Submitted:
25 January 2024
Posted:
26 January 2024
You are already at the latest version
Abstract
Clostridioides (Clostridium) difficile is an enteric pathogen of neonatal pigs with zoonotic transmission remaining a significant problem in humans, including nosocomial resurgence, following oral administration of broad-spectrum antibiotics. To date, the immune response to C. difficile has mostly focused on neutrophils and cytokine/chemokines, particularly in human infection.
We show that porcine monocytes respond to C. difficile differently compared with many other bacterial infections. Infection of porcine monocytes with human C. difficile strains CD630 (Ribotype 078) or R20291 (Ribotype 027) for 3 or 24h post-infection (pi) resulted in a lack of oxidative burst or nitrite ion production when compared to uninfected controls (P >0.05). The survival dynamics of both CD630 and R20291 in monocytes were similar with intracellular bacterial numbers being similar at 3h pi and 24h pi (P >0.05). However, we show that porcine monocytes entrap C. difficile via extracellular DNA traps. This process began as early as 3h pi and at 24h pi the nuclei appeared to be depleted of DNA, although extracellular DNA was associated with the cell membrane. Our preliminary study also suggests that entrapment of C. difficile by extracellular DNA may occur via a process of monocyte etosis.
Keywords:
Clostridiodes (Clostridium) difficile
; Monocyte
; DNA traps
; Etosis
; Pigs
1.0. Introduction
Clostridioides difficile is a Gram-positive, spore-forming, anaerobic enteropathogen of pigs, humans and other animals [1,2]. Pathology associated with C. difficile results from the production of Rho-glucosylating toxins A and B (TcdA and TcdB) which induce a significant inflammatory immune response and subsequent cellular pathology. However, in humans and pigs, C. difficile is also found in clinically normal individuals and development of clinical disease occurs usually as a result of dysbacteriosis, often following antibiotic intervention for other reasons.
Much of the focus on the immune response to C. difficile has centered on the role of neutrophils and inflammatory mediators such as cytokines, [3] particularly in the development of classical volcano lesions in colonic tissue [4]. However, monocytes play a key role during intestinal inflammation. In a dextran sulphate-induced murine model of colitis, F4/80+CD11b+CCR2+Ly6Chigh inflammatory monocytes were recruited to the colon and F4/80+CD11b+CCR2+Ly6Chigh colonic monocyte/macrophages correlated with colonic eosinophilic inflammation [5]. In experimental murine C. difficile infections, innate cells including monocytes are recruited to the large intestine [6,7] in a CCR2-dependent manner [8]. This indicates that monocyte chemoattractant protein 1 (CCL2) and its receptor are essential in this process, although CCR deficiency does not alter the course of infection [8].
Although relatively little is known about the immunological function of monocytes in the colonic epithelium during C. difficile infection, a study by McDermott et al. [9] reported that, in mice, CCR2 and IL-23 are required for monocyte recruitment and this increases inducible nitric oxide synthase (INOS) expression in colonic tissues. INOS is a precursor required for reactive nitrogen species (RNS) such as NO2- which is a known effector in many different types of bacterial infection. However, toxin A from C. difficile strain 10463 has been shown to inhibit reactive oxygen species (ROS) production by TNF-α-stimulated human neutrophils [10] and ROS is another common killing pathway utilized by innate immune cells during bacterial infection. C. difficile in pigs is now recognized as an important source of human infection [11] and could spread antimicrobial resistance into the human food chain. However, nothing has been reported regarding the response of porcine monocytes to human C. difficile strains.
The aim of this study was to describe the behavior porcine monocytes in response to C. difficile CD630 (Ribotype 078) or R20291 (Ribotype 027) infection, both of which have been shown to be pathogenic in pigs and humans [12].
2.0. Materials and Methods
2.1. C. difficile Strains and Culture
Two strains of C. difficile were used, CD630 and R20291, received from Professor Nigel Minton (School of Molecular Medical Science, The University of Nottingham). CD630 was initially sourced from a patient in Switzerland suffering from severe pseudomembranous colitis in 1982. R20291 (a hypervirulent strain) was sourced from an outbreak in 2006 at Stoke Mandeville hospital, UK [13].
Strains were stored as spore preparations at 4oC which were cultured on C. difficile Agar Base (Oxoid Ltd, Hampshire UK), supplemented with cholic acid sodium salt (Acros Organics, Antwerp, Belgium) 0.1% (w/v) to aid germination, which was autoclaved and cooled to ~40˚C before 7% (v/v) pre-warmed defibrinated horse blood was added (Oxoid). The strains were incubated anaerobically at 37.4˚C for 2-3 days in a Bactron™ Environmental Chamber (Bactron II-2, Sheldon Manufacturing Inc., USA). Both strains were passaged twice-weekly for preparation of liquid cultures.
Liquid cultures for experimental use were prepared using anaerobe basal broth (Oxoid) which was taken immediately from the autoclave, with pressure released manually, to the anaerobic chamber, to prevent oxygen infusing back into the liquid. Broths were held in the port for 5 days, with loose lids, before inoculation from the passaged strains. Broths were incubated at 37˚C for 2 days prior to use.
Bacterial cell/spore concentration was determined by enumeration of colony-forming units (CFU/ml) [14]. Serial dilutions were plated out on unsupplemented C. difficile agar, cholic acid and defibrinated horse blood (CCH) plates in the environmental chamber. These were incubated for 24 hours and then counted.
2.2. Porcine Monocyte Isolation and Culture
Whole blood with lithium heparin was purchased from Matrix Biologicals Ltd. (Hull, UK) obtained from 8 clinically healthy, 5-6 months old Large White female pigs at slaughter. Standard density-gradient centrifugation of whole blood from each sample was performed using Histopaque (Sigma, Dorset UK) to obtain buffy coats. The resulting pellet was resuspended in PBS for cell enumeration, using a hemocytometer and Trypan Blue Solution 0.4% (Sigma, UK) to analyze cell viability. This showed that >99% cells were viable in all the buffy coat samples analyzed. 1x106 buffy coat cells were seeded into 500µl of RPMI 1640 (Gibco, UK), L-Glutamine (1% w/v, Gibco) combined with fetal calf serum (5% v/v) and penicillin and streptomycin, per well of a 24 well cell culture dish (Nunc, Naperville, Illinois, USA). Cells were cultured for 2h at 370C and 5% CO2, after which supernatants were removed and attached cells (monocytes) were then detached gently and cultured in fresh RPMI media at 1x105 monocytes per 500 μl for 24h prior to use.
2.3. Fluorescence-Activated Cell Sorting (FACS)
FACS was performed in order to determine CD14 expression (monocytes) within the buffy coat. Buffy coats were washed in PBS, centrifuged for 5 min, and resuspended in 100µl PBS to give 1x106 cells. 10µl (10µg/ml) of mouse anti-porcine CD14 antibody conjugated to fluorescein isothiocyanate (FITC) (AbD Serotec, Oxford, United Kingdom) was added to the sample and incubated in a dark room at 4oC for 30 min. Mouse IgG2β FITC (AbDSerotec, UK) was used as an isotype control. After 30 min, samples were washed in 5ml PBS and centrifuged for 5 min prior to being resuspended in 500µl of PBS for CD14 expression analysis by FACS (FACS Canto II™ Flow Cytometer, Becton Dickinson and Company, USA).
2.4. Monocyte Infection Studies
1x106 cells of each C. difficile strain was added to monocyte cultures to achieve a multiplicity of infection (MOI) of 10 and incubated for 2h; negative control cells were incubated with media only. After 2 h the wells were washed 3 x in PBS warmed at 37oC? and the monolayer was inspected using an upright microscope to assess cell integrity. Cells were then incubated with vancomycin for 60 min to kill extracellular bacteria as done previously with Salmonella and gentamicin [15]. Cells were incubated for either 3 or 24h post-infection (pi) prior to washing a further 3 x in PBS before all the liquid was removed from the cells. The plates were then transferred to the anaerobic environmental cabinet and lysed with 0.5% Triton-X-100 for 30 min. Serial dilutions were carried out plating on CCH plates. The test was done in triplicate for each of the 8 pigs.
2.5. Measurement of ROS in Monocytes
A standard nitroblue tetrazolium (NBT) reduction assay [16] was used to measure ROS activity in porcine monocytes at 3 and 24h pi. Briefly, infected and control monocytes were incubated for 45 min with 50 μl NBT (Sigma) (10 mg/ml NBT in PBS) at 37°C in 5% (vol/vol) CO2. The cells were then washed in PBS and incubated in 100 μl 1M hydrochloric acid for 10 min to stop the reaction. The cells were then washed in PBS, and 150 μl dimethyl sulfoxide (Sigma) was added and mixed thoroughly prior to addition of 10 μl 5M sodium hydroxide to develop the color. The optical density of the reaction mixture was determined at 620 nm with a plate reader (Anthos Labtech Instruments, Hamburg, Germany). As a positive control, monocytes were incubated with 1 μg/ml zymosan (Sigma, Poole, UK) for the same times.
2.6. Measurement of RNS in Monocytes
At 3h and 24h pi, supernatants from infected and uninfected monocytes were removed and stored at -80˚C prior to use. Greiss reagent (Promega, USA) was used to detect nitrite ions in the supernatants according to the manufacturer’s instructions measuring absorbance at 540 nm. As a positive control, monocytes were incubated with 10 μg/ml phorbol myristate acetate (PMA) (Sigma, Poole, United Kingdom). with the manufacturer’s guidelines and absorbance measured at an optical density of 540 nm.
2.7. Fluorescence Microscopy
Autoclaved, circular, glass coverslips were placed on the bottom of 24-well plates. Monocytes were seeded onto the glass as described in section 2.2, and C. difficile infection was carried out as described in section 2.3. At 3h and 24h pi the coverslips were removed from the wells and fixed in acetone for 15 min. The coverslips were then washed 3 x in PBS. Phalloidin (10mg/ml) was used to stain actin within the cells for 15 min. The coverslips were placed on microscope slides and mounted with ProLong Gold anti-fade reagent with DAPI (Invitrogen, UK) before covering with a square coverslip. All slides were examined using a TCS SP2 confocal microscope (Leica Microsystems, Heidelberg, Germany).
The C. difficile strains were also examined by fluorescence microscopy. A loop of each C. difficile broth culture was diluted 1:10 and smeared onto a microscope slide. The preparations were stained with phalloidin as detailed above and mounted on a slide using DPX-mountant (Sigma, UK) with a cover slip prior to examination using a TCS SP2 confocal microscope (Leica, Germany). This approach has been used regularly by this group [17,18].
2.8. Statistical Analysis
Statistical analysis was performed by Student’s t-test using GraphPad Prism version 5.00 for windows (GraphPad Software, San Diego, California, USA). Statistical significance was assessed at the 5% confidence limit (P <0.05)
3.0. Results
3.1. Phenotype and Morphology of Uninfected (Steady-State) Porcine Monocytes
Initial FACS experiments were performed to determine the morphology of the monocytes prior to infection with C. difficile strains CD630 or R20291. The cells were large (SSc high) and expressed CD14 (Figure 1A) and CD172a (not shown). The high SSc/CD14 staining indicated that the cells cultured were monocytes. The nucleus of the cells were typically C-shaped with densely packed DNA, shown by DAPI staining (Figure 1B).
3.2. Staining of C. difficile Vegetative Cells
Phalloidin staining of bacterial cells showed a clearly discernable, but mixed morphology consisting primarily of medium to long bacilli, which was observed consistently in preparations of both C. difficile CD630 and R20291 strains (Figure 1C). In contrast, phalloidin staining of the storage spore preparation was not at all clear which is likely consequence of the spores not having taken up the stain at all.
3.3. Survival Dynamics of C. difficile in Porcine Monocytes
Survival of both strains in porcine macrophages was good with very little change in the bacterial numbers recovered between 3h and 24h albeit with a very small non-significant increase at 24h. (Figure 2A).
3.4. Reactive Oxygen and Nitrogen Species Produced by Porcine Monocytes in Response to C. difficile Infection
Oxidative burst tends to be more acute than the production of nitric oxide. The product of the NBT reaction indicated that levels of reactive oxygen species produced were very similar to those of the uninfected monocytes whereas significantly more was produced by the positive zymosan control (Figure 2B).
Similar results were found for reactive nitrogen species with values increasing at 24h but no difference between the levels produced by the two strains compared with the uninfected negative control and the significantly greater amount produced by the PMA positive control (Figure 2C).
3.5. Porcine Monocyte form DNA Traps to Immobilize C. difficile
DAPI staining of macrophages 3h after infection with C. difficile CD630 showed a mass of DNA surrounding and connecting groups of monocytes (Figure 3A, solid arrows). The outline of the nuclei of the cells were less distinct than observed in uninfected cells and the cell morphology was also less clear (Figure 3B, broken arrows) when compared with Figure 1B. It was possible in the overlay to observe structures which may have been individual bacterial cells (Figure 3C, solid arrows). None was seen in uninfected cells (data not shown). Similar images were observed for R20291 (data not shown).
When individual monocytes were observed after 3h it was possible to discern DNA fibers protruding through pores or discrete areas of lysis in the monocyte cell membrane (Figure 4) indicating early-stage etosis. The outline of the c-shaped nucleus remained clear at this stage.
3.6. Following 24 h Culture of Porcine Monocytes with C. difficile, Complete Etosis Occurred but Extracellular DNA Was Still Able to Entrap CD630 or R20291
After 24h infection the nuclei were much less clear and, in some cases, not clearly discernible at all. Most of the DNA appeared to be extra-cellular (Figure 5A) with the macrophage cell outline very uneven (Figure 5B, E). The pattern of etosis induced by the two strains appeared to be different with more filamentous release of DNA by CD630 (Figure 5a, C) and more explosive release by R20291 (Figure 5D and F).
Figure 6 shows that individual bacterial cells (solid arrows) not directly in contact with a macrophage appeared nevertheless to be entangled by DNA possibly released by the adjacent monocyte although, in the case of the CD630 (Figure 6C) compared with R20291 (Figure 6F) bacterial cells, this was not completely clear. Uninfected monocytes showed no adverse effects at all at either 3 or 12h (data not shown).
4.0. Discussion
We report for the first time that porcine monocytes, exposed to the clinical C. difficile strains CD630 and R20291 [12], do not respond by the induction of common (ROS or RNS) innate cell killing pathways but are able to immobilize C. difficile via DNA traps. Neutrophil extracellular traps (NETS) were first shown to kill Gram-positive and Gram-negative bacteria in a study by Brinkmann et al. [19] and it has remained a contentious issue whether entrapment of bacteria occurs as a consequence of a cell death pathway elicited by bacteria and/or toxins (termed Netosis) or is a neutrophil-driven and evolutionary defense mechanism [20]. Since it is now recognized that other human myeloid immune cells, including macrophages [2] and plasmacytoid dendritic cells [22] can also undergo this process, it is now termed etosis. The proposed significance of etosis for myeloid cells remains unclear and its potential involvement in innate immunity has also been discussed [21]. In this case it remains unclear whether bacterial killing might also be taking place or whether the bacteria are simply immobilized facilitating further phagocytosis by neutrophils.
Our study indicates that the process of monocyte etosis with C. difficile CD630 or R20291 occurs as early as 3h post-infection, as indicated by nuclear DNA protruding through pores in the monocyte membrane [23]. At this time point, monocyte DNA surrounded the bacteria and although large numbers were entrapped between groups of monocytes, nuclear DNA remained largely intact and the characteristic ‘C’ shaped morphology was evident.
At 24h post-infection we show that most nuclear DNA was extracellular and although it was still associated with the cell membrane it appeared to be effectively lost from the cell nucleus. This occurred in monocytes which were not in contact with C. difficile and may thus be a result of toxin release by C. difficile. This would indicate a progressive etosis of porcine monocytes from around 3h to 24h pi when infected with either C. difficile CD630 or R20291. However, even by 24h post-infection extracellular monocyte DNA was still able to entrap C. difficile and our data, therefore, probably suggests that the process of etosis is pathogen-driven but a consequence of that is entrapment of some of the bacterial population. This was supported by the fact that there was no detectable decline in bacterial viability during this period.
There was no detectable ROS or RNS response by these monocytes at either 3 or 24h pi. The formation and effect of antibacterial free radicals and molecules such as H2O2 and NO have been studied for many years [24] and more recently their potential therapeutic use as antimicrobials has been explored [25,26]. However, some bacterial species are known to inhibit production of RNS and ROS by innate immune cells and α-toxin-induced inhibition of ROS has been reported in human neutrophils infected with C. difficile strain 10463 [10]. A correlation between human monocyte etosis and inhibition of ROS has also been reported by Webster et al., [27]. In that particular study it was shown that, when human monocytes were infected with either Escherichia coli or Klebsiella pneumoniae, etosis occurred with detectable extracellular DNA, but there was no detectable ROS and phagocytosis was inhibited at this point. In contrast, when human monocytes were infected with Neisseria meningitidis, etosis did not occur, there was a detectable ROS response and the monocytes showed prolonged phagocytic activity [28]. Results from our study would therefore suggest that porcine monocytes infected with human/pig C. difficile strains CD630 and R20291 undergo a similar process to human monocytes infected with E. coli or K. pneumoniae. The standard invasion assay used here was developed for pathogenic members of the Enterobacteriaceae, particularly, Salmonella and related organisms, for which an intra-cellular phase involving macrophages is a key stage in the disease process. This is not necessarily the case with C. difficile although it is certainly pertinent to the initiation of the immune response to infection. The two-hour exposure stage allows invasion to take place but exposes the monocytes to extra-cellular toxins produced by the initially extra-cellular bacteria. The bacterial cells observed at 24h entrapped by the DNA are extracellular and it is impossible to say at this juncture whether these were bacteria which had been released from dying monocytes or whether they were extra-cellular from the outset surviving the vancomycin treatment used to kill initially extra-cellular bacteria.
In conclusion, we show that when porcine monocytes are infected with pathogenic C. difficile strains, they form extracellular DNA traps which immobilize bacteria and that this occurs via a process of etosis, resulting in complete loss of nuclear DNA and monocyte death. We cannot say what the overall immunological effect of this response has on C. difficile infection in pigs or humans. However, studies in mice have previously reported that monocyte recruitment is not required for clearance of C. difficile but that this may increase the inflammatory response and clinical signs of disease [29]. It is, therefore, possible that monocyte etosis, which we report, may be important in this regard. Our study also shows a hitherto unknown fate of porcine monocytes infected with zoonotic C. difficile.
References
- Songer, J.G. The emergence of Clostridium difficile as a pathogen of food animals. Anim Health Res Rev 2004, 5, 321–326. [Google Scholar] [CrossRef]
- Squire, M.M.; Riley, T.V. Clostridium difficile infection in humans and piglets: a 'One Health' opportunity. Curr Top Microbiol Immunol 2013, 365, 299–314. [Google Scholar]
- Cowardin, C.A.; Petri, W.A., Jr. Host recognition of Clostridium difficile and the innate immune response. Anaerobe 2014, 30, 205–9. [Google Scholar] [CrossRef]
- Songer, J.G.; Uzal, F.A. Clostridial enteric infections in pigs. J Vet Diagn Invest 2005, 17, 528–36. [Google Scholar] [CrossRef]
- Waddell, A.; Ahrens, R.; Steinbrecher, K.; et al. Colonic eosinophilic inflammation in experimental colitis is mediated by Ly6C(high) CCR2(+) inflammatory monocyte/macrophage-derived CCL11. J Immunol 2011, 186, 5993–6003. [Google Scholar] [CrossRef]
- Chen, X.; Katchar, K.; Goldsmith, J.D.; Nanthakumar, N.; Cheknis, A.; Gerding, D.N.; Kelly, C.P. A mouse model of Clostridium difficile-associated disease. Gastroenterology 2008, 135, 1984–1992. [Google Scholar] [CrossRef] [PubMed]
- Sadighi Akha, A.A.; Theriot, C.M.; Erb-Downward, J.R.; McDermott, A.J.; Falkowski, N.R.; Tyra, H.M.; Rutkowski, D.T.; Young, V.B.; Huffnagle, G.B. Acute infection of mice with Clostridium difficile leads to eIF2α phosphorylation and pro-survival signalling as part of the mucosal inflammatory response. Immunology 2013, 140, 111–22. [Google Scholar] [CrossRef] [PubMed]
- Jarchum, I.; Liu, M.; Shi, C.; Equinda, M.; Pamer, E.G. Critical role for MyD88-mediated neutrophil recruit McDermott, A.J.; Falkowski, N.R.; McDonald, R.A.; et al. Role of interferon-γ and inflammatory monocytes in driving colonic inflammation during acute Clostridium difficile infection in mice. Immunology 2017, 150, 468–477. [Google Scholar]
- McDermott, A.J.; Falkowski, N.R.; McDonald, R.A.; et al. Role of interferon-γ and inflammatory monocytes in driving colonic inflammation during acute Clostridium difficile infection in mice. Immunology 2017, 150, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Brito, G.A.C.; Sullivan, G.W.; Cielsa, W.P.; Carper, H.T.; Mandell, G.L.; Guerrant, R.L. Clostridium difficile Toxin A alters in vitro-adherent neutrophil morphology and function. J Infect Dis 2002, 185, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Keessen, E.C.; Harmanus, C.; Dohmen, W.; Kuijper, E.J.; Lipman, L.J. Clostridium difficile infection associated with pig farms. Emerg Infect Dis 2013, 19, 1032–1034. [Google Scholar] [CrossRef]
- Goorhuis, A.; Bakker, D.; Corver, J.; Debast, S.B.; Harmanus, C.; Notermans, D.W.; Bergwerff, A.A.; Dekker, F.W.; Kuijper, E.J. Emergence of Clostridium difficile infection due to a new hypervirulent strain, polymerase chain reaction ribotype 078. Clin Infect Dis 2008, 47, 1162–1170. [Google Scholar] [CrossRef] [PubMed]
- Dawson, L.F.; Donahue, E.H.; Cartman, S.T.; Barton, R.H.; Bundy, J.; McNerney, R.; Minotn, N.P.; Wren, B.W. The analysis of para-cresol production and tolerance in Clostridium difficile 027 and 012 strains. BMC Microbiol 2011, 11, 86. [Google Scholar] [CrossRef] [PubMed]
- Burns, D.A.; Minton, N.P. Sporulation studies in Clostridium difficile. J Microbiol Methods 2011, 87, 133–138. [Google Scholar] [CrossRef]
- Ibrahim, H.; Askar, B.; Hulme, S.; Neilson, P.; Barrow, P.; Foster, N. Differential Immune Phenotypes in Human Monocytes Induced by Non-Host-Adapted Salmonella enterica Serovar Choleraesuis and Host-Adapted S. Typhimurium. Infect Immun 2018, 86, e00509–518. [Google Scholar] [CrossRef] [PubMed]
- Park, B.H.; Fikrig, S.M.; Smithwick, E.M. Infection and nitroblue-tetrazolium reduction by neutrophils. A diagnostic acid. Lancet ii 1968, 2, 532–534. [Google Scholar] [CrossRef] [PubMed]
- Mutamba, S.; Allison, A.; Barrow, P.A.; Mahida, Y.; Foster, N. Expression of IL-1Rrp2 by human myelomonocytic cells is unique to DCs and facilitates DC maturation by IL-1F8 and IL-1F9. European J Immunol 2012, 42, 607–17. [Google Scholar] [CrossRef] [PubMed]
- Penha, R.; Higgins, J.; Mutamba, S.; Barrow, P.; Mahida, Y.; Foster, N. IL-36 receptor is expressed by human blood and intestinal T lymphocytes and is dose-dependently activated via IL-36β and induces CD4+ lymphocyte proliferation. Cytokine. 2016, 85, 18–25. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Yipp, B.G.; Kubes, P. NETosis: how vital is it? Blood 2013, 122, 2784–2794. [Google Scholar] [CrossRef]
- Oboe, D.M.; Curtis, B.J.; Chen, M.M.; Ippolito, J.A.; Kovacs, E.J. Extracellular traps and macrophages: new roles for the versatile phagocyte. J Leukoc Biol 2015, 97, 1023–1035. [Google Scholar] [CrossRef]
- Loures, F.V.; Rohm, M.; Lee, C.K.; Santos, E.; Wang, J.P.; Specht, C.A.; Calich, V.L.G.; Urban, C.F.; Levitz, C.F. Recognition of Aspergillus fumigatus hyphae by human plasmacytoid dendritic cells is mediated by dectin-2 and results in formation of extracellular traps. PLoS Pathog 2015, 11, e1004643. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, B.E.; Grinstein, S. Unconventional roles of the NADPH oxidase: signaling, ion homeostasis, and cell death. Sci STKE 2007, 379, pe11. [Google Scholar] [CrossRef] [PubMed]
- Staerck, C.; Gastebois, A.; Vandeputte, P.; Calenda, A.; Larcher, G.; Gillmann, L.; Papon, N.; Bouchara, J.P.; Fleury, M.J.J. Microbial antioxidant defense enzymes. Microb Pathog 2017, 110, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Dryden, M.; Cooke, J.; Salib, R.; Holding, R.; Pender, S.L.F.; Brooks, J. Hot topics in reactive oxygen therapy: Antimicrobial and immunological mechanisms, safety and clinical applications. J Glob Antimicrob Resist 2017, 8, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Dryden, M. Reactive oxygen species: a novel antimicrobial. Int J Antimicrob Agents 2018, 51, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Webster, S.J.; Daigneault, M.; Bewley, M.A.; Preston, J.A.; Marriott, H.M.; Walmsley, S.R.; Read, R.C.; Whyte, M.K.B. Distinct cell death programs in monocytes regulate innate responses following challenge with common causes of invasive bacterial disease. J Immunol 2010, 185, 2968–2979. [Google Scholar] [CrossRef] [PubMed]
- Bassoe, C.F.; Smith, I.; Sørnes, S.; Halstensen, A.; Lehmann, A.K. (2000) Concurrent measurement of antigen- and antibody-dependent oxidative burst and phagocytosis in monocytes and neutrophils. Methods 2000, 21, 203–220. [Google Scholar] [CrossRef]
- Jarchum, I.; Liu, M.; Shi, C.; Equinda, M.; Pamer, E.G. Critical role for MyD88-mediated neutrophil recruitment during Clostridium difficile colitis. Infect Immun 2012, 80, 2989–2996. [Google Scholar] [CrossRef]
Figure 1.
Morphological appearance of porcine monocytes and C. difficile vegetative cells and spores.Figure 1(A) FACS analysis of uninfected porcine monocytes showing a SSc high/CD14high phenotype (large cells expressing CD14) typical of monocytes. Figure 1(B) DAPI staining showing the typical densely packed nuclear DNA in uninfected macrophages with a ‘C-shaped’ morphology with Phallodin marking the extent of the cell outlines. Figure 1(C) Phalloidin staining of cultures and spores of C. difficile strain CD630 and R20291 showing oval to elongated bacilli. Bacteria spores are notoriously difficult to stain and this was expected to be the case with C. difficile spores.
Figure 1.
Morphological appearance of porcine monocytes and C. difficile vegetative cells and spores.Figure 1(A) FACS analysis of uninfected porcine monocytes showing a SSc high/CD14high phenotype (large cells expressing CD14) typical of monocytes. Figure 1(B) DAPI staining showing the typical densely packed nuclear DNA in uninfected macrophages with a ‘C-shaped’ morphology with Phallodin marking the extent of the cell outlines. Figure 1(C) Phalloidin staining of cultures and spores of C. difficile strain CD630 and R20291 showing oval to elongated bacilli. Bacteria spores are notoriously difficult to stain and this was expected to be the case with C. difficile spores.
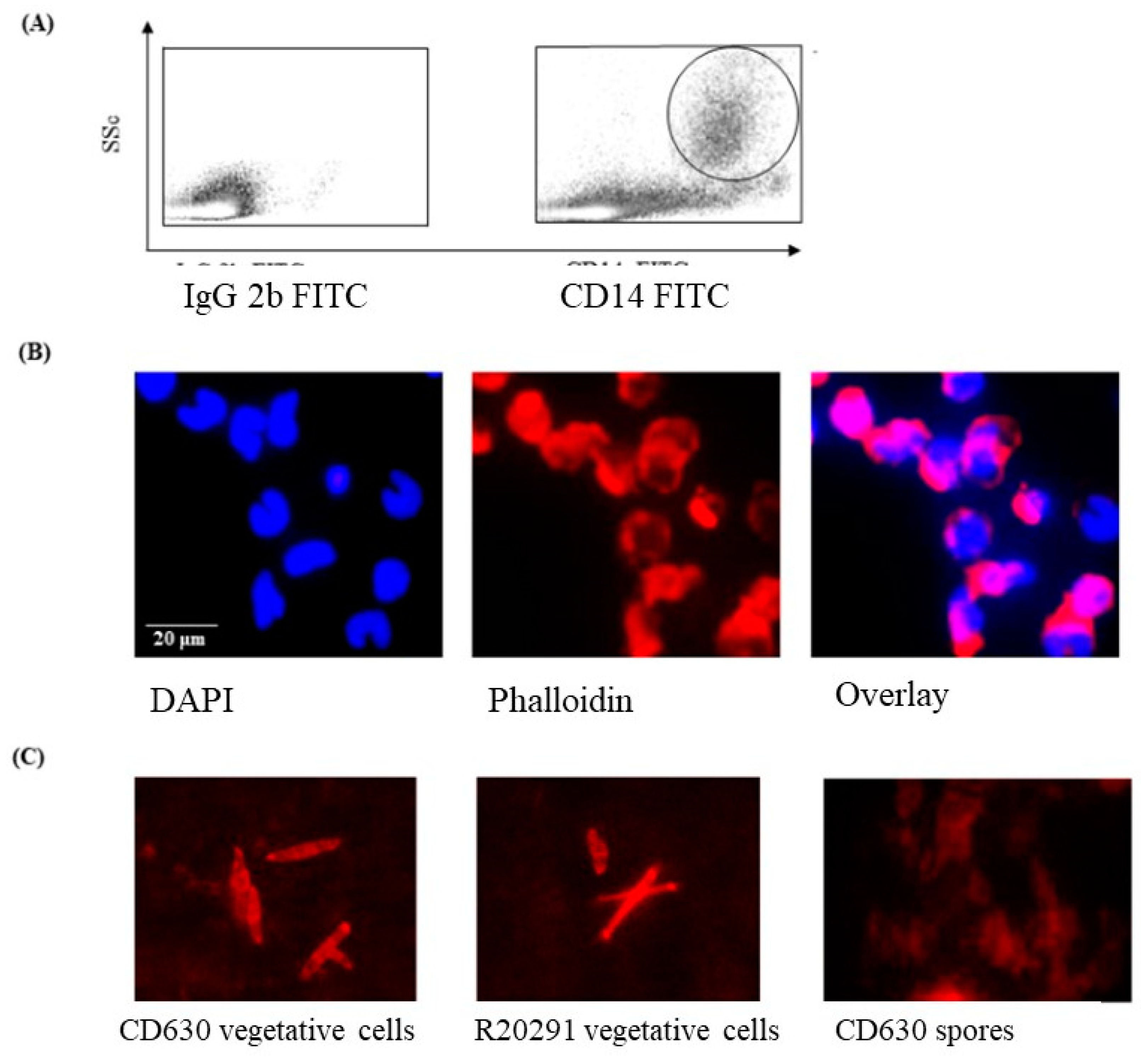
Figure 2.
Survival of C. difficile CD630 and R20291 in porcine monocytes and production of ROS and RNS.Figure 2(A) The numbers (CFU/ml lysate) of C. difficile CD630 and R20291 recovered after 3h and 24h infection of porcine macrophages. Figure 2B,C. Production of reactive oxygen species (2B) detected by NBT or reactive nitrogen species (2C) detected by Griess reagent. Black bars = 3h pi; White bars = 24h pi. Positive controls were zymosan (ROS) or PMA (RNS). Error bars show the standard deviation from the mean. * = significant difference (P<0.05) from uninfected/unstimulated control monocytes. Data shown were calculated from mean values obtained from 8 pigs, each performed in triplicate.
Figure 2.
Survival of C. difficile CD630 and R20291 in porcine monocytes and production of ROS and RNS.Figure 2(A) The numbers (CFU/ml lysate) of C. difficile CD630 and R20291 recovered after 3h and 24h infection of porcine macrophages. Figure 2B,C. Production of reactive oxygen species (2B) detected by NBT or reactive nitrogen species (2C) detected by Griess reagent. Black bars = 3h pi; White bars = 24h pi. Positive controls were zymosan (ROS) or PMA (RNS). Error bars show the standard deviation from the mean. * = significant difference (P<0.05) from uninfected/unstimulated control monocytes. Data shown were calculated from mean values obtained from 8 pigs, each performed in triplicate.
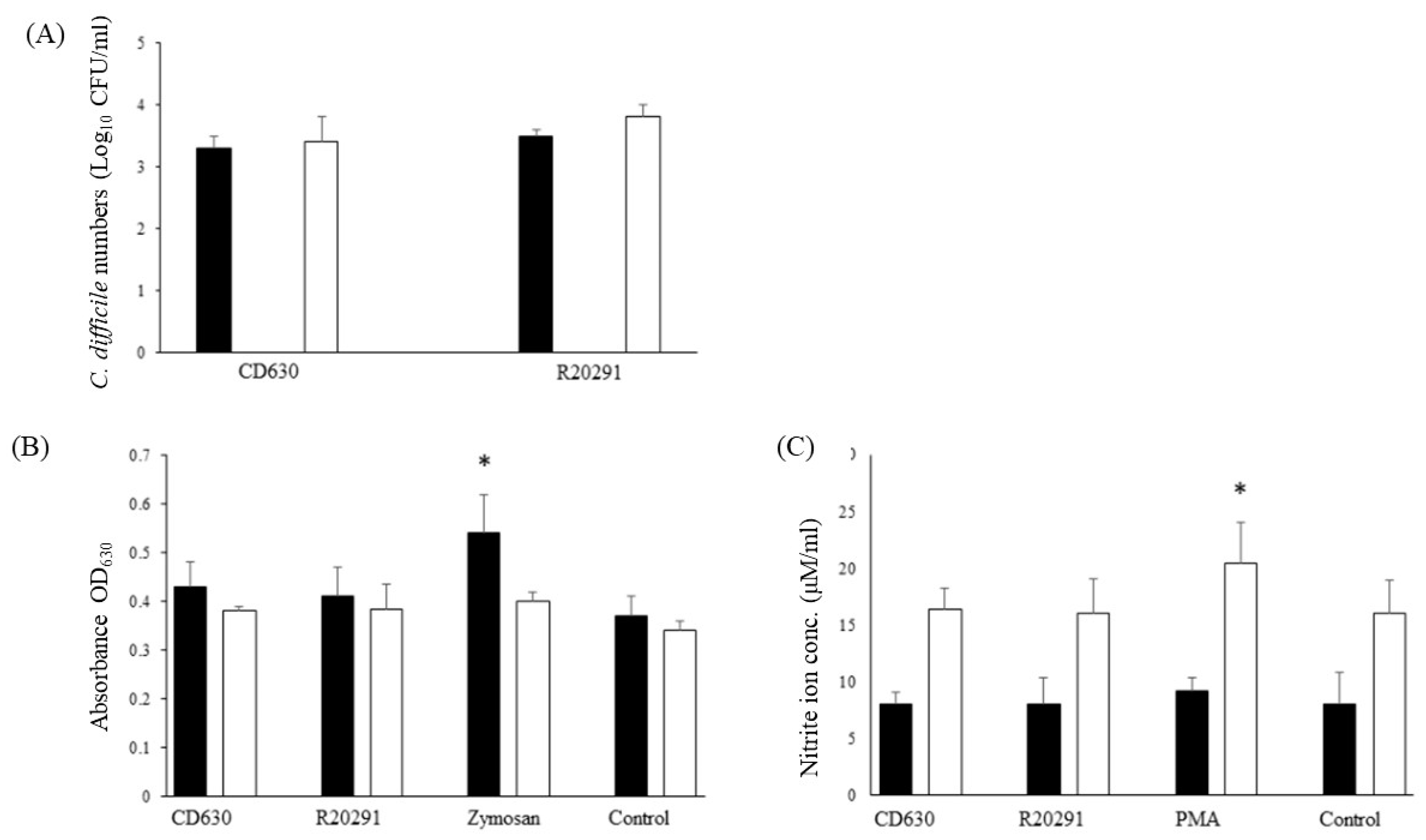
Figure 3.
Production of porcine extracellular monocyte DNA in response to C. difficile infection. DAPI (Figure 3A) and phalloidin (Figure 3B) staining at 3h pi. Network of DNA (A, solid arrows) inter-connecting infected cells with diffuse outlines (B, broken arrows) clearly visible. Individual bacterial cells possibly visible in overlay (Figure 3C, solid arrows). Images shown are of C. difficile CD630 infection and are representative of images taken from monocytes isolated from 3 individual pigs. Similar images observed for strain R20291. Scale bars (20 μm) are shown.
Figure 3.
Production of porcine extracellular monocyte DNA in response to C. difficile infection. DAPI (Figure 3A) and phalloidin (Figure 3B) staining at 3h pi. Network of DNA (A, solid arrows) inter-connecting infected cells with diffuse outlines (B, broken arrows) clearly visible. Individual bacterial cells possibly visible in overlay (Figure 3C, solid arrows). Images shown are of C. difficile CD630 infection and are representative of images taken from monocytes isolated from 3 individual pigs. Similar images observed for strain R20291. Scale bars (20 μm) are shown.
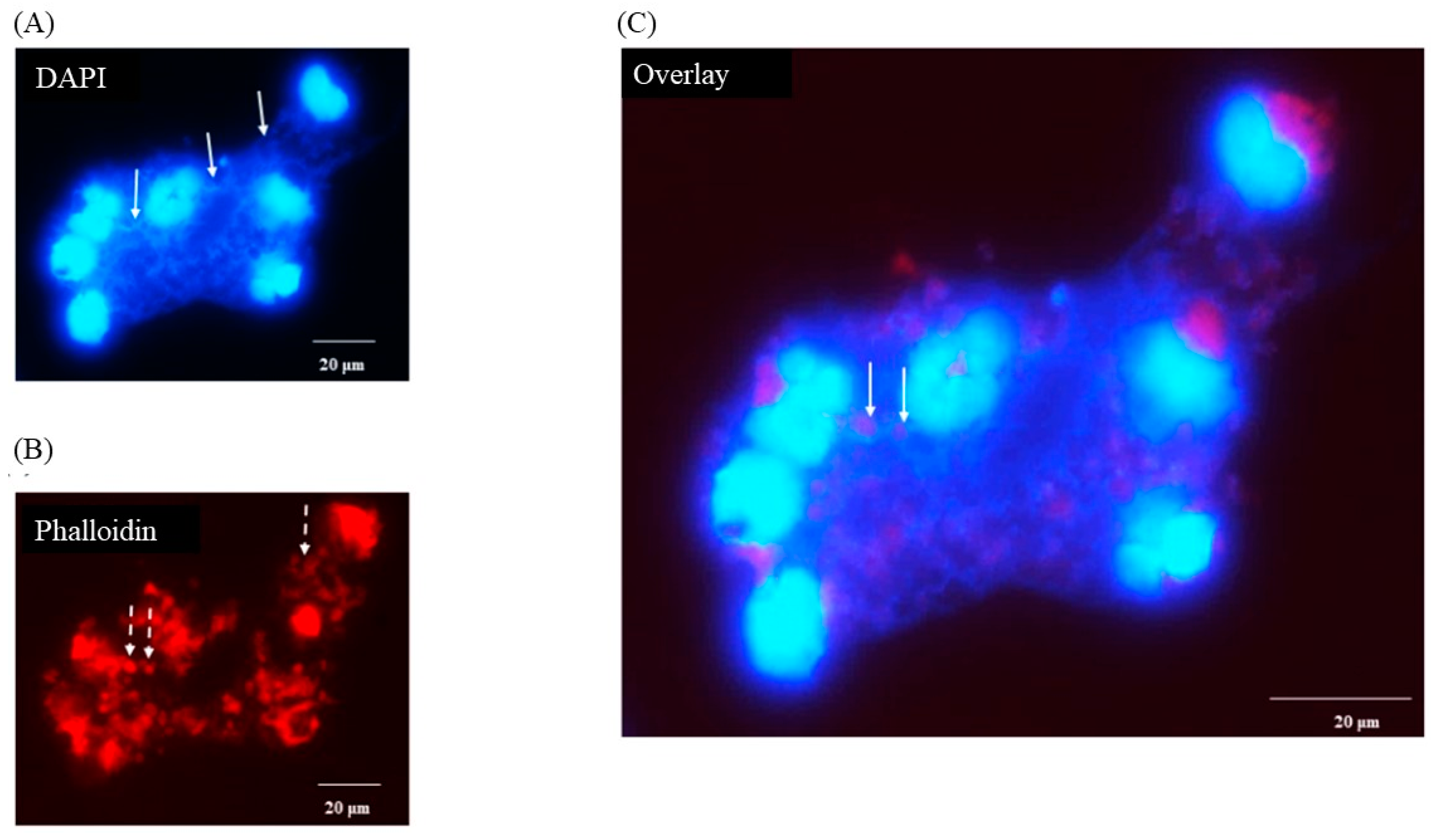
Figure 4.
Porcine monocyte etosis at 3h post-infection with C. difficile CD630. Higher resolution (Grey scale) image of individual infected macrophage stained with DAPI. Fibres of DNA visible protruding through pores in monocyte cell membrane. The classic ‘C-shaped’ morphology was still evident. The scale bar (5um) is shown.
Figure 4.
Porcine monocyte etosis at 3h post-infection with C. difficile CD630. Higher resolution (Grey scale) image of individual infected macrophage stained with DAPI. Fibres of DNA visible protruding through pores in monocyte cell membrane. The classic ‘C-shaped’ morphology was still evident. The scale bar (5um) is shown.
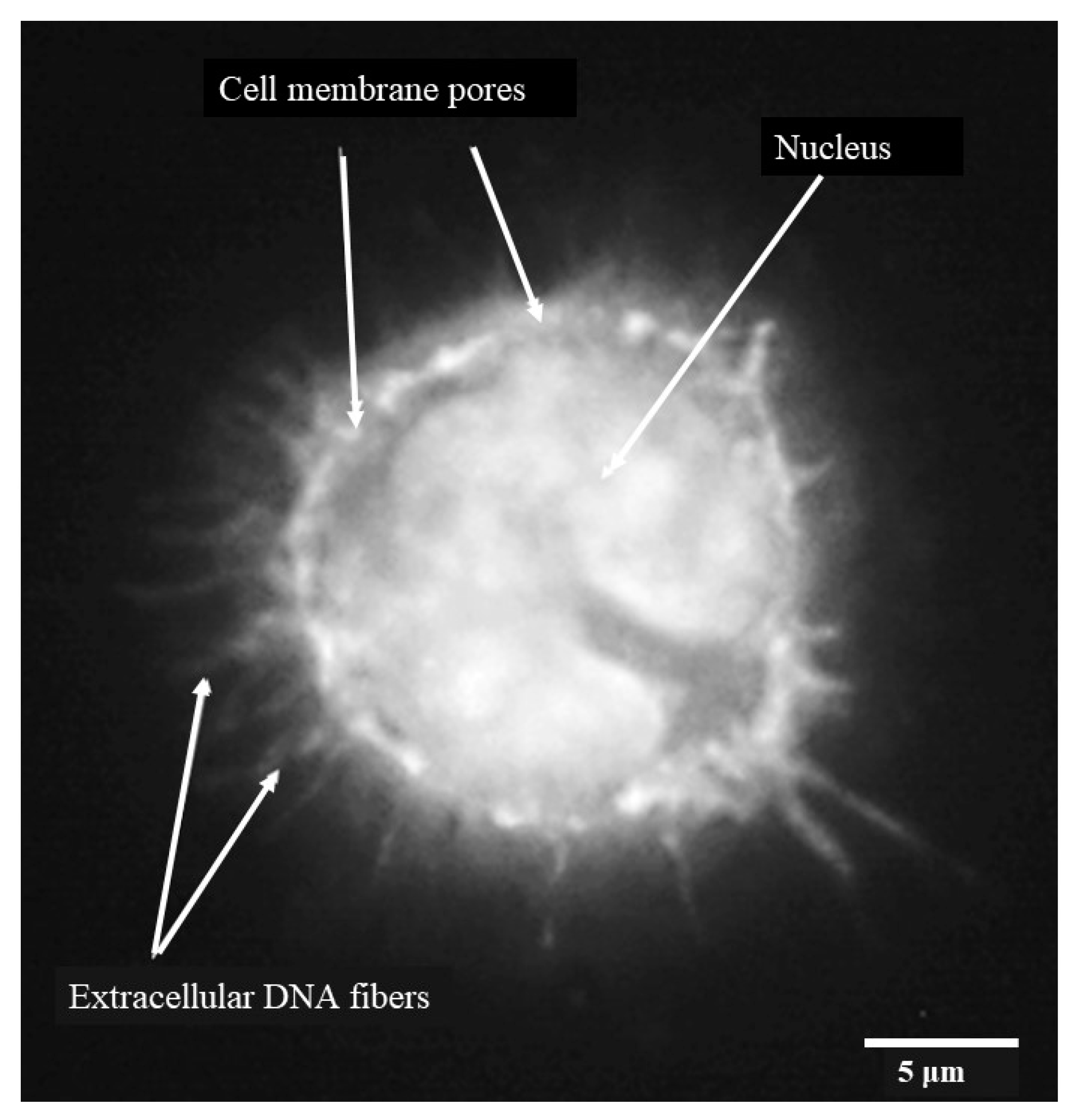
Figure 5.
Porcine monocyte etosis at 24h post-infection with C. difficile. DAPI (Figure 5A, D) and phalloidin (Figure 5B, E) staining of porcine macrophages after 24h infection with C. difficile strains CD630 (Figure 6A-C) and R20291 (Figure 6D-F). Images shown are representative of images taken from monocytes isolated from 3 individual pigs. Scale bars (20 μm) are shown.
Figure 5.
Porcine monocyte etosis at 24h post-infection with C. difficile. DAPI (Figure 5A, D) and phalloidin (Figure 5B, E) staining of porcine macrophages after 24h infection with C. difficile strains CD630 (Figure 6A-C) and R20291 (Figure 6D-F). Images shown are representative of images taken from monocytes isolated from 3 individual pigs. Scale bars (20 μm) are shown.
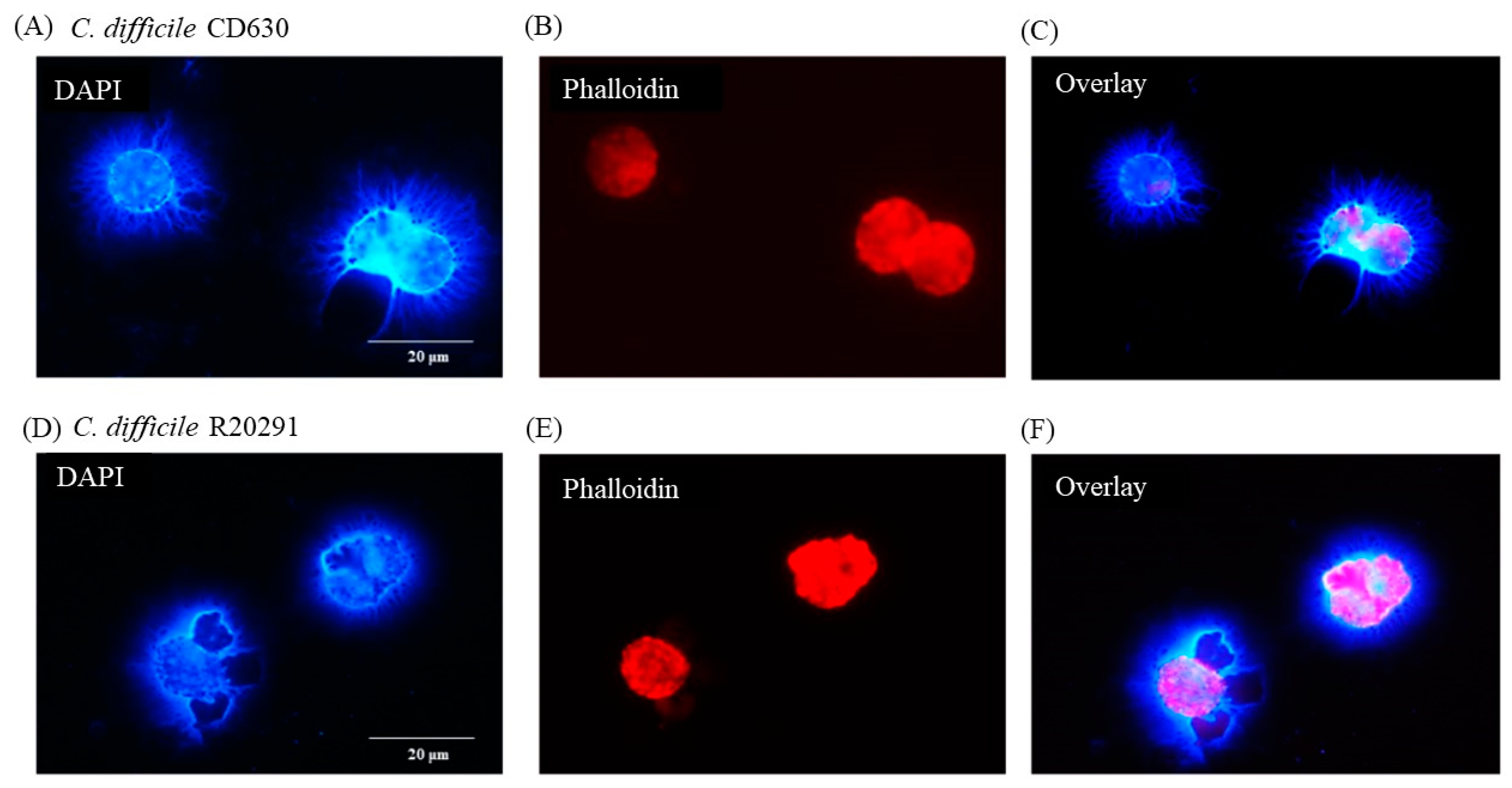
Figure 6.
C. difficile apparently entrapped by DNA released from infected monocytes showing etosis. DAPI (Figure 6A, D) and phalloidin (Figure 6B, E) staining of porcine macrophages 24 h post-infection with C. difficile CD630 (Fig, 6A-C) and R20291 (Figure 6D-F). Bacteria entrapped by DNA released from macrophages marked by arrows. Images shown are representative of images taken from monocytes isolated from 3 individual pigs. Scale bars (20 μm) are shown.
Figure 6.
C. difficile apparently entrapped by DNA released from infected monocytes showing etosis. DAPI (Figure 6A, D) and phalloidin (Figure 6B, E) staining of porcine macrophages 24 h post-infection with C. difficile CD630 (Fig, 6A-C) and R20291 (Figure 6D-F). Bacteria entrapped by DNA released from macrophages marked by arrows. Images shown are representative of images taken from monocytes isolated from 3 individual pigs. Scale bars (20 μm) are shown.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



