angelakis, E.; Bechah, Y.; Raoult, D. The History of Epidemic Typhus. Microbiol. Spectr. 2016 Aug;4(4). doi: 10.1128/microbiolspec.PoH-0010-2015. PMID: 27726780. [CrossRef] [PubMed]
- Semenza, J.C.; Suk, J.E. Vector-borne diseases and climate change: a European perspective. FEMS Microbiol. Lett. 2018, 365, fnx244. [Google Scholar] [CrossRef]
- Pan American Health Organization/World Health Organization (PAHO/WHO). Response to the epidemic of Zika virus in the Americas, December 2015–2016. https://www.paho.org/en/topics/zika. Accessed 5th December 2020.
- Lemaitre, B.; Nicolas, E.; Michaut, L.; Reichhart, J.M.; Hoffmann, J.A. The dorsoventral regulatory gene cassette spaetzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell 1996, 86, 973–983. [Google Scholar] [CrossRef]
- Rock, F.; Hardiman, G.; Timans, J.; Kastelein, R.; Bazan, J. A family of human receptors structurally related to Drosophila Toll. Proc. Natl Acad. Sci. USA 1998, 95, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Baldwin, J.; Brar, D.; Salunke, D.B.; Petrovsky, N. Toll-like receptor (TLR) agonists as a driving force behind next-generation vaccine adjuvants and cancer therapeutics. Curr. Opin. Chem. Biol. 2022, 70, 102172. [Google Scholar] [CrossRef]
- Van de Leemput, J. , Han, Z. Drosophila, a powerful model to study virus-host interactions and pathogenicity in the fight against SARS-CoV-2. Cell Biosci. 2021, 11, 110. [Google Scholar]
- Piatek, M.; Sheehan, G.; Kavanagh, K. Galleria mellonella: the versatile host for drug discovery, in vivo toxicity testing and characterising host-pathogen interactions. Antibiotics 2021, 10, 1545. [Google Scholar] [CrossRef]
- Lehane, M.J. The Biology of Blood-Sucking in Insects. Cambridge University Press: Second Ed. Cambridge, UK, 2009. ISBN: 9780521543958.
- Van Nouhuys, S.; Niemikapee, S.; Hanski, I. Variation in a host–parasitoid Interaction across independent populations. Insects 2012, 3, 1236–1256. [Google Scholar] [CrossRef]
- Black, J.L.; Clark, M.K.; Sword, G.A. Physiological and transcriptional immune responses of a non-model arthropod to infection with different entomopathogenic groups. PLoS ONE 2022, 17, e0263620. [Google Scholar] [CrossRef]
- Antonelli, P. , Duval, P., Luis, P.; Minard, G.; Moro, C.V. Reciprocal interactions between anthropogenic stressors and insect microbiota. Environ. Sci. Pollut. Res. 2022; 29, 64469–64488. [Google Scholar]
- Ferreira, A.B.B.; Bahia, A.C.; Pitaluga, A.N.; Barros, E.; dos Santos, D.G.; Bottino-Rojas, V.; Kubota, M.S.; de Oliveira, P.L.; Pimenta, P.F.P.; Traub-Csekö, Y.M.; Sorgine, M.H.F. Sexual dimorphism in immune responses and infection resistance in Aedes aegypti and other hematophagous insect vectors. Front. Trop. Dis. 2022, 3 Article –847109, 1–17. [Google Scholar]
- Booth, K.; Cambron, L.; Fisher, N.; Greenlee, K.J. Immune defense varies within an instar in the tobacco hornworm, Manduca sexta. Physiol. Biochem. Zool. 2015, 88, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Candian, V.; Tedeschi, R. Impact of the diet on the mortality and on gene expression of the antimicrobial peptide Tenecin 3 in Tenebrio molitor larvae infected by Beauveria bassiana. Insects 2023, 14, 359. [Google Scholar] [CrossRef] [PubMed]
- Vilcinskas, A. Evolutionary plasticity of insect immunity. J. Insect Physiol. 2013, 59, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Eleftherianos, I.; Tafesh-Edwards, G.; Mohamed, A. Pathogen infection routes and host innate immunity: lessons from insects. Immunol. Lett. 2022, 247, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, G.; Farrell, G.; Kavanagh, K. Immune priming: the secret weapon of the insect world. Virulence 2020, 11, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Baxter, R.H.G. , Contet, A., Krueger, K. Arthropod innate immune systems and vector-borne diseases. Biochemistry 2017, 56, 907–918. 56,.
- Dyer, N.A.; Rose, C.; Ejeh, N.O.; Acosta-Serrano, A. Flying tryps: survival and maturation of trypanosomes in tsetse flies. Trends Parasitol. 2013, 29, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Hillyer, J.F. Insect immunology and hematopoiesis. Dev. Comp. Immunol. 2016, 58, 102–118. [Google Scholar] [CrossRef]
- Moure, U.A.E; Tan, T.; Sha, L.; Lu, X.; Shao, Z.; Yang, G.; Wang, Y.; Cui, H. Advances in the immune regulatory role of non-coding RNAs (miRNAs and lncRNAs) in insect-pathogen interactions. Front. Immunol. 2022, 13, 856457. [Google Scholar] [CrossRef] [PubMed]
- Rosetto, M.; Engstrom, Y.; Baldari, C.T.; Telford, J.L.; Hultmark, D. Signals from the IL-1 receptor homolog, Toll, can activate an immune response in a Drosophila hemocyte cell line. Biochem. Biophys. Res. Commun. 1995, 209, 111–116. [Google Scholar] [CrossRef]
- Hoffmann, J.A. The immune response of Drosophila. Nature 2003, 426(6962), 33–38. [Google Scholar] [CrossRef]
- Yu, S.; Luo, F.; Xu, Y.; Zhang, Y.; Jin, L.H. Drosophila innate immunity involves multiple signaling pathways and coordinated communication between different tissues. Front. Immunol. 2022, 13, 905370. [Google Scholar] [CrossRef]
- Collins, F.H.; Sakai, R.K.; Vernick, K.D.; Paskewitz, S.; Seeley, D.C.; Miller, L.H.; Collins, W.E.; Campbell, C.C.; Gwadz, R.W. Genetic selection of a Plasmodium-refractory strain of the malaria vector Anopheles gambiae. Science 1986, 234(4776), 607–610. [Google Scholar] [CrossRef]
- Dimopoulos, G. , Richman, A., Muller, H.-M., Kafatos, F. C. Molecular immune responses of the mosquito Anopheles gambiae to bacteria and malaria parasites. Proc. Natl Acad. Sci. USA 1997, 94, 11508–11513. [Google Scholar] [CrossRef] [PubMed]
- Stower, H. Anti-mosquito immunity. Nat. Med. 2020, 26, 1009. [Google Scholar] [CrossRef] [PubMed]
- Hultmark, D.; Steiner, H.; Rasmuson, T.; Boman, H.G. Insect immunity. Purification and properties of three inducible bactericidal proteins from hemolymph of immunized pupae of Hyalophora cecropia. Eur. J. Biochem. 1980, 106, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Dunn, P.E.; Dai, W.; Kanost, M.R.; Geng, C.X. Soluble peptidoglycan fragments stimulate antibacterial protein synthesis by fat body from larvae of Manduca sexta. Dev. Comp. Immunol. 1985, 9, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Shan, T.; Wang, Y.; Dittmer, N.T.; Kanost, M.R.; Jiang, H. Serine protease networks mediate immune responses in extra-embryonic tissues of eggs in the tobacco hornworm, Manduca sexta. J. Innate Immun. 2022, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- De Verno, P.J.; Chadwick, J.S.; Aston, W.P.; Dunphy, G.B. The in vitro generation of an antibacterial activity from the fat body and hemolymph of non-immunized larvae of Galleria mellonella. Dev. Comp. Immunol. 1984, 8, 537–546. [Google Scholar] [CrossRef]
- Sheehan, G.; Margalit, A.; Sheehan, D.; Kavanagh, K. Proteomic profiling of bacterial and fungal induced immune priming in Galleria mellonella larvae. J. Insect Physiol. 2021, 131, 104213. [Google Scholar] [CrossRef]
- Kordaczuk, J.; Sułek, M.; Mak, P.; Zdybicka-Barabas, A.; Śmiałek, J.; Wojda, I. Cationic protein 8 plays multiple roles in Galleria mellonella immunity. Sci. Rep. 2022, 12, 11737. [Google Scholar] [CrossRef]
- Ashida, M.; Ohnishi, E. Activation of pre-phenol oxidase in hemolymph of the silkworm, Bombyx mori. Arch. Biochem. Biophys. 1967, 122, 411–416. [Google Scholar] [CrossRef]
- Wang, R.J.; Chen, K.; Xing, L.S.; Lin, Z.; Zou, Z.; Lu, Z. Reactive oxygen species and antimicrobial peptides are sequentially produced in silkworm midgut in response to bacterial infection. Dev. Comp. Immunol. 2020, 110, 103720. [Google Scholar] [CrossRef] [PubMed]
- Komano, H.; Mizuno, D.; Natori, S. Purification of lectin induced in the hemolymph of Sarcophaga peregrina larvae on injury. J Biol Chem. 1980. 255, 2919–29124. [CrossRef]
- Masova, A.; Sanda, M.; Jiracek, J.; Selicharova, I. Changes in the proteomes of the hemocytes and fat bodies of the flesh fly Sarcophaga bullata larvae after infection by Escherichia coli. Proteome Sci. 2010, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Azambuja, P.; Freitas, C.C.; Garcia, E.S. Evidence and partial characterization of an inducible antibacterial factor in the haemolymph of Rhodnius prolixus. J. Insect Physiol. 1986, 32, 807–812. [Google Scholar] [CrossRef]
- Salcedo-Porras, N.; Oliveira, PL.; Guarneri, A.A.; Lowenberger, C. A fat body transcriptome analysis of the immune responses of Rhodnius prolixus to artificial infections with bacteria. Parasit. Vectors 2022, 15, 269. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.C. Current concepts concerning insect hemocytes. American Zoologist, 1962; 2, 209–246. [Google Scholar]
- Price, C,D. ; Ratcliffe, N.A. A reappraisal of insect haemocyte classification by the examination of blood from fifteen insect orders. Z. Zellforsch. mikrosk. Anat. 1974, 147, 537–549.
- Ribeiro, C.; Brehélin, M. Insect haemocytes: what type of cell is that? J. Insect Physiol. 2006, 52, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Mead, G.P.; Ratcliffe, N.A.; Renwrantz, L.R. The separation of insect haemocyte types on Percoll gradients: methodology and problems. J. Insect Physiol. 1986, 32, 167–177. [Google Scholar] [CrossRef]
- Anggraeni, T.; Ratcliffe, N.A. Studies on cell-cell co-operation during phagocytosis by purified haemocyte populations of the wax moth, Galleria mellonella. J. Insect Physiol. 1991, 37, 453–460. [Google Scholar] [CrossRef]
- Mullett, H.; Ratcliffe, N.A.; Rowley, A.F. Analysis of immune defences of the wax moth, Galleria mellonella, with anti-haemocytic monoclonal antibodies. J. Insect Physiol. 1993, 39, 897–902. [Google Scholar] [CrossRef]
- Tattikota, S.G.; Cho, B.; Liu, Y.; Hu, Y.; Barrera, V.; Steinbaugh, M.J.; Yoon, S.H.; Comjean, A.; Li, F.; Dervis, F.; et al. A single-cell survey of Drosophila blood. eLife 2020, 9, e54818. [Google Scholar] [CrossRef]
- Hultmark, D.; Andó, I. Hematopoietic plasticity mapped in Drosophila and other insects. Elife, 2022, 11, e78906. [Google Scholar] [CrossRef]
- Severo, M.S.; Landry, J.J.M.; Lindquist, R.L.; Goosmann, C.; Brinkmann, V.; Collier, P.; Hauser, A.E.; Benes, V.; Henriksson, J.; Teichmann, S.A.; Levashina, E.A. Unbiased classification of mosquito blood cells by single-cell genomics and high-content imaging. Proc. Natl Acad. Sci. USA. 2018, 115, E7568–E7577. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.; Mohammed, M.; Franzén, O.; Ankarklev, J.; Smith, R. Correction: Single-cell analysis of mosquito hemocytes identifies signatures of immune cell subtypes and cell differentiation. Elife 2022, 11, e85158. [Google Scholar] [CrossRef] [PubMed]
- Gorbet, M.B.; Sefton, M.V. Biomaterial-associated thrombosis: roles of coagulation factors, complement, platelets and leukocytes. Biomaterials 2004, 25, 5681–5703. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, H.I.; John, A.B.; Ichwan, S.J.A.; Kamaruzzaaman, B.Y. Effect of prolonged captivity on the hemolymph profile of Tachypleus gigas using the various anticoagulant formulations. Aquac. Rep. 2021, 20, 1–9. [Google Scholar] [CrossRef]
- Hall, D.W. Mosquito hemocytes: a review. Dev. Comp. Immunol. 1983, 7, 1–12. [Google Scholar] [CrossRef]
- Castillo, J.C.; Robertson, A.E.; Strand, M.R. Characterization of hemocytes from the mosquitoes Anopheles gambiae and Aedes aegypti. Insect Biochem. Mol. Biol. 2006, 36, 891–903. [Google Scholar] [CrossRef]
- Hillyer, J.F.; Strand, M.R. Mosquito hemocyte-mediated immune responses. Curr. Opin. Insect Sci. 2014, 3, 14–21. [Google Scholar] [CrossRef]
- 103. Silva, J.C.; Pessoa, F.A.C., Ríos-Velásquez, C.M., Araújo, H.R.C., Feitosa, A.P.S., Alves, L.C., Brayner, F.A., Eds.; Medeiros, J.F. 2015. Morphological characterization of hemocytes in Ectemnaspis rorotaense (Floch & Abonnenc) and Ectemnaspis trombetense (Hamada, Py-Daniel & Adler) (Diptera: Simuliidae). EntomoBrasilis, 2015; Volume 8, pp. 209–213. [Google Scholar]
- Luckhart, S.; Cupp, M.S.; Cupp, E.W. Morphological and functional classification of the hemocytes of adult female Simulium vittatum (Diptera: Simuliidae), J. Med. Entomol. 1992, 29, 457–466. [Google Scholar] [CrossRef]
- Cupp, M.S.; Chen, Y.; Cupp, E.W. Cellular hemolymph response of Simulium vittatum (Diptera: Simuliidae) to intrathoracic injection of Onchocerca lienalis (Filarioidea: Onchocercidae) microfilariae. J. Med. Entomol. 1997, 34, 56–63. [Google Scholar] [CrossRef] [PubMed]
- De Albuquerque, J.R.F. Identificação das Espécies de Flebotomíneos no Município de Timbaúba/pe e Caracterização dos Hemócitos de Lutzomyia migonei (França, 1920) (Diptera: Psychodidae) Vetor de Leishmania spp. 1918. Fundação Oswaldo Cruz Instituto Aggeu Magalhães, Mestrado Acadêmico em Biociências e Biotecnologia em Saúde, Recife, 68 pages.
- East, J.; Molyneux, D.H.; Hillen, N. Haemocytes of Glossina. Ann. Trop. Med. Parasitol. 1980, 74, 471–474. [Google Scholar] [CrossRef] [PubMed]
- Kaaya, G.P.; Ratcliffe, N.A. Comparative study of hemocytes and associated cells of some medically important dipterans. J. Morphol. 1982, 173, 351–365. [Google Scholar] [CrossRef] [PubMed]
- Coulaud, P.J.; Lepolard, C.; Bechah, Y.; Berenger, J.M.; Raoult, D.; Ghigo, E. Hemocytes from Pediculus humanus humanus are hosts for human bacterial pathogens. Front. Cell Infect. Microbiol. 2015, 4, 183. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, M.P.; Nadeina, V.P.; Chumakova, I.V. Kletki gemolimfy blokh i ikh fagotsitarnaia aktivnost' [Hemolymph cells of fleas and their phagocytic activity. Parazitologiia 1988, 22–321-8. [Google Scholar]
- Muñoz, M.; Lin, N.; Lin, R.; King, B.; Brown, L.D. Immune defense mechanisms against a systemic bacterial infection in the cat flea (Ctenocephalides felis). J. Invert. Pathol. 2022, 195, 107850. [Google Scholar] [CrossRef] [PubMed]
- Azambuja, P.; Garcia, E.S.; Ratcliffe, N.A. Aspects of classification of Hemiptera hemocytes from six triatomine species. Mem.Inst. Oswaldo Cruz 1991, 86, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Moyetta, N.R.; Ramos, F.O.; Leyria, J.; Canavoso, L.E.; Fruttero, L.L. Morphological and ultrastructural characterization of hemocytes in an insect model, the hematophagous Dipetalogaster maxima (Hemiptera: Reduviidae). Insects 2021, 12, 640. [Google Scholar] [CrossRef]
- Agianian, B.; Lesch, C.; Loseva, O.; Dushay, M.S. Preliminary characterization of hemolymph coagulation in Anopheles gambiae larvae. Dev. Comp. Immunol. 2007, 31, 879–888. [Google Scholar] [CrossRef]
- Kim, J.H.; Min, J.S.; Kang, J.S.; Kwon, D.H.; Yoon, K.S.; Strycharz, J.; Koh, Y.H.; Pittendrigh, B.R.; Clark, J.M.; Lee, S.H. Comparison of the humoral and cellular immune responses between body and head lice following bacterial challenge. Insect Biochem. Mol. Biol. 2011, 41, 332–339. [Google Scholar] [CrossRef]
- Dziedziech, A.; Shivankar, S.; Theopold, U. Drosophila melanogaster responses against entomopathogenic nematodes: focus on hemolymph clots. Insects 2020, 11, 62. [Google Scholar] [CrossRef]
- Arai, I.; Ohta, M.; Suzuki, A.; Tanaka, S.; Yoshizawa, Y.; Sato, R. Immunohistochemical analysis of the role of hemocytin in nodule formation in the larvae of the silkworm, Bombyx mori. J. Insect Sci. 2013, 13, 125. [Google Scholar] [CrossRef]
- Nazario-Toole, A.E. , Wu, L. P. Phagocytosis in insect immunity. Adv. Insect Physiol. 2017, 52, 35–82. [Google Scholar]
- Salcedo-Porras, N.; Noor, S.; Cai, C.; Oliveira, P.L.; Lowenberger, C. Rhodnius prolixus uses the peptidoglycan recognition receptor rpPGRP-LC/LA to detect Gram-negative bacteria and activate the IMD pathway. Curr. Res. Insect Sci. 2020; 13, 1:100006. [Google Scholar]
- Blandin, S.A.; Levashina, E.A. Phagocytosis in mosquito immune responses. Immunol. Rev. 2007, 219, 8–16. [Google Scholar] [CrossRef] [PubMed]
- League, G.P.; Hillyer, J.F. Functional integration of the circulatory, immune, and respiratory systems in mosquito larvae: pathogen killing in the hemocyte-rich tracheal tufts. BMC Biol. 2016, 14, 78. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Srivastava, P.; Sirisena, P.; Dubey, S.K.; Kumar, R.; Shrinet, J.; Sunil, S. Mosquito innate immunity. Insects 2018, 9, 95. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.; Smith, R.C. Chemical depletion of phagocytic immune cells in Anopheles gambiae reveals dual roles of mosquito hemocytes in anti-Plasmodium immunity. Proc. Natl Acad. Sci. USA. 2019, 116, 14119–14128. [Google Scholar] [CrossRef] [PubMed]
- Leite, T.H.J.F.; Ferreira, Á.G.A.; Imler, J.L.; Marques, J.T. Distinct roles of hemocytes at different stages of infection by dengue and Zika viruses in Aedes aegypti mosquitoes. Front. Immunol. 2021, 12, 660873. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Sigle, L.T.; Rinker, D.C.; Estévez-Lao, T.Y.; Capra, J.A.; Hillyer, J.F. The immune deficiency and c-Jun N-terminal kinase pathways drive the functional integration of the immune and circulatory systems of mosquitoes. Open Biol. 2022, 12, 220111. [Google Scholar] [CrossRef] [PubMed]
- Hillyer, J.F. Mosquito immunity. Adv. Exp. Med. Biol. 2010, 708, 218–238. [Google Scholar]
- Tinoco-Nunes, B.; Telleria, E.L.; da Silva-Neves, M.; Marques, C.; Azevedo-Brito, D.A.; Pitaluga, A.N.; Traub-Csekö. The sandfly Lutzomyia longipalpis LL5 embryonic cell line has active Toll and Imd pathways and shows immune responses to bacteria, yeast and Leishmania. Parasit Vectors 2016, 9, 222. [Google Scholar] [CrossRef]
- Perotti, M.A.; Allen, J.M.; Reed, D.L.; Braig, H.R. Host-symbiont interactions of the primary endosymbiont of human head and body lice. FASEB J. 2007, 21, 963–1284. [Google Scholar] [CrossRef]
- Previte, D.; Olds, B.P.; Yoon, K.; Sun, W.; Muir, W.; Paige, K.N.; Lee, S.H.; Clark, J.; Koehler, J.E.; Pittendrigh, B.R. Differential gene expression in laboratory strains of human head and body lice when challenged with Bartonella quintana, a pathogenic bacterium. Insect Mol. Biol. 2014, 23, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Russell, C.W.; Johnson, K.L.; Mortensen, R.D.; Erickson, D.L. Gene expression analysis of Xenopsylla cheopis (Siphonaptera: Pulicidae) suggests a role for reactive oxygen species in response to Yersinia pestis infection. J. Med. Entomol. 2012, 49, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Azambuja, P.; Garcia, E.S. Trypanosoma rangeli interactions within the vector Rhodnius prolixus - A mini review. Mem. Inst. Oswaldo Cruz, 2005, 100, 567–572. [Google Scholar] [CrossRef]
- Weiss, B.L.; Maltz, M.; Aksoy, S. Obligate symbionts activate immune system development in the tsetse fly. J. Immunol. 2012, 188, 3395–3403. [Google Scholar] [CrossRef]
- Matetovici, I. , Van Den Abbeele, J. Thioester-containing proteins in the tsetse fly (Glossina) and their response to trypanosome infection. Insect Mol. Biol. 2018; 27, 414–428. [Google Scholar]
- Hao, Z.; Kasumba, I.; Lehane, M.J.; Gibson, W.C.; Kwon, J.; Aksoy, S. Tsetse immune responses and trypanosome transmission: implications for the development of tsetse-based strategies to reduce trypanosomiasis. Proc. Natl Acad; Sci. USA. 2001; 98, 12648–12653. [Google Scholar]
- Azambuja, P.; Garcia, E.S.; Waniek, P.J.; Vieira, C.S.; Figueiredo, M.B.; Gonzalez, M.S.; Mello, C.B.; Castro, D.P.; Ratcliffe, N.A. Rhodnius prolixus: from physiology by Wigglesworth to recent studies of immune system modulation by Trypanosoma cruzi and Trypanosoma rangeli. J. Insect Physiol. 2017, 97, 45–65. [Google Scholar] [CrossRef] [PubMed]
- Salcedo-Porras, N,; Guarneri, A. ; Oliveira, P.L,.Lowenberger, C. Rhodnius prolixus: Identification of missing components of the IMD immune signaling pathway and functional characterization of its role in eliminating bacteria. PLoS One, 2019, 14, e0214794. [Google Scholar]
- Salcedo-Porras, N.; Lowenberger, C. The innate immune system of kissing bugs, vectors of chagas disease. Dev. Comp. Immunol. 2019, 98, 119–128. [Google Scholar] [CrossRef]
- Schaub, G.A. An update on the knowledge of parasite–vector interactions of Chagas Disease. Res. rep. trop. med. 2021, 12, 63–76. [Google Scholar] [CrossRef]
- Alejandro, A.D.; Lilia, J.P.; Jesús, M.B.; Henry, R.M. The IMD and Toll canonical immune pathways of Triatoma pallidipennis are preferentially activated by Gram-negative and Gram-positive bacteria, respectively, but cross-activation also occurs. Parasit. Vectors 2022, 15, 256. [Google Scholar] [CrossRef]
- Borges, A.; Santos, P.N.; Furtado, A.F.; Figueiredo, R.C. Phagocytosis of latex beads and bacteria by hemocytes of the triatomine bug Rhodnius prolixus (Hemiptera: Reduvidae). Micron 2008, 39, 486–494. [Google Scholar] [CrossRef]
- Dubovskiy, I.M.; Kryukova, N.A.; Glupov, V.V.; Ratcliffe, N.A. Encapsulation and nodulation in insects. Invertebr. Surviv. J. 2016, 13, 229–246. [Google Scholar]
- Sato, R. Mechanisms and roles of the first stage of nodule formation in lepidopteran insects. J. Insect Sci. 2023, 23, 3–1-14. [Google Scholar] [CrossRef]
- Ni, W.; Bao, J.; Mo, B.; Liu, L.; Li, T.; Pan, G.; Chen, J.; Zhou, Z. Hemocytin facilitates host immune responses against Nosema bombycis. Dev. Comp. Immunol. 2020, 103, 103495. [Google Scholar] [CrossRef] [PubMed]
- Shiao, S-H. ; Whitten, M.M.A.; Zachary, D.; Hoffmann, J.A.; Levashina, E.A. Fz2 and Cdc42 mediate melanization and actin polymerization but are dispensable for Plasmodium killing in the mosquito midgut. PLoS Pathog. 2006, 2, e133. [Google Scholar]
- Camacho, E.; Dong, Y.; Anglero-Rodriguez, Y.; Smith, D.; Jacomini, R.S.; Dimopoulos, G.; Casadevall, A. Analysis of melanotic Plasmodium spp. capsules in mosquitoes reveal eumelanin-pheomelanin composition and identify AgMesh as a modulator of parasite infection. bioRxiv, 2021.05.07.443077.
- Edgerton, E.B.; McCrea, A.R.; Berry, C.T.; Kwok, J.Y.; Thompson, L.K.; Watson, B.; Povelones, M. Activation of mosquito immunity blocks the development of transmission-stage filarial nematodes. Proc. Natl Acad; Sci. USA. 2020, 117, 3711-3717.
- Liu, C.T.; Hou, R.F.; Chen, C.C. Formation of basement membrane-like structure terminates the cellular encapsulation of microfilariae in the haemocoel of Anopheles quadrimaculatus. Parasitology 1998, 116, 511–518. [Google Scholar] [CrossRef]
- Infanger, L.-C.; Rocheleau, T.A.; Bartholomay, L.C.; Johnson, J.K.; Fuchs, J.; Higgs, S.; Christensen, B.M. (2004). The role of phenylalanine hydroxylase in melanotic encapsulation of filarial worms in two species of mosquitoes. Insect Biochem. Mol. Biol. 2004, 34, 1329–1338. [Google Scholar] [PubMed]
- Binggeli, O.; Neyen, C.; Poidevin, M.; Lemaitre, B. Prophenoloxidase activation Is required for survival to microbial infections in Drosophila. PLoS Pathog. 2014, 10, e1004067. [Google Scholar] [CrossRef]
- Zou, Z.; Shin, S.W.; Alvarez, K.S.; Bian, G.; Kokoza, V.; Raikhel, A.S. Mosquito RUNX4 in the immune regulation of PPO gene expression and its effect on avian malaria parasite infection. Proc. Natl Acad; Sci. USA. 2008; 105, 18454–18459. [Google Scholar]
- Eleftherianos, I.; Heryanto, C. Transcriptomic insights into the insect immune response to nematode infection. Genes 2021, 12, 202. [Google Scholar] [CrossRef]
- Loghry, H.J.; Kwon, H.; Smith, R.C.; Sondjaja, N.A.; Minkler, S.J.; Young, S.; Wheeler, N.J.; Zamanian, M.; Bartholomay, L.C.; Kimber, M.J. ; Extracellular vesicles secreted by Brugia malayi microfilariae modulate the melanization pathway in the mosquito host. Sci. Rep. 2023, 13, 8778. [Google Scholar] [CrossRef]
- Castillo, J.C.; Reynolds, S.E.; Eleftherianos, I. Insect immune responses to nematode parasites. Trends Parasitol. 2011, 27, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Beerntsen, B.T.; Bartholomay, L.C.; Lowery, R.J. Penetration of the mosquito midgut is not required for Brugia pahangi microfilariae to avoid the melanotic encapsulation response of Armigeres subalbatus. Vet. Parasitol. 2007, 144, 371–374. [Google Scholar] [CrossRef] [PubMed]
- Mello, C.B. , Azambuja, P., Garcia, E.S., Ratcliffe, N.A. Differential in vitro and in vivo behavior of three strains of Trypanosoma cruzi in the gut and hemolymph of Rhodnius prolixus. Exp. Parasitol. 1996, 82, 112–121. [Google Scholar] [CrossRef]
- Mello, C.B. , Nigam, Y., Garcia, E.S., Azambuja, P., Newton, R.P., Ratcliffe, N.A. Studies on a haemolymph lectin isolated from Rhodnius prolixus and its interaction with Trypanosoma rangeli. Exp. Parasitol. 1999, 91, 289–296. [Google Scholar] [CrossRef]
- Araújo, C.A.C.; Pacheco, J.P.F.; Waniek, P.J.; Geraldo, R.B.; Sibajev, A.; Dos Santos, A.L.; Evangelho, V.G.O.; Dyson, P.J.; Azambuja, P.; Ratcliffe, N.A.; Castro, H.C.; Mello, C.B. A rhamnose-binding lectin from Rhodnius prolixus and the impact of its silencing on gut bacterial microbiota and Trypanosoma cruzi. Dev. Comp. Immunol, 2021; 114, 103823. [Google Scholar]
- Kaaya, G.P. , Ratcliffe, N.A., Alemu, P. Cellular and humoral defenses of Glossina (Diptera: Glossinidae): reactions against bacteria, trypanosomes, and experimental implants. J. Med. Entomol. 1986; 23, 30–43. [Google Scholar]
- Lee, W.S.; Webster, J.A.; Madzokere, E.T.; Stephenson, E.B.; Herrero, L.J. Mosquito antiviral defense mechanisms: a delicate balance between innate immunity and persistent viral infection. Parasites Vectors 2019, 12, 165. [Google Scholar] [CrossRef]
- Coste Grahl, M.V.; Perin, A.P.A.; Lopes, F.C.; Porto, B.N.; Uberti, A.F.; Canavoso, L.E.; Stanisçuaski, F.; Fruttero, L.L. The role of extracellular nucleic acids in the immune system modulation of Rhodnius prolixus (Hemiptera: Reduviidae). Pestic. Biochem. Physiol. 2020, 167, 104591. [Google Scholar] [CrossRef]
- Holt, R.A.; Subramanian, G.M.; Halpern, A.; Sutton, G.G.; Charlab, R.; Nusskern, D.R.; Wincker, P.; Clark, A.G.; Ribeiro, J.M.; Wides, R.; et al. The genome sequence of the malaria mosquito Anopheles gambiae. Science 2002, 298(5591), 129–149. [Google Scholar] [CrossRef] [PubMed]
- Nene, V.; Wortman, J.R.; Lawson, D.; Haas, B.; Kodira, C.; Tu, Z.J.; Loftus, B.; Xi, Z.; Megy, K.; Grabherr, M.; et al. Genome sequence of Aedes aegypti, a major arbovirus vector. Science 2007, 316(5832), 1718–1723. [Google Scholar] [CrossRef]
- Arensburger, P.; Megy, K.; Waterhouse, R.M.; Abrudan, J.; Amedeo, P.; Antelo, B.; Bartholomay, L.; Bidwell, S.; Caler, E.; Camara, F.; et al. Sequencing of Culex quinquefasciatus establishes a platform for mosquito comparative genomics. Science, 2010; 330(6000), 86–88. [Google Scholar]
- Boissière, A. , Tchioffo, M.T., Bachar, D., Abate, L., Marie, A., Nsango, S.E., Shahbazkia, H.R.; Awono-Ambene, P.H.; Levashina, E.A.; Christen, R.; Morlais, I. Midgut microbiota of the malaria mosquito vector Anopheles gambiae and interactions with Plasmodium falciparum infection. PLoS Pathog. 2012, 8, e1002742. [Google Scholar]
- Bartholomay, L.C.; Waterhouse, R.M.; Mayhew, G.F.; Campbell, C.L.; Michel, K.; Zou, Z.; Ramirez, J.L.; Das, S.; Alvarez, K.; Peter Arensburge, P.; et al. Pathogenomics of Culex quinquefasciatus and meta-analysis of infection responses to diverse pathogens. Science 2010, 330(6000), 88–90. [Google Scholar] [CrossRef]
- Zhang, G.; Niu, G.; Franca, C.M.; Dong, Y.; Wang, X.; Butler, N.S.; Dimopoulos, G.; Li, J. Anopheles midgut FREP1 mediates Plasmodium invasion. J. Biol. Chem. 2015, 290, 16490–16501. [Google Scholar] [CrossRef]
- Erlandson, M.A.; Toprak, U.; Hegedus, D.D. Role of the peritrophic matrix in insect-pathogen interactions. J, Insect Physiol. 2019, 117, 103894. [Google Scholar] [CrossRef] [PubMed]
- Kato, N. , Mueller, C. R., Fuchs, J. F., McElroy, K., Wessely, V., Higgs, S., Christensen, B.M. Evaluation of the function of a type I peritrophic matrix as a physical barrier for midgut epithelium invasion by mosquito-borne pathogens in Aedes aegypti. Vector-Borne Zoonotic Dis. 2008; 8, 701–712. [Google Scholar]
- Bai, L. , Wang, L., Vega-Rodríguez, J., Wang, G., and Wang, S. A gut symbiotic bacterium Serratia marcescens renders mosquito resistance to Plasmodium infection through activation of mosquito immune responses. Front. Microbiol. 2019, 10, 1580. [Google Scholar] [CrossRef]
- Harsh, S.; Eleftherianos, I. Flavivirus infection and regulation of host immune and tissue homeostasis in insects. Front. Immunol. 2020, 11, 618801. [Google Scholar] [CrossRef]
- Rodgers, F.H.; Gendrin, M.; Christophides, G.K. Chapter 6 - the mosquito immune system and its interactions with the microbiota: implications for disease transmission. In: Wikel, S.K.; Aksoy, S.; Dimopoulos, G, editors. Arthropod Vector: Controller of Disease Transmission, Volume 1 [Internet]: Academic; 2017, pp. 101–22. [cited 2020 Oct 29]. Available from: http://www.sciencedirect.com/science/article/pii/B9780128053508000064.
- Lowenberger, C.A. Ferdig, M.T., Bulet, P., Khalili, S., Hoffmann, J.A., Christensen, B.M. Aedes aegypti-Induced antibacterial proteins reduce the establishment and development of Brugia malayi. Exp. Parasitol. 1996, 83, 191–201.
- Ratcliffe, N.A.; Whitten, M.M.A. Vector immunity in microbe–vector interactions. Microbe–Vector Interactions in Vector-borne Diseases (ed. by Gillespie, S.H.; Smith, G.L.; Osbourn, A.), 2004, pp. 199–262, SGM Symposium 63. Cambridge University Press, Cambridge, U.K.
- Sousa, G.L.; Bishnoi, R.; Baxter, R.H.G.; Povelones, M. The CLIP-domain serine protease CLIPC9 regulates melanization downstream of SPCLIP1, CLIPA8, and CLIPA28 in the malaria vector Anopheles gambiae. PLoS Pathog. 2020, 16, e1008985. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Cheng, J.; Mu, X.; Kuang, X.; Li, Z.; Wu, J.A. C-type lectin in saliva of Aedes albopictus (Diptera: Culicidae) bind and agglutinate microorganisms with broad spectrum. J. Insect Sci. 2023, 23, 1. [Google Scholar] [CrossRef]
- Chen, C.; Durrant, H.J.; Newton, R.P.; Ratcliffe, N.A. A study of novel lectins and their involvement in the activation of the prophenoloxidase system in Blaberus discoidalis. Biochem J. 1995, 310, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Simões, M.L.; Dong, Y.; Mlambo, G.; Dimopoulos, G. C-type lectin 4 regulates broad-spectrum melanization-based refractoriness to malaria parasites. PLoS Biol, 2022, 20, e3001515. [Google Scholar] [CrossRef]
- Bukhari, T.; Aimanianda, V.; Bischoff, E.; Brito-Fravallo, E.; Eiglmeier, K.; Riehle, M.M.; Vernick, K.D.; Mitri, C. Genetics and immunity of Anopheles response to the entomopathogenic fungus Metarhizium anisopliae overlap with immunity to Plasmodium. Sci. Rep. 2022, 12, 6315. [Google Scholar] [CrossRef] [PubMed]
- Molina-Cruz, A.; DeJong, R.J.; Charles, B.; Gupta, L.; Kumar, S.; Jaramillo-Gutierrez, G.; Barillas-Mury, C. Reactive oxygen species modulate Anopheles gambiae immunity against bacteria and Plasmodium. J. Biol. Chem. 2008, 283, 3217–3223. [Google Scholar] [CrossRef]
- Barillas-Mury, C. CLIP proteases and Plasmodium melanization in Anopheles gambiae. Trends Parasitol. 2007, 23, 297–299. [Google Scholar] [CrossRef]
- Gonçalves, R.L.; Oliveira, J.H.; Oliveira, G.A.; Andersen, J.F.; Oliveira, M.F.; Oliveira, P.L.; Barillas-Mury, C. Mitochondrial reactive oxygen species modulate mosquito susceptibility to Plasmodium infection. PLoS One 2012, 7, e41083. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulos, G. , Seeley, D., Wolf, A.; Kafatos, F.C. Malaria infection of the mosquito Anopheles gambiae activates immune-responsive genes during critical transition stages of the parasite life cycle. EMBO J. 1998, 17, 6115–6123. [Google Scholar] [CrossRef] [PubMed]
- Luckhart, S.; Vodovotz, Y.; Cui, L.; Rosenberg, R. The mosquito Anopheles stephensi limits malaria parasite development with inducible synthesis of nitric oxide. Proc. Natl Acad. Sci. USA 1998, 95, 5700–5705. [Google Scholar] [CrossRef]
- 184. Oliveira, Gde. A.; Lieberman, J.; Barillas-Mury, C. Epithelial nitration by a peroxidase/NOX5 system mediates mosquito antiplasmodial immunity. Science, 2012; 335(6070), 856–859.
- Castillo, J.C.; Ferreira, A.B.B.; Trisnadi, N.; Barillas-Mury, C. Activation of mosquito complement antiplasmodial response requires cellular immunity. Sci. Immunol. 2017, 2, eaal1505. [Google Scholar] [CrossRef]
- Keleta, Y. , Ramelow, J., Cui, L.; Li, J. Molecular interactions between parasite and mosquito during midgut invasion as targets to block malaria transmission. npj Vaccines 2021, 6, 140. [Google Scholar] [CrossRef]
- McCoy, K.D. , Weldon, C.T., Ansumana, R.; Lamin, J.M.; Stenger, D.A.; Ryan, S.J.; Bardosh, K.; Jacobsen, K.H.; Dinglasan, R.R. Are malaria transmission-blocking vaccines acceptable to high burden communities? Results from a mixed methods study in Bo, Sierra Leone. Malar. J. 2021; 20, 183. [Google Scholar]
- Molina-Cruz, A.; Canepa, G.E.; Alves, E.; Silva, T.L.; Williams, A.E.; Nagyal, S.; Yenkoidiok-Douti, L.; Nagata, B.M.; Calvo, E.; Andersen, J.; Boulanger, M.J.; Barillas-Mury, C. Plasmodium falciparum evades immunity of anopheline mosquitoes by interacting with a Pfs47 midgut receptor. Proc. Natl Acad. Sci. USA. 2020, 117, 2597–2605. [Google Scholar] [CrossRef]
- Vogel, K.J.; Coon, K.L. Chapter Seven - Functions and Mechanisms of Symbionts of Insect Disease Vectors, Editor(s): Oliver, K.M.; Russell, J.A. Advances in Insect Physiology, Academic Press, 2020, 58, 233–275. [Google Scholar]
- Ramirez, J.L. , Souza-Neto, J., Torres Cosme, R., Rovira, J., Ortiz, A., Pascale J.M., Dimopoulos, G. Reciprocal tripartite interactions between the Aedes aegypti midgut microbiota, innate immune system and dengue virus influences vector competence. PLoS Negl. Trop. Dis. 2012, 6, e1561. [Google Scholar] [CrossRef]
- Gómez, M.; Martinez, D.; Muñoz, M.; Ramírez, J.D. Aedes aegypti and Ae. albopictus microbiome/virome: new strategies for controlling arboviral transmission? Parasit. Vectors 2022, 15, 287. [Google Scholar] [CrossRef]
- Scolari, F.; Casiraghi, M.; Bonizzoni, M. Aedes spp. and their microbiota: a review. Front. Microbiol. 2019, 10, 2036. [Google Scholar] [CrossRef]
- Gendrin, M.; Christophides, G.K. The Anopheles Mosquito Microbiota and Their Impact on Pathogen Transmission. In: Manguin, S., editor. Anopheles mosquitoes-new insights into malaria vectors. London: IntechOpen; 2013. pp. 525–48.
- Guégan, M.; Zouache, K.; Démichel, C.; Minard, G.; Van, T.V.; Potier, P.; Mavingui, P.; Moro, C.V. The mosquito holobiont: fresh insight into mosquito–microbiota interactions. Microbiome 2018, 6, 49. [Google Scholar] [CrossRef]
- Nilsson, L.K.J.; de Oliveira, M.R.; Marinotti, O.; Rocha, E.M.; Håkansson, S.; Tadei, W.P.; de Souza, A.Q.L.; Terenius, O. Characterization of bacterial communities in breeding waters of Anopheles darlingi in Manaus in the Amazon Basin malaria endemic area. Microb. Ecol. 2019, 78, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Bongio, N.J.; Lampe, D.J. Inhibition of Plasmodium berghei development in mosquitoes by effector proteins secreted from Asaia sp. bacteria using a novel native secretion signal. PLoS ONE 2015, 10, e0143541. [Google Scholar] [CrossRef]
- Cappelli, A.; Damiani, C.; Mancini, M.V.; Valzano, M.; Rossi, P. ; Serrao, A, Ricci, I.; Favia, G. Asaia activates immune genes in mosquito eliciting an anti-Plasmodium response: implications in malaria control. Front. Genet, 2019; 10, 836. [Google Scholar]
- Rossi, P.; Ricci, I.; Cappelli, A.; Damiani, C.; Ulissi, U.; Mancini, M.V.; Capone, A.; Epis, S.; Crotti, E.; Chouaia, B.; et al. Mutual exclusion of Asaia and Wolbachia in the reproductive organs of mosquito vectors. Parasit. Vectors 2015, 8, 278. [Google Scholar] [CrossRef]
- Wang, S.; Jacobs-Lorena, M. Genetic approaches to interfere with malaria transmission by vector mosquitoes. Trends Biotechnol. 2013, 31, 185–193. [Google Scholar] [CrossRef]
- Adler, P.H. , McCreadie, J.W. Black Flies (Simuliidae). Med. Vet. Entomol. 2019, 237–259. https://api.semanticscholar.org/CorpusID:91385523.
- Chagas, A.C.; Calvo, E.; Pimenta, P.F.; Ribeiro, J.M. An insight into the sialome of Simulium guianense (DIPTERA: SIMUlIIDAE), the main vector of River Blindness Disease in Brazil. BMC Genomics 2011, 12, 612. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Mu, L.; Zhuang, L.; Han, Y.; Liu, T.; Li, J.; Yang, Y.; Yang, H.; Wei, L. A cecropin-like antimicrobial peptide with anti-inflammatory activity from the black fly salivary glands. Parasit. Vectors 2015, 8, 561. [Google Scholar] [CrossRef]
- Ham, P.J.; Albuquerque, C.; Baxter, A.J.; Chalk, R.; Hagen, H.E. Approaches to vector control: new and trusted. 1. Humoral immune responses in blackfly and mosquito vectors of filariae. Trans. R. Soc. Trop. Med. Hyg. 1994, 88, 132–135. [Google Scholar] [CrossRef] [PubMed]
- Kläger, S.L.; Watson, A.; Achukwi, D.; Hultmark, D.; Hagen, H.-E. Humoral immune response of Simulium damnosum s.l. following filarial and bacterial infections. Parasitology 2002, 125, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Ham, P.J. Vector immunity: new potential for disease control? Annales de la Societe Beige de Medecine Tropicale 1991, 71, 179–l87. [Google Scholar]
- Efon Ekangouo, A.; Nana Djeunga, H.C.; Sempere, G.; Kamgno, J.; Njiokou, F.; Moundipa Fewou, P.; Geiger, A. Bacteriome diversity of blackflies' gut and association with Onchocerca volvulus, the causative agent of onchocerciasis in Mbam Valley (Center Region, Cameroon). Pathogens 2021, 11, 44. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Leishmaniasis. 2023. https://www.who.int/news-room/fact-sheets/detail/leishmaniasis. Accessed 15 Nov 2023.
- Dillon, R.J.; Ivens, A.C.; Churcher, C.; Holroyd, N.; Quail, M.A.; Rogers, M.E.; Soares, M.B.; Bonaldo, M.F.; Casavant, T.L.; Lehane, M.J.; Bates, P.A. Analysis of ESTs from Lutzomyia longipalpis sand flies and their contribution toward understanding the insect-parasite relationship. Genomics 2006, 88, 831–840. [Google Scholar] [CrossRef]
- Boulanger, N.; Lowenberger, C.; Volf, P.; Ursic, R.; Sigutova, L.; Sabatier, L.; Svobodova, M.; Beverley, S.M.; Späth, G.; Reto Brun, R.; et al. Characterization of a defensin from the sand fly Phlebotomus duboscqi induced by challenge with bacteria or the protozoan parasite Leishmania major. Infect. Immun. 2004, 72, 7140–7146. [Google Scholar] [CrossRef] [PubMed]
- Coutinho-Abreu, I.V. , Serafim, T.D., Meneses, C.; Kamhawi, S.; Oliveira, F.; Valenzuela, J.G. Leishmania infection induces a limited differential gene expression in the sand fly midgut. BMC Genomics, 2020; 21, 608. [Google Scholar]
- Kykalová, B.; Tichá, L.; Volf, P.; Loza Telleria, E. Phlebotomus papatasi antimicrobial peptides in larvae and females and a gut-specific defensin upregulated by Leishmania major Infection. Microorganisms 2021, 9, 2307. [Google Scholar] [CrossRef]
- Omondi, Z.N.; Arserim, S.K.; Töz, S.; Özbel, Y. Host-parasite interactions: Regulation of Leishmania infection in sand fly. Acta Parasitol. 2022, 67, 606–618. [Google Scholar] [CrossRef]
- Campolina, T.B.; Villegas, L.E.M.; Monteiro, C.C.; Pimenta, P.F.P.; Secundino, N.F. C Tripartite interactions: Leishmania, microbiota and Lutzomyia longipalpis. PLoS Negl. Trop. Dis. 2020, 14, e0008666. [Google Scholar] [CrossRef]
- Dostálová, A. , Volf, P. Leishmania development in sand flies: parasite-vector interactions overview. Parasites Vectors, 2012; 5, 276. [Google Scholar]
- Wijerathna, T.0; Gunathunga, S.; Gunathilaka, H.N. Recent developments and future directions in the paratransgenesis based control of Leishmania transmission. Biol. Control 2020, 145, 104260. [Google Scholar] [CrossRef]
- Kelly, P.H.; Bahr, S.M.; Serafim, T.D.; Ajami, N.J.; Petrosino, J.F.; Meneses, C.; Kirby, J.R.; Valenzuela, J.G.; Kamhawi, S.; Wilson, M.E. The gut microbiome of the vector Lutzomyia longipalpis is essential for survival of Leishmania infantum. mBio, 2017; 38, e01121-16. [Google Scholar]
- Jeffries, C.L.; Rogers, M.E.; Walker, T. Establishment of a method for Lutzomyia longipalpis sand fly embryo microinjection: The first step towards potential novel control strategies for leishmaniasis. Wellcome Open Res. 2018, 3, 55. [Google Scholar] [CrossRef] [PubMed]
- Serafim, T.D.; Coutinho-Abreu, I.V.; Dey, R.; Kissinger, R.; Valenzuela, J.G.; Oliveira, F.; Kamhawi, S. Leishmaniasis: the act of transmission. Trends Parasitol. 2021, 37, 976–987. [Google Scholar] [CrossRef] [PubMed]
- Aoki, V.; Abdeladhim, M.; Li, N.; Cecilio, P.; Prisayanh, P.; Diaz, L.A.; Valenzuela, J.G. Some good and some bad: Sand fly salivary proteins in the control of leishmaniasis and in autoimmunity. Front. Cell. Infect. Microbiol. 2022, 12, 839932. [Google Scholar] [CrossRef] [PubMed]
- Krafsur, E.S. Tsetse fies: genetics, evolution, and role as vectors. Infect. Genet. Evol. 2009, 9, 124–141. [Google Scholar] [CrossRef] [PubMed]
- International Glossina Genome Initiative. Genome sequence of the tsetse fly (Glossina morsitans): vector of African trypanosomiasis. Science. 2014 Apr 25; 344(6182): 380-386. PMID: 24763584; PMCID: PMC4077534. [CrossRef]
- Attardo, G.M. , Abd-Alla, A.M.M., Acosta-Serrano, A.; Allen, J.E.; Bateta, R.; Benoit, J.B.; Bourtzis, K.; Caers, J.; Caljon, G.; Christensen, M.B.; et al. Comparative genomic analysis of six Glossina genomes, vectors of African trypanosomes. Genome Biol. 2019, 20, 187. [Google Scholar] [CrossRef]
- Weiss, B.L.; Wang, J.; Aksoy, S. Tsetse immune system maturation requires the presence of obligate symbionts in larvae. PLoS Biol. 2011, 9, e1000619. [Google Scholar] [CrossRef] [PubMed]
- Benoit, J.B.; Vigneron, A.; Broderick, N. A, Wu, Y.; Sun, J.S.; Carlson, J.R.; Aksoy, S.; Weiss, B.L. Symbiont-induced odorant binding proteins mediate insect host hematopoiesis. Elife 2017, 6, e19535. [Google Scholar] [CrossRef]
- Wang, J.; Wu, Y.; Yang, G.; Aksoy, S. Interactions between mutualist Wigglesworthia and tsetse peptidoglycan recognition protein (PGRP-LB) influence trypanosome transmission. Proc. Natl Acad. Sci. USA. 2009, 106, 2133–2138. [Google Scholar] [CrossRef]
- Wang, J.; Aksoy, S. PGRP-LB is a maternally transmitted immune milk protein that influences symbiosis and parasitism in tsetse's offspring. Proc. Natl Acad. Sci. USA. 2012, 109, 10552–10557. [Google Scholar] [CrossRef]
- Amanzougaghene, N. , Mediannikov, O., Ly, T.D.A.; Gautret, P.; Davoust, B.; Fenollar, F.; Izri, A. Molecular investigation and genetic diversity of Pediculus and Pthirus lice in France. Parasites Vectors 2020, 13, 177. [Google Scholar] [CrossRef]
- Kirkness, E.F.; Haas, B.J.; Sun, W.; Braig, H.R.; Perotti, M.; Clark, J.M.; Lee, S.H.; Robertson, H.M.; Kennedy, R.C.; Elhaik, E. et al. Genome sequences of the human body louse and its primary endosymbiont provide insights into the permanent parasitic lifestyle. Proc. Natl Acad. Sci. USA. 2010, 107, 12168-12173. doi:10.1073/pnas.1003379107. Epub 2010 Jun 21. Erratum in: Proc Natl Acad Sci USA. 2011 Apr 12, 108(15), 6335. Krause, E. [corrected to Kraus, E. PMID: 20566863; PMCID: PMC2901460. [CrossRef] [PubMed]
- Amanzougaghene, N.; Fenollar, F.; Raoult, D.; Mediannikov, O. Where are we with human lice? A review of the current state of knowledge. Front. Cell, Infect. Microbiol, 2020; 9, 474. [Google Scholar]
- Kim, J.H. , Previte, D.J., Yoon, K.S., Murenzi, E., Koehler, J.E., Pittendrigh, B.R., Lee, S.H.; Clarke, J.M. Comparison of the proliferation and excretion of Bartonella quintana between body and head lice following oral challenge. Insect Mol. Biol. 2017, 26, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Burkhart, C.N.; Burkhart, C.G. Bacterial symbiotes, their presence in head lice, and potential treatment avenues. J. Cutan. Med. Surg. 2006, 10, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.E. Resume of the Siphonaptera (Insecta) of the World. J. Med. Entomol. 1999, 35, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Bitam, I.; Dittmar, K.; Parola, P.; Whiting, M.F.; Raoult, D. Fleas and flea-borne diseases. Int. J. Infect. Dis. 2010, 14, e667–76. [Google Scholar] [CrossRef]
- Miarinjara, A.; Bland, D.M.; Belthoff, J.R.; Hinnebusch, B.J. Poor vector competence of the human flea, Pulex irritans, to transmit Yersinia pestis. Parasites Vectors 2021, 14, 317. [Google Scholar] [CrossRef]
- Danchenko, M.; Laukaitis, H.J.; Macaluso, K.R. Dynamic gene expression in salivary glands of the cat flea during Rickettsia felis infection. Pathog. Dis. 2021, 79, ftab020. [Google Scholar] [CrossRef]
- Driscoll, T.P.; Verhoeve, V.I.; Gillespie, J.J.; Johnston, J.S.; Guillotte, M.L.; Rennoll-Bankert, K.E.; Rahman, S.; Hagen, D.; Elsik, C.G.; Macaluso, K.R.; Azad, A.F. A chromosome-level assembly of the cat flea genome uncovers rampant gene duplication and genome size plasticity. BMC Biology 2020, 18. [Google Scholar] [CrossRef]
- Bland, D.M.; Martens, C.A.; Virtaneva, K.; Kanakabandi, K.; Long, D.; Rosenke, R.; Saturday, G.A.; Hoyt, F.H.; Bruno, D.P.; Ribeiro, J.M.; et al. Transcriptomic profiling of the digestive tract of the rat flea, Xenopsylla cheopis, following blood feeding and infection with Yersinia pestis. PLoS Negl. Trop. Dis. 2020, 14, e0008688. [Google Scholar] [CrossRef]
- Vadyvaloo, V.; Viall, A.K.; Jarrett, C.O.; Hinz, A.K.; Sturdevant, D.E.; Joseph Hinnebusch, B. Role of the PhoP-PhoQ gene regulatory system in adaptation of Yersinia pestis to environmental stress in the flea digestive tract. Microbiology (Reading, England), 2015; 161, 1198–1210. [Google Scholar]
- Dreher-Lesnick, S.M.; Ceraul, S.M.; Lesnick, S.C.; Gillespie, J.J.; Anderson, J.M.; Jochim, R.C.; Valenzuela, J.G.; A. F. Azad, A.F. Analysis of Rickettsia typhi-infected and uninfected cat flea (Ctenocephalides felis) midgut cDNA libraries: deciphering molecular pathways involved in host response to R. typhi infection. Insect Mol. Biol. 2010, 19, 229–241. [Google Scholar] [CrossRef]
- Laukaitis, H.J.; Macaluso, K. Unpacking the intricacies of Rickettsia–vector interactions. Trends Parasitol. 2021, 37, 734–746. [Google Scholar] [CrossRef]
- Beliavskaia, A.; Tan, K-K. ; Sinha, A.; Husin, N.A.; Lim, F.S.; Loong, S.K.; Bell-Sakyi, L.; Carlow, C.K.S.; AbuBakar, S.; Darby, A.C.; Makepeace, B.L.; Khoo, J.J. Metagenomics of culture isolates and insect tissue illuminate the evolution of Wolbachia, Rickettsia and Bartonella symbionts in Ctenocephalides spp. Fleas. Microbial Genomics 2023, 9, 001045. [Google Scholar]
- Coura, J.R.; Viñas, P.A. Chagas Disease: A New Worldwide Challenge. Nature, 2010; 465(7301), S6–7. [Google Scholar] [CrossRef]
- Vieira, C.B.; Praça, Y.R.; Bentes, K.L.; da Santos Santiago, P.B.; Silva, S.M.M.; Silva, G.D.S.; Motta, F.N.; Bastos, I.M.D.; Santana, J.M.; de Araújo, C.N.; et al. Triatomines: trypanosomatids, bacteria, and viruses potential vectors? Front. Cell. Infect. Microbiol. 2018, 8, 405. [Google Scholar] [CrossRef]
- Dario, M.A.; Pavan, M.G.; Rodrigues, M.S.; Lisboa, C.V.; Kluyber, D.; Desbiez, A.L.J.; Herrera, H.M.; Roque, A.L.R.; Lima, L.; Teixeira, M.M.G.; Jansen, A.M. Trypanosoma rangeli genetics, mammalian hosts, and geographical diversity from five Brazilian biomes. Pathogens 2021, 10, 736. [Google Scholar] [CrossRef]
- Tobie, E.J. Observation on the development of Trypanosoma rangeli in the haemocoel of Rhodnius prolixus. J. Invertebr. Pathol, 1970, 15, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Dias, J.C.P. , Schofield, C.J. Controle da transmissão transfusional da doença de Chagas na Iniciativa do Cone Sul. Rev. Soc. Bras. Med. Trop. 1998, 31, 373–383. [Google Scholar] [CrossRef]
- Wigglesworth, V.B. The Principles of Insect Physiology, (seventh ed.), Chapman and Hall, London (1974).
- Mesquita, R.D.; Vionette-Amaral, R.J.; Lowenberger, C.; Rivera-Pomar, R.; Monteiro, F.A.; Minx, P.; Spieth, J.; Carvalho, A.B.; Panzera, F.; Lawson, D. et al. Genome of Rhodnius prolixus, an insect vector of Chagas disease, reveals unique adaptations to hematophagy and parasite infection. Proc. Natl Acad. Sci. USA, 2015; 112(48), 14936–14941. [Google Scholar]
- Zumaya-Estrada, F.A.; Martínez-Barnetche, J.; Lavore, A.; Rivera-Pomar, R.; Rodríguez, M.H. Comparative genomics analysis of triatomines reveals common first line and inducible immunity-related genes and the absence of Imd canonical components among hemimetabolous arthropods. Parasit. Vectors 2018, 11, 48. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Stanley, D. Eicosanoid signaling in insect immunology: new genes and unresolved Issues. Genes (Basel) 2021, 12, 211. [Google Scholar] [CrossRef]
- Carmona-Peña, S.P.; Contreras-Garduño, J.; Castro, D.P.; Manjarrez, J.; Vázquez-Chagoyán, J.C. The innate immune response of triatomines against Trypanosoma cruzi and Trypanosoma rangeli with an unresolved question: Do triatomines have immune memory? Acta Trop. 2021, 224, 106108. [Google Scholar] [CrossRef]
- Eberhard, F.E. , Klimpel, S., Guarneri, A.A.; Tobias, N.J. Exposure to Trypanosoma parasites induces changes in the microbiome of the Chagas disease vector Rhodnius prolixus. Microbiome 2022, 10, 45. [Google Scholar] [CrossRef]
- Barbosa, H.J.; Quevedo, Y.S.; Torres, A. M, Veloza, G.A.G.; Martínez, J.C.C.; Urrea-Montes, D.A.; Robello-Porto, C.; Vallejo, G.A. Comparative proteomic analysis of the hemolymph and salivary glands of Rhodnius prolixus and R. colombiensis reveals candidates associated with differential lytic activity against Trypanosoma cruzi I and T. cruzi II. bioRxiv, 2023. [Google Scholar] [CrossRef]
- Whitten, M.M.A. , Mello, C.B., Gomes, S.A.O., Nigam, Y., Azambuja, P., Garcia, E.S., Ratcliffe, N.A. Role of superoxide and reactive nitrogen intermediates in Rhodnius prolixus (Reduviidae)/Trypanosoma rangeli interactions. Exp. Parasitol, 2001; 98, 44–57. [Google Scholar]
- Whitten, M.; Sun, F.; Tew, I.; Schaub, G.; Soukou, C.; Nappi, A.; Ratcliffe, N. Differential modulation of Rhodnius prolixus nitric oxide activities following challenge with Trypanosoma rangeli, T. cruzi and bacterial cell wall components. Insect Biochem. Mol. Biol. 2007, 37, 440–452. [Google Scholar] [CrossRef]
- Castro, D.P. , Moraes, C.S., Gonzalez, M.S., Ratcliffe, N.A., Azambuja, P., Garcia, E.S. Trypanosoma cruzi immune response modulation decreases microbiota in Rhodnius prolixus gut and is crucial for parasite survival and development. PLoS One 2012, 7, e36591. [Google Scholar]
- Batista, K.K.D.S.; Vieira, C.S.; Florentino, E.B.; Caruso, K.F.B.; Teixeira, P.T.P.; Moraes, C.D.S.; Genta, F.A.; Azambuja, P.; Castro, D.P. Nitric oxide effects on Rhodnius prolixus's immune responses, gut microbiota and Trypanosoma cruzi development. J. Insect Physiol. 2020, 126, 104100. [Google Scholar] [CrossRef]
- Gomes, S.A.; Feder, D.; Garcia, E.S.; Azambuja, P. Suppression of the prophenoloxidase system in Rhodnius prolixus orally infected with Trypanosoma rangeli. J. Insect Physiol. 2003, 49, 829–837. [Google Scholar] [CrossRef]
- Paranaiba, L.F.; Guarneri, A.A.; Torrecilhas, A.C.; Melo, M.N.; Soares, R.P. Extracellular vesicles isolated from Trypanosoma cruzi affect early parasite migration in the gut of Rhodnius prolixus but not in Triatoma infestans. Mem. Inst. Oswaldo Cruz, 2019; 114, e190217. [Google Scholar]
- Mantel, P.Y.; Marti, M. The role of extracellular vesicles in Plasmodium and other protozoan parasites. Cell. Microbiol. 2014, 16, 344–354. [Google Scholar] [CrossRef]
- Stanley, D.W. , Kim, Y. Eicosanoid signaling in insects: from discovery to plant protection. Crit. Rev. Plant Sci. 2014, 33, 20–63. [Google Scholar] [CrossRef]
- Stanley, D.; Kim, Y. Prostaglandins and other eicosanoids in insects: biosynthesis and biological actions. Front. Physiol. 2019, 9, 1927. [Google Scholar] [CrossRef]
- 263. . Barletta, A.B.F., Trisnadi, N., Ramirez, J.L., Barillas-Mury, C. Mosquito midgut prostaglandin release establishes systemic immune priming. iScience 2019, 19, 54–62. [Google Scholar] [CrossRef]
- Garcia, E.S.; Machado, E.M.; Azambuja, P. Effects of eicosanoid biosynthesis inhibitors on the prophenoloxidase-activating system and microaggregation reactions in the hemolymph of Rhodnius prolixus infected with Trypanosoma rangeli. J. Insect Physiol. 2004, 50, 157–165. [Google Scholar] [CrossRef]
- Tunctan, B.; Korkmaz, B.; Cuez, T.; Kemal Buharalioglu, C.; Sahan-Firat, S.; Falck, J.; Malik, K.U. Contribution of vasoactive eicosanoids and nitric oxide production to the effect of selective cyclooxygenase-2 inhibitor, NS-398, on endotoxin-induced hypotension in rats. Basic Clin, Pharmaco.l Toxicol. 2010, 107, 877–882. [Google Scholar] [CrossRef]
- Wang, P.H.; He, J.G. Nucleic acid sensing in invertebrate antiviral immunity. Int. Rev. Cell Mol. Biol. 2019, 345, 287–360. [Google Scholar]
- Zhang, W.; Tettamanti, G.; Bassal, T.; Heryanto, C.; Eleftherianos, I.; Mohamed, A. Regulators and signalling in insect antimicrobial innate immunity: Functional molecules and cellular pathways. Cell Signal. 2021, 83, 110003. [Google Scholar] [CrossRef]
- Zhao, L.; Niu, J.; Feng, D.; Wang, X.; Zhang, R. Immune functions of pattern recognition receptors in Lepidoptera. Front. Immunol. 2023, 14, 1203061. [Google Scholar] [CrossRef]
- Michel, T. , Reichhart, J.M., Hoffmann, J.A.; Royet, J. Drosophila Toll is activated by Gram-positive bacteria through a circulating peptidoglycan recognition protein. Nature, 2001; 414, 756–759. [Google Scholar]
- Monahan, A.; Kleino, A.; Silverman, N. Readapting the role of PGRP-SD in bacterial sensing and Immune activation. Immunity 2016, 45, 951–953. [Google Scholar] [CrossRef]
- Iatsenko, I.; Kondo, S.; Mengin-Lecreulx, D.; Lemaitre, B. PGRP-SD, an extracellular pattern-recognition receptor, enhances peptidoglycan-mediated activation of the Drosophila Imd pathway. Immunity 2016, 45, 1013–1023. [Google Scholar] [CrossRef]
- Choe, K.M.; Werner, T.; Stöven, S.; Hultmark, D.; Anderson, K.V. Requirement for a peptidoglycan recognition protein (PGRP) in Relish activation and antibacterial immune responses in Drosophila. Science 2002, 296(5566), 359–362. [Google Scholar] [CrossRef]
- Gottar, M.; Gobert, V.; Michel, T.; Belvin, M.; Duyk, G.; Hoffmann, J.A.; Ferrandon, D.; Royet, J. The Drosophila immune response against Gram-negative bacteria is mediated by a peptidoglycan recognition protein. Nature 2002, 416(6881), 640–644. [Google Scholar] [CrossRef]
- Rämet, M.; Manfruelli, P.; Pearson, A.; Mathey-Prevot, B.; Ezekowitz, R.A. Functional genomic analysis of phagocytosis and identification of a Drosophila receptor for E. coli. Nature 2002, 416(6881), 644–648. [Google Scholar] [CrossRef]
- Takehana, A.; Katsuyama, T. ; Yano T, Oshima, Y.; Takada, H.; Aigaki, T.; Kurata, S. Overexpression of a pattern-recognition receptor, peptidoglycan-recognition protein-LE, activates imd/relish-mediated antibacterial defense and the prophenoloxidase cascade in Drosophila larvae. Proc. Natl Acad. Sci. USA, 2002; 99, 13705–13710. [Google Scholar]
- Takehana, A.; Yano, T.; Mita, S.; Kotani A, Oshima Y, Kurata S. Peptidoglycan recognition protein (PGRP)-LE and PGRP-LC act synergistically in Drosophila immunity. EMBO J. 2004, 23, 4690–4700. [Google Scholar] [CrossRef]
- Yano, T.; Mita, S.; Ohmori, H.; Oshima, Y.; Fujimoto, Y.; Ueda, R.; Takada, H.; Goldman, W.E.; Fukase, K.; Silverman, N.; Yoshimori, T.; Kurata, S. Autophagic control of listeria through intracellular innate immune recognition in Drosophila. Nat. Immunol. 2008, 9, 908–916. [Google Scholar] [CrossRef]
- Das, S.; Dong, Y.; Garver, L.; Dimopoulos, G. Specificity of the innate immune system: a closer look at the mosquito pattern-recognition receptor repertoire'. In Rolff J., Reynolds, S. (eds), Insect Infection and Immunity: Evolution, Ecology, and Mechanisms, 2009, 69-85 (Oxford University Press, 2009; online edn, Oxford Academic, 1 Sept. 2009, accessed 30 Dec. 2023. [CrossRef]
- Zhu, Y.; Yu, X.; Cheng, G. Insect C-type lectins in microbial infections. Adv. Exp. Med. Biol. 2020, 1204, 129–140. [Google Scholar]
- Dong, Y.; Manfredini, F.; Dimopoulos, G. Implication of the mosquito midgut microbiota in the defense against malaria parasites. PLoS Pathog. 2009, 5, e1000423. [Google Scholar] [CrossRef]
- Waterhouse, R.M.; Kriventseva, E.V.; Meister, S.; Xi, Z.; Alvarez, K.S.; Bartholomay, L.C.; Barillas-Mury, C.; Bian, G.; Blandin, S.; Christensen, B.M.; et al. Evolutionary dynamics of immune-related genes and pathways in disease-vector mosquitoes. Science 2007, 316(5832), 1738–1743. [Google Scholar] [CrossRef]
- Dong, Y.; Dimopoulos, G. Anopheles fibrinogen-related proteins provide expanded pattern recognition capacity against bacteria and malaria parasites. J. Biol. Chem. 2009, 284, 9835–9844. [Google Scholar] [CrossRef]
- Blandin, S.; Levashina, E.A. Thioester-containing proteins and insect immunity. Mol. Immunol. 2004, 40, 903–908. [Google Scholar] [CrossRef]
- Shokal, U.; Eleftherianos, I. Evolution and function of thioester-containing proteins and the complement system in the innate immune response. Front. Immunol. 2017, 8, 759. [Google Scholar] [CrossRef]
- Dostálová, A.; Rommelaere, S.; Poidevin, M.; Lemaitre, B. Thioester-containing proteins regulate the Toll pathway and play a role in Drosophila defence against microbial pathogens and parasitoid wasps. BMC Biol. 2017, 15, 79. [Google Scholar] [CrossRef]
- Shokal, U.; Eleftherianos, I. Thioester-containing protein-4 regulates the Drosophila immune signaling and function against the pathogen Photorhabdus. J. Innate Immun. 2017, 9, 83–93. [Google Scholar] [CrossRef]
- Levashina, E.A.; Moita, L.F.; Blandin, S.; Vriend, G.; Lagueux, M.; Kafatos, F.C. Conserved role of a complement-like protein in phagocytosis revealed by dsRNA knockout in cultured cells of the mosquito, Anopheles gambiae. Cell 2001, 104, 709–718. [Google Scholar] [CrossRef]
- Blandin, S.; Shiao, S.H.; Moita, L.F.; Janse, C.J.; Waters, A.P.; Kafatos, F.C.; Levashina, E.A. Complement-like protein TEP1 is a determinant of vectorial capacity in the malaria vector Anopheles gambiae. Cell 2004, 116, 661–670. [Google Scholar] [CrossRef]
- Yassine, H.; Kamareddine, L.; Osta, M.A. The mosquito melanization response Is implicated in defense against the entomopathogenic fungus Beauveria bassiana. PLoS Pathog. 2012, 8, e1003029. [Google Scholar] [CrossRef]
- Weng, S.C.; Li, H.H.; Li, J.C.; Liu, W.L.; Chen, C.H.; Shiao, S.H. A thioester-containing protein controls dengue virus infection in Aedes aegypti through modulating immune response. Front. Immunol. 2021, 12, 670122. [Google Scholar] [CrossRef]
- Imler, J.L.; Ferrandon, D.; Royet, J.; Reichhart, J.M.; Hetru, C.; Hoffmann, J.A. Toll-dependent and Toll-independent immune responses in Drosophila. J. Endotoxin Res. 2004, 10, 241–246. [Google Scholar] [CrossRef]
- Valanne, S.; Kallio, J.; Kleino, A.; Rämet, M. Large-scale RNAi screens add both clarity and complexity to Drosophila NF-κB signaling. Dev, Comp. Immunol, 2012; 37, 9–18. [Google Scholar]
- Parker, J.S.; Mizuguchi, K.; Gay, N.J. A family of proteins related to Spätzle, the toll receptor ligand, are encoded in the Drosophila genome. Proteins 2001, 45, 71–80. [Google Scholar] [CrossRef]
- Weber, A.N.; Tauszig-Delamasure, S.; Hoffmann, J.A.; Lelièvre, E.; Gascan, H.; Ray, K.P.; Morse, M.A.; Imler, J.L.; Gay, N.J. Binding of the Drosophila cytokine Spätzle to Toll is direct and establishes signaling. Nat. Immunol. 2003, 4, 794–800. [Google Scholar] [CrossRef]
- Sun, H.; Bristow, B.N.; Qu, G.; Wasserman, S.A. A heterotrimeric death domain complex in Toll signaling. Proc. Natl Acad. Sci. USA 2002, 99, 12871–12876. [Google Scholar] [CrossRef]
- Wu, L.P.; Anderson, K.V. Regulated nuclear import of Rel proteins in the Drosophila immune response. Nature 1998, 392(6671), 93–97. [Google Scholar] [CrossRef]
- Frolet, C.; Thoma, M.; Blandin, S.; Hoffmann, J.A.; Levashina, E.A. Boosting NF-kappaB-dependent basal immunity of Anopheles gambiae aborts development of Plasmodium berghei. Immunity 2006, 25, 677–685. [Google Scholar] [CrossRef]
- Georgel, P.; Naitza, S.; Kappler, C.; Ferrendon, D.; Zachary, D.; Swimmer, C.; Kopczynski, C.; Duyk, G.; Reichhart, J.-M.; Hoffmann, J.A.J.D.C. Drosophila immune deficiency (IMD) is a death domain protein that activates antibacterial defense and can promote apoptosis. Dev. Cell 2001, 1, 503–514. [Google Scholar] [CrossRef]
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef]
- Ertürk-Hasdemir, D.; Broemer, M.; Leulier, F.; Lane, W.S.; Paquette, N.; Hwang, D.; Kim, C.H.; Stöven, S.; Meier, P.; Silverman, N. Two roles for the Drosophila IKK complex in the activation of Relish and the induction of antimicrobial peptide genes. Proc. Natl Acad. Sci. USA. 2009, 106, 9779–9784. [Google Scholar] [CrossRef]
- Barletta, A.B.F.; Nascimento-Silva, M.C.L.; Talyuli, O.A.C.; Oliveira, J.H.; Pereira, L.O.; Oliveira, P.L.; Sorgine, M.H. Microbiota activates IMD pathway and limits Sindbis infection in Aedes aegypti. Parasites Vectors 2017, 10, 103. [Google Scholar] [CrossRef]
- Garver, L.S.; Bahia, A.C.; Das, S.; Souza-Neto, J.A. , Shiao, J.; Dong, Y.; Dimopoulos, G. Anopheles Imd pathway factors and effectors in infection intensity-dependent anti- Plasmodium action. PLoS Pathog, 2012; 8, e1002737. [Google Scholar]
- Telleria, E.L.; Sant'Anna, M.R.; Ortigão-Farias, J.R.; Pitaluga, A.N.; Dillon, V.M.; Bates, P.A.; Traub-Csekö, Y.M.; Dillon, R.J. Caspar-like gene depletion reduces Leishmania infection in sand fly host Lutzomyia longipalpis. J. Biol. Chem. 2012, 287, 12985–12993. [Google Scholar] [CrossRef]
- Vomáčková Kykalová, B.; Sassù, F.; Volf, P.; Telleria, E.L. RNAi-mediated gene silencing of Phlebotomus papatasi defensins favors Leishmania major infection. Front. Physiol. 2023, 14, 1182141. [Google Scholar] [CrossRef]
- Rennoll, S.A.; Rennoll-Bankert, K.E.; Guillotte, M.L.; Lehman, S.S.; Driscol, l T. P.; Beier-Sexton, M.; Rahman, M.S.; Gillespie, J.J.; Azad, A.F. The cat flea (Ctenocephalides felis) immune deficiency signaling pathway regulates Rickettsia typhi infection. Infect. Immun. 2017, 86, e00562–17. [Google Scholar]
- Rebeil, R.; Jarrett, C.O.; Driver, J.D.; Ernst, R.K.; Oyston, P.C.; Hinnebusch, B.J. Induction of the Yersinia pestis PhoP-PhoQ regulatory system in the flea and its role in producing a transmissible infection. J. Bacteriol. 2013, 195, 1920–1930. [Google Scholar] [CrossRef]
- Hu, C.; Aksoy, S. Innate immune responses regulate trypanosome parasite infection of the tsetse fly Glossina morsitans morsitans. Mol. Microbiol. 2006, 60, 1194–1204. [Google Scholar] [CrossRef]
- Batista, K.K.S.; Vieira, C.S.; Figueiredo, M.B.; Costa-Latgé, S.G.; Azambuja, P.; Genta, F.A.; Castro, D.P. Influence of Serratia marcescens and Rhodococcus rhodnii on the humoral immunity of Rhodnius prolixus. Int. J. Mol. Sci. 2021, 22, 10901. [Google Scholar] [CrossRef]
- Vieira, C.S.; Moreira, O.C.; Batista, K.K.S.; Ratcliffe, N.A.; Castro, D.P.; Azambuja, P. The NF-κB inhibitor, IMD-0354, affects immune gene expression, bacterial microbiota and Trypanosoma cruzi infection in Rhodnius prolixus midgut. Front. Physiol. 2018, 9, 1189. [Google Scholar] [CrossRef]
- Alves-Bezerra, M.; Cosentino-Gomes, D.; Vieira, L.P.; Rocco-Machado, N.; Gondim, K.C.; Meyer-Fernandes, J.R. Identification of uncoupling protein 4 from the blood-sucking insect Rhodnius prolixus and its possible role on protection against oxidative stress. Insect Biochem. Mol. Biol. 2014, 50, 24–33. [Google Scholar] [CrossRef]
- Yoon, K.A.; Lee, D.E.; Lee, S.H.; Kim, J.H. Exploring the potential role of defensins in differential vector competence of body and head lice for Bartonella quintana. Parasit. Vectors 2023, 16, 183. [Google Scholar] [CrossRef] [PubMed]
- Nishide, Y.; Kageyama, D.; Yokoi, K.; Jouraku, A.; Tanaka, H.; Futahashi, R.; Fukatsu, T. Functional crosstalk across IMD and Toll pathways: insight into the evolution of incomplete immune cascades. Proc. R. Soc. B. 2019, 286, 20182207. [Google Scholar] [CrossRef]
- Gulia-Nuss, M. , Nuss, A., Meyer, J.; Sonenshine, D.E.; Roe, R.M.; Waterhouse, R.M.; Sattelle, D.B.; de la Fuente, J.; Ribeiro, J.M.; Megy, K.; et al. Genomic insights into the Ixodes scapularis tick vector of Lyme disease. Nat. Commun. 2016, 7, 10507. [Google Scholar] [CrossRef]
- Sidak-Lofti, L.C.; Rosche, K.L.; Pence, N.; Ujczo, J.K.; Hurtado, J.; Fisk, E.A.; Goodman, A.G.; Noh, S.M.; Peters, J.W.; Shaw, D.K. The unfolded protein response triggers the immune deficiency pathway in ticks. mBio. 2022, 13(6), e0292222. [Google Scholar]
- Cabral, S.; de Paula, A.; Samuels, R.; da Fonseca, R.; Gomes, S.; Silva, J.R.; Mury, F. Aedes aegypti (Diptera: Culicidae) immune responses with different feeding regimes following infection by the entomopathogenic fungus Metarhizium anisopliae. Insects 2020, 11, 95. [Google Scholar] [CrossRef]
- Millymaki, H.; Ramet, M. JAK/STAT pathway in Drosophila immunity. Scand. J. Immunol. 2014, 79, 377–385. [Google Scholar] [CrossRef]
- Brown, S. , Hu, N., & Hombria, J.C. Novel level of signalling control in the JAK/STAT pathway revealed by in situ visualisation of protein-protein interaction during Drosophila development. Development, 2003; 130, 3077–3084. [Google Scholar]
- Bina, S.; Zeidler, M. JAK/STAT Pathway signalling in Drosophila melanogaster. Landes Bioscience 2009. www.ncbi.nlm.nih.gov › books › NBK6034.
- Zeidler, M. , Bach, E.; Perrimon, N. The roles of the Drosophila JAK/STAT pathway. Oncogene, 2000; 19, 2598–2606. [Google Scholar]
- Gupta, L.; Molina-Cruz, A.; Kumar, S.; Rodrigues, J.; Dixit, R.; Zamora, R.E.; Barillas-Mury, C. The STAT pathway mediates late-phase immunity against Plasmodium in the mosquito Anopheles gambiae. Cell Host Microbe 2009, 5, 498–507. [Google Scholar] [CrossRef]
- Bahia, A.C.; Kubota, M.S.; Tempone, A.J.; Araujo, H.R.C.; Guedes, B.A.M.; Orfano, A.S.; Tadei, W.P.; Rıos-Velasquez, C.M.; Han, Y.S.; Nagila, F.C.; et al. The JAK-STAT pathway controls Plasmodium vivax load in early stages of Anopheles aquasalis infection. PLoS Negl. Trop. Dis. 2011, 5, e1317. [Google Scholar] [CrossRef]
- Souza-Neto, J.A.; Sim, S.; Dimopoulos, G. An evolutionary conserved function of the JAK-STAT pathway in anti-dengue defense. Proc. Natl Acad. Sci. USA. 2009, 106, 17841–17846. [Google Scholar] [CrossRef]
- Telleria, E.L.; Azevedo-Brito, D.A.; Kykalová, B.; Tinoco-Nunes, B.; Pitaluga, A.N.; Volf, P. Traub-Csekö, Y.M. Leishmania infantum infection modulates the JAK-STAT pathway in Lutzomyia longipalpis LL5 embryonic cells and adult females, and affects parasite growth in the sand fly. Front. Trop. Dis, 2021. [Google Scholar] [CrossRef]
- Weston, C.R.; Davis, R.J. The JNK signal transduction pathway. Curr. Opin. Genet. Dev. 2002, 12, 14–21. [Google Scholar] [CrossRef]
- Tafesh-Edwards, G. , Eleftherianos, I. JNK signaling in Drosophila immunity and homeostasis. Immunology Letters 2020, 226, 7–11. [Google Scholar] [CrossRef]
- Garver, L.S.; de Almeida, O.G.; Barillas-Mury, C. The JNK pathway is a key mediator of Anopheles gambiae antiplasmodial immunity. PLoS Pathog. 2013, 9, e1003622. [Google Scholar] [CrossRef]
- Chowdhury, A.; Modahl, C.M.; Tan, S.T.; Wong, W.X.B; Missé, D.; Vial, T.; Kini, R.M.; Pompon, J.F. JNK pathway restricts DENV2, ZIKV and CHIKV infection by activating complement and apoptosis in mosquito salivary glands. PLoS Pathog. 2020, 16, e1008754. [Google Scholar] [CrossRef]
- Yajima, M.; Takada, M.; Takahashi, N.; Kikuchi, H.; Natori, S.; Oshima, Y.; Kurata, S. A newly established in vitro culture using transgenic Drosophila reveals functional coupling between the phospholipase A2-generated fatty acid cascade and lipopolysaccharide-dependent activation of the immune deficiency (imd) pathway in insect immunity. Biochem. J. 2003, 371 (Pt 1), 205–210. [Google Scholar] [CrossRef]
- Kwon, H.; Hall, D.R.; Smith, R.C. Prostaglandin E2 signaling mediates oenocytoid immune cell function and lysis, limiting bacteria and Plasmodium oocyst survival in Anopheles gambiae. Front. Immunol. 2021, 12, 680020. [Google Scholar] [CrossRef]
- Barletta, A.B.F.; Alves, E.S.T.L.; Talyuli, O.A.C.; Luna-Gomes, T.0; Sim, S.; Angleró-Rodríguez, Y.; Dimopoulos, G.; Bandeira-Melo, C.; Sorgine, C.M.H.F. Prostaglandins regulate humoral immune responses in Aedes aegypti. PLoS Negl.Trop Dis. 2020, 14, e0008706. [Google Scholar] [CrossRef]
- Figueiredo, M.B.; Genta, F.A.; Garcia, E.S.; Azambuja, P. Lipid mediators and vector infection: Trypanosoma rangeli inhibits Rhodnius prolixus hemocyte phagocytosis by modulation of phospholipase A2 and PAF-acetylhydrolase activities. J. Insect Physiol. 2008, 54, 1528–1537. [Google Scholar] [CrossRef]
- Mussabekova, A.; Daeffler, L.; Imler, J.L. Innate and intrinsic antiviral immunity in Drosophila. Cell. Mol. Life Sci. 2017, 74, 2039–2054. [Google Scholar] [CrossRef]
- Avadhanula, V.; Weasner, B.P.; Hardy, G.G.; Kumar, J.P.; Hardy, R.W. A novel system for the launch of Alphavirus RNA synthesis reveals a role for the Imd pathway in arthropod antiviral response. PLoS Pathog. 2009, 5, e1000582. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Zhou, G.; Wu, J. ;Xi, Z. Wolbachia induces reactive oxygen species (ROS)-dependent activation of the Toll pathway to control dengue virus in the mosquito Aedes aegypti. Proc. Natl Acad. Sci. USA, 2012; 109, E23-31. [Google Scholar]
- Behura, S.K.; Gomez-Machorro, C.; Harker, B.W.; DeBruyn, B.; Lovin, D.D.; Hemme, R.R.; Mori, A.; Romero-Severson, J.; Severson, D.W. Global cross-talk of genes of the mosquito Aedes aegypti in response to dengue virus infection. PLoS Negl. Trop. Dis. 2011, 5, e1385. [Google Scholar] [CrossRef] [PubMed]
- Angleró-Rodríguez, Y.I.; MacLeod, H.J.; Kang, S.; Carlson, J.S.; Jupatanakul, N.; Dimopoulos, G. Aedes aegypti molecular responses to Zika Virus: modulation of Infection by the Toll and Jak/Stat immune pathways and virus host factors. Front. Microbiol. 2017, 8, 2050. [Google Scholar] [CrossRef]
- Dong, S.; Dimopoulos, G. Aedes aegypti Argonaute 2 controls arbovirus infection and host mortality. Nat. Commun. 2023, 14, 5773. [Google Scholar] [CrossRef]
- Goto, A.; Okado, K.; Martins, N.; Cai, H.; Barbier, V.; Lamiable, O.; Troxler, L.; Santiago, E. ; Kuhn L, Paik, D.; et al. The kinase IKKβ regulates a STING- and NF-κB-dependent antiviral response pathway in Drosophila. Immunity, 2018; 49, 225-234, e4. [Google Scholar]
- Hartman, R.S.; Karp, R.D. Short-term immunologic memory in the allograft response of the American cockroach, Periplaneta americana. Transplantation 1989, 47, 920–922. [Google Scholar] [CrossRef]
- Faulhaber, L.M.; Karp, R.D. A diphasic immune response against bacteria in the American cockroach. Immunology 1992, 75, 378–381. [Google Scholar]
- Lanz-Mendoza, H. , Garduño, J.C. Insect Innate Immune Memory. In: Cooper, E. (eds), 2018, Advances in Comparative Immunology. Springer, Cham. [CrossRef]
- Sheehan, G.; Farrell, G.; Kavanagh, K. Immune priming: the secret weapon of the insect world. Virulence 2020, 11, 238–246. [Google Scholar] [CrossRef]
- 243. Vilcinskas, A. Mechanisms of transgenerational immune priming in insects. Dev. Comp. Immunol. 2021, 124, 104205. [Google Scholar] [CrossRef]
- Prakash, A.; Khan, I. Why do insects evolve immune priming? A search for crossroads. Dev. Comp. Immunol. 2022, 126, 104246. [Google Scholar] [CrossRef]
- Gomes, F.M.; Silva, M.; Molina-Cruz, A.; Barillas-Mury, C. Molecular mechanisms of insect immune memory and pathogen transmission. PLoS Pathog. 2022, 18, e1010939. [Google Scholar] [CrossRef]
- Vargas, V.; Cime-Castillo, J.; Lanz-Mendoza, H. Immune priming with inactive dengue virus during the larval stage of Aedes aegypti protects against the infection in adult mosquitoes. Sci Rep. 2020, 10, 6723. [Google Scholar] [CrossRef]
- Cabrera, K.; Hoard, D.S.; Gibson, O.; Martinez, D.I.; Wunderlich, Z. Drosophila immune priming to Enterococcus faecalis relies on immune tolerance rather than resistance. PLOS Pathogens 2023, 19, e1011567. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



