Submitted:
25 January 2024
Posted:
26 January 2024
You are already at the latest version
Abstract
Chive (Allium schoenoprasum L.) has a strong antioxidant property as it contains abundant phenolic compounds and ascorbic acid (AsA). In the present study, we investigated the metabolism of phenolic compounds and the change in antioxidant activity in different tissue parts of post-harvest chive. The results showed that compared with the bottom white part (BW), the round green part (RG) exhibited significantly higher content of phenolic compounds, increased enzyme activities and antioxidant activities, indicating that phenolic compounds were mainly synthesized in RG. The expression levels of genes (phenylalanine ammonia lyase (PAL), cinnamate 4-hydroxylase (C4H) and 4-coumaroyl-CoA ligase (4CL)) and their corresponding enzymes activities rapidly decreased in RG, whereas they were maintained in BW, suggesting that senescence occurred more rapidly in RG compared with that in BW. Our study provides a theoretical foundation for further research and development of different parts of Allium plants and offers a basis for consumers' nutritional considerations.
Keywords:
Allium plants
; Antioxidant
; Chive
; Metabolism
; Phenolic compounds
; Postharvest
1. Introduction
The genus Allium, comprising several hundred species, is an important food plant that is used as a drug in traditional and modern medicine for its anti-microbial, lipid-lowering, cardiovascular, hypocholesterolaemic, anti-thrombotic, hypoglycaemic and anti-tumorigenic activities (Abdelrahman et al., 2020). The development of most of the abovementioned diseases is believed to be related to oxidative stress. Organosulfur compounds, abundant in Allium plants, have been extensively studied for their bioactive properties. However, our previous studies (Dai et al., 2023), using different measurement methods, showed that these compounds exhibited weak antioxidant activities. In contrast, phenolic compounds and ascorbic acid (AsA) were found to significantly contribute to the antioxidant activities in chive.
Phenolic compounds with one or more hydroxyl groups exhibit antioxidant activity, which is closely related to the scavenging of reactive oxygen species and stress response (Zhang et al., 2022). Studies have shown that total phenolic content is significantly and positively correlated with antioxidant activity in A. cepa and A. semenovii (Kumar and Kumar, 2023), A. scorodoprasum (Mollica et al., 2018), leek (A. porrum L.) (Biernacka et al., 2021) and garlic (A. sativum L.) (Naheed et al., 2017).
Globally, chive, Allium schoenoprasum L., is cultivated as vegetables or seasoning herbs. According to the literature (Beretta et al., 2017), chives exhibit the highest antioxidant activity compared with other Allium plants (e.g. onion, garlic, shallot, leek and bunching onions). Apart from phenolic compounds, chive also contains ample amount of AsA, which is highly associated with antioxidant activity (Dai et al., 2023).
Allium species have different tissue parts. For instance, chive, leek, Chinese chives (Allium tuberosum) and Welsh onion (Allium fistulosum L.) have a round green part (RG) and a bottom white part (BW), whereas garlic and onion have green leaves and cloves or bulbs. Thus, understanding the phytochemical and physiological properties of different tissues of Allium plants is crucial for developing and utilising these important medicinal plants. Different tissue parts have been proven to possess different concentrations of phenolic compounds in A. scorodoprasum (Mollica et al., 2018), A. galanthum (Kadyrbayeva et al., 2021) and A. ursinum (Lachowicz et al., 2017). The dynamics of phenolic compounds and antioxidant activities have also been studied in post-harvest onion (Gouda et al., 2023), garlic (Naheed et al., 2017) and leek (Bernaert et al., 2013). However, information regarding these dynamics in different parts of post-harvest Allium plants is scarce. Our previous study showed that a higher content of organosulfur compounds was mainly present in BW in chive, and these compounds were continuously translocated from the predominantly biosynthesised tissue (RG) of chive to BW during post-harvest storage (Dai et al., 2022a; Dai and Yu, 2022b). However, the mechanism of the biosynthesis of phenolic compounds that occurs in different parts of Allium plants during post-harvest storage is unclear.
This study aimed to accomplish the following: (i) characterise the dynamics of phenolic compounds and antioxidant activities in different parts of post-harvest chive and (ii) elucidate the mechanism underlying the biosynthesis of phenolic compounds in different parts of Allium plants during post-harvest storage. Our study will provide new insights into phenolic compound biosynthesis in different tissue parts of post-harvest chive and offer a theoretical basis for further research and development of Allium plants, considering different tissues containing various bioactive compounds.
2. Materials and Methods
2.1. Materials and treatments
Chive (A. schoenoprasum L. cv. Xinghua), grown in Zhujia village, Xinghua County, Jiangsu Province, China, was used for this study. Harvested chives (day 0) were transported to the laboratory within 3 h at 20 °C. Senescent leaves were removed, and those with uniform sizes were selected and dried using a fan. The chives were then stored at room temperature (RT, 20 °C) or low temperature (LT, 4 °C).
At each testing point, 3 kg of chives were sampled in each group, with three biological replicates (1 kg per replicate). The RG and BW of the chive were cut into pieces of 0.5 – 1 cm, separately, and immediately frozen in liquid nitrogen and stored at −80 °C for further analysis.
2.2. Preparation of extracts
The samples were frozen with liquid nitrogen and ground into a powder. A 1 g sample was extracted with 5 mL of 70 % ethanol for 1 h at 35 °C in a water bath, with continuous shaking at 120 rpm. The mixture was centrifuged at 12,000 × g for 10 min at 4 °C, and the supernatant was collected. The residue was extracted with another 3 mL of 70 % ethanol under the same conditions. Subsequently, the supernatants were combined. These extracts were used for the Folin–Ciocalteu assay, AsA content determination and antioxidant activity assessment.
The extracts used to determine the phenolic compounds (including free and bound) were prepared using the method described by (Beato et al., 2011), with modifications. Fresh chive powder (5 g), ground in liquid nitrogen, was weighed into a 250-ml flask, and 25 mL of 2 M HCl was added. The mixture was refluxed at 80 °C for 1.5 h in the dark using a shaker at 100 rpm. After cooling, the sample extract was adjusted to pH 2 with 6 N NaOH. Phenolic compounds were extracted three times with 20 ml of ethyl acetate, each in a shaker at 250 rpm and 25 °C for 20 min in the dark. The combined extracts were evaporated to 3 – 5 mL, transferred to a 10 mL tube and completely dried with nitrogen. Subsequently, 1 mL of methanol was added to the tube to dissolve the phenolic compounds. The extract was filtered and subjected to ultra-high-performance liquid chromatography coupled to tandem mass spectrometry (UHPLC-MS/MS) to test the phenolic compounds.
2.3. Folin–Ciocalteu assay and determination of ascorbic acid
Total phenolic content was determined using the Folin–Ciocalteu method (TPC-FC) described by (Dai et al., 2023). Absorption was measured at 760 nm against a blank, and the amount of TPC-FC was expressed as gallic acid equivalents (mg gallic acid/g sample) through the calibration curve of gallic acid.
To determine AsA content, 1 g of sample powder ground with liquid nitrogen was homogenised in 10 mL of 5 % (w/v) trichloroacetic acid and then centrifuged at 12,000 × g for 10 min at 4 °C. The supernatant was collected to determine the AsA content, which was measured following the method outlined by (Kampfenkel et al., 1995).
2.4. UHPLC-MS/MS method
Phenolic compounds were determined using the UHPLC-MS/MS method reported by (Han et al., 2016) with modifications. These compounds were quantified using a model Exion LC system (AB SCIEX, USA) interfaced with a Triple Quad 5500 mass spectrometer (AB SCIEX, USA) equipped with a Turbo electrospray ion source. Analyst LC/MS Data Acquisition software was used for instrument control and data acquisition, whereas SCIEX OS Qualitative and Quantitative Analysis software were used for data evaluation.
The phenolic compounds were separated using a Kinetex F5 LC column (2.1 × 100 mm, 2.6 μm, 100 Å) (Phenomenex, USA) at 40 °C. Chromatographic conditions comprised 0.2 % acetic acid in distilled water (solvent A) and acetonitrile containing 0.2 % acetic acid (solvent B) as the mobile phase, with a flow rate of 0.15 mL min−1 and a gradient of 2 % B (0 min), 70 % B (12 min), 95 % B (12.5 min), 95 % B (13.5 min), 2 % B (14 min) and 2 % B (17 min). The injection volume was 1 μL, and the column temperature was maintained at 40 °C for chromatographic separation. The LC effluent was connected to the interface through a divert valve to prevent clogging of the mass spectrometer cone.
Mass spectrometry employed electrospray ionisation in the negative ion mode. Quantification was conducted using multiple reaction monitoring modes to monitor precursor–product ion transitions, with corresponding parameters detailed in Table S1. The general settings employed included curtain gas at 35 psi, collision gas at 8 psi, ion source gas (1) at 55 psi, ion source gas (2) at 55 psi, temperature at 550 °C and ion spray voltage at 5500 V.
2.5. Determination of enzyme activities involved in phenolic compounds biosynthesis and degradation
Frozen powder (1 g) was homogenised using 5.0 mL of 100 mM phosphate buffer (pH 6.0) and 2 % polyvinylpolypyrrolidone, then centrifuged at 10,000 × g for 30 min at 4 °C. The activities of phenylalanine ammonia-lyase (PAL), cinnamate 4-hydroxylase (C4H) and 4-coumaroyl-CoA ligase (4CL) were determined using ELISA kits (Mallbio, Nanjing Mallbio Biological Technology Co., Ltd.), following the methods described by (Olivares et al., 2017). The absorbance readings of the reaction mixtures were taken at 290 nm (PAL), 333 nm (4CL) and 340 nm (C4H), with one unit (U) of enzyme activity defined as a 0.01 increase in absorbance per hour per gram.
Peroxidase (POD) and polyphenol oxidase (PPO) activities were determined following the method proposed by (Zhang et al., 2022). One unit of POD and PPO activity was defined as an increase of 1 in absorbance per minute per gram at 460 nm and 420 nm, respectively. Units for all enzymes were expressed as U g−1 based on fresh weight.
2.6. Determination of relative gene expression
Real-time quantitative PCR (qPCR) was used to test the relative expression levels of the targeted genes in the phenylpropanoid metabolism pathway in RG and BW. Twenty-three unigenes associated with the phenylpropanoid metabolism pathway were analysed using an SYBR Premix Ex Taq II (TaKaRa, Dalian, China) kit, and the reaction was conducted using the StepOne Real-Time PCR System (Applied Biosystems, Foster City, CA, USA). Three replicates were conducted, and the relative expression levels of the selected unigenes were normalised (with the ACTIN gene as an internal control) and determined using the 2−△△Ct method. The primers used for qPCR analysis are presented in Supplementary Table S2.
2.7. Determination of antioxidant activities
We followed the procedures described by (Dai et al., 2023) to determine the ferric-reducing antioxidant power (FRAP), scavenging activity towards 1,1- diphenyl-1-picrylhydrazyl (DPPH), superoxide anion (O2•-) and hydroxyl radical (•OH). Absorbance was measured at 593 nm (FRAP), 517 nm (DPPH), 510 nm (•OH) and 320 nm (O2•-). The relative FRAP capacity and DPPH scavenging capacity of the chive extracts were expressed in µmol of Trolox equivalents per gram of fresh weight (µmol TE g−1). The relative •OH and O2•- scavenging capacity of the chive extracts were expressed in mg of AsA equivalents per gram of fresh weight (mg VE g−1).
3. Results and Discussion
3.1. Principal component analysis
In this study, principal component analysis was performed to visualise the variation and correlation among 20 individual phenolic compounds, AsA content, antioxidant activities and the activities of the enzymes involved in the biosynthesis and degradation of phenolic compounds in different parts of chive during storage (Figure 1). The loading plot (Figure 1B) shows that AsA, antioxidant indexes, enzyme indexes and most of the phenolic compounds (trans-cinnamic acid (TCA), p-coumaric acid (PCA), ferulic acid (FA), caffeic acid (CA), gallic acid (GA), p-hydroxybenzoic acid (HBA), sinapic acid (SA), isorhamnetin (IH), quercetin (QC), vanillic acid (VA), dihydroquercetin (DHQC), naringenin chalcone (NGNC) and the sum of total phenolic compounds (TPC-LC)) significantly contributed to PC1. In contrast, neochlorogenic acid (NCGA), chlorogenic acid (CGA), rutin, hesperetin (HPT), luteolin (LL) and apigenin (API) significantly contributed to PC2. PC1 mainly differentiated between the RG and BW groups, indicating that part specificity was the main contributor to clustering. PC2 primarily differentiated between the RT and LT groups. The RG group exhibited higher content of most phenolic compounds, AsA, antioxidant activities and enzyme activities compared with those in the BW, whereas the BW group exhibited higher content of DHQC, VA and phenylalanine (PA).
3.2. Dynamics of antioxidant compounds
3.2.1. Dynamics of TPC-FC and ascorbic acid
TPC-FC was 3- to 12-fold higher in RG than in BW during storage (Figure 2A). In RG, TPC-FC declined on day 1 and gradually increased thereafter under room and low temperatures, with 10.77 % – 14.75 % higher content observed in chives under RT than under LT. In BW, TPC-FC continuously increased during storage, with 28.59 % –46.45 % higher content observed in chives under RT than under LT.
AsA content was 1.1- to 4.0-fold higher in RG than that in BW (Figure 2B). In RG, AsA decreased on day 5 under RT, whereas it decreased on day 1 under LT, followed by an increase. Furthermore, in RG, AsA content was 15.13 % higher on day 1 but 12.51 % lower on day 5 compared with that under LT. Conversely, in BW, AsA content was observed to gradually rise under both temperatures, with no significant difference between the content under RT and LT.
3.2.2. Dynamics of phenolic compounds determined using LC–MS
Twenty-one phenolic compounds were detected in the chives (Figure 3 and Table S3), and the LC–MS chromatograph is shown in Figure S1. Regarding RG, FA (22.22 – 28.79 mg kg−1) and PCA (11.63 – 17.40 mg kg−1) were the most abundant, followed by HBA (2.26 – 6.15 mg kg−1), QC (1.52 – 3.81 mg kg−1), SA (1.39 – 2.81 mg kg−1), VA (1.24 – 1.80 mg kg−1), IH (0.63 – 1.69 mg kg−1) and CA (0.95 – 1.45 mg kg−1). For BW, VA (1.60 – 2.30 mg kg−1), FA (0.85 – 1.93 mg kg−1) and HBA (0.90 – 1.16 mg kg−1) were the major phenolic compounds, followed by PCA (0.22 – 0.39 mg kg−1) and SA (0.24 – 0.34 mg kg−1). Rutin, HPT, API and LL were detected in trace amounts in chives.
Currently, FA is utilised as an additive antioxidant with pharmacological applications for human health (Biernacka et al., 2021). In this study, FA and PCA were predominant in chives, consistent with a previous study (Ferreira et al., 2022). It appeared that different phenolic compounds dominated in different Allium species, for example, GA in A. stylosum (Emir and Emir, 2021), PCA in A. subhirsutum (Emir et al., 2020a), protocatechuic acid and VA in A. semenovii (Kumar and Kumar, 2023), FA in leek (Biernacka et al., 2021) and QC in onion (Cecchi et al., 2020) were reported as the most dominant phenolic compounds in these species.
HBA, VA and GA, which are hydroxybenzoic acids, were detected in chives. HBA content in RG was 1.62- to 4.94-fold and 1.52- to 2.81-fold higher than those in BW under RT and LT, respectively. HBA content increased in chives under both temperatures. The HBA content in RG was higher under RT than under LT, especially on day 5, with a 127.15 % increase in content. The HBA content in BW was 9.19 % – 16.64 % higher under RT than under LT. VA is a natural antioxidant present in fruits and vegetables (Castillejo et al., 2021). Unlike other phenolic compounds, VA content was observed to be 13.35 % – 70.23 % and 13.35 % – 52.75 % higher in BW than in RG under RT and LT, respectively. VA content increased in BW during storage, with a 14.34 % – 22.27 % higher content under RT compared with that at LT. In RG, VA content increased during the late storage stage, with a 45.66 % higher content under RT than that under LT on day 5. GA was 1.37- to 15.07-fold and 2.68- to 15.07-fold higher in RG than in BW under RT and LT, respectively. HBA, VA and GA content were lower in chives under LT than under RT, suggesting that LT suppressed hydroxybenzoic acid biosynthesis in RG and BW.
PA is the initial substance in the phenylpropanoid pathway and can be catalysed into phenolic acids (Zhou et al., 2023). The concentration of PA in RG and BW increased continuously during storage, with a more pronounced increase in BW than in RG under both temperatures. PA content was higher in both RG and BW under RT than under LT. TCA content in RG were 35.78 % – 168.96 % and 49.72 % – 205.00 % higher than in BW under RT and LT, respectively. The TCA content in RG was 8.36 % and 60.35 % higher under RT than under LT on days 1 and 3, respectively. The TCA content in BW was 143.40 % and 52.79 % higher under RT than under LT on days 1 and 3, respectively. PCA content was 36.09- to 73.70-fold and 32.83- to 62.86-fold higher in RG than in BW under RT and LT, respectively. Furthermore, in RG, PCA content increased under RT but declined under LT. The PCA content in RG was 19.46 % – 47.88 % higher under RT than under LT. Conversely, in BW, PCA content initially increased and then decreased under both temperatures. Additionally, PCA content in BW was higher on day 1 but lower on day 3 and 5 under RT compared with LT.
NGNC is a key compound for the biosynthesis of downstream flavonoids. NGNC content was 23.68- to 51.69-fold and 27.85- to 71.75-fold higher in RG than in BW under RT and LT, respectively. NGNC content in RG was higher on day 1 but lower on day 5 under RT compared with LT. NGNC content in BW was higher under RT than under LT. Moreover, dihydrokaempferol (DHKF) content in RG were 13.74 % –36.99 % higher under LT than under RT. However, DHKF was not detected in BW. DHQC content was 158.24 % – 184.81 % and 65.38 % – 186.35 % higher in BW than in RG under RT and LT, respectively. DHQC content in RG and BW showed a gradually decreasing trend under both temperatures, with a more rapid decline under RT than under LT. NGNC is a precursor of DHKF, which is further converted to DHQC in the phenylpropanoid pathway (Zhou et al., 2020). NGNC content was significantly lower in BW than in RG, whereas DHQC content was higher in BW than in RG. These results suggest that flavonoids are rapidly biosynthesised in BW during storage.
QC, the main flavonoid detected, was 22.22- to 222.81-fold and 20.63- to 235.98-fold higher in RG than in BW under RT and LT, respectively. In both RG and BW, QC increased under both temperatures. The QC content in RG was 10.93 % – 92.06 % higher under RT than under LT. The QC content in BW was 244.53 % – 572.83 % higher under RT than under LT. IH content was 32.69- to 91.85-fold and 36.90- to 91.85-fold higher in RG than in BW under RT and LT, respectively. IH showed an increasing trend in RG and BW, except for a decline observed on day 1 in RG under LT. The IH content in RG was 28.02 % – 85.47 % higher under RT than under LT. The QC content in BW was 92.18 % – 195.51 % higher under RT than under LT.
Flavonoids are important antioxidant compounds that protect fruits and vegetables against oxidative damage throughout the post-harvest period (Castillejo et al., 2021). The observed higher content of QC and IH in RG than in BW were in agreement with a previous study reporting the green shaft of leek containing a higher flavonoid content than the white shaft (Biernacka et al., 2021). Moreover, the increased content (the difference in the values obtained at the end of storage and day 0) of QC and IH was much higher in RG than that in BW under both temperatures, suggesting that flavonoids were mainly biosynthesised in RG during storage. However, notably, the increased percentage ((value obtained at the end of storage − value obtained on day 0)/value obtained on day 0) of flavonoids was significantly higher in BW than in RG. The increased percentage of QC in RG was 97.01 % and 92.33 % under RT and LT, respectively, whereas in BW, it was 1028.57 % and 1890.39 % under RT and LT, respectively. Similarly, the percentage increase of IH in RG was 69.28 % and 59.74 % under RT and LT, respectively, whereas in BW, it was 366.50 % and 201.37 % under RT and LT, respectively. Furthermore, the flavonoid content in chives were higher under RT than under LT, indicating that LT suppressed their biosynthesis.
CA content was 46.06- to 93.38-fold and 13.95- to 51.63-fold higher in RG than in BW under RT and LT, respectively. However, CA content in RG were observed to fluctuate during storage, with 10.84 % – 39.11 % higher content under RT than under LT. In contrast, CA content in BW initially increased and then declined under both temperatures, with 186.36 % and 126.67 % higher content under LT than under RT on days 3 and 5, respectively. This result was contrary to that observed in RG. FA was 16.80- to 32.42-fold and 11.01- to 32.42-fold higher in RG than in BW under RT and LT, respectively. In RG, FA decreased initially and then increased under both temperatures, with 8.28 % – 24.35 % higher content observed under RT than under LT. Conversely, FA in BW initially increased and then declined. FA in BW under RT was higher on day 1 but was lower on day 5 as compared with that under LT. SA in RG was 6.98- to 8.46-fold and 4.39- to 8.46-fold higher than in BW under RT and LT, respectively. In RG, SA showed a gradual increase under RT, whereas it sharply dropped on day 1 under LT and then increased to the initial value on day 5. SA in RG was higher under RT than under LT, especially on day 1, with an 86.26 % higher content. In BW, SA initially increased and then decreased during the late storage stage, with a higher content under RT than under LT.
TPC-LC in RG was 8.71- to 12.32-fold and 7.52- to 12.32-fold higher than in BW under RT and LT, respectively. In RG, TPC-LC increased under RT, whereas it initially decreased and then gradually increased from day 3 till the end of storage under LT. TPC-LC in RG was 17.92 % – 33.37 % higher under RT than under LT. In BW, TPC-LC increased under both temperatures, with a 10.85 % – 25.36 % higher content under RT on days 1 and 3 but a lower content on day 5 compared with that under LT.
Hydroxycinnamic acids, such as PCA, CA, FA and SA are important antioxidants and precursors of monolignols, directly linked to lignin biosynthesis (Metsämuuronen and Sirén, 2019; Zhang et al., 2022). As TCA, PCA, CA and FA decreased on day 1 and/or 3 in RG, a corresponding increase occurred in BW. These phenolic acids, along with SA and TPC-LC in RG, exhibited lower content under LT than under RT, suggesting that LT inhibited their biosynthesis. However, in BW, some of these compounds (e.g. PCA and CA on days 3 and 5 and FA on day 5) were higher under LT than under RT. Additionally, the increased content of these compounds in RG were significantly higher under RT than under LT, including FA (0.36, −0.63 mg kg−1), PCA (1.93, −1.85 mg kg−1), HBA (2.94, 1.24 mg kg−1), VA (0.40, 0.10 mg kg−1), total phenolic acids (6.46, −0.81 mg kg−1) and TPC-LC (8.90, 1.48 mg kg−1). However, the increased levels of these compounds in BW were much less under RT than under LT for FA (0.24, 0.81 mg kg−1), PCA (−0.01, 0.05 mg kg−1), HBA (0.09, 0.22 mg kg−1), VA (0.57, 0.66 mg kg−1), total phenolic acids (1.04, 1.85 mg kg−1) and TPC-LC (1.07, 1.93 mg kg−1). The biosynthesis of phenolic compounds in RG was inhibited under LT. Conversely, BW showed much more increased content of phenolic acids under LT than under RT. One possible explanation could be that LT maintained the quality of chive and promoted the transfer of phenolic compounds from RG to BW.
The inhibitory effect of LT on phenolic compounds in post-harvest chive aligns with previous studies, which reported reduced anthocyanin content in blood oranges stored at 4 °C compared with 9 °C (Carmona et al., 2017). This alignment is further supported by the continuous accumulation of anthocyanin and phenolic compounds in Purple-Pericarp Supersweet Sweetcorn during storage at 23 °C while remaining unchanged at 4 °C (Hong et al., 2021) and the lower content of phenolic compounds in mango fruits stored at 5 °C compared with those stored at 13 °C (Vithana et al., 2018).
Overall, the content of most phenolic compounds in RG was significantly higher than that in BW, except for VA, PA, DHQC, CGA and NCGA. The difference in the composition and concentration of phenolic compounds between RG and BW aligns with several previous studies reporting (1) the bulb extract of A. scorodoprasum as inferior in phenolic compounds compared with the flower and stem (Mollica et al., 2018), (2) a much higher amount of the major phenolic kaempferol in leaves than in the bulb of Allium galanthum (Kadyrbayeva et al., 2021), (3) higher phenolic content in leaves than in bulb extracts of A. ursinum (Lachowicz et al., 2017) and (4) the green shaft of leek containing a higher content of phenolic acids compared with that in the white shaft (Biernacka et al., 2021).
3.3. Dynamics of enzyme activities involved in the metabolism of phenolic compounds in post-harvest chive
Generally, phenylpropanoid biosynthesis initiates with the deamination of phenylalanine by PAL, yielding cinnamic acid and downstream phenylpropanoids (Castillejo et al., 2021). C4H catalyses the conversion of TCA to PCA (Zheng et al., 2021). Furthermore, 4CL catalyses the conversion of PCA to p-coumaroyl CoA, a central intermediate for various phenylpropanoid metabolites (Zheng et al., 2021). PAL activity in RG was 57.39 % – 95.30 % higher than in BW (Figure 4A). The PAL activity was 11.34 % – 21.62 % higher in RG under RT than under LT. The PAL activity was 11.40 % – 20.70 % higher in BW under RT than under LT on days 1 and 3. C4H activity in RG was 28.21 % – 83.42 % higher than that in BW (Figure 4B). On days 1 and 5, C4H activity was 18.03 % and 17.30 % higher in RG under RT than under LT and 15.43 % and 15.77 % higher in BW under RT than under LT, respectively. 4CL activity in RG was 11.28 % – 131.38 % higher than in BW (Figure 4C). In RG, 4CL activity was 14.85 % – 24.80 % higher under RT than under LT, with no significant difference observed between both temperatures in BW. The activities of these enzymes in RG significantly declined in the first three days, exhibiting slight changes in BW during the same period. These findings suggest that physiological changes occurred more rapidly in RG compared with BW, indicating faster senescence in RG.
The main enzymes involved in the catabolism of phenolic compounds in fruits and vegetables are PPO and POD (Castillejo et al., 2021). POD is involved in the final step of monolignol polymerisation, forming lignin and eliminating H2O2 (Zhang et al., 2022). PPO activity was 40.13 % – 86.67 % higher in RG than in BW (Figure 4D). It decreased in chives under both temperatures, being 18.75 % and 9.60 % higher on days 1 and 3 but 9.54 % lower on day 5 in RG under RT compared with that under LT. PPO activity in BW was 13.73 % and 9.04 % higher under RT than under LT on days 1 and 3, respectively. POD activity was 1.9- to 3.9-fold higher in RG than in BW (Figure 4E). It increased in chives under both temperatures, except for a decline on day 5 in RG under RT. On days 1 and 3, it was 24.33 % and 20.56 % higher in RG under RT but 18.12 % lower on day 5 compared with that under LT. Conversely, it was 20.92 % higher on day 5 in BW under RT than under LT.
The enzymes associated with the biosynthesis and degradation of phenolic compounds showed higher activities in RG compared with BW. LT inhibited both the biosynthesis and degradation of phenolic compounds in post-harvest chive. This finding is consistent with reduced PAL activity in sweet cherry stored at LT (1 °C) compared with RT (20 °C) (Tsaniklidis et al., 2017) and lower POD activity in chives stored at 3 °C compared with 20 °C (Dai et al., 2021).
3.4. Dynamics of gene expression
As shown in Figure 5, the relative expression of PAL in RG under both RT and LT initially declined and then increased during the late storage stage. Conversely, PAL in BW remained constant under RT but declined initially and then increased under LT. PAL generally maintained a higher level of expression in BW than in RG during storage, with a 0.82- and 1.89-fold increase in expression levels on days 1 and 3 under RT and 0.23- to 0.70-fold higher expression from days 1–12 under LT. LT inhibited PAL expression during storage. C4H1 in chives showed different trends under RT and LT, decreasing under RT but increasing under LT. Its expression was higher in RG compared with BW under LT, showing a 0.26- to 0.76-fold increase. Conversely, C4H2 showed a declining trend, with elevated expression in BW than in RG, with a 0.54- to 0.90-fold increase under RT, and 0.15- to 0.80-fold increase under LT. LT inhibited C4H2 expression on days 3 and 5 compared with RT. Both 4CL1 and 4CL2 showed decreasing trends during storage. BW showed a higher expression level in 4CL2 than RG, with a 0.56- to 0.88-fold increase under RT and a 1.85- to 3.57-fold increase from days 5 to 19 under LT. Notably, LT inhibited the expression of 4CL1 and 4CL2.
PAL, C4H2 and 4CL2 exhibited higher expression levels in BW than in RG post-harvest, indicating that BW maintained gene expression in the phenylpropanoid pathway. LT almost suppressed the expression of PAL, C4H2, 4CL1 and 4CL2, indicating that LT inhibited the pathway. These findings align with previous studies reporting that LT suppresses the expression levels of genes related to phenolic compound biosynthesis in sweet cherry (Tsaniklidis et al., 2017) and blood orange (Carmona et al., 2017).
CHS converts 4-coumaroyl-CoA to naringenin chalcone, which is further converted to naringenin by CHI (Tao et al., 2022). Naringenin, which contains a flavonoid skeleton, can be modified and transformed into different flavonoids (e.g. DHQC, DHKF, QC and IH) (Zhang et al., 2022) by several enzymes, including flavanone 3-hydroxylase (F3H), flavonoid 3'-hydroxylase (F3'H), flavonoid 3',5'-hydroxylase (F3'5'H) and flavonol synthase (FLS).
CHS in chives initially decreased and then increased at the later stage of storage under both temperatures, with significantly higher expression in BW on days 1 and 3 under RT (2.19-fold and 4.66-fold) and on days 1 and 12 under LT (5.35-fold and 1.45-fold) compared with that in RG. CHI1 and CHI3 in chives under both temperatures decreased, except CHI3 in BW under RT, which remained constant. CHI1 exhibited significantly higher levels in BW than in RG under both temperatures, with a 2.93- to 10.90-fold increase on days 1 and 3 under RT and from 2.83- to 5.85-fold increase during LT storage. CHI3 also showed significantly higher levels in BW than in RG under both temperatures, with a 2.1- to 7.9-fold increase under RT and a 0.89- to 4.69-fold increase under LT. LT increased the expression of CHI1 on day 1 but decreased its expression on days 3 and 5. Additionally, LT increased CHI3 expression on day 1 in BW but decreased its level on days 3 and 5 in BW.
F3H in chives initially declined and then increased under RT, whereas it gradually decreased under LT. BW exhibited a significantly higher expression level of F3H on days 1 and 3 under RT (3.26- and 16.43-fold) and on day 1 under LT (4.79-fold) compared with that in RG. LT suppressed its expression in RG and BW on day 1, but enhanced F3H expression in BW on day 3 and in both RG and BW on day 5. F3'H, responsible for DHQC synthesis, exhibited decreased expression levels in chives at both temperatures. F3'H showed significantly higher expression levels in BW than in RG, with a 13.00- to 45.32-fold increase under RT and a 3.55- to 68.20-fold increase under LT. Moreover, LT promoted F3'H expression. Generally, the expression levels of F3'5'H in chives showed a declining trend under both temperatures. RG exhibited a higher expression of F3'5'H during the late storage stage under RT and LT compared with that in BW. LT inhibited F3'5'H expression during storage. FLS expression showed a declining trend in chives under both temperatures. FLS was expressed more in BW than RG, with a 1.91- to 8.86-fold increase under RT and a 1.08- to 5.16-fold increase under LT. FLS expression level was higher on day 1 in both RG and BW under LT than under RT. The FLS expression level in BW was higher on days 3 and 5 under RT than under LT.
CHS, CHI1, CHI 3, F3H, F3'H and FLS showed higher expression levels in BW than in RG during storage, indicating that the genes in BW related to flavonoid biosynthesis were more active than those in RG. LT generally increased the expression of CHS, CHI1, CHI3, F3H and FLS in RG and BW on day 1 but suppressed them on days 3 and/or 5. These findings were consistent with previous studies reporting (1) a lower expression level of genes in the flavonoid pathway in blood oranges during cold storage at 4 °C than at 9 °C (Carmona et al., 2017) and (2) the reduced transcription of genes involved in anthocyanin biosynthesis in sweet cherry during cold storage compared with room storage (Tsaniklidis et al., 2017).
The genes coding shikimate O-hydroxycinnamoyltransferase (HCT1 and HCT2) in chives decreased during storage at both temperatures, except for the stable expression of HCT1 in BW under RT and an increase in HCT2 expression in BW under RT. HCT1 demonstrated significantly higher expression levels in RG than in BW, with 3.51- to 6.67-fold increase under RT, and with 4.67- to 9.23-fold increase under LT. The expression level of HCT2 was generally higher in RG than in BW, with a 0.30- to 3.70-fold increase under RT and a 0.18- to 1.34-fold increase under LT. LT inhibited HCT1 and HCT2 expression in RG and BW during storage. The gene coding ferulate 5-hydroxylase (F5H) in chives declined during RT and LT storage, with a higher expression level in RG than in BW. LT increased F5H in RG on day 1 but decreased its expression on days 3 and 5. Under both temperatures, 3-O-methyltransferase (COMT) in chives initially increased and then declined, except for a fluctuation in RG under LT. COMT expression was higher in RG than in BW, with a 1.95- to 2.42-fold increase under RT and a 0.90- to 3.24-fold increase under LT. Notably, LT increased COMT expression in BW on day 1 but suppressed its expression in RG on days 3 and 5.
Cinnamoyl CoA reductase (CCR) is the primary enzyme responsible for producing monolignols from phenylpropanoid metabolites in chives. Generally, CCR exhibited an increasing trend under RT and LT conditions during storage, with higher expression in RG than in BW, with a 0.25- and 0.51-fold increase on days 1 and 3 under RT, and 0.15- to 0.50-fold increase from days 5 – 19 under LT. Cinnamyl alcohol dehydrogenase (CAD) catalyses the final step in the monolignol pathway. Overall, RG showed a higher expression level of CAD1 compared with that in BW, with a 0.15- to 0.81-fold increase from days 1 to 12 under LT. CAD6 expression was significantly higher in RG than in BW, with a 0.93- to 2.67-fold increase during RT storage and a 0.25- to 2.80-fold increase during LT storage. LT inhibited CAD6 expression in chives on day 5.
POD1 expression in RG declined at both temperatures, whereas it remained largely unchanged in BW during storage. A significantly higher expression of POD1 was observed in RG than in BW, with a 3.43- to 8.10-fold increase under RT and a 6.33- to 16.59-fold increase under LT. LT induced its expression in RG on days 1 and 3. POD42 expression increased under both temperatures, with a higher expression level in BW than in RG, showing a 10.80- to 19.12-fold increase under RT and a 11.84- to 50.84-fold increase under LT. This result indicates that POD1 dominated in RG, whereas POD42 dominated in BW. PPO1 in RG showed a declining trend under RT and LT during storage, whereas it fluctuated in BW under RT, and decreased in BW under LT. The expression of PPO3 in RG and BW exhibited an increasing trend under RT and LT during storage, with a significantly higher expression in RG than in BW, showing a 0.90- to 2.0-fold increase under RT and a 1.02- to 4.51-fold increase under LT. LT inhibited the expression of PPO1 and PPO3 in RG and BW.
RG showed higher expression levels of HCT1, HCT2, F5H, COMT, CAD1, CAD6, POD1 and POD6 than BW, which are related to lignin biosynthesis (Liu et al., 2018), indicating that the genes related to lignin biosynthesis were more active in RG than in BW. Moreover, HCT1, HCT2, F5H and COMT are also involved in phenolic acid biosynthesis, indicating that the genes related to phenolic acid biosynthesis were more active in RG than in BW. The expression of HCT1, HCT2, F5H, COMT and CAD6 were suppressed under LT, indicating that LT inhibited the biosynthesis of phenolic acids and lignin.
3.5. Dynamics of antioxidant activities in post-harvest chive
As shown in Figure 6, FRAP content was 2.9- to 8.9-fold higher in RG than in BW. Moreover, in RG, they were 25.88 % higher on day 1 and 8.39 % lower on day 5 under RT compared with that under LT. DPPH content was 1.3- to 4.9-fold higher in RG than in BW. DPPH scavenging in RG was 40.70 % and 15.56 % higher on days 1 and 3, respectively, under RT than under LT, whereas it was 8.64 % lower on day 5 under RT than under LT. •OH scavenging was 2.3- to 4.4-fold higher in RG compared with BW. In RG, it was 19.39 % and 10.35 % higher under RT on days 1 and 3, respectively, than under LT. In BW, it was 18.58 % higher on day 5 under RT than under LT. O2•- scavenging was 2.0- and 3.9-fold higher in RG than in BW. Furthermore, in RG, it was 17.23 % and 11.57 % higher under RT than under LT on days 1 and 3. The antioxidant activities in RG slightly changed under RT and remarkably decreased on day 1, followed by a gradual increase to the initial level under LT. In contrast, the activities in BW continuously increased under both temperatures. These findings suggest that the performance of antioxidant activity in post-harvest chive differed in different tissue parts.
RG exhibited significantly higher antioxidant activity than BW. This aligns with observations of (1) significantly higher DPPH scavenging and reducing ability in the green part of leek than in the white part (Biernacka et al., 2021), (2) higher antioxidant potential in leaves than in bulb extracts in Allium ursinum L. (Lachowicz et al., 2017) and (3) increased antioxidant activity in the aerial parts of all Allium species compared with the bulbs (Rocchetti et al., 2022). LT suppressed the antioxidant activities in RG on days 1 and/or 3, and inhibited •OH scavenging in BW on day 5. This is consistent with reduced antioxidant activity in mango fruits stored at 5 °C than at 13 °C (Vithana et al., 2018) and lower total antioxidant activity in eggplants stored at 1 °C than at 10 °C (Kantakhoo et al., 2022).
3.6. Dynamics of other nutrients
In our previous work (Dai et al., 2022a) (Table S4), S-alk(en)ylcysteine sulfoxides (CSOs) content were higher in BW than in RG. CSOs, primarily biosynthesised in RG (Dai and Yu, 2022b), remained largely unchanged in RG, whereas they continued to accumulate in BW. Free amino acids (FAAs) have also been found to significantly accumulate in BW (Dai et al., 2022a), primarily when proteolysis occurs in RG. These findings suggest that compounds for sulphur and nitrogen storage accumulated in BW in post-harvest chive.
3.7. Correlation analysis
The correlation analysis is presented in Figure 7. Tissue type showed a high relationship with each parameter (|r| > 0.8**, P < 0.01), indicating a correlation between parameters and tissue types. VA displayed a negative correlation with other phenolic compounds. Antioxidant activity parameters strongly correlated with HBA, QC, IH, FA, SA, TPC-LC, TPC-FC and AsA (r > 0.8**, P < 0.01). This was in alignment with reports that showed increased AsA and TPC content in onions treated with licorice root extract and Trichoderma album thereby enhancing total antioxidant activity (Younes et al., 2023; Younes et al., 2021) and the highest phenolics coinciding with the highest DPPH scavenging and FRAP in stems compared with bulbs and flowers in Allium pallens L. (Emir and Emir, 2020b). Enzymes involved in the biosynthesis and degradation of phenolic compounds exhibited a positive correlation with each phenolic compound, TPC-LC and TPC-FC (r > 0.65**, P < 0.01), except for a negative correlation with VA.
A comparison of gene expression and enzyme activities in the phenylpropanoid pathway in different parts of the post-harvest chive is shown in Figure 8A. Enzymes related to the biosynthesis and degradation of phenolic compounds showed significantly higher activities in RG than in BW, indicating that the biosynthesis and degradation of phenolic compounds primarily occurred in RG. The activities of biosynthetic enzymes in RG remarkably declined compared with those in BW, indicating a substantial physiological change in RG. Genes related to phenolic acids and lignin biosynthesis, including HCT, COMT, F5H, CCR and CAD, showed higher expression levels in RG than in BW, suggesting a rapid lignin biosynthesis in RG. The expression levels of most genes related to flavonoid biosynthesis greatly decreased in RG and showed lower expression levels than in BW, indicating that BW maintained the expression of genes related to flavonoid biosynthesis.
Overall, phenolic compounds were primarily synthesised in RG, but BW maintained the expression of biosynthetic genes, the activities of biosynthetic enzymes for phenolic compounds and the expression of genes related to flavonoid biosynthesis (Figure 8B). AsA content and antioxidant activities measured using different methods gradually increased in BW, whereas they remained constant in RG. Other nutrients, such as CSOs and FAAs, were constantly transferred from RG to BW, accumulating in BW. These results suggest that RG, as the main organ for nutrient biosynthesis in the development and growth of chive, undergoes rapid senescence post-harvest. BW, as the primary storage organ, undergoes senescence much more slowly than RG. This aligns with the distinct appearance of different tissue parts in post-harvest chive.
4. Conclusion
To the best of our knowledge, this study represents the first investigation on the metabolism of phenolic compounds and changes in antioxidant activity in different tissues of post-harvest chive. The results show that RG exhibited significantly higher content of AsA, phenolic compounds, metabolic enzyme activities and antioxidant activities compared with those of BW, indicating that phenolic compounds were mainly synthesised in RG. The expression of related genes and enzyme activities rapidly decreased in RG, whereas they were maintained in BW, suggesting a rapid occurrence of senescence in RG than that in BW. LT inhibited gene expression, enzyme activities and antioxidant activities but promoted the transfer of phenolic compounds from RG to BW. Our previous work demonstrated a constant transfer of CSOs and FAAs from RG to BW, resulting in their accumulation in BW. Combined with this finding, we can conclude that RG, as the primary organ for nutrient biosynthesis in the development and growth of chive, undergoes faster senescence post-harvest compared with that in BW, which was the main storage organ. The present study provides a theoretical foundation for further research and development of different parts of Allium plants and offers a basis for consumers' nutritional considerations.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Xiaomei Dai: Conceptualization, Methodology, Software, Writing an original draft. Chonglei Jia: Methodology, Writing—review & editing. Jiaqi Lu: Investigation, validation. Zhifang Yu: Supervision, Project administration.
Acknowledgments
This research was funded by the Priority Academic Program Development of Jiang Su Higher Education Institutions (PAPD) and the Key project of Jiang Su Province Modern Agriculture (BE2018382).
Conflicts of Interest
The authors declare that we do not have any commercial or associative interest that represents a conflict of interest in connection with the work submitted. All authors have read and agreed to the published version of the manuscript.
References
- Abdelrahman, M., Ariyanti, N.A., Sawada, Y., Tsuji, F., Hirata, S., Hang, T.T.M., Okamoto, M., Yamada, Y., Tsugawa, H., Hirai, M.Y., Shigyo, M., 2020. Metabolome-based discrimination analysis of shallot landraces and bulb onion cultivars associated with differences in the amino acid and flavonoid profiles. Molecules 25. [CrossRef]
- Beato, V.M., Orgaz, F., Mansilla, F., Montano, A., 2011. Changes in phenolic compounds in garlic (Allium sativum L.) owing to the cultivar and location of growth. Plant Foods for Humman Nutrition 66, 218-223. [CrossRef]
- Beretta, H.V., Bannoud, F., Insani, M., Berli, F., Hirschegger, P., Galmarini, C.R., Cavagnaro, P.F., 2017. Relationships Between Bioactive Compound Content and the Antiplatelet and Antioxidant Activities of Six Allium Vegetable Species. Food Technology and Biotechnology 55, 266-275. [CrossRef]
- Bernaert, N., De Clercq, H., Van Bockstaele, E., De Loose, M., Van Droogenbroeck, B., 2013. Antioxidant changes during postharvest processing and storage of leek (Allium ampeloprasum var. porrum). Postharvest Biology and Technology 86, 8-16. [CrossRef]
- Biernacka, B., Dziki, D., Kozlowska, J., Kowalska, I., Soluch, A., 2021. Dehydrated at Different Conditions and Powdered Leek as a Concentrate of Biologically Active Substances: Antioxidant Activity and Phenolic Compound Profile. Materials (Basel) 14. [CrossRef]
- Carmona, L., Alquezar, B., Marques, V.V., Pena, L., 2017. Anthocyanin biosynthesis and accumulation in blood oranges during postharvest storage at different low temperatures. Food Chemistry 237, 7-14. [CrossRef]
- Castillejo, N., Martínez-Zamora, L., Gómez, P.A., Pennisi, G., Crepaldi, A., Fernández, J.A., Orsini, F., Artés-Hernández, F., 2021. Postharvest yellow LED lighting affects phenolics and glucosinolates biosynthesis in broccoli sprouts. Journal of Food Composition and Analysis 103. [CrossRef]
- Cecchi, L., Ieri, F., Vignolini, P., Mulinacci, N., Romani, A., 2020. Characterization of Volatile and Flavonoid Composition of Different Cuts of Dried Onion (Allium cepa L.) by HS-SPME-GC-MS, HS-SPME-GCxGC-TOF and HPLC-DAD. Molecules 25. [CrossRef]
- Dai, X., Jia, C., Lu, J., Yu, Z., 2023. The dynamics of bioactive compounds and their contributions to the antioxidant activity of postharvest chive (Allium schoenoprasum L.). Food Research International 174. [CrossRef]
- Dai, X., Lu, Y., Yang, Y., Yu, Z., 2021. 1-Methylcyclopropene preserves the quality of chive (Allium schoenoprasum L.) by enhancing its antioxidant capacities and organosulfur profile during storage. Foods 10. [CrossRef]
- Dai, X., Yu, H., Zhu, L., Yu, Z., 2022a. S-alk(en)ylcysteine sulfoxides biosynthesis and free amino acids profile in different parts of postharvest chive (Allium schoenoprasum L.). Scientia Horticulturae 303. [CrossRef]
- Dai, X., Yu, Z., 2022b. Transcriptome analysis reveals the genes involved in S-alk(en)ylcysteine sulfoxide biosynthesis and its biosynthetic location in postharvest chive (Allium schoenoprasum L.). Food Research International 158, 111548. [CrossRef]
- Emir, A., Emir, C., 2020b. Chemical profiles and biological properties of methanol extracts of Allium pallens L. from different localities in Turkey. Archives of Biological Sciences 72, 193-201. [CrossRef]
- Emir, A., Emir, C., Yildirim, H., 2020a. Characterization of phenolic profile by LC-ESI-MS/MS and enzyme inhibitory activities of two wild edible garlic: Allium nigrum L. and Allium subhirsutum L. Journal of Food Biochemistry 44, e13165. [CrossRef]
- Emir, C., Emir, A., 2021. Phytochemical analyses with LC-MS/MS and in vitro enzyme inhibitory activities of an endemic species "Allium stylosum O. Schwarz" (Amaryllidaceae). South African Journal of Botany 136, 70-75. [CrossRef]
- Ferreira, F.S., de Oliveira, V.S., Chavez, D.W.H., Chaves, D.S., Riger, C.J., Sawaya, A., Guizellini, G.M., Sampaio, G.R., Torres, E., Saldanha, T., 2022. Bioactive compounds of parsley (Petroselinum crispum), chives (Allium schoenoprasum L) and their mixture (Brazilian cheiro-verde) as promising antioxidant and anti-cholesterol oxidation agents in a food system. Food Research International 151, 110864. [CrossRef]
- Gouda, M., Nassarawa, S.S., Gupta, S.D., Sanusi, N.I., Nasiru, M.M., 2023. Evaluation of carbon dioxide elevation on phenolic compounds and antioxidant activity of red onion (Allium cepa L.) during postharvest storage. Plant Physiology and Biochemistry 200, 107752. [CrossRef]
- Han, C., Ji, Y., Li, M., Li, X., Jin, P., Zheng, Y., 2016. Influence of wounding intensity and storage temperature on quality and antioxidant activity of fresh-cut Welsh onions. Scientia Horticulturae 212, 203-209. [CrossRef]
- Hong, H.T., Phan, A.D.T., O'Hare, T.J., 2021. Temperature and Maturity Stages Affect Anthocyanin Development and Phenolic and Sugar Content of Purple-Pericarp Supersweet Sweetcorn during Storage. Journal of Agricultural and Food Chemistry 69, 922-931. [CrossRef]
- Kadyrbayeva, G., Zagorska, J., Grzegorczyk, A., Gawel-Beben, K., Strzepek-Gomolka, M., Ludwiczuk, A., Czech, K., Kumar, M., Koch, W., Malm, A., Glowniak, K., Sakipova, Z., Kukula-Koch, W., 2021. The Phenolic Compounds Profile and Cosmeceutical Significance of Two Kazakh Species of Onions: Allium galanthum and A. turkestanicum. Molecules 26. [CrossRef]
- Kampfenkel, K., Vanmontagu, M., Inze, D., 1995. Extraction and Determination of Ascorbate and Dehydroascorbate from Plant Tissue. Analytical biochemistry 225, 165-167. [CrossRef]
- Kantakhoo, J., Ose, K., Imahori, Y., 2022. Effects of hot water treatment to alleviate chilling injury and enhance phenolic metabolism in eggplant fruit during low temperature storage. Scientia Horticulturae 304. [CrossRef]
- Kumar, R., Kumar, D., 2023. Comprehensive metabolomics and antioxidant activity of Allium species viz. Allium semenovii, A. sativum and A. cepa: An important spice. Food Research International 166, 112584. [CrossRef]
- Lachowicz, S., Kolniak-Ostek, J., Oszmiański, J., Wiśniewski, R., 2017. Comparison of Phenolic Content and Antioxidant Capacity of Bear Garlic (Allium ursinum L.) in Different Maturity Stages. Journal of Food Processing and Preservation 41. [CrossRef]
- Liu, Q., Luo, L., Zheng, L., 2018. Lignins: Biosynthesis and Biological Functions in Plants. International Journal of Molecular Sciences 19. [CrossRef]
- Metsämuuronen, S., Sirén, H., 2019. Bioactive phenolic compounds, metabolism and properties: a review on valuable chemical compounds in Scots pine and Norway spruce. Phytochemistry Reviews 18, 623-664. [CrossRef]
- Mollica, A., Zengin, G., Locatelli, M., Picot-Allain, C.M.N., Mahomoodally, M.F., 2018. Multidirectional investigations on different parts of Allium scorodoprasum L. subsp. rotundum (L.) Stearn: Phenolic components, in vitro biological, and in silico propensities. Food Research International 108, 641-649. [CrossRef]
- Naheed, Z., Cheng, Z., Wu, C., Wen, Y., Ding, H., 2017. Total polyphenols, total flavonoids, allicin and antioxidant capacities in garlic scape cultivars during controlled atmosphere storage. Postharvest Biology and Technology 131, 39-45. [CrossRef]
- Olivares, D., Contreras, C., Munoz, V., Rivera, S., Gonzalez-Aguero, M., Retamales, J., Defilippi, B.G., 2017. Relationship among color development, anthocyanin and pigment-related gene expression in 'Crimson Seedless' grapes treated with abscisic acid and sucrose. Plant Physiology and Biochemistry 115, 286-297. [CrossRef]
- Rocchetti, G., Zhang, L., Bocchi, S., Giuberti, G., Ak, G., Elbasan, F., Yildiztugay, E., Ceylan, R., Picot-Allain, M.C.N., Mahomoodally, M.F., Lucini, L., Zengin, G., 2022. The functional potential of nine Allium species related to their untargeted phytochemical characterization, antioxidant capacity and enzyme inhibitory ability. Food Chemistry 368, 130782. [CrossRef]
- Tao, X., Wu, Q., Li, J., Huang, S., Cai, L., Mao, L., Luo, Z., Li, L., Ying, T., 2022. Exogenous methyl jasmonate regulates phenolic compounds biosynthesis during postharvest tomato ripening. Postharvest Biology and Technology 184. [CrossRef]
- Tsaniklidis, G., Kafkaletou, M., Delis, C., Tsantili, E., 2017. The effect of postharvest storage temperature on sweet cherry (Prunus avium L.) phenolic metabolism and colour development. Scientia Horticulturae 225, 751-756. [CrossRef]
- Vithana, M.D.K., Singh, Z., Johnson, S.K., 2018. Cold storage temperatures and durations affect the concentrations of lupeol, mangiferin, phenolic acids and other health-promoting compounds in the pulp and peel of ripe mango fruit. Postharvest Biology and Technology 139, 91-98. [CrossRef]
- Younes, N.A., Anik, T.R., Rahman, M.M., Wardany, A.A., Dawood, M.F.A., Tran, L.P., Abdel Latef, A.A.H., Mostofa, M.G., 2023. Effects of microbial biostimulants (Trichoderma album and Bacillus megaterium) on growth, quality attributes, and yield of onion under field conditions. Heliyon 9, e14203. [CrossRef]
- Younes, N.A., Rahman, M.M., Wardany, A.A., Dawood, M.F.A., Mostofa, M.G., Keya, S.S., Abdel Latef, A.A.H., Tran, L.P., 2021. Antioxidants and Bioactive Compounds in Licorice Root Extract Potentially Contribute to Improving Growth, Bulb Quality and Yield of Onion (Allium cepa). Molecules 26. [CrossRef]
- Zhang, F., Xie, Y., Shi, J., Jiang, L., 2022. Effects of 1-methylcyclopropene treatment on phenolic metabolism in postharvest Gynura bicolor DC. Scientia Horticulturae 293. [CrossRef]
- Zheng, X., Jiang, H., Silvy, E.M., Zhao, S., Chai, X., Wang, B., Li, Z., Bi, Y., Prusky, D., 2021. Candida Oleophila Proliferated and Accelerated Accumulation of Suberin Poly Phenolic and Lignin at Wound Sites of Potato Tubers. Foods 10. [CrossRef]
- Zhou, D., Liu, Q., Peng, J., Tu, S., Pan, L., Tu, K., 2020. Metabolic analysis of phenolic profiles reveals the enhancements of anthocyanins and procyanidins in postharvest peach as affected by hot air and ultraviolet C. Postharvest Biology and Technology 167. [CrossRef]
- Zhou, D., Sun, R., Zhu, W., Shi, Y., Ni, S., Wu, C., Li, T., 2023. Impact of dielectric barrier discharge cold plasma on the quality and phenolic metabolism in blueberries based on metabonomic analysis. Postharvest Biology and Technology 197. [CrossRef]
Figure 1.
(A) Sample scattered plot of chive samples and (B) the component loading plot of 32 parameters (including phenolic compounds, ascorbic acid, related enzymes and antioxidant parameters) obtained through principal component analysis. RL, the round green part (RG) stored at 4 oC (LT); BL, the bottom white part (BW) stored at LT; RR, RG stored at 20 oC (RT); BR, BW stored at RT. PA, phenylalanine; TCA, trans-cinnamic acid; PCA, p-coumaric acid; CA, caffeic acid; FA, ferulic acid; SA, sinapic acid; HBA, p-hydroxybenzoic acid; VA, vanillic acid; GA, gallic acid; QC, quercetin; IH, isorhamnetin; DHQC, dihydroquercetin; HPT, hesperitin; NCGA, neochlorogenic acid; CGA, chlorogenic acid; RUT, rutin; API, apigenin; LL, luteolin; NGNC, naringenin chalcone; TPC-FC, total phenolic content determined using Folin-Ciocalteu method; TPC-LC, the sum of phenolic compounds determined by LC-MS; AsA, ascorbic acids.
Figure 1.
(A) Sample scattered plot of chive samples and (B) the component loading plot of 32 parameters (including phenolic compounds, ascorbic acid, related enzymes and antioxidant parameters) obtained through principal component analysis. RL, the round green part (RG) stored at 4 oC (LT); BL, the bottom white part (BW) stored at LT; RR, RG stored at 20 oC (RT); BR, BW stored at RT. PA, phenylalanine; TCA, trans-cinnamic acid; PCA, p-coumaric acid; CA, caffeic acid; FA, ferulic acid; SA, sinapic acid; HBA, p-hydroxybenzoic acid; VA, vanillic acid; GA, gallic acid; QC, quercetin; IH, isorhamnetin; DHQC, dihydroquercetin; HPT, hesperitin; NCGA, neochlorogenic acid; CGA, chlorogenic acid; RUT, rutin; API, apigenin; LL, luteolin; NGNC, naringenin chalcone; TPC-FC, total phenolic content determined using Folin-Ciocalteu method; TPC-LC, the sum of phenolic compounds determined by LC-MS; AsA, ascorbic acids.
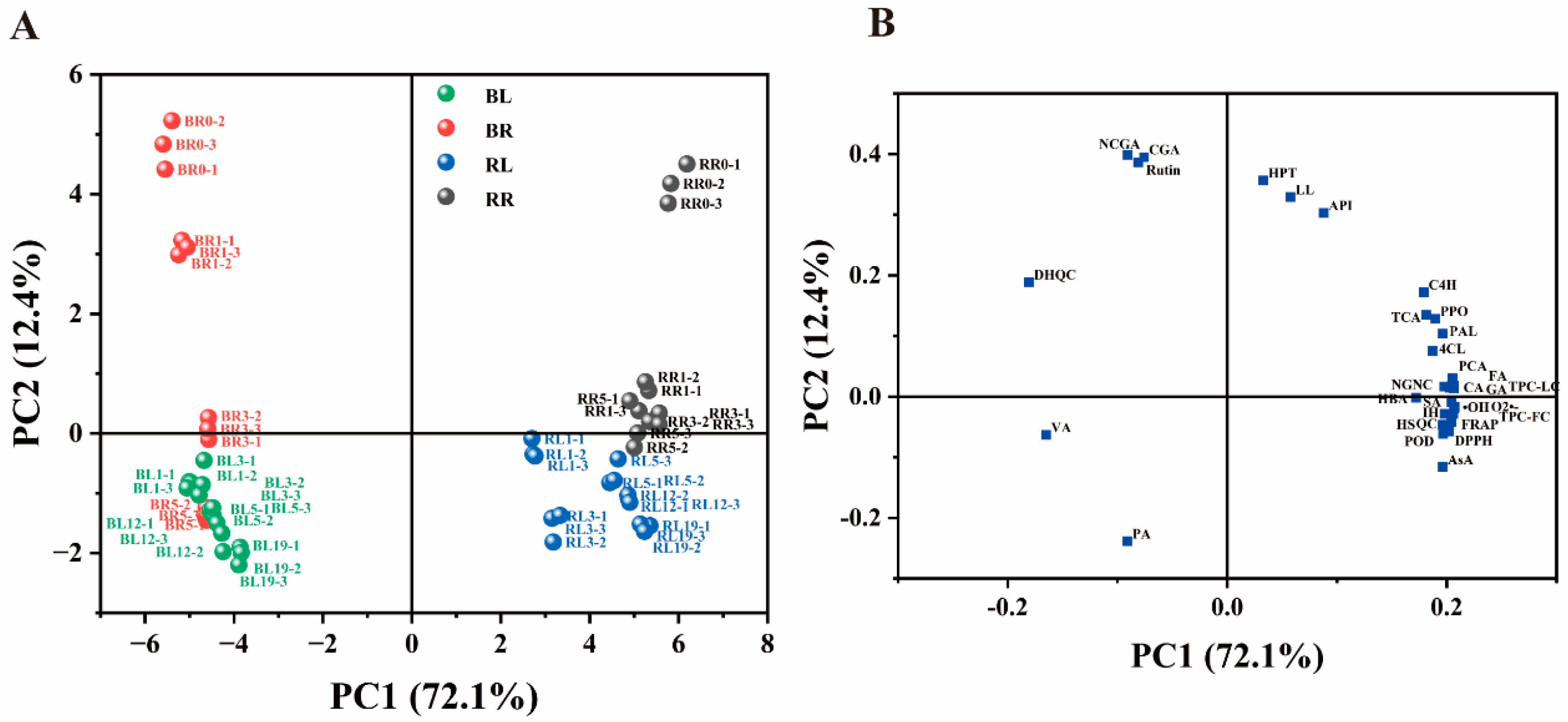
Figure 2.
(A) The content of TPC-FC and (B) Ascorbic acid in different parts of chives during storage at 20 °C (RT) and 4 °C (LT). RG, the round green part. BW, the bottom white part. Data are the means of the three independent measurements. Letters represent statistical differences among different tissues under RT and LT at each time point.
Figure 2.
(A) The content of TPC-FC and (B) Ascorbic acid in different parts of chives during storage at 20 °C (RT) and 4 °C (LT). RG, the round green part. BW, the bottom white part. Data are the means of the three independent measurements. Letters represent statistical differences among different tissues under RT and LT at each time point.
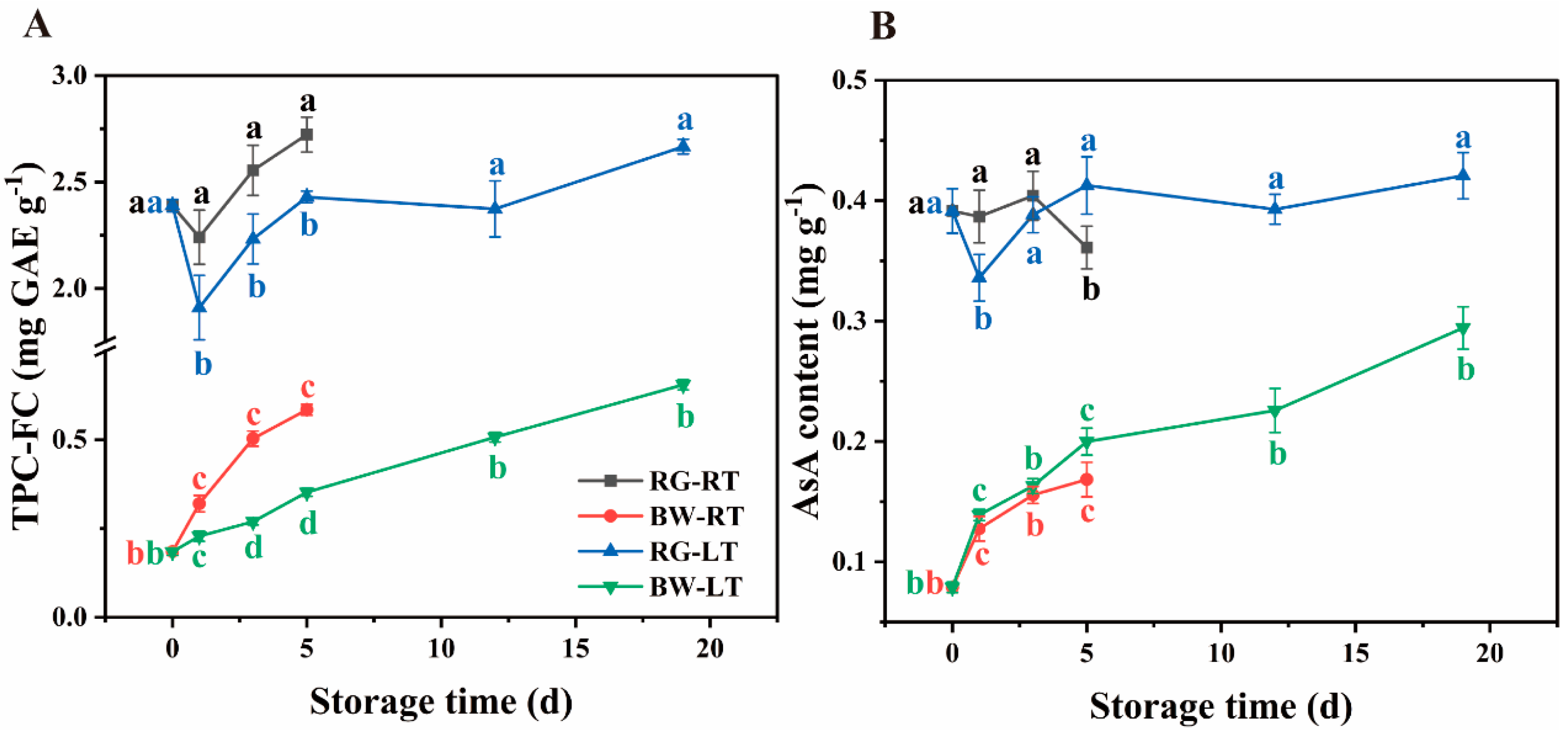
Figure 3.
The phenolic compound profiles in different parts of chive during storage at 20 °C (RT) and 4 °C (LT). RG, the round green part. BW, the bottom white part. Data are the means of the three independent measurements. Letters represent statistical differences among different tissues under RT and LT at each time point.
Figure 3.
The phenolic compound profiles in different parts of chive during storage at 20 °C (RT) and 4 °C (LT). RG, the round green part. BW, the bottom white part. Data are the means of the three independent measurements. Letters represent statistical differences among different tissues under RT and LT at each time point.
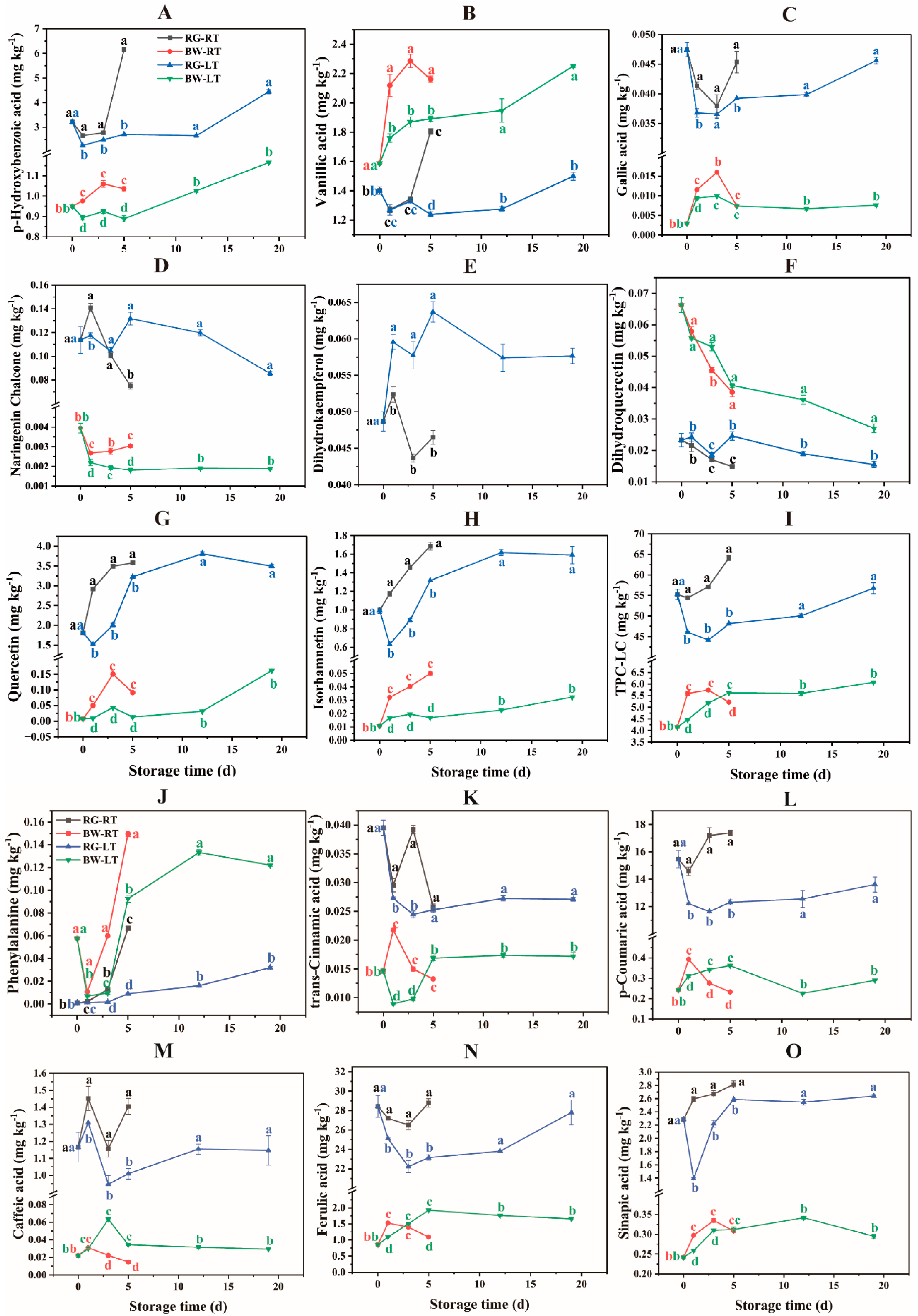
Figure 4.
The activities of enzymes involved in the biosynthesis and degradation of phenolic compounds in different parts of chive during storage at 20 °C (RT) and 4 oC (LT). (A) PAL, (B) C4H, (C) 4CL, d (D) PPO, (E) POD. RG, the round green part. BW, the bottom white part. Data are the means of the three independent measurements. Letters represent statistical differences among different tissues under RT and LT at each time point.
Figure 4.
The activities of enzymes involved in the biosynthesis and degradation of phenolic compounds in different parts of chive during storage at 20 °C (RT) and 4 oC (LT). (A) PAL, (B) C4H, (C) 4CL, d (D) PPO, (E) POD. RG, the round green part. BW, the bottom white part. Data are the means of the three independent measurements. Letters represent statistical differences among different tissues under RT and LT at each time point.
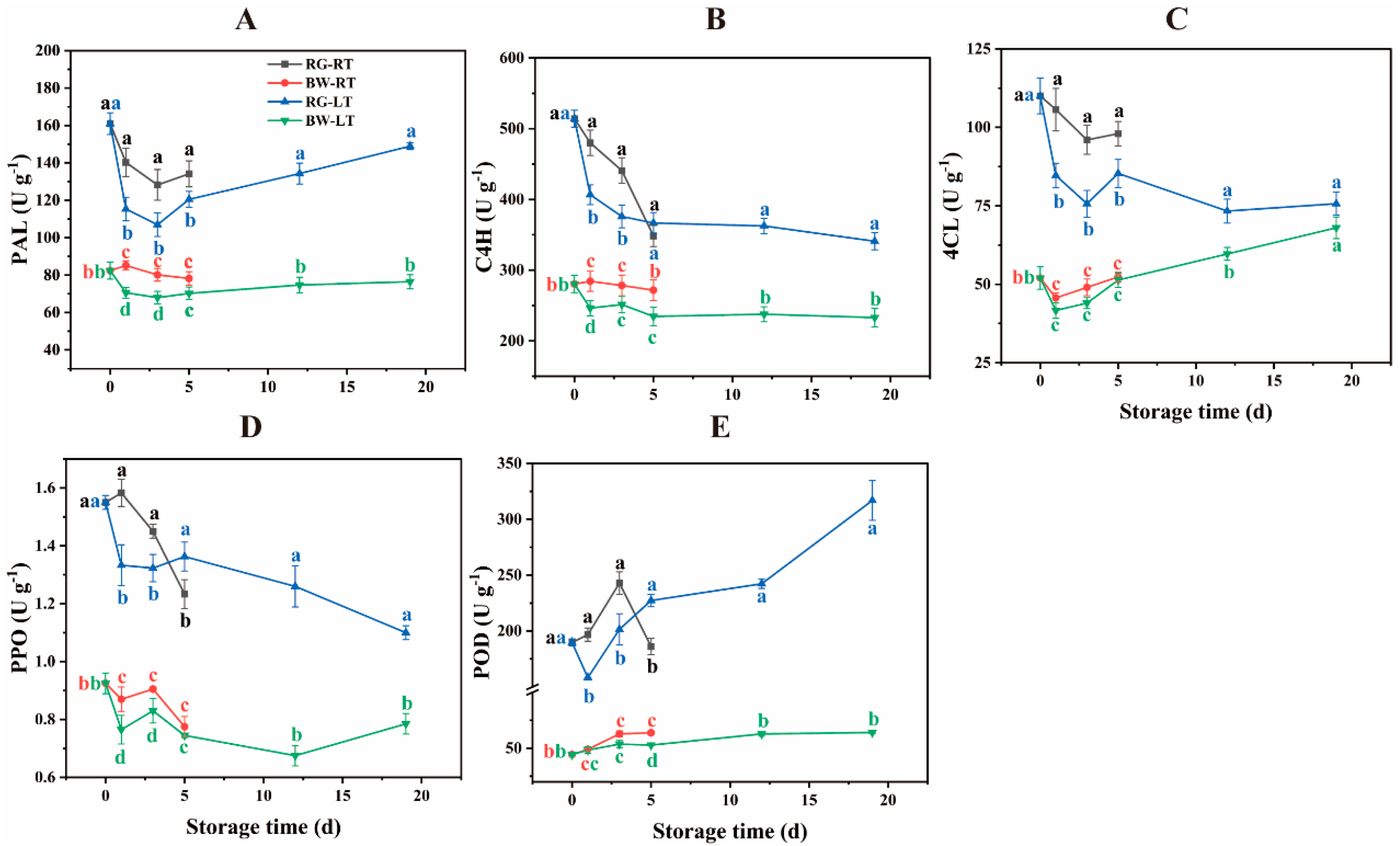
Figure 5.
Expression pattern of genes (qPCR) involved in the biosynthesis and degradation of phenolic compounds in different parts of chive during storage at 20 °C (RT) and 4 °C (LT). RG, the round green part. BW, the bottom white part. Data are the means of the three independent measurements. Letters represent statistical differences among different time points and tissues under RT or LT.
Figure 5.
Expression pattern of genes (qPCR) involved in the biosynthesis and degradation of phenolic compounds in different parts of chive during storage at 20 °C (RT) and 4 °C (LT). RG, the round green part. BW, the bottom white part. Data are the means of the three independent measurements. Letters represent statistical differences among different time points and tissues under RT or LT.
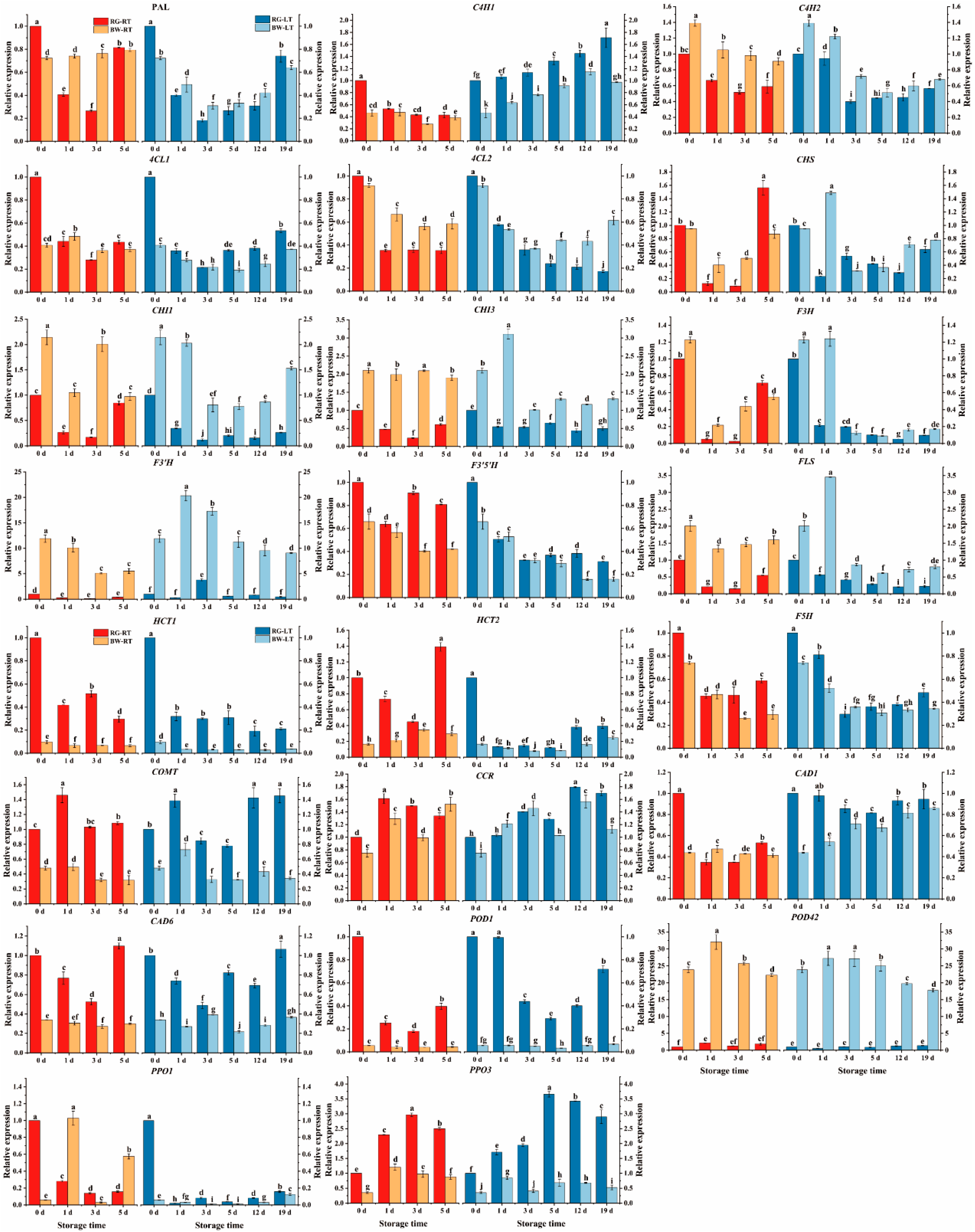
Figure 6.
(A) FRAP activity, (B) DPPH scavenging, (C) •OH scavenging and (D) O2•- scavenging in different parts of chive during storage at 20 °C (RT) and 4 °C (LT). RG, the round green part. BW, the bottom white part. Data are the means of the three independent measurements. Letters represent statistical differences among different tissues under RT and LT at each time point.
Figure 6.
(A) FRAP activity, (B) DPPH scavenging, (C) •OH scavenging and (D) O2•- scavenging in different parts of chive during storage at 20 °C (RT) and 4 °C (LT). RG, the round green part. BW, the bottom white part. Data are the means of the three independent measurements. Letters represent statistical differences among different tissues under RT and LT at each time point.
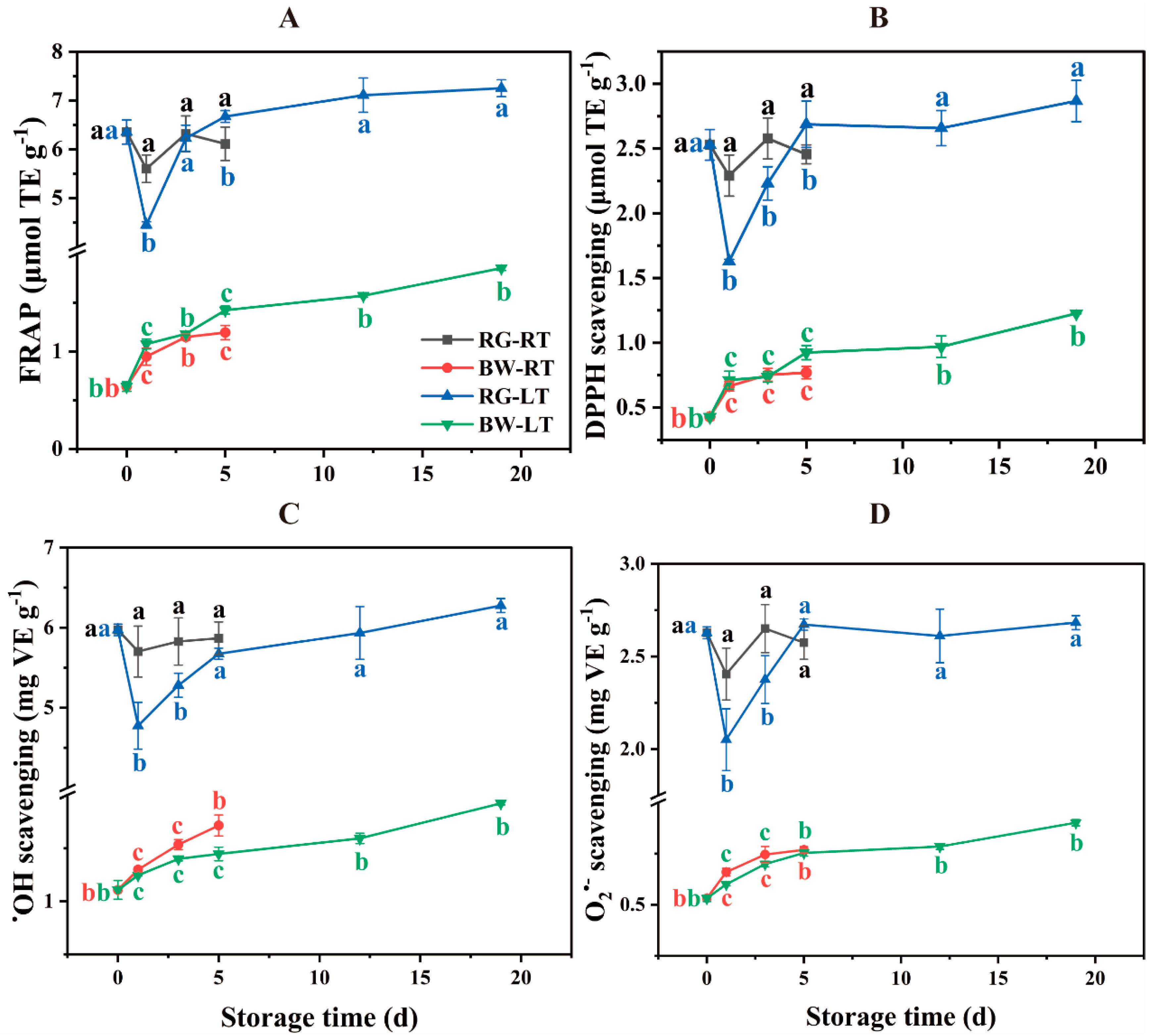
Figure 7.
Spearman’s correlation among tissue parts, phenolic compounds, AsA, related enzymes and antioxidant parameters in different parts of chive during storage at 20 °C (RT) and 4 °C (LT).
Figure 7.
Spearman’s correlation among tissue parts, phenolic compounds, AsA, related enzymes and antioxidant parameters in different parts of chive during storage at 20 °C (RT) and 4 °C (LT).
Figure 8.
(A) Comparison of gene expression and enzyme activities in the phenylpropanoid pathway in different parts of post-harvest chive and (B) the mechanism of biosynthesis, change and transfer of different nutrients in different parts of chive during storage. R, the round green part; B, the bottom white part. CSOs, S-alk(en)ylcysteine sulfoxides; FAAs, free amino acids. R and B in italics represent the expression of related genes in the corresponding part. R and B without italics represent the activity of related enzymes in the corresponding part. R and B in red denote that the tissue has a higher gene expression level or enzyme activity compared with that in other tissues. 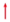 means increase.
means increase.  means decrease.
means decrease.  means change not too much. The thickness of the arrows indicates the extent of the change.
means change not too much. The thickness of the arrows indicates the extent of the change.  means inhibited by low temperature.
means inhibited by low temperature.
means increase. means decrease. means change not too much. The thickness of the arrows indicates the extent of the change. means inhibited by low temperature.
Figure 8.
(A) Comparison of gene expression and enzyme activities in the phenylpropanoid pathway in different parts of post-harvest chive and (B) the mechanism of biosynthesis, change and transfer of different nutrients in different parts of chive during storage. R, the round green part; B, the bottom white part. CSOs, S-alk(en)ylcysteine sulfoxides; FAAs, free amino acids. R and B in italics represent the expression of related genes in the corresponding part. R and B without italics represent the activity of related enzymes in the corresponding part. R and B in red denote that the tissue has a higher gene expression level or enzyme activity compared with that in other tissues. means increase. means decrease. means change not too much. The thickness of the arrows indicates the extent of the change. means inhibited by low temperature.
means increase. means decrease. means change not too much. The thickness of the arrows indicates the extent of the change. means inhibited by low temperature.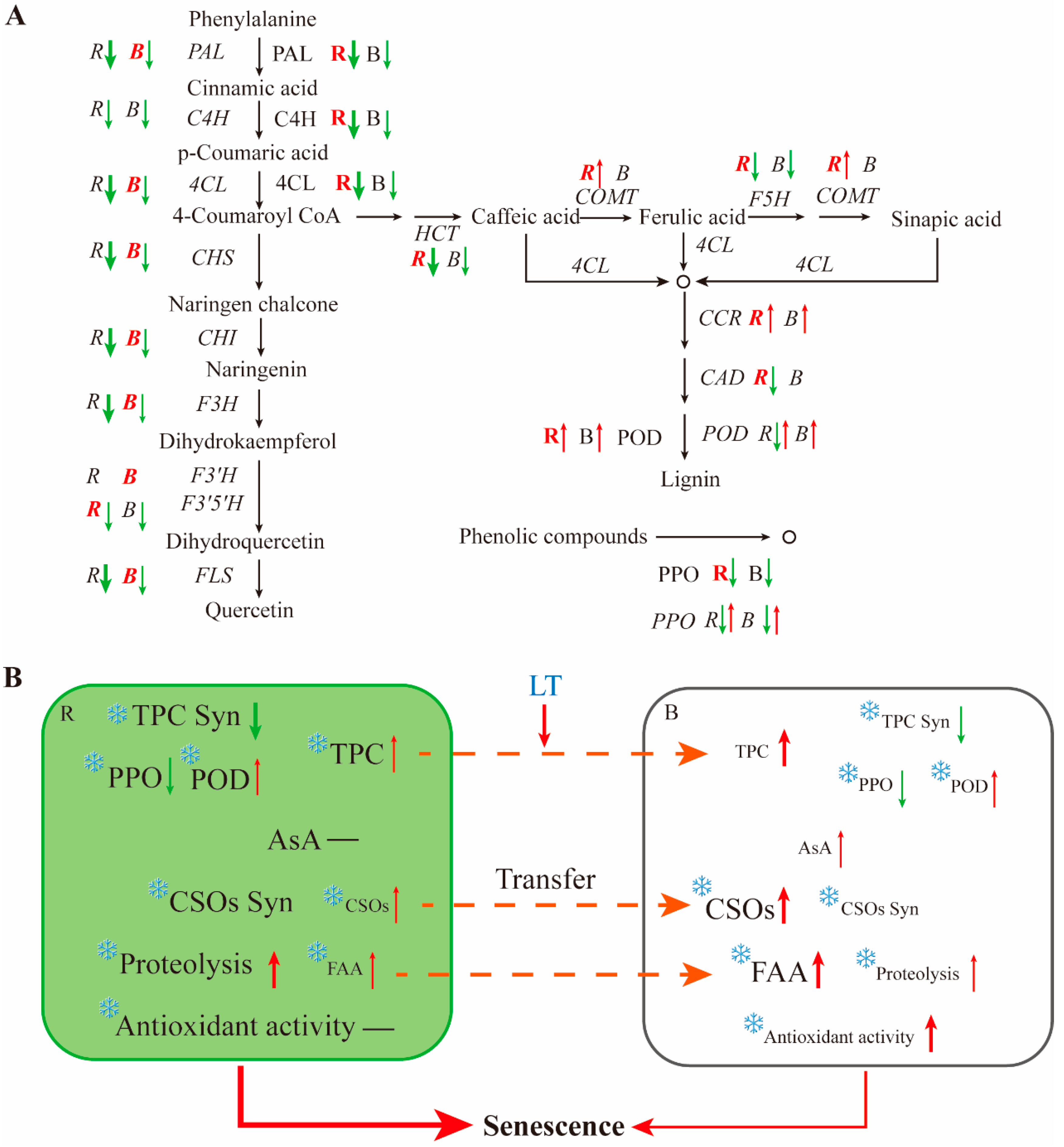
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



